
MICROBIAL GENETICS · Cytoplasmic inheritance, therefore, can be understood based on...
24
MICROBIAL GENETICS Extra-chromosomal inheritance Prof. Arif Ali Professor & Coordinator Biotechnology Program Jamia Millia Islamia (A Central University) Maulana Mohammad Ali Johar Marg Jamia Nagar, New Delhi - 1100 25, India E-mail: [email protected] 02-Dec-2006 (Revised 07-Aug-2007) CONTENTS Introduction Chloroplast Genome structure of chloroplast Chloroplast genetics in higher plants RFLP proof that chloroplast DNA can be maternally inherited Examples of chloroplast inheritance Non chromosomal genes in Chlamydomonas Mitochondria Structure of mitochondria Mitochondrial Genome RFLP proof that mitochondrial DNA can be maternally inherited Examples of mitochondrial inheritance Plasmids Infective particles Maternal effects Origin of cytoplasmic genetic systems Keywords Transmission, nuclear markers, cytoplasmic markers, endonucleases, isogamous, symbiotic backcross, petites, susceptible, syngamy, promiscuous DNA
Transcript of MICROBIAL GENETICS · Cytoplasmic inheritance, therefore, can be understood based on...

MICROBIAL GENETICS
Extra-chromosomal inheritance
Prof. Arif Ali Professor & Coordinator Biotechnology Program
Jamia Millia Islamia (A Central University) Maulana Mohammad Ali Johar Marg
Jamia Nagar, New Delhi - 1100 25, India E-mail: [email protected]
02-Dec-2006 (Revised 07-Aug-2007)
CONTENTS Introduction Chloroplast Genome structure of chloroplast Chloroplast genetics in higher plants RFLP proof that chloroplast DNA can be maternally inherited Examples of chloroplast inheritance Non chromosomal genes in Chlamydomonas Mitochondria Structure of mitochondria Mitochondrial Genome RFLP proof that mitochondrial DNA can be maternally inherited Examples of mitochondrial inheritance Plasmids Infective particles Maternal effects Origin of cytoplasmic genetic systems Keywords Transmission, nuclear markers, cytoplasmic markers, endonucleases, isogamous, symbiotic backcross, petites, susceptible, syngamy, promiscuous DNA

Introduction
The existence of genes as segments of nucleic acid, located in chromosomes and controlling phenotypes is known and predictable, and has been amply demonstrated on sound, observable, verifiable bases. But the firm establishment of such a chromosomal mechanism of inheritance does not necessarily preclude a role for other extra nuclear cell parts. In fact, when certain mutants demonstrated an inheritance very different from normal Mendelian inheritance patterns, geneticists were forced to look to places other than the nucleus to explain the results. Abnormal segregation patterns showing inheritance from only a single parent, uniparental inheritance, or more specially maternal inheritance, where the genotype of the female parents was preferably inherited by the progeny led to considering the cytoplasm as an additional genetic systems. Maternal inheritance has a great influence on the offspring’s gene fitness and evolution and it is involved in a wide variety of taxa. Based on the pattern of hereditary transmission, there are four observations that definitely point to the existence of cytoplasmic genetic system: (i) Because the female gamete contributes almost all of the cytoplasm to the zygote and the
male gamete contributes only a nucleus, an inheritance pattern that differs between reciprocal crosses suggest a cytoplasmic involvement. This is clearly the basis for uniparental or maternal inheritance where the progeny always resemble one parent, most commonly the female parent.
(ii) Whenever traits fail to demonstrate classical segregation patterns and deviate from standard
ratios, the conclusion is again a cytoplasm-based type of inheritance. This is particularly evident when presumptive cytoplasmic markers do not segregate along with known nuclear markers.
(iii) When the traits failed to show linkage to any known nuclear linkage groups, and assorts
independently from nuclear genes, a cytoplasmic mode of inheritance is suggested.
(iv) Many types of mutants that fit the above criteria will show segregation during mitotic division. This is very common in variegated plants that carry more than one type of plastid (chloroplast) per cell. This leads to variegation, suggesting somatic or vegetative segregation of the plastid types. In addition, it may actually be possible to either observe by microscopy or to detect by biochemical methods the actual transmission of the cytoplasmic markers associated with the organelles (plastids and mitochondria).
Cytoplasmic inheritance, therefore, can be understood based on cytoplasmically located, independent, self–replicating nucleic acids, which differ from chromosomal genes by their location within the cell, and have their own unique nucleotide sequences. The basis for identifying cytoplasmic transmission in most cases is the differential contribution of cytoplasm, and the organelles therein, of the male and female gametes. Two major organelles associated with examples of cytoplasmic inheritance are chloroplast and mitochondria. Interestingly, both share a lot of similarities with a prokaryotic organization lending support to endosymbiotic origin of these organelles from a prokaryotic progenitor.
2

