
MAKlO, a Glucose-Repressible Gene Necessary for ... · PDF fileduction of killer toxin led to...
10
Copyright 0 1992 by the Genetics Society of America MAKlO, a Glucose-Repressible Gene Necessary for Replication of a dsRNA Virus of Saccharomyces cerevisiue, Has T Cell Receptor a-Subunit Motifs Yang-Ja Lee and Reed B. Wickner Section on Genetics of Simple Eukaryotes, Laboratory of Biochemical Pharmacology, National Institute of Diabetes Dagestive and Kid%eyDiseases, National Institutes of Health, Bethesda, Maryland 20892 Manuscript received April 8, 1992 Accepted for publication May 18, 1992 ABSTRACT The MAKlO gene is necessary for the propagation of the L-A dsRNA virus of the yeast Saccharomyces cererrisiae. We haveisolated MAKlO from selectedphage X genomic DNA clones that map near MAKJO. This gene encodes a 733-amino acid protein with several regions of similarity to T cell receptor a-subunit V (variable) regions. We show that MAKlO is essential for optimal growth on nonfermentable carbon sources independent of its effect on L-A. Although loss of L-A by makl0-1 mutants is partiallysuppressed by loss of themitochondrialgenome,nosuchsuppression of a makl0::URAP mutation was observed. Using MAKlO-lacZ fusions we show that MAKlO is expressed at a very low level and that it is glucose repressed.The highest levels of expression were seen in tup1 and cyc8 mutants, known to be defective in glucose repression. These results suggest that the mitochondrial genome and L-A dsRNA compete for the MAKlO protein. L -A is a 4.6-kb linear, single-segment dsRNA virus of Saccharomyces cerevisiae (reviewed in WICKNER 199 1,1992). Viral (+) strands are produced by con- servative intraviral transcription and extruded from the viral particles into the cytoplasm where they are translated and packaged in viral proteins to form new virus particles. The newly packaged (+) strands are then converted within the particles to double-stranded form completing the cycle. The L-A (+) strand en- codes two proteins: the 5’ open reading frame (ORF) (gag) encodes the viral major coat protein (76 kD) and a fusion of the 5’ ORF and the overlapping 3’ ORF (pol) encodes a fusion protein (170 kD) formed by a -1 ribosomal frameshift (FUJIMURA and WICKNER 1988b; ICHO and WICKNER 1989; DINMAN, ICHO and WICKNER 1991). The pol domain of the fusion protein has the consensus sequences diagnostic of viral RNA- dependent RNA polymerases (ICHO and WICKNER 1989; RIBAS and WICKNER 1992), and a sRNA-bind- ing activity thought to be involved in packaging (Fu- JIMURA and WICKNER 1988b; FUJIMURA et al. 1990). Using template-dependent invitro replication, tran- scription and RNA-binding activities of the isolated opened empty viral particles, the cis-acting sites needed for viral replication and packaging have been determined (FUJIMURA and WICKNER, 1988a, 1989; ESTEBAN, FUJIMURA and WICKNER 1988, 1989; FUJI- MURA et al. 1990). MI dsRNA is a satellite of L-A that encodes a secreted protein “killer” toxin and immunity to that toxin (reviewed by BUSSEY 1988; WICKNER 1991). Studies of the chromosomal genes necessary for pro- duction of killer toxin led to the discovery of the KEXl and KEX2 genes (killer expression) (WICKNER Genetics 132: 87-96 (September, 1992) and LEIBOWITZ 1976a; LEIBOWITZ and WICKNER 1976), encoding proteases that process the prepro- toxin (JULIUS et al. 1984; DMOCHOWSKA et al. 1987), and,from these, to homologous mammalian genes encoding proteases responsible for processing prohor- mones (ROEBROEK et al. 1986;FULLER,BRAKE and THORNER 1989; SMEEKINS and STEINER 1990; SMEEK- INS et al. 1991). The killer phenotype was used to identify several chromosomal genes (MAK for maintenance of killer) which are required for viral propagation, and others (SKI for superkiller) that negatively control viral copy number. The propagation of the L-A virus requires the products of the MAK3, MAKlO and PET18 chro- mosomal loci (WICKNER and TOH-E 1982; SOMMER and WICKNER 1982). MAK3 encodes a protein N- acetyltransferase that modifies the amino terminus of the gag protein, a modification necessary for particle formation (J. C. TERCERO, L. RILES and R. B. WICK- NER, submitted; J. C. TERCERO and R. B. WICKNER, submitted). The pet1 8 mutation is a large deletion that includes two ORFs, MAK31 and MAK32, both of which are necessary for propagation of L-A, but only at temperatures above 30 O . The biochemical function of PET18 is not known (TOH-E and SAHASHI 1985). There is presumably a fourth host gene that encodes the “host factor” that is required in vitro in the repli- cation [(-) strand synthesis] step (FUJIMURA and WICK- MAKlO was actually the first of the MAK genes discovered (SOMERS and BEVAN 1969). T h e inability of mak25 mutants to maintain L-A led to the discovery that there were two unrelated L-sized dsRNAs, L-A and L-BC (WICKNER and TOH-E 1982; SOMMER and NER 1988a).
Transcript of MAKlO, a Glucose-Repressible Gene Necessary for ... · PDF fileduction of killer toxin led to...

Copyright 0 1992 by the Genetics Society of America
M A K l O , a Glucose-Repressible Gene Necessary for Replication of a dsRNA Virus of Saccharomyces cerevisiue, Has T Cell Receptor a-Subunit Motifs
Yang-Ja Lee and Reed B. Wickner Section on Genetics of Simple Eukaryotes, Laboratory of Biochemical Pharmacology, National Institute of Diabetes Dagestive and
Kid%ey Diseases, National Institutes of Health, Bethesda, Maryland 20892 Manuscript received April 8, 1992
Accepted for publication May 18, 1992
ABSTRACT The MAKlO gene is necessary for the propagation of the L-A dsRNA virus of the yeast Saccharomyces
cererrisiae. We have isolated MAKlO from selected phage X genomic DNA clones that map near MAKJO. This gene encodes a 733-amino acid protein with several regions of similarity to T cell receptor a-subunit V (variable) regions. We show that MAKlO is essential for optimal growth on nonfermentable carbon sources independent of its effect on L-A. Although loss of L-A by makl0-1 mutants is partially suppressed by loss of the mitochondrial genome, no such suppression of a makl0::URAP mutation was observed. Using MAKlO-lacZ fusions we show that MAKlO is expressed at a very low level and that it is glucose repressed. The highest levels of expression were seen in tup1 and cyc8 mutants, known to be defective in glucose repression. These results suggest that the mitochondrial genome and L-A dsRNA compete for the MAKlO protein.
L -A is a 4.6-kb linear, single-segment dsRNA virus of Saccharomyces cerevisiae (reviewed in WICKNER
199 1, 1992). Viral (+) strands are produced by con- servative intraviral transcription and extruded from the viral particles into the cytoplasm where they are translated and packaged in viral proteins to form new virus particles. The newly packaged (+) strands are then converted within the particles to double-stranded form completing the cycle. The L-A (+) strand en- codes two proteins: the 5’ open reading frame (ORF) (gag) encodes the viral major coat protein (76 kD) and a fusion of the 5’ ORF and the overlapping 3’ ORF ( p o l ) encodes a fusion protein (170 kD) formed by a -1 ribosomal frameshift (FUJIMURA and WICKNER 1988b; ICHO and WICKNER 1989; DINMAN, ICHO and WICKNER 199 1). The pol domain of the fusion protein has the consensus sequences diagnostic of viral RNA- dependent RNA polymerases (ICHO and WICKNER 1989; RIBAS and WICKNER 1992), and a sRNA-bind- ing activity thought to be involved in packaging (Fu- JIMURA and WICKNER 1988b; FUJIMURA et al. 1990). Using template-dependent in vitro replication, tran- scription and RNA-binding activities of the isolated opened empty viral particles, the cis-acting sites needed for viral replication and packaging have been determined (FUJIMURA and WICKNER, 1988a, 1989; ESTEBAN, FUJIMURA and WICKNER 1988, 1989; FUJI- MURA et al. 1990).
MI dsRNA is a satellite of L-A that encodes a secreted protein “killer” toxin and immunity to that toxin (reviewed by BUSSEY 1988; WICKNER 1991). Studies of the chromosomal genes necessary for pro- duction of killer toxin led to the discovery of the K E X l and K E X 2 genes (killer expression) (WICKNER Genetics 132: 87-96 (September, 1992)
and LEIBOWITZ 1976a; LEIBOWITZ and WICKNER 1976), encoding proteases that process the prepro- toxin (JULIUS et al. 1984; DMOCHOWSKA et al. 1987), and, from these, to homologous mammalian genes encoding proteases responsible for processing prohor- mones (ROEBROEK et al. 1986; FULLER, BRAKE and THORNER 1989; SMEEKINS and STEINER 1990; SMEEK- INS et al. 199 1).
