
Determinacion Experimental de Biomoleculas II Carbohidratos y Lipidos
Upload
estudiantes-de-medicina-fundacion-barceloCategory
view
9.380download
2
1
BIOSINTESIS DE ÁCIDOS GRASOS

• Cuando la ingesta supera las necesidades energéticas, el exceso se almacena como reserva en forma de grasas.
• Los restos de acetil-CoA provenientes de la β-oxidación y de la degradación de glucosa o de las cadenas carbonadas de algunos aa, pueden utilizarse para sintetizar nuevos ac. Grasos.
• Estos se incorporan al glicerol para ser almacenados como grasa de depósito.
• La síntesis de ac. Grasos de hasta 16 C ocurre en el citoplasma y se conoce como SINTESIS DE NOVO.
• La elongación de ac. Grasos preexistentes se realiza en las mitocondrias.
2

3
SINTESIS DE NOVO(SINTESIS CITOPLASMÁTICA DE AC. GRASOS)
• Los ac. grasos se sintetizan en el citosol a partir de acetil-CoA que se produce en la mitocondria por lo tanto es necesario que estos últimos sean transportados afuera de las mitocondrias.
• La membrana mitocondrial interna es impermeable a acetil-CoA.
• La manera en que salen es como citrato mediante un transportador de tricarboxilatos.
• Se usa NADPH como fuente de H para las reducciones.

SALIDA DE ACETILOS DE LA MITOCONDRIA AL CITOSOL
4

CARACTERÍSTICAS DE LAS VÍAS PARA LA BIOSÍNTESIS
Los intermediarios en la síntesis están unidos a grupos sulfihidrilos de una proteína portadora de grupos de acilos .
Las enzimas de la síntesis de los ácidos grasos están integrados en una única cadena polipeptídica llamada ACIDO GRASO SINTASA.
La cadena de ácido graso en crecimiento se alarga por adición secuencial de dos carbonos derivados de la molécula de Acetil-Coa.
La elongación catalizada por el complejo ÁCIDO GRASO SINTASA se detiene con la formación del palmitato. La elongación posterior y la inserción de dobles enlaces se lleva a cabo por otros sistemas enzimáticos

Etapas de la Síntesis de Ac. Grasos
Comprende: Formación de malonil-CoA. Reacciones catalizadas por
el complejo multienzimático de la Ácido graso sintasa.
6

1)Formación de malonil-CoA• Es una carboxilación que requiere HCO3
- como fuente de CO2.
• Cataliza: acetil-CoA carboxilasa que usa biotina (vit B7) como coenzima.
• Es el principal sitio de regulación de la síntesis de ac. Grasos.
7
H3C C
O
S CoA + CO2
ATP ADP + Pi
H2C C
O
S CoA
COO-
acetil-CoAcarboxilasa
acetil-CoA malonil-CoA

2)Reacciones de la acido graso sintetasa
• Cataliza la síntesis de ac. Grasos de hasta 16 C.• Posee 7 proteínas con actividad catalítica.• Formada por 2 subunidades, cada una con 3
dominios:• Dominio 1: ingreso de sustratos y unidad de
condensación. Contiene 3 enzimas:• Acetil transferasa (AT)• Malonil transferasa (MT)• Enzima condensante (KS) con resto de Cys.
• Dominio 2: unidad de reducción. Contiene 3 enzimas:• Cetoacil reductasa (KR)• Hidroxiacil deshidratasa (HD)• Enoil reductasa (ER)
– Posee la porción transportadora de acilos ACP( contiene un residuo de ácido pantoténico en su estructura a través del cual se forma un enlace tioester con la molécula de ácido graso en crecimiento.)
• Dominio 3: liberación de ácidos grasos. Posee la enzima:• Deacilasa.
8
Una subunidad de Acido Graso Sintetasa.

1)Transferencia de acetato.
9
Una molécula de acetil-CoA ingresa y la acetil transferasa (AT) transfiere el resto acetilo al sitio activo de la enzima condensante (KS).