Chloroplast
Chloroplasts are specialized organelles found in all higher plant cells. These organelles contain the chlorophyll hence provide green color. They are very important for the plant, because they constitute the sites of photosynthesis. These may be spherical, ovoid, or discs shaped and are commonly 4 to 6µm thick. In some algal cells there is only single chloroplast whereas in higher plants the number is much larger (usually 20-40) per cell. The chloroplast is bounded by two membranes, an outer and an inner membrane with an intermembrane space between them. The inner membrane encloses a stroma that contains several enzymes. In higher plants, the stroma contains small cylindrical structures called grana. Most chloroplasts contain 10-100 grana (Fig. 1), for example, in spinach chloroplast contains 40-60 grana, and tobacco chloroplast may have 40-80 grana. Each granum consists of disc-shaped membranous sacs called thylakoids (80-120Å across) piled on top of each other. Regions of the thylakoid membranes in contact with each other are known as the stacked regions, while regions not in contact are known as the unstacked regions. The number of thylakoids in a granum may vary from a few to more than 50. The grana are interconnected by a network of anastamosing tubules called intergrana or stroma lamellae. Single thylakoids, called stroma thylakoids are found in chloroplast. Electron dense bodies called globuli are found in the stroma. Chloroplasts thus have three different membrane types, the outer membrane the inner membrane and the thylakoid membrane. Thylakoids are formed by invagination of the inner chloroplast membrane. The outer membrane of the chloroplast is freely permeable to small molecules, but the inner chloroplast membrane is impermeable to small molecules, even to H+ ions. It, however, has translocators for a number of compounds, and for passage of substances across the membrane, both in the inward and outward direction. These substances include dicarboxylic acids, ATP and other organic phosphates. The thylakoid membrane contains all the enzymatic components required for photosynthesis. Interaction between chlorophyll, electron carriers, coupling factors and other components takes place within the thylakoid membrane. Light energy is used for splitting water to oxygen. Reduction of NADP+ and phosphorylation of ADP take place simultaneously. The resulting products NADPH and ATP, respectively, are utilized by the stroma enzymes that fix carbon dioxide, synthesize and break down starch and catalyze the oxidative pentose pathway.
Genome structure of Chloroplast
Chloroplast genomes resemble large bacterial plasmids or small chromosomes. Their size ranges from 110,000 bp to 160,000 base pairs, depending on the species. Variation rarely appears in size or structure within an individual.
3

Fig. 1: Structure of chloroplast A) Cross section through a chloroplast B) Grana and intergrana C) Section through a granum D) Details of lamellar structure E) A model for the molecular organization of a granum
The chloroplast genome was the first plant genome to be characterized. Its small size and limited number of repeat elements made it a prime candidate for characterization. Its abundance in foliar tissue made it easy to isolate. Complete chloroplast genome sequences have been reported for maize, rice, liverwort, Chlamydomonas, Euglena, and several other interesting plant species.
4

DNA could be easily isolated from chloroplast and appears in the highly super coiled form (Fig. 2). A large number of investigations have provided definitive evidence that cpDNA is a closed circular double stranded molecule. In maize, each molecule of cpDNA is about 139,00 base pairs long and there are about 50 copies of this DNA per chloroplast. Each molecule is long enough to encode approximately 140 proteins, although very few have actually been identified till now. Plastids contain their own DNA polymerases and the DNA is replicated in a semi-conservative fashion. RNA polymerase found in plastids most closely resembles RNA polymerase II of the nucleus. The RNA polymerase transcribes several messenger RNAs, unique chloroplast ribosomal RNAs, and a full set of chloroplast tRNAs. Several proteins encoded on the cpDNA are synthesized on the chloroplast ribosomes. A partial list of these proteins is given in Table 1.
Fig. 2: Electron micrographs of chloroplast DNA from green alga Acetabularia
Table 1: Chloroplast proteins known to be coded by cp DNA and synthesized in the chloroplast
Proteins Function
Ribulose biphosphate carboxylase oxygenase (RUBISCO) large subunit
Part of CO2-fixing enzyme
Subunit of coupling factor 1 Membrane-bound photosynthetic protein
Ubiquinone-binding protein Membrane-bound protein binds ubiquinone
Light-induced proteins Unknown functions – but synthesized when plant is transferred from dark to light
5

Extensive work carried out on maize cp DNA has led to the construction of a genetic map illustrating the location of many genes (Fig. 3). The most striking feature of the map is the large inverted repeat area. This inverted repeat area contains the rRNA genes and some tRNA genes in the same order but in reverse orientation. While there is much to learn, the chloroplast clearly represents a separate genetic system in the cell, but not totally independent from the nucleus.
Fig. 3: A genetic map of the circular maize chloroplast DNA
Chloroplast genetics in higher plants
We know that chloroplast of lower eukaryotes and higher plants like Zea mays contain DNA. Chloroplast genomes are relatively large (with a narrow range) usually ranging from 120kb to 210 kb except in Acetabularia (400kb). This is comparable to the size of a large bacteriophage like T4 (~110 kb). Each cell may have several chloroplasts and each chloroplast may have several circular DNA molecules, so that the chloroplast DNA (cpDNA) may make up to 14% of cellular DNA in lower eukaryotes. In higher eukaryotes, however, the cpDNA makes a small proportion of total DNA in the cell. These features of chloroplast DNA are presented in Table 2.
There is evidence that genes controlling specific traits or those controlling the synthesis of specific molecules of RNA and protein, are found on the chloroplast DNA. The genes could be located by a technique called restriction fragment analysis or restriction mapping. In this technique, using specific restriction endonucleases (enzymes which cut DNA at specific sites), DNA can be cut into fragments. Each enzyme will cut the DNA into a definite number of segments of definite sizes and the different enzymes differ in their pattern of cutting. Using
6

