N-Terminal Arm of Mcm1 Is Required for Transcription of...
12
EUKARYOTIC CELL, Nov. 2005, p. 1808–1819 Vol. 4, No. 11 1535-9778/05/$08.000 doi:10.1128/EC.4.11.1808–1819.2005 Copyright © 2005, American Society for Microbiology. All Rights Reserved. N-Terminal Arm of Mcm1 Is Required for Transcription of a Subset of Genes Involved in Maintenance of the Cell Wall Deepu S. Abraham and Andrew K. Vershon* Waksman Institute of Microbiology and Department of Molecular Biology and Biochemistry, Rutgers University, Piscataway, New Jersey 08854 Received 11 August 2005/Accepted 16 August 2005 The yeast Mcm1 protein is a member of the MADS box family of transcription factors that interacts with several cofactors to differentially regulate genes involved in cell-type determination, mating, cell cycle control and arginine metabolism. Residues 18 to 96 of the protein, which form the core DNA-binding domain of Mcm1, are sufficient to carry out many Mcm1-dependent functions. However, deletion of residues 2 to 17, which form the nonessential N-terminal (NT) arm, confers a salt-sensitive phenotype, suggesting that the NT arm is required for the activation of salt response genes. We used a strategy that combined information from the mutational analysis of the Mcm1-binding site with microarray expression data under salt stress conditions to identify a new subset of Mcm1-regulated genes. Northern blot analysis showed that the transcript levels of several genes encoding associated with the cell wall, especially YGP1, decrease significantly upon deletion of the Mcm1 NT arm. Deletion of the Mcm1 NT arm results in a calcofluor white-sensitive phenotype, which is often associated with defects in transcription of cell wall genes. In addition, the deletion makes cells sensitive to CaCl 2 and alkaline pH. We found that the defect caused by removal of the NT arm is not due to changes in Mcm1 protein level, stability, DNA-binding affinity, or DNA bending. This suggests that residues 2 to 17 of Mcm1 may be involved in recruiting a cofactor to the promoters of these genes to activate transcription. The MADS box family of proteins contains a highly con- served DNA-binding and dimerization domain, named after the founding members, Mcm1, Agamous, Deficiens, and serum response factor (SRF) (34, 36, 46, 51). MADS box proteins have been identified in most eukaryotic organisms, and these proteins are involved in the transcriptional regulation of a wide array of cellular and developmental processes (3, 26, 34, 46, 49). Many MADS box proteins are able to bind DNA as dimers with high affinity on their own in vitro. However, these proteins often interact with other DNA-binding cofactors to regulate transcription in vivo. Their interaction with different cofactors helps specify the DNA target sites bound by these proteins and thus the genes that they regulate. The conserved 80-amino-acid MADS box, DNA-binding, and dimerization domain is often sufficient for the interaction with these cofactors (6, 10, 37, 45). However, many MADS box proteins also appear to use regions outside of the conserved domain to interact with cofactors. For example, the 28-amino-acid-long myocyte enhancer factor (MEF) domain of MEF2 proteins in animals is found immediately C terminal to the MADS box domain and enables these proteins to heterodimerize with other members of the MEF2 family (32). In plants, the I region and K box outside the MADS box are involved in homo- and heterodimerization (11, 16, 39). Mcm1 is a MADS box transcriptional regulatory protein in the yeast Saccharomyces cerevisiae. The 286-amino-acid-long Mcm1 protein binds to a 10-bp palindromic sequence, fre- quently called the CArG box [CC(A/T) 6 GG] (31). In combi- nation with different cofactors, Mcm1 bound to CArG sites activates or represses the transcription of a variety of genes, including those involved in cell type determination, arginine metabolism, pheromone response, mating, and cell cycle pro- gression (20, 27, 29, 36). It is also involved in Ty-mediated transcription and minichromosome maintenance (36, 52). Mcm1 was recently found to bind to yeast autonomous repli- cating sequences (designated ARS) and initiate replication (8, 9). Mcm1 is an essential protein in yeast, and the conserved MADS box domain (residues 18 to 96; designated Mcm1-18- 96) is sufficient to maintain viability, DNA binding, dimeriza- tion, interaction with several cofactors, and transcriptional reg- ulation of many of its target genes (6, 10, 28). However, deletion of the nonessential N-terminal extension (residues 2 to 17) of Mcm1 causes cells to be sensitive to high-salt levels, indicating that regions outside the conserved MADS box are important for this function (24). In this paper, we investigate the role of the Mcm1 N-termi- nal (NT) arm in regulating cellular osmotolerance. We have found that deletion of the NT arm confers strong sensitivity to CaCl 2 . Deletion of the arm also confers sensitivity to cell wall- disrupting agents like calcofluor white (CFW) and alkaline pH. Importantly, we show here that the NT arm of Mcm1 is re- quired for the proper regulation of a subset of genes that encode components of the cell wall, such as YGP1 (for yeast glycoprotein 1). Our results suggest that the NT arm of Mcm1 may be needed for interaction with another cofactor that is required for the proper regulation of these cell wall genes. MATERIALS AND METHODS Plasmids and strains. Yeast strain JM01 (MAT leu2-3,112 trp1-1 ura3 his3- 11,15 ade2-1 can1-100 mcm1::LEU2/pSL1574-CEN/ARS MCM1 URA3) was used for all the in vivo assays (28). Plasmids with different mcm1 mutations were transformed into strain JM01 and selected on synthetic medium lacking histidine. Colonies that had lost the wild-type MCM1 on plasmid pSL1574 were selected by growth on 5-fluoroorotic acid plates. Mutations of the putative Mcm1 and Rlm1 * Corresponding author. Mailing address: Waksman Institute, 190 Frelinghuysen Rd., Piscataway, NJ 08854-8020. Phone: (732) 445-2905. Fax: (732) 445-5735. E-mail: [email protected]. 1808 on June 7, 2018 by guest http://ec.asm.org/ Downloaded from
Transcript of N-Terminal Arm of Mcm1 Is Required for Transcription of...

EUKARYOTIC CELL, Nov. 2005, p. 1808–1819 Vol. 4, No. 111535-9778/05/$08.00�0 doi:10.1128/EC.4.11.1808–1819.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
N-Terminal Arm of Mcm1 Is Required for Transcription of a Subsetof Genes Involved in Maintenance of the Cell Wall
Deepu S. Abraham and Andrew K. Vershon*Waksman Institute of Microbiology and Department of Molecular Biology and Biochemistry,
Rutgers University, Piscataway, New Jersey 08854
Received 11 August 2005/Accepted 16 August 2005
The yeast Mcm1 protein is a member of the MADS box family of transcription factors that interacts withseveral cofactors to differentially regulate genes involved in cell-type determination, mating, cell cycle controland arginine metabolism. Residues 18 to 96 of the protein, which form the core DNA-binding domain of Mcm1,are sufficient to carry out many Mcm1-dependent functions. However, deletion of residues 2 to 17, which formthe nonessential N-terminal (NT) arm, confers a salt-sensitive phenotype, suggesting that the NT arm isrequired for the activation of salt response genes. We used a strategy that combined information from themutational analysis of the Mcm1-binding site with microarray expression data under salt stress conditions toidentify a new subset of Mcm1-regulated genes. Northern blot analysis showed that the transcript levels ofseveral genes encoding associated with the cell wall, especially YGP1, decrease significantly upon deletion of theMcm1 NT arm. Deletion of the Mcm1 NT arm results in a calcofluor white-sensitive phenotype, which is oftenassociated with defects in transcription of cell wall genes. In addition, the deletion makes cells sensitive toCaCl2 and alkaline pH. We found that the defect caused by removal of the NT arm is not due to changes inMcm1 protein level, stability, DNA-binding affinity, or DNA bending. This suggests that residues 2 to 17 ofMcm1 may be involved in recruiting a cofactor to the promoters of these genes to activate transcription.
The MADS box family of proteins contains a highly con-served DNA-binding and dimerization domain, named afterthe founding members, Mcm1, Agamous, Deficiens, and serumresponse factor (SRF) (34, 36, 46, 51). MADS box proteinshave been identified in most eukaryotic organisms, and theseproteins are involved in the transcriptional regulation of a widearray of cellular and developmental processes (3, 26, 34, 46,49). Many MADS box proteins are able to bind DNA as dimerswith high affinity on their own in vitro. However, these proteinsoften interact with other DNA-binding cofactors to regulatetranscription in vivo. Their interaction with different cofactorshelps specify the DNA target sites bound by these proteins andthus the genes that they regulate. The conserved 80-amino-acidMADS box, DNA-binding, and dimerization domain is oftensufficient for the interaction with these cofactors (6, 10, 37, 45).However, many MADS box proteins also appear to use regionsoutside of the conserved domain to interact with cofactors. Forexample, the 28-amino-acid-long myocyte enhancer factor (MEF)domain of MEF2 proteins in animals is found immediately Cterminal to the MADS box domain and enables these proteins toheterodimerize with other members of the MEF2 family (32). Inplants, the I region and K box outside the MADS box are involvedin homo- and heterodimerization (11, 16, 39).
Mcm1 is a MADS box transcriptional regulatory protein inthe yeast Saccharomyces cerevisiae. The 286-amino-acid-longMcm1 protein binds to a 10-bp palindromic sequence, fre-quently called the CArG box [CC(A/T)6GG] (31). In combi-nation with different cofactors, Mcm1 bound to CArG sitesactivates or represses the transcription of a variety of genes,
including those involved in cell type determination, argininemetabolism, pheromone response, mating, and cell cycle pro-gression (20, 27, 29, 36). It is also involved in Ty-mediatedtranscription and minichromosome maintenance (36, 52).Mcm1 was recently found to bind to yeast autonomous repli-cating sequences (designated ARS) and initiate replication (8,9). Mcm1 is an essential protein in yeast, and the conservedMADS box domain (residues 18 to 96; designated Mcm1-18-96) is sufficient to maintain viability, DNA binding, dimeriza-tion, interaction with several cofactors, and transcriptional reg-ulation of many of its target genes (6, 10, 28). However,deletion of the nonessential N-terminal extension (residues 2to 17) of Mcm1 causes cells to be sensitive to high-salt levels,indicating that regions outside the conserved MADS box areimportant for this function (24).
