
lib.ugent.belib.ugent.be/fulltxt/RUG01/000/518/853/RUG01-000518853_2010_0001_AC.pdf · This theory...
89
Transcript of lib.ugent.belib.ugent.be/fulltxt/RUG01/000/518/853/RUG01-000518853_2010_0001_AC.pdf · This theory...






Preface This ecological handbook (Part II in a series of handbooks for students in marine
sciences) was developed in the framework of the Belgian-Russian collaboration
project ‘Joint Curriculum Development in Geo-Ecological Surveying in areas of
development of Natural resources’. Part I, a geological handbook, has been written in
2000 and is entitled “Gas and fluids in marine sediments and related phenomena”.
The geological handbook contains a series of exercises on the interpretation of
stratigraphy, high-resolution seismics and side-scan sonar of gas hydrates and mud
volcanoes.
The purpose of the present ecological handbook is:
q To give graduate and undergraduate students the opportunity to put their
theoretical knowledge about ecological principles into practice
q To get acquainted with the ecosystems formed by cold-water corals
This handbook is mainly based on material from Lophelia reefs because they are
very diverse, concerning both faunal and structural diversity. Moreover, deep-water
corals form a very interesting and extremely beautiful deep-sea habitat, which makes
them very suitable for educational purposes.
The handbook provides an introduction to cold-water coral ecosystems and
ecological sampling techniques. Frequently used methods in the study of biodiversity
and community structure are briefly introduced and clarified with examples. Then the
user can apply these methods in the exercises at the end of each chapter. All the
exercises are based on photographs and video footage collected in cold-water coral
reefs off Norway and Ireland:
q The Norwegian oil company STATOIL provided a 56 min. long video film of
the Haltenpipe coral reef, reef A (see Hovland & Mortensen, 1999). The video
footage was recorded with the ROV “Solo” in the summer of 1997 at a depth
of 275-320 m. They reveal a Lophelia reef of 100 m in diameter and 31 m
high, and a large variety of associated megafauna. STATOIL also provided 80
digital stereo photographs from the HDP-reef cluster taken in 1993 and 1997.
q IFREMER, the French Institute for Exploitation of the Sea, provided digital
photographs of Lophelia and its associated fauna on Theresa Mound, Ireland.
These pictures were taken during the CARACOLE cruise with the research
ship L’ Atalante in the summer of 2001 (30 Juli-15 August) using the deep-
water remotely operated vehicle (ROV) Victor 6000 to study coral

occurrences in the Porcupine Seabight, the Porcupine Bank and the Rockall
Trough.
Figure 0.1 Maps showing the origin of the visual material used in this handbook. The left map shows cold-water coral sites (black dots) around the British Isles. The material from IFREMER comes from Theresa Mound, which is a part of the ‘Belgica' mound province on the eastern slope of Porcupine Basin. The map on the right shows the coast of Norway with the Sula Ridge indicated by the yellow square. The STATOIL material was collected on the Haltenpipe-reefs, south of the Sula Ridge.
Porcupine

List of contents
1. Introduction
1.1 Deep-water corals
1.2 Distribution of Lophelia pertusa
1.2.1 Substrate
1.2.2 Water-masses, currents and their influence on food-availability
1.2.3 Seepage of hydrocarbons
1.3 Lophelia and human activities
2. Material and Methods
2.1 Locating and Mapping
2.2 Sampling
2.2.1 Dredge
2.2.2 Grab and boxcorer
2.2.3 Video and photo
3. Species associated with living and dead Lophelia
3.1 Introduction
3.2 Tree of megafaunal life
3.3 Megafaunal groups, their characteristics and examples
3.3.1 Porifera (sponges)
3.3.2 Cnidaria (hydras, jellyfish, sea anemones and corals)
3.3.3 Annelida: Polychaeta (bristle worms)
3.3.4 Mollusca (clams, oysters, squids, octopods and snails)
3.3.5 Arthropoda: Crustacea (crabs, shrimps, lobsters, etc.)
3.3.6 Echinodermata (sea stars, sea urchins, sea cucumbers, brittle
stars and sea lilies/feather stars)
3.3.7 Vertebrata: Pisces (fish)
4. Diversity
4.1 General
4.1.1 How to calculate species diversity
4.1.2 Shannon-Wiener diversity index
4.1.3 K-dominance curve
4.1.4 Rarefaction or Hurlbert’s (1971) expected number of species
4.1.5 Hill’s diversity numbers
4.2 Biodiversity associated with Lophelia reefs
4.2.1 Deep-sea coral reefs versus tropical reefs
4.2.2 Biodiversity within Lophelia reefs

5. Communities
5.1 Definition
5.2 How are communities defined?
5.2.1 Cluster analysis
5.2.2 TWINSPAN or Two Way INdicator SPecies Analysis
5.2.3 Ordination
5.2.4 Example of community identification based on Haltenpipe data
5.3 Characterising communities
5.3.1 Mean and standard deviation
5.3.2 Example of community characterisation based on Haltenpipe data
5.4 Indicator species and habitat preferences
5.4.1 Indicator species
5.4.2 Taxonomic and trophic organization
5.4.3 Example based on Haltenpipe data
6. Conclusions
7. Glossary
8. References

1. Introduction Nowadays, the whole world is acquainted with the beauty and diversity of shallow-
water tropical and subtropical coral reefs. The deep-water reef-forming corals from
the Atlantic region, since long known by local fishermen, are less known to the public.
Nevertheless, the intensive research of the last years points out that these systems
may have diversities similar to those of tropical reefs; and their beauty is anything but
inferior.
1.1. Deep-water corals
Along the NE-Atlantic margin, the dominant reef-forming corals are the scleractinians
Lophelia pertusa (Linnaeus, 1758), Madrepora oculata (Linnaeus, 1758) and
Desmophyllum cristagalli (Milne-Edwards and Haime, 1848) (Rogers, 1999). The
most remarkable feature of deep-water species is the absence of the algal
endosymbionts (zooxanthellae) characteristic for tropical corals. Being independent
from photosynthetic activity of algae, cold-water corals have different environmental
requirements (further discussed below) and a wider bathymetrical and geographical
distribution (Mortensen, 2000).
In the North-Atlantic, the species Lophelia pertusa (family Caryophilliidae, Gray 1846)
is the most important coral framework builder. This species is characterized by a
variable growing habit (Freiwald et al, 1997) and colour, which may be influenced by
oceanographic conditions and the degree of disturbance and turbulence at each site
(Tasker et al, 2001).
Figure 1.1 The scleractinian coral Lophelia pertusa with visible polyps (left) and retracted polyps (right), Theresa Mound, Ireland. Copyright IFREMER
Copyright IFREMER

1.2. Distribution of Lophelia pertusa
The species Lophelia pertusa was firstly recognised in the early nineteenth century
after the recovery of specimens by Scottish fishermen (Wilson, 1979). Up till now,
this coral has been found throughout the North Atlantic but also more southward,
along the coasts of Africa and Brazil. Records show that the species occurs in parts
of the Mediterranean, in the Pacific and the Indian Ocean (Mortensen, 2000 and
Rogers, 1999). Globally, Lophelia can be found at depths from 39 to 3380 m
(Mortensen, 2000) with an optimum between 250 and 450 m (Frederiksen et al,
1992). Water temperature ranges from 4 to 12 degrees (Rogers, 1999) and salinity
from 31.85 to 37 PSU (Practical Salinity Unit) (Strömgren, 1971; Le Danois, 1948
and Freiwald, 1998).
Several hypotheses have been proposed about the habitat preferences of the
species (Rogers, 1999 and Mortensen, 2000):
1.2.1 Substrate
Lophelia needs hard substrates to settle on; this can be morainic boulders, pebbles
or even shell debris and polychaete tubes. The dead coral branches of the first
colony form new settling substrates.
1.2.2 Water-masses, currents and their influence on food-availability
The depth at which Lophelia is found varies largely. On the Norwegian/Scottish shelf
and around the Faroe Islands, the coral occurs most frequently between 200m and
400m depth (Teichert, 1958 & Frederiksen et al, 1992). Around western Ireland and
in the Bay of Biskay, the coral occurs between 200 and 1000m depth (Le Danois,
1948 and Kenyon et al, 1998). Frederiksen et al (1992) studied the occurrence of
Lophelia pertusa off the Faroe Islands. They suggested that the depth distribution of
Figure 1.2 Global distribution of the scleractinian coral Lophelia pertusaaccording to Rogers (1999).

Lophelia could depend on the maximal wave action, as this coral is very fragile and
can tip over at low current speeds. They also stress the importance of temperature
as a limiting factor, as North-Atlantic Lophelia occurs in the presence of cool Atlantic
water. Freiwald (1998) found a relation between the occurrence of the thermocline
and the growth rate of Lophelia in Norwegian water (Stjernsund Sill). Above the
thermocline, the fjord waters show seasonal fluctuations with respect to temperature
and salinity, while below the inflowing Atlantic water it remains stable throughout the
year.
According to Frederiksen et al (1992), the occurrence of Lophelia around the Faroe
Islands is correlated with bottom slopes critical to internal tidal waves. The breaking
of the waves can increase food supply to the corals by resuspension of organic
matter from the bottom and by increasing the vertical nutrient flux through the
thermocline. This flux can in turn increase the phytoplankton production and the
vertical flux of flocculent organic particulate matter.
Mortensen et al (in press) showed that the density of reefs on the Norwegian shelf is
positively correlated with the seafloor topography, with reefs occurring on local
elevations. This could be due to the local increase in encounter rate of food particles
caused by a higher current velocity (Frederiksen et al, 1992). The currents also
remove waste and keep corals and substratum free of sediments, which can impair
feeding and larval settlement.
1.2.3 Seepage of hydrocarbons
Hovland and colleagues proposed a theory stating that Lophelia reefs are formed at
locations characterized by high concentrations of seeping fluids and gasses. These
substances provide the energy and carbon source for deep-water ecosystems that
grow independent from photosynthesis. Hydrocarbons can feed a variety of
microorganisms, on which the Lophelia ecosystem can grow. Indicators for
hydrocarbon seepage are pockmarks and high values of light hydrocarbons like
methane and ethane in the sediment. Gas-charged layers can be detected on
seismic records (e.g. Hovland & Thomsen, 1997; see also Geological handbook).
This theory would explain the conical growth pattern (corals grow symmetrically
around the energy source, instead of linear as expected in the case of current
dependency) of mounds like they can be found in the Norwegian Sula reef and
Haltenpipe reefs (Hovland et al, 1998). However, Lophelia does not grow on every
location with hydrocarbon seepage, so other controlling factors have to be taken into
account.