different restriction enzymes and through an analysis of fragment produced, sites could be mapped on chloroplast genome. The mapping of specific genes was also facilitated by hybridization of restriction fragments with RNA transcripts, produced in the cell and suspected to have originated from chloroplast genome. In this manner genes for ribosomal RNA, tRNA and some proteins were located. Ribulose bisphosphate carboxylase (rubisco) is one such important enzyme, which is abundant in green leaves. Each molecule of this enzyme has 16 polypeptides, eight small (100 amino acids long) and eight large (50 amino acids long). Although small subunit is synthesized through nuclear genes, the bigger is under the control of cpDNA. The map of maize chloroplast genome known at present is shown in Fig. 3.
Table 2: Features of chloroplast DNA (cp DNA) in lower eukaryotes and higher plants
Species cpDNA
(in kb pairs)
No. genomes per organelle
No. Organelles per cell
cp DNA
as % total DNA
Chlamydomonas reinhardii
195 70-110 1 11%
Euglena gracilis 135 40 15 3%
Higher plants 120-210 variable variable variable
RFLP proof that chloroplast DNA can be maternally inherited
To demonstrate that a trait is maternally inherited specific crosses need to be made to generate the required offspring. A number of crosses were made between cultivated tomato (L. esculentum) as a female parent and a number of wild species. Chloroplast DNA was obtained from F1 plants of these crosses and digested with different enzymes. As can be seen in Fig. 4, in each case the F1 restriction fragment pattern was identical to L. esculentum (sample 8). This is conclusive evidence that chloroplast DNA is inherited in a maternal manner. Examples of Chloroplast Inheritance
There are ample examples of cytoplasmic inheritance (Fig. 5) found in the higher plants. The chloroplast inheritance in the four O’clock plant (Mirabilis jalapa) and corn are well documented in the literature. This Mirabilis jalapa plant may exist in three forms. Normal green variegated (patches of green and or nongreen tissue), and white (no chlorophyll). Egg cells from green plants carry normal green plastids, those from white plants may have both plastid types or just one type. Pollen (which produces the sperm and rarely contains any plastids) has no effect on progeny phenotype. All that is really involved here is the type of proplastids and plastids present in the egg cytoplasm. If the plastids are defective with regard to chlorophyll synthesis, the F1 plant will be nongreen, if they are not defective, the F1 plant will be green. Variegated parents produce eggs that contain both normal and defective plastids. These eggs give rise to plants, whose cell fortuitously receive a majority of green plastids or whose cells receive larger number of white
7

plastids, which results in variegated plants. Recall that division of the cytoplasm following mitosis is not a quantitatively exact process. Result from three kind of branches used as shown in Fig. 6.
Fig. 4: RFLP analysis of chloroplast DNA Iojap trait is characterized by contrasting stripes of green and white colors on leaves. This character is controlled by gene (ij) located on chromosome 7 in corn. The dominant allele Ij is responsible for normal green plastids and its recessives allele ij for iojap (striped) character, so that IjIj is green and ijij is iojap. Female gametes from ijij can therefore, carry either only green plastids or only abnormal white plastids or both. Consequently if ijij is used as female parent and IjIj as male parent the F1 individuals (Ijij) will be green, white or stripped (Fig. 7). It is therefore, obvious that Iojap character once inherited through female parent will be able to maintain itself in the cytoplasm irrespective of genotype of plant.
8

Fig. 5: The four o’clock plant, Mirabilis jalapa, green-leaved variety
Fig. 6: Plasmid inheritance in four o’clock plant Mirabilis jalapa
Non chromosomal genes in Chlamydomonas
Although, the first non-Mendelian gene controlling streptomycin resistance was described by Ruth Sager in Chlamydomonas, as early as 1954, but its location on chloroplast genome became known only later. Several genes located on chloroplast genome were reported and a circular genetic map was proposed by Sager in 1972 in her book “Cytoplasmic Genes and Organelles”. Later she presented further results and discussed methods used in the genetic analysis of these non-Mendelian genes.
9

Fig. 7: Inheritance of iojap
(a) Uniparental inheritance, as several characters show maternal inheritance both in plants and animals. This was earlier explained to be due to greater amount of cytoplasm in egg than in sperm or male gamete. In Chlamydomonas, which is a haploid organism, sexual reproduction is isogamous conjugation, both gametes being similar in morphology and thus having equal amount of cytoplasm, Despite this lack of difference in cytoplasmic content in Chlamydomonas, some characters are transferred only from one parent exhibiting what is called uniparental inheritance (Fig. 8) rather than maternal inheritance, because in Chlamydomonas, two sexes as two mating types, i.e. mt+ and mt-, rather than female and male are described. One may notice that in diploids, the male gametes do not contribute cytoplasm (Fig. 9). This way one may also distinguish between the patterns of inheritance between non-chromosomal and chromosomal genes. The inheritance patterns for chromosomal genes in haploid and diploid organisms are shown in Fig. 9 and 10.
Fig. 8: Uniparental inheritance in a haploid organism like Chlaydomonas
10

Fig. 9: Segregation of nuclear genes in a haploid organism (1:1 ratio)
Fig. 10: Segregation of nuclear genes in a diploid organism (1:2:1 ratio)
(b) Segregation, independent assortment and gene mapping. In earlier years (before 1960), a large number of cases were known, where non-chromosomal inheritances, as evident from uniparental or maternal inheritance, were described. However, further detailed study of these traits was not possible for two reasons: First the mutant non-chromosomal genes could not be artificially produced when desired and spontaneous mutations for these genes were rare thus making their study difficult. Second, since these traits were transmitted from only one parent, standard techniques of genetic analysis could not be applied to study segregation and independent assortment as studied in chromosomal genes. Ruth Sager was successful in overcoming both these difficulties in Chlamydomonas and was thus able to study the non-chromosomal genes in this organism in some detail. Ruth Sager found that if streptomycin sensitive cells are grown in a medium containing streptomycin, several kind of induced mutations in chloroplast including streptomycin resistant mutations were produced. Streptomycin resistance gene could be found on chromosomes also and there it may be inherited in a Mendelian manner, but in some strains this character showed uniparental inheritance as shown in Fig. 11. It may be noticed that when mt+ strain was streptomycin resistant, all the progeny were streptomycin resistant, and when it was sensitive, all the progeny were sensitive. Streptomycin was used for inducing not only resistance to
11

