
Metabolite profiling of mutants for genes induced by ...
31
Metabolite profiling of mutants for genes induced by osmotic stress in germinated Arabidopsis seeds Author: Brend Kemperman [email protected] MSc student Plant Biotechnology Reg: 910319427030 Supervisors: Anderson Silva Wilco Ligterink Laboratory of Plant Physiology
Transcript of Metabolite profiling of mutants for genes induced by ...

MetaboliteprofilingofmutantsforgenesinducedbyosmoticstressingerminatedArabidopsisseeds
Author: Brend Kemperman
MSc student Plant Biotechnology
Reg: 910319427030
Supervisors: Anderson Silva
Wilco Ligterink
Laboratory of Plant Physiology

2
TableofContentsAbstract ............................................................................................................................................................................................. 3
Keywords ........................................................................................................................................................................................... 3
Introduction ....................................................................................................................................................................................... 4
LEA ................................................................................................................................................................................................. 4
HSP................................................................................................................................................................................................. 4
ABA ................................................................................................................................................................................................ 4
Metabolomics ................................................................................................................................................................................ 5
Mutant lines ................................................................................................................................................................................... 5
Material and methods ....................................................................................................................................................................... 8
Selection of homozygous mutant lines .......................................................................................................................................... 8
Plant material and Re‐establishment of DT ................................................................................................................................... 8
Re‐establishment of DT by treating with PEG ................................................................................................................................ 9
Metabolomics ................................................................................................................................................................................ 9
Dehydration curve; natural re‐establishment of DT ................................................................................................................... 10
Natural re‐establishment of DT ................................................................................................................................................... 10
Results ............................................................................................................................................................................................. 11
Genotyping .................................................................................................................................................................................. 11
Re‐establishment of DT by treating with and without PEG ......................................................................................................... 11
Metabolomics .............................................................................................................................................................................. 12
Natural re‐establishment of DT ................................................................................................................................................... 15
Discussion ........................................................................................................................................................................................ 16
Genotyping .................................................................................................................................................................................. 16
Re‐establishment of DT by treating with PEG .............................................................................................................................. 16
Metabolomics .............................................................................................................................................................................. 16
Natural re‐establishment of DT ................................................................................................................................................... 17
Possible future projects ................................................................................................................................................................... 18
Acknowledgements ......................................................................................................................................................................... 18
References ....................................................................................................................................................................................... 19
Appendix .......................................................................................................................................................................................... 23
A. Protocol: DNA extraction ................................................................................................................................................ 23
B. Composition of Hyponex ................................................................................................................................................. 24
C. Mutant lines .................................................................................................................................................................... 25
D. Protocol: Extraction of metabolites (polar + apolar phase) for GC‐TOF‐MS .................................................................... 26
E. Batches greenhouse ........................................................................................................................................................ 27
F. Re‐establishment of DT with natural drying .................................................................................................................... 29
G. Metabolomics graphs ...................................................................................................................................................... 30

3
AbstractDesiccation tolerance is the ability of an organism to survive a 0.1g H2O per gram of dry weight and be able to
be rehydrated without any lethal damage to the organism itself. The aim of this study is to search for a
metabolic profile in preselected Arabidopsis thaliana mutant lines which can explain desiccation tolerance.
Mutant lines that showed a greater or lower establishment of desiccation tolerance were selected. Sod1, daa1
and di19 showed a greater establishment and atvip1 a lower establishment of desiccation tolerance. GC‐TOF‐
MS analysis of these four lines showed a general trend of increase upon treatment with polyethylene glycol.
Specific plant metabolites such as valine, isoleucine and leucine which are metabolites that form a defence
mechanism against drought stress were increased. This also applies for raffinose and proline which both act as
a protectant against reactive oxygen species. Asparagine and threonine which are involved in salt tolerance are
also increased for all lines. However other metabolites such as alanine, glutamine and glutamic acid which are
expected to increase upon salt stress and tryptophan and phenylalanine which are expected to increase upon
drought stress, were either not increased for all lines, no change occurred or were even decreased in
abundance upon treatment with polyethylene glycol. In general, no clear metabolic profile has been found that
could explain desiccation tolerance for germinated A. thaliana seeds. Furthermore a general trend of the lines
upon treatment with polyethylene glycol was found and results partly correspond to previous reported results
in literature.
Keywords ABA Abscisic Acid DAP Days after pollination DW Dry Weight FW Fresh weight GC Green Cotyledon HSP Heat Shock Protein sHSP Small Heat Shock Protein LEA Late Embryogenesis Abundant PEG polyethylene glycol RH Relative humidity RH Root Hair ROS Reactive oxygen species RP Radicle Protrusion TR Testa Rupture WC Water content

4
Introduction Desiccation tolerance (DT) is the ability of a plant to be dehydrated until water content around molecules is lost
and to be rehydrated without causing lethal damage to the cells (Hoekstra, Golovina et al. 2001). DT is different
from drought tolerance. In drought tolerant conditions, cells have a minimal amount of water available to keep
specific tissues hydrated to stay alive. Water content of cells that are drought tolerant is ~0.3g of water at
minimum (H2O/g dry weight), whereas for DT it is ~0.1g (H2O/g dry weight). DT is based on the stability of
tissues to replace water with sugars, metabolites and other molecules that form hydrogen bonds, which
stabilize the proteins and membranes. During desiccation a glassy state of the cytoplasm is formed, which is a
state where molecular diffusion and chemical reactions are reduced (Hoekstra, Golovina et al. 2001). Other
macromolecules that play a role in establishing (DT) are: late embryogenesis abundant proteins (LEA’s), heat
shock proteins (HSP’s) and abscisic acid (ABA).
LEALEA’s are already studied for years and are not plant specific proteins (Hundertmark and Hincha 2008) because
they have also been found in: bacteria (Bacillus subtilis) (Stacy and Aalen 1998) and nematodes (Aphelenchus
avenae) (Browne, Tunnacliffe et al. 2002). Precise function, even when a LEA76 homologue shows that it is
involved in establishing DT, is unknown (Battista, Park et al. 2001). In A. thaliana, the ABRE (ABA responsive
element) core motif is present in the promoters of 82% of all LEA genes (Hundertmark and Hincha 2008). This
high percentage of ABREs in LEA promoters suggest that ABA is also involved in activating LEA proteins.
Furthermore LEA proteins have been shown to prevent aggregation of dehydration‐sensitive proteins for which
the LEA proteins have a peptide profile that is shared with some chaperones (Wise and Tunnacliffe 2004, Goyal,
Walton et al. 2005, Tunnacliffe and Wise 2007). A simpler explanation for the function of LEA proteins is that
they improve glass stability (Wolkers, Tetteroo et al. 1999). This function is called a ‘molecular shield’ where
the interaction between partially denatured proteins is decreased due to a physical barrier that is formed by
the LEA proteins which have a very flexible structure (Wise and Tunnacliffe 2004, Goyal, Walton et al. 2005,
Tunnacliffe and Wise 2007).
HSPHeat shock proteins (HSPs) are proteins ranging from 60 to 110 kDa in size. The HSPs family has subgroups of
small heat shock proteins (sHSPs) (14‐43 kDa), which act as molecular chaperones (Hendrick and Hartl 1993,
Sun and MacRae 2005). HSPs and sHSPs bind to a partially folded or denatured substrate of proteins and it can
promote correct substrate folding and prevent irreversible aggregation (Sun, Van Montagu et al. 2002, Sun and
MacRae 2005). HSP17.4, for instance, is expressed in the whole embryo in mature seeds (Wehmeyer and
Vierling 2000), which might indicate that HSPs have an important role in protecting the whole embryo during a
period of dehydration. Furthermore, abscisic acid (ABA) can induce the expression of HaHSP17.9‐CII in water
stressed sunflowers (Coca, Almoguera et al. 1994, Coca, Almoguera et al. 1996). To strengthen this argument,
in desiccation tolerant callus of the resurrection plant Craterostigma plantagineum treated with ABA, cytosolic
sHSPs were found which are homologous to HaHSP17.9‐CII (Alamillo, Almoguera et al. 1995).
ABAABA is involved in multiple pathways that are involved in establishment of DT in seeds. It is involved in
enhancing transcription of HSP and LEA proteins, however function or mechanisms that LEA proteins and HSPs
follow, are still unknown (Coca, Almoguera et al. 1994, Alamillo, Almoguera et al. 1995, Coca, Almoguera et al.
1996, Hundertmark and Hincha 2008). ABA itself is needed to develop DT in seeds and it inhibits germination of
precocious seeds (Meurs, Basra et al. 1992). It was found that ABA is involved in establishing DT in the seeds 13
days after pollination (DAP) (Koornneef, Hanhart et al. 1989). ABA sensitivity correlates with the ability to re‐
establish DT during germination in A. thaliana and reduced re‐establishment of DT is observed for ABA‐
insensitive and ABA–deficient mutants (Maia, Dekkers et al. 2014). ABA therefore, is an important hormone
involved in (re) establishment of DT.

