
Meiotic Nondisjunction and Recombination of Chromosome ZZZ ...
10
Copyright 0 1993 by the Genetics Society of America Meiotic Nondisjunction and Recombination of Chromosome ZZZ and Homologous Fragments in Saccharomyces cerevisiae Martin Goldway, Tamar Arbel and Giora Simchen Department of Genetics, The Hebrew University of Jerusalem, Jerusalem 91904, Israel Manuscript received May 2 1, 1992 Accepted for publication October 16, 1992 ABSTRACT A yeast strain, in which nondisjunction of chromosome ZIZ at the first meiotic division could be assayed, was constructed. Using chromosome fragmentation plasmids, chromosomal fragments (CFs) were derived in isogenic strains from six sites along chromosome ZZZ and onesite on chromosome VU. Whereas the presence of the CFs derived from chromosome ZZI increased considerably the meiosis I nondisjunction of that chromosome, the CF derived from chromosome VIZ had no effect on chro- mosome ZIZ segregation. The effects of the chromosome ZZZ-derived fragments were not linearly related to fragment length. Two regions, one of 12 kb in size located at the left end of the chromosome, and the other of 5 kb, located at the center of the right arm, were found to have profound effects on chromosome ZZZ nondisjunction. Most disomics arising from meioses in strains containing chromosome IZI CFs did notcontain the CF; thus it appears that the two chromosome IZZ homologs had segregated away from the CF. Among the disomics, recombination between the homologous chromosomes 1Zl was lower than expected from the genetic distance, while recombination between one of the chromosomes IZZ and the fragment was frequent. We suggest that there are sites along the chromosome that are more involved than others in the pairing of homologous chromosomes and that the pairing between fragment and homologs involves recombination among these latter elements. A synthesis, eukaryotic cell entering meiosis undergoes DNA by which chromosomes are duplica- ted, followed by reductional (meiosis I) and equational (meiosis 11) divisions. In meiosis I, homologous chro- mosomes pair,recombine, disjoin and segregate to opposite poles. In meiosis 11, the sister chromatids segregate from one another. From normal meiosis of a diploid cell four haploid cells (gametes) are pro- duced. Pairing of homologs initiates at the beginning of zygotene and is completed at pachytene. Initiation of pairing is associated with theappearance of early recombination nodules and the beginning of synap- tonemal complex (SC) formation (ALBINI and JONES 1987; ANDERSON and STACK 1988; CARPENTER 1987) (reviewed by ROEDER 1990). It has been proposed (CARPENTER 1987) that early recombination nodules, which are distributed almost randomly along the chro- mosome, participate in the search for homology by mediating a recombination-like interaction, and that the SCs stabilize this interaction when a long stretch of homology is found. According to electron microscopic observations, ini- tiation of pairing and SC formation in some organisms tend to begin at subtelomericregions (ALBINI and JONES 1987; JONES and CROFT 1986; HOLM and RAS- MUSSEN 1980; GILLES 1985), while in other organisms they are randomly distributed along the chromo- somes (RASMUSSEN and HOLM 1980; ANDERSON and Genetics 133: 149-158 (February, 1992) STACK 1988). Experiments using chromosome re- arrangements, such as translocations and deletions (LINDSLEY and SANDLER 1957; ROSENBLUTH and BAILLIE 1990; HERMAN and CLAIRE 1988; MCKIM, HOWELL and ROSE 1988; HAWLEY 1980), suggest that specific sites along the chromosome have a relatively high tendency to pair and recombine and that these may be the sites for initiation of pairing.In male meiosis of Drosophila melanogaster, a 12-kb single rDNA repeat from the X chromosome was found to be important for X-Y pairing (MCKEE and KARPEN In this paper we present experiments in which yeast fragmentation vectors (VOLLRATH et al. 1988) were used to add chromosomal fragments (CFs) to the Saccharomyces cerevisiae genome. The effects of the CFs on meiotic chromosome segregation and recom- bination were tested. The results suggest that specific regions along the chromosome are especially impor- tant for meiotic disjunction, through involvement in pairing. 1990). MATERIALS AND METHODS Plasmids: YCFT4: Chromosome fragmentation plasmids for the generation of CFs were all based on the vector YCFT4. This 8.3-kb long plasmid was derived from the plasmid YCM, obtained from P. HIETER (VOLLRATH et al. 1988), fromwhich the SUP11 gene was removed by replac- ing the NdeI fragment with an analogous NdeI fragment from plasmid YIp5 (BOTSTEIN et al. 1979). Six fragments,
Transcript of Meiotic Nondisjunction and Recombination of Chromosome ZZZ ...

Copyright 0 1993 by the Genetics Society of America
Meiotic Nondisjunction and Recombination of Chromosome ZZZ and Homologous Fragments in Saccharomyces cerevisiae
Martin Goldway, Tamar Arbel and Giora Simchen Department of Genetics, The Hebrew University of Jerusalem, Jerusalem 91904, Israel
Manuscript received May 2 1 , 1992 Accepted for publication October 16, 1992
ABSTRACT A yeast strain, in which nondisjunction of chromosome ZIZ at the first meiotic division could be
assayed, was constructed. Using chromosome fragmentation plasmids, chromosomal fragments (CFs) were derived in isogenic strains from six sites along chromosome ZZZ and one site on chromosome VU. Whereas the presence of the CFs derived from chromosome ZZI increased considerably the meiosis I nondisjunction of that chromosome, the CF derived from chromosome VIZ had no effect on chro- mosome ZIZ segregation. The effects of the chromosome ZZZ-derived fragments were not linearly related to fragment length. Two regions, one of 12 kb in size located at the left end of the chromosome, and the other of 5 kb, located at the center of the right arm, were found to have profound effects on chromosome ZZZ nondisjunction. Most disomics arising from meioses in strains containing chromosome IZI CFs did not contain the CF; thus it appears that the two chromosome IZZ homologs had segregated away from the CF. Among the disomics, recombination between the homologous chromosomes 1Zl was lower than expected from the genetic distance, while recombination between one of the chromosomes IZZ and the fragment was frequent. We suggest that there are sites along the chromosome that are more involved than others in the pairing of homologous chromosomes and that the pairing between fragment and homologs involves recombination among these latter elements.
A synthesis, eukaryotic cell entering meiosis undergoes DNA
by which chromosomes are duplica- ted, followed by reductional (meiosis I) and equational (meiosis 11) divisions. In meiosis I, homologous chro- mosomes pair, recombine, disjoin and segregate to opposite poles. In meiosis 11, the sister chromatids segregate from one another. From normal meiosis of a diploid cell four haploid cells (gametes) are pro- duced.
Pairing of homologs initiates at the beginning of zygotene and is completed at pachytene. Initiation of pairing is associated with the appearance of early recombination nodules and the beginning of synap- tonemal complex (SC) formation (ALBINI and JONES 1987; ANDERSON and STACK 1988; CARPENTER 1987) (reviewed by ROEDER 1990). It has been proposed (CARPENTER 1987) that early recombination nodules, which are distributed almost randomly along the chro- mosome, participate in the search for homology by mediating a recombination-like interaction, and that the SCs stabilize this interaction when a long stretch of homology is found.