streptomycin but also to induce mutations for several other non-chromosomal traits thus making it possible to study their inheritance.
Fig. 11: Reciprocal cross showing uniparental inheritance
The second difficulty of getting the transmission of non-chromosomal genes from only one parent was overcome when it was observed by Ruth Sager that in very rare cases streptomycin resistance and streptomycin sensitivity from parents of both mating types, mt+ and mt- could be transmitted. It was found that, when streptomycin-dependent strain (sd, which will not survive without streptomycin) was used as mt+ strain and progeny grown in a medium containing streptomycin, only sd progeny will be produced (Fig. 12A). However, if the same progeny is grown in a medium lacking streptomycin, then biparental exceptional zygotes will grow, because the uniparental sd strains will not survive in the absence of streptomycin (Fig. 12B). Similarly in the reciprocal cross, using ss sd as mt-, only ss strains will be available in the absence of streptomycin showing uniparental inheritance (Fig. 12D), but if the progeny is grown in the presence of streptomycin, then biparental zygotes will be produced (Fig. 12C). Later it was discovered that if mt plus parent is irradiated with ultraviolet (UV) irradiation prior to mating, frequency of biparental zygotes would greatly increase. In this manner, techniques for obtaining exceptional biparental zygotes could be standardized.
12

Fig. 12: Results of reciprocal crosses Mitochondria
All living cell except bacteria cyanobacteria, and mature erythrocytes contain mitochondria- small, self–replicating organelles of considerable internal structural complexity, which is the center of aerobic respiration. Mitochondria are small granular or filamentous bodies that are called the powerhouses of the cell. They are associated with cellular respiration and are the sources of energy for all metabolic functions. Filamentous mitochondrion often swells at the end giving it a club shaped appearance. Pleomorphic forms may contain swellings at both ends. The average length of the mitochondrion is 3-4 µ and the average diameter 0.5 –1.0 micron. The number of mitochondria varies in different cell types. It is however, rather constant for a particular cell type. The number of mitochondria depends upon the metabolic activity of the cells; cells with high metabolic activity have a high number, while those with low metabolic activity have a lower number. In a normal live cell there are 1000-1600 mitochondria, while renal tubules have 300-400. In sperm they are as far as 20-24. However, the size, number and intracellular volume of the mitochondria undergo variations in response to a variety of physiological and pathological stimuli. Mitochondria may move freely in some cells, carrying ATP wherever required, or may be localized in a particular region, as in muscles and in reptilian venom glands. Movement is less frequent in animals than in plants. Often the movements are rhythmical.
Structure of Mitochondria
The mitochondrion is bounded by two membranes, the outer membrane and the inner membrane. The space between the two membranes is called the outer chamber or intermembrane space. It is filled with a watery fluid, and is 40-70Å in width. The space bounded by the inner membrane is called the inner chamber or inner membrane space. The inner membrane space is filled with a matrix, which contains dense granules (300-500Ǻ), ribosome and mitochondrial DNA. The
13