In addition to the theory of Hovland & Thomsen (1997), Henriet et al (1998)
suggested that the decay of a gas hydrate horizon in the “Magellan” mound province,
Porcupine Seabight, might have fuelled methane seeps, which in turn could support
coral growth.
1.3. Lophelia and human activities
Ecosystems formed by Lophelia have been studied intensively during the last ten
years during programs like ACES (Atlantic Coral Ecosystem Study), Geomound and
Ecomound. This is mainly due to the increasing exploitation of hydrocarbons (oil and
gas) and fish (e.g. fish in figure
1.4) on the continental margins
(Mortensen, 2000). These
activities pose a serious threat to
the survival of Lophelia
ecosystems.
Deep-sea fish trawling nets
break up the coral colonies and
broken lines and entangled nets
keep killing fish, crabs and other
animals that get stuck in them
(ghost fishing). Next to the direct
impacts, the trawling also causes
indirect damage by changing the hydrodynamic and sedimentary conditions. Trawls
resuspend sediment that can clog up the respiratory and feeding structures of
Lophelia and associated fauna downstream. Trawling may also level off the seabed
by scraping off high points, filling in depressions and moving boulders. Since
Lophelia seems to need high points to grow on (see 1.2), trawling can make the
seabed unsuitable for growth (Tasker et al, 2001). Another result of fishing can be an
alteration of the amount of reef- eroding species, which can influence the balance
between reef building and reef-erosion (Rogers, 1999).
The consequences of trawling and other activities can be studied using an ROV or
other remote sensing techniques like side scan sonar, which can reveal trawl marks
on the seafloor. The net on figure 1.3 was found during a dive on Theresa Mound
with ROV Victor 6000 from the IFREMER institute.
As for the hydrocarbon exploitation, the main menaces are:
q Release of mud, sand and water contaminated with chemicals and oil
Figure 1.3 Abandoned net on Theresa Mound, Ireland.
Copyright IFREMER

q Mechanical damage to the seafloor due to construction works and
anchoring
Drilling mud does not only increase the sedimentation rate, which disturbs the
feeding of Lophelia; it also influences calcification, physiology and polyp expansion
and retraction (Rogers, 1999).
Copyright IFREMER
Figure 1.4 Phycis blennoides (forkbeard) at the left is an exploited deep-sea species, found living among Lophelia (Theresa mound, Ireland). The picture on the right is the commercial fish Pollachius virens or saithe (Haltenpipe, Norway).
STATOIL

2. Material and methods
2.1 Locating and mapping
The knowledge of local fisherman has proved to be very useful for locating cold-
water reefs unknown to the scientific world (Wilson, 1979 and Mortensen, 2000). The
subsequent precise positioning and mapping by scientists is done using side scan
sonar images, high resolution sub bottom profilers, seismic profiles and multi-beam
bathymetry (e.g. De Mol et al, in press; Mortensen, 2000; Hovland et al, 1998;
Hovland et al, in press). Coral-banks can be recognized by their anomalously steep
surface gradients and isolated occurrence (Hovland et al, 1998). Chapter II of the
geological handbook (gas and fluids in marine sediments and related phenomena)
gives a detailed overview of the functioning and outputs of side scan sonar, high-
resolution sub bottom profilers and seismic systems.
Copyright IFREMER
Figure 2.2 High resolution sub bottom profile from Hovland Mounds, northern slope of Porcupine Basin, Ireland (Veerle Huvenne). Sub bottom imagesgive information about the upper layers of the seafloor.
Figure 2.1 3D model of Theresa mound, Ireland, based on multi-beam bathymetry. Bathymetry is basically the measurement of depths by recording the travel time from the acoustic source to the seafloor and back. The colours represent differences in height.
Copyright IFREMER
Figure 2.3. Adapted side scan sonar image from the Hovland Mounds, Ireland. The white side of the mounds are directed towards the towfish, the black side is the acoustic shadow (positive sonograph). Side scan sonar images give information about topography and seafloor characteristics. (image Veerle Huvenne)
Cross-section

2.2 Sampling
Sampling is necessary for the interpretation of the acquired data and for detailed
geological and biological investigations. Different gears and methods have been
used during the study of the distribution and morphology of Lophelia and the
composition of the associated fauna. The choice of sampling device depends on a
large number of factors like the available vessel, the location of and access to the
sampling site, the number of samples to be taken, the weather and of course the
available budget. In this chapter, only the devices that are frequently used for
biological sampling will be discussed.
2.2.1 Dredge
A dredge sampler consists of a cutting edge furnished with hardened steel teeth.
During dragging, the edge penetrates into the sediment. Biological or mineralogical
material is collected in a nylon or canvas bag of variable mesh size. Dredging mostly
yields large qualitative samples but these are not representative for a single point
and the finer fraction of the sample may be washed out during recovery (Larson et al,
Figure 2.4 2D high resolution sparker profile (seismic profile) of Theresa Mound (BEL35) and her neighbour (BEL36), Ireland. The upper part of the figure shows the original seismic profile, the lower part shows the interpretation. The vertical scale indicates time in seconds. This Two Way Travel Time (TWT or TTWT) represents the time needed by the sound wave to travel from the sparker to a horizon in thebottom and back (image Ben De Mol).
Theresa

1997). Although dredge samples are/were frequently used in the research regarding
Lophelia reefs (Stetson et al, 1962; Wilson, 1979; Mortensen, 2000; Jensen &
Frederiksen, 1992, De Mol et al, in press, Freiwald et al, 1997), alternative sampling
devices are preferable because of the destructive effects of dredging and the
inadequate sampling of mobile fauna (Gage & Tyler, 1996 and Mortensen, 2000).
2.2.2 Grab and Box-corer
When one aims to study temporal and spatial patterns in the distribution of fauna,
quantitative samples are needed. A basic tool is
the grab sampler (e.g. Van Veen grab, figure 2.5),
which consists of two opposing, articulate scoop-
shaped jaws that are lowered to the bottom in an
open position and are then closed by various trip
mechanisms to retrieve the sample (Larson et al,
1997). A grab sampler can be used for sampling
Lophelia reefs, providing that the closing
mechanism is strong enough to break off coral
branches (Stetson et al, 1962 and Mortensen,
2000). If not, the jaws will not close completely and the sample will be lost. Moreover,
grab sampling is less destructive than dredges and facilitates precise sampling
(Mortensen, 2000 and Gordon et al, 2000), especially in combination with an
underwater video system (TV-grab). This system allows checking the closing of the
grab before retrieval.
Another quantitative sampling device, used in the study of cold-water reefs and
especially the soft sediment around and between the reefs, is the box-corer (Kenyon
et al, in press; Freiwald et al, 1997 and Raes, unpublished data). The box-corer
(figure 2.6) consists of an open-ended stainless steel box attached to a weighted
column and a stabilizing frame. The box penetrates the sediment under influence of
its own weight. A spade, which was held horizontally during descent, closes off the
box upon retrieval (Gage & Tyler, 1996).
The samples of both devices can be seriously biased when used in fine sediments
(Bett et al, 1994 and Blomqvist, 1991) mainly due to the bow-wave effect, skewed
penetration and, in the case of the box-corer, compaction of the sample or core
shortening. Attempts have been made to minimize these biasing effects.
q Bow-wave effect: This effect is caused by water displacement during
the descent, which displaces the top layer of the sediment. This effect
Figure 2.5 Van Veen grab

can already be avoided by slow and gentle lowering of the box-core
towards the sediment surface. In both grab and box-corer, valves can
be used to allow the flow-through of the water during the descent.
These valves will also close off the device after sampling and prevent
the loss of material during ascent.
q Skewed penetration due to bottom currents or ship movements can be
avoided by using a heavier grab or corer. In the case of a box-corer, a
stabilizing frame can be used that touches the bottom before the core.
That way, samples can even be taken on steep slopes.
q Compaction of the sample is related to penetration velocity and core
diameter. This effect is difficult to correct, but it is important to be
aware of the phenomenon (Blomqvist, 1991).
Adaptation of the box-corer towards more efficient sampling in fine deep-sea
sediments (this device is not suitable for coral sampling!) resulted in the multiple
corer (Barnett et al, 1984; Bett et al, 1994, Gage & Tyler, 1996 and Blomqvist, 1991)
with stabilizing frame and a hydraulic system to slow down the penetration. This
corer takes up to nine, practically undisturbed samples at the same time, but these
samples are small and can be used for meiobenthic or microbiological analysis.
2.2.3 Video and Photo
Video footage has proved to be a very useful tool for studying deep-sea in general
and Lophelia reefs in particular (Gage & Tyler, 1996; Mortensen, 2000; Kenyon et al;
in press, Freiwald et al, 1997). Its major advantage is the minimal disturbance and
damage, which makes it a perfect system to study the fragile and slow-growing cold-
water corals. Video recordings were recently also used for monitoring and for
Figure 2.6. The mechanism of a box-corer: the frame touches the seafloor, the box penetrates the sediment and is sealed off by the spade.
Figure 2.7 Picture of a box-core sample from the Belgica cruise (1005 m depth, june 2000) with sediment, Lophelia pertusa and pieces of the sponge Aphrocallistes bocasei . (photo David Van Rooy)