According to electron microscopic observations, ini- tiation of pairing and SC formation in some organisms tend to begin at subtelomeric regions (ALBINI and JONES 1987; JONES and CROFT 1986; HOLM and RAS- MUSSEN 1980; GILLES 1985), while in other organisms they are randomly distributed along the chromo- somes (RASMUSSEN and HOLM 1980; ANDERSON and
Genetics 133: 149-158 (February, 1992)
STACK 1988). Experiments using chromosome re- arrangements, such as translocations and deletions (LINDSLEY and SANDLER 1957; ROSENBLUTH and BAILLIE 1990; HERMAN and CLAIRE 1988; MCKIM,
HOWELL and ROSE 1988; HAWLEY 1980), suggest that specific sites along the chromosome have a relatively high tendency t o pair and recombine and that these may be the sites for initiation of pairing. In male meiosis of Drosophila melanogaster, a 12-kb single rDNA repeat from the X chromosome was found to be important for X-Y pairing (MCKEE and KARPEN
In this paper we present experiments in which yeast fragmentation vectors (VOLLRATH et al. 1988) were used to add chromosomal fragments (CFs) to the Saccharomyces cerevisiae genome. The effects of the CFs on meiotic chromosome segregation and recom- bination were tested. The results suggest that specific regions along the chromosome are especially impor- tant for meiotic disjunction, through involvement in pairing.
1990).
MATERIALS AND METHODS
Plasmids: YCFT4: Chromosome fragmentation plasmids for the generation of CFs were all based on the vector YCFT4. This 8.3-kb long plasmid was derived from the plasmid YCM, obtained from P. HIETER (VOLLRATH et al. 1988), from which the SUP11 gene was removed by replac- ing the NdeI fragment with an analogous NdeI fragment from plasmid YIp5 (BOTSTEIN et al. 1979). Six fragments,

150 M. Goldway, T. Arbel and G. Simchen
from various sites along chromosome IIZ (see Figure l), and one fragment from chromosome VZI, were each cloned into the polylinker of YCFT4 in the appropriate orientation. Upon linearization and transformation, these plasmids led to the formation of chromosomal fragments extending dis- tally from the given chromosomal sites to their respective telomeres.
YCFT3: Similar to YCFT4, but with no centromere. The length of YCFT3 is 6.9 kb and it was derived from YCF3 (VOLLRATH et al. 1988) in the same way as YCFT4 was obtained from YCF4.
pMG390: A 1.4-kb HindIII-BamHI fragment, next to HML (Y. KASSIR, personal communication), was inserted between the HindIII and BamHI sites of YCFT4.
pMG391: A 2.3-kb HindIII-Hind111 fragment, harboring THR4 (GOLDWAY 1991), was inserted into the HindIII site of YCFT4.
pMG397: A 1 .9-kb HindIII-BglII fragment, harboring the 5’ region of the HIS4 gene (DONAHUE, FARABAUCH and FINK 1982), was inserted between the HindIII and BamHI sites of YCFT4.
PMG8OO: A 2.6-kb HindIII-EcoRI fragment harboring part of the D8B fragment (NEWLON et al. 1991) on the left arm of chromosome ZZI, next to the centromere, was in- serted between the HindIII and EcoRI sites of YCFT4.
pMG803: A 1.6-kb SalI-Sal1 fragment, located 3.5 kb proximal to THR4 (GOLDWAY 1991), was first inserted into the SalI site of the pUC18 polylinker, then cut with BamHI and HindIII and inserted between the HindIII and BamHI sites of YCFT4.
pMG806: A 2.6-kb HindIII fragment next to HMR (GOLD- WAY 1991) was inserted into YCFT4 at the HindIII site.
pTA705: A 1.8-kb BglII-BglII fragment with the gene CUP2 (GOLDWAY 1984) from chromosome VIZ was inserted between the BamHI and BglII sites of YCFT4.
pMG357: A 3.0-kb KpnI-ClaI fragment from plasmid pYe(CDCIO)l, containing the gene CDClO (CLARKE and CARBON 1980), was ligated into plasmid pUC18 between the sites KpnI and AccI. The derived plasmid was then digested with BamHI and EcoRI (partial digest of the latter) and a 0.85-kb EcoRI-BglII fragment carrying the gene TRPl from plasmid YRp7 (STINCHCOMB, STRUHL and DAVIS 1979) was inserted between the EcoRI and BamHI sites that are just 5’ to CDCIO. These EcoRI and BamHI sites next to CDClO are 0.25 kb apart.
pMG811: A pBR322 derivative from which the EcoRI- SalI segment was deleted and then cut with PvuII and ligated with a 2.4-kb PuuII-PuuII fragment from HIS#. The 0.7-kb HindIII-Hind111 insert in HIS4 was then replaced by a 1.1- kb HindIII fragment containing URA3.
Generation of CFs: A CF (VOLLRATH et al., 1988) is a duplication of a chromosomal segment that is attached to a vector containing a centromere, a marker and a telomere. The duplication consists of a part of a chromosomal arm running from a site of choice to the distal telomere on the same arm. Chromosome fragmentation was carried out as described (VOLLRATH et al. 1988), using the fragmentation plasmids presented above. The fragmentation plasmids de- rived from YCFT4 contain the marker URA3 and therefore the derived CFs are marked with this gene near their cen- tromeres. The plasmids were cut between the chromosome 111 (or VU)-derived segment and the subtelomeric Y’. The diploid strain 2210 was transformed with the linearized plasmids and transformant colonies with stable Ura+ phe- notype were sporulated and dissected (strains with the short CFs had mild instability). Usually, when such transformation is done, an addition of a CF to the full diploid complement is obtained rather that a substitution of one of the homologs
by the CF. In the latter cases, two spores in every tetrad are expected to be nonviable, because they do not contain the homolog of the CF. When tetrad analysis showed four viable spores in most tetrads and 2:2 segregation for uracil proto- trophy (the segregation behavior expected when a CF was added to the diploid complement), DNA was prepared from the transformants and electrophoresed (SCHWARTZ and CANTOR 1984) on a Bio-Rad CHEF I1 apparatus, to confirm the length of the CF. The gels were Southern blotted (SOUTHERN 1975) and hybridized with a S2P-labeled URA3 probe. Following each transformation, two independent transformants carrying the desired CF were selected and used in subsequent experiments.
Yeast strains: 2210: ade2-IOl/ade2-101, ura3’-52/ura3- 52, trpl A 1 /trp 1 AI, cyhZ’/CYHZ; chromosome 111 HZS4/his4- 912, leu2/LEU2, CDCIO/cdclO”::TRPl, MATalMATa, thr4/ THR4 (see Figure 2).
TRPl was deleted from its original genomic position by integrative transformation and recombination excision with plasmid pl3, which carries this deletion (HIETER et al. 1985). Subsequently, TRPl was inserted next to cdcl0’” by trans- formation with the KpnI-XbaI fragment of plasmid pMG357.
The following strains are isogenic to strain 22 10, except that each carries a different CF.
2204: The CF begins at D8B on the left arm of chromo- some IIZ and was generated by transformation with the fragmentation plasmid pMG800. In colony #2 the CF was derived from chromosome 111”, whereas in colony #5 the CF was derived from its homolog, chromosome 111‘ (Figure
2205: The CF begins at HIS4, on the left arm of chro- mosome 111 and extends to the left telomere; it was gener- ated by transformation with pMG397.
2206: Contains a short CF that begins at HML, on the left arm of chromosome 111 and extends to the left telomere; it was generated by transformation with pMG390.
2207: The CF begins 5 kb proximal to THR4 on the right arm of chromosome 111 and extends to the right telomere; it was generated by transformation with pMG803. In colony #5 the CF was derived from chromosome IZZ” and in colony #8 the CF was derived from 111’ (Figure 2).