5
MetabolomicsAmong responses of plants to stress due to a change in environmental conditions, is a change in the
accumulation of sugars, amino acids, betaines, organic acids and other osmolytes (compounds affecting
osmosis) in the cytoplasm. Most amino acids show a linear increase when drought stress is induced and a
decline of amino acids is observed upon rehydration (Delauney and Verma 1993). Branched chain amino acids
(BCAA’s) like leucine, isoleucine and valine are amino acids that are involved in plant defence mechanisms
against abiotic stresses and are actively involved in osmotic, drought and salt stress tolerance (Nambara,
Kawaide et al. 1998). Free amino acids as leucine, isoleucine, proline, valine and phenylalanine are significantly
increased in leaves, flowers and siliques in A. thaliana during drought stress (Nambara, Kawaide et al. 1998)
similar results were found in (Joshi, Joung et al. 2010). Continuing, a recent study showed that a feedback‐
insensitive form of a key enzyme of methionine synthesis resulted in an increase of BCAA’s, sugars, proteins,
starch and other amino acids, which resulted, in an increased germination rate of the transgenic seeds under
salt and osmotic stresses. This also occurred when ethylene or abscisic acid was added to A. thaliana seeds
(Cohen, Israeli et al. 2014).
Sugars also play a role in establishing DT. It has been found in soybean (Glycine max L. Merr. Cv Williams) corn
(Zea mays L. cv Merit) and pea (Pisum sativum L. cv Alaska) that oligosaccharides are present in the seeds
during DT induction and DT is lost when the concentration of oligosaccharides are decreasing in the seeds
(Koster and Leopold 1988). It also has been found that increases of stachyose, raffinose and sucrose in soybean
(Glycine max L. Merril cv Chippewa 64) and red oak (Quercus rubra L.) seeds is correlated with establishment of
DT (Blackman, Obendorf et al. 1992, Sun, Irving et al. 1994). A glassy state, or formation of intracellular glass
found in corn seeds, occurs when the water content in the seeds is decreased and could help to protect the
embryos from damage caused by DT (Williams and Leopold 1989, Koster 1991). The glassy state is a liquid
solution with a viscosity of a solid present in the cells of the seed that consists of a high concentration of
solutes, for instance soluble sugars (Williams and Leopold 1989, Koster 1991). The glassy state is lost again
upon imbibition on which biochemical activity is resumed in the seed (Williams and Leopold 1989). Other
compounds such as proline, glutamate, glycine‐betaine, carnitine, mannitol, sorbitol, fructans, polyols,
trehalose and sucrose are also helping to keep proteins from denaturation and membranes from fusing
together(Hoekstra, Golovina et al. 2001).
Mutantlines
To investigate the role of genes induced by osmotic stress,
re‐establishment of DT and metabolite profiling were
performed in germinated A. thaliana seeds. Eight mutants
with T‐DNA insertions were tested (atcc, at1g614304, daa1,
asn1, di19, at1g07728, atvip1 and jmjd5) which showed
greater expression in re‐establishment of DT. ATCC is
responsible for CuZnSOD activity (Chu, Lee et al. 2005).
CuZnSOD, or copper/zinc superoxide dismutase, found in
the chloroplast is an enzyme that is responsible for
catalysing reactive oxygen species (ROS) (fig 2). These ROS
include superoxide, hydrogen peroxide and hydroxyl
radicals. These molecules are generated as a by‐product
during metabolism in cells and they might cause damage to
the organism itself (Imlay and Linn 1988). There are two
other SOD systems found in plants, namely manganese SOD
(MnSOD) and iron SOD (FeSOD) which can catalyse the ROS
(Bowler, Van Camp et al. 1994, Zelko, Mariani et al. 2002,
Gao, Ren et al. 2003). Furthermore, other molecules as the
organic compound ascorbate acid (AsA) which is oxidized
Figure 2 image taken from Allen (1995), which describesthe dismutation/disproportionation of superoxideradicals (O2
‐) by SOD associated with PSI. The H2O2 is thencatalysed by tAPX and sAPX into harmless H2O. Themonodehydroascorbate radicals (MDA) produced by theAPX are converted, with reactions with Fd ormomodehydroascorbate reductase (MDHAR), to ascorbicacid (AsA). Dehydroascorbate reductase (DHAR) iscatalyzing the DHA to AsA.

6
form dehydroascorbic acid (DHA), ascorbate peroxidase (APX) and glutathione reductase (GR) are active in
chloroplasts. In the cytosol there are also hydrogen peroxide scavenging enzymes present, which help to
protect the cells from damage caused by hydrogen peroxide and hydroxyl radicals (Smith, Vierheller et al. 1989,
Asada 1992, Smirnoff and Wheeler 2000). No expression of ATCC is observed during germination and it is mildly
down regulated during abiotic stress in the root and down regulated during osmotic and salt stress in the shoot
(Table 1).
At1g614304 is a S‐Locus lectin protein kinase family protein involved in protein amino acid phosphorylation.
Not much is known about this protein family. It is expressed in the root during osmotic, salt and drought stress,
but not during germination or abiotic stress in the shoot (Table 1).
DAA1 an ATPase protein that is activated by the R2R3 MYB (a gene family in A. thaliana) (Stracke, Werber et al.
2001) transcription factor DUO1. Up regulation is observed in 14 day old seedlings upon drought, cold, heat
and salt stress. Also in syncytia At1g64110 is upregulated (Ali, Plattner et al. 2013). Furthermore, this gene is
induced in germ cells by DUO1 for differentiation (Borg, Brownfield et al. 2011). The gene is down regulated
during germination but highly upregulated in the root during osmotic stress and in the shoot during both
osmotic and salt stress (Table 1).
ASN1 transcribes a glutamine‐dependent asparagine synthase 1, which is one of three ASN genes in A. thaliana.
It is predominantly expressed in shoot tissues and asparagine plays an important role in transport and storage
of assimilated nitrogen (Lam, Peng et al. 1994, Lam, Hsieh et al. 1998). Asparagine itself can enhance the
tolerance of seedlings and plants, at least when added exogenous, to NaCl (Lehle, Chen et al. 1992). The gene is
down regulated during germination but highly upregulated during osmotic and salt stress in the root, whereas
in the shoot it is down regulated during salt and osmotic stress (Table 1).
DI19 encodes the drought induced 19 protein which acts as a transcriptional regulator. DI19 regulates several
pathogenesis related (PR) genes (PR1, PR2 and PR5) which are involved in plant responses to drought stress.
DI19 is regulated by CPK11 which acts as a positive upstream regulator (Liu, Zhang et al. 2013) and CPK11 itself
is again regulated by ABA (Zhu, Yu et al. 2007). DI19 is down regulated during germination and osmotic stress in
the root. It is however upregulated during both osmotic and drought stress in the shoot and mildly upregulated
during salt stress in the shoot (Table 1).
AtG07728 transcribes a potential natural antisense gene, and the locus overlaps with At1G07725 which is a
protein that is a member of the EXO70 gene family which is important for cell and organ differentiation (Synek,
Schlager et al. 2006). No expression data was found in the Arabidopsis eFP
Browser (Table 1).
AtVIP1 transcribes a myo‐inositol hexakisphosphate kinase (AtVIP1) that,
when overexpressed, shows hypersensitivity to ABA at seed germination
and seedling stages is observed and significantly enhanced drought
tolerance in transgenic plants (see fig 3). AtVIP1 is a regulator of the
transcription factor AtMYB44(Xu, Chen et al. 2015). When AtMYB44 is
overexpressed, an increased sensitivity to ABA for seeds germination and
enhanced drought tolerance is observed. This drought tolerance is
probably obtained by stomatal closure (Jung, Seo et al. 2008). Expression
of the gene is down regulated during germination and no up or down
regulation of the gene is observed during abiotic stress in both root and
shoot (Table 1).
At3g20810 transcribes JMJD5 and JMJ30 which are jumonji domain
containing 5 and jumonji c domain‐containing protein 30 respectively,
Figure 3: Schematic drawing from Xu, Chen et al. (2015). VIP1 is affected by MPK6 which upregulates MYB44 which increases drought tolerance and sensitivity to ABA.

7
which are histone demethylases that are involved in regulating chromatin structure and gene expression.
JMJD5 is co‐regulated with evening‐phased clock components and it affects the expression of clock genes that
are induced at dawn. JMJ30 is a protein that regulates the circadian clock in association with the central
oscillator and it is repressed when it is bound with CIRCADIAN CLOCK ASSOCIATED1 and LATE ELONGATED
HYPOCOTYL, which are suggested to form a feedback loop in the circadian clock. (Jones, Covington et al. 2010,
Lu, Knowles et al. 2011). As mutant line atvip1, expression of the gene is down regulated during germination
and no differential regulation of the gene is observed during abiotic stress in both root and shoot (Table 1).
The objective was to study genes that play a role in re‐establishment of DT upon polyethylene glycol (PEG)
treatment. Re‐establishment of DT was tested on four developmental stages of germinated A. thaliana seeds:
when seeds where at testa rupture (TR), radicle protrude (RP), first appearance of root hairs (RH) and when
cotyledons turned green (GC). Metabolomics
analysis of selected mutant lines atcc, daa1, di19
and atvip1 for the RH stage was performed to
search for a metabolic profile with a correlation
with DT. Mutant lines showed no clear metabolic
profile that could explain desiccation tolerance
for germinated A. thaliana seeds. A general trend
of the lines upon treatment with PEG was found.
Figure 1 Developmental stages of germinating Arabidopsisseeds; Testa Rupture (TR), Radical Protrusion (RP), Root Hair (RH),Cotyledons Greening (GC)

8
Materialandmethods
SelectionofhomozygousmutantlinesOne leaf of the rosette or flower of each plant was used for DNA extraction. The DNA extraction protocol can
be found in Appendix A. Primers were designed (http://signal.salk.edu/tdnaprimers.2.html) to identify the
insertion in each line. Forward‐ plus reverse primer master mix and forward‐ ,reverse plus border (SALK or SAIL)
primer master mix were used to check location of the insertions. The first combination was used to check if the
WT allele was present in that specific line, whereas the second combination was to check if the SALK or SAIL T‐
DNA insertion was present. Each A. thaliana plant that showed only one amplicon smaller than the wild type
amplicon was considered as being homozygous plant. Heterozygous plants would show two bands, one for the
wild type and one band for the inserted T‐DNA. Heterozygous plants were discarded and a second check of the
selected homozygous plants were performed by checking it on a gel with new extracted DNA from a leaf or
flower. When homozygosis was again confirmed, the selected plants were grown until the seeds were fully
matured.
PlantmaterialandRe‐establishmentofDTMutant line were growth in the greenhouse on a table that has a flood feeding function. Relative humidity (RH)
in the greenhouse was set at 80% with 16/8 hour light/dark controlled conditions. All plants received Hyponex
(Appendix B) as nutrient solution. Each homozygote plant was harvested separately and the seeds were stored
at room temperature. Eight homozygous mutants were selected (Table 1). Each of these eight mutant lines was
tested for their ability to re‐establish DT upon polyethylene glycol (PEG) treatment. Other mutant lines of
different genes also induced by osmotic stress were selected as being homozygous (Appendix C), but due to
lack of plant material for these lines, they have not been used for experiments during this thesis.