granules consist of insoluble inorganic salts and are believed to be the binding sites of divalent ions like Mg++ and Ca++. In some cases they apparently contain polymers of sugars. The side of the inner membrane facing the matrix side is called the M-side while that facing the outer chamber is called the C-side. Two to six circular DNA molecules have been identified within each mitochondrion. These rings may either be in the open or in the twisted configuration. They may be present free in the matrix or may be attached to the membrane. The enzymes of the Krebs cycle are located in the matrix. The inner membrane has highly convoluted folding called cristae, which project into the inner chamber. They greatly expand surface area enhancing its ability to generate ATP. In typical liver mitochondria, for example, the area of the inner membrane is about five times that of the outer membrane. The cavity of the cristae is called the intercristae space, and is continuous with the intermembrane space (Fig. 13).
Fig. 13: Section through mitochondria
Mitochondrial Genome
As we discussed in chloroplast genome, a similar case can be made for the mitochondria. The presence of DNA in mitochondrion has been clearly established. The interesting finding about mitochondrial DNA (mtDNA) is its tremendous variation in size. The three major systems studied so far are human, yeast, and higher plant mtDNA. The human mtDNA contains 16,568 base pairs and has been completely sequenced consisting of 37 genes. Yeast mtDNA is about five times larger than human mtDNA, and maize is about five times larger than yeast mtDNA. The mtDNA encodes several proteins as well as rRNA and tRNAs. A list of mitochondrial proteins and RNAs known to be encoded by the mt DNA is found in Table 3. The human mitochondrial genome has been fully sequenced and mapped. The genome is highly compact such that every base pair is involved in coding for either an mtRNA for a protein, rRNA or a tRNA. The genes appear to be very close, with no noncoding sequences between them. The one DNA strand is rich in guanosine and is called H strand, and the other strand rich in cytosine is referred to as L strand. Furthermore, there is only one major promoter on each strand. The primary transcription product is a full–length copy of each strand. The transcript is then cleaved into the various RNA molecules. At least some of the genes contain introns. It appears that tRNA genes flank almost every major gene and are actually used as spacers between these genes (Fig. 14).
14
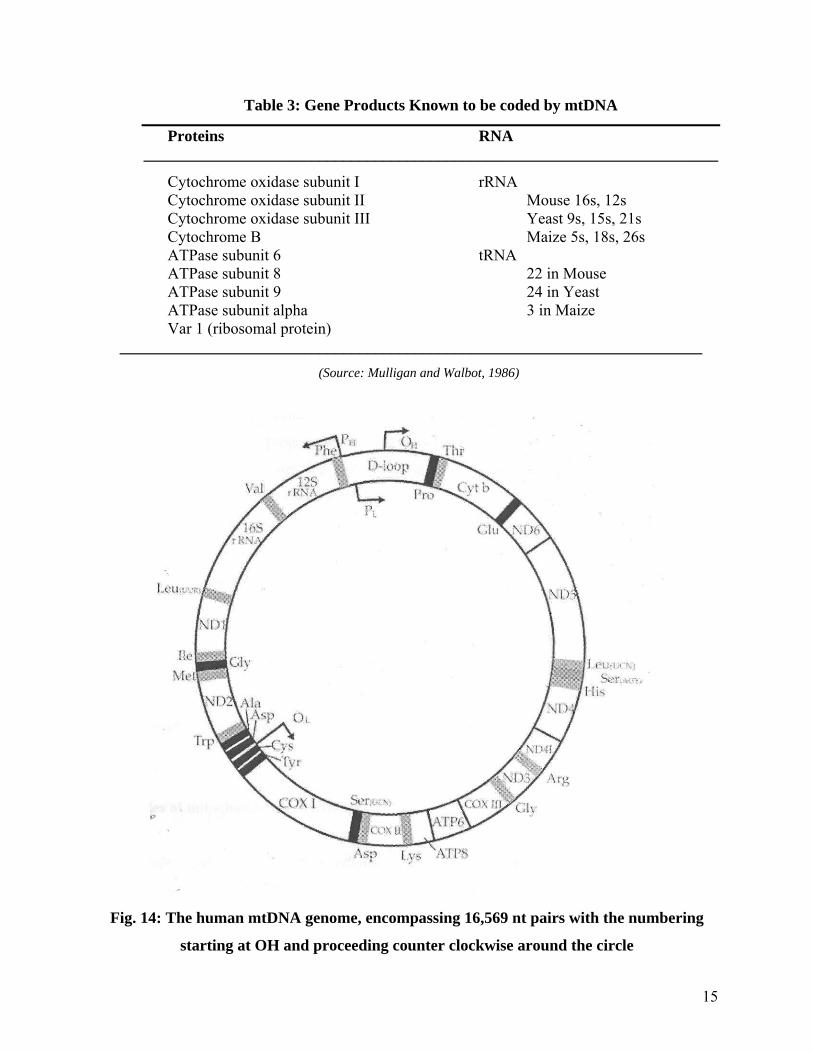
Table 3: Gene Products Known to be coded by mtDNA
Proteins RNA ________________________________________________________________________
Cytochrome oxidase subunit I rRNA Cytochrome oxidase subunit II Mouse 16s, 12s Cytochrome oxidase subunit III Yeast 9s, 15s, 21s Cytochrome B Maize 5s, 18s, 26s ATPase subunit 6 tRNA ATPase subunit 8 22 in Mouse ATPase subunit 9 24 in Yeast ATPase subunit alpha 3 in Maize Var 1 (ribosomal protein)
_________________________________________________________________________ (Source: Mulligan and Walbot, 1986)
Fig. 14: The human mtDNA genome, encompassing 16,569 nt pairs with the numbering
starting at OH and proceeding counter clockwise around the circle
15

RFLP proof that mitochondrial DNA can be maternally inherited
Experiments designed to show that mitochondrial DNA was maternally inherited were analogous to those, which demonstrated that chloroplast DNA was maternally inherited. Mouse is the experimental organism used to demonstrate this principle here. Reciprocal crosses were made between the species Mus domesticus, common mouse, and the wild species Mus spretus, (Algerian mouse). F1 progeny were then backcrossed, with one of the two species serving as females. The restriction enzyme HindII is diagnostic for the mitochondrial DNA of the two species. M. domesticus mtDNA contains five restriction sites for the enzyme that generate four fragments whereas M. spretus mtDNA contains eight restriction sites which generate seven fragments. Only two of the fragments are the same size and presumably contain the same sequence information. The fig. 15 depicts the results of the backcross experiments. As one can see the female contributes the mitochondrial genome. The information derived from human mitochondrial gene inheritance also confirmed that the females contribute the mitochondrial genome.
Fig. 15: RFLP analysis of mitochondrial DNA
(Source: Gyllestein et al 1985. J. of heredity 76:321)
Examples of mitochondrial inheritance
Poky mutant of Neurospora
Mutants with impaired mitochondrial function demonstrate a simple case of cytoplasmic inheritance. A classical example is the poky mutant of the fungus Neurospora. Poky is characterized by its very slow growth compared to the normal one. This slow growth has been
16