2)Transferencia de malonilo.
10
El malonil-CoA formado ingresa y se une al residuo de Fosfopanteteína de la Proteína Transportadora de Acilos (ACP) por acción de la malonil transferasa (MT).

3)Condensación de acetilo con malonilo
11
•El carboxilo libre del malonilo se separa como CO2.
•Se produce la unión de acetilo y malonilo catalizada por la enzima condensante (KS) para formar ceto-acil ACP.
•Se libera el acetilo de la enzima condensante.

4) Primera reducción( reducción del grupo ceto)
12
El ceto-acil ACP formado se reduce a hidroxi-acil ACP por acción de la ceto-acil reductasa (KR).

5)Deshidratación
13
Se pierde una molécula de agua, reacción catalizada por la hidroxi acil deshidratasa (HD).

6)Segunda reducción(Saturación del enlace C-C)
14
El compuesto insaturado es hidrogenado por acción de la enoil reductasa (ER).

Translocación
15
La cadena en elongación unida al grupo fosfopanteteína de la ACP es translocada al residuo de cisteína de la enzima condensante (KS).
El grupo fosfopanteteína queda libre para la unión a malonilo comenzando un nuevo ciclo.
El ciclo se repite hasta llegar a ac. Grasos de 16 C.
Los H necesarios para las reducciones provienen de NADPH que se obtiene en la vía de las pentosas y en menor cantidad por la enzima málica que convierte el piruvato en malato para su salida al citosol (transporte de acetilos)

RESUMEN: Pasos de la biosíntesis de Ac. Grasos.
16

REGULACION DE LA LIPOGENESIS
• La enzima Acetil Coa carboxilasa en la enzima reguladora de la síntesis de los ácidos grasos.
• Ésta enzima es regulada:4.Mecanismo covalente (citrato estimula a la
enzima y el palmitato la inhibe)5.Mecanismo alostérico (forma activa la
desfosforilada)

Balance de la síntesis del AcidoPalmítico
acetyl-CoA + 7 malonil-CoA + 14 NADPH palmitate + 7 CO2 + 14 NADP+ + 8 CoA
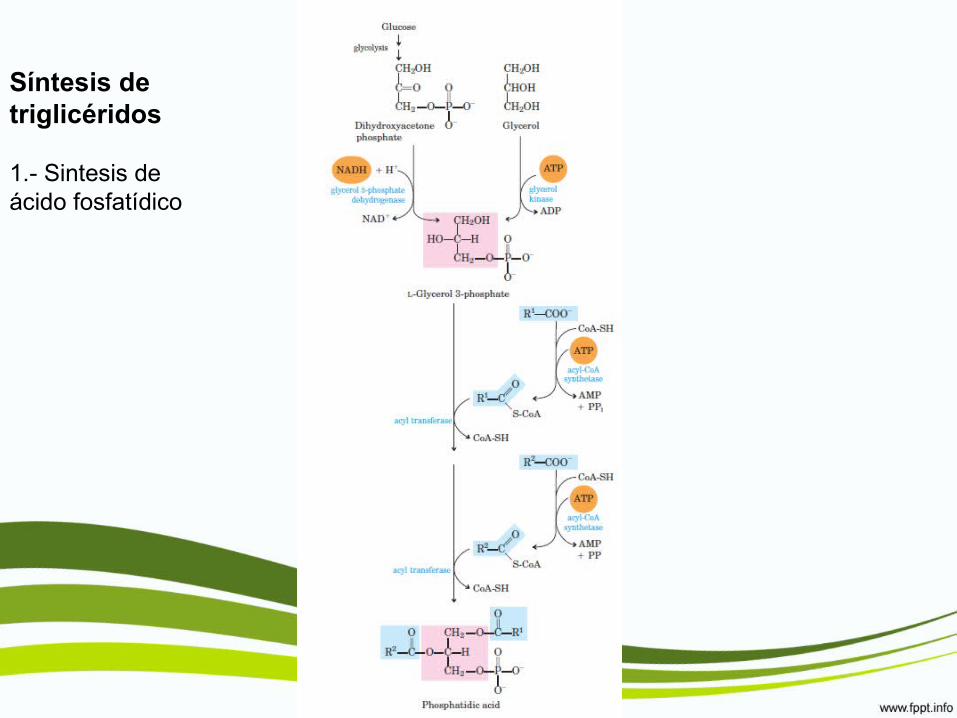
Síntesis de triglicéridos
1.- Sintesis de ácido fosfatídico