2208: The CF begins at THR4 on the right arm of chromosome III; it was generated by transformation with pMG39 1.
2209: Contains a short CF that begins near HMR on the right arm of chromosome III; it was generated by transfor- mation with pMG806.
2202: The CF begins at CUP2 on left arm of chromosome VIZ and extends to the left telomere of that chromosome: it was generated by transformation with pTA705.
2203: This strain is also isogenic to 2210, carrying an insertion of YCFT3 at the NcoI site of the mutant gene urd-52 on chromosome V.
2212 and 2213: Derivatives of strain 2204 #5 with two changes in the genetic markers on the CF as follows. The centromere marker URA3 was replaced by the gene ADE2 by transformation with the plasmid pTA738 (ARBEL 1992) and the URA3 gene was inserted into the his4 locus by transformation with a 2.8-kb PuuII-PvuII fragment from plasmid pMG811. In 2212 the URA3 insert is into his4 on the chromosome (IZZ”) whereas in 2213 the insert is into his4 on the CF.
Media for yeast: YEPD medium (1% yeast extract 2% glucose, 2% bacto peptone, plus 1.5% agar for solid me- dium) was used for nonselective growth. For selective growth, dropout synthetic media (SHERMAN, FINK and HICKS 1986) lacking the components selected for were used.
2).

Nondisjunction and Recombination 151
SPO solid sporulation medium has been described (HICKS and HERSKOWITZ 1976). CYH medium is YEPD with 1.8 mglliter cycloheximide.
Medium for Escherichia coli: LB+Amp (1 % Bacto-tryp- tone, 0.5% yeast extract, 1% NaC1, 50 mglliter ampicillin, plus 1.5% agar for plates), was used for growth of plasmid- containing bacteria.
Preparation of chromosome-size yeast DNA molecules in solid agarose: This was done according to SCHWARTZ and CANTOR (1 984).
Monitoring nondisjunction of meiosis I: Strain 2210 was constructed so that nondisjunction of chromosome ZIZ at the first meiotic division could be monitored genetically, as follows. First, the native TRPl gene was deleted from its original location on chromosome N. Then, the gene TRPl was inserted next to the mutation cdclO". Thus, among haploid progeny, TRPl segregated as an allele of CDCIO, i.e., in each tetrad two spores were Trp+Cdc- and two were Trp-Cdc+. For detection of spore haploidization the strain was also made heterozygous for the recessive cycloheximide resistance allele c y h l . Following meiosis and sporulation, spores that formed colonies at the restrictive temperature (3 1 ") on medium lacking tryptophan and containing cyclo- heximide were expected to be haploid and disomic for chromosome ZZZ. CDClO is less than 1 cM from the centrom- ere and therefore the disomics selected as described above are expected to be the outcome of nondisjunction in meiosis I.
Quantitative assays of disomic production: Quantitative assays of disomic production were carried as follows. The strains were sporulated for 3 days at 31 ". Asci were sus- pended in a 1:25 dilution of Glusulase (Du Pont) and incu- bated at 37" for 3 hr with shaking, followed by vigorous vortexing for 30 min. The spores were then spread on plates containing cycloheximide and incubated at 3 1 O . As CYH2 and CDClO map to different chromosomes, the genes were expected to segregate independently from each other, and therefore one fourth of the plated haploid spores was ex- pected to be Cyh'Cdc+ ( c y h l CDCIO). Thus on average one spore per meiosis could form a colony under these condi- tions.
After 3 days of incubation the plates were replica plated to medium lacking tryptophan and containing cyclohexi- mide, and incubated overnight at 3 1 O . Colonies that grew at this stage were haploid and disomic for chromosome ZZZ, or diploid, homozygous for c y h l . These colonies were picked, 50 to a plate, and after a day at 3 1 " were replicated to SPO plates (sporulation medium) and later examined microscopically for their sporulation phenotypes. ala disom- ics showed aberrant sporulation (ROTH and FOCEL 1971), which was easily distinguished from normal sporulation of ala diploids, whereas ala and ala disomics were Spo-. The latter were further identified by mating to tester strains with the opposite mating types and checking their progeny for sporulation. Those that had ala disomics among their prog- eny were identified as disomics homozygous at MAT.
RESULTS
Our working hypothesis was as follows. Pairing of homologous chromosomes in meiosis I is essential for their proper segregation. Interfering with proper pairing may result in malsegregation (nondisjunction of the two homologs), giving rise to disomic ( N + 1) or nullisomic ( N - 1) products (spores). T h e presence of added CFs may interfere with pairing of the native
pMG390 pMG397 pMG8OO pMG803 pMG391 pMG806
t i t + # t I I I l l I T I
HML HIS4 LEU2 DEB CDClO MAT THR4 HMR
FIGURE 1 ."Sites along chromosome ZZZ from which clones were used as inserts in the fragmentation plasmids (indicated by arrows).
homologs from which the CFs originated. By moni- toring each CF for its effect on nondisjunction and by comparing the CFs and their effects, it should be possible to identify sites involved in the pairing of homologs.
Fragmentation of chromosome ZZZ: T h e yeast chro- mosome fragmentation system (VOLLRATH et al. 1988) was used to add CFs to the diploid strain 22 10. The fragmentation plasmids were derived from plasmid YCFT4, which is YCF4 (VOLLRATH et al. 1988) with- out the suppressor gene SUP1 1 (see MATERIALS AND METHODS). Strain 2210 was transformed with seven fragmentation plasmids (MATERIALS AND METHODS and Figure l), six for fragmentation at different sites along chromosome ZZZ and one, which served as a control, for fragmentation of the left arm of chro- mosome VZZ, at CUP%. The starting point of each CF was directed by the sequence inserted into the frag- mentation vector. Tetrad analysis of transformants that were presumed to carry CFs has shown that in most cases the CF has been added to the full diploid chromosome complement. Two transformants were chosen to represent each CF, and the segregation of the CF was tested in a limited-scale tetrad analysis (20-40 tetrads). In all cases, except for strains 2206 and 2209, the CFs segregated 2:2 in these tetrads, indicating that a single CF was present in every cell entering meiosis. T h e instability of the short CFs in strains 2206 and 2209 is shown and discussed below. T h e sizes of the CFs, as given in Table 1, were determined by separating chromosome-length DNA of the transformants by pulse-field gel electrophoresis (on a CHEF apparatus).
Effects of the chromosomal fragments on meiosis I nondisjunction: The strains with the various CFs, as well as the controls, were tested for the frequency of chromosome ZZZ disomics generated by nondisjunc- tion at meiosis I, as described in MATERIALS AND METHODS. T h e results are presented in Table 1. The rate of spontaneous nondisjunction for chromosome ZZZ was determined to be 1.05 x lo-' per meiosis. This is the average of nondisjunction values obtained from strain 22 10, which has no added CF or plasmid, and strain 2203, which is isogenic to 2210, but has plasmid YCFT3 integrated into chromosome V (YCFT3 is identical to YCFT4 except that it does not contain a centromere).