In this paper, we investigate the role of the Mcm1 N-termi-nal (NT) arm in regulating cellular osmotolerance. We havefound that deletion of the NT arm confers strong sensitivity toCaCl2. Deletion of the arm also confers sensitivity to cell wall-disrupting agents like calcofluor white (CFW) and alkaline pH.Importantly, we show here that the NT arm of Mcm1 is re-quired for the proper regulation of a subset of genes thatencode components of the cell wall, such as YGP1 (for yeastglycoprotein 1). Our results suggest that the NT arm of Mcm1may be needed for interaction with another cofactor that isrequired for the proper regulation of these cell wall genes.
MATERIALS AND METHODS
Plasmids and strains. Yeast strain JM01 (MAT� leu2-3,112 trp1-1 ura3 his3-11,15 ade2-1 can1-100 mcm1::LEU2/pSL1574-CEN/ARS MCM1 URA3) was usedfor all the in vivo assays (28). Plasmids with different mcm1 mutations weretransformed into strain JM01 and selected on synthetic medium lacking histidine.Colonies that had lost the wild-type MCM1 on plasmid pSL1574 were selected bygrowth on 5-fluoroorotic acid plates. Mutations of the putative Mcm1 and Rlm1
* Corresponding author. Mailing address: Waksman Institute, 190Frelinghuysen Rd., Piscataway, NJ 08854-8020. Phone: (732) 445-2905.Fax: (732) 445-5735. E-mail: [email protected].
1808
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

sites in the YGP1-lacZ reporters were constructed by site-directed mutagenesisusing the Stratagene QuikChange kit. Plasmids used in this study are listed inTable 1. Sequences of the oligonucleotides used for creating probes, constructionof plasmids, and site-directed mutagenesis are available on request.
Biological assays. JM01 strains transformed with wild-type or mutant mcm1on HIS3 plasmids were grown overnight in liquid SD-His-Leu medium (28). Theconcentration of the cells in each culture was normalized after their opticaldensity at 600 nm was measured. Threefold serial dilutions (each, 3 �l) of thenormalized strains were spotted onto yeast extract-peptone-dextrose (YEPD)medium supplemented with the indicated concentrations of calcofluor white,KCl, sorbitol, dithiothreitol (DTT), and CaCl2. The pH of alkaline YEPD me-dium was adjusted to 8.5 with NaOH before being autoclaved. The �-galactosi-dase activities of YGP1 promoter-lacZ fusion reporters were determined in strainJM01 cotransformed with wild-type or mutant derivatives of pJM231 (MCM1HIS3 CEN) and the different lacZ reporter plasmids (2� URA3). Cells weregrown to log phase in minimal medium lacking leucine, histidine, and uracil;liquid �-galactosidase assays were performed as described previously (20).
Protein purification. The Mcm1 proteins used in the in vitro DNA-binding andDNA-bending studies were purified from Escherichia coli BL21 cells transformedwith pTBA25 or pTBA22 or maltose-binding protein fusion expression vectorswith sequences encoding Mcm1 residues 1 to 96 and 18 to 96, respectively. Theproteins were purified from 50-ml cultures as described previously (2). Proteinconcentrations were determined by Bradford assays, normalized, and verified bysodium dodecyl sulfate-polyacrylamide gel electrophoresis.
Northern blot analysis. Yeast strain JM01 was transformed with wild-type andmutant derivatives of pJM231 (MCM1 HIS3 CEN) or pAB76 (MCM1 HIS3 2�),and cultures were grown at 30°C in SD-His-Leu medium to mid-log phase(optical density at 600 nm, �0.6). The cells were split into two aliquots. One halfwas resuspended in dropout medium supplemented with 1.4 M NaCl and grownat 30°C for 1.5 h, while the other half was harvested after growth in mediumwithout a high salt level. Total RNA was extracted from both samples by hotacid-phenol extraction (5). RNA was separated by molecular weight on a 1.5%agarose gel containing formaldehyde and transferred to a Nytran membrane(Schleicher and Schuell) by capillary transfer. The membrane was prehybridizedin a 10-ml hybridization solution (50% formamide, 5� SSC [1� SSC is 0.15 MNaCl plus 0.015 M sodium citrate], 50 mM NaPO4 [pH 7.0], 1� Denhardt’sreagent, 0.1% SDS, 100-�g/ml sheared single-stranded salmon sperm DNA).DNA probes to open reading frames (ORFs) were generated by PCR andlabeled with [�-32P]dCTP by random priming (MegaPrime DNA Labeling Kit,Amersham). After overnight hybridization to labeled DNA probes and washingwith 2� SSC and 0.1% SDS at 42°C, specific mRNAs were detected by exposureto a phosphorscreen and imaging on a Molecular Dynamics phosphorimager.
Western blot analysis. Cultures of yeast strain JM01 transformed with C-termi-nal V5 epitope tagged full-length MCM1 and mcm1-�2-17 were grown to mid-logphase in SD-His-Leu medium. Proteins were extracted by glass bead lysis, and theprotein concentration was determined with the Bio-Rad Protein Assay kit. Proteins
were separated on 12% SDS-polyacrylamide gel, transferred to a nitrocellulosemembrane, probed with mouse antibody directed against the V5 epitope (Invitro-gen), and detected with sheep anti-mouse immunoglobulin G horseradish peroxi-dase (Amersham Pharmacia). The blot was stripped and probed with rabbit anti-Rpa1 antibodies (a gift from Steve Brill) and goat anti-rabbit immunoglobulin Ghorseradish peroxidase (Bio-Rad). Proteins were detected with the ECL Westernblot detection kit (Amersham Pharmacia Biotech). Protein levels were quantitatedwith FluorChem software. To determine the rate of degradation of Mcm1, cells weregrown to mid-log phase, and protein synthesis was arrested by the addition ofcycloheximide to a final concentration of 20 �g/ml (14). Cells were harvested every30 min for 4 h after the addition of cycloheximide. Protein extraction and detectionwere performed as described above.
Electrophoretic mobility shift assay (EMSA). Oligonucleotides with P(PAL)or YGP1 M1M2 sites were end labeled with [�-32P]ATP using polynucleotidekinase and purified using QIAGEN nucleotide removal columns. The labeledoligonucleotides were incubated at 90°C for 5 min with a threefold excess of thematching strand and allowed to anneal by being cooled slowly to 25°C. Thepurified Mcm1-1-96 and Mcm1-18-96 proteins were incubated for 1 h at roomtemperature with the labeled DNA probes in 20 mM Tris (pH 7.6), 5 mM MgCl2,0.1 mM EDTA, 0.1% NP-40, 5% glycerol, 10-mg/ml bovine serum albumin,10-mg/ml single-stranded DNA, and labeled oligonucleotide pairs (300 cpm/�l)in a total volume of 24 �l at room temperature for 1 h. All protein dilutions weremade in 50 mM Tris (pH 7.6), 500 mM NaCl, 1 mM EDTA, 10 mM �-mercap-toethanol, and 1-mg/ml bovine serum albumin. Samples were loaded on a 6%polyacrylamide gel (run in 0.5� Tris-borate-EDTA buffer for 1 h at 200 V). Gelswere dried after electrophoresis, exposed to a phosphor screen, and scanned witha Molecular Dynamics Phosphorimager.
Circular permutation assay. A circularly permuted plasmid, pDA192, with a80-bp fragment containing the R1, M2, and M3 sites from the YGP1 promotercloned into the SacI-XbaI polylinker site of pGD579 (2) was digested withBamHI, NheI, HindIII, or EcoRI to yield probes with the R1-M2-M3 regionlocated at different positions along the length of each DNA fragment. EMSAswere performed using position-permuted fragments and purified Mcm1-1-96 andMcm1-18-96. The bend angle for each protein-DNA complex was calculatedusing the Thompson and Landy relationship (48).
ChIP assay. Transformants of strain JM01 with wild-type and mutant deriva-tives of MCM1 with or without the V5 epitope tag were used for chromatinimmunoprecipitation (ChIP) assays. Proteins bound to DNA were cross-linkedby the addition of 1.35 ml of 37% formaldehyde to 50 ml of log-phase culturesand shaking at room temperature for 15 min. The cells were then harvested andlysed with glass beads. The crude lysate was sonicated to shear the protein-boundDNA to approximately 500-bp-long fragments. After cell debris was removed bycentrifugation, the total chromatin was precleared by nutation with proteinG-agarose for 1 h and centrifugation. The supernatant was then incubated withthe anti-V5 antibody overnight at 4°C. Protein G-agarose was added to immu-noprecipitate the sample. Reversal of formaldehyde cross-links was carried out
TABLE 1. Plasmids used in this study
Plasmid Description Referenceor source
pJM231 MCM1 CEN HIS3 1pDA126 mcm1-�2-17 CEN HIS3 This studypDA135 C-terminally-V5 tagged MCM1 CEN HIS3 This studypDA136 C-terminally-V5 tagged mcm1-�2-17 CEN HIS3 This studypDA135-T35A C-terminally-V5 tagged mcm1-T35A CEN HIS3 This studypDA181 700 bp of YGP1 promoter lacZ reporter This studypDA181-r1 pDA181 with mutation in site R1 This studypDA181-m2 pDA181 with mutation in site M2 This studypDA181-m3 pDA181 with mutation in site M3 This studypDA181-m4 pDA181 with mutation in site M4 This studypDA181-r1m2 pDA181 with mutation in sites R1, M2 This studypDA181-m2m3 pDA181 with mutation in sites M2, M3 This studypDA181-m1m2m3 pDA181 with mutation in sites M1, M2, M3 This studypDA181-r1m2m3 pDA181 with mutation in sites R1, M2, M3 This studypDA181-m1m2m3m4 pDA181 with mutation in sites M1, M2, M3, M4 This studypDA181-r1m1m2m3 pDA181 with mutation in sites R1, M1, M2, M3 This studypDA192 R1M2M3 site cloned into pGD579 This studypTBA22 Mcm1-18-96 fused to maltose-binding protein 2pTBA25 Mcm1-1-96 fused to maltose-binding protein 2
VOL. 4, 2005 Mcm1 REGULATES CELL WALL GENES 1809
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from
as described previously (22). The DNA fragments precipitated with theV5-tagged Mcm1 proteins were detected by PCR using primers that annealed tothe target promoter regions. Each PCR used three sets of primers: one set toamplify the positive control for Mcm1 binding, another for the test promoter,and a third for the negative control.