The killer phenotype was used to identify several chromosomal genes (MAK for maintenance of killer) which are required for viral propagation, and others ( S K I for superkiller) that negatively control viral copy number. The propagation of the L-A virus requires the products of the M A K 3 , M A K l O and PET18 chro- mosomal loci (WICKNER and TOH-E 1982; SOMMER and WICKNER 1982). MAK3 encodes a protein N- acetyltransferase that modifies the amino terminus of the gag protein, a modification necessary for particle formation (J. C. TERCERO, L. RILES and R. B. WICK- NER, submitted; J. C. TERCERO and R. B. WICKNER, submitted). The pet1 8 mutation is a large deletion that includes two ORFs, MAK31 and M A K 3 2 , both of which are necessary for propagation of L-A, but only at temperatures above 30 O . The biochemical function of PET18 is not known (TOH-E and SAHASHI 1985). There is presumably a fourth host gene that encodes the “host factor” that is required in vitro in the repli- cation [(-) strand synthesis] step (FUJIMURA and WICK-
M A K l O was actually the first of the M A K genes discovered (SOMERS and BEVAN 1969). The inability of mak25 mutants to maintain L-A led to the discovery that there were two unrelated L-sized dsRNAs, L-A and L-BC (WICKNER and TOH-E 1982; SOMMER and
NER 1988a).

88 Y.-J. Lee and R. B. Wickner
TABLE 1
Strains of Saccharomyces cerevisiae
Strain Genotype Source
5x47 MATaIMATa hisl/ + trpl/ + ura3/ + [KIL-o]
3063 2629 2842 2842 Bltfl 2842 B;tf2 Cross 4291 Cross 4292 Cross 4303
Crosses
Y L42 RTY235 RTY363 RTY4 18
Y003, YO04
MATS makl0-1 trpl adel his3 po L-A-o M-o MATa leu1 karl-1 L-A-HNB M I p+ 2601 (MATa ura3-52 his5 M-o) X 2602 (MATa ura3-52 his6 leu2 L-A-HN MI) 2842 heterozwous for maklO::URA3 disruvtion 2842 heterozbEous for maklO::URA3 disrubtion Segregants 019842 Bltfl Se re ants of 2842 Bltf2 426l-qB (MATa his(5,6) ura3maklO::lJRA3) X 2944 (MATa ura3 thrl L-A-o M-o 20s-0) 18 (MATa lysl L-A-HN MI) X 1053 (MATa iIv3 ura3 can1 makl0-1 L-A-o M-o)
MATa his5 ura3-52 L-A-o "o L-BC MATa his4-519 leu2-3, 112 trpl-289 ura3-52 MATa his4-519 leu2-3,112 ura3-52 cycBAl::LEU2 MATa his4-519 leu2-3,112 trpl-289 ura3-52 tupl-A1::TRPl
RIDLEY, SOMMER and WICKNER (1 984) WICKNER et al. (1991) WICKNER et al. (1991) RHEE, ICHO and WICKNER (1989) This work This work This work This work This work
This work
This work WILLIAMS and TRUMBLY (1 990)
WILLIAMS and TRUMRLY (1 990) WILLIAMS and TRUMBLY (1 990)
clwl *PRB1 ~
?lo 16 CWl
Genetlc Map CM I I 1 I I FIGURE 1 .-Genetic and physical
kb 7 10 20 30 40 50 60 maps of the region around MAKlO and complementation tests of subclones.
Phy8lcal Map I mal I , 1 1 1 1 I 1 1 1 1 I I wn I II 111 d l 11, I I a The genetic map of part of the left arm
between genes in centiMorgans. The
I I I I I I t I I I I I
I S I I I I I I 1 I 1 I I 1 I 1 I I of V is shown at the top with distances
"e- -.. centromere is to the right. The physi- -- . cal map is shown with the locations of
"""". L 2 -" ""
-_ ._ "-" -. Hind111 or EcoRI sites that are known (vertical lines that go through the hor- izontal line) and those for which frag- ments are only partially localized (ver- tical lines that only meet the horizontal line) (data from L. RILFS and M. OL-
EjdentdlheMAKlOgene SON, personal communication). There- striction map of X6662 is shown with - , the region deleted and replaced by
region sequenced in this work and the extent of the M A K l O gene based on the subclones shown below. The open
H - - rnakl 0- " URA3 in the disruption mutation, the - 1 I H 2 kb - M
Smk box is the minimum extent of the gene - and the hatched box is the maximum - extent. H = HindII1; E = EcoRI. - M A K ~ O+
WICKNER 1982). A maklO& mutation results in insta- drial function. Deletion of the major mitochondrial bility of particles containing L-A dsRNA, but the outer membrane porin led to poor growth on glycerol particles from the Same cells containing L-A (+) and accumulation of L-A virus particles, suggesting sRNA a re relatively stable (FUJIMURA and WICKNER that it was the loss of L-A that led to the poor growth 1987). This result suggested that the MAKIO protein OfmaklO strains on glycerol O r ethanol (DIHANICH et
was involved in the assembly process or was actually 1989). part of the particle. Supplying L-A-encoded proteins We show here that MAKlO encodes a glucose-re- from an L-A cDNA vector supresses the mak10-1 pressible protein necessary for normal growth on non- mutation for M~ replication, but not for L-A replica- fermentable carbon sources independent of its effect tion, suggesting that the requirement of MAKlO for On the L-A virus and that the MAK1o product has L-A propagation is more stringent than for MI (WICK- several local similarities to the a-subunits of T cell
NER et al. 199 1). receptors.
The makl0-1 mutation is partially suppressed by loss of mitochondrial DNA (WICKNER 1977). This connection with mitochondria was reinforced by DI- Strains and media: YPAD, YPG (glycerol), YPE
(ethanol), SD and synthetic complete (= H) medium (SHER-
grow slowly on glycerol or ethanol, nonfermentable 1982) and 4.7MB (for the killer test, RIDLEY, SOMMER and carbon sources whose utilization requires mitochon- WICKNER 1984) have been described. Most of the strains
MATERIALS AND METHODS
HANICH et (lg8') finding that maklO-l MAN 1991), LB medium (MANIATIS, FRI~CH and SAMBROOK

Yeast MAKlO Gene 89 h01 ~ G T A T C Q A Q T T A C C A A T T T T G Q A T A C A T C C A C A T A C Q C C C C A A T Q T T C T T A Q Q T T T Q C C A C Q Q C T A A A T A 7 5
C Q G A T T T A A G A C C * T T C A A C C A A C C G C C A A C T T Q ~ A Q ~ A T T Q Q Q T Q T ~ T T Q A C A A C C Q Q A Q Q T A C A T A T T 1 5 0
TAACCAQTQAATCATACAAATCCCCTAAQQAAQAQATQATACCACQCTQATATQTATCATCCCQTAQCTTCCCCA 2 2 5
ACTTCACCAQTCTQTCQTACTCTTQCAAQQQCQTCAQACACTTATTTQACTTCQTAQATTQATACCQACAAATQC 3 0 0
Q A A A Q C T Q C A A T Q C T G C A C T T Q T C Q Q A A Q Q T T C Q A A C A ~ C A T T Q Q Q C T T C A A A Q C Q A T C A T Q A T T T T A A C C A Q ~ A a 7 5
CTCCCTCCCACTTTAAAG~~AATCCQQAQATTCCCQTTATTACCTTACATATCQAAATATQACCCATAAAAQATQ 5 2 5
T Q T A A A A T T A C G T A A Q A C A A A Q Q A A C T C Q Q C A C A C C T A T T Q A A T A Q Q T A A T A Q T Q Q A A T C A Q Q T Q C C T A A A T A A A 8 0 0
A T Q T T T T ~ : A T T A T Q A Q A T Q Q A Q T Q A A A T T T ~ A ~ Q A A Q A Q A T A ~ A A Q T A A A A T A Q ~ A Q C T C T T C A A ~ A 7 5 0
QCAAQAAQAQTQQCQCTQAQATTQAAAACAACTQTQCTAQTQTAQAQCAACTTTQATAATAQCTQQTACQTTTCC 8 2 5
Q A Q A A T Q Q A A G T A Q A C A C T A T A T T A Q Q T T C C T T Q A Q C A T A A C T Q A T Q A T T T T Q A C C A A T T Q Q T T Q A C Q ~ C A ~ A A ~ 9 ; :
T C T Q T T T Q A T Q A A T T Q T Q T T C A A A A C T A A A A C C A Q A Q Q C T A T C Q T Q A A A Q A T C ~ C A Q Q T ~ T Q A T T T Q T T C Q ~ Q Q ~ 9 i z
T A C A C A T T C T T T A Q A G Q T C A A C A A C T C C A A A T T A Q A C T C C A Q T C T Q A T A Q A A T T Q A C A Q C A Q A A Q A A A T A Q A A ~ T l o 5 0
AATACTATGTTTCACTGCTTTACCTATTTATTCTTTQTTCTTTCQQAQCACCCQCAAATCAQAAQAAAQQAACTT 4 5 0