Addition of chromosome ZZZ CFs resulted in in- creased chromosome ZZZ nondisjunction. In general, the effect of the CF on nondisjunction is greatest in

152 M. Goldway, T. Arbel and G. Simchen
TABLE 1
Meiotic nondisjunction levels for strains carrying CFs
Beginning site Chromosomal CF Nondisjunction Size of
Strain of CF arm (kb) x10”
2210 1 . 1 (7/6258) 2203” 1 .o (2/2002) 2206 HML I11 L 20 5.1b (45/11298) 2205 HIS4 111 L 70 16.2 (112/6901) 2204 D8B III L 105 40.4 (255/6317) 2209 HMR I11 R 40 2.7’(33/13261) 2208‘ THR4 I11 R 110 30.7 (359/11698) 2207c THR4 + 5 kb I I I R 115 38.3 (519/13547) 2202“ CUP2 VI1 L 160 1.8 (1417767)
” Strains serving as controls: 2203 harbors the yeast fragmenta- tion vector YCFT3, integrated into chromosome V , near the gene URA3. 2202 contains a CF derived from chromosome VII.
The levels of nondisjunction for the short CFs were corrected for CF instability as shown in Table 4. For strain 2206 the observed value was corrected by multiplying it by a factor of 1/0.78, and for strain 2209 the correction factor was 1/0.91. The figures 0.78 and 0.91 are the frequencies of meioses (tetrads) which contain CFs (T2ble 4).
The rates of nondisjunction for 2207 and 2208 were compared statistically by x* test and were found to be significantly different from each other ( P < 0.01).
strains with longer CFs. For the 40-kb CF (beginning at HMR) nondisjunction occurred in 0.27% of the meiotic divisions or at 2.6 times the spontaneous rate, and for the 110-kb fragment (beginning at THR4) it occurred in 3% of the meiotic divisions, 29 times the spontaneous rate.
Two regions with exceptionally high effects were identified by comparing the effects on nondisjunction of the various CFs (Table 1). One is the 12-kb region between HML and the left telomere, and the other is the 5-kb region proximal to the gene THR4, in the middle of the right arm of chromosome ZZZ. The effect of the region adjoining the left telomere is seen by comparing strains 2206 and 2209, which contain the shortest CFs, derived from the left and right chro- mosomal arms, respectively. Although the CF in strain 2206 is half as long as the CF in the strain 2209, the frequency of disomics produced by the former was 1.9 times that produced by the latter (Table 1).
The effect of the THR4-adjacent region can be seen by comparing strains 2207 and 2208. The CF in strain 2207 is 5 kb longer than the CF in 2208, a relative increase in size of less than 5%. Nevertheless, the frequency of disomics produced by strain 2207 was 38 X lo-’, compared with a value of 30 X lop3 for strain 2208 (a significant difference by x‘ test). A detailed analysis of this 5-kb region is reported else- where (GOLDWAY et al. 1993). Briefly, it was shown to be a “hot spot” for meiotic recombination, as well as to contain a preferred site for meiosis-induced double strand breaks in DNA. The recombinogenic activity of this region and its influence on meiotic
TABLE 2
Spore viability in isogenic strains with and without a CF
No. of tetrads; viable spores per tetrad Spore
Strain 4 3 2 1 0 Total viability(%)
2210 122 65 1 1 1 0 199 88.7 2204 159 70 21 1 0 251“ 88.5
“ Not included seven tetrads in which nondisjunction for chro- mosome I11 occurred. Six of these had two viable spores disomic for chromosome 111, and one tetrad had only one viable spore, which was also disomic for chromosome I l l .
chromosome segregation were shown to be interre- lated (GOLDWAY et al. 1993).
Strain 2204, which gave the highest yield of disom- ics (Table I), was also subjected to tetrad analysis (Table 2). Out of 258 tetrads dissected, seven gave rise to chromosome ZZZ disomics. Two other strains, 2212 and 221 3, were derived from strain 2204 by inserting the gene URA3 into his4, either on one of the chromosomes ZZZ, or on the CF, respectively. Out of the 300 tetrads dissected from these two strains, ten gave rise to chromosome ZZZ disomics. Out of 17 tetrads with disomic spores (from the three strains), 13 had two viable spores per tetrad, both disomic, and four had only a single viable spore each, all of which were disomic. The absence of any viable nondisomic spores in these tetrads, indicates that the disomics have arisen from nondisjunction of the two homologs in the first meiotic division and not from precocious separation of one pair of sister chromatids (of one of the chromosome ZZZ homologs) during meiosis I. The latter would have yielded tetrads with one disomic, one nullisomic (dead) and two normal (and viable) haploid spores. The frequency of nondisjunction events obtained from the tetrad analyses (17/558) is not very different from the frequency of 4% obtained from the large-scale random-spore analysis (Table 1). CF does not cause a general failure of meiotic
disjunction: If an added CF caused general nondis- junction in meiosis I, rather than specific nondisjunc- tion of the chromosome from which it was derived, we would expect the following. (1) A CF derived from one chromosome would elevate the nondisjunction frequency of another chromosome. (2) Among tetrads of a strain carrying a CF we would find a marked increase in the frequency of tetrads with two nonvia- ble spores (nullisomic for any one of the sixteen chro- mosomes), and an increase in the frequency of tetrads with four nonviable spores (presumably because more than one pair of homologs had not properly disjoined).
As shown in Table 1, a 160-kb long CF derived from chromosome VZZ, which is longer than any of the CFs derived from chromosome 111, has no effect on the nondisjunction frequency of chromosome ZZZ (by x* test, the difference between nondisjunction values of strains 2202 and 2210 is not significant).
Nondisjunction and Recombination 153
TABLE 3
Ura+ phenotype of disomics
Frequency of Fragment Disomics
Strain size (kb) checked Ura+ disomics
(%)
2206 20 32 1 35.8 2209 40 25 1 33.1 2205 70 338 2.6 2204 105 605 3.6 2208 110 660 1.7 2207 115 572 1.9
T o examine the effect of a CF on spore viability, strain 2204, which gave the highest level of chromo- some ZZZ nondisjunction found among the strains in this work, was subjected to tetrad analysis and com- pared to the isogenic strain 22 10, which does not contain a CF (Table 2). For strain 2210, 199 tetrads were analyzed and the spore viability was 88.7%, whereas for strain 2204, spore viability was 88.5% in 251 tetrads. The distribution of viability among te- trads in strain 2204 was similar to the distribution found in strain 22 10 and no tetrads with four nonvi- able spores were found. A CF similar to the one in strain 2204, which gave a 40-fold increase in chro- mosome ZZZ nondisjunction, did not affect the meiotic disjunction of chromosomes VZZ (ARBEL 1992). From these results we conclude that the CF originating from chromosome ZZZ affects disjunction of chromosome ZZZ only and not of other chromosomes.
Presence of the chromosomal fragment among the disomics: When a CF is present in diploid cells, it is expected to migrate to one of the poles in meiosis I and be present in two sister spores in every tetrad. Usually, such segregation was indeed found. The pres- ence or absence of the fragment among the disomics was expected to shed light on its interaction with the chromosome ZZZ homologs. If the CF was not involved in pairing with the major chromosomes ZZZ, it would be expected to segregate randomly at meiosis I and to appear in 50% of the disomics. On the other hand, if the CF was involved in pairing with one of the chromosomes ZZZ, this chromosome and the CF would be expected to disjoin from each other and the result- ing disomics should not contain the CF. The presence of the CFs, which were marked with URA3, was indi- cated by growth on medium lacking uracil. With reference to the results presented in Table 3, the strains can be divided into two groups. In the first group, consisting of strains 2204, 2205, 2207 and 2208, the majority of disomics are indeed Ura-. These strains contain CFs of size 70 kb or greater. The second group includes strains 2206 and 2209 that carry short CFs of 20-kb and 40-kb, respectively, and show a high frequency of Ura+ disomics.