defining the successional stages in deep-sea ecosystems (Gutt & Starmans, 2001).
Vetter and Dayton (1998 & 1999) used video images for studying submarine
canyons. The quality of the images has significantly improved over the last decades,
among other things by using ROV ‘s (Remotely Operated Vehicles), with which
kilometre-long transects with a large amount of high resolution information can be
obtained in a short period of time. The information about bottom sediment type and
epibenthic fauna complements other sampling
techniques like dredges, grabs or corers
(Vorberg & van Bernem, 1998).
In situ samples can be taken with the ROV ’s
robotic arms (figure 2.8). However, ROV ‘s are
very expensive and heavy, so they require a
ship with adapted winches and cables. A
cheaper alternative is the already mentioned
TV-grab, which is easier to operate
(Mortensen, 2000).
Analysis of underwater video images,
however, brings about some difficulties and imperfections.
q Only the recognisable epibenthic megafauna (> 5cm or more, depending on
the height of the camera above the seabed and the quality of the film) can be
identified. A part of these organisms will not be visible on the image because
they can be hidden between branches, under rocks, etc. Accordingly, one has
to anticipate an underestimation of both diversity and density.
q When identifying living organisms, one has to take into account the different
appearances of an organism (e.g. coral with or without its polyps sticking out).
Moreover, only one side of the animal is visible on film. It can therefore be
difficult to identify an organism when the determinative features are situated
on the ventral side of the organism. A similar problem appears while trying to
identify sponges: spicules (=skeletal elements) are needed for identification.
Morphology alone does not suffice.
q The density of individuals of a species can be so high on a certain location
Figure 2.8 In situ sampling with arm of ROV Victor during the Caracole cruise (2001), Theresa Mound, Ireland.
Copyright IFREMER
Figure 2.9. Field of crinoids from Haltenpipe, Norway
STATOIL

that discerning the separate organisms and counting them can become a
difficult task (e.g. crinoids in figure 2.9).
q The size of the recorded area will not be the same along the whole transect,
due to vertical movements of the camera or the ROV, and changes in towing
speed. Therefore, standardizing the number of organisms will be impossible,
unless some measure of the size of the area is included on the image. A lead
weight (e.g. TTR-TVAT) or laser beams (Gutt & Starmans, 2001 and
Starmans et al, 1999) can indicate the width of the video line.
q In order to define zonation patterns using video footage, the trajectory of the
ROV or the camera has to be established using GPS (Global Positioning
System).
In addition to, or apart from video images, still photographs can be taken (Stetson,
1962; Gutt & Starmans, 2001 and Starmans et al, 1999). These photos can facilitate
the identification of the megafauna and usually have a higher resolution than
underwater video (Gordon et al, 2000). Systems like ‘Bathysnap’ (Gage & Tyler,
1996), which are put on the seafloor for a long period of time and take pictures at
regular intervals, allow the determination of temporal changes in deep-sea
environments.

3. Species associated with living and dead Lophelia
3.1 Introduction
The main aim of this chapter is to briefly introduce the large megafaunal taxa and
their characteristics to non-biologists and to bring you into contact with some
representative species that were observed on Lophelia reefs. In that way, you will be
able to extract more information from images and species lists, and you will better
understand the ecological exercises.
As there are large differences in faunal composition between different sites,
examples will be given from both Norwegian reefs (in particular the Haltenpipe reef)
and Irish reefs (in particular Theresa Mound).
3.2 Tree of megafaunal life (adapted from Ruppert & Barnes, 1994)
Figure 3.1 A simplified version (only megafaunal taxa) of the evolution of animal life. The groups at the base of the tree (e.g. sponges) are the most primitive. The blue groups will be discussed in detail in the following text.

3.3 Megafaunal groups, their characteristics and examples
3.3.1 Porifera (sponges)
Sponges are the most primitive of
multicellular organisms. They lack
organs but they have a well-
developed connective tissue that
builds up the sessile body around a
system of water canals. The water
flow through the body, created by the
beating of flagella, provides oxygen
and food and removes waste, sperm
and larvae.
Feeding type: mainly filter feeders
Taxonomy: subdivision mainly based
on shape and composition of the
skeletal elements => calcareous,
siliceous (glass sponges), spongin
fibers or combinations of the former
3.3.2 Cnidaria (hydras, jellyfish, sea anemones and corals)
Cnidarians are aquatic, radially symmetrical animals with tentacles encircling the
mouth. Food is caught using tentacles and is immobilized by specialized cells
(cnidocytes). Unlike the sponges, these animals do have a gut. Cnidarians can
exhibit two body forms: the pelagic medusa (jellyfish) and the benthic, sessile polyp.
Feeding type: carnivores – suspension feeders
Taxonomy: subdivision based on importance of one of the two body forms (medusa
or polyp) and on morphological features (e.g. type of gametes and cnidocytes,
number of tentacles, presence and characteristics of a skeleton).
The species Lophelia pertusa, the key species in Atlantic deep-water reefs, is a
scleractinian coral with a branching calcium carbonate skeleton. Lophelia is a
pseudocolonial species, meaning that it produces a skeleton by cloning but the
polyps are not interconnected with a neural network (Mortensen, 2000). Little is
known about reproduction, larval biology and recruitment (Rogers, 1999).
Figure 3.2. The upper species is the glass sponge Aphrocallistes bocasei, which is a dominant element in the biological assemblage of Theresa Mound, Ireland. The left picture shows Mycale lingua, which can overgrow Lophelia colonies (Haltenpipe, Norway).
Copyright IFREMER
STATOIL

3.3.3 Annelida: Polychaeta (bristle worms)
The body form of polychaetes varies widely, reflecting a range of life styles from
swimming, through crawling, to active burrowing or tube dwelling. The generalized
polychaete body plan is segmented, with each segment bearing a pair of lateral
appendages (parapods). The anterior end of the worm is the prostomium, which
bears sense organs. The mouth is situated ventrally, following the prostomium. The
terminal unsegmented region, the pygidium, carries the anus.
A large number of species have already been found in samples from Lophelia reefs
(e.g. Eunice norvegica, which shows an interspecific relationship with Lophelia
pertusa (Mortensen, 2000)). In video-footage and still photographs, only the biggest
species can be discerned and those are mainly belonging to the group of Sabellidae
or fan worms, which are filter feeders and tube builders.
Feeding type: carnivores, filter feeders, herbivores, omnivores, scavengers, browsers
and deposit feeders.
Taxonomy: subdivisions are based on morphological adaptations to the type of
feeding and locomotion (e.g. shape of parapods, presence of gills, shape of
prostomial appendages, presence of jaws) The sabellid worm Sabella pavonina can be seen on video segment 4 on the CD in the back of the
handbook.
3.3.4 Mollusca (clams, oysters, squids, octopods and snails)
Moluscs generally have a muscular foot, a calcareous shell that is secreted by the
underlying body wall (the mantle), and a feeding organ (the radula). They can
possess several pairs of gills in their mantle cavity.
Figure 3.3. A. The stony coral Lophelia pertusa, note the redfish
Sebastes viviparus (Haltenpipe, Norway) B. The octocoral (8 tentacles) Paragorgia arborea, with
red and white variety (left) from Haltenpipe, Norway C. A little anemone on octocoral Paramuricea placomus ?
(Theresa Mound, Ireland) D. Two anemones (Bolocera thuediae?) – in the middle
the crab Lithodes maja (Haltenpipe, Norway)
A B C
D
STATOIL STATOIL
STATOIL
Copyright IFREMER

On the investigated video footage and photographs from the Haltenpipe A reef in
Norway and Theresa Mound in Ireland, only representatives of the subgroups
Gastropoda (snails) and Bivalvia (clams) were encountered.
Gastropoda usually have a coiled shell and they use their foot for locomotion.
Feeding type: highly variable (herbivores,
carnivores, scavengers, deposit feeders,
suspension feeders and parasites).
Taxonomy: subdivisions are based on the
shape of the shell and on internal features like
the number and position of gills, the shape of
the teeth on the radula etc.
Bivalvia are characterized by a
reduced head and a laterally compressed
body covered by a bivalved shell. They do not
possess a radula because they are mostly
filterfeeders. Their foot is compressed and
adapted for burrowing.
Feeding type: deposit feeders or filter feeders
Taxonomy: the different groups can be
recognized by the shape of their shells
3.3.5 Arthropoda: Crustacea (crabs, shrimps, lobsters, etc.)
Crustaceans are mainly marine
organisms with 5 pairs of head
appendages, a segmented trunk, which
can be covered with a shield (= the
carapace) and trunk appendages; all
with varying degrees of specialization.
Within the megafauna, only benthic
Decapods (with five pairs of legs) were
observed on film and photographs
(figures 3.3 and 3.5). Their first pair of
legs, like in the genus Munida, can be
enlarged and used for grabbing prey.
Decapod gills are very well enclosed in the carapace.
Feeding type: predators and scavengers
Figure 3.4. The upper picture shows an unidentified gastropod on Lophelia, characterized by its coiled shell (Theresa Mound, Ireland). The lower picture shows a cluster of bivalves of the species Acesta excavata (Haltenpipe, Norway). See also video fragment 1 on the CD: Acesta excavata.
Copyright IFREMER
STATOIL
Figure 3.5. Decapods from Theresa Mound, Ireland. The picture shows one crab (bottom) and two squat lobsters from the genus Munida. This animal is also very common in Norwegian reefs.
Copyright IFREMER

Taxonomy of Decapoda: identification based on shape of legs and carapace,
development of the abdomen, shape of antennae, etc.
3.3.6 Echinodermata (sea stars, sea urchins, sea cucumbers, brittle stars and
sea lilies / feather stars)
Echinoderms are marine bottom dwelling organisms with a pentamerous radial
symmetry. All groups have a skeleton that consists of articulating or fused calcareous
plates that bear spines or tubercles.
Asteroidea or sea stars mostly have five arms, which are grading into a
central disc. They can move using their large number of
tube feet at the ventral side of the arms. This group is
also known for its high regenerative power: arms can
grow back and even a new star can grow from one arm
with a piece of disc.
Feeding type: most species are carnivores and
scavengers
Taxonomy: based on features like the shape of the tube
feet, skeletal plates, disc and spines
Ophiuroidea or brittle stars differ from sea stars with their long arms that are
more sharply set off from the disc. The arms are more solid and the tube feet only
play a little role in locomotion. The mouth is situated on the ventral side of the disc
and is surrounded by large plates and five jaws for chewing.
Feeding type: scavenging, deposit feeding and filter feeding
Taxonomy: based on presence and shape of papillae on jaws and structure of
skeletal plates
Echinoidea or sea urchins are circular or oval and have no arms. Their
skeleton is fused into a solid armour and covered with movable spines. These
spines, together with tube feet, are used for locomotion.
Figure 3.7. The sea urchin on the right is a specimen of the species Cidaris cidaris, which is commonly found on Lophelia reefs (Haltenpipe, Norway). The picture on the left shows Echinus sp., observed on Theresa Mound, Ireland.
Copyright IFREMER STATOIL
Figure 3.6. On the left a sea star (Asteroidea) and on the right a brittle star (Ophiuroidea), both from Haltenpipe, Norway.
STATOIL STATOIL