T Q A A T A A Q A Q T A C T Q T A T T A T C T A Q T A Q T T Q T T A T A T T A T C T A A C C T T T A C A T A A T A C Q A T T A T T T C A C A T T Q T T 6 7 5
M E V D S I L Q S L S I T O D F D ~ L V D
L F D E L C S K L K P E A I V L O R O L F
T H S L E V N N S K L D S S L I E L T A E E I E F 7 4
TQATQTQAACGTQQCCTACGATCCACCQTTQQCAAQTQTAQCAQCCATAQCQQATAQATTACTTAQATQTQTTAT 1 1 2 5 D V N V A Y O P P L A S V A A I A D R L L R C V I 9 9
CTCCTQQCTQAATGACTACCAQACTTTACCCACTACTQTATTAAQCTQCAQQTATACAQ~ ~ T T A T C O T C 1 2 0 0 8 W L N D Y 0 T L P T T Y _ _ _ _ L . 5 _ _ _ _ ' L . 8 _ _ _ _ Y...L L 8 5 1 2 4
ATTAQTTAAQGQAACAACTQCCQQTTCQTCCTQQTQTACAQQQAATATTTTQTATQATAAAQTACTAQQAAQTTQ 1 2 7 s
C A T A C T T Q C C Q T T T Q T T A T T T A A C A A A A T T T Q T Q C A Q A A A C T C T T Q A Q T Q C T Q Q T A T T Q T T T T T Q A A Q A Q G A Q Q A 1 8 5 0 I L Q V C Y L T K F V ~ K L L S A Q I V F E E E D ~ ~ ~
L V U Q T T A Q S S W C T Q N I L Y O K V L Q S ~ l ~ V
T T T A A A T T T T A A T A A C A T Q Q Q Q T T T A A T A C A T T C Q A T A A T T T Q C C Q Q Q Q C A A Q A T Q T T Q T T A T C A A T T C T C T C A C 1 4 2 5 L N F N N M Q F N T F D N...l...P _ _ _ _ Q...B...Q-..V-~-Y~~-+...N....8....c....r 1 9 9
C Q A Q A Q C C T T C A Q A T A T T A Q A Q Q C A T A T T C T Q A T Q A T A Q C T T A C A T T T A A C T A T Q C T Q A A Q C A T A T A C T A A A A A T 1 6 0 0 ~ . ~ ~ ~ ~ S L O I L E A Y S D D D L H L T M L U H I L K I ~ ~ ~
TATCATTTQCTTAQTCCACTTAQAAQACCATCTQACAQACTATTCCACAAAQACAAQCCACCTQQATQAQCTAAT 1 5 7 5
T G A A A A C Q C T A A C T C Q Q T T A A T Q Q C A T A T T T C C A C A A T T A C A Q C T Q T C T C C A C C A A A Q Q Q T Q C Q T ~ C T C A A ~ A T ~ 1;;: E N A N S V N G I F P Q L ~ L S P P K Q A 8
T A T A C A A A A G C A C A G Q T C T A A T C A A T T C C C T C C C A Q Q A A Q A T T A C Q A A Q T T A C C Q A C Q Q A T T A C A Q C Q Q C T T T A T 1 7 2 5 I ~ K H R S N O F P P R K I T K L P T O Y S Q F I ~ O O
T L A H D V U T I L L V D U A E S A L E T Y ~ F A ~ ~ ~
C A A A T ~ C T T C A A C A A Q T T C Q A A C A T O T C A T T O C * A Q Q A T C T ~ A T T C C C C T T A T ~ T T T C A T T A Q A Q A C Q ~ 1 8 7 5
C A G A A C A Q T T T T Q Q Q T A A A T T T T C A T A T A C C C A A T T T T A T T T ~ C T A C A T Q T C A A Q Q A G T T T T C A Q C A C A A A C T C C 1 9 0 0 R T V L Q K F S Y T ~ F Y L L H V K E F S A ~ T P ~ ~ ~
S E F E S S I Q N E L I ~ E S S N M L L E W Y ~ N ~ ~ ~
C T G C T C C C A A A A C A C A T G T A Q A T A T A Q A C A A Q Q T T T T A A C A Q Q C A A C T T A T T C T A T Q Q Q A T T C T C T T C A A Q C C ~ A 2 1 0 0 C S ~ N T C R Y R ~ Q F N R O L I L W D S L ~ A ~ ~ ~ ~
Q T T T Q A A A Q T Q T C A A T T C A C A A Q T T T A C T Q T T C Q T Q Q A C A T A T T T T A T Q A A A C T A T C A A Q T A T Q A T C ~ T ~ 2 1 7 5 F E S V N --L.O---Y"-V _ _ _ _ L - 5 _ _ _ _ V?---LL. F M K L 8 5 M I 4 4 9
T T T A A A Q G Q Q T T T G A T T T Q Q A T A T T T A C A A A C C C T T T Q A A Q C A T A C T C T A T Q T T T T Q Q T A T Q T T T A T T A T T T A A Q 2 2 5 0
T C A C C A C T T A Q A A A C C T T T T T Q A A A Q A T A Q T C A A A A T Q A T A T T Q A A T C A A A T A T T A A T Q C C A T T C A T T C C A T Q A A 2 3 2 6 H H L E T F L K D S ~ N D I E S N I . N A I H S M N ~ ~ Q
C A A ~ A A A C T T A A A A A A T T Q A A A Q C T Q Q T Q A A A A Q A A Q Q A C C A A T T Q A Q G C T A A A Q T A C A ~ Q T T ~ Q C T A T Q Q A T A ~ 2 4 0 0 K K L K K L K A Q E K K D ~ L R L K Y F A M D 5 2 4
T Q A A A T Q Q A A C A A T T A C A A G C T A C C A A A C A A T T T T T A A A C T A T C T T T T Q A A Q Q A A A T T A A C A T T A C C A A A T C T T T 2 4 7 5 E U E Q L Q A T K ~ F L H Y L L U E I N I T K S L ~ ~ Q
A T Q T C T A A T A G A Q Q T T T T C C A A T T T Q C A A T T T T A A A Q T C A T T T Q Q T T T A A T T Q A C A ~ C A A Q ~ ~ T ~ Q T ~ C T C C Q ~ C 2 5 5 0 C L X E V F ~ F A I L K S F Q L I D N K N S T P S ~ ~ ~
A A A A T T C T C T A A T G A A C Q Q T T A A T C C A T A A T T T Q A Q A T T C A A Q C C A T T T ~ ~ T ~ ~ A A T T Q Q T Q T ~ C C T Q ~ Q T T A C C 2 5 2 5 K f S N E R L l H N L R F K P F N S I Q v p E L P 5 g g
A Q A A T ~ T G A A G T A T T T C A A C A A A C T T T A A A A Q A T T T T Q ~ C A T T Q A A Q ~ ~ ~ ~ ~ Q Q A Q C T Q C A T ~ ~ Q ~ T ~ ~ ~ ~ ~ Q T T 2 7 0 0 E E V F ~ O T L K D F V I E E K Q A A F O I K L ~ Z ~
A Q A A A Q Q Q C C A C T A A C T T T A T C Q A A A C T Q A A Q T Q C Q C ~ ~ T Q T C Q T ~ A Q T ~ C ~ ~ ~ T Q ~ ~ Q ~ Q ~ ~ ~ ~ ~ Q C ~ ~ Q Q ~ ~ ~ 2 7 7 5 E R A T N F I E T E V R N V V S S I D E I M ~ Q I ~ ~ ~
C*~QQQAGQCQ*TA*TAACGQCQTTCTAQTAACAQQAACAAQACTQQTTCAAQAQCTCTCTCTQQAQTACTATTQ 2 0 5 0 K Q Q D N N Q V L V T Q T R L V ~ E L S L E ~ ~ ~ ~ ~ ~
A K l4 -.?---R--.K-"! _ _ _ _ C...&...Y.. N 8 K V I V N T L K I( N I K 8 9 9 T ~ ~ * C T ~ A * ~ C A T * C T T C T ~ A A Q C A T T A T C A Q T C A A C T C A A A A Q T T A T C Q T C A A C A C T T T Q A A A A A Q A A C A T T A A 2 9 2 5
~ ~ ~ C A ~ Q Q A C T C C C ~ C Q A A T A C A A Q Q T T Q A A T T A Q T T C A T A C A A C A G A A Q Q A T Q Q A A C T A T T T C C C C A T A C A A A C 3 0 0 0 N K D S H E Y K V E L V H T T E Q W N ~ F ~ I ~ T ~ ~ ~
* ~ ~ Q ~ Q A A T A A * Q C * A ~ A C C Q C T A T A A A T ~ A T C T Q Q A A T T T A C C C T A A C T Q T A C A A A A T A T A T T T A Q C A T T A T A T 3 0 7 5
*CCCQCTATTTAATTATTATAAAQQAQACCTTTATACTCAAACAAATQAAQCCACCTTATQCCATTATCCCATTC 3 7 ~ 0
733
Q T ~ C C * Q T C T A * * A C A * G * T A A A T A C C C T Q T T T A A C A T C C Q A A C A C C A A A T A T Q T ~ T Q Q Q T Q T C A A A C A A A Q A Q A 3 2 2 5
C A A T Q ~ A A T Q * Q * C A * T Q T A C T Q C T T Q T A A A C T A T A A C A Q T A T C A Q C T C A A T T T C A A T Q C C A T C T A C T C C A Q A T A 3 3 0 0
T C Q T T P C Q A T T Q T A C C T A C A A Q C C Q C C T A C A A C Q Q C Q T T A T T Q T T Q T A A Q C T Q Q T A A A A T Q C A Q T Q C Q T T C ~ ~ ~ C 3 3 7 1
Q T T Q T Q A Q T T C Q Q Q C C A A A C T T A C T T Q Q C T Q Q Q A Q T T C C Q T C C Q T A C A Q T T Q T Q Q C T T Q C T Q C C T A Q A C T ~ A Q Q C a 4 s a QQACQCQQCQQCAATTACTCCTQQQCTCTCCCTAQAQCTQQAQAQACACTCTTQQTAACCGAAQQQAAAATCCCQ 3 5 2 5
~ * * ~ ~ * ~ A T T T ~ Q A A T C C G T T A A C A C T Q C T Q Q A A A T Q Q A C A T ~ T C C T A A T A ~ T Q A A A A A A T T A T T ~ A A A Q ~ ~ ~ ~ ~
I I C L V H L E D H L T D Y S T K T S H L D E L I Z ~ V
TAChCfQQCAAATQATQTTAAAACAATACTCCTTQTAQATAAAQCCQAQTCTQCQTTQQAQACCTACCAQTlT~~ 1 8 0 0
K F N K L E Q R H V I A R I F P L F I R D 3 4 9
A T C T G A A T T T Q A A T C C T C T A T T Q Q T A A T Q A A C T T A T C C A A Q A A A Q C T C T A A T A T Q C T T T T A Q A A T Q Q T A C C A A A A 2 0 2 5
M I
L U G F D L D I Y K P F E A Y S M F W Y V Y ~ L ~ 4 7 4
L R I K O D R Y K
FIGURE 2.-Sequence of the MAKIO gene. Regions found to have motifs present in T cell receptor a-subunits (Figure 3) are underlined. Solid underlines are used for the two regions for which the motifs were shown to be unique to T cell receptor &-subunits, while dotted underlines indicate similarities of uncertain statistical signif- icance. A potential REBI-binding site is also in- dicated. The orientation of the gene is shown by an arrow in Figure 1.