RESULTS
Identification of genes requiring the NT arm of Mcm1 foractivation. Deletion of the NT arm of Mcm1 (residues 2 to 17)makes cells osmosensitive (24). We therefore reasoned thatthe NT arm may be required for the transcriptional activationof genes required for tolerance to high salt levels. To identifypotential Mcm1-activated salt response genes, we scanned thepromoter regions of genes that showed increased expressionunder high-salt conditions for sites that are similar to theconsensus sequence ccywwwxxrg (where y T/C, w A/T, x any base, and r A/G) that was derived from mutationalanalysis of an Mcm1-binding site (2, 50). Among the 570 salt-induced genes that were induced twofold or more in the mi-croarray analysis, 163 genes had one or more putative Mcm1-binding sites in the 500-bp region upstream of their start codon(50). Among this group, we focused on genes that containedmultiple Mcm1-binding sites or that were known to be involvedin the osmotic stress response. To test if the NT arm of Mcm1
was required for the regulation of the selected salt-inducedgenes, we compared their expression by Northern blot analysisin strains containing wild-type Mcm1 or Mcm1-�2-17 grownunder high- and low-salt conditions. Deletion of Mcm1 resi-dues 2 to 17 resulted in a decrease in the mRNA levels ofYGP1, PNC1, DAK1, and YHR087W (Fig. 1). The most dra-matic effect was observed for YGP1, in which both the nonin-duced and salt-induced transcription decreased to 10% ofwild-type levels. In addition to causing a decrease in expressionfor some genes, the deletion of the NT arm caused an increasein the expression of URA1 and SIT1. This suggests that the NTarm might be required for Mcm1 to function as a repressor at
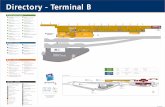
FIG. 1. The N-terminal arm of Mcm1 is required for the regulationof a specific subset of salt-induced and cell wall genes. The mRNAlevels of genes with putative Mcm1-binding sites were measured underlow- and high-salt conditions by Northern blot analysis of strain JM01transformed with either wild-type MCM1 (pDA135) or mcm1-�2-17(pDA136). The CAR2 and CLB2 genes are regulated by Mcm1 but donot require the NT arm for activation. ACT1 is shown as a loadingcontrol.
FIG. 2. Strains containing mcm1-�2-17 are sensitive to CaCl2, KCl,CFW, and pH 8.5, but not sorbitol or DTT. Serial dilutions (threefold)of yeast strain JM01 transformed with plasmids containing MCM1 ormcm1-�2-17 were spotted onto the indicated media and grown at 30°Cfor 2 to 5 days.
1810 ABRAHAM AND VERSHON EUKARYOT. CELL
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

these loci. In contrast to YGP1, the mRNA level of CAR2, asalt-induced arginine catabolic gene that is activated by theMcm1-Arg80 complex, was unaffected by deletion of the NTarm (Fig. 1) (4). Deletion of the NT arm also did not affectexpression of CLB2, a target of the Mcm1-Fkh1 complex, orSTE3, an �-specific gene activated by the Mcm1-�1 complex(Fig. 1 and data not shown) (21, 43). Taken together, theseresults indicate that Mcm1 residues 2 to 17 are required for theactivation of a specific subset of genes that are induced by salt.
Among the genes that were assayed, deletion of the NT armhad the greatest effect on YGP1, which encodes a protein thatis secreted from protoplasts during cell wall regeneration.Ygp1 is highly glycosylated, and its synthesis is induced inresponse to glucose, nitrogen, phosphate starvation, and cellwall disruptions (12, 35). We therefore wanted to determine ifother stress-induced genes that were coinduced with YGP1under these conditions also required the Mcm1 NT arm foractivation. YGP1 is one of 25 genes that are regulated by Rlm1,another MADS box transcription factor (18). Twenty of thesegenes had putative Mcm1 sites in the 1-kb region upstream oftheir start sites. However, Northern blot analysis showed thatonly the salt-induced expression of SPS100 was dependent onthe presence of the Mcm1 NT arm (Fig. 1 and data not shown).Since YGP1 is induced in response to disruption of the cellwall, we also examined the promoters of genes that are inducedin response to various cell wall stresses (25). Among the 80genes categorized as the cell wall compensatory cluster, 38 hadputative Mcm1-binding sites in their promoters. The expres-sion of 15 of these genes was assayed under high-salt condi-tions in Mcm1 and Mcm1-�2-17 strains by Northern blotting.Among them, only SPI1 and YBR071W expression was depen-dent on the presence of the NT arm by Northern analysis(Fig. 1 and data not shown).
Deletion of the NT arm causes cell wall defects. Deletion ofthe Mcm1 NT arm causes sensitivity to increased NaCl levels(24). Since the Mcm1 NT arm is required for the properregulation of a subset of cell wall biosynthesis genes, we wereinterested in determining if this mutation affected sensitivity toother cell wall related stresses. While the mcm1-�2-17 strainwas very sensitive to YEPD medium with 0.35 M CaCl2(Fig. 2B), it was only mildly sensitive to 1.3 M KCl (Fig. 2C)and hardly sensitive to 0.4 M LiCl (data not shown).
Since the NT arm of Mcm1 was required for the properexpression of genes that encode components of the cell wall,we tested cells with mcm1-�2-17 for sensitivity to several cellwall-disrupting agents. The drug CFW is a fluorescent dye thatbinds to chitin in the cell wall and is thought to weaken the cell
wall by preventing microfibril formation (15, 33, 38). CFWsensitivity is commonly used as an indicator of defects in cellwall biogenesis. Deletion of the Mcm1 NT arm conferred astrong sensitivity to CFW (Fig. 2D). Deletion of the Mcm1 NTarm also caused the cells to be sensitive to alkaline pH(Fig. 2E). However, the mutant was not sensitive to otheragents that are commonly used to monitor defects in the cellwall, such as 2 M sorbitol, 50 mM DTT, and acidic pH (Fig. 2Fand G and data not shown). These data suggest that Mcm1 isrequired for the transcriptional regulation of a subset of cellwall genes that are required for CFW and high pH resistancebut not DTT or sorbitol resistance.
CFW interacts with chitin in the yeast cell wall, and exposureto CFW induces chitin synthesis (40–42). CHS1, CHS2, andCHS3 encode the catalytic subunits of chitin synthetases.There are three putative Mcm1-binding sites in the 1-kb regionupstream of the CHS2 start codon. However, Northern anal-ysis showed that CHS1, CHS2, and CHS3 mRNA levels werenot altered by the Mcm1 NT arm deletion, suggesting that theCFW-sensitive phenotype is not caused by improper expres-sion of these genes (data not shown).
Effect of Mcm1 point mutations on YGP1 transcription andCFW and alkaline pH tolerance. Although there are multiplephosphorylated isoforms of Mcm1, residues S2 and T8 in theMcm1 NT arm are the two major phosphorylation sites in theprotein (24). Under normal conditions, residue S2 in the NTarm is phosphorylated with nearly 100% efficiency and aserine-to-alanine mutation at position S2, which mimics theconstitutively dephosphorylated form of S2, has a salt-sensitivephenotype. This result suggested that phosphorylation of S2was required for the induction of Mcm1-dependent salt re-sponse genes. Therefore, it is possible that phosphorylation ofresidues in the NT arm also has a role in YGP1 regulation. Wetested this hypothesis by making alanine (A) and aspartate (D)substitutions of S2, T8, and T10 to mimic constitutive dephos-phorylation and phosphorylation, respectively, of these resi-dues. If differential phosphorylation states of the NT arm wereimportant for the activation of YGP1, either A or D substitu-tions would be expected to have antagonistic effects on itstranscription. However, Northern analysis showed that boththe A and D substitutions of residues in the NT arm onlyresulted in slightly lower levels of the YGP1 transcript(Fig. 3A). Different combinations of these mutations also didnot significantly affect YGP1 expression. In contrast, point mu-tations (V34A, T35A, S37A, K40A, and T66A) in the MADSbox domain of Mcm1 that were previously shown to decreaseDNA-binding affinity or bending and thus affect transcriptional
FIG. 3. The presence of the NT arm rather than its phosphorylation state regulates YGP1 transcription. Northern blots are shown for theexpression of YGP1 and ACT1 (loading control) for strains containing wild-type MCM1 and the indicated mcm1 mutants.
VOL. 4, 2005 Mcm1 REGULATES CELL WALL GENES 1811
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from
activation were defective in YGP1 transcription (see Fig. 5Band data not shown).
We next tested the effects of the amino acid substitutions inthe NT arm and MADS box domain on resistance to cellwall-disrupting agents. Although deletion of the Mcm1 NTarm conferred a pronounced growth defect on rich mediumsupplemented with CaCl2 (Fig. 2B), single amino acid substi-tutions in the NT arm or MADS box domain were not sensitiveto CaCl2 (data not shown). However, although significantly lessthan the 243-fold decrease in growth caused by deletion of theNT arm, A and D substitutions of residues S2, T8, and T10caused 3- to 27-fold decreases in growth on medium with CFWor alkaline pH (Fig. 4A and B). Alanine replacements of otherresidues in the arm that are not phosphorylated also made thecells sensitive to CFW. Since deletion of the NT arm showed amuch stronger phenotype than individual point mutations inthe arm, it appears that the entire arm is important for itsfunction.