The low proportion of Ura+ disomics in the first group of strains indicates that the CF affects nondis-
TABLE 4
Meiotic stability of short chromosomal fragments
No. of 4-spored tetrads; Ura+:Ura- segregations Length of
Strain CF (kb) 4+:0- 3+:1- 2+:2- 1+:3- 0+:4- Total
2206 20 2 3 (8) 13 (36) 10 (28) 8 (22) 36 2209 40 0 0 20 (46) 19 (45) 4 (9) 43 2204 105 0 2 (1.5) 128 (97) l (0 .8) l (0 .8) 132
a Numbers in parentheses are percent of total.
HIS4 leu2 CDCJO MATa thr4 _I
L - I 111' 340-kb
Genetic distances: 16cM 7cM 25CM 44cM
.̂ 1 Y I 111"
his4 LEU2 MA Toc THR4 325-kb cdcJ0::TRPJ
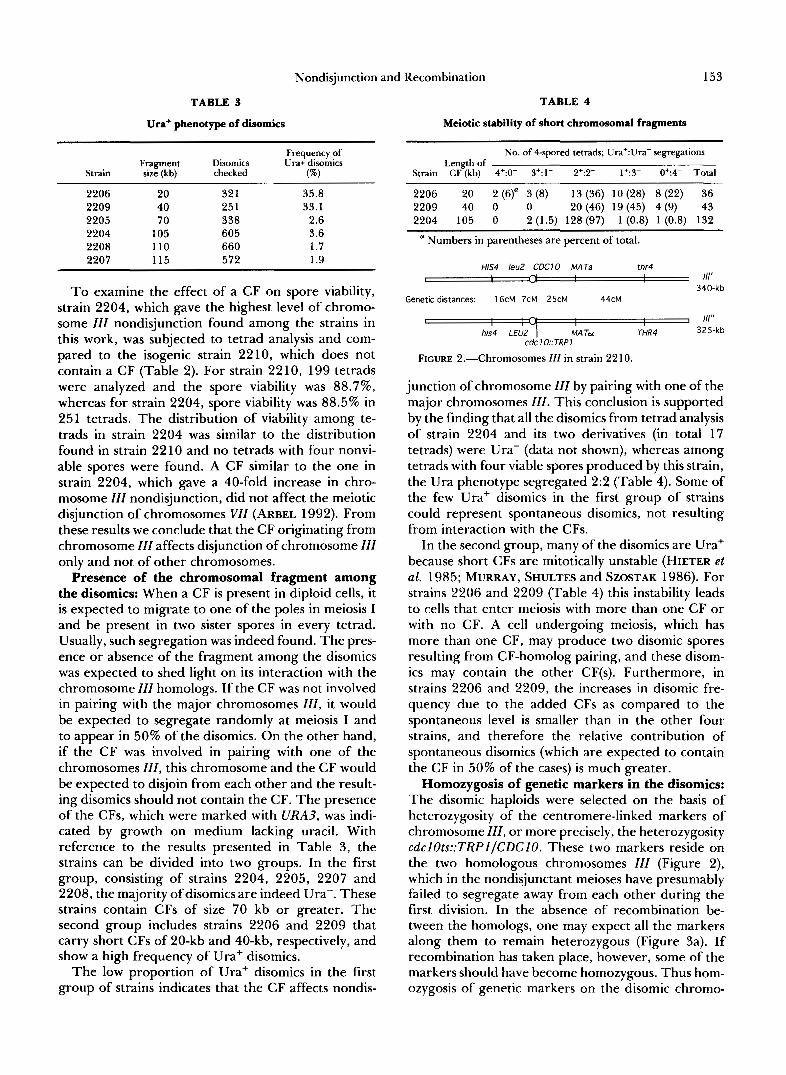
FIGURE 2.-Chromosomes III in strain 2210.
junction of chromosome ZZZ by pairing with one of the major chromosomes ZZZ. This conclusion is supported by the finding that all the disomics from tetrad analysis of strain 2204 and its two derivatives (in total 17 tetrads) were Ura- (data not shown), whereas among tetrads with four viable spores produced by this strain, the Ura phenotype segregated 2:2 (Table 4). Some of the few Ura+ disomics in the first group of strains could represent spontaneous disomics, not resulting from interaction with the CFs.
In the second group, many of the disomics are Ura+ because short CFs are mitotically unstable (HIETER et al. 1985; MURRAY, SHULTES and SZOSTAK 1986). For strains 2206 and 2209 (Table 4) this instability leads to cells that enter meiosis with more than one CF or with no CF. A cell undergoing meiosis, which has more than one CF, may produce two disomic spores resulting from CF-homolog pairing, and these disom- ics may contain the other CF(s). Furthermore, in strains 2206 and 2209, the increases in disomic fre- quency due to the added CFs as compared to the spontaneous level is smaller than in the other four strains, and therefore the relative contribution of spontaneous disomics (which are expected to contain the CF in 50% of the cases) is much greater.
Homozygosis of genetic markers in the disomics: The disomic haploids were selected on the basis of heterozygosity of the centromere-linked markers of chromosome ZZZ, or more precisely, the heterozygosity cdclOts::TRPl/CDClO. These two markers reside on the two homologous chromosomes ZZZ (Figure 2), which in the nondisjunctant meioses have presumably failed to segregate away from each other during the first division. In the absence of recombination be- tween the homologs, one may expect all the markers along them to remain heterozygous (Figure 3a). If recombination has taken place, however, some of the markers should have become homozygous. Thus hom- ozygosis of genetic markers on the disomic chromo-

154
a
M. Goldway, T. Arbel and G . Simchen
b C
MEIOSIS I
MEIOSIS II
TETRAD
N + l N + l N - 1 N - 1 N + l N + l N - 1 N - 1 FIGURE 3.-Nondisjunction at meiosis I of two homologous chromosomes in the presence of a CF derived from one chromosomal arm.
(a) No recombination between the homologs or between the homologs and the CF; the disomic products are heterozygous for the markers on the homologs. @) Recombination between the two homologs on one arm and between the CF and one of the homologs on the other arm. The fragment carries a recessive marker. Segregation at meiosis I1 leads to homozygosis of both markers. ( E ) Same recombination events as in b, but segregation at meiosis 11 leads to homozygosis of one recessive marker only, the one originating on the CF.
somes can tell us about recombination events between these homologs, and in some cases also between the homologs and the CF (Figures 3, b and c). If the two homologous chromosomes that failed to disjoin had paired and recombined normally, homozygosis of markers would have appeared among the disomics at a level expected from the genetic distances between the markers and their centromeres. Some cases of homozygosis could originate from recombination be- tween a CF carrying a recessive marker and one of the homologs. It should be noted that for all markers, except MAT, only homozygosis of the recessive alleles could be readily recognized.
In the disomics produced by the strain without a CF, 2210, the rate of marker homozygosis is lower than that expected from the genetic distances and normal meiotic recombination (Table 5); for some markers the reduction is to half and for others it is to less than a quarter of the expected level. These low homozygosis levels imply that many of the sponta- neous disomics resulted from meiotic divisions in which the two chromosomes ZZZ did not recombine and probably did not pair (see DISCUSSION).