2.- Transformación de ác. Fosfatídico en triacilglicéridos

CATABOLISMO DE LOS ÁCIDOS GRASOS
21

β-Oxidación de Ácidos Grasos
• Ocurre en tejidos como: Hígado, músculo esquelético, corazón, riñón, tej. Adiposo, etc.
• Comprende la oxidación del carbono β del ácido graso.
• Ocurre en las MITOCONDRIAS.• Antes debe ocurrir:
1. Activación del ácido graso (requiere energía en forma de ATP)
2. Transporte al interior de la mitocondria
22
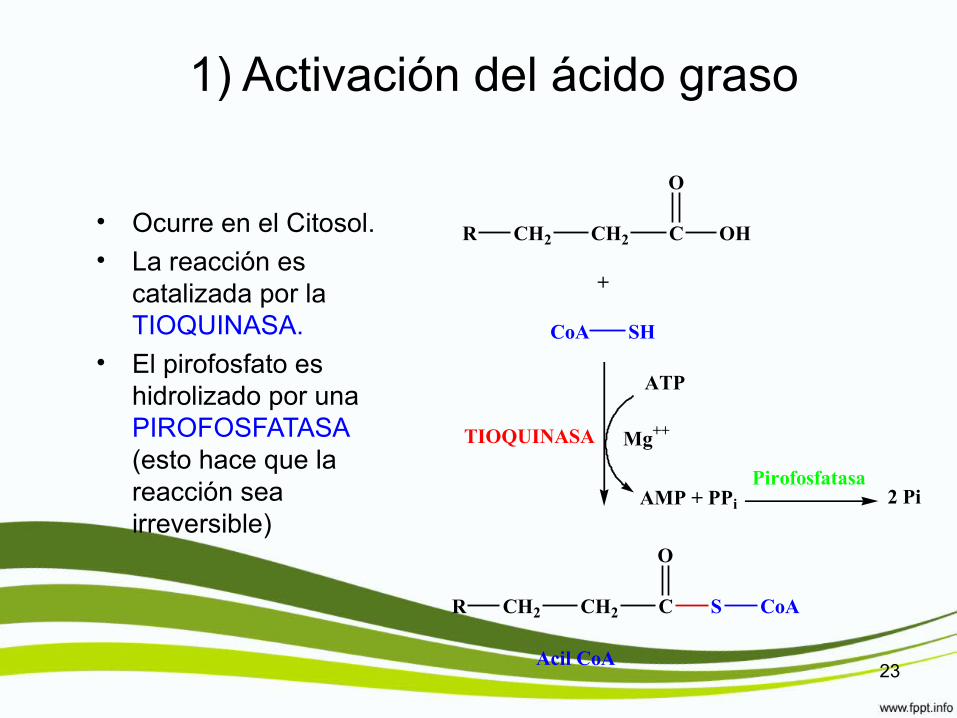
1) Activación del ácido graso
• Ocurre en el Citosol.• La reacción es
catalizada por la TIOQUINASA.
• El pirofosfato es hidrolizado por una PIROFOSFATASA (esto hace que la reacción sea irreversible)
R CH2 CH2 C
O
OH
+
CoA SH
ATP
AMP + PPi
Mg++TIOQUINASA
R CH2 CH2 C
O
S CoA
Acil CoA
2 PiPirofosfatasa
23