Feeding type: sea urchins feed by scraping off detritus or encrusting organisms and
algae, using a complex scraping apparatus: grazing
Taxonomy: identification based on development of scraping system, position of tube
feet, number of spines, etc.
Crinoidea are primitive echinoderms that can be stalked
(sea lilies) or free-living (feather stars). The latter can attach
themselves to the substrate with grasping organs (cirri). Unlike
sea stars, sea urchins and brittle stars, crinoids have an upwards-
directed mouth. Crinoids feed by filtering particles out of the water
column, using a crown of featherlike arms surrounding the mouth.
Feeding type: suspension feeders
Taxonomy: subdivisions are based on the presence/absence of a
stalk, the length of the arms and the shape of the cirri and the
basal structure (centro-dorsal) to which the cirri are attached.
Holothuroidea or sea cucumbers were not observed on
the studied films and pictures, although they are very abundant
on soft deep-sea bottom.
3.3.7 Vertebrata: Pisces (fish)
Fish are aquatic vertebrates that breathe by means of gills. This group comprises
jawless fish (Agnatha), cartilaginous fish (Chondrichthyes, e.g. sharks and rays) and
bony fish
(Osteichthyes,
e.g. cod and
salmon).
Feeding type:
various, but all
the species
observed on
the studied
film and
photographs
are carnivorous.
Taxa: subdivisions are based on the shape of the whole body, of the fins, the scales,
the lateral line system etc. The redfish Sebastes viviparus can be seen on video fragment 2 on the CD, together with the
anemone Bolocera thuediae
Figure 3.8. Crinoids (Hathrometra sarsi) on the underside of a buoy on the Ha ltenpipe reef, Norway.
STATOIL
Figure 3.9. Bony fish A. Lepidion eques (Theresa
Mound, Ireland) B. Angler fish Lophius sp.
(Theresa Mound, Ireland) C. Tusk (Brosme brosme)
from Haltenpipe, Norway
A B
Copyright IFREMER
Copyright IFREMER
C
STATOIL

4. Diversity 4.1 General
Biodiversity is a term used to describe the rich variation of life on earth. Diversity can
concern gene pools, species communities or landscapes, which are respectively
composed of genes, species communities and habitats (Heip et al, 1998).
Hill (1973) defined the diversity of a community as ‘a parameter describing the
complexity of the environment, the interspecific relations and the stability of the
community’.
When describing the diversity of a biological community, one has to consider two
components:
• Species richness representing the number of species encountered in the
investigated community
• Evenness representing the spreading of the individuals in the community
over the different species
Over the years, a high variety of indices and visual plots have been developed for
use in different circumstances and with more emphasis on either species richness or
evenness. When choosing an index or plot to describe a certain community, several
questions have to be asked:
• What is the weight you want to give to dominant and/or rare species in
the community?
• Do you want to compare with other studies and what statistics do you
want to apply?
• Are the samples of the same size and what was the sampling
strategy?
Different diversity-indices summarise slightly different aspects of species
assemblages. Therefore it is useful to calculate and compare different indices
obtained using species abundances from the assemblages (Heip et al, 1998).
4.1.1. How to calculate species diversity? The basis of a community analysis is primarily a matrix containing stations/samples
as columns and species as rows. The entries of the matrix are mostly abundances.
Together with the appropriate formulae, these data in a worksheet (e.g. Microsoft
Excel) can be used for calculating diversities or for making a plot that allows

comparing diversities of different samples or stations. In the following paragraphs,
the most frequently used diversity indices and plots will be briefly discussed. They
will be further elaborated in the exercises.
4.1.2. Shannon-Wiener diversity index:
The Shannon-Wiener index is a measure of the uncertainty in predicting to what
species an individual drawn from a collection of S species and N individuals will
belong (Ludwig & Reynolds, 1988). The higher the index, the higher the biodiversity.
ð Can be used to compare habitats based on different sample sizes
ð Sensitive about rare species
ð Commonly used index in the study of Lophelia’s associated fauna (e.g.
Jensen & Frederiksen, 1992 and Mortensen, 2000)
The evenness can be expressed as: J = H’/ ln S where S represents the number of
species in the sample. Evenness assumes a value between 0 and 1.
4.1.3. K-dominance curve
This method graphically displays the distribution (rare or common) of species in
different samples. The K-dominance or cumulative abundance (i.e. the percentage
made up by the k-th-most dominant species plus all more dominant species) is
with Ni: number of individuals of the species I in
the sample, and N: total number of individuals,
(Ni/N) = pi
)/ln()/(' NNNNH ii∑−=
species abundances
samples
Figure 4.1. Excel spreadsheet with species (first column), samples (first row) and species abundances per sample (entries of matrix).
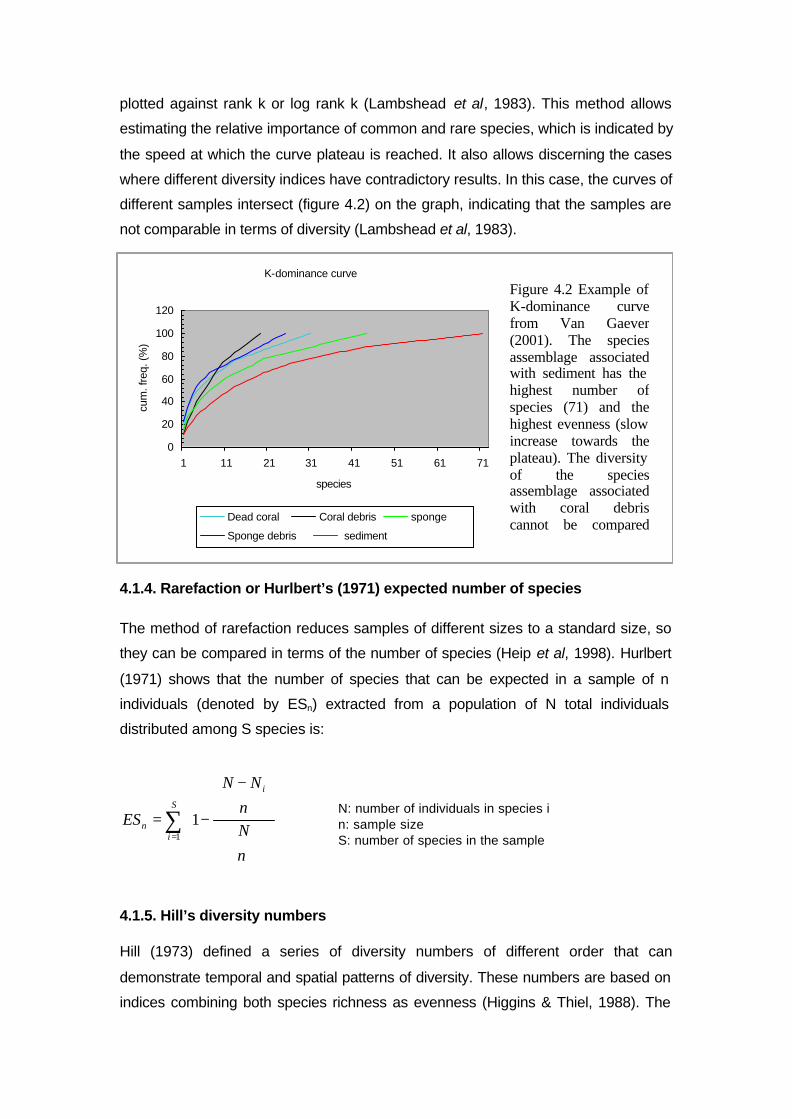
plotted against rank k or log rank k (Lambshead et al, 1983). This method allows
estimating the relative importance of common and rare species, which is indicated by
the speed at which the curve plateau is reached. It also allows discerning the cases
where different diversity indices have contradictory results. In this case, the curves of
different samples intersect (figure 4.2) on the graph, indicating that the samples are
not comparable in terms of diversity (Lambshead et al, 1983).
4.1.4. Rarefaction or Hurlbert’s (1971) expected number of species The method of rarefaction reduces samples of different sizes to a standard size, so
they can be compared in terms of the number of species (Heip et al, 1998). Hurlbert
(1971) shows that the number of species that can be expected in a sample of n
individuals (denoted by ESn) extracted from a population of N total individuals
distributed among S species is:
4.1.5. Hill’s diversity numbers Hill (1973) defined a series of diversity numbers of different order that can
demonstrate temporal and spatial patterns of diversity. These numbers are based on
indices combining both species richness as evenness (Higgins & Thiel, 1988). The
N: number of individuals in species i n: sample size S: number of species in the sample
∑=
−
−=S
i
i
n
n
Nn
NN
ES1
1
K-dominance curve
0
20
40 60
80
100
120
1 11 21 31 41 51 61 71 species
cum
. fre
q. (%
)
Dead coral Coral debris sponge Sponge debris sediment
Figure 4.2 Example of K-dominance curve from Van Gaever (2001). The species assemblage associated with sediment has the highest number of species (71) and the highest evenness (slow increase towards the plateau). The diversity of the species assemblage associated with coral debris cannot be compared with the species

diversity number of order a is defined as: ( ) ( )aan
aa pppN1/1
21 ... +++= with pn the
proportional abundance of species n in the sample. Different diversity numbers can
be used:
ð N0 represents the number of species in the sample. No distinction is made
between rare and dominant species.
ð N1 = exp (H) where H is the Shannon-Wiener index, which attaches less
importance to the rare taxa than N0:
ð N2 = the reciprocal of Simpson’s diversity index:
∑=
=S
iip
D
1
2
1
Simpson’s index, which varies from 0 to 1, gives the probability that two
individuals drawn at random from a population belong to the same species
(Ludwig & Reynolds, 1988).
ð Ninf = the reciprocal of the proportional abundance of the most dominant taxa
and is also named the ‘dominance-index’. This number only takes into
account the most dominant species: Ninf = 1−ip
Soetaert and Heip (1990) tested the sample size dependency of different diversity
indices, including Hill’s diversity numbers. They concluded that high-diversity
communities and the use of indices, which are sensitive to rare species, require
larger samples.
4.2 Biodiversity associated with Lophelia reefs
Several studies on the associated fauna of cold-water corals have already been
carried out, with samples and images from different areas (Mortensen et al, 1995;
)ln()(' ii ppH ∑−=
Number of individuals
Num
ber o
f spe
cies
N0
N1
N2
Ninf
Figure 4.3 Hill’s diversity numbers and sample size dependency (adapted from Soetaert & Heip, 1990). This graph indicates that the indices N0 and N1 are the most dependent on sample size: with increasing sampling effort, more and more rare species will be found. Ninf is independent from sample size: only the most common species, which will also be present in small samples, is considered. Sample size dependency increases with diversity: more rare species, thus larger samples have to be taken in order to estimate diversity.
ES