9
Table 1: Mutant lines used in this thesis *Gene expression after 6‐24 hours of imbibition in water. +: increase, ‐: decrease, O: No increase or decrease in gene expression. ND: No Data. **Expression of abiotic stress, osmotic( 300mM Mannitol), salt (150mM NaCl) and drought stress (plants were exposed for 15 min with loss of app.10% fresh weight) in both root and shoot. O: No difference compared to the wild type (WT). ‐/O: 25‐50% negative difference compared to the WT. O/+:25‐50% positive difference compared to the WT. +: 50‐500% positive difference compared to the WT. ++: 500‐1000%: positive difference compared to the WT. +++:1000+ % positive difference compared to the WT. If expression is below a factor 100 a O (no change) will be noted. The ‐, ‐‐ and ‐‐‐ correspond to the same percentage as +, ++ and +++, but as a decrease in expression. Data obtained from: http://bar.utoronto.ca/efp/cgi‐bin/efpWeb.cgi
Abbreviation ID code Function Expression during germination*
Expression of abiotic stress in the root**
Expression of abiotic stress in the shoot**
Osmotic Salt Drought Osmotic Salt Drought
ATCC At1g12520 Copper Chaperone for SOD1
O ‐/O ‐/O ‐/ O ‐ ‐ ‐/O
At1g61430 At1g61430
Protein phosphorylation (S‐Locus lectin protein kinase family protein)
O ‐/O ‐ ‐ O O O
DAA1 At1g64110 Gene coding for an ATPase
‐ +++ O O +++ +++ O
ASN1 At3g47340
Glutamine‐dependent asparagine synthetase1
‐ +++ +++ O ‐ ‐‐‐ O
DI19 At1g56280 Drought induced gene family
‐ ‐/O O O + O/+ +
At1g07728 At1g07728 Putativeexocytosis
ND ND ND ND ND ND ND
ATVIP1 At3g01310 Phosphate metabolism
‐ O O O O O O
JMJD5 At3g20810 Reverse histone methylation
‐ O O O O O O
Re‐establishmentofDTbytreatingwithPEGA duplicate experiment of four replicas of 30 seeds each in the stages RP and RH for the mutants with T‐DNA
insertion; atcc, at1g614304, daa1, asn1, di19, at1g07728, atvip1, jmjd5 and a wild type (Col‐0) were incubated
for 3d in a 1ml polyethylene glycol (PEG 8000) solution with an osmotic potential of ‐2.5 MPa on one layer of
filter paper in a 6‐cm petri dish as described (Maia, Dekkers et al. 2011). After incubation, seeds were washed
with distilled water to remove the remaining PEG and dried for 3d above a saturated CaCl2 solution on a new
white filter paper inside a desiccator (42% RH at 20OC) (Maia, Dekkers et al. 2014). After drying, seeds were first
pre‐humidified (relative humidity of 100%) for 24H at 220C under constant white light and further rehydrated
on a Copenhagen table. Viability of the seeds was checked after 7 days. Seeds that continued their
developmental program and turned into a seedling were considered as being DT tolerant.
MetabolomicsMetabolites were measured of seeds at RH stage, treated in PEG for 3d. Four biological replicates for the
mutant lines atcc, daa1, di19 atvip1 and wild type in the RH stage were used with 200 seeds for each biological
replicate and treatment. Each line was treated with and without 1ml ‐2.5MPa polyethylene glycol (PEG 8000)
solution. Metabolites were extracted from the seeds according to the Extraction protocol (polar + apolar phase)
for GC‐TOF‐MS analysis (Appendix D). Samples were put in the GC‐TOF‐MS and obtained data was analyzed
using MetaboAnalyst, a web‐based platform for comprehensive analysis of quantitative metabolomics data (Xia
and Wishart 2011).

10
Dehydrationcurve;naturalre‐establishmentofDTTo access a more natural way to re‐establish DT in germinated seeds, four developmental stages were used to
induce DT without PEG. Four developmental stages are: testa rupture (TR), radical protrusion (RP), first
appearance of root hair (RH) and green cotyledons (GC). These stages occur after approximately 24, 28, 32 and
36 hours respectively, germinating in a 220C light chamber after stratification of at least 72h in a 4oC dark
chamber. A dehydration curve was measured, for each developmental stage, during 72 hours in nine time
points (0, 2, 4, 6, 8, 10, 24, 48 and 72 hours). Four replicas of Col‐0 with 36 seeds for each developmental stage
and time point were selected and dried in a dark 22oC cabinet with a RH of 32% on a black membrane in a seed
tray containing one blue filters and 25ml of demi‐water to represent a natural drying. The fresh weight (FW) of
the seeds after drying for the different durations was measured. The seeds were thereafter placed in a 105oC
oven to fully dry overnight. The seed dry weight (DW) was measured and the WC (water content) (g g‐1 dw) was
calculated and put in a graph.
Naturalre‐establishmentofDTA duplicate experiment of four replicas of 30 seeds each in the stages RP and RH for the mutant lines atcc,
at1g614304, daa1, asn1, di19, at1g07728, atvip1, jmjd5 and wild type (Col‐0) were selected, put on one black
membrane and incubated in a seed tray with one blue filter and 25 ml of water. After drying, seeds were first
pre‐humidified (100% RH) for 24H at 220C (Maia, Dekkers et al. 2014) in the light and subsequently further
rehydrated on a Copenhagen table. Viability of the seeds was checked after 7 days.

11
Results
GenotypingPlants were genotyped individually. Mutant lines At2G01100, At5G58720, At3G52180, NAKR3 and NTMC2T6.2
showed a homozygous genotype (mutant lines correspond to the numbers 6, 10, 22, 23 and 34 in appendix E).
Other mutant lines showed heterozygous or wild type alleles. A few siliques of each mutant line were
harvested and seeds were checked under a microscope in search for missing seeds that indicates a lethal allele,
this was not found. Furthermore, mutant line 9, even after multiple attempts, did not germinate in the
greenhouse.
Re‐establishmentofDTbytreatingwithandwithoutPEGAs described in Maia, Dekkers et al. (2011), DT can be re‐established by treating the seeds using a PEG (‐
2.5MPa) solution. This experiment was a follow‐up of an experiment performed by the BSc student Josephine
van Eggelen. Only mutant line atvip1 showed a significant decrease in re‐establishing DT for the RP stage
(Appendix F). Whereas when the same mutants were treated with PEG (‐2.5 MPa ) solution to re‐establish DT in
germinated seeds, each mutant was able to re‐establish DT at RP.
It was observed that the mutants atcc, daa1 and di19 show a greater DT compared with Col‐0, whereas mutant
line atvip1 just as mutant At1g07728 showed no differential re‐establishment of DT. The same experiment with
the same conditions was performed again and mutant line atvip1 did show a significant lower re‐establishment
of DT. Mutant lines atcc, daa1, di19 and atvip1 were selected for metabolomics analysis.
Figure 4: Survival of ‐2.5 MPa PEG treated mutant lines after a 72H dehydration,duplicate experiment of 4 replicates per line, 36 seeds per replicate.

12
MetabolomicsMutant lines atcc, daa1, di19 and atvip1 were investigated further by performing a metabolomics analysis. A
total of 50 different metabolites were measured. Figure 5 shows the strongest increase and decrease upon
treatment in seeds treated with PEG. Xylose and 4‐hydroproline show both a significant decrease, whereas
raffinose shows an increase. In table 2 a summary of each metabolite and the general response of the PEG
treated mutant lines compared to the untreated mutant lines is shown.
Figure 5. Metabolites of the Mutant lines without PEG treatment (coloured columns) and with PEG (‐2.5 MPa) treatment (striped columns), which show the highest change between with and without treatment.
As shown in table 2, which summarized a comparison of mutant seeds treated with PEG (‐2.5 MPa) solution
compared to their own non‐treated seeds. The abundance of 16 metabolites for the mutant seeds are
increased, (asparagine, aspartic acid, benzoic acid, beta‐D‐methylfrutofuranoside, isoleucine, lysine, phophoric
acid, proline, pyroglutamic acid, raffinose, serine, s‐methyl‐Lcysteine, sucrose, threitol, threonine and valine). 5
metabolites have no significant change (citric acid, methionine, phenylalanine, tryptophan, and tyrosine). 10
metabolites, are decreased (4‐hydroxyproline, alfa‐ketoglutaric acid, ethanolamine, fructose, galactinol,
glyceric acid, malic acid, myo inositol succinic acid and xylose). 9 metabolites showed no significant increase,
but for some mutants these metabolites were upregulated (alanine, ascorbic acid GABA, glutamine, glycine,
glycolic acid, leucine, monomethylphosphate and nonanoic acid). The last 10 metabolites showed no significant
increase in the WT, but for some mutants these metabolites were down regulated. (urea, beta alanine, gumaric
acid, glucopyranose, glucose, glucose‐6‐phosphate, glutamic acid, N‐acetylglutamic acid, putrescine, and
pyruvic acid) Some lines showed a significant difference for some metabolites in a treatment compared to the
wild type (Col‐0) for the same treatment (see Appendix G). Mutant line atcc showed an increase of
monomethylphosphate for the untreated seeds. Mutant line daa1 showed a decrease in both urea and
putrescine for the treated seeds and for the untreated seeds an increase in glucose . Mutant line di19 showed
for treated seeds an increase in threitol and a decrease in myo inositol. The untreated seeds for this line
showed only an increase in glucose. Most significant changes were found for mutant line atvip1. The treated
seeds show an increase in sucrose and an decrease for GABA, putrescine and myo inositol. Untreated seeds
showed an increase in abundance in phosphoric acid and glutamine. A decrease in abundance, for untreated
seeds, was found for in putrescine, beta‐D‐methylfructofuranoside, beta‐alanine, 4‐hydroxyproline and myso
inositol.
Metabolites, that are important or are involved in establishing DT are shown in figure 6 (CHIANG and Dandekar
1995, Nambara, Kawaide et al. 1998, Joshi, Joung et al. 2010). The metabolites valine, isoleucine, proline, lysine,
asparagine and threonine are all increased for all mutant lines upon treatment with PEG (‐2.5 MPa) solution. All
mutant lines have increased levels for leucine, except atvip1 which does not show an increase or decrease.
Tryptophan is increased for mutant line atvip1 and not increased or decreased for the other mutant lines.
Alanine shows for both daa1 and atvip1 no increase or decrease, but for sod1 and di19 levels are higher.
Glutamine is not increased in mutant line sod1, while the other mutant lines show a higher level. Finaly,
glutamic acid shows a decrease for sod1 and atvip1 but this is not found for the other mutant lines.

