
Mediating -Opioid-Initiated Heart Protection via the ... fileDepartment of Pathology (PJB), ......
27
1 Mediating -Opioid-Initiated Heart Protection via the 2 -Adrenergic Receptor: Role of the Intrinsic Cardiac Adrenergic Cell Short Title: Mediating Cardioprotection by the Intrinsic Cardiac Adrenergic Cell Ming-He Huang, Hui-Qun Wang, William R Roeske, Yochai Birnbaum, Yewen Wu, Ning-Ping Yang, Yu Lin, Yumei Ye, David J McAdoo, Michael G Hughes, Scott D Lick, Paul J Boor, Charles Y Lui, Barry F Uretsky Department of Internal Medicine, Cardiology Division (M-HH, YB, YW, YL, YY, CL, BFU), Research Histopathology Core (H-QW, N-PY), Department of Neuroscience and Cell Biology (DJM, MGH), Department of Surgery, Division of Cardiothoracic Surgery (SDL), and Department of Pathology (PJB), University of Texas Medical Branch, Galveston, Texas Department of Medicine, University of Arizona, College of Medicine, Sarver Heart Center (WRR), Tucson, Arizona Address for correspondence: Dr. Ming-He Huang, University of Texas Medical Branch, Department of Internal Medicine, 5.106 John Sealy Annex, 301 University Boulevard, Galveston, TX 77555-0553, Tel: (409) 772-2410, Fax: (409) 772-4982, Email: [email protected] Page 1 of 27 Copyright Information Articles in PresS. Am J Physiol Heart Circ Physiol (March 16, 2007). doi:10.1152/ajpheart.01195.2006 Copyright © 2007 by the American Physiological Society.
-
Upload
dinhkhuong -
Category
Documents
-
view
214 -
download
0
Transcript of Mediating -Opioid-Initiated Heart Protection via the ... fileDepartment of Pathology (PJB), ......

1
Mediating δ-Opioid-Initiated Heart Protection via the β2-Adrenergic Receptor:
Role of the Intrinsic Cardiac Adrenergic Cell
Short Title: Mediating Cardioprotection by the Intrinsic Cardiac Adrenergic Cell
Ming-He Huang, Hui-Qun Wang, William R Roeske, Yochai Birnbaum, Yewen Wu, Ning-Ping
Yang, Yu Lin, Yumei Ye, David J McAdoo, Michael G Hughes, Scott D Lick, Paul J Boor,
Charles Y Lui, Barry F Uretsky
Department of Internal Medicine, Cardiology Division (M-HH, YB, YW, YL, YY, CL, BFU),
Research Histopathology Core (H-QW, N-PY), Department of Neuroscience and Cell Biology
(DJM, MGH), Department of Surgery, Division of Cardiothoracic Surgery (SDL), and
Department of Pathology (PJB), University of Texas Medical Branch, Galveston, Texas
Department of Medicine, University of Arizona, College of Medicine, Sarver Heart Center
(WRR), Tucson, Arizona
Address for correspondence: Dr. Ming-He Huang, University of Texas Medical Branch,
Department of Internal Medicine, 5.106 John Sealy Annex, 301 University Boulevard,
Galveston, TX 77555-0553, Tel: (409) 772-2410, Fax: (409) 772-4982, Email:
Page 1 of 27
Copyright Information
Articles in PresS. Am J Physiol Heart Circ Physiol (March 16, 2007). doi:10.1152/ajpheart.01195.2006
Copyright © 2007 by the American Physiological Society.

2
Abstract
Stimulation of cardiac β2-adrenergic receptor (β2-AR) or δ-opioid receptor (DOR) exerts a
similar degree of cardioprotection against myocardial ischemia in experimental models. We
hypothesized that δ-opioid-initiated cardioprotection is mediated by the intrinsic cardiac
adrenergic (ICA) cell via enhanced epinephrine release. Using immunohistochemical and in situ
hybridization methods, we detected in situ tyrosine hydroxylase (TH) mRNA and TH
immunoreactivity which was co-localized with DOR immunoreactivity in ICA cells in human
and rat hearts. Western blot analysis detected DOR protein in ICA cells isolated from rat
ventricular myocytes. The physiology of DOR expression was examined by determining changes
of cytosolic [Ca2+]i transients in isolated rat ICA cells using fluorescence spectrophotometry.
Exposing the selective δ-opioid agonist DPDPE to ICA cells increased [Ca2+]i transients in a
concentration-dependent manner. Such an effect was abolished by the Ca2+ channel blocker
nifedipine. HPLC-electrochemical detection demonstrated a 2.4-fold increase in epinephrine
release from ICA cells following DPDPE application. The significance of ICA cell and its
epinephrine release in δ-opioid-initiated cardioprotection was demonstrated in the rat myocardial
infarction model and ICA cell-ventricular myocyte co-culture. DPDPE administered before
coronary artery occlusion or simulated ischemia-reperfusion reduced left ventricular infarct size
by 54±15% or myocyte death by 26±4%, respectively. β2-AR blockade markedly attenuated δ-
opioid-initiated infarct-size-limiting effect and abolished δ-opioid-initiated myocyte survival
protection in rat ICA cell-myocyte co-culture. Furthermore, δ-opioid agonist exerted no myocyte
survival protection in the absence of co-cultured ICA cells during ischemia-reperfusion. We
conclude that δ-opioid-initiated myocardial infarct-size-reduction is primarily mediated via
endogenous epinephrine-β2-AR signaling pathway as a result of ICA cell activation.
Key Words: β2-adrenergic receptor, delta opioid receptor, epinephrine, ICA cell, myocardial
ischemia
Page 2 of 27
Copyright Information