Jensen & Frederiksen, 1992). A few results and trends seem to be common and will
be briefly discussed in this chapter.
4.2.1 Deep-sea coral reefs versus tropical reefs Generally, Lophelia reefs are considered to be very diverse. In fact, the diversity is
almost equal to that of tropical, hermatypic (containing zooxanthellae) scleractinians
(Jensen and Frederiksen, 1991). This high diversity is mainly caused by the following
factors:
q The complex 3D structure of the corals, which creates a high variety of
suitable habitats for the associated fauna. Mortensen (1995) discerns
four different microhabitats: (1) the smooth surface of living Lophelia,
(2) the detritus laden surface of dead Lophelia, (3) the cavities inside
dead Lophelia made by boring organisms and (4) the space between
the coral branches.
q The age and size of the colonies, and the stability of the deep-sea
environment allowed the development of complex interspecies
interactions and specialisations.
However, the diversity of tropical coral reefs is still higher than that of Lophelia reefs.
This can be explained by the high variety of Scleractinia, each with their associated
fauna. Additionally, deep-sea coral reefs lack macro-algae, which contribute largely
to the structural habitat complexity of tropical reefs and are important food sources
for associated organisms. Finally, tropical reefs are older than deep-water reefs so
there was more time for establishing a high diversity of associated organisms
(Mortensen, 2000).
4.2.2 Biodiversity within Lophelia reefs Mortensen et al (1995) demonstrated that the diversity of megafauna is higher in
areas of dead Lophelia compared to that of living coral. The same pattern appeared
in the case of macrofaunal (smaller, more individuals and more species) diversity
(Jensen & Frederiksen, 1992). The low number of species found on living Lophelia
may be due to antifouling properties of the living tissue; or the living substrate may be
to unstable for settlement. Next to the differences in diversity between living and
dead coral, there is also a remarkable difference between the reefs and the
surrounding sediment. Mortensen (2000) found that the Shannon-Wiener index was
three times higher on the reefs.

5. Communities
5.1 Definition
A community can be defined as “ an assemblage of species populations that occur
together in space and time.” (Begon et al, 1996) or “ the total set of organisms in an
ecological unit (biotope)” (Heip et al, 1998). However, a community is more than only
an assemblage of species in a certain area. An important part of the community
structure is made up by interactions between the species and their populations, like
predation and competition. Examples of how these interactions can influence the
community structure are frequently encountered in tropical coral reefs:
q Anemonefish have a special symbiotic relationship with sea anemones.
Normally, the nematocysts (stinging cells) of an anemone's tentacles would
discharge when the fish brushed against the tentacles, paralyzing the fish. But
anemonefish appear to be at home among the tentacles, even hiding among
them as predators approach. The anemone and the fish are not found
together because they require the same environmental conditions, but
because fish can avoid predation by hiding among anemone tentacles.
q Competition for space is a very important factor in tropical coral
ecosystems. At night, corals can compete for space by reaching out to their
neighbours with long filaments, which literally dissolve the coral tissue next
door, making room for them to expand their own surface area. The threatened
coral will not disappear due to unfavourable environmental conditions but due
to competitive exclusion.
Communities are entities composed of individuals and populations. They have
identifiable collective properties, such as species diversity, community biomass and
productivity (Begon et al, 1996). It is the aim of most ecological studies to verify the
existence of differences in species assemblages and to link them with measured
environmental variables like substrate type, temperature, depth and salinity. If
different communities can be discerned, the ecologist can investigate:
q Habitat preferences of the different faunal groups and species
q Significance of variations in density (numbers of individuals per volume or
surface), biomass, relative abundances (%) and biodiversity over the different
communities

5.2 How are communities defined?
The analysis of communities begins with collecting data on the species assemblages
and the environmental variables of the investigated area. This results in complex
data matrices with information about the encountered species (number of individuals
per volume or surface, see figure 4.1) and different environmental variables (depth,
salinity, current velocity, seafloor characteristics, etc.) per sample like in figure 5.1
where substrate for epifaunal communities is characterized as % debris and
sediment, % living Lophelia and % dead Lophelia.
Finding a pattern in this kind of large matrices containing different variables is very
difficult. Therefore, multivariate analysis is used in ecology because its statistical
methods allow sorting and visualizing data from community studies in an objective
way. The output of these statistical techniques can help the ecologist to find patterns
in species compositions (which samples have the same composition and which
variables are responsible for that?) and to identify indicator species or taxa (which
taxa dominate in which samples?).
There are two essentially different methods: classification and ordination.
Classification is a discontinuous technique that defines groups or clusters. It can, for
example, be used to analyze data from samples taken in an area where sand
patches are clearly distinct from rocky zones (figure 5.2, yellow dots). Ordination
analyzes continuous data and is used to recognize gradients in the environment.
Ordination is, for example, preferred when studying species assemblages along
gradients of height and grain size, in the sediment on a sandy beach (figure 5.2, red
stars). In most cases, both techniques are used to structure the dataset and to
determine whether the data are continuous or discontinuous. The analyses can be
done with presence/absence data of species (e.g. Sørensen index in cluster analysis,
see exercise 2) or with species densities.
Figure 5.1. Part of Excel spreadsheet with environmental variable data matrix representing, per sample, the coverage of the seafloor with living Lophelia, dead Lophelia and coral debris with sediment (based on video Haltenpipe). During processing of the film, each minute was regarded as a sample. This data matrix and the one from figure 4.1 will be used in the example of section 5.2.4 to verify if there are different species communities associated with different substrate types.

In the next pages, more information will be provided about some frequently used
multivariate techniques. These techniques can be applied with programs like PC-
ORD and CANOCO.
5.2.1 Cluster analysis
Cluster analysis is an exploratory data analysis tool for solving classification
problems. Its object is to sort cases (people, samples, stations, etc) into groups or
clusters, so that the degree of association is strong between members of the same
cluster and weak between members of different clusters. Cluster analysis is an
agglomerative method, starting with separate samples and grouping them together
based on their (dis)similarities (figure 5.3). Clusters/samples can then be fused into a
larger unit using different group linkage methods. The Group Average Sorting
method, for example, calculates the mean distance between all samples of one
group and all samples of the other group. The final result is a dendrogram.
species 1 species 2 species 3 species 4
sample 1 35 1 15 0
sample 2 0 18 0 27
sample 3 0 11 0 21
sample 4 22 1 8 0
Figure 5.2. Use of classification and ordination.
compare samples of beach and rocky substrate: there are two classes of substrates but no gradients => classification is preferred. compare samples from the higher to the lower beach. There is a gradient in height => ordination is preferred
sample 3
sample 2
sample 4
sample 1
sample 1 sample 4
sample 2 sample 3
sample 1 sample 4 sample 2 sample 3Figure 5.3
Example of cluster analysis:
agglomerative classification with dendrogram as
result

5.2.2 TWINSPAN or Two Way INdicator SPecies ANalysis TWINSPAN is a divisive classification method, meaning that it starts with a large
group of samples and divides it into smaller groups. Samples and species are
classified simultaneously. The output of TWINSPAN is a two-way ordered table with
species names on the left side of the table and sample numbers along the top. The
patterns of zeros and ones on the right and bottom sides define the dendrogram of
the classifications of species and samples. The interior of the table contains the
abundance class of each species in each sample. Using this table, a dendrogram
can be drawn with divisions of samples and indicator species for each division.
species 1 species 2 species 3 species 4
sample 1 35 1 15 0
sample 2 0 18 0 27
sample 3 0 11 0 21
sample 4 22 1 8 0
TWO-WAY ORDERED TABLE 1423 1 species 11-- 0 3 species 11-- 0 4 species --11 10 2 species 1111 11 0011
5.2.3 Ordination
Ordination is a generic term for multivariate techniques that order samples along
axes based on their species compositions. Samples are ordered in such a way that
those with similar species compositions are grouped together in the graph. There are
two ways to order data. The first method arranges samples using only species data.
For the interpretation, one uses measured environmental gradients and background
knowledge about the species; this is called indirect gradient analysis (e.g. PCA and
CA1, which are respectively based on a linear or unimodal response model). Direct
gradient analysis (e.g. CCA2) integrates environmental data and species
1 PCA: Principal Components Analysis, CA: Correspondence Analysis 2 CCA: Canonical Correspondence Analysis
samples
species
Sample 2 Sample 3
Sample 1 Sample 4
Figure 5.4 Construction of TWINSPAN dendrogram from a two-way ordered table: divisive classification based on presence or absence (1= present and -= absent)
Zeros and ones represent samples on the left and right side of the division. They can be linked to the sample numbers on top of the two-way ordered table: the zeros are the samples 1 & 4 and the ones are the samples 2 & 3.