used are shown in Table 1. Strain 5x47 was the indicator- sensitive strain for the killer test (RIDLEY, SOMMER and WICKNER 1984). Transformation of yeast was by the lithium acetate method (ITOH et al. 1983). The diploid 2842 was used to disrupt the MAKlO gene by the method of ROTH- STEIN (1991). The isogenic strains RTY235 (wild type), RTY363 (cyc8Al::LEUS) and RTY418 (tupl-A1::TRPl) kindly provided by R. J. TRUMBLY (WILLIAMS and TRUMBLY 1990; WILLIAMS, VARANASI and TRUMBLY 1991) were used to study regulation of MAKlO expression, along with the wild-type strain YL42 and its pa derivative.
Testing plasmids for the M A K l O gene: The makl0-1 strain 3063 was transformed with the plasmid to be tested.
Cytoplasm carrying the mitochondrial genome (p'), as a cytoduction marker, and the L-A and MI viruses, was trans- ferred from the karl killer strain 2629 by cytoduction as described (RIDLEY, SOMMER and WICKNER 1984). The cy- toduction mix was plated on H-ura-leu plates to select against the donor strain and for retention of the plasmid in the recipient strain. After 2 days, colonies were replica- plated on SD to screen out diploids, YPG to detect cytoduc- tants by the presence of p , and 4.7MB plates with a lawn of a sensitive strain to detect killers. The plasmid was inferred to contain MAKIO if the p + , Ura+, SD- cytoductants were killers.
Sequence analysis: ssDNA templates from deletion mu-

Y.3. Lee and R. B. Wickner 90
A25733 F24402 E24891 D27557 RUMS58
A32537 C25733 A32536 E29774 M A K l O
A25733 F24402 E24891 027557 RUMS58 A32537 C25733 A32536 E29774 M A K l O
A25733 F24402 ~ 2 4 8 9 1 D27557 RUMS58 A32537 C25733 A32536 E29774 M A K l O
74 77 77 54 74 73 74 74 79
488
l v k g i k C F E A E F l K S L F S F N L R K P S V ~ U S D - - - - - - - T A E Y F C A V N E y ~ k l - - - - - - - - - - - - - S F C A C T T V T V R a - - - - - - - - - r tehq-GFHATLHKSSSSFHLOKSSAPLSD-- - - - - -SALYYCALCTg~nf- - - - - - - - - - - - -YFCKGTSLTVIp- - - - - - - - -
vvqgvnCFEAEFSKSNSSFHLRKASVHUSD-------UAVYFCAVSLny~gkl-----------IFCPCTKLSIKpniqnpep--
vvqgvnCFEAEFSKSNSSFHLRKASVHUSO------SAVYFCAVSCfasal------------- TFCSGTKVIVL---------- lvkgsyGFFAEFNKSPTSFHLKKPSALVSD-------SALYFCADSAlyfcavr~rgstlgr--LYCRCTPLTVV~iqn----- ik - - - -GFTADLNKGETSFHLKKPFAQEED-- - - - - -SAHYYCALSV~ggkl - - - - - - - - - - - IFCPCTELSVKp-- - - - - - - - lvkginGFEAEFKKSETSFHLTKPSAHHSD-------AAEYFCADMeyfcavsdlepnssaskiIFCSCTRLSIRpniqn-----
esninaihsrmkklkklkagekkdqlrlkyrf ... e l S L E Y Y C K L K h - - - - - - - - - - - - - - - - - - - - - TSKALSVnskvivntl..
r f e h q - ~ F H A T L H K S S S S F H L ~ K S S A ~ L ~ D - - - - - - - S A L y y C A L S D - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
""" P G R Y N H T Y E R F S S S L L I L P V R E A D - - - - ~ - - A A V Y Y C A V E V ~ t d k l - - - - - - - - - - - - I F C T G T R L P V F p " ~ - - - - - -
R e g i o n E * *** Region F
tants generated by the exonuclease 111 method (HENIKOFF 1984), and the dideoxy method using Sequenase (TABOR and RICHARDSON 1987) were used to sequence all of both strands of the region indicated in Figures 1 and 2. Analysis and presentation of the sequence utilized the FASTA, BLASTS, DNADRAW and MACAW programs (PEARSON and LIPMAN 1988; ALTSCHUL and LIPMAN 1990; SHAPIRO 1990; SCHULER, ALTSCHUL and LIPMAN 1991) and a pat- tern-matching program, FPAT (C. BUCKLER, personal com- munication).
Plasmid constructions: Bacteriophage X was purified by the NaC1-polyethylene glycol method of YAMAMOTO et al . (1970). Subclones of X6662 shown in Figure 1 were made in the centromere-containing shuttle vectors pTI 15 (ICHO and WICKNER 1988) and pRS316 (SIKORSKI and HIETER 1989). The 3.5-kb XbaI-XhoI fragment carrying the MAKlO gene was inserted between the XbaI and XhoI sites of the Bluescript plasmids KS+ and KS- (Stratagene) forming p402 and p400, respectively. These plasmids were the source of single-stranded DNA used for sequencing. To disrupt the MAKlO gene, the 1 .O-kb HindIII-EcoRI fragment within the MAKJO ORF of p402 was replaced with the URA3 gene on a 1.2-kb HindIII-EcoRI fragment from pACl00-2 (kindly provided by J. HILL) to make p406. The disruption was introduced into the diploid strain 2842 by transforming with Xhol-Not1 digested p406 DNA and selecting for Ura+ colonies.
The makJ0::lacZ in-frame fusion genes were made by inserting the 1.9-kb Sall-EcoRI fragment into YEp356R
+ *** R e g i o n C
131 133 116 120 130 145 129 148 131 694
FIGURE 3.-Similarity of MAKlO and T cell receptor a-subunits. The most conserved residues are indi- cated by a "*" while a "+" denotes those that are less well conserved. Dots (. . .) indicate part of the se- quence is left out. Dashes (- - -) indi- cate a gap has been inserted in the sequence. These lineups were pro- duced using MACAW (SCHULER, AL- TSCHUL and LIPMAN 1991). The NBRF database access numbers for the T cell receptor a-subunit V re- gions are shown on the left: A25733, E29774, C25733 (YOSHIKAI et al. 1986); F24402 (BECKER et al. 1985); E24891 (CHOU et al. 1986); D27557 (IWAMOTO et al. 1987); RWMS58 (SAITO et al. 1984); A32537, A32536 (KIMURA et al. 1987). The probability of finding as good or better a three- way match as that among MAKIO, E24891 and A2573 is given by the BLAST3 program as 0.00084.