We also examined the relative contribution of different res-idues in the MADS box domain toward CFW and alkaline pHtolerance. A comparison of the mutant strains showed that thesame regions in Mcm1 are important for tolerance to both
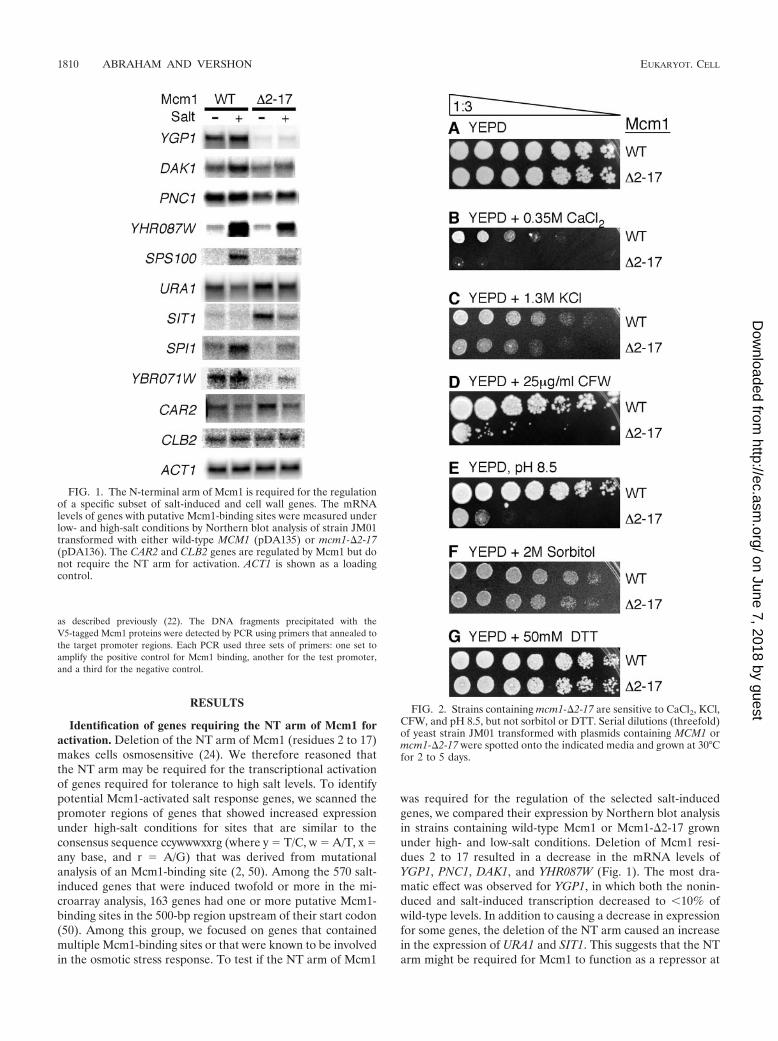
CFW (Fig. 4A) and alkaline pH (Fig. 4B). In general, themutations had a more severe growth defect on CFW than highpH, underscoring the sensitivity of the CFW phenotype overthe pH phenotype. Interestingly, the mutations that were themost defective in these assays mapped to three locations on thecrystal structure of the Mcm1 dimer bound to DNA. One classof mutants, exemplified by L60, L61, V62, and V63, consists ofresidues along the dimer interface. It is not surprising thatmutations of these residues affect Mcm1-mediated functions,most likely by adversely affecting Mcm1 homodimerization.The second class of mutants, represented by V81A, G86A, andR87A, form a hydrophobic groove on the surface of Mcm1(Fig. 4C). In the crystal structure of the Mcm1-�2-DNA com-plex, this region of Mcm1 interacts with residues in the �2protein and it is possible that Mcm1 uses these same residuesto interact with another cofactor required for induction of thecell wall-associated genes identified (47). The third class ofmutants, I23A, F48A, and E49A, are positioned near the NTarm in the crystal structure (Fig. 4D). These mutants havepreviously been shown to affect the expression of promotersdriven by the Mcm1-�1 and Mcm1-Ste12 complexes, but not byMcm1 alone or repression by the Mcm1-�2 complex (28). The
FIG. 4. Effects of amino acid substitutions in Mcm1 on CFW and alkaline pH tolerance. Each of the indicated mutants was assayed for growthon YEPD medium with 25-mg/ml CFW (A) and alkaline pH 8.5 (B). Bars represent the fold decrease in growth of the Mcm1 mutations indicatedat the bottom, relative to wild-type MCM1. X, a mutant that was not tested. Mutants that affect activation in complex with �1 (dark circles), Ste12(gray circles), or Mcm1 on its own (open circles) are shown for comparison (28). (C) Model of the Mcm1 dimer bound to DNA and positions ofmutations (highlighted in black) that result in significant growth defects on CFW and alkaline pH. Residues 15 to 22 that form part of the NT armare shaded gray. The protein is shown in spacefill and the DNA is shown as a wireframe.
1812 ABRAHAM AND VERSHON EUKARYOT. CELL
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

sensitivity of the mutants belonging to the second and thirdcategories suggests that Mcm1 may be interacting with a co-factor to activate the transcription of genes required for resis-tance to CFW and alkaline pH.
Mutations that suppress the CFW, alkaline pH, and CaCl2sensitivity to deletion of the Mcm1 NT arm do not suppressthe transcriptional defect of YGP1. The T35A mutation in theMADS box domain of Mcm1 caused cells to grow better thanthe wild-type strain in the presence of high salt levels (23). Thismutation also suppressed the effect of the NT arm deletion.We therefore wanted to determine if the T35A mutation wasable to suppress the effects of the Mcm1 NT arm deletion onsensitivity to cell wall-disrupting agents. On its own, the T35AMcm1 mutant grew as well as the wild-type MCM1 strain onmedium with CaCl2 (Fig. 5). In contrast, this mutant was de-fective in growth on plates with CFW and alkaline pH, al-though not to the same extent as mcm1-�2-17. As was ob-served for the NaCl-sensitive phenotype, the T35A mutant wasable to partially suppress the CaCl2-sensitive phenotype of theMcm1 NT arm deletion. Similarly, the T35A mutation partiallysuppressed the growth defect of mcm1-�2-17 on alkaline pHand very weakly on CFW.
Since YGP1 transcription was induced with high salt levels andwas adversely affected by the deletion of the Mcm1 NT arm, wewondered if the T35A mutation suppressed the effect of Mcm1-�2-17 on expression of the gene. However, Northern blot analysisshowed that both Mcm1-T35A and Mcm1-�2-17,T35A causeddecreases in YGP1 expression (Fig. 5B). Taken together, thesedata suggest that the mechanism of Mcm1-dependent regulationof genes required for growth on CFW and with alkaline pH andCaCl2 is different than Mcm1 regulation of YGP1.
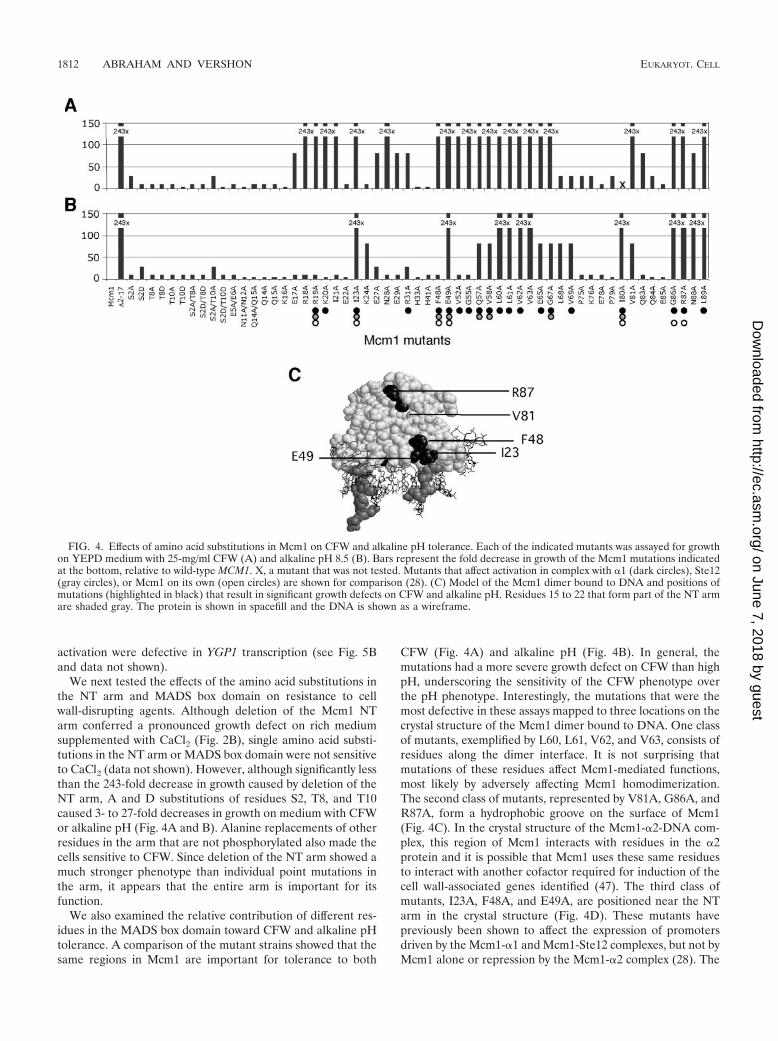
The YGP1 promoter is regulated by two MADS box proteins,Mcm1 and Rlm1. Since deletion of the Mcm1 NT arm showedthe largest effect on the transcription of YGP1, we decided toanalyze the YGP1 promoter to understand the mode of regu-lation of this gene by Mcm1. To monitor expression of thegene, we fused the 700-bp promoter region 5� to the YGP1ORF to the lacZ gene. The reporter was strongly expressed ina wild-type strain, but its expression was decreased almost 300fold in an mcm1-�2-17 strain (Fig. 6). This result showed thatthe regulatory elements that responded to the deletion of theMcm1 NT arm were located in this region of the YGP1 pro-moter. Sequence inspection of the YGP1 reporter predictedfour potential Mcm1-binding sites, which we called M1, M2,M3, and M4. To determine if the putative Mcm1-binding siteswere functional, these sites were mutated individually and incombination in the context of the YGP1-lacZ reporter, andtheir effect on YGP1 expression was measured by assaying�-galactosidase activity in cells with wild-type Mcm1. Individ-ual disruption of each of these sites caused a partial decreasein expression of the reporter, indicating that these sites arerequired for regulation of YGP1 and are likely to be functionalMcm1 targets. The relative effect of each of the mutants var-ied, suggesting that these sites may be bound by Mcm1 withdiffering affinities. Mutation of all four Mcm1-binding sites(M1, M2, M3, and M4) resulted in a significant decrease inactivation, similar to �-galactosidase levels obtained by dele-tion of the NT arm.