Similar levels of homozygosis were found among
the disomics produced by the strains carrying CFs (Table 5). Two aspects of the homozygosis among disomics of these strains should be noted. First, the homozygosis frequencies on one arm seem to be re- lated to the length of the CF that was derived from the other arm (Table 5). Disomics induced by longer fragments show higher levels of homozygosis com- pared to those induced by shorter ones (compare, for instance, the frequency of thr4 homozygosis in strains 2204,2205 and 2206 in Table 5). Second, strains that contain CFs with a recessive marker, show higher frequencies of homozygosis for this marker among the disomic progeny (see Table 5 , homozygosis fre- quencies for leu2 and his4 in the two different isolates of strain 2204, and homozygosis for thr4 in strain 2207). These latter additional homozygosis events probably result from recombination between the CF and one of the homologs.
Fragment-chromosome recombination in nondis- junctant meiotic divisions: T o further analyze recom- bination between the CF and its homologs, two strains, 2212 and 221 3 were constructed, in which the two chromosomes ZZZ and the CF were differentially marked at the HIS4 locus on the left arm. Strains

Nondisjunction and Recombination 155
TABLE 5
Homozygosis of markers among disomic progeny
Added CF Homozygosis (%); marker
Arm of Homology Recessive Strain Colony origin (W' marker his4 leu.? MAT lhr4 disomics
No. of
2210
2204 2204
2205 2205
2206 2206
2207 2207
2208 2208
2209 2209
Expectedd
#2 #5
#2 #4
#5 5 #58
#5 #8
#26 #33
#5 #8
111' L 1II"L
IIIL IIIL
IIIL IIIL
111' R 1II"R
III 'R 111' R
IIIR IIIR
95 90
60 60
12 12
105 115
100 100 30 30
leu2 his4
None None
None None
thr4 None
None None
None None
4.80
7.60' 12.50'
8.90' 7.25'
6.10 6.00
5.05 4.40
3.50 6.25
3.80 1.65
11.67
0.95 4.55
3.30' 4.50 1 .OOb 3.95
0.70 4.10 0.50 3.10
1.35 1.70 0.60 2.00
0.78 4.08 0.67 3.45
0 2.20 1.25 2.33
0.75 1.50 0.83 4.15
3.48 24.0
8.15
14.10 11.85
8.20 6.75
4.10 5.35
17.12' 8.80'
5.60' 7.15'
3.80 2.50
34.0
208
301 304
146 193
147 167
257 318
34 1 320 131 120
' The lengths given do not include the vector portion of the CF. The differences between the CFs in colonies 2204 #2 and 2204 #5 result from a length polymorphism between the two original chromosomes 111 in strain 2210 (Figure 2). Likewise for the difference in length between 2207 #5 and 2207 #8. The two 2208 colonies were derived from the same chromosome 111 as 2207 #5.
These values may be slight underestimates because some recombination events could be masked by recombination with the overlapping CF (which carries the dominant allele). However, tetrad analysis has shown that such double crossovers (homolog-homolog as well as CF- homolog) are very rare (data not shown).
Recombination with the CF, which carries the recessive allele, could have contributed to these homozygosis values. Expected values, calculated on the basis of genetic distances in strain 2210 (from tetrad analysis). For any marker, the expected value is
the genetic distance between the centromere and the marker, multiplied by the probability of the homozygotes being recognized as such; for a recessive marker this probability is 0.5 and for a codominant marker (MAT) it is 1.0.
22 12 and 22 13 were derived from strain 2204, which contains a 105-kb CF that consists of most of the left arm of chromosome ZZZ, from D8B to the left telom- ere. Strain 2204 was chosen for the following con- structions because it gave a high proportion of disom- ics (Table l), thus enabling detection of meiosis I nondisjunction in tetrad analysis.
The gene URA3 on the CF in strain 2204 #5 was replaced by the gene ADEP (see MATERIAL AND METH- ODS) and subsequently URA3 was inserted into the his4 allele, either on the CF, to generate strain 2213, or on chromosome ZZZ", to generate 2212. In the latter two strains, almost all recombination events in the CEN-HIS4 interval may be detected by tetrad analysis (except 2-strand double crossovers), and many events among the random-spore disomics may also be rec- ognized.
Among disomics produced by strains 22 13 (Table 6) each chromosome IZZ recombined with the CF 50% more frequently than with its homolog, whereas in normal meiosis (analysis of 108 tetrads, not shown), each of the chromosomes ZZZ recombined with its homolog 4.5 times more frequently than with the CF. This preference for chromosome-CF recombination observed among disomics is even more striking if one takes into account the physical distance between CEN4 and his4 on the CF, which is 10 kb shorter than
TABLE 6
Recombination between the HIS4 locus and the centromere among disomics produced by strain 2213
Phenotype of disomic spore Recombination in the No. of Percent of
colonies HIM-CEN interval disomlcs total meioses
Ura-His-Ade- Chromosomes 111'-111" 1 1 20.8' Ura+His+Ade- Chromosome 111"-CF 31 29.2' Ura+His-Ade- Chromosome 111'-CF 32 30.2' Ura-His'Ade- 138 Total 212
' A crossover event involves two (out of four) chromatids which, following nondisjunction in meiosis I and segregation in meiosis 11, may either be included in the same disomic spore, or each be included in a different disomic spore. In the former case the two disomics will be Ura- His- and Ura- His', whereas in the latter the two disomics will be Ura- His+ (but each will be heterozygous for HIS4/his4). As we regard each disomic as representing an inde- pendent meiosis, the frequency of the recognizable recombinant disgmics has been multiplied by 4.
A crossover event between chromosome 111 and the CF yields one full-length chromatid with the marker URA3. One of the two disomics resulting from a nondisjunctant meiosis with such a cross- over will show the Ura+ phenotype. Therefore, the frequency of recognizable recombinant disomics has been multiplied by 2. Note that none of the disomics contained the CF (marked by ADE2 near CEN4).
the distance between CEN3 and his4 on the homologs. Similar results were obtained from the individual
nondisjunction tetrads, which contained only disomic spores. Among 10 such tetrads (7 with two viable

156 M. Goldway, T. Arbel and G. Simchen
spores each, both disomic, and 3 with one viable disomic spore), recombination between the CF and one or both homologs occurred in four tetrads, while no recombination events between the two homologs were detected. The latter events were looked for in the whole region between HIS4 and THR4, which covers about 60% of the chromosome.
The conclusion from these results is that in nondis- junctant meioses, recombination between the CF and chromosomes ZZZ is much higher than in normal meiosis, while recombination between the two homo- logs is greatly reduced. It follows that nondisjunction of chromosomes ZZZ might have resulted from pairing (and recombination) between the CF and one of the chromosomes ZZZ.
DISCUSSION
The presence of CFs in yeast cells entering meiosis occasionally leads to nondisjunction in meiosis I and to the production of disomics. This has been originally demonstrated by SUROSKY and TYE ( 1 988), who used CFs of either the entire left arm of chromosome ZZZ ( 1 20 kb) or the left arm plus 30 kb of the right arm. We find that from a basal level of spontaneous chro- mosome ZZZ nondisjunction of 1 . O - 1 . 1 X loe3 per meiosis, the added CFs derived from chromosome ZZZ result in increases of up to 4%, depending on the length and nature of the CF (Table 1) . SUROSKY and TYE (1988) report that a CF consisting of the entire left arm resulted in 4.2% meiosis I nondisjunction of chromosome ZZZ, a value similar to the one we ob- tained with strain 2204, which contains a somewhat shorter CF ( 1 05-kb) derived from the same arm. The effect of CFs on nondisjunction is chromosome-spe- cific, as demonstrated by the finding that a CF derived from chromosome VZZ does not increase nondisjunc- tion of chromosomes ZZZ (Table l) , by the lack of effect of the CF on spore viability (Table 2) and by the study of chromosomes VZZ nondisjunction that is induced by various CFs derived from chromosome VZZ but not by a chromosome ZZ-derived CF (ARBEL 1992).