(MEYERS et al. 1986) cut with the same two enzymes forming pYJ41 or the 2.3-kb EcoRI-EcoRI fragment into EcoRI-cut YEp357 (MEYERS et al. 1986) forming pYJ20. fi-Galactosid- ase was measured in permeabilized cells as described by GUARENTE (1 983).
Southern blots were carried out as described (MANIATIS, FRISCH and SAMBROOK 1982).
RESULTS
Cloning of MAKIO: The slightly slowed growth of malt10 mutants on nonfermentable carbon sources was not useful for selecting or screening clones of the MAKlO gene. However, OLSON et al. (1986) have constructed, in a X vector, a bank of yeast DNA covering most of the genome. Using the known map location of MAKlO on chromosome V between PRBl (protease B) and CYCI (cytochrome CI) (BEVAN and THEIVENDIRARAJAH, cited in WICKNER and LEIBOW- ITZ 1976b; Figure 1, top), we selected X clone X66.52,
that covers the region expected to include MAKlO. Plasmid subclones were introduced into a makl0-1 mutant and the L-A and MI viruses were transferred by cytoplasmic mixing (cytoduction) into the plasmid- bearing makl0-1 strain. If the plasmid had the MAK 10 gene, then L-A and MI would be maintained and the

Yeast MAKlO Gene 91
a
0 0 ) " E 0 Tetrad 1 Tetrad 2 a ? 2 2 1A 1B 1C 1D 2A 28 2C 2D a a
Intact +
Disrupted$
- 9.6 - 6.6 - 4.4
- 2.3 - 2.0
FIGURE 4.-Deletion-insertion of the MAKlO gene. The EcoRI- HindlIl fragment shown in Figure 1 was replaced with a 1.2-kb fragment carrying URA3 bounded by XbaI sites. DNA was extracted from the parent diploid strain 2842, the original disruptant diploid and from the spore clones of two tetrads derived from it (cross 4291). Samples were digested with XbaI and Xhol, analyzed on a 1% agarose gel, blotted to nitrocellulose and hybridized with T 7 RNA polymerase transcripts of p402. In this plasmid the T 7 pro- moter transcribes from the 3' end of the MAKlO gene. The disrup tion construct should produce a 5' fragment of 1.2 kb (hybridizing poorly because the probe should have fewer transcripts in this region) and a 3' fragment of 1.5 kb. The intact fragment is 3.6 kb. Segregants 4291-1A, lB, 2A and 2C were Mak- Ura+.
strain would be a killer. Several subclones of A6662 did complement (Figure I), defining the minimum extent of the gene. The fact that a clone complementing makl0-l has the same physical linkage to PRBZ and CYCl as the MAKlO gene has genetic linkage to these genes shows that this is indeed a clone of the MAKlO gene.
MaklOp has T cell receptor a-subunit motifs: The sequence of this region showed an open reading frame
of 733 amino acids (MAKIO ORF in Figure 1; Figure 2) whose location relative to the region necessary for complementation indicates that it is the MAKlO ORF. Searching the databases with FASTA did not produce any dramatic pairwise matches with known proteins. BLAST3, which detects sequence patterns among groups of proteins, found that Mak 1 Op shares a region that is common among a large number of T cell receptor a-subunit V (variable) regions (region D in Figure 3). Although the T cell receptor a-subunits differ in their V regions, they form a related family. To test the significance of the similarity, the well known common regions among T cell receptor a- subunits were sought using MACAW. The presence of such regions in MaklOp was then tested using the same program. Of the seven such regions in T cell receptor a-subunits, some level of match was found for six in Makl Op, and in the same order in which they were found in the T cell receptor a-subunits (Figure 3).
The consensus of the T cell receptor a-subunits and MaklOp in region D (Figure 3) is [LMF]FWY[LV] .Y. . . .L..[LF][LV]K. Using FPAT to search the pro- tein databases for just part of this, the pattern "FWY[LV].Y," produced matches only with T cell receptor a-subunits (64 matches). In another similar search, all but one of the 45 matches with the pattern "S..YYC.[LV]," the consensus for region F, were with T cell receptor a-subunits or other members of the immunoglobulin family. These two patterns are in distinct parts of the molecules, so their presence in MaklOp are independent events. That either alone is unique to T cell receptor a-subunits indicates that this similarity with MaklOp is significant. Although no proteins showed dramatic similarity to Mak 1 Op, 10 of the top 52 matches using FASTA were with T cell receptor cy-subunits.
maRZO::URA3 disruption mutation: A diploid strain heterozygous for a maklO::URA3 disruption mu-
TABLE 4
makl0::URAJ strains grow slowly on nonfermentable carbon sources
Growth on Growth on Growth on Strain Nuclear genotype Viruses dextrose ethanol glycerol
2842 2842 Bltfl 4291-1A 4291-1B 4291-1C 4291-1D 4303-12A 4303-12B 4303-12C 4303-12D
MAKlOIMAKlO MAKlOlmaklO::URA3 maklO::URA3 maklO::URA3 MAKlO MAKlO maklO::URA3 maklO::URA3 MAKlO MAKlO
L-A-HN MI L-A-HN MI L-A-o M-o L-A-o M-o LA-HN MI L-A-H N MI L-A-o M-o L-A-o M-o L-A-o M-o L-A-o M-o
+++ +++ +++ +++ +++ +++ +++ +++ +++ +++
++ ++ + + ++ ++ + + ++ ++
++ ++ + + ++ ++ + + ++ ++
MAKlO is necessary for optimal growth on glycerol independent of the presence of L-A. Strains were streaked for single colonies on YPAD. YPG or YPE and the size of colonies formed was compared after 2 days. Data shown for crosses 4291 and 4303 are typical of that observed in 7 and 10 tetrads, respectively.
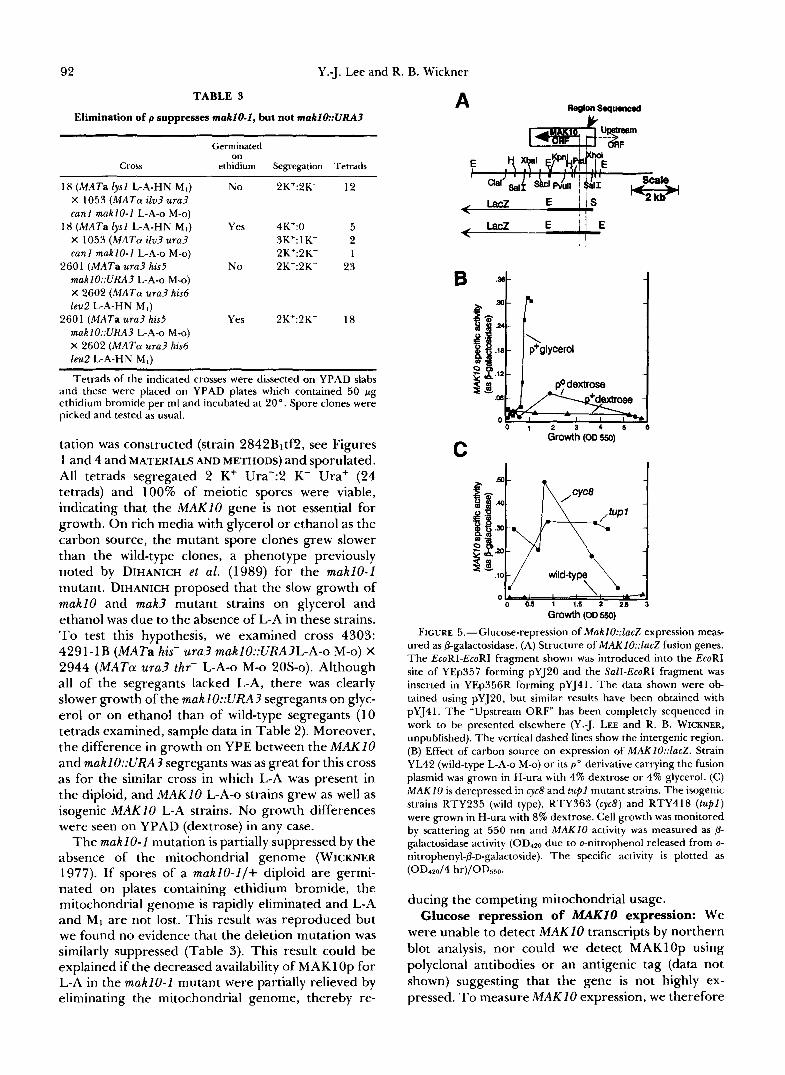
92 Y.-J. Lee and R. B. Wickner
T A B L E 3
Eliminat ion of p suppresses makl0-1, bu t no t maklO::URA3
Germinated on
Cross ethidium Segregation Tetrads
18 (MATa lysl L-A-HN M I ) No 2K+:2K- 12 X 1053 (MATa ilu3 ura3 can1 makl0-1 L-A-o M-o)
18 (MATa lysl L-A-HN M I ) Yes 4 K+: 0 5 X 1053 (MATa ilv3 ura3 3K+: 1 K- 2 can1 makl0-1 L-A-o M-o) 2K+:2K- 1
2601 (MATa ura3 his5 No 2K':2K- 23 maklO::URA3 L-A-o M-o) X 2602 (MATa ura3 his6 leu2 L-A-HN M I )
maklO::URA3 L-A-o M-o) X 2602 (MATa ura3 his6 leu2 L-A-HN M I )
Tetrads of the indicated crosses were dissected on YPAD slabs and these were placed on YPAD plates which contained 50 r g ethidium bromide per ml and incubated at 20". Spore clones were picked and tested as usual.