13
Figure 6: Measured metabolites that are important or involved in establishing DT or are found to be increased upon drought or salinity stress. Mutant lines without PEG treatment (coloured columns) and with PEG (‐2.5 MPa) treatment (striped columns).

14
Table 2: An schematic overview of 50 measured metabolites which show the trend that the mutant lines atcc, daa1, di19 and atvip1 follow when they are treated with PEG. A + indicates that all 4 lines show a significant increase for that particular metabolite when they are treated with a ‐2.5 MPa PEG solution compared to their own non treated mutant wild type. A – indicates a decrease and a O indicates no significant increase was found. The combination of ‐/O and O/+ indicates that both an increase or decrease together with no significant change was found for that particular metabolite.
Metabolite Metabolite Metabolite Metabolite Metabolite
Asparagine + Serine + Glycine 3tms O/+ Urea ‐/O 4‐Hydroxyproline
‐
Aspartic Acid + S‐Methyl‐L‐cysteine
+ Glycolic acid O/+ Beta alanine ‐/O Alfa‐Ketoglutaric Acid
‐
Benzoic Acid + Sucrose + Leucine 2tms O/+ Fumaric acid ‐/O Ethanolamine
‐
Beta‐D‐Methylfructofuranoside
+ Threitol + Monomethylphosphate 2tms
O/+ Glucopyranose
‐/O Fructose ‐
Isoleucine + Threonine 3tms
+ Nonanoic acid O/+ Glucose ‐/O Galactinol ‐
Lysine + Valine + Citric acid O Glucose‐6‐phosphate
‐/O Glyceric Acid ‐
Phosphoric Acid
+ Alanine O/+ Methionine O Glutamic acid
‐/O Malic Acid ‐
Proline + Ascorbic Acid
O/+ Phenylalanine O N‐acetylglutamic acid
‐/O Myo Inositol ‐
Pyroglutamic acid
+ GABA O/+ Tryptophan O Putrescine ‐/O Succinic Acid ‐
Raffinose + Glutamine O/+ Tyrosine 3tms O Pyruvic Acid ‐/O Xylose ‐

15
Naturalre‐establishmentofDTTo perform a more natural way of re‐establishment of DT, a dehydration curve has been made for each stage:
TR, RP, RH and GC. Dehydration curves are shown in figure 7 .
Figure 7: Dehydration curve of the different stages TR, RP, RH and GC altogether and each of them separately. Inside the combined dehydration curve, another curve is shown that represents the survival rate of the different developmental stages when dried in natural conditions. TR and RP stage survival ration is 100% upon re‐hydration whereas for the RH a 20% and GC stage a 0% survival rate is observed. All stages have reached the same water content after 72H of drying.
The dehydration curves show a similar decrease of moisture during a drying period of 72H in a 220C 32% RH
environment for all developmental stages. Within 6h all stages, except TR, have lost more than 90% of their
moisture. After 10h the slope of all the stages has stabilized. The stages reached the same water content after
72H of drying and only TR and RP have a 100% survival rate upon re‐hydration. The stages lost 90 to 96 % of
their total weight after 72H of drying.

16
Discussion
GenotypingMost of the mutant lines showed heterozygosity. Even after multiple attempts and several batches, no
homozygous genotype was found for these lines but only heterozygous or wild type genotypes. It is a strange
occurrence that out of 25 plants and for the second batch which was grown out of heterozygous seeds, not a
single homozygous plant was found when mendel’s law of segregation suggest that 25% of the 25 plants should
be homozygous. This raised the question if, for at least one line, a lethal allele could exist. This was not the case
when the siliques were checked; all seeds were present for all lines.
For mutant line 9, the knocked‐out gene encodes a RING‐finger type E3 ligase. This ligase is necessary for
mitotic cell cycle progression during gametogenesis in A. thaliana (Liu, Zhang et al. 2008). This could explain
why none of the seeds germinated in the greenhouse.
Re‐establishmentofDTbytreatingwithPEGLines that showed an increased or decreased survival rate with PEG at RH, were chosen to continue research by
performing a MS‐TOF‐GC analysis. Mutant line atcc, daa1, di19 and atvip1 were chosen to investigate more
thoroughly. Mutant lines asn1 and Atg07728 were not chosen, since they did not show a significant difference
when compared to col‐0. Mutant line atvip1 was chosen, due to a significant score in the second experiment
and due to the fact that it showed a lower establishment of DT. It was preferred to use the phenotypes that
showed greater or lower re‐establishment of DT, so a possibility to check if a specific metabolic profile is
present that could explain the re‐establishment of DT in the seeds.
MetabolomicsTo check which metabolites would have a role in re‐establishment of DT in A. thaliana germinated seeds,
mutants with greater re‐establishment at RH (atcc, daa1 and di19) and lower (atvip1) were used for metabolite
analysis. Change in valine, leucine and isoleucine are observed upon treatment with PEG (fig 6). These free
amino acids are responsible to act as a defensive mechanism against abiotic stresses, in this case drought stress
(Nambara, Kawaide et al. 1998, Joshi, Joung et al. 2010). This is seen in the results except for mutant line atvip1,
which does not show an increase in leucine. In plant material these three amino acids, together with
phenylalanine, proline, tryptophan and lycine (fig 6), are increased several fold during dehydration (Nambara,
Kawaide et al. 1998). This trend is also seen for threonine, asparagine, glutamate, glutamine, proline and
alanine at increasing NaCl concentrations with 2 week old plant material (fig 6) (CHIANG and Dandekar 1995).
Proline, lycene, asparagine and threonine are increased for all treated seeds. In A. thaliana seeds proline can
enclose 17% of the total free amino acids. The same accounts for asparagine, which encloses 22%. However in
tissues with a higher water content, a lower level of proline and asparagine is found (CHIANG and Dandekar
1995). In general proline acts as a protectant and can scavenge ROS. In many species it can be activated by
ABA (KISHOR, POLAVARAPU et al. 2014). Alanine, tryptophan, phenylalanine and glutamate (fig 6), however, do
not show an increase in most of the mutants and for some mutant lines even a decrease is observed compared
to col‐0. For instance, tryptophan and phenylalanine could both be involved in providing protein stability (Costa,
Righetti et al. 2015). Tryptophan is increased for mutant line atvip1 which shows a negative effect on re‐
establishment of DT, whereas phenylalanine shows a decrease for mutant line daa1 which shows a greater re‐
establishment of DT at RH. So a change in one metabolite does not have to correlate with the phenotypical
behaviour of that specific line. Studies correlate in most cases with the obtained metabolic results. However
most metabolite results in other studies are obtained from 2 week old plant material and not from the seed
itself. It is possible that therefore results obtained in this thesis do not correlate with results from other studies.
Three metabolites, raffinose, xylose and 4‐hydroxyproline showed the most pronounced change in abundance.
Raffinose together with galactinol and stachyose is accumulated in the seeds of A. thaliana. Raffinose and
galactinol are detected in stressed plants, whereas stachyose is not (Taji, Ohsumi et al. 2002). Two of seven

17
genes for galactinol synthase AtGolS1 and AtGolS2 are induced during drought or salt stresses (Taji, Ohsumi et
al. 2002). Overexpression of AtGOLS2 showed an increase in the accumulation of galactinol and raffinose
together with an increase of drought‐stress tolerance (Taji, Ohsumi et al. 2002). It is suggested that both
galactinol and raffinose are acting as a protectant just as proline (Taji, Ohsumi et al. 2002, Stoyanova, Geuns et
al. 2011). Xylose is important for a plant, because plant N‐linked glycan’s which contain β1.2‐xylose and α1,3‐
fucose have the function to affect the co and post‐translational folding of proteins (Rayon, Lerouge et al. 1998).
The N‐glycan proteins, present in the Golgi apparatus, are essential for root salt tolerance and both β1.2‐xylose
and α1,3‐fucose might have an overlapping function in salt tolerance (Kang, Frank et al. 2008) and if one of the
two is disturbed, salt sensitivity is increased (Kaulfürst‐Soboll, Rips et al. 2011). In this study, interestingly,
xylose drops upon treatment with PEG, but a higher re‐establishment of DT is observed for mutant lines atcc,
daa1 and di19. A lower amount of xylose is observed for mutant line atvip1 in non‐treated seeds. An amino
acid (4‐Hydroxyproline) is used by the cell to produce hydroxyproline‐rich glycoproteins , which is necessary for
a normal embryo development in A. thaliana (Hall and Cannon 2002). Without this protein, the developing cells
of for instance the root will show abnormal shapes, cell differentiation occurs to some degree but cell
expansion does not occur and cell growth is in random directions (Tierney and Varner 1987, Hall and Cannon
2002). It is a logical result that this metabolite is decreased in abundance since no cell differentiation is
necessary during periods of drought stress in A. thaliana seeds.
It is noteworthy to mention mutant line atvip1, which shows a phenotype with a lower re‐establishment of DT.
This line has the most differences in metabolites between treatments compared to the three other mutants
(atcc, daa1 and di19). There are 12 differences, of which eight for untreated and four for treated seeds. Two
metabolites, putrescine and myo‐inositol, show for untreated and treated seeds a decrease in abundance.
There are indications that putrescine can influence the transpiration rate and stomatal conductance as
response for dehydration. This was shown in transgenic A. thaliana plants where Arginine decarboxylase 2 gene
was overexpressed and an overproduction of putrescine was observed (Alcázar, Planas et al. 2010). Results
showed that putrescine plays a role in re‐establishment of DT in germinated seeds. Myo‐inositol is an amino
acid which is used as a substrate for rafinose, which is a oligosacharide that can protect a plant against drought
stress (Taji, Seki et al. 2004). That a decrease of both metabolites would cause a drastic lower re‐establishment
of DT for mutant line atvip1 is unlikely. Sucrose is increased for mutant line atvip1 when treated with PEG
which has also been shown for the sucrose content of both cucumber and Impatiens when treated with PEG
(Bruggink and van der Toorn 1995). This is probably due to breakdown of food reserves, which might also have
a function in stabilizing cell membranes and proteins in the absence of water (Bruggink and van der Toorn
1995).
It is important to mention that not only the T‐DNA insertion in the genome of these lines are the cause of the
metabolic profiles that were found. Point mutations, insertions, deletions or other T‐DNA fragments could have
altered the genome and therefore altered the accumulation of amino acids and sugars during DT.
Naturalre‐establishmentofDT
A dehydration curves show a similar decrease of moisture during a drying period of 72H in a 220C 32% RH
environment for all developmental stages. Within 6h all stages, except TR, have lost more than 90% of
moisture content. This is caused by a lower influence of the seed coat which encloses the embryo in the TR
stage almost completely and therefore inhibits the evaporation of water present in the embryo at a higher
degree (Debeaujon, Léon‐Kloosterziel et al. 2000). For the GC 0h the highest FW has been found, which is a
logical result of a germinating seed that grown longer in favourable conditions. After drying the samples for
72H the DW of the seeds is <0.1 mg. This curve can be used as a standard to search for mutant lines that show
a different trend in losing moisture.