3
INTRODUCTION
Stimulation of β2-adrenergic receptor (β2-AR) before myocardial ischemia confers
cardioprotection via infarct-size-reduction (13, 14) and facilitated ventricular contractile
recovery (25) during ischemia-reperfusion in experimental models. Numerous studies indicate
that δ-opioid agonists precondition the heart against ischemia (2, 7, 19, 22, 23). The extent of
heart protection conferred by δ-opioids is comparable to those elicited by myocardial β2-AR
stimulation. It has been suggested that δ-opioid-initiated cardioprotection is partially mediated by
β2-AR in the murine heart (20). Although δ-opioid agonists exert potent cardioprotection, it is
unclear which type of cardiac cells actually express δ-opioid receptor (DOR). Despite receptor-
binding study of DOR (11, 27, 33) and its gene expression in human and animal hearts (2, 28),
no clear immunohistochemical evidence has demonstrated in situ DOR expression in cardiac
myocytes. The intrinsic cardiac adrenergic (ICA) cell is a neuroendocrine cell that expresses
genes and enzyme proteins for epinephrine biosynthesis (5, 9, 10). ICA cells possess an oxygen-
sensing function suggesting an intimate involvement in ischemia-reperfusion (10). In the present
study, we have hypothesized that ICA cells express DOR mediating ICA cell function. We have
also hypothesized that ICA cells participate in δ-opioid-initiated cardioprotection against
ischemia. We examined the following objectives: 1) ICA cells express DOR immunoreactivity in
human and rat hearts; 2) DOR regulates ICA cell function via [Ca2+]i transients modulation; 3) δ-
opioid stimulation enhances endogenous epinephrine release from isolated ICA cells; and 4) δ-
opioid-initiated cardioprotection depends on ICA cell activation via enhanced epinephrine
liberation and consequent myocardial β2-AR stimulation during ischemia-reperfusion in vivo and
in vitro.
Page 3 of 27
Copyright Information

4
MATERIALS AND METHODS
Immunohistochemical studies: Immunohistochemical labeling of ICA cells was performed on 4-
µm paraffin sections of 4% formaldehyde-fixed cardiac tissue. Human heart tissues (n=6) were
obtained from recipient’s hearts during heart transplant surgery (n=4) or autopsy (n=2). For rat
(Sprague-Dawley) immunohistochemical studies, 4 adult rat hearts were used. The protocols for
using human and animal tissue were approved by the Institutional Review Board and the
Institutional Animal Care and the Use Committee of the University of Texas Medical Branch.
Human tissues were taken from left ventricular (LV) free wall, sinoatrial (SA) and
atrioventricular (AV) nodal regions. Immunoperoxidase (9) and immunofluorescent labeling was
performed with antibodies against tyrosine hydroxylase (TH) and phenylethanolamine-N-
methyltransferase (PNMT), markers of ICA cells (5, 9). The dilutions for mouse anti-human TH
(Neuromics, Northfield, MN) and mouse anti-rat TH (ImmunoStar Inc, Hudson, WI) were 1:40.
The dilution for rabbit anti-human PNMT (ImmunoStar Inc, Hudson, WI ) was 1:500. To co-
localize DOR and TH immunoreactivity, immunofluorescent double labeling methods were used.
The concentrations for rabbit anti-human DOR (US Biological, Swampscott, MA) and rabbit
anti-rat DOR (Calbiochem, San Diego, CA) were 1:200 and 1:250, respectively. The specificity
of mouse anti-TH and rabbit anti-DOR antibodies was tested by substituting these antibodies
with Universal Negative Controls for Mouse and Rabbit IgG (DAKO Corporation, Carinteria,
CA), respectively. Immunofluorescent double labeling was also used to determine whether ICA
cells express neuronal marker PGP 9.5 or muscle marker myosin-heavy-chain. The dilutions for
PGP 9.5 (Chemicon International Inc., Temecula, CA) and myosin-heavy-chain (Abcam Inc,
Cambridge, MA) were 1:3000 and 1:500, respectively. The double staining included four steps:
(1) rabbit anti-DOR served as the first primary antibody and was stained with goat anti-rabbit
Alexa Fluor 594 followed by amplification with donkey anti-goat Alexa Fluor 594; (2) slides
were then incubated with biotin-labeled goat anti-rabbit for 30 min to saturate unbound rabbit
IgG; (3) mouse anti-TH served as the second primary antibody and was stained sequentially with
rabbit anti-mouse Alexa Fluor 488 and goat anti-rabbit Alexa Fluor 488 (Signal-Amplification
Kit for mouse antibodies, Molecular Probes, Inc., Eugene, OR). A control slide with omitted
mouse anti-TH treatment was stained with Streptavidin-Alexa Fluor 488 after step (2) to test
possible cross-reaction between goat anti-rabbit biotin used in step (2) and rabbit anti-DOR
Page 4 of 27
Copyright Information

5
antibody used in step (1); (4) Slides were counterstained with DAPI. Double labeling of TH with
PNMT or TH with PGP 9.5 was performed in the same fashion. Image-IT TM FX Signal
Enhancer (Molecular Probes, Inc., Eugene, OR) and autofluorescence eliminator reagent were
used before and after staining to block nonspecific background or autofluorescence.
Immunohistochemical co-localization of TH and DOR immunoreactivity was performed in rat
hearts and in isolated rat ICA cells. Finally, to determine whether both ICA cells and ventricular
myocytes express DOR, we performed simultaneous immunofluorescent double detection of
immunoreactivity of DOR (rabbit anti-DOR diluted in 1:600) and myosin-heavy-chain (mouse
anti-myosin-heavy-chain diluted in 1:1000) in dissociated and mixed rat ventricular cardiocytes.
Detection of TH mRNA in human ICA cells by in situ hybridization: In situ hybridization was
performed on two human LV tissue samples. TH oligonucleotide probes were synthesized
(Sigma-Genosys, St. Louis, MO) per the published sequence (accession number: NM 012740).
The antisense sequence was GCATAGTTCCTGAGCT TGTCCT, and sense was
CGTATCAAGGACTCGAACAGGA. Both were labeled with fluorescein at 5'. Paraffin sections
of LV were rehydrated and treated in a microwave oven at 1000C for 6 min then postfixed with
2% paraformaldehyde followed by graded dehydration. FITC-labeled TH mRNA oligonucleotide
probes of sense and antisense were applied at the same concentration and sealed with Hybri-Well
Press-Seal Hybridization Chambers (Sigma). Hybridization was performed at 850C for 5 min
then 2.5 hr at 500C in the HybriteTM (Vysis, Inc; Downers Groove, IL). After post-hybridization
wash, goat anti-FITC was applied followed by donkey anti-goat IgG Alexa Fluor 594. The slides
were treated with Image-ITTM Enhancer before applying mouse anti-human TH antibody used
for labeling TH immunoreactivity. Dual detection of chicken anti-mouse Alexa Fluor 488 and
rabbit anti-mouse Alexa Fluor 488 was performed.
[Ca2+]i recording in isolated ICA cells: Rat ICA cell isolation was made possible by the original
observation that live ICA cells but not other cardiocytes preferentially bind to iron oxide-based
paramagnetic beads after they have been dissociated from myocardial tissue (9). Ventricular
cardiocytes were enzymatically dissociated and resuspended in 4 ml Tyrode’s solution
containing 50-µl paramagnetic beads that were not coated with any primary antibody
(Polysciences Inc., Warrington, PA). ICA cells attached to the beads were recovered by magnetic
Page 5 of 27
Copyright Information