traced to impaired mitochondrial function related to certain cytochromes (proteins needed for oxidation of food and generation of ATP energy). Cyt a, b, c all are found in wild type, while cyt a and b are absent and cyt c is in excess in poky mutants. The pattern of inheritance of poky is strictly maternal and is illustrated in the two reciprocal crosses:
POKY (female) X NORMAL (male)
Progeny: all poky
.
NORMAL (female) X POKY (male)
Progeny: all normal
In each cross, the phenotype of the progeny is identical to the phenotype of the female parent.
Respiratory deficient strain of Saccharomyces cerevisiae
Studies of respiratory-deficient strain of baker’s yeast (Saccharomyces cerevisiae) indicate clearly the inheritance of mitochondrial genes. Yeasts are unicellular ascomycete fungi. In the life cycle of some species, diploid and haploid adults alternate. The former species reproduces by meiospores called ascospores and the later reproduces by budding. Respiratory deficient strains are able to respire only anaerobically and “on agar, produce characteristically small colonies known as petite”. Both nuclear and cytoplasmic controls could affect this trait. The petite mutant can be segregational that follow Mendelian segregation and therefore, presumably controlled by chromosomal genes or may be more often ‘vegetative’ (non-segregational) or extra-chromosomal. The genetic basis of petite character is a cytoplasmic factor p+ (rho), which may be absent or defective in petites. In segregational petite, a recessive nuclear gene causes respiratory deficiencies. Crosses between haploid petite and normal result in all normal diploid F1 progeny, but ascospores produced by the latter segegate1: 1 for petite and normal. On the other hand, another strain (neutral petite po, which completely lack p+) has defective mitochondrial DNA. More will be said about mitochondrial DNA in the next section. In this trait, the petite character fails to segregate and F1 and F2 of the cross petite x normal are all-normal; the cytoplasm containing normal mitochondria is incorporated into F1 zygotes and vegetative cells and distributed to ascospores and haploid vegetative cells of the next generation. In a third type of petite (suppressive petite p-), crosses with normal produce a highly variable fraction of petite in the progeny. It is though that suppressive petites have rapidly replicating, abnormal mitochondrial DNA. So petite may lack mitochondrial function because of either mutant nuclear DNA or mutant mitochondrial DNA, or both. Here respiratory enzymes (except for cytochromes) are under a double genetic control.
17

Cytoplasmic male sterility
An interesting example of a mitochondrially determined trait found in some higher plants is cytoplasm male sterility. A plant showing lame sterility simply does not produce functional pollen. If the trait shows maternal inheritance, it is called cytoplasmic male sterility or CMS. The best example studied so far is in maize. There are three different systems of CMS, in maize: CMS-S, CMS-C, and CMS-T. A detailed account of CMS can be found in any standard book of Genetics. Plasmids
Extrachromosomal genetic elements composed of circular, closed DNA molecules occur in many bacteria and have been the most clearly characterized in some cases such as E.coli. These are now generally and collectively termed as plasmids, although the term episome was originally introduced by Jacob and his colleagues in 1960 to designate genetic material such as the F (fertility or sex) factor which can exist either integrated into the bacterial chromosome or separately and autonomously in the cytoplasm. The term plasmid was reserved for those bits of genetic material that exist only extrachromosomally and cannot be integrated into the genome. For simplicity sake, all bacterial extrachromosoml genetic material, whether integrable or not, will be designated plasmids. We shall deal with three important classes of plasmids: F (fertility) factor, R (resistance) factor, and col (colicinogenic) factor. F factors
A lot of work has been directed to the F factor and its important function in bacterial conjugation. In a strict sense, the F factor is an episome, for it could either be integrated into the chromosome or remain autonomous. When integrated, the donor bacterial cell is designated as a high-frequency recombinant (Hfr) strain and when it exists as a separate cytoplasmic entity, the donor cell is designated F+. The latter behaves differently from the Hfr in conjugation. The fertility factor in E. coli is composed of about 1x105 base pairs. It determines that the cell in
which it resides will serve as a donor in conjugation, and by its location (that is, whether integrated or not) whether that cell will be Hfr or F+. Cells without the F factor are recipient in conjugation. Therefore, the fertility factor does fulfill the criteria outlined at the opening of this chapter for a true cytoplasmic genetic determinant. A separate chapter is devoted to the plasmids where a detailed account is provided. R factors
The R factors are plasmids. They range in size from an apparent low 1.5x104 to a high of 1x105
deoxyribonucleotide pairs. They are most often present in one to three copies per cell, although somewhat higher numbers have been reported for a few R factor. These plasmids carry genes for resistance to one or more chemicals that include many drug used to combat infection (for example, chloramphenicol, neomycin, penicillin, streptomycin, sulfonamides, and tetracyclines). Resistance to one or more (usually several) drugs is infectious in the sense that the plasmids carrying resistance markers are often transferred during conjugation.
18