18
PossiblefutureprojectsThere are multiple possible future projects that could help to continue the study on desiccation tolerance. First
of all, an ABA measurement could be performed. Seeds are already treated and stored a the ‐800C freezer. It is
expected that upon drying an increase of ABA is shown and upon rehydration of 24 hours after 72 hours drying
an decrease of ABA is expected to be seen. This curve can be used as a standard for the behaviour of seeds
upon germination. Similar ABA measurements can be made to study ABA in the mutant lines. If an alteration of
ABA behaviour is observed, further investigation could be advised.
Second, a sugar measurement in germinated A. thaliana seeds is another experiment that is going to be
performed. The seeds were prepared and are stored in a ‐800C freezer until they can be put into the machine. It
is interesting to see if treatment with PEG alters the amount of specific sugars. That alteration happens has
already been shown for raffinose, fructose and sucrose in the metabolomics analysis for different treatments.
However, there are more sugars, for instance trehalose, that are involved in desiccation tolerance and can
therefore be investigated. Trehalose is necessary for the survival of desiccation over a long period of time in
Saccharomyces cerevisiae (Tapia and Koshland 2014). Overexpression of yeast trehalose‐6‐phosphate in tobaco
plants increases its tolerance to drought (Romero, Bellés et al. 1997). In A. thaliana, 11 trehalose‐6‐phosphate
synthase genes were found which synthesize UDP glucose and glucose‐6‐phosphate to trehalose‐6‐
phosphosphate and Tre6P phosphatase (TPP) can dephosphorylate it to trehalose (Leyman, Van Dijck et al.
2001). Sugars are, as introduced, important in establishing DT. It would be interesting to see the ratios of
sugars that are present in the mutant lines and it would be interesting if differences in sugar ratios between the
mutant lines can be found.
Third, as introduced, LEA proteins and HSPs are also important in helping a seed to establish DT. It would be
interesting to check which LEA and HSP proteins are transcribed for the developmental stages RP and RH and
how ABA is correlated to the transcription of these proteins.
Finally as mentioned in the discussion, the RH stage of the ‘natural’ drying experiment performed by Josephine
van Eggelen should be repeated (appendix H). Results show a low survival rate, below 10% in average, for the
developmental stage RH, whereas figure 7 shows that survival of at least 20% is expected for the wild type.
Furthermore, line atvip1 shows an positive establishment of DT, whereas for RP (appendix G) and for RH when
treated with PEG a negative establishment of DT is shown. There is a possibility that the seeds were not picked
at the right moment of germination. If the seeds show even the tiniest bit of green, there is no chance of
survival of that seed. The time frame of picking the seeds is roughly 3 to 4 hours. Which can be difficult if a high
number of seeds have to be picked.
AcknowledgementsFirst of all, I would like to thank my direct supervisor Anderson Silva, who guided me through my thesis, helped
me in the laboratory and reviewed this report. Second I would like to thank my other supervisor Wilco Ligterink
who gave me the opportunity to work in the Laboratory of Plant Physiology. I would like to thank the
technicians Leo Willems for helping me with the GC‐TOF‐MS and Juriaan Rienstra for answering my questions
in the laboratory. Finally, I would like to thank Josephine van Eggelen and Adane Demissie for their help in the
laboratory and greenhouse.

19
ReferencesAlamillo,J.,C.Almoguera,D.BartelsandJ.Jordano(1995)."ConstitutiveexpressionofsmallheatshockproteinsinvegetativetissuesoftheresurrectionplantCraterostigmaplantagineum."Plantmolecularbiology29(5):1093‐1099.Alcázar,R.,J.Planas,T.Saxena,X.Zarza,C.Bortolotti,J.Cuevas,M.Bitrián,A.F.TiburcioandT.Altabella(2010)."PutrescineaccumulationconfersdroughttoleranceintransgenicArabidopsisplantsover‐expressingthehomologousArgininedecarboxylase2gene."PlantPhysiologyandBiochemistry48(7):547‐552.Ali,M.A.,S.Plattner,Z.Radakovic,K.Wieczorek,A.Elashry,F.M.Grundler,M.Ammelburg,S.SiddiqueandH.Bohlmann(2013)."AnArabidopsisATPasegeneinvolvedinnematode‐inducedsyncytiumdevelopmentandabioticstressresponses."ThePlantJournal74(5):852‐866.Allen,R.D.(1995)."Dissectionofoxidativestresstoleranceusingtransgenicplants."PlantPhysiology107(4):1049.Asada,K.(1992)."Ascorbateperoxidase–ahydrogenperoxide‐scavengingenzymeinplants."PhysiologiaPlantarum85(2):235‐241.Battista,J.R.,M.‐J.ParkandA.E.McLemore(2001)."InactivationoftwohomologuesofproteinspresumedtobeinvolvedinthedesiccationtoleranceofplantssensitizesDeinococcusradioduransR1todesiccation."Cryobiology43(2):133‐139.Blackman,S.A.,R.L.ObendorfandA.C.Leopold(1992)."Maturationproteinsandsugarsindesiccationtoleranceofdevelopingsoybeanseeds."PlantPhysiology100(1):225‐230.Borg,M.,L.Brownfield,H.Khatab,A.Sidorova,M.LingayaandD.Twell(2011)."TheR2R3MYBtranscriptionfactorDUO1activatesamalegermline‐specificregulonessentialforspermcelldifferentiationinArabidopsis."ThePlantCellOnline23(2):534‐549.Bowler,C.,W.VanCamp,M.VanMontagu,D.InzéandK.Asada(1994)."Superoxidedismutaseinplants."CriticalReviewsinPlantSciences13(3):199‐218.Browne,J.,A.TunnacliffeandA.Burnell(2002)."Anhydrobiosis:plantdesiccationgenefoundinanematode."Nature416(6876):38‐38.Bruggink,T.andP.vanderToorn(1995)."Inductionofdesiccationtoleranceingerminatedseeds."SeedScienceResearch5(01):1‐4.CHIANG,H.H.andA.Dandekar(1995)."RegulationofprolineaccumulationinArabidopsisthaliana(L.)Heynhduringdevelopmentandinresponsetodesiccation."Plant,Cell&Environment18(11):1280‐1290.Chu,C.‐C.,W.‐C.Lee,W.‐Y.Guo,S.‐M.Pan,L.‐J.Chen,H.‐m.LiandT.‐L.Jinn(2005)."Acopperchaperoneforsuperoxidedismutasethatconfersthreetypesofcopper/zincsuperoxidedismutaseactivityinArabidopsis."PlantPhysiology139(1):425‐436.Coca,M.A.,C.AlmogueraandJ.Jordano(1994)."Expressionofsunflowerlow‐molecular‐weightheat‐shockproteinsduringembryogenesisandpersistenceaftergermination:localizationandpossiblefunctionalimplications."Plantmolecularbiology25(3):479‐492.Coca,M.A.,C.Almoguera,T.L.ThomasandJ.Jordano(1996)."Differentialregulationofsmallheat‐shockgenesinplants:analysisofawater‐stress‐inducibleanddevelopmentallyactivatedsunflowerpromoter."Plantmolecularbiology31(4):863‐876.Cohen,H.,H.Israeli,I.MatityahuandR.Amir(2014)."Seed‐SpecificExpressionofaFeedback‐InsensitiveFormofCYSTATHIONINE‐γ‐SYNTHASEinArabidopsisStimulatesMetabolicandTranscriptomicResponsesAssociatedwithDesiccationStress."Plantphysiology166(3):1575‐1592.Costa,M.C.D.,K.Righetti,H.Nijveen,F.Yazdanpanah,W.Ligterink,J.BuitinkandH.W.Hilhorst(2015)."Ageneco‐expressionnetworkpredictsfunctionalgenescontrollingthere‐establishmentofdesiccationtoleranceingerminatedArabidopsisthalianaseeds."Planta:1‐15.Debeaujon,I.,K.M.Léon‐KloosterzielandM.Koornneef(2000)."Influenceofthetestaonseeddormancy,germination,andlongevityinArabidopsis."PlantPhysiology122(2):403‐414.