6
separation and cultured for 48-72 hr. The cultured ICA cells were loaded with 4 mmol/L fura 2-
AM for 30 min. A ratio-based microscopic fluorescent spectrometer was used to measure the
[Ca2+]i transients generated by ICA cells (10).
Western blot analysis and epinephrine assay: To determine whether both ICA cells and
ventricular myocytes express DOR protein, freshly isolated rat ICA cells and ventricular
myocytes with depletion of ICA cells were lysed separately to extract their protein. Protein of rat
brain tissue served as a positive control. Western blot analysis was performed as described (10).
The concentration for rabbit anti-DOR was 1:500. In vitro epinephrine release assay was
performed in isolated rat ICA cells. ICA cells isolated from adult rat hearts were pooled and
plated on 12-well culture plates (2 hearts /well) and cultured for 72 hr. Epinephrine release from
ICA cells in culture was determined using an HPLC-electrochemical detection system as
described (4).
Experimental Design
DOR mechanism: We studied the effects of a potent selective δ1-opioid agonist [D-Pen25]-
enkephalin (DPDPE) on [Ca2+]i transients generated by isolated ICA cells in culture. After
obtaining baseline [Ca2+]i transients of ICA cells for 15 min, DPDPE at different doses (0.1
nmol/L, 1 nmol/L, 10 nmol/L, and 100 nmol/L) was administered in random order to the ICA
cell for 15 min. Cells were then washed for 20 min with continuously recording [Ca2+]i transients
until the activity reached a plateau. The specificity of DPDPE was examined by exposing ICA
cells to the DOR antagonist naltrindole (10 µmol/L) for 10 min followed by application of
DPDPE (100 nmol/L) in the continuous presence of naltrindole for 15 min. To determine
whether the L-type Ca2+ channels were responsible for altered [Ca2+]i transients following δ-
opioid stimulation, the effects of DPDPE on ICA cells in the presence of the L-type Ca2+ channel
blocker was tested by applying nifedipine (1 µmol/L for 15 min) to the cells when the
enhancement in [Ca2+]i was elicited with DPDPE.
Effect of δ-opioid agonist on epinephrine release: After 72 hr in culture, ICA cells were switched
to 500 µL of Tyrode’s solution and incubated for 1 hr. The sample solution was collected for
basal epinephrine release. Then the culture wells were treated for 30 min with 500 µl Tyrode’s
Page 6 of 27
Copyright Information

7
solution with or without DPDPE (100 nmol/L). Conditioned solutions were collected at the end
of 30 min and the culture wells were washed 3 times. After washing, 500 µl of Tyrode’s solution
was reintroduced to the culture wells for 1 hr (recovery phase). The samples were recollected and
the cells were lysed for protein quantification.
In vivo myocardial ischemia study: The rat myocardial infarct model has been described
previously (30). Briefly, following anesthesia and ventilation support, the chest is opened and the
left coronary artery encircled with a suture and ligated for 30 min. The snare is then released and
myocardium reperfused for 4 hr. The left coronary artery is then reoccluded and Evan's blue dye
injected into the right ventricle. The LV is sliced into 6-7 sections. Tissue slices are incubated for
10 min in 1% 2,3,5-triphenyl-tetrazolium-chloride (TTC), fixed in 10% formaldehyde, and
photographed to identify the ischemic myocardium at risk (uncolored by the blue dye), the
necrotic zone (unstained by TTC) and the nonischemic zones (colored by blue dye). The areas of
ischemia and necrosis in each slice are determined by planimetry, converted into percentages of
the whole for each slice, and multiplied by the weight of the slice. The examiner who analyzed
infarct size was blinded to treatment assignment. To demonstrate δ-opioid-initiated
cardioprotection, DPDPE (200 µg/kg) was intravenously infused 30 min before the coronary
artery occlusion (n=9). To determine whether the infarct-size-limiting effect of DPDPE
depended on endogenous epinephrine, the nonselective AR antagonist labetalol (2 mg/kg) was
intravenously infused over 30 min followed by DPDPE at 30 min before the coronary artery
occlusion (n=9). To determine the relative contribution of β2-AR in δ-opioid-initiated infarct-
size-reduction, the effect of DPDPE in the presence of the β2-AR antagonist ICI-118,551 (ICI,1
mg/kg) was tested (n=9). ICI was intravenously infused over 30 min followed by DPDPE
infusion at 30 min before coronary artery occlusion. The effects of saline, labetalol or ICI alone
on infarct size were determined (n= 8/group). The specificity of DPDPE was tested in the
presence of the DOR antagonist naltrindole (NTI, 200 µg/kg, n=5).
In vitro simulated ischemia in ventricular cardiocytes: To confirm the role of ICA cells in δ-
opioid-initiated cardioprotection in vitro, we utilized a simulated ischemia-reperfusion model
(18) of adult rat ICA cell-ventricular myocyte co-culture. This model allowed us to determine
ICA cell-ventricular myocyte interactions in the absence of sympathetic nerve innervation. ICA
Page 7 of 27
Copyright Information