2) Transporte de Acil-CoA al interior de la mitocondria.
24

β- Oxidación de Ac. Grasos
25

Después de la activación, los ésteres de ac. grasos con CoA entran a la mitocondria para ser procesados.
β-Oxidación• Los ácidos grasos son procesados por las mismas 5
etapas cíclicas.• Se remueven 2 carbonos por ciclo• Se produce una molécula de Acetil-CoA en cada
ciclo.• El acetil-CoA producido entra en el ciclo de
Krebs para producir energía.
26

¿Porqué se llama β-Oxidación?En este proceso el carbono β del ác. Graso se
oxida a una cetona y luego a un tioéster.
27

28
Se obtienen 4-5 ATP por ciclo de β-Oxidación

29


INTERRELACION CON EL CICLO DE KREBS
31
•El acetil-CoA formado en la β-OXIDACIÓN ingresa al CICLO DE KREBS para su oxidación total a CO2.
•Los NADH y FADH2 producidos en el CICLO DE KREBS forman ATP en la cadena de transporte de e- (FOSFORILACIÓN OXIDATIVA)

32
•En cada ciclo se pierden 2 átomos de C en forma de Acetil-CoA.
•Para degradar completamente un ac. Graso de 16 C hacen faltan 7 ciclos de β-Oxidación.
Nº de ciclos = (nº de C) – 1
2
•En cada ciclo se produce 1 molécula de FADH2 y otra de NADH:
FADH2= 2ATP
NADH= 3ATP
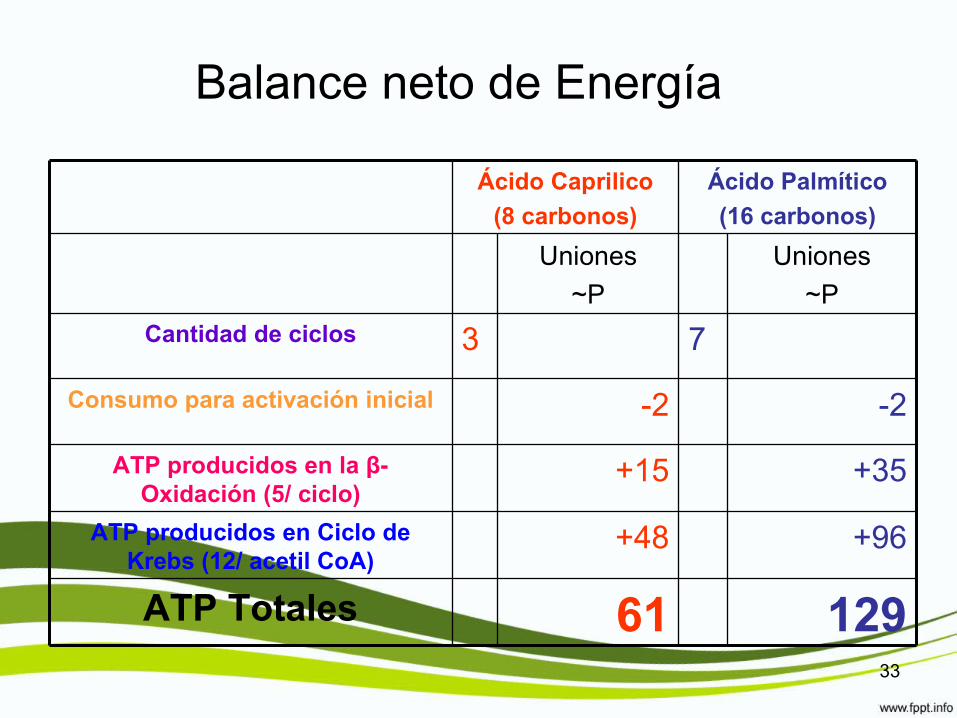
Balance neto de Energía
Ácido Caprilico
(8 carbonos)
Ácido Palmítico
(16 carbonos)
Uniones
~P
Uniones
~P
Cantidad de ciclos 3 7
Consumo para activación inicial -2 -2
ATP producidos en la β-Oxidación (5/ ciclo)
+15 +35
ATP producidos en Ciclo de Krebs (12/ acetil CoA)
+48 +96
ATP Totales 61 12933