R factors are composed of two parts: the resistance transfer factor (RTF) responsible for the transfer, and various drug resistance genes. The latter includes all drug-resistance factors that may be present, except the factor conferring tetracycline resistance, which appears to be in a different location. Lewin and others have described the discovery of bacterial drug resistance during treatment of cases of bacilllary dysentery (caused by species of the bacterial genus Shigella) in Japan in the late 1950s. Four drugs commonly used for treatment of this disease are chloramphenicol, streptomycin, sulfonamides, and tetracycline. Strains of the causal organism, isolated from human cases of disease, were observed to acquire resistance to one or, more commonly, to all four drugs simultaneously. Japanese investigators in the late 1950s and late 1960s established that Shigella strains with multiple drug resistance arose in the human gut by conjugation with resistance strains of E.coli. Drug resistance can be transferred only by cell-to-cell contact but may be passed both from E.coli to Shigella and from Shigella to E.coli. Intergeneric transfer of this kind is widespread in bacteria, especially in the members of the family Enterobacteriaceae, to which both E.coli and Shigella belong. Second, at least a low level of resistant E.coli cells is often present in the intestinal tract, and treatment of dysentery with any of the four drugs kills off all the susceptible E.coli and Shigella. The remaining resistants multiply and “trade” RTFs by conjugation in the gut. Colicinogenic factors
The colicinogenic (Col) factors are plasmids that carry genes for productions of colicins, which are highly specific proteins produced by some strains of intestinal bacteria. Other strains are susceptible if their walls bear specific colicin receptor sites and resistant if the receptors are absent. Colicinogenic cells are resistant to the colicins that they themselves synthesize. There are many kinds of colicins, each having their own killing action. One of the most interesting, colicin K, inhibits DNA replication, RNA transcription, and polypeptide synthesis. Many but not all of these plasmids are transferable in conjugation. They are closed, double-stranded, circular DNA molecules ranging roughly between 6,000 and 21,000 base pairs in size. Detection of plasmids
The occurrence of plasmids in a given cell can be detected in several ways. To begin with, their presence was speculated when a bacterial cell was found to carry a phenotypes/s of selective advantage such as antibiotic resistance, degradation of xenobiotic compounds etc. Subsequently methods were developed to isolate them from a cell and most convincingly, they can be photographed as an independent entity by use of an electron microscope. Also, some are “infectious,” that is, they can be transferred through conjugation and with that all the phenotypic markers that they carried. This is notably true in the case of fertility and drug resistance plasmid. Because they replicate independently of the chromosome, they may be lost from a cell (plasmid curing) resulting in the loss of phenotype associated. The inheritance of the genes carried by the plasmid will show no linkage to any markers on the bacterial chromosome.
19

Infective particles
Cytoplasmic inheritance may also be observed in the transmission of invading microorganisms or cytoplasmic particles that have definite phenotypic effects. Two classical examples have been reported from the fly, Drosophila. Carbon dioxide sensitivity in Drosophila
A certain strain of D. melanogaster shows a high degree of sensitivity to CO2. Whereas the wild type can be exposed for long periods to pure carbon dioxide without permanent damage, the sensitive strain quickly becomes uncoordinated with even brief exposure to even low concentrations. This trait is transmitted primarily, but not exclusively, through the maternal parent. Tests have disclosed that sensitivity is dependent on an infective virus-like particle, called sigma in the cytoplasm. It is normally transmitted via the eggs’ cytoplasm but occasionally through the sperms as well. Sensitivity may even be induced by injection of a cell-free extract from sensitives. Sigma contains DNA and is mutable, but it is clearly an infective “foreign” particle. Multiplication is independent of any nuclear gene, but the mechanism of its sensitizing action is unknown. Sex Ratio In Drosophila
In another strain of Drosophila, almost all males die at zygotic stage itself. Studies have disclosed this trait to be dependent on a spirochete transmitted through the cytoplasm of the female. The maintenance of the spirochete is dependent on the nuclear genes. Killer Trait in Paramecium
Some races (“killers”) of the common ciliate Paramecium aurelia produce a substance called paramecin that is lethal to other individuals (“sensitives”). Paramecin is water soluble and diffusible, and its production depends on particles called kappa in the cytoplasm. Kappa contains DNA and RNA and is mutable, but its presence is dependent on the nuclear gene K. Animals of nuclear genotype kk are unable to harbor kappa. K-individuals do not possess kappa unless and until it is introduced through a cytoplasmic bridge during conjugation (Fig.16). Non-killer k-animals may also be derived from killers by decreasing the number of kappa particles. This may be accomplished either by starving the culture or subjecting it to low temperatures or, in contrast, by causing killers to multiply more rapidly than kappa. Kappa particles can be seen with the microscope. Electron microscope reveals them to have minute amounts of cytoplasm and to be bounded by a membrane. In addition, kappa contains DNA and protein. The particles can be transferred to other ciliates by feeding. Far from being the illustration of the cytoplasmic gene, it was first surmised to be, as infectious organism that has attained a high degree of symbiosis with the host. Similarly, the mate-killer trait in Paramecium is imparted by a Mu particle that, in turn, exists only in those cells whose micronucleus contains at least one dominant allele of a nuclear gene. Mu particles too, appear to be endosymbionts.
20
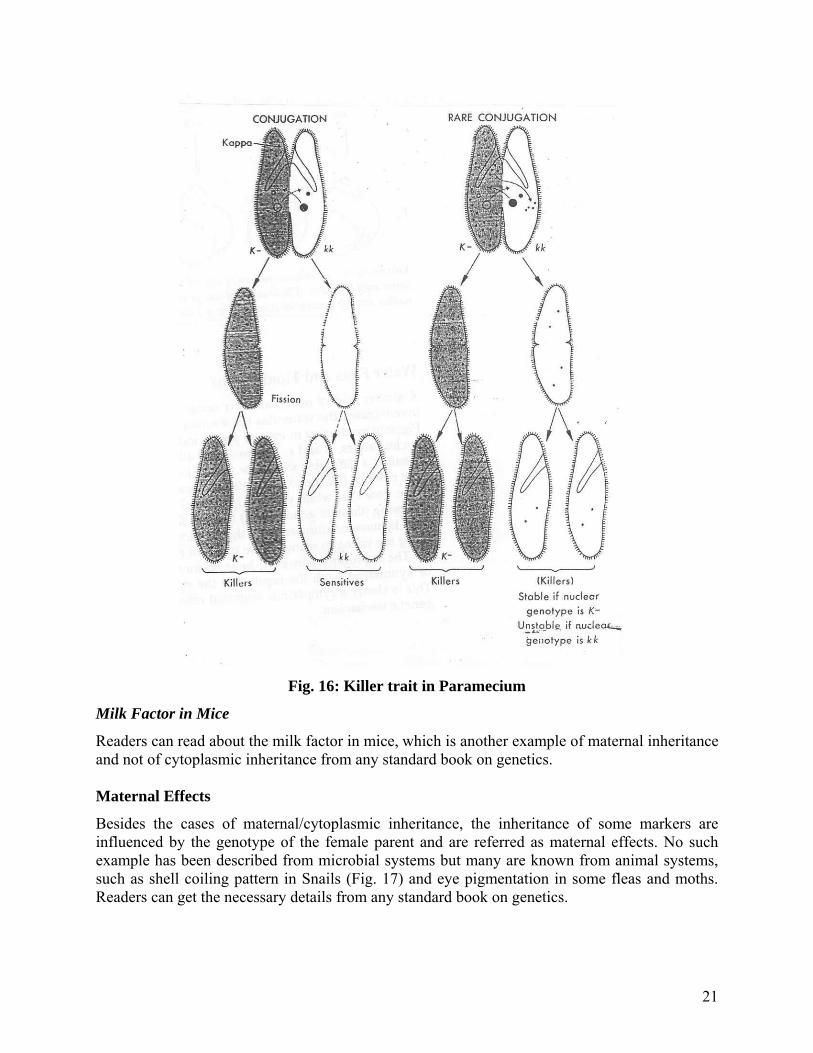
Fig. 16: Killer trait in Paramecium
Milk Factor in Mice
Readers can read about the milk factor in mice, which is another example of maternal inheritance and not of cytoplasmic inheritance from any standard book on genetics. Maternal Effects
Besides the cases of maternal/cytoplasmic inheritance, the inheritance of some markers are influenced by the genotype of the female parent and are referred as maternal effects. No such example has been described from microbial systems but many are known from animal systems, such as shell coiling pattern in Snails (Fig. 17) and eye pigmentation in some fleas and moths. Readers can get the necessary details from any standard book on genetics.
21