compositions. However, you have to be sure that the most important structuring
environmental variables are measured.
The axes in an ordination graph represent hypothetical gradients, which are
constructed based on the species compositions. During interpretation, these axes
can be related to gradients in measured variables. The importance of every axis is
expressed in the ‘eigenvalue’, which reflects the percentage of variation in the data
matrix explained by the axis in question. The first axis always accounts for most
variation in the distribution of the data.
You can already try to solve the first two exercises at the end of this chapter. It
might help you to understand the following sections better!
5.2.4 Example of community identification based on Haltenpipe data
This example on community analysis is based on the 56 min. long video film of the
Haltenpipe A coral reef, provided by the Norwegian oil company STATOIL (see
preface). This video was analysed with an SVHS video recorder system (Panasonic
AG-7330) in order to identify the recognisable megafauna to the lowest possible
taxonomic level. The data matrix (every minute of film is regarded as a sample) can
be found in the appendix. Encrusting sponges were disregarded during species
identifications because it was impossible to discern individuals. These sponges were
regarded as a part of the substrate. This means of course that there will be an
underestimation in epifaunal diversity. Colonies of gorgonians were regarded as
individuals.
Samples (minutes of film) were labeled based on the substrate type: samples were
regarded as dead coral samples if there was 25% or more dead coral visible among
the coral debris and sediment. Samples were labeled as living coral samples if they
contained 25% or more living coral between the dead coral, the coral debris and the
sediment. The purpose of this community analysis is to find out if there are
differences in species assemblages depending on the substrate (living coral, dead
coral and debris + sediment).
REMARK! Taking every minute of film as a sample is not an optimal sampling technique. First of all, samples are not taken independently from each other, although independent sampling is a basic condition for the use of many statistical analyses. But more important is the fact that the samples do not have the same sample size due to changes in the speed of the ROV and its height above the seabed.

A. Adapting the data matrix
The species abundances were expressed in percentages (relative abundance in the
sample) in order to limit the effect of varying sample sizes. Since most ecological
data are not normally distributed, it is necessary to transform the data prior to
statistical analysis. In the case of percentage data, an arcsine transformation should
be used.
Three multivariate analysis techniques were applied: (1) cluster analysis using the
Sørensen index as a distance measure and Group Average Sorting as Group
Linkage Method, (2) TWINSPAN analysis
using the presence/absence criterion, and (3)
ordination, more precisely a CA or
Correspondence analysis (based on linear
response model; also known as RA or
Reciprocal Averaging). The initial results
revealed some outliers, which can be
observed in the ordination graph of figure 5.5
(samples in red circle). TWINSPAN analysis
also mentions misclassified samples, i.e.
samples that cannot unequivocally be
assigned to a certain division. Outliers and
misclassified samples were excluded from
further analysis. Furthermore, all species observed in only one sample and
represented by one single individual, were equally removed from the dataset
because they are too rare and usually have no influence on the analysis.
Video fragment 3 on the CD in the back of this handbook shows a part of the Haltenpipe video with the three substrates: coral debris, dead coral and living Lophelia.
CA
Axis 1
Axi
s 2
Figure 5.5 Initial ordination with outliers: indirect gradient analysis

B. Output of cluster analysis and ordination
C. Interpretation
From figure 5.6, we can conclude that cluster analysis shows an inclination towards
grouping the samples with the same seafloor characteristics but this trend is not very
3:00-4:022:00-236:00-7:016:00-1720-00:2115:00-1621:00-2234:00-3535:00-3613:00-14 42:00-4317:00-1826:00-2745:00-4646:00-4751:00-5214:00-1518:00-1919:00-2025:00-2647:00-4844:00-458:00-9:038:00-3912:00-1341:00-4224:00-259:00-10:23:00-2410:00-1111:00-1227:00-2828:00-2933:00:3452:00-5339:00-4029:00-3030:00-3131:00-3240:00-4150:00-51
Mainly samples with species on dead coral
Mixed: living, dead and debris & sediment
Mixed: living and dead
Mixed: dead and debris & sediment
Figure 5.6 Cluster dendrogram (Sørensen index and Group Average Sorting) with indication of different substrate types using different colours: green for dead coral, red for living coral and blue for debris plus sediment.
Dead coral
Living coral
Debris & sediment
Axis 1
Axi
s 2
Figure 5.7 Ordination (CA) from the adapted matrix. Three groups of samples can be discerned. Eigenvalue axis 1: 0.49 Eigenvalue axis 2: 0.34
100% D&S 80% D&S
70% D&S
100%LC
85%LC
40%LC
80%DC
100%LC
50%DC
99%DC

clear. The ordination on the other hand reveals groups with their extreme values on
the outside of the graph (100% living coral, 100% debris & sediment, extremes not as
clear in the case of dead coral) and a mixture of these groups in the center. The clear
results from the ordination are due to the continuity of the dataset. A film reflects
gradual changes in species compositions across the site through which the camera is
moving. So, it could be anticipated from the beginning that a technique like
ordination, which orders samples and species in a continuous multidimensional
space, would be more appropriate for the community analysis.
From the ordination (figure 5.7), we can conclude that the communities change
gradually into 3 extreme species assemblages with increasing importance of each of
the three identified substrate types: the living Lophelia, the dead Lophelia and the
coral debris with sediment.
5.3 Characterizing communities
5.3.1 Mean and standard deviation
If different communities can be identified based on species compositions, the
ecologist will compare other characteristics (density, biomass, productivity, diversity,
etc.) of the communities and he/she will search for significant differences (differences
are significant if they are so big that they cannot result from sampling errors or
coincidence) between the established groups.
A rough idea of differences between communities can be obtained by calculating the
mean and standard deviation of the studied characteristics per community.
1−= ∑
n
xMeanµ : The mean can be calculated as the sum of the values of all
individual observations divided by the total number of observations n. N-1 is used
instead of n because the use of n would result in an underestimation, especially if n is
small.
( )1
2
−
−= ∑
n
xsd
µ: The standard deviation is a measure of how spread out a
distribution is. It is computed as the square root of the average squared deviation of
each number from its mean.
These calculated values can be displayed graphically per community; the
significance of the differences can be verified using more complex statistical
procedures and tests, which will not be further elaborated in this handbook.

5.3.2 Example of community characterization based on Haltenpipe data
The graph in figure 5.8 represents the variation in diversity between the three
communities that were defined in section 5.2.4. This graph indicates that the mean
diversity is the highest in the species community on dead coral (mean ES (100):
8.63) and the lowest in the species community on debris and sediment (mean ES
(100): 5.83). The community with living Lophelia shows an intermediate value (mean
ES (100): 6.55). These data confirm the statements of Mortensen (2000) and Jensen
& Frederiksen (1992) that the diversity is higher in areas of dead coral compared to
areas of living coral (see section 2.2.2), and that the reef shows a higher diversity
than the surrounding sediment (in this case, the sediment is mixed with debris).
The standard deviation gives an indication about the significance of these differences
in biodiversity: when there is a clear overlap between the whiskers, the difference will
probably be non-significant (e.g. between the diversities of the living coral community
and the debris/sediment community).
In this case, non-parametric statistical tests (Mann-Whitney U test and Kruskall-
Wallis test, for those who are familiar with statistical analyses) confirmed that the
difference between the dead coral community and the living coral community is
significant. That was also the case between the dead coral community and the
debris/sediment community but not between the living coral community and the
debris/sediment community.
Figure 5.8 Graph showing mean and standard deviation of the expected number of species per community. The squares represent the mean, the whiskers the standard error.
Mean+SD Mean-SD
Mean
Box Plot expected number of species
ES
__10
0_
3
5
7
9
11
dead
debris/sediment
Living coral coral

5.4 Indicator species and habitat preferences
5.4.1 Indicator species
A very common goal in community analysis is to detect and describe the value of
different species for indicating environmental conditions. These indicators can be
derived from data about the occurrence of species and from background information
about the environmental requirements of the species. The best indicators will be
those species with a narrow range of tolerance concerning a certain environmental
variable.
Example: Indicator species are a widely used tool to monitor the state of habitats
vulnerable to pollution (figure 5.9). Indicator species are species with a very low
tolerance for toxicants, turbidity of the water column or eutrophication. The presence
or absence of indicator species will tell you if the water is polluted or not, while this
information cannot be obtained from the presence of species with a very high
tolerance for pollution.
Species typically occurring in a certain community, which can possibly give an
indication about the environmental conditions, can be searched for with multivariate
analysis (see section 5.2).
Typical species for each community can be found in an ordination graph where the
species are plotted as points representing their optima of the response curve
concerning a certain environmental variable (example in section 5.4.2).
Another technique is called Indicator Species Analysis (Mc Cune & Mefford, 1999).
This method calculates the significance of an indicator by giving it a value between 0
(no indication) and 100 (perfect indication). Two factors are calculated:
Tolerance: response curve
0 20 40 60 80
100 120
0 50 100 150 200 concentration of polluting substance
num
ber
of in
divi
dual
s range
optimum
Figure 5.9 Example of response curve showing tolerance and range of occurrence in the presence of a polluting substance. The concentration that coincides with the highest number of individuals is the optimum.
# individuals of sensitive species # individuals of tolerant species