In addition to the Mcm1-binding sites, there is a putativebinding site for the Rlm1 MADS box protein in the YGP1
promoter. Rlm1 is activated through phosphorylation by theMpk1 MAP kinase in response to cell wall integrity signaling(13). Northern blot analysis of a strain with constitutively ac-tivated Rlm1 showed that YGP1 expression was decreased,suggesting that Rlm1 functions as a repressor of this gene (18).However, mutation of the Rlm1-binding site in the YGP1-lacZfusion reporter caused a slight decrease in expression both onits own and in combination with mutations in the Mcm1 sites(Fig. 6). This result suggests that Rlm1 may have a direct rolein activating YGP1 expression under the noninducing growthcondition used.
FIG. 5. (A) The T35A mutation in Mcm1 suppresses the growthdefect of Mcm1-�2-17 on media containing CFW, pH 8.5, and CaCl2.Serial dilutions (threefold) of yeast strain JM01 transformed withplasmids containing MCM1, mcm1-�2-17, mcm1-T35A, or mcm1-�2-17,T35A were spotted onto the indicated media and grown at 30°C for2 to 5 days. (B) The T35A mutation does not suppress the defectiveYGP1 transcription of Mcm1-�2-17. Northern blots are shown for theexpression of YGP1 and ACT1 (loading control) for strains containingwild-type MCM1 and the indicated mcm1 mutants.
VOL. 4, 2005 Mcm1 REGULATES CELL WALL GENES 1813
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from
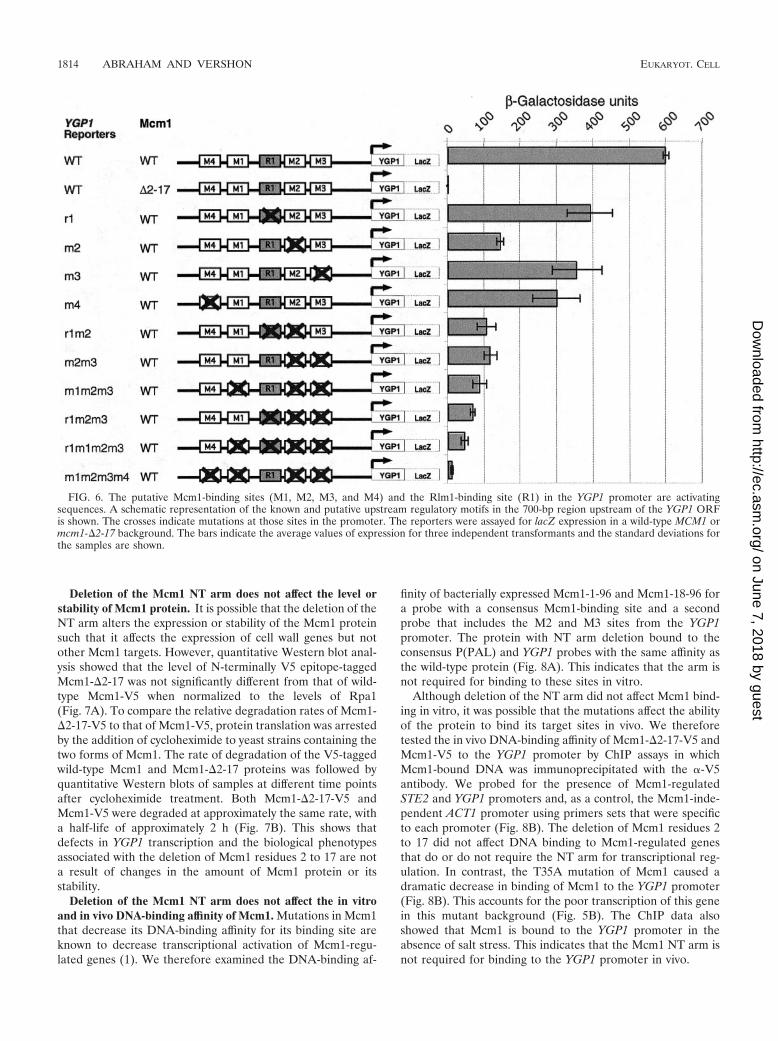
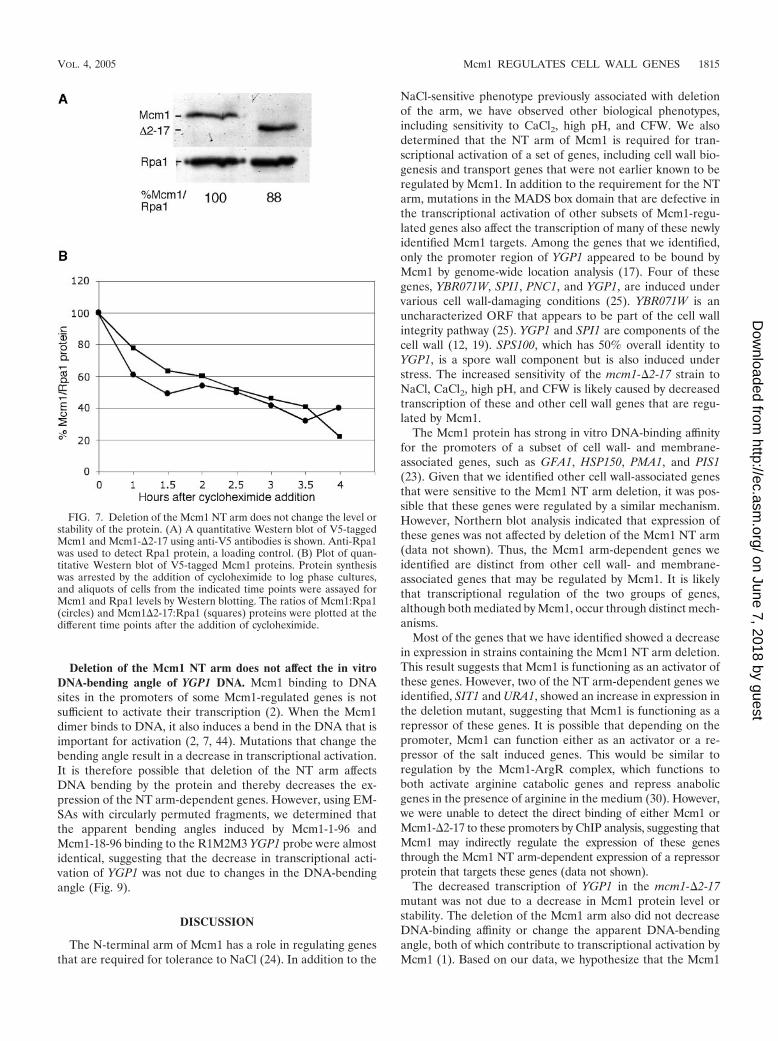
Deletion of the Mcm1 NT arm does not affect the level orstability of Mcm1 protein. It is possible that the deletion of theNT arm alters the expression or stability of the Mcm1 proteinsuch that it affects the expression of cell wall genes but notother Mcm1 targets. However, quantitative Western blot anal-ysis showed that the level of N-terminally V5 epitope-taggedMcm1-�2-17 was not significantly different from that of wild-type Mcm1-V5 when normalized to the levels of Rpa1(Fig. 7A). To compare the relative degradation rates of Mcm1-�2-17-V5 to that of Mcm1-V5, protein translation was arrestedby the addition of cycloheximide to yeast strains containing thetwo forms of Mcm1. The rate of degradation of the V5-taggedwild-type Mcm1 and Mcm1-�2-17 proteins was followed byquantitative Western blots of samples at different time pointsafter cycloheximide treatment. Both Mcm1-�2-17-V5 andMcm1-V5 were degraded at approximately the same rate, witha half-life of approximately 2 h (Fig. 7B). This shows thatdefects in YGP1 transcription and the biological phenotypesassociated with the deletion of Mcm1 residues 2 to 17 are nota result of changes in the amount of Mcm1 protein or itsstability.
Deletion of the Mcm1 NT arm does not affect the in vitroand in vivo DNA-binding affinity of Mcm1. Mutations in Mcm1that decrease its DNA-binding affinity for its binding site areknown to decrease transcriptional activation of Mcm1-regu-lated genes (1). We therefore examined the DNA-binding af-
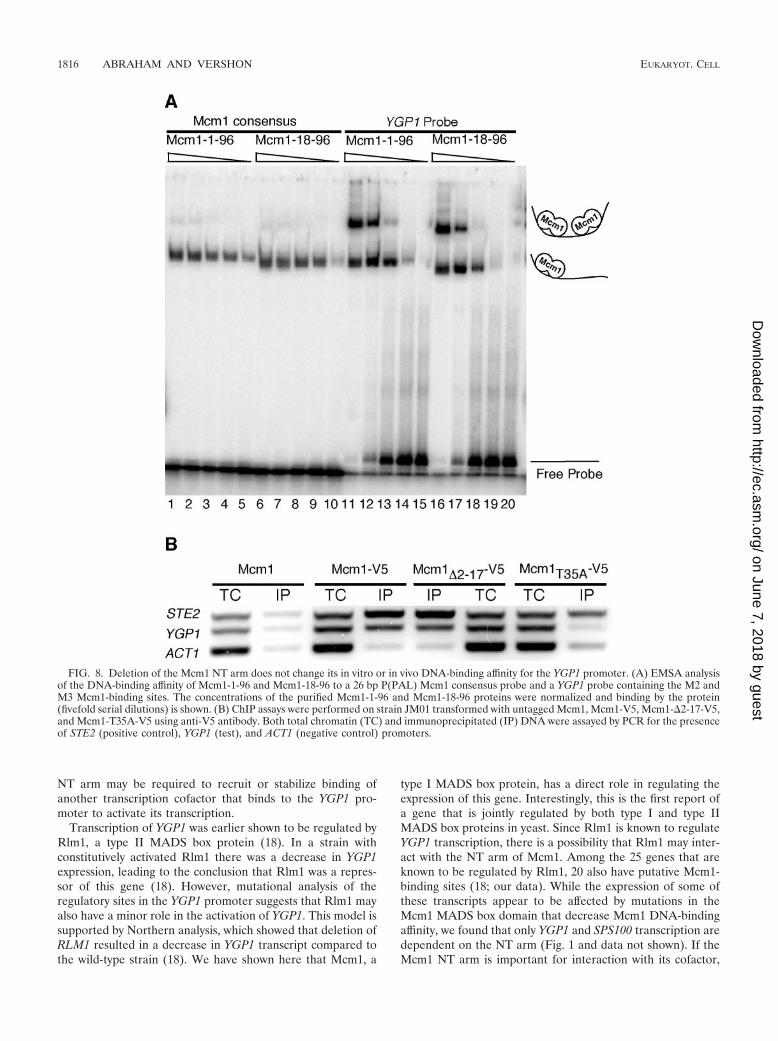
finity of bacterially expressed Mcm1-1-96 and Mcm1-18-96 fora probe with a consensus Mcm1-binding site and a secondprobe that includes the M2 and M3 sites from the YGP1promoter. The protein with NT arm deletion bound to theconsensus P(PAL) and YGP1 probes with the same affinity asthe wild-type protein (Fig. 8A). This indicates that the arm isnot required for binding to these sites in vitro.