8
cell-myocyte co-culture was prepared by directly plating the dissociated ventricular cardiocytes
to 24-well plates for 24 hr without undergoing magnetic ICA cell purification. Ischemia was
induced by layering mineral oil (0.5 ml for 2 hr) over a thin film of hypoxic media (pre-bubbled
with N2 gas) covering the cells followed by 60 min of "reperfusion" in normal Tyrode’s solution.
The treatments included 1) saline, 2) labetalol (1 µmol/L), 3) ICI (1 µmol/L), 4) NTI (10
µmol/L), 5) DPDPE (100 nmol/L), 6) labetalol+DPDPE, 7) ICI+ DPDPE, and 8) NTI+DPDPE.
Saline, labetalol, ICI, NTI, and DPDPE were applied individually to the cell culture for 30 min
before ischemia. For drug combination treatment, labetalol, ICI and NTI were added
individually10 min prior to the application of DPDPE. Finally, we tested whether DPDPE
exerted any myocyte protection in the absence of co-cultured ICA cells. The ICA cell-free
myocytes were prepared by depleting the ICA cells from dissociated mixed ventricular
cardiocytes with the magnetic purification method. This ICA cell depletion procedure was
repeated 4 times to ensure maximum effect. Myocyte death was quantified by counting trypan
blue-stained cells and expressed as a percentage of the total cells counted. All experiments (n= 5
rats/group) were carried out at 37°C. The examiner who counted cells was blinded to the
treatment assignment.
Data analysis: For semi-quantification of ICA cells in human LV tissue, ICA cells were defined
as cells displaying TH immunofluorescence. Those cardiocytes with nuclei stained with DAPI
(blue fluorescence) without TH signal were classified as non-ICA cells. The percentage of
ICA/non-ICA cells was counted from 20 consecutive views (100X lens) for each slide. At least 3
slides were examined for each LV sample. Data are presented as mean ± SD. For quantification
of [Ca2+]i transients, the firing frequency of [Ca2+]i spikes of ICA cells was determined based on
an average of the last 5-min duration for the baseline and each drug treatment. The outcome
measure was the number of spikes/min. ANOVA was used for analyzing changes in [Ca2+]i
transients, epinephrine release, and myocardial ischemia studies with the Bonferroni adjustment
for intra-group comparisons. Paired t-test was used to determine the effects of naltrindole and
nifedipine on DPDPE-induced [Ca2+]i changes. The significance level α is 0.05.
Page 8 of 27
Copyright Information

9
RESULTS
Identification of human ICA cells ICA cells were identified in human hearts (Figure 1). In situ
expression of TH mRNA was identified in ICA cells but not in the sympathetic nerve endings
(Figure 2). ICA cells expressed TH, PNMT, and PGP 9.5 immunoreactivity (Figure 2) and were
distributed diffusely throughout the LV. Perivascular distribution was a common feature.
Abundant ICA cells were also observed in the smooth muscle layer of epicardial coronary artery
(Figure 1E). The density of ICA cells varied considerably constituting as many as 20% to as little
as 0% of total cardiocytes/microscopic view (100X lens) with an average of 7±2%. ICA cells
were identified in the SA (Figure 1G) and AV nodal regions (data not shown).
Identification of DOR in ICA cells DOR immunoreactivity was exclusively co-localized with
TH in human and rat ICA cells (Figures 3 and 4). Ventricular myocytes expressed myosin-
heavy-chain immunoreactivity (Figure 4G). Over 90% of isolated ICA cells co-expressed TH
and DOR immunoreactivity yielding extremely high ICA cell purity. DOR-positive ICA cells
constituted approximately 13±4% of total rat ventricular cardiocytes based on the cell counting.
No DOR immunoreactivity was identified in human and rat ventricular myocytes in situ and in
vitro. No immunoreactivity was detected in IgG control slides. The control slides for double
labeling that was stained with only Streptavidin-Alexa Fluor 488 after step 2 showed no cross-
reactivity to DOR confirming immunohistochemical specificity of DOR expression in ICA cells.
Western blot analysis detected DOR protein in rat ICA cell isolates, which was identical to that
found in rat brain. No DOR protein was detected in ventricular myocytes in the absence of ICA
cells following ICA cell depletion (Figure 4H).
Modulation of [Ca2+]i transients and epinephrine release of rat ICA cells Application of
DPDPE to ICA cells increased [Ca2+]i transient spikes (n=11 cells) in a concentration-dependent
manner (Figure 5A). Increases in [Ca2+]i transients were sometimes preceded by a brief
quiescent phase. DPDPE (100 nmol/L) did not change [Ca2+]i transient activity significantly in
the presence of the DOR antagonist naltrindole (10 µmol/L, n=4, p<0.01). DPDPE-enhanced
[Ca2+]i transients were abolished in the presence of nifedipine (1 µmol/L, n=4, p<0.01). Basal
endogenous epinephrine release was detected from isolated ICA cells in culture. Application of
Page 9 of 27
Copyright Information

10
DPDPE (100 nmol/L) increased epinephrine release 2.4-fold (p<0.01, n=4 duplicates from 8 rats,
Figure 5B). Norepinephrine was not detected during the baseline, DPDPE’s treatment or during
the recovery phase.
δ-opioid-initiated myocardial and myocyte protection: Intravenous infusion of DPDPE prior to
coronary artery occlusion reduced LV infarct size by 54±15% compared to control (Figure 6A).
Infusion of labetalol prior to ischemia tended to increase (statistically insignificant) the infarct
size compared to control. DPDPE-initiated infarct-size-reduction was markedly attenuated in the
rats pretreated with labetalol. The β2-AR blocker ICI alone significantly increased infarct size
compared to control. The infarct size following DPDPE infusion in the presence of labetalol or
ICI was not significantly different than those treated with saline infusion. There was no
significant difference in body and LV weight among animal groups. There was a small reduction
in arterial blood pressure in the rats following labetalol infusion
In vitro simulated ischemia-reperfusion markedly increased myocyte death compared to the non-
ischemic state (p<0.001). Pretreatment of cardiocytes with DPDPE reduced ischemia-induced
cell death by 26±4% (p<0.001, n=5 rats). In the presence of labetalol, ICI, or NTI, DPDPE no
longer exerted protection on ischemia-induced myocyte death (Figure 6B). In myocytes in the
absence of co-cultured ICA cells, ischemia resulted in 95±2% myocyte death compared to that of
56±3% in ICA cell-ventricular myocyte co-culture (p<0.001). Application of DPDPE prior to the
ischemia exerted no myocyte protection during subsequent ischemia-reperfusion in ventricular
myocytes in the absence of co-cultured ICA cells (Figure 6C). A 48% and 70% increase
(p<0.001) in myocyte death was seen at rest and during ischemia-reperfusion, respectively, in
myocytes without co-cultured ICA cells, compared to those with co-cultured ICA cells.
Page 10 of 27
Copyright Information