β-oxidación de ácidos grasos insaturados
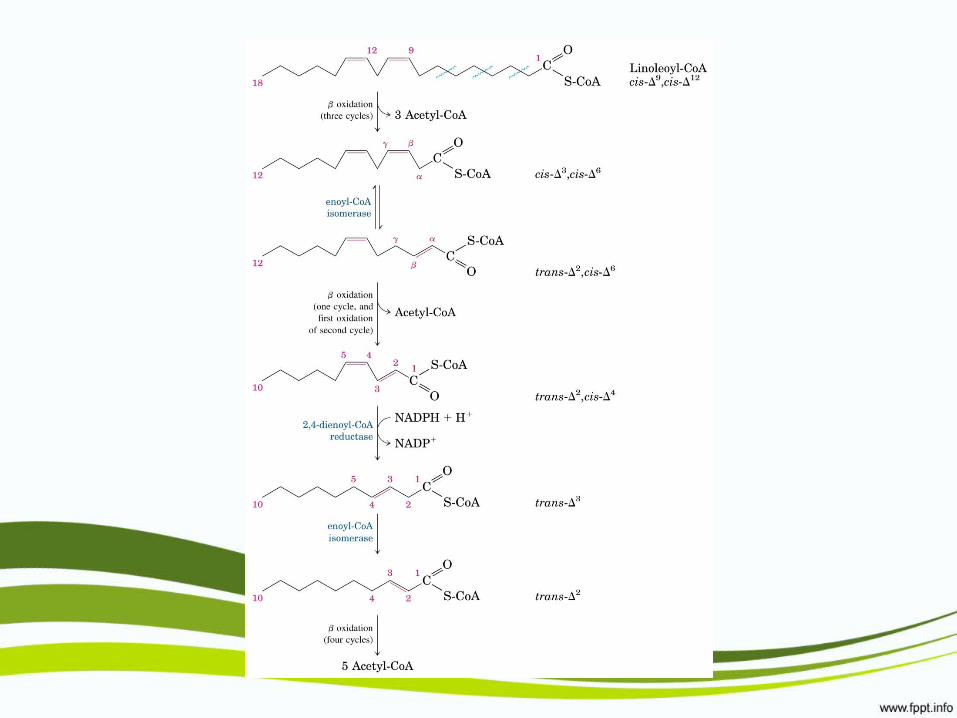

β-oxidación de ácidos grasos de cadena impar
Posible entrada en vía gluconeogénica

Formación de Cuerpos Cetónicos (Cetogénesis)
• Después de la degradación de los ac. Grasos, el Acetil-CoA es oxidado en el Ciclo de Krebs.
• Para esto es necesaria la presencia de oxalacetato (1er intermediario del ciclo de Krebs). Si la cantidad de éste es insuficiente, las unidades de acetil-CoA son utilizadas mediante una vía alternativa en la que se producen “Cuerpos Cetónicos”
• Estos compuestos se forman principalmente en el hígado, a partir de acetil-CoA mediante una serie de etapas.
H3C C
O
CH3
acetona
H3C C
O
CH2 C
O
O-
acetoacetato
H3C CH
OH
CH2 C
O
O-
3-OH-butirato
39

CETOGENESIS
1. El 1er paso es la inversa de la última etapa de la b-oxidación.
2. El acetoacetatil-CoA se condensa con otro acetil-CoA para dar HMG-CoA.
3. El HMG-CoA se desdobla formando acetoacetato y Ac-CoA.
4. El Acetoacetato origina los otros cuerpos cetónicos.
5. La conversión a β-OH-butirato depende de NADH y por ende del estado redox de la célula.
40
Ocurre en HÍGADO (mitocondria)