Although deletion of the NT arm did not affect Mcm1 bind-ing in vitro, it was possible that the mutations affect the abilityof the protein to bind its target sites in vivo. We thereforetested the in vivo DNA-binding affinity of Mcm1-�2-17-V5 andMcm1-V5 to the YGP1 promoter by ChIP assays in whichMcm1-bound DNA was immunoprecipitated with the �-V5antibody. We probed for the presence of Mcm1-regulatedSTE2 and YGP1 promoters and, as a control, the Mcm1-inde-pendent ACT1 promoter using primers sets that were specificto each promoter (Fig. 8B). The deletion of Mcm1 residues 2to 17 did not affect DNA binding to Mcm1-regulated genesthat do or do not require the NT arm for transcriptional reg-ulation. In contrast, the T35A mutation of Mcm1 caused adramatic decrease in binding of Mcm1 to the YGP1 promoter(Fig. 8B). This accounts for the poor transcription of this genein this mutant background (Fig. 5B). The ChIP data alsoshowed that Mcm1 is bound to the YGP1 promoter in theabsence of salt stress. This indicates that the Mcm1 NT arm isnot required for binding to the YGP1 promoter in vivo.
FIG. 6. The putative Mcm1-binding sites (M1, M2, M3, and M4) and the Rlm1-binding site (R1) in the YGP1 promoter are activatingsequences. A schematic representation of the known and putative upstream regulatory motifs in the 700-bp region upstream of the YGP1 ORFis shown. The crosses indicate mutations at those sites in the promoter. The reporters were assayed for lacZ expression in a wild-type MCM1 ormcm1-�2-17 background. The bars indicate the average values of expression for three independent transformants and the standard deviations forthe samples are shown.
1814 ABRAHAM AND VERSHON EUKARYOT. CELL
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from
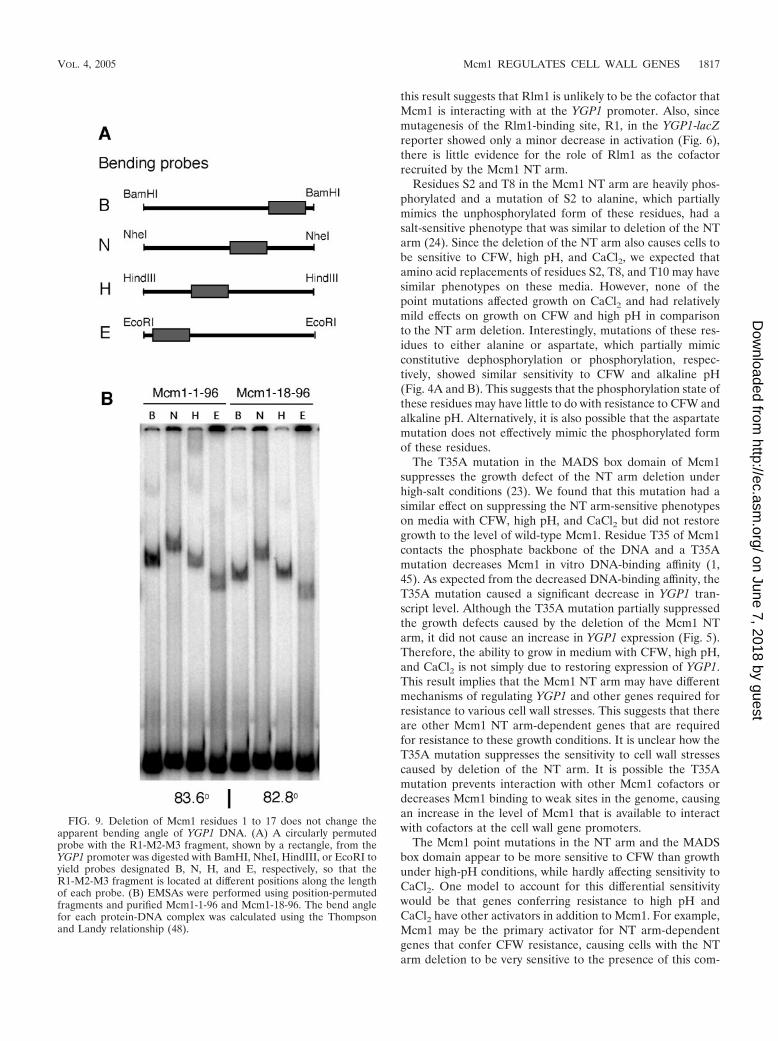
Deletion of the Mcm1 NT arm does not affect the in vitroDNA-bending angle of YGP1 DNA. Mcm1 binding to DNAsites in the promoters of some Mcm1-regulated genes is notsufficient to activate their transcription (2). When the Mcm1dimer binds to DNA, it also induces a bend in the DNA that isimportant for activation (2, 7, 44). Mutations that change thebending angle result in a decrease in transcriptional activation.It is therefore possible that deletion of the NT arm affectsDNA bending by the protein and thereby decreases the ex-pression of the NT arm-dependent genes. However, using EM-SAs with circularly permuted fragments, we determined thatthe apparent bending angles induced by Mcm1-1-96 andMcm1-18-96 binding to the R1M2M3 YGP1 probe were almostidentical, suggesting that the decrease in transcriptional acti-vation of YGP1 was not due to changes in the DNA-bendingangle (Fig. 9).
DISCUSSION
The N-terminal arm of Mcm1 has a role in regulating genesthat are required for tolerance to NaCl (24). In addition to the
NaCl-sensitive phenotype previously associated with deletionof the arm, we have observed other biological phenotypes,including sensitivity to CaCl2, high pH, and CFW. We alsodetermined that the NT arm of Mcm1 is required for tran-scriptional activation of a set of genes, including cell wall bio-genesis and transport genes that were not earlier known to beregulated by Mcm1. In addition to the requirement for the NTarm, mutations in the MADS box domain that are defective inthe transcriptional activation of other subsets of Mcm1-regu-lated genes also affect the transcription of many of these newlyidentified Mcm1 targets. Among the genes that we identified,only the promoter region of YGP1 appeared to be bound byMcm1 by genome-wide location analysis (17). Four of thesegenes, YBR071W, SPI1, PNC1, and YGP1, are induced undervarious cell wall-damaging conditions (25). YBR071W is anuncharacterized ORF that appears to be part of the cell wallintegrity pathway (25). YGP1 and SPI1 are components of thecell wall (12, 19). SPS100, which has 50% overall identity toYGP1, is a spore wall component but is also induced understress. The increased sensitivity of the mcm1-�2-17 strain toNaCl, CaCl2, high pH, and CFW is likely caused by decreasedtranscription of these and other cell wall genes that are regu-lated by Mcm1.
The Mcm1 protein has strong in vitro DNA-binding affinityfor the promoters of a subset of cell wall- and membrane-associated genes, such as GFA1, HSP150, PMA1, and PIS1(23). Given that we identified other cell wall-associated genesthat were sensitive to the Mcm1 NT arm deletion, it was pos-sible that these genes were regulated by a similar mechanism.However, Northern blot analysis indicated that expression ofthese genes was not affected by deletion of the Mcm1 NT arm(data not shown). Thus, the Mcm1 arm-dependent genes weidentified are distinct from other cell wall- and membrane-associated genes that may be regulated by Mcm1. It is likelythat transcriptional regulation of the two groups of genes,although both mediated by Mcm1, occur through distinct mech-anisms.
Most of the genes that we have identified showed a decreasein expression in strains containing the Mcm1 NT arm deletion.This result suggests that Mcm1 is functioning as an activator ofthese genes. However, two of the NT arm-dependent genes weidentified, SIT1 and URA1, showed an increase in expression inthe deletion mutant, suggesting that Mcm1 is functioning as arepressor of these genes. It is possible that depending on thepromoter, Mcm1 can function either as an activator or a re-pressor of the salt induced genes. This would be similar toregulation by the Mcm1-ArgR complex, which functions toboth activate arginine catabolic genes and repress anabolicgenes in the presence of arginine in the medium (30). However,we were unable to detect the direct binding of either Mcm1 orMcm1-�2-17 to these promoters by ChIP analysis, suggesting thatMcm1 may indirectly regulate the expression of these genesthrough the Mcm1 NT arm-dependent expression of a repressorprotein that targets these genes (data not shown).
The decreased transcription of YGP1 in the mcm1-�2-17mutant was not due to a decrease in Mcm1 protein level orstability. The deletion of the Mcm1 arm also did not decreaseDNA-binding affinity or change the apparent DNA-bendingangle, both of which contribute to transcriptional activation byMcm1 (1). Based on our data, we hypothesize that the Mcm1
FIG. 7. Deletion of the Mcm1 NT arm does not change the level orstability of the protein. (A) A quantitative Western blot of V5-taggedMcm1 and Mcm1-�2-17 using anti-V5 antibodies is shown. Anti-Rpa1was used to detect Rpa1 protein, a loading control. (B) Plot of quan-titative Western blot of V5-tagged Mcm1 proteins. Protein synthesiswas arrested by the addition of cycloheximide to log phase cultures,and aliquots of cells from the indicated time points were assayed forMcm1 and Rpa1 levels by Western blotting. The ratios of Mcm1:Rpa1(circles) and Mcm1�2-17:Rpa1 (squares) proteins were plotted at thedifferent time points after the addition of cycloheximide.
VOL. 4, 2005 Mcm1 REGULATES CELL WALL GENES 1815
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from
NT arm may be required to recruit or stabilize binding ofanother transcription cofactor that binds to the YGP1 pro-moter to activate its transcription.