We find that CFs elevate nondisjunction of the chromosomes they were derived from in a nonlinear fashion. We were able to identify two regions that have profound effects on meiotic nondisjunction, the 12 kb from HML to the telomere and the 5 kb prox- imal to THR4.
The effect of the 12-kb telomeric region is consist- ent with cytological studies (ALBINI and JONES 1987; JONES and CROFT 1986; HOLM and RASMUSSEN 1980; GILLES 1985), in which initiation of pairing and of the synaptonemal complexes was seen at subtelomeric re- gions. However, our data show no strong effect of the right subtelomeric region on nondisjunction (strain 2209). One way to explain the difference between the
effects on nondisjunction of the two short CFs is that in strain 2206, the left telomeric CF is present at a higher number of copies per cell than the right telom- eric CF in strain 2209. Short CFs are indeed more unstable than long ones. However, although the insta- bility of the CF in strain 2206 is somewhat more pronounced than in 2209 (Table 4), both strains seem to have on average one CF per meiosis.
The 5-kb region near THR4 has been analyzed elsewhere in detail (GOLDWAY et al. 1993). Besides its roles in meiosis I segregation, it was found to be a “hot spot” for meiotic recombination (that is, to re- combine more frequently than most other regions of the genome) and to contain a preferred site for meiosis-induced double strand breaks. Such double stand breaks were previously found in other locations in the yeast genome (CAO, ALANI and KLECKNER 1990; SUN et al. 1989). A plasmid carrying this region integrates into chromosome ZZZ in 2.4% of normal meioses, whereas such integrations occur in 15.5% of meioses with nondisjunction of chromosomes ZZZ (GOLDWAY et al. 1993). Thus high recombination activity (integration) and the role of this region in meiotic chromosome segregation are interrelated in this case. Another association between recom- bination and nondisjunction is found in the present study. Disomic progeny of strains carrying a CF ap- pear to have resulted from nondisjunctant meioses in which the CF has recombined with one of the homo- logs, thus interfering with pairing between the two homologs and their eventual segregation from each other.
Regions of chromosome ZZZ with pronounced effects on nondisjunction were identified only when relatively small intervals could be evaluated. This, and the dif- ferences between the effects of CFs of similar sizes (for instance, strains 2204 and 2208, Table l ) , suggest that there are several additional regions along the chromosome that are involved in pairing. These re- gions may be identified by constructing additional CFs, that will focus on short intervals, as reported here for the 5 kb proximal to THR4. We suspect that these regions will also associate with “hot spots” for meiotic recombination (GOLDWAY et al. 1993).
When evaluating the effect of a region on nondis- junction, the following points need to be considered. ( 1 ) If the effect on nondisjunction is due to a short region, it may not be detected when evaluated as a part of a relatively long one. (2) The same size region may have a greater effect when it is part of a long CF than when it is part of a short one, because of a cooperative interaction between several pairing ele- ments within the same CF (T. ARBEL, M. GOLDWAY and G. SIMCHEN, in preparation). (3) As discussed below, the relative location of the centromere on the fragment, in comparison to its “native” location on

Nondisjunction and Recombination 157
the chromosome, may also have an effect on nondis- junction.
The mechanism of nondisjunction is somewhat il- luminated by the analysis of recombination patterns among the disomics. We may assume that nondisjunc- tion could arise from failure of chromosomes to pair and/or to recombine, giving rise to nonrecombinant disomics. Alternatively, nondisjunction could arise from the functional failure of the machinery that pulls the chromosomes to the poles; such nondisjunction products could be recombinant disomics. Recombi- nation among disomics (measured by the homozygosis of recessive genetic markers) originating from spon- taneous nondisjunction was lower than would have been expected if pairing and recombination were normal. This suggests that nondisjunction was due partly to failure of the homologs to pair, and partly to malsegregation in cases in which pairing and recom- bination took place.
When CFs were added to the strains, nondisjunction was higher but the levels of recombination among the disomic spores were similar to the levels found for spontaneous nondisjunction (Table 5). Thus addition of CFs affected both mechanisms; it raised the number of meiotic divisions in which the homologs did not pair and at the same time it also raised the number of meiotic divisions with effective pairing, in which the segregational machinery failed.
The two chromosomes ZZZ in the disomics recom- bined with each other only rarely, at a level about one quarter of that expected from the frequency of recom- bination in normal meioses (Table 5). Chromosome ZZZ is a relatively short chromosome, with a linkage map of about 150 cM. The low levels of recombina- tion among the disomics imply that in many of the nondisjunctant meioses the two homologs did not recombine with each other. These data are supported by tetrad analysis of strains 2212 and 2213, which carry the long CF from D8B to the left telomere. The two chromosome ZZZ homologs in these strains are differentially marked over the interval HZS4-THR4, which covers about 90 cM of the genetic map of that chromosome. From normal-level recombination be- tween the homologs in meioses with nondisjunction one would expect to recognize almost one recombi- nation event in each tetrad (1.8 events per tetrad, only half of which may be recognized, depending on the segregation mode of the four chromatids among the two disomic products). However, among the 10 nondisjunctant tetrads (7 with two viable spores and 3 with one viable spore, all disomic, out of 300 dis- sected) none showed recombination between the hom- ologs in the HZS4-THR4 interval.
In contrast, recombination between the CF and one of the homologs appears to have been rather high in the nondisjunctant meioses. Among the disomics this
is seen in the high level of homozygosis for recessive markers that are carried by the CFs (Table 5), as well as in the transfer of a CF-implanted dominant marker to one of the homologs (Table 6). About 60% of the nondisjunctant meioses were associated with recom- bination between the CF and one of the homologs (Table 6). Recombination between the CF and one of the normal chromosomes ZZZ was similarly found to be very high (1 3 out of 2 1 tetrads) in nondisjunctant meioses in another study (SUROSKY and TYE 1988). As recombination could be scored over less than half of the length of the CF in our study, and over a somewhat longer interval in SUROSKY and TYE’S work, we may conclude that the great majority of nondis- junction events are associated with CF-homolog pair- ing and recombination. The very same meioses are also characterized by a marked reduction in recom- bination between the two homologs, from which we may further conclude that pairing between the CF and one of the chromosomes ZZZ precludes pairing and recombination between the two homologs. Subse- quently, the two unpaired chromosomes ZZZ segregate independently from each other in meiosis I, resulting in disomic products.
An alternative interpretation of the prevalence of disomics in which the chromosome ZZZ homologs did not recombine with each other is that the CF and the two homologs usually pair in meiosis as a trivalent. When the CF recombines with one of the homologs, the probability of homolog-homolog recombinatio.1 is reduced. If the two homologs fail to recombine, they are not held together by chiasmata during their seg- regation in anaphase I and therefore are likely to migrate to the same pole, resulting in meiosis I non- disjunction.