2601 (MATa ura3 his5 Yes 2K+:2K- 18
tation was constructed (strain 2842Bltf2, see Figures 1 and 4 and MATERIALS AND METHODS) and sporulated. All tetrads segregated 2 K+ Ura-:2 K- Ura' (24 tetrads) and 100% of meiotic spores were viable, indicating that the MAK10 gene is not essential for growth. On rich media with glycerol or ethanol as the carbon source, the mutant spore clones grew slower than the wild-type clones, a phenotype previously noted by DIHANICH et al. (1989) for the makl0-1 mutant. DIHANICH proposed that the slow growth of maklO and mak3 mutant strains on glycerol and ethanol was due to the absence of L-A in these strains. To test this hypothesis, we examined cross 4303: 4291-1B (MATa his- ura3 maklO::URA3L-A-o M-o) X 2944 (MATa u r d thr- L-A-o M-o 20s-0). Although all of the segregants lacked L-A, there was clearly slower growth of the makl O::URA3 segregants on glyc- erol or on ethanol than of wild-type segregants (10 tetrads examined, sample data in Table 2). Moreover, the difference in growth on YPE between the MAKlO and maklO::URA3 segregants was as great for this cross as for the similar cross in which L-A was present in the diploid, and MAKlO L-A-o strains grew as well as isogenic MAKlO L-A strains. No growth differences were seen on Y P A D (dextrose) in any case.
The makl0-1 mutation is partially suppressed by the absence of the mitochondrial genome (WICKNER 1977). If spores of a maklO-l/+ diploid are germi- nated on plates containing ethidium bromide, the mitochondrial genome is rapidly eliminated and L-A and MI are not lost. This result was reproduced but we found no evidence that the deletion mutation was similarly suppressed (Table 3). This result could be explained if the decreased availability of MAKl Op for L-A in the makl0-1 mutant were partially relieved by eliminating the mitochondrial genome, thereby re-
B 1
0 1 2 3 4 5 5 Growth (OD 5 5 0 )
C
d
0 0.5 1 1.6 2 2.1 3 0 -1
Growth (OD sso)
FIGURE 5.-Glucose-repression of Makl0::lacZ expression meas- ured as &galactosidase. (A) Structure of MAK1O::lacZ fusion genes. The EcoRI-EcoRI fragment shown was introduced into the EcoRI site of YEp357 forming pYJ2O and the Sall-EcoRI fragment was inserted in YEp356R forming pYJ41. The data shown were ob- tained using pYJ20, but similar results have been obtained with pYJ41. The "Upstream ORF" has been completely sequenced in work to be presented elsewhere (Y..J. LEE and R. B. WICKNER, unpublished). The vertical dashed lines show the intergenic region. (B) Effect of carbon source on expression of MAK10::lacZ. Strain YL42 (wild-type L-A-o M-o) or its p o derivative carrying the fusion plasmid was grown in H-ura with 4% dextrose or 4% glycerol. (C) M A K I O is derepressed in cyc8 and tup1 mutant strains. The isogenic strains RTY235 (wild type), RTY363 (cycb) and RTY418 (tupl) were grown in H-ura with 8% dextrose. Cell growth was monitored by scattering at 550 nm and MAKIO activity was measured as P- galactosidase activity (OD420 due to o-nitrophenol released from o- nitrophenyl-b-D-galactoside). The specific activity is plotted as (OD4d4 hr)/ODm.
ducing the competing mitochondrial usage. Glucose repression of M A K l O expression: We
were unable to detect MAKlO transcripts by northern blot analysis, nor could we detect MAKlOp using polyclonal antibodies or an antigenic tag (data not shown) suggesting that the gene is not highly ex- pressed. To measure MAK 10 expression, we therefore

Yeast MAKlO Gene 93
constructed makl0::LacZ fusion genes (Figure 5A). D-Galactosidase activity was in general very low,
particularly on glucose media (Figure 5B). In some strains, some activity was observed in late log phase when cells were grown on 2% glucose, with a decrease later in stationary phase, suggesting that MAKlO might be glucose repressed and unstable. Indeed, specific activity was higher in glycerol-grown cells and lower in P O cells grown on glucose. Mutants in tup1 and cyc8 (ssn6) are defective in glucose repression (SCHAMHART, TEN BERGE and VAN DE POLL 1975; ROTHSTEIN and SHERMAN 1980; STARK, FUGIT and MOWSHOWITZ 1980; CARLSON et al. 1984; TRUMBLY 1986; reviewed by JOHNSTON 1987; TRUMBLY 1992) and in functions controlled by the al/a2 and a2 repressors (WICKNER 1974; ROTHSTEIN and SHERMAN 1980; MUKAI, HARASHIMA and OSHIMA 1991). We found that tup1 and cyc8 mutants gave the highest levels of MAKlO activity, even when grown on glucose (Figure 5C). No differences between MATa and MATa strains in makl0::LacZ activity were seen (data not shown). The same glucose-repression of D-galactosid- ase activity was seen with fusions containing the 2.6- kb EcoRI fragment that extends from 1285 bases 5' of the MAKIO translational start site to base 1338 (amino acid 446) of the MAKlO ORF or with fusions of the 2.0 kb from the Sal1 site 719 bases 5' of the MAKlO ORF to the same EcoRI site inside the ORF (Figure 5A).
L-A copy number is negatively controlled by the SKI genes, but no effect of ski2 or ski3 mutations on makl0::LacZ activity was observed on either glucose or glycerol media. Nor was activity affected by the mak3-1 or makl0-1 mutations. The presence or ab- sence of L-A or M, dsRNA likewise had no consistent effect on makl0::LacZ activity. Introducing the MAKlO gene on a high copy vector did not make cells superkillers or affect cell growth.
DISCUSSION
Relation of W I O , the L-A virus, glucose repres- sion and mitochondria: Although neither is necessary for the expression or replication of the other, there are many connections between mitochondrial DNA and the L-A virus system. The absence of mitochon- drial DNA largely suppresses the makl0-1 mutation (WICKNER 1977), but we show here that there is no suppression of a makl0::URA;f disruption. This sug- gests that ,both the mitochondria and L-A use the MAKlO product, and that elimination of mitochon- drial DNA eliminates mitochondrial use making the presumably reduced supply in the makl0-1 strain ad- equate for L-A. Germination of spores on ethanol or glycerol might be expected to suppress makl0-1 by increasing the supply, but this was not the case (data not shown), perhaps because the mitchondrial usage also increases on these carbon sources.
DIHANICH et al. (1989) showed that makZO and mak3 mutants (both of which lose L-A) grow slowly on nonfermentable carbon sources. They also showed that cells growing slowly on glycerol because of a defect in the major outer membrane porin gene ac- cumulate large amounts of L-A virions. It was sug- gested that the presence of the L-A virus helps respi- ration, that its loss in the mak3 and maklO mutants led to the partial respiration defect and its accumulation in the porin mutants was an adaptive response that helps these cells grow. We show here that the growth defect in maklO mutants is not due to its loss of L-A, and the same conclusion has been established for mak3 strains (J. C. TERCERO, L. RILES and R. B. WICKNER, submitted). Furthermore, isogenic strains with and without L-A grow equally well on nonfermentable carbon sources. MAKlO is apparently directly needed for optimum respiration.
Consistent with this idea was the finding that expres- sion of MAKlO was glucose-repressed and was dere- pressed in tup1 and cyc8 mutants. L-A copy number is glucose-repressed (OLIVER et al. 1977), so we consid- ered the possibility that glucose repression of L-A is due to glucose repression of MAKlO. However, the tup1 mutation that derepresses MAKlO in glucose does not derepress L-A (W. R. WIDNER and R. B. WICKNER, unpublished). The MIGl gene product, that actually binds to the upstream regions of target genes, is thought to mediate most glucose repression that in- volves the Tuplp-Cyc8p complex (NEHLIN and RONNE 1990; NEHLIN, CARLBERG and RONNE 199 1). The Miglp recognition sites in the SUC2, GAL4 and GAL1 genes have been determined and shown to be of the form [AT],T[GC][CT]GGGG. There are, how- ever, no sites of this form from the upstream Sal I site to the beginning of the MAKlO ORF. However, at bases 683-689 there is a site for binding of the REBl protein [see Ju, MORROW and WARNER (1990) and references therein], a general transcription factor whose physiological function is not yet clear.
Like the porin defect, absence of the major mito- chondrial nuclease ( n u c l ) results in derepressed L-A replication (LIU and DIECKMANN 1989). Thus, the relation of viral replication with the mitochondria is complex. The MAKlO gene is necessary for optimal mitochondrial function and is needed by L-A, while porin and NUCl are mitochondrial-associated genes that repress L-A copy number.