1. the proportional abundance of the species in a particular group relative to the
abundance of that species in all groups
2. the percentage of samples in each group that contain that species
Example: There are two sites with different environmental conditions; 4 samples
were taken at the first site and 2 samples were taken on the second site.
condition 1 condition 2 sample 1 sample 2 sample 3 sample 4 sample 1 sample 2
species 1 4 5 4 6 8 0 species 2 18 0 0 0 20 36
The first species will be a good indicator for the first environmental condition because
it has a higher abundance under that condition and the species is found in all
samples of the site, which is not the case the second site. Species 2 is a good
indicator for the other environmental condition.
Remark: The presence of an indicator species for a certain environmental variable in
a sample, does not necessarily give you an unequivocal indication about that
variable. If, for example, brittle stars would appear to be indicators for a substrate of
dead coral, it would be
impossible to say what substrate
the sample came from if you only
found one brittle star in your
sample. On the other hand, if
you found that brittle stars were
abundant and dominant in the
sample, then you could say that there is a large possibility that the sample came from
a site with a dead coral substrate.
See also exercise 6
5.4.2 Taxonomic and trophic organization
Community analysis is not always limited to the species level. The characterization of
the faunal communities can also be done on other levels: on a higher taxonomical
level, on the level of trophic organization (feeding types), on the types of locomotion,
etc. These data can give indications about the environmental settings additional to
those obtained from species data and they can provide insights in the functioning of
the different communities.
Figure 5.10 An ophiuroid (Ophiactis abyssicola) from the Porcupine Seabight (photo Kai Kaszemeik, from Van Gaever, 2001)

q Higher taxonomical level: Taxonomy is that field of science that
classifies life in well-defined groups and which is based on the species-
concept. Species are grouped into genera, families, orders, classes, and
phyla, depending upon similarities and inferred evolutionary relationships.
Example: The classification of the crab species Cancer pagurus (Hayward &
Ryland, 1996): Phylum Crustacea
Class Malacostraca
Order Decapoda
Family Cancridae
Genus Cancer
Species Cancer pagurus
The faunal composition of a community can be analyzed on each of these
levels. You can find a video fragment of Cancer pagurus (fragment 5) on the CD.
q Feeding types: Due to the absence of light in the deep-sea, there can
be no primary production from photosynthesis. Therefore, most of the
organisms depend on sinking surface material, mostly phytodetritus (the
remains of dead phytoplankton), faecal pellets and animal carcasses. A lot of
deep-sea animals are suspension feeders, meaning that they capture
particles suspended in the water (e.g.sponges and sabellid worms). Other
animals, the deposit feeders, ingest sediment and digest the nutritional
fraction. Typical examples are sea cucumbers and echiurid worms. A last
group of animals consists of the scavengers, which feed on carcasses, and
predators.
q Types of
locomotion: The deep-sea
megafauna includes both
mobile and sessile
organisms. The former
includes many
echinoderms, mollusks
(inclusing octopods), sea
spiders (pycnogonids), true
crabs, hermit crabs,
shrimps, squat lobsters and
the benthic fishes. The
sessile organisms include
Copyright IFREMER
Figure 5.11 Assemblage of sessile fauna from Theresa Mound, Ireland: little anemones, Lophelia pertusa and a gorgonian octocoral (probably Paramuricea placomus)
Copyright IFREMER

sponges, anemones, corals, sea lilies, barnacles, mussels and ascidians
(Herring, 2002)
5.4.3 Example based on Haltenpipe data
A. Indicator species
The ordination graph on figure 5.13 shows samples and
species simultaneously plotted against the ordination axes.
The position of the species points on the graph can be used
to define indicator species for each of the three
communities. In this case, the following species will probably
be indicators:
• Sebastes viviparus, the unidentified teleost (juvenile
Sebastes?) and the anemone Bolocera tuediae for
the species community on living Lophelia
• Pollachius virens for the species community on coral rubble and sediment
• 2 species of Sabellidae, ophiuroids, Hathrometra sarsi, Cidaris cidaris and
Munida sp. for the species community on dead coral. See Sebastes viviparus and Bolocera tuediae on video fragment 2 on the CD.
The significance of these species as indicators for the three communities was verified
with the technique of Indicator Species Analysis. Only three species seemed to be
significant indicators:
• Sebastes viviparus: highly significant indicator for the species community on
living Lophelia: 86% of the individuals were observed in association with living
Lophelia and 82% of the living coral samples contained Sebastes viviparus.
• Pollachius virens: highly significant indicator for the species community on
coral debris and sediment: 77% of the individuals was observed in
association with coral debris and sediment and all samples (100%) with
debris and sediment are characterized by the presence of saithe (Pollachius
virens).
• Cidaris cidaris: significant indicator for the species community on dead coral:
all individuals (100%) were found associated with dead coral and 33% of the
dead coral samples contained the species Cidaris cidaris.
These three species are indicators for environmental variables in the communities,
respectively for the coverage of the seafloor with living coral, dead coral and coral
Figure 5.12 Norway Redfish (Sebastes viviparus) from Haltenpipe, Norway
STATOIL

rubble plus sediment. Now it is possible to derive the substrate type from data on the
species composition (see exercises).
The species in the center of the graph, like Mycale lingua and Paragorgia arborea,
are common in all three communities. They are certainly not indicators.
B. Taxonomic and trophic organization
The following figures show differences in the amount of sessile and mobile fauna, the
amount of suspension feeders, deposit feeders, scavengers and predators, the
amount of individuals per higher taxon, and the amount of individuals in the 10 most
common species, per community.
1. Figure 5.14 shows the distribution of the relative abundances of the 10
most common species over the three communities.
Axis 1
Mycaling
Axinspec
sponspe4
Geodspec
sponspe6
sponspe8
Poeccomp
Hydrspec
Protsimp
Bolotued
PrimPara
Sabespe1
Sabepavo
Sabespe8
Acesexca
Decaspe
c
Munispec
Cidacida
Hathsars
Astespe1
Ophispec
Sebavivi
Brosbros
Pollvire
Telespec
Axi
s 2
Living coral
Dead coral
Debris & sediment
Figure 5.13 Ordination graph (CA) with samples and species scores. The position of the species can be used to define indicator species for each of the three communities
Paraarbo

The most obvious feature on this graph is the dominance of saithe (Pollachius
virens) and the sponge Mycale lingua in the community associated with debris
and sediment. The living coral community is characterized by Sebastes
viviparus, octocorals, the anemone Prothantea simplex and the sponge
Mycale lingua. The situation is similar in the dead coral community but the fish
Sebastes viviparus is less common.
2. Figure 5.15 shows that the species community associated with coral debris
and sediment is characterized by a higher percentage (mean 77.8%) of
mobile fauna compared to the other communities in which the majority of the
fauna are sessile.
Figure 5.15 Occurrence of mobile and sessile fauna
0% 20% 40% 60% 80%
100%
debris & sediment
Living coral
Dead coral
mea
n p
erce
nta
ge
mobile sessile
0%
20%
40%
60%
80%
100%
Dead coral
Living coral
debris & sediment
rest
Pollachius virens
Sebastes viviparus
Ophiuroidea spec
Hathrometra sarsi
Munida spec
Sabella pavonina
Paragorgia arborea
Primnoa resedaeformis & Paramuricea placomus
Protanthea simplex
Mycale lingua
Figure 5.14 Distribution of the mean relative abundances of the 10 most common species over the three substrate types

3. Figure 5.16 shows the distribution of the feeding types per community. The
living coral community and the dead coral community are again very similar
with 60-80% mean percentage of suspension feeders, while the community
associated with debris and sediment is characterized by almost 75% of
scavengers and predators. Deposit feeders (only Echiuroidea) are scarce.
4. Figure 5.17 shows the distribution of the relative abundance of higher taxa
over the communities. The distribution is different compared to those of the
separate species. In this case the communities on living Lophelia and coral
debris and sediment are the most similar. The dead coral community contains
less fish but more echinoderms and crustaceans.
C. Conclusion The community associated with living Lophelia is characterized by:
• The presence of the indicator species Sebastes viviparus
• Dominance of suspension feeders and sessile fauna
Figure 5.16 Feeding types
0%
20%
40%
60%
80%
100%
Dead coral
Living coral
debris & sediment
mea
n p
erce
nta
ge
scavengers & predators deposit feeders suspension feeders
Figure 5.17 Preferences of higher taxa
0%
20%
40%
60%
80%
100%
dead living debris & sediment
communities
mea
n p
erce
nta
ge
fish
echinoderms
crustaceans
molluscs
Echiuroids
polychaetes
cnidarians
spongescoral
coral

The community associated with dead Lophelia is characterized by:
• The presence of the indicator species Cidaris cidaris
• Dominance of suspension feeders and sessile fauna
• A low abundance of fish and a high abundance of echinoderms and
crustaceans
The community associated with coral debris and sediment is characterized by:
• The presence of the indicator species Pollachius virens
• Dominance of mobile fauna
• Dominance of scavengers and predators

6. Conclusions
The main objective of this handbook was to introduce the concepts of biodiversity
and community analysis to graduate and undergraduate students. The following list
gives an overview of things to remember from this handbook:
The sampling method is an essential element in ecological investigations.
Think carefully before you choose a sampling technique: make it as cheap as
possible but always keep the objectives of your study in mind. Minimize the
disturbance and bias of your samples and choose the sample size carefully.
Your samples must be representative for the studied species community. The
same goes for the measurement of environmental variables: make sure you
measure the most important structuring variables.
Nowadays, there is a large variety in biodiversity indices. When choosing an
index, you have to consider a lot of factors, especially the dependency on
sample size and the weight you want to give to rare or dominant species.
Community analysis is a complex process that can be facilitated using
multivariate and statistical analysis. But keep in mind that statistics are not the
goal in community analyses; they are only a medium to describe the variation
in species assemblages. After processing the data on species and their
abundances, also try looking at your data on other levels, like the level of
trophic organisation, the mobility of the species and higher taxonomical levels.
Another objective of this handbook was to acquaint you with cold-water coral
ecosystems. This was done using exercises and examples based on data from
Norwegian and Irish coral reefs. For those who want to know more or want to see
more beautiful pictures, the following list gives you a few interesting websites:
=> http://www.uni-tuebingen.de/geo/gpi/ag-freiwald/proj/aces/ : ACES project
=> http://www.geomar.de/projekte/ecomound/ecomoundstart: ECOMOUND
=> www.kystnett.no/~emil/skarnsundet/skarnsundet.htm : Trondheimsfjord
=> www.sams.ac.uk/dml/projects/benthic/lophns.htm : Lophelia on oil platforms
=> www.ukbap.org.uk/plans/habitats/NBNSYS0000004650.htm : protection
=> privat.egersund.com/erling/Nesledyr/page4.htm : pictures of species
=> www.pcweb.liv.ac.uk/ocean/org_geochem/aces.html: geochemistry