20
Delauney,A.J.andD.P.S.Verma(1993)."Prolinebiosynthesisandosmoregulationinplants."Theplantjournal4(2):215‐223.Gao,X.,Z.Ren,Y.ZhaoandH.Zhang(2003)."OverexpressionofSOD2increasessalttoleranceofArabidopsis."PlantPhysiology133(4):1873‐1881.Goyal,K.,L.WaltonandA.Tunnacliffe(2005)."LEAproteinspreventproteinaggregationduetowaterstress."Biochem.J388:151‐157.Hall,Q.andM.C.Cannon(2002)."Thecellwallhydroxyproline‐richglycoproteinRSHisessentialfornormalembryodevelopmentinArabidopsis."ThePlantCellOnline14(5):1161‐1172.Hendrick,J.P.andF.Hartl(1993)."Molecularchaperonefunctionsofheat‐shockproteins."Annualreviewofbiochemistry62(1):349‐384.Hoekstra,F.A.,E.A.GolovinaandJ.Buitink(2001)."Mechanismsofplantdesiccationtolerance."Trendsinplantscience6(9):431‐438.Hundertmark,M.andD.K.Hincha(2008)."LEA(lateembryogenesisabundant)proteinsandtheirencodinggenesinArabidopsisthaliana."BMCgenomics9(1):118.Imlay,J.andS.Linn(1988)."DNAdamageandoxygenradicaltoxicity."Science(NewYork,NY)240(4857):1302‐1309.Jones,M.A.,M.F.Covington,L.DiTacchio,C.Vollmers,S.PandaandS.L.Harmer(2010)."JumonjidomainproteinJMJD5functionsinboththeplantandhumancircadiansystems."ProceedingsoftheNationalAcademyofSciences107(50):21623‐21628.Joshi,V.,J.‐G.Joung,Z.FeiandG.Jander(2010)."Interdependenceofthreonine,methionineandisoleucinemetabolisminplants:accumulationandtranscriptionalregulationunderabioticstress."Aminoacids39(4):933‐947.Jung,C.,J.S.Seo,S.W.Han,Y.J.Koo,C.H.Kim,S.I.Song,B.H.Nahm,Y.DoChoiandJ.‐J.Cheong(2008)."OverexpressionofAtMYB44enhancesstomatalclosuretoconferabioticstresstoleranceintransgenicArabidopsis."PlantPhysiology146(2):623‐635.Kang,J.S.,J.Frank,C.H.Kang,H.Kajiura,M.Vikram,A.Ueda,S.Kim,J.D.Bahk,B.TriplettandK.Fujiyama(2008)."SalttoleranceofArabidopsisthalianarequiresmaturationofN‐glycosylatedproteinsintheGolgiapparatus."ProceedingsoftheNationalAcademyofSciences105(15):5933‐5938.Kaulfürst‐Soboll,H.,S.Rips,H.Koiwa,H.Kajiura,K.FujiyamaandA.vonSchaewen(2011)."ReducedimmunogenicityofArabidopsishgl1mutantN‐glycanscausedbyalteredaccessibilityofxyloseandcorefucoseepitopes."JournalofBiologicalChemistry286(26):22955‐22964.KISHOR,K.,B.POLAVARAPUandN.SREENIVASULU(2014)."Isprolineaccumulationpersecorrelatedwithstresstoleranceorisprolinehomeostasisamorecriticalissue?"Plant,cell&environment37(2):300‐311.Koornneef,M.,C.J.Hanhart,H.W.HilhorstandC.M.Karssen(1989)."InvivoinhibitionofseeddevelopmentandreserveproteinaccumulationinrecombinantsofabscisicacidbiosynthesisandresponsivenessmutantsinArabidopsisthaliana."PlantPhysiology90(2):463‐469.Koster,K.L.(1991)."Glassformationanddesiccationtoleranceinseeds."PlantPhysiology96(1):302‐304.Koster,K.L.andA.C.Leopold(1988)."Sugarsanddesiccationtoleranceinseeds."PlantPhysiology88(3):829‐832.Lam,H.‐M.,S.S.PengandG.M.Coruzzi(1994)."Metabolicregulationofthegeneencodingglutamine‐dependentasparaginesynthetaseinArabidopsisthaliana."PlantPhysiology106(4):1347‐1357.Lam,H.M.,M.H.HsiehandG.Coruzzi(1998)."ReciprocalregulationofdistinctasparaginesynthetasegenesbylightandmetabolitesinArabidopsisthaliana."ThePlantJournal16(3):345‐353.Lehle,F.R.,F.ChenandK.R.Wendt(1992)."EnhancementofNaCltoleranceinArabidopsisthalianabyexogenousL‐asparagineandD‐asparagine."PhysiologiaPlantarum84(2):223‐228.Leyman,B.,P.VanDijckandJ.M.Thevelein(2001)."AnunexpectedplethoraoftrehalosebiosynthesisgenesinArabidopsisthaliana."Trendsinplantscience6(11):510‐513.

21
Liu,J.,Y.Zhang,G.Qin,T.Tsuge,N.Sakaguchi,G.Luo,K.Sun,D.Shi,S.AkiandN.Zheng(2008)."Targeteddegradationofthecyclin‐dependentkinaseinhibitorICK4/KRP6byRING‐typeE3ligasesisessentialformitoticcellcycleprogressionduringArabidopsisgametogenesis."ThePlantCellOnline20(6):1538‐1554.Liu,W.‐X.,F.‐C.Zhang,W.‐Z.Zhang,L.‐F.Song,W.‐H.WuandY.‐F.Chen(2013)."ArabidopsisDi19functionsasatranscriptionfactorandmodulatesPR1,PR2,andPR5expressioninresponsetodroughtstress."Molecularplant6(5):1487‐1502.Lu,S.X.,S.M.Knowles,C.J.Webb,R.B.Celaya,C.Cha,J.P.SiuandE.M.Tobin(2011)."TheJumonjiCdomain‐containingproteinJMJ30regulatesperiodlengthintheArabidopsiscircadianclock."Plantphysiology155(2):906‐915.Maia,J.,B.J.Dekkers,M.J.Dolle,W.LigterinkandH.W.Hilhorst(2014)."Abscisicacid(ABA)sensitivityregulatesdesiccationtoleranceingerminatedArabidopsisseeds."NewPhytologist203(1):81‐93.Maia,J.,B.J.Dekkers,N.J.Provart,W.LigterinkandH.W.Hilhorst(2011)."There‐establishmentofdesiccationtoleranceingerminatedArabidopsisthalianaseedsanditsassociatedtranscriptome."PloSone6(12):e29123.Meurs,C.,A.S.Basra,C.M.KarssenandL.C.vanLoon(1992)."RoleofabscisicacidintheinductionofdesiccationtoleranceindevelopingseedsofArabidopsisthaliana."Plantphysiology98(4):1484‐1493.Nambara,E.,H.Kawaide,Y.KamiyaandS.Naito(1998)."CharacterizationofanArabidopsisthalianamutantthathasadefectinABAaccumulation:ABA‐dependentandABA‐independentaccumulationoffreeaminoacidsduringdehydration."Plantandcellphysiology39(8):853‐858.Rayon,C.,P.LerougeandL.Faye(1998)."TheproteinN‐glycosylationinplants."JournalofExperimentalBotany49(326):1463‐1472.Romero,C.,J.M.Bellés,J.L.Vayá,R.SerranoandF.A.Culiáñez‐Macià(1997)."Expressionoftheyeasttrehalose‐6‐phosphatesynthasegeneintransgenictobaccoplants:pleiotropicphenotypesincludedroughttolerance."Planta201(3):293‐297.Smirnoff,N.andG.L.Wheeler(2000)."Ascorbicacidinplants:biosynthesisandfunction."CriticalReviewsinBiochemistryandMolecularBiology35(4):291‐314.Smith,I.K.,T.L.VierhellerandC.A.Thorne(1989)."Propertiesandfunctionsofglutathionereductaseinplants."PhysiologiaPlantarum77(3):449‐456.Stacy,R.A.andR.B.Aalen(1998)."IdentificationofsequencehomologybetweentheinternalhydrophilicrepeatedmotifsofGroup1late‐embryogenesis‐abundantproteinsinplantsandhydrophilicrepeatsofthegeneralstressproteinGsiBofBacillussubtilis."Planta206(3):476‐478.Stoyanova,S.,J.Geuns,E.HidegandW.VanDenEnde(2011)."Thefoodadditivesinulinandsteviosidecounteractoxidativestress."Internationaljournaloffoodsciencesandnutrition62(03):207‐214.Stracke,R.,M.WerberandB.Weisshaar(2001)."TheR2R3‐MYBgenefamilyinArabidopsisthaliana."Currentopinioninplantbiology4(5):447‐456.Sun,W.,M.VanMontaguandN.Verbruggen(2002)."Smallheatshockproteinsandstresstoleranceinplants."BiochimicaetBiophysicaActa(BBA)‐GeneStructureandExpression1577(1):1‐9.Sun,W.Q.,T.C.IrvingandA.C.Leopold(1994)."Theroleofsugar,vitrificationandmembranephasetransitioninseeddesiccationtolerance."PhysiologiaPlantarum90(4):621‐628.Sun,Y.andT.H.MacRae(2005)."Smallheatshockproteins:molecularstructureandchaperonefunction."CellularandMolecularLifeSciencesCMLS62(21):2460‐2476.Synek,L.,N.Schlager,M.Eliáš,M.Quentin,M.T.HauserandV.Žárský(2006)."AtEXO70A1,amemberofafamilyofputativeexocystsubunitsspecificallyexpandedinlandplants,isimportantforpolargrowthandplantdevelopment."ThePlantJournal48(1):54‐72.Taji,T.,C.Ohsumi,S.Iuchi,M.Seki,M.Kasuga,M.Kobayashi,K.Yamaguchi‐ShinozakiandK.Shinozaki(2002)."Importantrolesofdrought‐andcold‐induciblegenesforgalactinolsynthaseinstresstoleranceinArabidopsisthaliana."ThePlantJournal29(4):417‐426.