W I O and viral propagation: Previous studies showed that a MAKlO" mutant could maintain L-A at the permissive temperature, but the L-A dsRNA- containing particles were unstable, consistent with the idea that the MAKlO protein was involved in packag- ing or was even a part of the particle (FUJIMURA and WICKNER 1987). In this study, we tried to detect the MAKlO protein in isolated viral particles without suc- cess (data not shown). In the earlier study, L-A (+)

94 Y . 3 . Lee and R. B. Wickner
sRNA-containing particles were relatively stable (Fu- JIMURA and WICKNER 1987). We have also found that proteins produced from an L-A cDNA expression vector can support M I dsRNA propagation in a maklO-1 host, but cannot support L-A in the same host (WICKNER et al. 1991). These results could be explained if M A K l O were necessary for the stability of full particles, since both L-A (+) ssRNA and MI should occupy less space than L-A dsRNA. A parallel to this is the gene D protein of bacteriophage X, which is necessary for particle stability only of full size X, but not of deletion mutants less than 82% of the size of wild-type X (STERNBERG and WEISBERC 197’7).
Similarity with T cell receptor a-subunits: The presence of several regions of similarity between the M A K l O protein and a class of T cell receptor a- subunits lends credence to the view that the similarity is not coincidental. Searching the databases for the pattern “S..YYC.[LV]” in region F or for the pattern “FWY[LV].Y,” which is only part of the pattern in region D, produces only the T cell receptor a-subunit V regions. The M A K l O protein is much larger than the T cell receptor a-subunit V regions and the re- gions of similarity are spread farther apart in M A K l O . The M A K l O protein does not have a hydrophobic amino terminus and only two moderately hydropho- bic stretches, and our efforts to localize it have thus far been frustrated by the apparently low levels in cells. There is thus no evidence that M A K l O has any membrane association. The similarity to T cell recep- tor a-subunit V regions may signify a distant evolu- tionary connection, but the functional significance, if any, is as yet unclear. The crystal structure of T cell receptors has not yet been determined, but conserved regions among different T cell receptors are believed to form the scaffolding or framework that holds in place the hypervariable complement-determining re- gions. Thus, the similarity noted here may not rep- resent a functional similarity, but rather a structural analogy.
The authors thank LINDA RILES and MAYNARD OLSON for X clones and MARK BOGUSKI and MARK JOHNSTON for helpful discus- sions.
LITERATURE CITED
ALTSCHUL, S. F., and D. J. LIPMAN, 1990 Protein database searches for multiple alignments. Proc. Natl. Acad. Sci. USA
BECKER, D. M., P. PATTEN, Y. CHIEN, T . YOKOTA, Z. ESHHAR, M. GIEDLIN, N. R. J. GASCOIGNE, C. GOODNOW, R. WOLF, K. ARAI and M. M. DAVIS, 1985 Variability and repertoire size of T- cell receptor V-(Y gene segments. Nature 317: 430-434.
BUSSEY, H., 1988 Proteases and the processing of precursors to secreted proteins in yeast. Yeast 4: 17-26.
CARLSON, M., B. C. OSMOND, L. NEIGEBORN and D. BOTSTEIN, 1984 A suppressor of snfl mutations causes constitutive high- level invertase synthesis in yeast. Genetics 107: 19-32.
CHOW, H. S., M. A. BEHLKE, S. A. GODAMBE, J. H. RUSSELL, C. G . BROOKS and D. Y. LOH, 1986 T cell receptor genes in an alloreactive CTL clone: implications for rearrangement and
87: 5509-5513.
germline diversity of variable gene segments. EMBO J. 5
DIHANICH, M., E. VAN TUINEN, J. D. LAMBRIS and B. MARSHALL- SAY, 1989 Accumulation of viruslike particles in a yeast mu- tant lacking a mitochondrial pore protein. Mol. Cell. Biol. 9:
DINMAN, J. D., T. ICHO and R. B. WICKNER, 1991 A-1 ribosomal frameshift in double-stranded RNA virus of yeast forms a gag- pol fusion protein. Proc. Natl. Acad. Sci. USA 88: 174-178.
DMOCHOWSKA, A,, D. DIGNARD, D. HENNING, D. Y. THOMAS and H. BUSSEY, 1987 Yeast K E X l gene encodes a putative pro- tease with a carboxypeptidase B-like function involved in killer toxin and (Y factor precursor processing. Cell 5 0 573-584.
ESTEBAN, R., T . FUJIMURA and R. B. WICKNER, 1988 Site-specific binding of viral plus single-stranded RNA to replicase-contain- ing open virus-like particles of yeast. Proc. Natl. Acad. Sci.
ESTEBAN, R., T . FUJIMURA and R. B. WICKNER, 1989 Internal and terminal cis-acting sites are necessary for in vitro replication of the L-A double-stranded RNA virus of yeast. EMBO J. 8: 947- 954.
FUJIMURA, T., and R. B. WICKNER, 1987 L-A double-stranded RNA viruslike particle replication cycle in Saccharomyces cere- uisiae: particle maturation in vitro and effects of maklO and pet18 mutations. Mol. Cell. Biol. 7: 420-426.
FUJIMURA, T., and R. B. WICKNER, 1988a Replicase of L-A virus- like particles of Saccharomyces cerevisiae. In vitro conversion of exogenous L-A and MI single-stranded RNAs to double- stranded form. J. Biol. Chem. 263: 454-460.
FUJIMURA, T., and R. B. WICKNER, 198813 Gene overlap results in a viral protein having an RNA binding domain and a major coat protein domain. Cell 5 5 663-671.
FUJIMURA, T., and R. B. WICKNER, 1989 Reconstitution of tem- plate-dependent in vitro transcriptase activity of a yeast double- stranded RNA virus. J. Biol. Chem. 264 10872-10877.
FUJIMURA, T., R. ESTEBAN, L. M. ESTEBAN and R. B. WICKNER, 1990 Portable encapsidation signal of the L-A double- stranded RNA virus of S. cerevisiae. Cell 62: 819-828.
FULLER, R. S., A. J. BRAKE and J. THORNER, 1989 Intracellular targeting and structural conservation of a prohormone-proc- essing endoprotease. Science 2 4 6 482-486.
GUARENTE, L., 1983 Yeast promoters and lacZ fusions designed to study expression of cloned genes in yeast. Methods Enzymol. 101: 181-191.
HENIKOFF, S., 1984 Unidirectional digestion with exonuclease 111 creates targeted breakpoints for DNA sequencing. Gene 28:
ICHO, T. , and R. B. WICKNER, 1988 The M A K l l protein is essential for cell growth and replication of M double-stranded RNA and is apparently a membrane associated protein. J. Biol. Chem. 263: 1467-1475.
ICHO, T., and R. B. WICKNER, 1989 The double-stranded RNA genome of yeast virus L-A encodes its own putative RNA polymerase by fusing two open reading frames. J. Biol. Chem.
ITO, H., Y. FUKUDA, K. MURATA and A. KIMURA, 1983 Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 153: 163-168.
IWAMOTO, A,, P. S. OHASHI, H. PIRCHER, C. L. WALKER, E. E. MICHALOPOULOS, F. RUPP, H. HENCARTNER and T . W. MAK, 1987 T cell receptor variable gene usage in a specific cyto- toxic T cell response. Primary structure of the antigen-MHC receptor of four hapten-specific cyotoxic T cell clones. J. Exp. Med. 165: 591-600.
JOHNSTON, M., 1987 A model fungal gene regulatory mechanism: the GAL gens of Saccharomyces cerevisiae. Microbiol. Rev. 51:
Ju, Q., B. E. MORROW, and J . R. WARNER, 1990 REB1, a yeast DNA-binding protein with many targets, is essential for cell growth and bears some resemblance to the oncogene myb. Mol. Cell. Biol. 1 0 5226-5234.
JULIUS, D., A. BRAKE, L. BLAIR, R. KUNISAWA and J. THORNER,
2149-2155.
1100-1 108.
USA 85: 441 1-4415.
351-359.
2 6 4 6716-6723.
458-476.

Yeast MAKlO Gene 95
1984 Isolation of the putative structural gene for the lysine- arginine-cleaving endopeptidase required for the processing of yeast prepro-a-factor. Cell 36: 309-3 18.
KIMURA, N., B. TOYONACA, Y. YOSHIKAI, R. P. Du and T. W. MAK, 1987 Sequences and repertoire of the human T cell receptor alpha- and beta- chain variable region genes in thymocytes. Eur. J. Immunol 17: 375-383.
LEIBOWITZ, M. J., and R. B. WICKNER, 1976 A chromosomal gene required for killer plasmid expression, mating and spore mat- uration in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA
LIU, Y., and C. L. DIECKMANN, 1989 Overproduction of yeast virus-like particle coat protein genome in strains deficient in a mitochondrial nuclease. Mol. Cell. Biol. 9 3323-3331.
MANIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982 Molecular C1oning:A LAboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
MEYERS, A. M., A. TZACOLOFF, D. M. KINNEY and C. J. LUSTY, 1986 Yeast shuttle and integrative vectors with multiple clon- ing sites suitable for construction of lacZ fusions. Gene 4 5
MUKAI, Y., S. HARASHIMA and Y. OSHIMA, 1991 AARl/TUPI protein, with a structure similar to that of the @-subunit of G proteins, is required for al-a2 and a2 repression in cell type control of Saccharomyces cerevisiae. Mol. Cell. Biol. 11: 3773- 3779.
NEHLIN, J. O., M. CARLBERC and H. RONNE, 1991 Control of yeast GAL genes by MIGl repressor: a transcriptional cascade in the glucose response. EMBO J. 1 0 3373-3377.
NEHLIN, J. O., and H. RONNE, 1990 Yeast MIGl repressor is related to the mammalian early growth response and Wilms’ tumour finger proteins. EMBO J. 9 2891-2898.
OLIVER, S. G., S. J. MCCREADY, C. HOLM, P. A. SUTHERLAND, C. S. MCLAUCHLIN and B. S. Cox, 1977 Biochemical and phys- iological studies of the yeast virus-like particle. J. Bacteriol.
OLSON, M. V., J. E. DUTCHIK, M. Y. GRAHAM, G. M. BRODEUR, C. HELMS, M. FRANK, M. MACCOLLIN, R. SCHEINMAN and T. FRANK, 1986 Random-clone strategy for genomic restriction mapping in yeast. Proc. Natl. Acad. Sci. USA 83: 7826-7830.
PEARSON, W. R., and D. J. LIPMAN, 1988 Improved tools for biological sequence analysis. Proc. Natl. Acad. Sci. USA 85: 2444-2448.
KHEE, S.-K., T. ICHO and R. B. WICKNER, 1989 Structure and nuclear localization signal of the SKI3 antiviral protein of Saccharomyces cereuisiae. Yeast 5 149-158.
RIBAS, J. C., and R. B. WICKNER, 1992 RNA-dependent RNA polymerase consensus sequence of the L-A double-stranded RNA virus: definition of essential domains. Proc. Natl. Acad. Sci. USA 8 9 2 185-2 189.
RIDLEY, S. P., S. S. SOMMER and R. B. WICKNER, 1984 Superkiller mutations in Saccharomyces cerevisiue suppress exclusion of M2 double-stranded RNA by L-A-HN and confer cold-sensitivity in the presence of M and L-A-HN. Mol. Cell. Biol. 4: 761-770.
ROEBROEK, A. J., J. A. SCHALKEN, J. A. LEUNISSEN, C. ONNEKINK, H. P. BLOEMERS and W. J. VAN DE VEN, 1986 Evolutionary conserved close linkage of the c-fes/fps proto-oncogene and genetic sequences encoding a receptor-like protein. EMBO J.
ROTHSTEIN, R. J., 1991 Targeting, disruption, replacement and allele rescue: integrative DNA transformation in yeast. Meth- ods Enzymol. 194: 281-301.
ROTHSTEIN, R. J., and F. SHERMAN, 1980 Genes affecting the expression of cytochrome c in yeast: genetic mapping and genetic interactions. Genetics 94: 87 1-889.
SAITO, H., D. M. KRANZ, Y. TAKACAKI, A. C. HAYDAY, H. N. EISEN and S. TONECAWA, 1984 A third rearranged and expressed gene in a clone of cytotoxic T lymphocytes. Nature 312: 36- 40.
SCHAMHART, D. H. J., A. M. A. TEN BERCE and K. W. VAN DE POLL, 1975 Isolation of a catabolite repression mutant of yeast as a revertant of a strain that is maltose negative in the
73: 2061-2065.
299-3 10.
130 1303-1309.
5: 2197-2202.
respiratory-deficient state. J. Bacteriol. 121: 747-752. SCHULER, G. D., S. F. ALTSCHUL and D. J. LIPMAN, 1991 A
workbench for multiple alignment construction analysis. Pro- teins Struct. Funct. Genet. 9 180-190.
SHAPIRO, M., 1990 DNAdraw-a program for drawing DNA se- quences. Binary 2: 187-1 90.
SHERMAN, F., 1991 Getting started with yeast. Methods Enzymol.
SIKORSKI, R. S., and P. HIETER, 1989 A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 122: 19-27
SMEEKENS, S. P., and D. F. STEINER, 1990 Identification of a human insulinoma cDNA encoding a novel mammalian protein structurally related to the yeast dibasic processing protease, KEX2. J. Biol. Chem. 265: 2997-3000.
SMEEKENS, S. P., A. S. AVRUCH, J. LAMENDOLA, S. J. CHAN and D. F. STEINER, 1991 Identification of a cDNA encoding a second putative prohormone convertase related to PC2 in AtT20 cells and islets of Langerhans. Proc. Natl. Acad. Sci. USA 88: 340- 344.
SOMERS, J., and E. A. BEVAN, 1969 The inheritance of the killer character in yeast. Genet. Res. 13: 71-83.
SOMMER, S. S., and R. B. WICKNER, 1982 Yeast L dsRNA consists of at least three distinct RNAs; evidence that the non-Mende- lian genes [HOK], [NEX] and [EXL] are on one of these dsRNAs. Cell 31: 429-441.
STARK, H. C., D. FucITand D. B. MOWSHOWITZ, 1980 Pleiotropic properties of a yeast mutant insensitive to catabolite repression. Genetics 9 4 921-928.
STERNBERG, N., and R. WEISBERG, 1977 Packaging of coliphage lambda DNA 11. The role of the gene D protein. J. Mol. Biol. 117: 733-759.
TABOR, S., and C. C. RICHARDSON, 1987 DNA sequence analysis with a modified bacteriophage T7 DNA polymerase. Proc. Natl. Acad. Sci. USA 84: 4767-4771.
TOH-E, A., and Y. SAHASHI, 1985 The PET18 locus of Saccharo- myces cereuisiae: a complex locus containing multiple genes. Yeast 1: 159-172.
TRUMBLY, R. J., 1986 Isolation of Saccharomyces cerevisiae mutants constitutive for invertase synthesis. J. Bacteriol. 166 1123- 1127.
TRUMBLY, R. J., 1992 Glucose repression in the yeast Saccharo- myces cerevisiae. Mol. Microbiol. 6 15-2 1.
WICKNER, R. B., 1974 Mutants of Saccharomyces cereuisiae that incorporate deoxythymidine-5’-monophosphate into deoxyri- bonucleic acid in vivo. J. Bacteriol. 117: 252-260.
WICKNER, R. B., 1977 Deletion of mitochondrial DNA bypassing a chromosomal gene needed for maintenance of the killer plasmid of yeast. Genetics 87: 441-452.
WICKNER, R. B., 1991 Yeast RNA virology: the killer systems, pp. 263-296 in The Molecular and Cellular Biology of the Yeast Saccharomyces, edited by J. BROACH, E. JONES and J. PRINCLE. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
WICKNER, R. B., 1992 Double-stranded and Single-stranded RNA Viruses of Saccharomyces cerevisiae. Annu. Rev. Microbiol. 4 6 347-375.
WICKNER, R. B., and M. J. LEIBOWITZ, 1976a Two chromosomal genes required for killing expression in killer strains of Saccha- romyces cereuisiae. Genetics 82: 429-442.
WICKNER, R. B., and M. J. LEIBOWITZ, 1976b Chromosomal genes essential for replication of a double-stranded RNA plasmid of Saccharomyces cereuisiae: the killer character of yeast. J. Mol. Biol. 105: 427-443.
WICKNER, R. B., and A. TOH-E, 1982 [HOK], a new yeast non- Mendelian trait, enables a replication-defective killer plasmid to be maintained. Genetics 100 159-174.
WICKNER, R. B., T. ICHO, T . FUJIMURA and W. R. WIDNER, 1991 Expression of yeast L-A double-stranded RNA virus proteins produces derepressed replication: a ski- phenocopy. J. Virol. 65: 155-161.
WILLIAMS, F. E., and R. J. TRUMBLY, 1990 Characterization of
194 3-2 1.

96 Y.-J. Lee and R. B. Wickner
TUPl: a mediator of glucose repression in Saccharomyces cere- the presence of polyethylene glycol and its application to large- uisiae. Mol. Cell. Biol. 10 6500-6511. scale virus purification. Virology 4 0 734-744.
WILLIAMS, F. E., U . VARANASI and R. J. TRUMBLY, 1991 The YOSHIKAI, Y., N. KIMURA, B. TOYONAGA and T. W. MAK, CYCB and TUPl proteins involved in glucose repression in 1986 Sequences and repertoire of human T cell receptor Saccharomyces cerevisiae are associated in a protein complex. alpha chain variable region genes in mature T lymphocytes. J. Mol. Cell. Biol. 11: 3307-3316.
and G . TREIBER, 1970 Rapid bacteriophage sedimentation in Communicating editor: M. JOHNSTON YAMAMOTO, K. R., B. M. ALBERTS, R. BENZINGER, L. LAWHORNE
Exp. Med. 164: 90-103.


