Transcription of YGP1 was earlier shown to be regulated byRlm1, a type II MADS box protein (18). In a strain withconstitutively activated Rlm1 there was a decrease in YGP1expression, leading to the conclusion that Rlm1 was a repres-sor of this gene (18). However, mutational analysis of theregulatory sites in the YGP1 promoter suggests that Rlm1 mayalso have a minor role in the activation of YGP1. This model issupported by Northern analysis, which showed that deletion ofRLM1 resulted in a decrease in YGP1 transcript compared tothe wild-type strain (18). We have shown here that Mcm1, a
type I MADS box protein, has a direct role in regulating theexpression of this gene. Interestingly, this is the first report ofa gene that is jointly regulated by both type I and type IIMADS box proteins in yeast. Since Rlm1 is known to regulateYGP1 transcription, there is a possibility that Rlm1 may inter-act with the NT arm of Mcm1. Among the 25 genes that areknown to be regulated by Rlm1, 20 also have putative Mcm1-binding sites (18; our data). While the expression of some ofthese transcripts appear to be affected by mutations in theMcm1 MADS box domain that decrease Mcm1 DNA-bindingaffinity, we found that only YGP1 and SPS100 transcription aredependent on the NT arm (Fig. 1 and data not shown). If theMcm1 NT arm is important for interaction with its cofactor,
FIG. 8. Deletion of the Mcm1 NT arm does not change its in vitro or in vivo DNA-binding affinity for the YGP1 promoter. (A) EMSA analysisof the DNA-binding affinity of Mcm1-1-96 and Mcm1-18-96 to a 26 bp P(PAL) Mcm1 consensus probe and a YGP1 probe containing the M2 andM3 Mcm1-binding sites. The concentrations of the purified Mcm1-1-96 and Mcm1-18-96 proteins were normalized and binding by the protein(fivefold serial dilutions) is shown. (B) ChIP assays were performed on strain JM01 transformed with untagged Mcm1, Mcm1-V5, Mcm1-�2-17-V5,and Mcm1-T35A-V5 using anti-V5 antibody. Both total chromatin (TC) and immunoprecipitated (IP) DNA were assayed by PCR for the presenceof STE2 (positive control), YGP1 (test), and ACT1 (negative control) promoters.
1816 ABRAHAM AND VERSHON EUKARYOT. CELL
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from
this result suggests that Rlm1 is unlikely to be the cofactor thatMcm1 is interacting with at the YGP1 promoter. Also, sincemutagenesis of the Rlm1-binding site, R1, in the YGP1-lacZreporter showed only a minor decrease in activation (Fig. 6),there is little evidence for the role of Rlm1 as the cofactorrecruited by the Mcm1 NT arm.
Residues S2 and T8 in the Mcm1 NT arm are heavily phos-phorylated and a mutation of S2 to alanine, which partiallymimics the unphosphorylated form of these residues, had asalt-sensitive phenotype that was similar to deletion of the NTarm (24). Since the deletion of the NT arm also causes cells tobe sensitive to CFW, high pH, and CaCl2, we expected thatamino acid replacements of residues S2, T8, and T10 may havesimilar phenotypes on these media. However, none of thepoint mutations affected growth on CaCl2 and had relativelymild effects on growth on CFW and high pH in comparisonto the NT arm deletion. Interestingly, mutations of these res-idues to either alanine or aspartate, which partially mimicconstitutive dephosphorylation or phosphorylation, respec-tively, showed similar sensitivity to CFW and alkaline pH(Fig. 4A and B). This suggests that the phosphorylation state ofthese residues may have little to do with resistance to CFW andalkaline pH. Alternatively, it is also possible that the aspartatemutation does not effectively mimic the phosphorylated formof these residues.
The T35A mutation in the MADS box domain of Mcm1suppresses the growth defect of the NT arm deletion underhigh-salt conditions (23). We found that this mutation had asimilar effect on suppressing the NT arm-sensitive phenotypeson media with CFW, high pH, and CaCl2 but did not restoregrowth to the level of wild-type Mcm1. Residue T35 of Mcm1contacts the phosphate backbone of the DNA and a T35Amutation decreases Mcm1 in vitro DNA-binding affinity (1,45). As expected from the decreased DNA-binding affinity, theT35A mutation caused a significant decrease in YGP1 tran-script level. Although the T35A mutation partially suppressedthe growth defects caused by the deletion of the Mcm1 NTarm, it did not cause an increase in YGP1 expression (Fig. 5).Therefore, the ability to grow in medium with CFW, high pH,and CaCl2 is not simply due to restoring expression of YGP1.This result implies that the Mcm1 NT arm may have differentmechanisms of regulating YGP1 and other genes required forresistance to various cell wall stresses. This suggests that thereare other Mcm1 NT arm-dependent genes that are requiredfor resistance to these growth conditions. It is unclear how theT35A mutation suppresses the sensitivity to cell wall stressescaused by deletion of the NT arm. It is possible the T35Amutation prevents interaction with other Mcm1 cofactors ordecreases Mcm1 binding to weak sites in the genome, causingan increase in the level of Mcm1 that is available to interactwith cofactors at the cell wall gene promoters.
The Mcm1 point mutations in the NT arm and the MADSbox domain appear to be more sensitive to CFW than growthunder high-pH conditions, while hardly affecting sensitivity toCaCl2. One model to account for this differential sensitivitywould be that genes conferring resistance to high pH andCaCl2 have other activators in addition to Mcm1. For example,Mcm1 may be the primary activator for NT arm-dependentgenes that confer CFW resistance, causing cells with the NTarm deletion to be very sensitive to the presence of this com-
FIG. 9. Deletion of Mcm1 residues 1 to 17 does not change theapparent bending angle of YGP1 DNA. (A) A circularly permutedprobe with the R1-M2-M3 fragment, shown by a rectangle, from theYGP1 promoter was digested with BamHI, NheI, HindIII, or EcoRI toyield probes designated B, N, H, and E, respectively, so that theR1-M2-M3 fragment is located at different positions along the lengthof each probe. (B) EMSAs were performed using position-permutedfragments and purified Mcm1-1-96 and Mcm1-18-96. The bend anglefor each protein-DNA complex was calculated using the Thompsonand Landy relationship (48).
VOL. 4, 2005 Mcm1 REGULATES CELL WALL GENES 1817
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

pound. On the other hand, genes required for growth underhigh-pH conditions may be regulated by other activators inaddition to Mcm1 and are therefore less sensitive to particularMcm1 mutations.
While deletion of the NT arm made cells sensitive to CaCl2,in contrast to the CFW and high pH phenotypes, growth onCaCl2 was not sensitive to any of the point mutations in theMADS box domain, suggesting that a very different mechanismmay be at play in regulating genes that confer resistance to thisgrowth condition. It is possible that another cofactor that bindsstrongly to these promoters and may be tethering Mcm1 to theDNA mainly via interactions with its NT arm. Thus, DNA-binding defects in Mcm1 may not have significant effects onactivation of these genes as long as Mcm1 is tethered to thepromoter through interactions with this cofactor.
ACKNOWLEDGMENTS
We thank Steve Brill for the anti-Rpa1 antibodies and members ofthe Vershon laboratory for plasmids and helpful discussions.
This work was supported by the Charles and Johanna Busch Fel-lowship (Rutgers University) to D.S.A. and by a grant from the Na-tional Institutes of Health (GM49265) to A.K.V.
REFERENCES
1. Acton, T. B., J. Mead, A. M. Steiner, and A. K. Vershon. 2000. Scanningmutagenesis of Mcm1: residues required for DNA binding, DNA bending,and transcriptional activation by a MADS-box protein. Mol. Cell. Biol.20:1–11.
2. Acton, T. B., H. Zhong, and A. K. Vershon. 1997. DNA-binding specificity ofMcm1: operator mutations that alter DNA-bending and transcriptional ac-tivities by a MADS box protein. Mol. Cell. Biol. 17:1881–1889.
3. Affolter, M., J. Montagne, U. Walldorf, J. Groppe, U. Kloter, M. LaRosa,and W. J. Gehring. 1994. The Drosophila SRF homolog is expressed in asubset of tracheal cells and maps within a genomic region required fortracheal development. Development 120:743–753.
4. Amar, N., F. Messenguy, M. El Bakkoury, and E. Dubois. 2000. ArgRII, acomponent of the ArgR-Mcm1 complex involved in the control of argininemetabolism in Saccharomyces cerevisiae, is the sensor of arginine. Mol. Cell.Biol. 20:2087–2097.
5. Ausubel, F. M. 1987. Current protocols in molecular biology, vol. 1. GreenePublishing Associates and Wiley-Interscience, New York, N.Y.
6. Bruhn, L., J.-J. Hwang-Shum, and G. F. Sprague, Jr. 1992. The N-terminal 96 residues of MCM1, a regulator of cell type-specific genes inSaccharomyces cerevisiae, are sufficient for DNA binding, transcriptionactivation, and interaction with �1. Mol. Cell. Biol. 12:3563–3572.
7. Carr, E. A., J. Mead, and A. K. Vershon. 2004. Alpha1-induced DNA bend-ing is required for transcriptional activation by the Mcm1-alpha1 complex.Nucleic Acids Res. 32:2298–2305.
8. Chang, V. K., J. J. Donato, C. S. Chan, and B. K. Tye. 2004. Mcm1 promotesreplication initiation by binding specific elements at replication origins. Mol.Cell. Biol. 24:6514–6524.
9. Chang, V. K., M. J. Fitch, J. J. Donato, T. W. Christensen, A. M. Merchant,and B. K. Tye. 2003. Mcm1 binds replication origins. J. Biol. Chem. 278:6093–6100.
10. Christ, C., and B. K. Tye. 1991. Functional domains of the yeast transcrip-tion/replication factor MCM1. Genes Dev. 5:751–763.
11. Davies, B., M. Egea-Cortines, E. de Andrade Silva, H. Saedler, and H.Sommer. 1996. Multiple interactions amongst floral homeotic MADS boxproteins. EMBO J. 15:4330–4343.
12. Destruelle, M., H. Holzer, and D. J. Klionsky. 1994. Identification andcharacterization of a novel yeast gene: the YGP1 gene product is a highlyglycosylated secreted protein that is synthesized in response to nutrientlimitation. Mol. Cell. Biol. 14:2740–2754.