11
DISCUSSION
We have demonstrated DOR expression in ICA cells in human and rat hearts. Consistent with
immunohistochemical findings, the physiological evidence provided herein demonstrates that
[Ca2+]i transients generated by ICA cells can be modified by δ-opioid stimulation. Activation of
ICA cells enhances endogenous epinephrine release and β2-AR stimulation that is essential for
the δ-opioid-initiated myocardial protection against ischemia.
ICA cells in the human heart: Human ventricular ICA cells express TH mRNA that encodes the
rate-limiting enzyme for epinephrine biosynthesis. The abundant ICA cells present in the SA and
AV nodes is consistent with an animal study reporting an intimate anatomic relationship between
ICA cells and cardiac pacemaker and conduction systems (5) suggesting a regulatory role of ICA
cells in impulse generation and conduction in human heart. The presence of ICA cells in
transplanted human LV myocardium (Figure 1H) is consistent with the concept that ICA cells
may provide an alternative adrenergic supply in the compromised sympathetic innervation (8).
This notion has been substantiated by the finding that the epinephrine producing gene PNMT
mRNA is upregulated in ventricular tissue of transplanted human hearts (6).
Cellular origin of DOR: The present study has located DOR immunoreactivity exclusively to the
ICA cells in human and rat hearts. Recently, Patel et al (18) reported DOR immunoreactivity in
the T-tubules of isolated rat ventricular myocytes in culture. However, the lack of corroborative
immunohistochemical evidence of in situ DOR immunoreactivity in rat myocardial tissue section
in that study (18) has made their claim inconclusive. The unequivocal in situ and in vitro
immunohistochemical evidence of DOR expression by ICA cells (Figures 3 and 4) is consistent
with our Western blot analysis demonstrating abundant DOR protein expression by ICA cells but
not myocytes (Figure 4H).
DOR signaling in ICA cells: Ca2+ influx is a fundamental mechanism for neurotransmitter
release from neuroendocrine cells (15). Increase in [Ca2+]i transients in ICA cells following δ-
opioid stimulation was mediated through Ca2+ influx via L-type Ca2+ channels, since the increase
in [Ca2+]i was abolished in the presence nifedipine (Figure 5). The cAMP-PKA pathway is the
Page 11 of 27
Copyright Information

12
best-characterized mechanism underlying opioid signaling in catecholaminergic neurons in the
locus ceruleus (12). Opioid exposure to these neurons up-regulates cAMP-pathway resulting in
increased Ca2+ influx (12, 16). DOR stimulation also activates cAMP pathway in olfactory bulb
neurons (17) and CHO cell line expressing human DOR (21). PKA phosphorylates L-type Ca2+
channels and enhances Ca2+ influx (26). The dependence of δ-opioid-enhanced [Ca2+]i transients
on L-type Ca2+ channels in ICA cells suggests an involvement of the cAMP-PKA pathway.
Mechanism involved in δ-opioid-initiated cardioprotection: We have demonstrated that δ-opioid-
initiated cardioprotection depends on ICA cells activation. Unlike sympathetic nerve endings that
release norepinephrine (a potent β1-AR agonist with negligible β2-AR effect), ICA cells release
epinephrine that is a potent endogenous β2-AR agonist. This is of particular importance since β2-
AR is essential in mediating cardioprotection against ischemia (14, 25) presumably via its anti-
apoptotic effect (3, 31). The increased infarct size in the presence of β2-AR blockade compared
to saline or labetalol (Figure 6A) is probably due to a fact that selective β2-AR blockade creates
unopposed β1-AR stimulation as a result of sympathetic nervous activation. Myocardial β1-AR
stimulation exerts detrimental proapoptotic effect (3, 31) hence increased infarct size. It appears
that β2-AR blockade did not completely abolish DPDPE-initiated infarct-size-limiting effect in
vivo, although this incompleteness is not statistically significant. Nevertheless, this incomplete
infarct-size-limiting effect is unlikely due to a direct δ-opioid effect on myocytes, as DPDPE-
initiated myocyte protection is lacking in the presence of β2-AR blockade in ICA cell-myocyte
co-culture or in myocytes in the absence of co-cultured ICA cells (Figure 6 B and 6C). It is well
known that systemic δ-opioid infusion elicits complex neuronal responses resulting in secondary
cardiovascular effects via non-adrenergic pathways. For instance, δ-opioid stimulation of sensory
neurons in dorsal root ganglia increases release of calcitonin gene-related peptide (1), a
neuropeptide exerting infarct-size-limiting effect during ischemia-reperfusion (29). It might also
be possible that enhanced epinephrine release from ICA cells might activate β-AR located on the
cardiac afferent nerve endings eliciting cardiocardiac reflex resulting in secondary cardiac effects
(24). Such secondary effects were not present in ICA cell-ventricular myocyte co-culture model
allowing direct assessment of δ-opioid signaling mechanisms in ICA cells. It is interesting to
note that in the absence of co-cultured ICA cells, ~95% myocytes died during ischemia-
Page 12 of 27
Copyright Information

13
reperfusion, suggesting a crucial role of ICA cells in maintaining myocyte vitality during
hypoxia. This finding is consistent with a murine study that targeted disruption of TH gene
causes lethal cardiomyopathy and fetal demise in the uterus where the hypoxic distress is
frequently encountered (32).
Limitations of the study: We acknowledge several limitations in the present study. The 7% of
ICA cells identified in human LV samples is relatively low compared to 13% in rat LV samples.
Since the human samples were largely derived from diseased hearts with end-stage heart failure,
it might not truly represent the percentage of ICA cells in normal human heart. The cellular
mechanisms underlying δ-opioid regulation of ICA cell [Ca2+]i transients were not explored.
Although DPDPE induces robust epinephrine release from isolated ICA cells in vitro, we cannot
totally exclude the possibility that endogenous epinephrine derived from the sources other than
ICA cells might also contribute, in part, to β2-AR-mediated cardioprotection in vivo. Finally, the
signaling pathways of β2-AR-mediated cardioprotection by epinephrine were not further
characterized. We presumed that pertussis toxin-sensitive Gi pathway was likely involved in β2-
AR-mediated cardioprotection as proposed previously (3, 31).
In conclusion, we have localized DOR expression in ICA cells in human and rat hearts. δ-opioid
stimulation of ICA cells enhances endogenous epinephrine liberation resulting in myocardial β2-
AR-mediated cardioprotection against ischemia. This study has demonstrated a novel δ-opioid
signaling and its cardioprotective mechanism that is dependent on neuroendocrine function of
ICA cells.
Page 13 of 27
Copyright Information