22
Taji,T.,M.Seki,M.Satou,T.Sakurai,M.Kobayashi,K.Ishiyama,Y.Narusaka,M.Narusaka,J.‐K.ZhuandK.Shinozaki(2004)."ComparativegenomicsinsalttolerancebetweenArabidopsisandArabidopsis‐relatedhalophytesaltcressusingArabidopsismicroarray."Plantphysiology135(3):1697‐1709.Tapia,H.andD.E.Koshland(2014)."Trehaloseisaversatileandlong‐livedchaperonefordesiccationtolerance."CurrentBiology24(23):2758‐2766.Tierney,M.L.andJ.E.Varner(1987)."Theextensins."Plantphysiology84(1):1‐2.Tunnacliffe,A.andM.J.Wise(2007)."ThecontinuingconundrumoftheLEAproteins."Naturwissenschaften94(10):791‐812.Wehmeyer,N.andE.Vierling(2000)."Theexpressionofsmallheatshockproteinsinseedsrespondstodiscretedevelopmentalsignalsandsuggestsageneralprotectiveroleindesiccationtolerance."PlantPhysiology122(4):1099‐1108.Williams,R.J.andA.C.Leopold(1989)."Theglassystateincornembryos."PlantPhysiology89(3):977‐981.Wise,M.J.andA.Tunnacliffe(2004)."POPPthequestion:whatdoLEAproteinsdo?"Trendsinplantscience9(1):13‐17.Wolkers,W.F.,F.A.Tetteroo,M.AlberdaandF.A.Hoekstra(1999)."Changedpropertiesofthecytoplasmicmatrixassociatedwithdesiccationtoleranceofdriedcarrotsomaticembryos.AninsituFouriertransforminfraredspectroscopicstudy."PlantPhysiology120(1):153‐164.Xia,J.andD.S.Wishart(2011)."Web‐basedinferenceofbiologicalpatterns,functionsandpathwaysfrommetabolomicdatausingMetaboAnalyst."Natureprotocols6(6):743‐760.Xu,D.,M.Chen,Y.Ma,Z.Xu,L.Li,Y.ChenandY.Ma(2015)."AG‐ProteinβSubunit,AGB1,NegativelyRegulatestheABAResponseandDroughtTolerancebyDown‐RegulatingAtMPK6‐RelatedPathwayinArabidopsis."PloSone10(1):e0116385.Zelko,I.N.,T.J.MarianiandR.J.Folz(2002)."Superoxidedismutasemultigenefamily:acomparisonoftheCuZn‐SOD(SOD1),Mn‐SOD(SOD2),andEC‐SOD(SOD3)genestructures,evolution,andexpression."FreeRadicalBiologyandMedicine33(3):337‐349.Zhu,S.‐Y.,X.‐C.Yu,X.‐J.Wang,R.Zhao,Y.Li,R.‐C.Fan,Y.Shang,S.‐Y.Du,X.‐F.WangandF.‐Q.Wu(2007)."Twocalcium‐dependentproteinkinases,CPK4andCPK11,regulateabscisicacidsignaltransductioninArabidopsis."ThePlantCellOnline19(10):3019‐3036.

23
Appendix
A. Protocol:DNAextraction1. 1 leaf per tube
2. Add 200 µL of extraction buffer, one stainless steel bullet and grind (1 min at 30 Hz)
3. 60 minutes at 60 ºC
4. Spin 15 minutes at 2.800 g
5. Take 75 µL of supernatant to new tube (Microtiter plate)
6. Add 75 µL of iso‐propanol and 30 µL of 10M of NH4Ac
7. 15 minutes at Room Temperature/ or over‐night
8. Spin 15 at room temperature at 2.500 g
9. Wash pellet with 70% of ethanol
10. Spin 5 minutes at 2.500 g, remove the ethanol
11. Quick spin on the plate upside‐down
12. Dissolve the pellet in 50 µL Milli‐Q water
13. Use up to 4 µL for the PCR
Extraction buffer 2M of NaCl (116.88 g per Litre)
200mM of Tris‐HCl pH8 (31.520 g per 1 Litre)
70mM EDTA (29.134 g per Litre)
20mM Na2S2O5 (3.802 g per Litre)

24
B. CompositionofHyponexMain Elements Concentration (mM/L) Trace Elements Concentration (μM/L)
NH4 1.7 Fe 21.0
K 4.131 Mn 3.4
Ca 1.97 Zn 4.7
Mg 1.239 B 14.0
NO3 4.141 Cu 6.9
SO4 3.411 MO 0.1
P 1.287

25
C. Mutantlines
Table 4: Mutant lines used in this thesis
*Gene expression after 6‐24 hours of imbibition in water. +: increase, ‐: decrease, O: No increase or decrease in gene expression. ND: No Data. **Expression of abiotic stress, osmotic( 300mM Mannitol), salt (150mM NaCl) and drought stress (plants were exposed for 15 min with loss of app.10% fresh weight) in both root and shoot. O: No difference compared to the wild type (WT). O/‐: 25‐50% negative difference compared to the WT. O/+:25‐50% positive difference compared to the WT. +: 50‐500% positive difference compared to the WT. ++: 500‐1000%: positive difference compared to the WT. +++:1000+ % positive difference compared to the WT. If expression is below a factor 100 a O (no change) will be noted. The ‐, ‐‐ and ‐‐‐ correspond to the same percentage as +, ++ and +++, but as a decrease in expression. Data obtained from: http://bar.utoronto.ca/efp/cgi‐bin/efpWeb.cgi
Number T‐DNA (agi) Expression during germination*
Expression of abiotic stress in the root** Expression of abiotic stress in the shoot**
Osmotic Salt Drought Osmotic Salt Drought
3 AT1G33970 1‐6h + 12‐24h ‐
O/+ O O ++ O O
6 AT2G01100 ‐ O O O + + O
8 AT5G01520 ‐ + + O +++ ++ O
9 AT5G22000 ‐ O/+ + O + + O
10 AT5G58720 ‐ O/+ O O O/+ O O
12 AT5G66400 1h + 3‐24h ‐
+++ +++ O +++ +++ +
13 AT4G01450 + + O O/+ O + O/+
18 AT1G04300 ‐ O/+ O O + O O
20 AT2G04240 1h,6h + 3,12,24h ‐
O O O + + O
21 AT3G14880 ‐ O O O O O O
22 AT3G52180 + O O O + + O
23 AT3G53530 ND ND ND ND ND ND ND
24 AT4G25800 O O O O O O O
25 AT5G03240 1‐6h + 12‐24h ‐
O O ‐/O + + O
29 AT5G57110 ‐ O O O O O O
30 AT1G53780 ‐ O O O + O/+ O
31 AT2G47850 ‐ O O O O O O
34 AT3G14590 ND ND ND ND ND ND ND
36 AT4G16160 1h + 3‐24h ‐
O O O O O O
38 AT5G04750 1h + 3‐24h ‐
+ O O + + O/+
39 AT5G52300 ‐ +++ +++ O +++ +++ O
40 AT1G30500 ‐ O O O + O/+ O
41 AT2G41070 ‐ O O O O O O
42 AT2G41710 ‐ O O O O O O
43 AT2G33830 1h + 3‐24h ‐
+ O O ++ O O
44 AT3G62090 ‐ O O O O O O
45 AT1G01240 ‐ + + O + O/+ +