Utilización de los cuerpos cetónicos
• El Hígado es el principal productor ya que posee todas las enzimas necesarias para ello, pero es incapaz de usarlos como combustible.
• Los órganos que los usan son: cerebro, músculo esquelético, corazón y riñón entre otros.
• Solo se usan como fuente de energía en situaciones metabólicas especiales. Ej: Diabetes, ayuno prolongado.
• El aumento de éstos puede llevar a una Acidosis Metabólica.
41
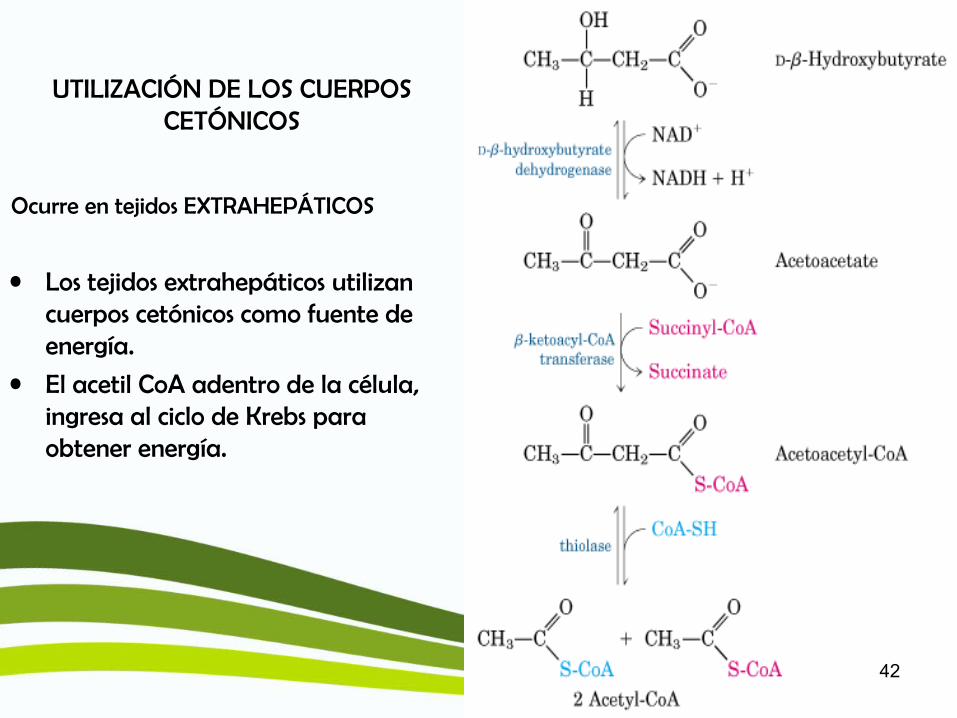
UTILIZACIÓN DE LOS CUERPOS CETÓNICOS
• Los tejidos extrahepáticos utilizan cuerpos cetónicos como fuente de energía.
• El acetil CoA adentro de la célula, ingresa al ciclo de Krebs para obtener energía.
42
Ocurre en tejidos EXTRAHEPÁTICOS

Formación y exportación de cuerpos cetónicos (hígado)
43
Los cuerpos cetónicos se forman y exportan desde el Hígado.
En condiciones energéticamente desfavorables, el oxalacetato se deriva hacia la Gluconeogénesis, para liberar glucosa a la sangre.
El ciclo de Krebs trabaja muy lentamente en el Hígado.
Gotas de lípidos
Hepatocito
Acetoacetato y β-hidroxibutirato exportados como energía para: corazón, músculo, riñón y cerebro.
Glucosa exportada como combustible para cerebro y otros tejidos.