14
ACKNOWLEDGEMENTS
We acknowledge the support of Gordon Ewy, MD in the early stage of this project. This study
was supported by the John Sealy Memorial Foundation from the University of Texas Medical
Branch.
Page 14 of 27
Copyright Information

15
REFERENCES
1) Bao L, Jin SX, Zhang C, Wang LH, Xu ZZ, Zhang FX, Wang LC, Ning FS, Cai HJ, Guan
JS, Xiao HS, Xu ZQ, He C, Hokfelt T, Zhou Z, Zhang X. Activation of delta opioid receptors
induces receptor insertion and neuropeptide secretion. Neuron. 37:121-133, 2003.
2) Bell SP, Sack MN, Patel A, Opie LH, Yellon DM. Delta opioid receptor stimulation mimics
ischemic preconditioning in human heart muscle. J Am Coll Cardiol. 36: 2296-2302, 2000.
3) Communal C, Singh K, Sawyer DB, Colucci WS. Opposing effects of β1- and β2-adrenergic
receptors on cardiac myocyte apoptosis: role of a pertussis toxin-sensitive G protein. Circulation.
100:2210-2212, 1999.
4) Cui M, Feng Y, McAdoo DJ, Willis WD. Periaqueductal gray stimulation-induced inhibition
of nociceptive dorsal horn neurons in rats is associated with the release of norepinephrine,
serotonin, and amino acids. J Pharmacol Exp Ther. 289: 868-876, 1999.
5) Ebert SN, Thompson RP. Embryonic epinephrine synthesis in the rat heart before innervation:
association with pacemaking and conduction tissue development. Circ Res. 88:117-124, 2001.
6) Goncalvesova E, Krizanova O, Micutkova L, Mravec B, Ksinantova L, Fabian J, Kvetnansky
R. Phenylethanolamine N-methyltransferase gene expression in transplanted human heart.
Transplant Proc. 37:1340-1342, 2005.
7) Guo Y, Stein AB, Wu WJ, Zhu X, Tan W, Li Q, Bolli R. Late preconditioning induced by
NO donors, adenosine A1 receptor agonists, and delta1-opioid receptor agonists is mediated by
iNOS. Am J Physiol-Heart Circ Physiol. 289:H2251-2257, 2005.
8) Huang MH, Ewy GA. Sympathetic reinnervation of transplanted heart. N Engl J Med (letter).
345:1914-1915, 2001.
Page 15 of 27
Copyright Information

16
9) Huang MH, Friend DS, Sunday ME, Singh K, Haley K, Austen KF, Kelly RA, Smith TW.
An intrinsic adrenergic system in mammalian heart. J Clin Invest. 98:1298-1303, 1996.
10) Huang MH, Bahl JJ, Wu Y, Larson DL, Hu FL, Ewy GA. Neuroendocrine properties of
intrinsic cardiac adrenergic cells in fetal rat heart. Am J Physiol-Heart Circ Physiol. 288:H497-
503, 2005.
11) Krumins SA, Faden A, Feuerstein G. Opiate binding in rat hearts: modulation of binding
after hemorrhagic shock. Biochem Biophys Res Commun. 127:120-128, 1985.
12) Lane-Ladd SB, Pineda J, Boundy VA, Pfeuffer T, Krupinski J, Aghajanian GK, Nestler E.
CREB (cAMP response element-binding protein) in the locus ceruleus: biochemical,
physiological, and behavioral evidence for a role in opiate dependence. J Neurosci. 17: 7890-
7901, 1997.
13) Lochner A, Genade S, Tromp E, Podzuweit T, Moolman JA. Ischemic preconditioning and
the beta-adrenergic signal transduction pathway. Circulation. 100:958-966, 1999.
14) Mieno S, Horimoto H, Sawa Y, Watanabe F, Furuya E, Horimoto S, Kishida K, Sasaki S.
Activation of beta2-adrenergic receptor plays a pivotal role in generating the protective effect of
ischemic preconditioning in rat hearts. Scand Cardiovasc J. 39: 313-319, 2005.
15) Mulkey RM, Zucker RS. Action potentials must admit calcium to evoke transmitter release.
Nature. 350:153-155, 1991.
16) Nestler ER, Aghajanian GK. Molecular and cellular basis of addiction. Science. 278: 58-63,
1997.
17) Olianas MC, Onali P. Participation of delta opioid receptor subtypes in the stimulation of
adenylyl cyclase activity in rat olfactory bulb. J Pharmacol Exp Ther. 275:1560-1567, 1995.
Page 16 of 27
Copyright Information

17
18) Patel HH, Head BP, Petersen HN, Niesman IR, Huang D, Gross GJ, Insel PA, Roth DM.
Protection of adult cardiac myocytes from ischemic cell death: role of caveolar microdomains
and opioid receptors. Am J Physiol-Heart Circ Physiol. 291:H344-350, 2006.
19) Peart JN, Gross GJ. Morphine-tolerant mice exhibit a profound and persistent
cardioprotective phenotype. Circulation. 109:1219-1222, 2004.
20) Peart JN, Gross GJ. Cardioprotective effects of acute and chronic opioid treatment are
mediated via different signaling pathways. Am J Physiol-Heart Circ Physiol. 291:H1746-1753,
2006.
21) Rubenzik M, Varga E, Stropova D, Roeske WR, Yamamura HI. Expression of alpha-
transducin in Chinese hamster ovary cells stably transfected with the human delta-opioid
receptor attenuates chronic opioid agonist-induced adenylyl cyclase superactivation. Mol
Pharmacol. 60:1076-1082, 2001.
22) Schultz JE, Hsu AK, Gross GJ. Ischemic preconditioning in the intact rat heart is mediated
by delta1- but not mu- or kappa-opioid receptors. Circulation. 97:1282-1289, 1998.
23) Sigg D, Coles JA, Oeltgen PR, Iaizzo PA. Role of δ-opioid receptor agonists on infarct size
reduction in swine. Am J Physiol-Heart Circ Physiol. 282:H1953-1960, 2002.
24) Thompson GW, Horackova M, Armour JA. Chemotransduction properties of nodose
ganglion cardiac afferent neurons in guinea pigs. Am J Physiol-Regul Integr Comp Physiol.
79:R433-439, 2000.
25) Tong H, Bernstein ME, Steenbergen C. The role of beta-adrenergic receptor signaling in
cardioprotection. FASEB J. 19:983-985, 2005.
26) Trautwein W, Hesheler J. Regulation of cardiac L-type calcium current by phosphorylation
and G proteins. Ann Rev Physiol. 52:257-274, 1990.
Page 17 of 27
Copyright Information