26
D. Protocol:Extractionofmetabolites(polar+apolarphase)forGC‐TOF‐MSmodified from Proteomics 2004, 4, 78-83
Internal standard stocks:
ribitol (adonitol) 1 mg/ml in water (polar phase)
nonanedecanoic acid methylester 1 mg/ml CHCl3 (organic phase)
Extraction mixes:
‐ Mix A: MeOH:CHCl3:internal standard (nonanedecanoicacid me 1 mg/ml): 400µl:280µl:20µl
‐ Mix B: Water ribitol: 130 µl water + 20 µl ribitol (1 mg/ml)
‐ Mix C: MeOH:CHCl3, 1:1 (v/v)
All steps kept on ice!
Extraction
‐ weigh ~20mg FW material in 2 ml Eppendorf tube and keep everything frozen!
‐ add 700 µl mix A
‐ vortex briefly
‐ add 150 µl mix B
‐ vortex thoroughly
‐ 10 min sonication in Ultrasonic bath
‐ add 200μL H2O
‐ vortex thoroughly
‐ centrifuge 5 min 15000 rpm
‐ transfer 90% of the upper phase to a 2mL Eppendorf tube
‐ add 0.5 mL mix C to the remaining organic/inter phase
‐ vortex thoroughly
‐ keep on ice ~10min
‐ add 200 μL H2O
‐ vortex thoroughly
‐ centrifuge 5 min 15000 rpm
‐ collect 90% of the upper phase and add to the previously collected polar phase
‐ collect the (lower) organic phase and transfer to a glass vial
‐ store the organic phase at ‐20°C until further processing
‐ transfer 25 µl of 10x diluted and 25‐100‐200 µl of the undiluted polar phase to inserts in 1.5 ml
glass GC‐vials and dry over night
‐ analyse on the GC‐TOF‐MS using the online derivatisation protocol.
100µl in insert
27
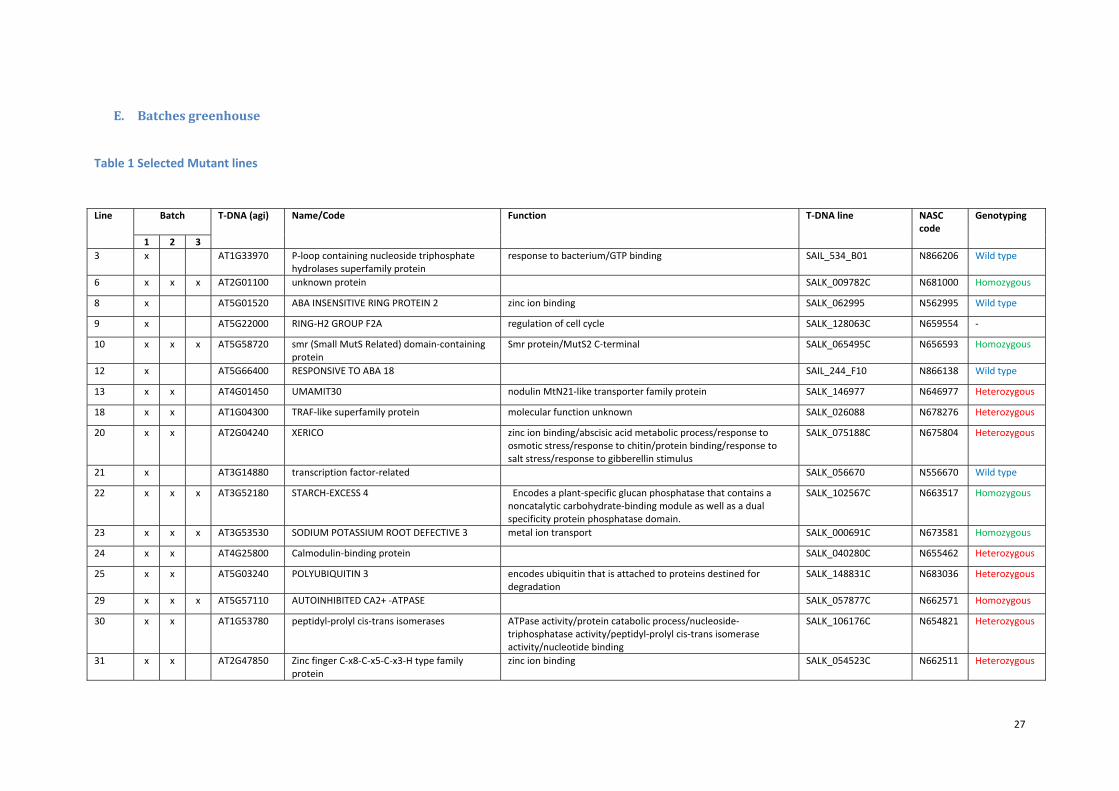
E. Batchesgreenhouse
Table 1 Selected Mutant lines
Line Batch
T‐DNA (agi) Name/Code Function T‐DNA line NASC code
Genotyping
1 2 3
3 x AT1G33970 P‐loop containing nucleoside triphosphate hydrolases superfamily protein
response to bacterium/GTP binding SAIL_534_B01 N866206 Wild type
6 x x x AT2G01100 unknown protein SALK_009782C N681000 Homozygous
8 x AT5G01520 ABA INSENSITIVE RING PROTEIN 2 zinc ion binding SALK_062995 N562995 Wild type
9 x AT5G22000 RING‐H2 GROUP F2A regulation of cell cycle SALK_128063C N659554 ‐
10 x x x AT5G58720 smr (Small MutS Related) domain‐containing protein
Smr protein/MutS2 C‐terminal SALK_065495C N656593 Homozygous
12 x AT5G66400 RESPONSIVE TO ABA 18 SAIL_244_F10 N866138 Wild type
13 x x AT4G01450 UMAMIT30 nodulin MtN21‐like transporter family protein SALK_146977 N646977 Heterozygous
18 x x AT1G04300 TRAF‐like superfamily protein molecular function unknown SALK_026088 N678276 Heterozygous
20 x x AT2G04240 XERICO zinc ion binding/abscisic acid metabolic process/response to osmotic stress/response to chitin/protein binding/response to salt stress/response to gibberellin stimulus
SALK_075188C N675804 Heterozygous
21 x AT3G14880 transcription factor‐related SALK_056670 N556670 Wild type
22 x x x AT3G52180 STARCH‐EXCESS 4 Encodes a plant‐specific glucan phosphatase that contains a noncatalytic carbohydrate‐binding module as well as a dual specificity protein phosphatase domain.
SALK_102567C N663517 Homozygous
23 x x x AT3G53530 SODIUM POTASSIUM ROOT DEFECTIVE 3 metal ion transport SALK_000691C N673581 Homozygous
24 x x AT4G25800 Calmodulin‐binding protein SALK_040280C N655462 Heterozygous
25 x x AT5G03240 POLYUBIQUITIN 3 encodes ubiquitin that is attached to proteins destined for degradation
SALK_148831C N683036 Heterozygous
29 x x x AT5G57110 AUTOINHIBITED CA2+ ‐ATPASE SALK_057877C N662571 Homozygous
30 x x AT1G53780 peptidyl‐prolyl cis‐trans isomerases ATPase activity/protein catabolic process/nucleoside‐triphosphatase activity/peptidyl‐prolyl cis‐trans isomerase activity/nucleotide binding
SALK_106176C N654821 Heterozygous
31 x x AT2G47850 Zinc finger C‐x8‐C‐x5‐C‐x3‐H type family protein
zinc ion binding SALK_054523C N662511 Heterozygous

28
34 x x x AT3G14590 NTMC2T6.2 Calcium‐dependent lipid‐binding SALK_131027 N631027 Homozygous
36 x x AT4G16160 ATOEP16‐S plastid outer membrane/mitochondrial inner membrane presequence translocase complex/P‐P‐bond‐hydrolysis‐driven protein transmembrane transporter activit
SALK_025109 N525109 Heterozygous
38 x x AT5G04750 F1F0‐ATPase inhibitor protein SALK_023158C N679430 Heterozygous
39 x x AT5G52300 73A HM Primers 926/927 Homozygous
40 x x AT1G30500 CBF/NF‐yA7 SALK_020063 Homozygous
41 x x AT2G41070 Bzip12/eel SALK_021965 Homozygous
42 x x AT2G41710 N658851 Homozygous
43 x x AT2G33830 dormancy/auxin associated family protein SALK_054451.53.45.x N554451 Homozygous
44 x x AT3G62090 PIL2 (PHYTOCHROME INTERACTING FACTOR 3‐LIKE 2); protein binding / transcription factor
SALK_090239C N661012 Homozygous
45 x x AT1G01240 unknown protein SALK_104275C N653407 Homozygous

29
F. Re‐establishmentofDTwithnaturaldrying
Figure 8: Re‐establishment of desiccation tolerance innatural drying conditions for the developmental stages RPand RH. After 72 hour drying seeds were put for 1 week onthe Copenhagen table. Seeds which germinated werecounted and are shown as a survival percentage.

30
G. Metabolomicsgraphs
c
a
c
a
c
a
c
a
b
a
0
4
8
12
16
Xylo se
abc c abc abc abc a ab bc abc abc
0
3
6
9
12
15
Urea
abcd
ad
bcd
ab
dc
d
0
3
6
9
12
15
Valine
abc c a abc ab abc ab abc abc bc
0
4
8
12
16
Tyrosine 3tms
ab
ab
ab
ab
ab
0
4
8
12
16
Th reonine 3tms
abc de bc
ef
bc
ef
abc d
f
a
de
0
4
8
12
Threito l
ab c b cb c b
ca
d
0
5
10
15
20
Sucrose
b
a
b
a
b
a
b
a
b
a
0
4
8
12
16
Su ccinic acid
a
b
a
b
a
b
a
b
a
b
0
4
8
12
S-M eth yl-L-cysteine
ab
c
a
c
ab
c
ab
cb
c
0
4
8
12
16
Ser ine
a
b
a
b
a
b
a
b
a
b
0
4
8
12
16
Raffinose
dabcd
c dabc d
d
a
bc d
abc
c d
ab
0
3
6
9
12
Pyru vic acid
abc
ab
cb
ca
cb
c
0
4
8
12
16
Pyro glu tamic acid
bde df bc def
bcde c bc e c def
a ab
0
3
6
9
12
Putrescine
a
bc
a
bc
ab
a
bca
c
0
5
10
15
20
Proline
a cab
c ab c ab c b c
0
5
10
15
20
Pho sphoric acid
bcab
bcabc
cab
aba
bcab
0
4
8
12
16
Ph enylalan ine
abc bc ab aabc
abc abc abc a c
0
3
6
9
12
No nanoic acid
bc
ab
cabc
c
abc
c
abc
a
0
4
8
12
16
N-Acetylglu tamic acid
d
b
d
ab
d
ab
c d
a
c
a
0
5
10
15
20
Myo ino sitol
a c bc bcab
abcabc
c ab abc
0
3
6
9
12
Mono methylpho sp hate (2TM S)ab
ab ab bb
aba ab
b
ab
0
2
4
6
8
10
Methio nin e
b
a
b
a
b
a
b
a
b
a
0
5
10
15
20
M al ic acid
abc
ac
abc
ac
a b
0
4
8
12
16
Lysine
a bc abc a bc a c
abbc
0
4
8
12
16
Leucine 2tms
ab
ab
a b ab a b
0
4
8
12
16
isoleu cine
acde
abd
cab
cea
cab bde
0
4
8
12
16
Glycolic acid
abc abcd abd b c d a bc d ab
bc
0
4
8
12
16
G lycin e (3tms)
b
a
b
a
b
a
b
a
b
a
0
4
8
12
16
Glyceric acid
a ca
abc a bc a c b c
0
5
10
15
20
Glutamine
bcdabc
c dab
dabc d
bc dab
cd
a
0
5
10
15
20
Glu tamic acid
cabc
cabc
cabc
c
abbc
a
0
4
8
12
16
G lucose-6-pho sphatebc
abc bc abc
abcbc
abc bca
0
5
10
15
20
Glucose
abab
abc da
cd
ab
d
abbc
ab
0
4
8
12
16
Glu copyrano se
b
ab
a
b
a
b
a
b
a
0
4
8
12
16
Galactinol
a deab bc de abc d abc e c de
abcdeab
0
4
8
12
16
GABA
bc dab
bca
c
abbc
ab
d
bc
0
4
8
12
16
Fu maric acid
c d
ab
c
a
c
a
c
a
bd
a
0
5
10
15
20
Fructose
c d
ab
c
a
c
a
c
a
bd
a
0
5
10
15
20
Ethano lamin e
aab
a aba
abab
abab
b
0
5
10
15
20
Citric acid
b
c
b
c
ab
c
ab
c
a
c
0
4
8
12
16
beta-D-Methyl fr uctofuranoside
cabc
c abcc
abcbc
abc ba
0
3
6
9
12
Beta alan ine
a ba
b a b a b a b
0
2
4
6
8
10
Ben zo ic acida b a b a b a b a b
0
4
8
12
16
aspartic acid
ab c da
cd a c d ab cd bc d
4
8
12
16
Asp ar agin e
abc abd bc d a bc d
abc d c d ab bc d
4
8
12
16
ascorb ic acid
bac
bac
b
a
bc
a
b
a
4
8
12
16
alfa-ketoglu tar ic acidc
a
c
a
c
a
c
a
b
a
4
8
12
16
4-Hydroxypr olineab
b a b ab ab a bab
ab
4
8
12
16
Alan ine
abc bca abc
ab bc ab abc b c
0
4
8
12
16
Tryp to phan

31


