13. Dodou, E., and R. Treisman. 1997. The Saccharomyces cerevisiae MADS-boxtranscription factor Rlm1 is a target for the Mpk1 mitogen-activated proteinkinase pathway. Mol. Cell. Biol. 17:1848–1859.
14. El Alami, M., F. Messenguy, B. Scherens, and E. Dubois. 2003. Arg82p is abifunctional protein whose inositol polyphosphate kinase activity is essentialfor nitrogen and PHO gene expression but not for Mcm1p chaperoning inyeast. Mol. Microbiol. 49:457–468.
15. Elorza, M. V., H. Rico, and R. Sentandreu. 1983. Calcofluor white alters theassembly of chitin fibrils in Saccharomyces cerevisiae and Candida albicanscells. J. Gen. Microbiol. 129:1577–1582.
16. Fan, H. Y., Y. Hu, M. Tudor, and H. Ma. 1997. Specific interactions betweenthe K domains of AG and AGLs, members of the MADS domain family ofDNA binding proteins. Plant J. 12:999–1010.
17. Harbison, C. T., D. B. Gordon, T. I. Lee, N. J. Rinaldi, K. D. Macisaac, T. W.Danford, N. M. Hannett, J. B. Tagne, D. B. Reynolds, J. Yoo, E. G. Jennings,J. Zeitlinger, D. K. Pokholok, M. Kellis, P. A. Rolfe, K. T. Takusagawa, E. S.Lander, D. K. Gifford, E. Fraenkel, and R. A. Young. 2004. Transcriptionalregulatory code of a eukaryotic genome. Nature 431:99–104.
18. Jung, U. S., and D. E. Levin. 1999. Genome-wide analysis of gene expressionregulated by the yeast cell wall integrity signalling pathway. Mol. Microbiol.34:1049–1057.
19. Kapteyn, J. C., B. ter Riet, E. Vink, S. Blad, H. De Nobel, H. Van Den Ende,and F. M. Klis. 2001. Low external pH induces HOG1-dependent changes inthe organization of the Saccharomyces cerevisiae cell wall. Mol. Microbiol.39:469–479.
20. Keleher, C. A., C. Goutte, and A. D. Johnson. 1988. The yeast cell-type-specific repressor alpha 2 acts cooperatively with a non-cell-type-specificprotein. Cell 53:927–936.
21. Kumar, R., D. M. Reynolds, A. Shevchenko, S. D. Goldstone, and S. Dalton.2000. Forkhead transcription factors, Fkh1p and Fkh2p, collaborate withMcm1p to control transcription required for M-phase. Curr. Biol. 10:896–906.
22. Kuo, M. H., and C. D. Allis. 1999. In vivo cross-linking and immunoprecipi-tation for studying dynamic protein:DNA associations in a chromatin envi-ronment. Methods 19:425–433.
23. Kuo, M. H., and E. Grayhack. 1994. A library of yeast genomic MCM1binding sites contains genes involved in cell cycle control, cell wall andmembrane structure, and metabolism. Mol. Cell. Biol. 14:348–359.
24. Kuo, M. H., E. T. Nadeau, and E. J. Grayhack. 1997. Multiple phosphory-lated forms of the Saccharomyces cerevisiae Mcm1 protein include an isoforminduced in response to high salt concentrations. Mol. Cell. Biol. 17:819–832.
25. Lagorce, A., N. C. Hauser, D. Labourdette, C. Rodriguez, H. Martin-Yken, J.Arroyo, J. D. Hoheisel, and J. Francois. 2003. Genome-wide analysis of theresponse to cell wall mutations in the yeast Saccharomyces cerevisiae. J. Biol.Chem. 278:20345–20357.
26. Lilly, B., S. Galewsky, A. B. Firulli, R. A. Schulz, and E. N. Olson. 1994.D-MEF2: a MADS box transcription factor expressed in differentiating me-soderm and muscle cell lineages during Drosophila embryogenesis. Proc.Natl. Acad. Sci. USA 91:5662–5666.
27. Lydall, D., G. Ammerer, and K. Nasmyth. 1991. A new role for MCM1 inyeast: cell cycle regulation of SW15 transcription. Genes Dev. 5:2405–2419.
28. Mead, J., A. R. Bruning, M. K. Gill, A. M. Steiner, T. B. Acton, and A. K.Vershon. 2002. Interactions of the Mcm1 MADS box protein with cofactorsthat regulate mating in yeast. Mol. Cell. Biol. 22:4607–4621.
29. Messenguy, F., and E. Dubois. 1993. Genetic evidence for a role for MCM1in the regulation of arginine metabolism in Saccharomyces cerevisiae. Mol.Cell. Biol. 13:2586–2592.
30. Messenguy, F., and E. Dubois. 2003. Role of MADS box proteins and theircofactors in combinatorial control of gene expression and cell development.Gene 316:1–21.
31. Minty, A., and L. Kedes. 1986. Upstream regions of the human cardiac actingene that modulate its transcription in muscle cells: presence of an evolu-tionarily conserved repeated motif. Mol. Cell. Biol. 6:2125–2136.
32. Molkentin, J. D., B. L. Black, J. F. Martin, and E. N. Olson. 1996. Muta-tional analysis of the DNA binding, dimerization, and transcriptional acti-vation domains of MEF2C. Mol. Cell. Biol. 16:2627–2636.
33. Murgui, A., M. V. Elorza, and R. Sentandreu. 1985. Effect of papulacandinB and calcofluor white on the incorporation of mannoproteins in the wall ofCandida albicans blastospores. Biochim. Biophys. Acta 841:215–222.
34. Norman, C., M. Runswick, R. Pollock, and R. Treisman. 1988. Isolation andproperties of cDNA clones encoding SRF, a transcription factor that bindsto the c-fos serum response element. Cell 55:989–1003.
35. Pardo, M., L. Monteoliva, J. Pla, M. Sanchez, C. Gil, and C. Nombela. 1999.Two-dimensional analysis of proteins secreted by Saccharomyces cerevisiaeregenerating protoplasts: a novel approach to study the cell wall. Yeast15:459–472.
36. Passmore, S., G. T. Maine, R. Elble, C. Christ, and B. K. Tye. 1988. Sac-charomyces cerevisiae protein involved in plasmid maintenance is necessaryfor mating of MAT alpha cells. J. Mol. Biol. 204:593–606.
37. Primig, M., H. Winkler, and G. Ammerer. 1991. The DNA binding andoligomerization domain of MCM1 is sufficient for its interaction with otherregulatory proteins. EMBO J. 10:4209–4218.
38. Ram, A. F., A. Wolters, R. Ten Hoopen, and F. M. Klis. 1994. A newapproach for isolating cell wall mutants in Saccharomyces cerevisiae byscreening for hypersensitivity to calcofluor white. Yeast 10:1019–1030.
39. Riechmann, J. L., B. A. Krizek, and E. M. Meyerowitz. 1996. Dimerizationspecificity of Arabidopsis MADS domain homeotic proteins APETALA1,APETALA3, PISTILLATA, and AGAMOUS. Proc. Natl. Acad. Sci. USA93:4793–4798.
40. Roncero, C., and A. Duran. 1985. Effect of Calcofluor white and Congo redon fungal cell wall morphogenesis: in vivo activation of chitin polymerization.J. Bacteriol. 163:1180–1185.
1818 ABRAHAM AND VERSHON EUKARYOT. CELL
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

41. Roncero, C., M. H. Valdivieso, J. C. Ribas, and A. Duran. 1988. Effect ofCalcofluor white on chitin synthases from Saccharomyces cerevisiae. J. Bac-teriol. 170:1945–1949.
42. Roncero, C., M. H. Valdivieso, J. C. Ribas, and A. Duran. 1988. Isolation andcharacterization of Saccharomyces cerevisiae mutants resistant to Calcofluorwhite. J. Bacteriol. 170:1950–1954.
43. Sengupta, P., and B. H. Cochran. 1990. The PRE and PQ box are functionallydistinct yeast pheromone response elements. Mol. Cell. Biol. 10:6809–6812.
44. Sharrocks, A. D., and P. Shore. 1995. DNA bending in the ternary nucleo-protein complex at the c-fos promoter. Nucleic Acids Res. 23:2442–2449.
45. Sharrocks, A. D., F. von Hesler, and P. E. Shaw. 1993. The identification ofelements determining the different DNA binding specificities of the MADSbox proteins p67SRF and RSRFC4. Nucleic Acids Res. 21:215–221.
46. Sommer, H., J. P. Beltran, P. Huijser, H. Pape, W. E. Lonnig, H. Saedler,and Z. Schwarz-Sommer. 1990. Deficiens, a homeotic gene involved in thecontrol of flower morphogenesis in Antirrhinum majus: the protein showshomology to transcription factors. EMBO J. 9:605–613.
47. Tan, S., and T. J. Richmond. 1998. Crystal structure of the yeast MATalpha2/MCM1/DNA ternary complex. Nature 391:660–666.
48. Thompson, J. F., and A. Landy. 1988. Empirical estimation of protein-induced DNA bending angles: applications to lambda site-specific recombi-nation complexes. Nucleic Acids Res. 16:9687–9705.
49. Trobner, W., L. Ramirez, P. Motte, I. Hue, P. Huijser, W. E. Lonnig, H.Saedler, H. Sommer, and Z. Schwarz-Sommer. 1992. GLOBOSA: a ho-meotic gene which interacts with DEFICIENS in the control of Antirrhinumfloral organogenesis. EMBO J. 11:4693–4704.
50. Yale, J., and H. J. Bohnert. 2001. Transcript expression in Saccharomycescerevisiae at high salinity. J. Biol. Chem. 276:15996–16007.
51. Yanofsky, M. F., H. Ma, J. L. Bowman, G. N. Drews, K. A. Feldmann, andE. M. Meyerowitz. 1990. The protein encoded by the Arabidopsis homeoticgene agamous resembles transcription factors. Nature 346:35–39.
52. Yu, G., and J. S. Fassler. 1993. SPT13 (GAL11) of Saccharomyces cerevisiaenegatively regulates activity of the MCM1 transcription factor in Ty1 ele-ments. Mol. Cell. Biol. 13:63–71.
VOL. 4, 2005 Mcm1 REGULATES CELL WALL GENES 1819
on June 7, 2018 by guesthttp://ec.asm
.org/D
ownloaded from