18
27) Villemagne PS, Dannals RF, Ravert HT, Frost JJ. Imaging of human cardiac opioid
receptors. Euro J Nuc Med Mol Imag. 29:1385-1388, 2002.
28) Wittert G, Hope P, Pyle D. Tissue distribution of opioid receptor gene expression in the rat.
Biochem Biophys Res Commun. 218:877-881, 1996.
29) Wolfrum S, Nienstedt J, Heidbreder M, Schneider K, Dominiak P, Dendorfer A. Calcitonin
gene related peptide mediates cardioprotection by remote preconditioning. Regul Pept. 127:217-
224, 2005.
30) Ye Y, Lin Y, Atar S, Huang M-H, Perez-Polo JR, Uretsky BF, Birnbaum Y. Myocardial
protection by pioglitazone, atorvastatin, and their combination: mechanisms and possible
interactions. Am J Physiol-Heart Circ Physiol. 291:H1158-1169, 2006.
31) Zhu WZ, Zheng M, Koch WJ, Lefkowitz RJ, Kobika BK, Xiao RP. Dual modulation of cell
survival and cell death by β2-adrenergic signaling in adult mouse cardiac myocytes. Proc Natl
Acad Sci USA. 98:1607-1612, 2001.
32) Zhou QY, Quaife CJ, Palmiter RD. Targeted disruption of the tyrosine hydroxylase gene
reveals that catecholamines are required for mouse fetal development. Nature. 374:640-643,
1995.
33) Zimlichman R, Gefel D, Eliahou H, Matas Z, Rosen B, Gass S, Ela C, Eilam Y, Vogel Z,
Barg J. Expression of opioid receptors during heart ontogeny in normotensive and hypertensive
rats. Circulation. 93:1020-1025, 1996.
Page 18 of 27
Copyright Information

19
FIGURE AND TABLE LEGENDS
Figure 1. Immunoperoxidase (panels A and C) and immunofluorescent (other panels) labeling of
ICA cells in human hearts. ICA cells expressing TH immunoreactivity (red) are distributed
diffusely throughout the LV myocardium. Perivascular location is a frequent feature of ICA
cells. Arrow (C) denotes a terminal arteriole. Panel E demonstrates abundant ICA cells in the
smooth muscle layers of epicardial circumflex coronary artery. TH-expressing sympathetic nerve
fiberss (arrows in D and G) can occasionally be seen in the field. Inserts are magnified ICA cell
images in panels B and D (arrows). TH immunoreactivity (green) was identified in ICA cells and
sympathetic nerve fibers in the SA nodal tissue (G). ICA cells are seen in transplanted human
LV tissue (H). All the scale bars =10 µm except for panel B (20µm).
Figure 2. Expression of TH mRNA and other neuroendocrine markers in human ICA cells. Two
ICA cells (A) that exhibit TH immunoreactivity (green) express TH mRNA (red, B) detected by
in situ hybridization in LV tissue. Inserts are magnified images of ICA cells. Panel C shows TH
immunoreactivity exhibited by a bundle of sympathetic nerve fibers that express no TH mRNA
(D). Panels E and F co-localize the immunoreactivity of TH (green) and PNMT (red) in an ICA
cell. Insert (F) is the overlay of E and F showing the co-localization of TH and PNMT. Panels G
and H co-localize the immunoreactivity of TH (green) and general neuronal marker PGP 9.5
(red) in a cluster of ICA cells. Scale bars =10 µm.
Figure 3. Expression of DOR immunoreactivity in human ICA cells. Immunofluorescent co-
localization of TH (green, A) and DOR immunoreactivity (red, B) in an ICA cell in human LV
tissue. Panel C: superimposed images of A and B exhibiting co-localized TH and DOR
immunoreactivity with TH concentrated on the opposite end. Inserts are magnified images.
Panels D and E show another ICA cell co-expressing immunoreactivity of TH and DOR. Panel F
demonstrates expression of DOR immunoreactivity (brown) by a perivascularly distributed ICA
cell in LV tissue by immunoperoxidase labeling (insert is the magnified cell image). Panel G
shows a TH-expressing sympathetic nerve fiber (green) in LV tissue. Double staining of the
same slide with anti-DOR antibody detects no DOR immunoreactivity in this nerve fiber and
ventricular myocytes (H). Scale bars =10 µm.
Page 19 of 27
Copyright Information