An unexplained result of our experiments was that among the disomics, recombination between the two homologs in one chromosomal arm appeared to be related to the length of homology between the CF and the other arm (Table 5). One possible explanation of this effect is that in a trivalent, with a long CF the position of the centromere of the CF is closer to the location of the centromeres of the two homologs than in a trivalent involving a short CF. Thus the long CF may induce nondisjunction of recombinant (paired) homologs by interfering with, or competing for the attachment of their centromeres to the spindle micro- tubules. T o test this explanation one needs to move the centromere on a given CF or to compare CFs of similar lengths with centromeres in different positions relative to the original centromeric location on the chromosome from which they were derived. Data pertinent to this issue will be presented elsewhere (T. ARBEL, M. GOLDWAY and G. SIMCHEN, in prepara- tion).
We thank AVI MORALI for technical assistance and SHOSHANA

158 M. Goldway, T. Arbel and G. Simchen
KLEIN for helpful comments on the manuscript. This research was supported by grant 87-00066 from the United States-Israel Bina- tional Science Foundation (BSF), Jerusalem, Israel.
LITERATURE CITED
ALBINI, S. M., and G. H. JONES, 1987 Synaptonemal complex spreading in Allium cepa and Alliumjstolosum. 1 . The initiation and sequence of pairing. Chromosoma 95: 324-338.
ANDERSON, L. K., and M. S. STACK, 1988 Nodules associated with axial core and synaptonemal complex during zygotene in Psi- lotum nudum. Chromosoma 97: 96-100.
ARBEL, T., 1992 Meiotic nondisjunction of homologous chromo- somes in the yeast S. cerevisiae. Ph.D. Thesis, The Hebrew University of Jerusalem, Israel.
BOTSTEIN, D., S. C. FALCO, S. E. STEWART, M. BRENNAN, S. CHERER, D. T . STINCHOMB, K. STRUHL and R. W. DAVIS, 1979 Sterile host yeast (SHY): a eukaryotic system of biolog- ical containment for recombinant DNA experiments. Gene 8:
CAO, L., E. ALANI and N. KLECKNER, 1990 A pathway of gener- ation and processing of double-strand breaks during meiotic recombination in S. cerevisiae. Cell 61: 1089-1 101.
CARPENTER, A. T. C., 1987 Gene conversion, recombination nod- ules and the initiation of meiotic synapsis. BioEssays 6 232- 236.
CLARKE, L., and J. CARBON , 1980 Isolation of a yeast centromere and construction of functional small cicular chromosomes. Na- ture 287: 504-509.
17-24.
DONAHUE, T. F., P. J. FARABAUGH and G. R. FINK, 1982 The nucleotide sequence of the HIS4 region of yeast. Gene 18: 47- 59.
GILLFS, C. B. , 1985 An electron microscopic study of synapto- nemal complex formation of zygotene in rye. Chromosoma 92:
GOLDWAY, M., 1984 Cloning and mapping of three copper-resist- ance genes in yeast. M.Sc. Thesis, The Hebrew University of Jerusalem, Israel.
GOLDWAY, M., 1991 Meiotic nondisjunction of chromosomes in S. cereuisiae. Ph.D. Thesis, The Hebrew University of Jerusa- lem, Israel.
GOLDWAY, M., A. SHERMAN, D. ZENVIRTH, T. ARBEL and G. SIMCHEN, 1992 A short chromosomal region with major roles in yeast chromosome ZIZ meiotic disjunction, recombination and double strand breaks. Genetics 133: 159- 169.
HAWLEY, R. S., 1980 Chromosomal sites necessary for levels of meiotic recombination in Drosophila melanogaster. 1. Evidence for and mapping the sites. Genetics 94: 625-646.
HERMAN, K. R., and K. K. CLAIRE, 1988 Recombination between small chromosome duplication and X chromosome in Caenor- habditis elegans. Genetics 121: 723-737.
HICKS, J. B., and 1. HERSKOWITZ , 1976 Interconversion of yeast mating types. I . Direct observation of the homothaly ( H O ) gene. Genetics 83: 445-462.
HIETER, P., C. MANN, M. SNYDER and R. W. DAVIS, 1985 Mitotic stability of yeast chromosomes: a colony color assay that meas- ures nondisjunction and chromosome loss. Cell 40: 381-392.
HOLM, P. B., and S. W. RASMUSSEN, 1980 Chromosome pairing,
165-175.
recombination nodules and chiasma formation int he Basidi- omycete Coprinus cinereus. Carlsberg. Res. Commun. 46: 300- 346.
JONES, G. H., and J. A. CROFT, 1986 Surface spreading of synap- tonemal complex in locusts. 11. Zygotene pairing behaviour. Chromosoma 93: 489-495.
MCKEE, D. B., and H. G. KARPEN, 1990 Drosophila ribosomal RNA genes function as an X-Y pairing site during male meiosis. Cell 61: 61-72.
MCKIM, S. K., M. HOWELL and M. A. ROSE, 1988 The effect of translocation on recombination frequency in Caenorhabditis elegans. Genetics 1 2 0 987-1001.
LINDSLEY, D. L., and L. SANDLER, 1957 The meiotic behavior of grossly deleted X chromosomes in Drosophila melanogaster. Genetics 43: 547-563.
MURRAY, A. W., N. P. SCHULTES and J. W. SZOSTAK, 1986 Chromosome length controls mitotic chromosome se- geregation in yeast. Cell 45: 529-536.
NEWLON, S. C., L. R. LIPCHITZ, I. COLLINS, A. DESHPANDE, R. J. DEVENISH, R. P. GREEN, H. L. KLEIN, T. G. PALZKIL, R. REN, S. SYNN and S. T . WOODY, 1991 Analysis of a circular deriv- ative of S. cereuisiae chromosome IZI A physical map and identification and location of ARS elements. Genetics 1 2 9 343-357.
Hereditas 93: 187-2 16. RASMUSSEN, S. W., and P. B. HOLM, 1980 Mechanics of meiosis.
ROEDER, G. S., 1990 Chromosome synapsis and genetic recombi-
ROSENBLUTH, E. R., and D. BAILLIE, 1990 Pairing for recombi- nation in LGV of Caenorhabditis elegans: a model based on recombination in deficiency heterozygotes. Genetics 124: 6 15- 625.
ROTH, R., and S. FOGEL, 1971 A system selective for yeast muta- tions defective in meiotic recombination. Mol. Gen. Genet. 112: 295-305.
SCHWARTZ, D. C., and C. R. CANTOR, 1984 Separation of yeast chromosome-sized DNAs by pulse field gradient gel electro- phoresis. Cell 37: 67-75.
SHERMAN, F., G. R. FINK and J. B. HICKS, 1986 Methods in Yeast Genetics: Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
SOUTHERN, E. M., 1975 Detection of specific sequences among DNA fragments by gel electrophoresis. J. Mol. Biol. 98: 503- 51 7.
STINCHCOMB, D. T., K. STRUHL and R. W. DAVIS, 1979 Isolation and characterization of a yeast chromosomal replicator. Nature
SUN, H., D. TRECO, N. P. SCHULTES and J. W. SZOSTAK, 1989 Double strand breaks as initiation site for meiotic gene conversions. Nature 338: 87-90.
SUROSKY, R. T., and B.-K. TYE, 1988 Meiotic disjunction of homologs in Saccharomyces cereuisiae is directed by pairing and recombination of the chromosome arms but not by pairing of the centromeres. Genetics 119: 273-287.
VOLLRATH, D., R. W. DAVIS, C. CONNELLY and P. HIETER, 1988 Physical mapping of large DNA by chromosome frag- mentation. Proc. Natl. Acad. Sci. USA 85: 6027-603 1 .
nation. Trends Genet. 6 385-389.
282: 39-43.
Communicating editor: F. WINSTON


