Fig. 17: Shell coiling in snail
Origin of cytoplasmic genetic systems
With the establishment of genetic systems in the cytoplasm, associated with the plastids and mitochondria, came the question of their origin. The currently accepted theory is the endosymbiont theory, which states that plastids and mitochondria originated with the invasion of a progenitor prokaryotic organism in a primitive eukaryotic cell. Eventually, a symbiotic relationship was established with each partner benefiting from the other. In time, they became dependent on each other and could not live separately. Certainly there is enough similarity between prokaryotes and organelles to support the theory. These similarities include: 1. Both have similar genome organization, with circular DNA, 70s ribosomes, absence of
histones, and lack of a membrane around DNA. 2. Both respond similarly to antibiotic that inhibit protein synthesis.
22

Additional support comes from the fact that these organelles code essential cellular protein not encoded by nuclear genes. This molecular cooperation indicates a symbiotic relationship for many cellular functions. For example, even though respiration occurs in the mitochondria, some respiratory enzymes are encoded by mitochondrial genes, and Ribulose bisphosphate carboxylase-oxygenase (RUBISCO) large subunit is encoded by a chloroplast gene and the small subunit is encoded by nuclear gene, even though the enzyme functions exclusively in the chloroplast. Also, when comparisons are made between organisms, transitional steps are observed. The subunit 9 protein of the mitochondrial ATPase complex is encoded by a nuclear gene in Neurospora and mammals, but by the mtDNA in yeast. This type of example has led some to propose the gene flow between organelles and the nucleus. Additional support for this gene flow hypothesis comes from the discovery of promiscuous DNA or the occurrence of the same nucleotide sequences in one or more of the three membrane bound compartments (nucleus, plastids, mitochondria) of eukaryotic cells. For example, sequences that encode the chloroplast 16s rRNA and several chloroplast tRNA also have been found in the mitochondrial DNA in maize. In yeast, the mitochondrial ATPase is encoded in the mitochondrial DNA, whereas in Neurospora the same gene is present in the nucleus; however, the mitochondrial genome of Neurospora does contain an inactive copy of the gene. This phenomenon certainly helps extend and support the conclusions of the endosymbiotic theory. Though the actual prokaryotic invader has not been identified, however, in the case of plastids, a primitive cynobacterium seems the likely candidate and purple sulfur bacteria are thought to be the progenitor of mitochondria. What appears true for now is that both types of organelles did not have a common origin. Finally, since mitochondrial DNA apparently has evolved quite rapidly, analysis of this DNA has been used to study possible evolutionary relationships. When human mtDNA was compared with four species of ape (chimpanzee, gorilla, orangutan, and gibbon), it was possible to construct in the fewest possible steps, an evolutionary tree with the five groups being derived from a common ancestor. The order of divergence was gibbon, orangutan, human, chimpanzee, and gorilla (Fig. 18). It was not possible to draw any conclusions using nuclear DNA.
Fig. 18: Evolutionary tree of mitochondrial DNA
23

In a similar study, chloroplast DNA has been used by Kung and his co-workers to form evolutionary relationships in the genus Nicotiana (tobacco). They found that by using a single restriction enzyme, SmaI they were able to identify each of the species in the genus Nicotiana. The results of this work are found in Fig. 19. This study was able to elucidate individual species relationships but also confirmed the origin of present day cultivated tobacco, Nicotiana tabaccum, as an interspecific hybrid between N. sylvestris and N. tomentosoformis. This origin had already been proposed based on other data.
Fig. 19: Phylogenetic tree for the species of Nicotiana
Suggested readings 1. Lodish et al., Molecular cell Biology, 2000. WH Freeman & Company 2. Richard Benedict & Gold Schmidt. Physiological Genetics. McGraw_Hill Book Company, 1938.
24


