Regulación de la cetogénesis.
1.- Movilización de lípidos de reserva
2.- Entrada de ácidos grasos activados a la mitocondria para β-oxidación (CAT-I)
3.- Disponibilidad de oxalacetato para entrada de acetil-CoA a Krebs.

Metabolismo del Colesterol
46

BIOSINTESIS DE COLESTEROL
47
La estructura de 27 C se obtiene a partir de moléculas de acetil-CoA.Se condensan moléculas de acetil-CoA para obtener ISOPRENOS activados.Principal órgano de síntesis: HIGADO
Los C negros derivan del grupo metilo del acetato.
Los C rojos derivan del carboxilo del acetato.

ETAPAS INICIALES
• Hasta la formación de β-OH-β-metil-glutaril CoA es similar a la cetogénesis aunque ocurre en citosol.
• La reducción para formar MEVALONATO es la etapa crítica que compromete a la formación de colesterol.
• Como muchos procesos biosintéticos, consume NADPH.
48
Estatinas

49
FORMACIÓN DE ISOPRENOIDES
ACTIVADOS A partir de
MEVALONATO se pueden obtener los ISOPRENOS ACTIVADOS, estos pasos implican gasto de ATP.
Los ISOPRENOS se obtienen por reacciones CABEZA-COLA.

CONDENSACIÓN DE LOS ISOPRENOS ACTIVADOS
• Los isoprenos activados reaccionan entre sí para formar ISOPRENOIDES de– 10 C: GERANIL-
PP– 15 C:
FARNESIL-PP
50

OBTENCIÓN DE ESCUALENO
51
2 moléculas de FARNESIL-PP se condensan para formar una de 30 C:ESCUALENO que se asimila a la estructura abierta del COLESTEROL.

CICLIZACIÓN DEL ESCUALENO
• El ESCUALENO se cicla para formar COLESTEROL en una serie de etapas que involucra otros esteroles como intermediarios
• Los vegetales no sintetizan colesterol
52
Hongos
Plantas
Animales

53
El colesterol se puede esterificar con ácidos grasos

Regulación de la síntesis de colesterol.
Mevalonato, colesterol

NUESTRO ORGANISMO NO DISPONE DE ENZIMAS PARA DEGRADAR EL COLESTEROL, DE MODO QUE ESTE ES EXCRETADO INTACTO O TRANSFORMADO EN SUSTANCIAS COMO LOS ÁCIDO BILIARES. EL HIGADO ES EL ÓRGANO DE ELIMINACIÓN DEL COLESTEROL DONDE BUENA PARTE SE TRANSFORMA EN ÁCIDOS BILIARES.
• ÁCIDOS BILIARES: el más abundante es el ácido cólico, en menor proporción se encuentra el ácido quenodesoxicólico.
• Son excretados en la bilis conjugados con glicina o taurina. Ej.: -ácido glicocólico
-ácido taurocólico
55
Ácido glicocólico
Ácido taurocólico

colesterol
Ácidos biliares

Ácidos biliares primarios (síntesis hepática)
•Ácido cólico•Ácido quenodeoxicólico
Ácidos biliares secundarios (síntesis intestinal)
•Ácido deoxicólico•Ácido litocólico

3 7
12

Destino de los ácidos biliares
15 % → excretados en las heces (500 mg/24 h)
15 % → reabsorbidos pasivamente a nivel del intestino
70 % → absorbidos activamente por el ileón terminal(cotransporte Na + - ácido biliar)

Circulación enterohepática de las sales biliares
Herrera, Fig.22.14

FUNCIÓN DE LOS ÁCIDOS BILIARES
• Aumentan la función de la Lipasa pancreática.
• Reducen la “Tensión Superficial” y con ello favorecen la formación de una EMULSIÓN de las grasas. Contribuyen a dispersar los lípidos en pequeñas partículas y por lo tanto hay mas superficie expuesta a la acción de la lipasa.
• Favorece la absorción de Vitaminas Liposolubles.
• Acción Colerética: estimulan la producción de bilis.
61


