20
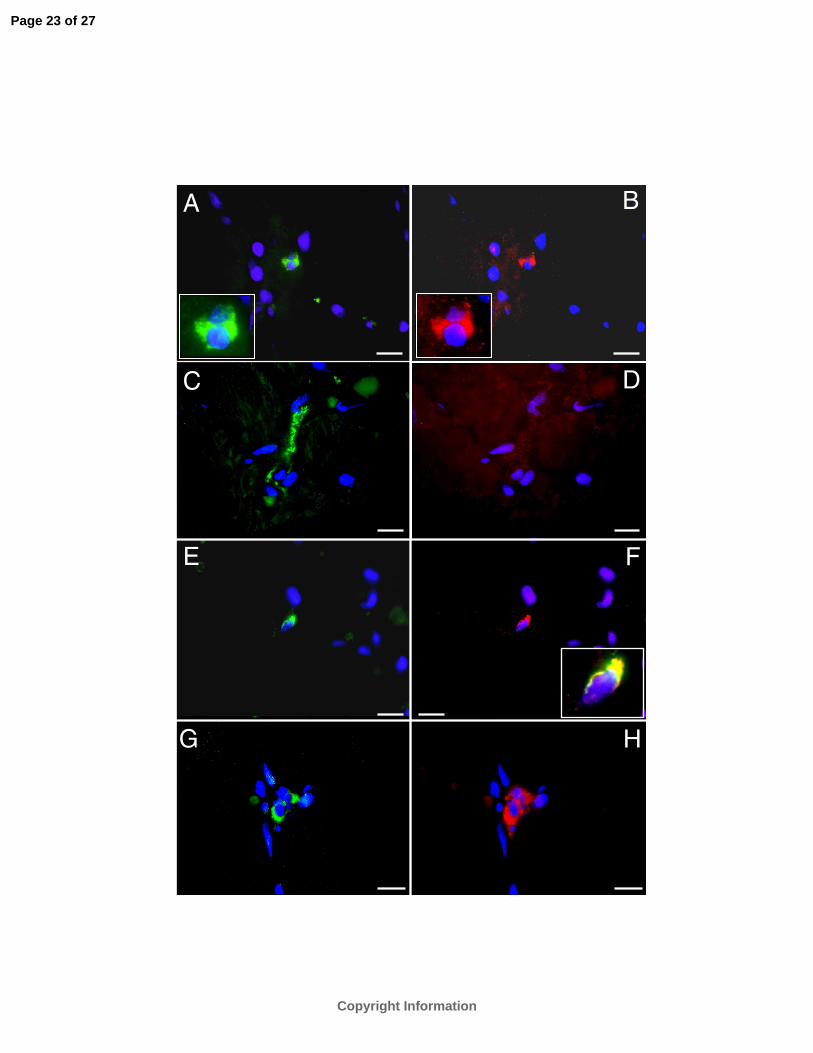
Figure 4. DOR expression in rat and human ICA cells. Immunofluorescent double labeling co-
localizes DOR (red, A) and TH (green, B) immunoreactivity in a cluster of ICA cells in rat
ventricular tissue. Panel C shows dissociated rat ventricular cardiocytes with only ICA cell
(arrow) exhibiting DOR immunoreactivity (green). Myocytes which display myosin-heavy-chain
immunoreactivity (red) express no detectable DOR immunoreactivity. Panel D shows >90%
isolated rat ICA cells expressing DOR immunoreactivity (green). Panels E and F show
immunofluorescent double labeling co-localizes the DOR (red) and TH (green)
immunoreactivity in isolated rat ICA cells. Insert (E) is the overlay of E-F showing co-localized
TH and DOR immunoreactivity in ICA cells. Panel G shows DOR activity (green) in two ICA
cells (arrow) abutted on myocytes expressing myosin-heavy-chain immunoreactivity (red) in
human LV tissue section. Calibration bar =10 µm. Panel H: Western blot analysis (repeated
twice) detects DOR protein in rat ICA cell isolates (lane 1) and brain tissue (lane 2) but not in
ventricular myocytes in the absence ICA cells (lane 3). Protein loading was 25 µg/lane. The
equivalency of protein loading was verified by the levels of β-actin.
Figure 5. Panel A: Modulation of [Ca2+]i transients generated by ICA cells by DPDPE. Bar
graph: DPDPE elicits a concentration-dependent increase (y-axis in fold) in [Ca2+]i transients
(n=11 cells). Top tracing shows that DPDPE (100 nmol/L) increases [Ca2+]i transients in an ICA
cell in culture. Middle tracing shows that increase in [Ca2+]i transients elicited by DPDPE is
abolished in the presence of nifedipine. Bottom tracing shows that DPDPE fails to increase
[Ca2+]i transients in the presence of the selective DOR antagonist naltrindole (NTI, 10 µmol/L).
Vertical scale bars represent 340/380 [Ca2+]i ratio. Panel B: DPDPE enhances epinephrine
release from ICA cells in culture. Basal epinephrine release at 1 hr is not different between the
two groups before the treatment of DPDPE or vehicle. Application of DPDPE (+DPDPE, 100
nmol/L for 30 min) to ICA cells increases epinephrine release by 2.4 fold (n=4 duplicates from 8
rats). There is no increase in epinephrine release when ICA cells are exposed to vehicle solution
(-DPDPE, n=4 duplicates from 8 rats). Enhanced epinephrine release persists during recovery
phase after the removal of DPDPE. **: p<0.01, *: p<0.05.
Figure 6. Panel A: Effects of DPDPE (200 µg/kg) on infarct-size-reduction in the absence and
presence of labetalol (Lab, 2 mg/kg), ICI-118,551 (ICI, 1 mg/kg), or NTI (200 µg/kg) in vivo.
Page 20 of 27
Copyright Information

21
Effects of labetalol, ICI, or saline (S) on infarct size are presented. Infarct size in DPDPE group
is markedly smaller that those of all other groups (p<0.001). Panel B: Effects of DPDPE (100
nmol/L) on ischemia-reperfusion-induced ventricular myocyte death in the absence and presence
of labetalol (1 µmol/L), ICI (1 µmol/L), or NTI (10 µmol/L) in ICA cell-ventricular myocyte co-
culture (n= 5/group). The myocyte death in ischemic group is significantly higher than that of
nonischemic one (p<0.001). Among ischemic groups, the myocyte death in DPDPE group is
significantly lower than those of all other groups (p<0.001). Panel C: Effects of DPDPE on
ischemia-reperfusion-induced cell death in ventricular myocytes in the absence of co-cultured
ICA cells (n=5/group). *: p<0.05, **: p<0.01, ***: p<0.001, ns: nonsignificant.
Page 21 of 27
Copyright Information

C
A
D
F
G H
B
E
Page 22 of 27
Copyright Information
A B
C D
E F
G H
Page 23 of 27
Copyright Information

A
B
C F
D
E
G H
Page 24 of 27
Copyright Information

A B
C D
E F1 2 3
DOR
H
β-actin
G
Page 25 of 27
Copyright Information

Nifedipine
DPDPE
DPDPE NTI
DPDPE
10 min
Baseline ± DPDPE Recovery
A
B
Page 26 of 27
Copyright Information

**ICI vs. (Lab+DPDPE)
***ICI vs. (ICI+DPDPE)
**ICI vs. Saline
PIntra-group
**Lab vs. ICI+DPDPE
*ICI vs. NTI
*ICI vs. Lab
*Saline vs.(NTI+DPDPE)
**Saline vs. (ICI+DPDPE)
*Saline vs. Lab
***Saline vs. ICI
PIntra-group
Panel B statistics
Panel A statistics
*** ns
***
A
B
C
Page 27 of 27
Copyright Information


