7. Glossary
ACES: Atlantic Coral Ecosystem Study, an international and multidisciplinary study
focussing on the cold- water corals from Galicia Bank, Spain (43°N), Porcupine
Seabight, Ireland (51°N), Rockall Trough, Ireland (59°N), Kosterfjord, Sweden (59°N)
and the Norwegian Shelf (64°N). This study is situated within the EC fifth framework
programme (1998-2002), just like Ecomound and Geomound. => http://www.uni-tuebingen.de/geo/gpi/ag-freiwald/proj/aces/
Benthic: ‘associated with the seafloor’ – benthic fauna include epibenthos (on the
seafloor), the infauna or endobenthos (in the sediment) and the benthopelagic fauna
(pelagic animals living in close association with the seafloor) (Herring, 2002).
CANOCO: for more information see
=> www.microcomputerpower.com/catalog/canoco.html
Clones: group of cells or organisms arising by asexual reproduction from a single
parent individual. Clones therefore have exactly the same genetic make-up (Lafferty
& Rowe, 1994)
Cnidocytes: stinging structures of cnidarians that are most abundant in tentacles. A
discharged cell consists of a capsule and a thread that can be armed with spines;
many cells can inject a toxin into the prey.
Deposit feeder: deposit feeders ingest sediment, selectively or not. The organic
portion is digested, the rest is eliminated.
ECOMOUND: Environmental Controls on Mound Formation along the European
Continental Margin. The objective of this programme is to define the environmental
controls and processes involved in the development and distribution of carbonate
mounds on the NW European continental margin.
=> http://www.geomar.de/projekte/ecomound/ecomoundstart
Words explained in glossary are underlined in the text

Eutrophication: a state of pollution resulting from introduction of sediments, sewage
or fertilizers into a waterway, causing an increase of the concentration of available
nutrients in that system.
Flagellum: small hairlike organelle on the surface of certain cells (e.g. single-celled
algae and sperm cells of higher animals). The movement of the flagellum can be
used for locomotion of the cell or to create water movements.
Gas hydrate: A gas hydrate is a crystalline solid similar to ice. Its building blocks
consist of a gas molecule surrounded by a cage of water molecules. Many gases
have molecular sizes suitable to form hydrate, including such naturally occurring
gases as carbon dioxide, hydrogen sulphide, and several low-carbon-number
hydrocarbons, but most marine gas hydrates that have been analysed are methane
hydrates.
GEOMOUND: This project focuses on the geological evolution of giant, deep-water
carbonate mounds in the Porcupine Basin and southeast Rockall Basin, off western
Ireland and the UK.
=> http://geomound.ucd.ie/
Lateral line system: system of sense organs in fishes that detects water movement.
It usually consists of a row of interconnected pores on either side of the body that
divide into a system of canals across the head.
Margin: boundary between coastal zone and open ocean, consists of the continental
shelf, slope and rise.
Normal distribution: important class of statistical distributions. All normal
distributions are symmetric and have bell-shaped density curves with a single peak.
The peak of the density coincides with the mean and the standard deviation
indicates the spread of the bell curve. In a normal distribution, 68% of the
observations fall within 1 standard deviation of the mean, that is, between and
.
PC-ORD: for more information see
=> www.ptinet.net/~mjm/pcordwin.htm

Pockmarks: craters developed in soft, fine-grained seafloor sediments by the action
of focused drainage of pore water and gases through the seafloor (Hovland & Judd,
1988)
Population: a group of individuals of one species in an area, though the size and
nature of the area is defined, often arbitrarily, for the purposes of the study being
undertaken (Begon et al, 1996)
Reef: In scientific literature, there has been some debate about the use of the term
‘reef’ for structures built by Lophelia, because Lophelia is an azooxanthellate coral.
Therefore, there are different terms for accumulations of azooxanthellate coral
colonies and skeletons: reef (e.g. Mortensen, 2000), massif (e.g. Le Danois, 1948),
bank (e.g. Jensen & Frederiksen, 1992), patch (e.g. Wilson, 1979), mound (e.g.
Hovland et al, 1994) and bioherm. In this handbook, the term ‘reef’ will be used.
Species: Groups of actually or potentially interbreeding natural populations which are
reproductively isolated from other such groups. May or may not be morphologically
distinct.
STATOIL: Statoil is a Norwegian company carrying out exploration, production,
transportation, refining and marketing of petroleum and petroleum-derived products.
Scleractinians: stony reef-building corals
Sessile: attached to a substrate, not mobile
Swimbladder: a hydrostatic organ used to obtain neutral buoyancy and facilitate
movement in the water column
Symbiosis: a close association between two species generally (with the exception of
parasitism) seeming to be unharmful to either member, or, more likely, beneficial to
one or both. Parasitism, commensalism, and mutualism are the three types of
symbiotic relationships.
Taxon (plural: taxa): unit of classification of living organisms (phylum, class, family,
genus, species)

Thermocline: the depth at which the temperature gradient is a maximum: generally a
layer of water with a more intensive vertical gradient in temperature than in the layers
either above or below it. It is an area of water within the water column in which the
warmer upper waters are prevented from mixing with those at a deeper level. This
barrier prevents the interchange of nutrients between the two.
Zooxanthellae: vegetative stadium of the dinoflagellate Gymnodium
microadriaticum, living inside the tissue of tropical corals. It provides oxygen
(promotes calcification), glucose and glycerol to the coral and uses nutrients
produced by the coral like nitrates, ureic acid and phosphates. These algae allow the
corals to grow faster.

8. References Barnett PRO, Watson J and Connely D (1984) A multiple corer for taking virtually undisturbed samples from shelf, bathyal and abyssal sediment. Oceanologica acta vol. 7(4) 399-408 Begon M, Harper JL & Townsend CR (1996) Ecology. Blackwell Science. 1068p.
Bett BJ, Vanreusel A., Vincx M, Soltwedel T, Pfannkuche O, Lambshead PJD, Gooday AJ, Ferrero T and Dinet A (1994) Sampler bias in the quantitative study of deep-sea meiobenthos. Mar. Ecol. Prog. Ser. Vol. 104: 197-203 Blomqvist S (1991) Quantitative sampling of soft-bottom sediments: problems and solutions. Mar. Ecol. Prog. Ser. Vol 72: 295-304 De Mol B, Van Rensbergen P, Pillen S, Van Herreweghe K, Van Rooij D, McDonell A, Huvenne V, Ivanov M, Swennen R & Henriet JP (in press) Large deep-water coral banks in the Porcupine Basin, southwest of Ireland. 42p Frederiksen R, Jensen A & Westerberg H (1992) The distribution of the scleractinian coral Lophelia pertusa around the Faroe islands and the relation to internal tidal mixing. Sarsia 77: 157-171 Freiwald A, Henrich R and Pätzold J (1997) Anatomy of a deep-water coral reef mound from Stjernsund, west Finnmark, northern Norway: in James NP & Clarke JAD eds., Cool-water Carbonates: Society of Sedimentary Geologists (SEPM), Special Publication, 56, p 140-161 Freiwald A (1998) Geobiology of Lophelia pertusa (Scleractinia) reefs in the North Atlantic. Habilitationsschrift zur Erlangung de venia legendi am Fachbereich Geowissenshaften der Universität Bremen, 116p Gage JD & Tyler PA (1996) Deep-sea biology. A natural history of organisms at the deep-sea floor. Cambridge University Press. 504p Gordon DC, Kenchington ELR, Gilkinson KD, McKeown DL, Steeves G, Chin-Yee M, Vass WP, Bentham K and Boudreau PR (2000) Canadian imaging and sampling technology for studying marine benthic habitat and biological communities. Ices 2000 Annual Science Conference. Gutt J & Starmans A (2001) Quantification of iceberg impact and benthic recolonisation patterns in the Weddell Sea (Antarctica). Polar Biology 24: 615-619 Hayward PJ & Ryland JS (1996) Handbook of the Marine Fauna of North-West Europe. Oxford University Press. 800p Heip CHR, Herman MJP & Soetaert K (1983) Indices of biodiversity and evenness. Océanis 24 (4) 61-87p Heip C, Herman PMJ and Soetaert K (1998) Indices of diversity and evenness. Océanis 24 (4): 61-87

Henriet JP, de Mol B, Pillen S, Vanneste M, Van Rooij D, Versteeg W, Croker PF, Shannon PM, Unnithan V, Bouriak S, Chachkine P & Porcupine-Belgica 97 Shipboard Party (1998) Gas hydrate crystals may help build reefs. Nature 391: 648-649 Herring P (2002) The Biology of the Deep Ocean. Oxford University Press. 314p Higgins RP & Thiel H (1988) Introduction to the study of meiofauna. Smithsonian Institution Press Washington/London. Hill MO (1973) Diversity and evenness: a unifying notation and its consequences. Ecology, 54: 427-432 Hovland M & Thomsen E (1997) Cold-water corals – are they hydrocarbon seep related? Marine Geology, 137: 159-164 Hovland M, Mortensen PH, Brattegard T, Strass P and Rokoengen K (1998) Ahermatypic Coral Banks off Mid-Norway: Evidence for a Link with Seepage of Light Hydrocarbons. Palaios, V.13 189-200p Hovland M & Mortensen PB (1999) Norske korallrev og processer i havbunnen. John Grieg Forlag. 155p Hovland M, Vasshus S, Indreeide A, Austdal L and Nilsen ∅ (in press) Mapping and
imaging deep-sea coral reefs off Norway, 1982-2000
Hurlbert SH (1971) The nonconcept of species diversity: a critique and alternative
parameters. Ecology vol. 52, 4: 577-586
Jensen A & Frederiksen R (1992) The fauna associated with the bank-forming deepwater coral Lophelia pertusa (Scleractinia) on the Faroe shelf. Sarsia 77: 53-63 Kenyon NH, Ivanov MI and Akhmetzanov AM (1998) Cold water carbonate mounds and sediment transport on the Northeast Atlantic margin. Preliminary results of the geological and geophysical investigations during the TTR-7 cruise of R/V Professor Logachev in co-operation with the CORSAIRES and ENAM2 Programmes July-August, 1997. Intergovernmental Oceanographic Commission Technical Series, 52. 179p. UNESCO, Paris, France. Kenyon NH, Akhmetzanov AM, Wheeler AJ, van Weering TCE, de Haas H and Ivanov MI (in press) Giant carbonate mud mounds in the southern Rockall Trough. Marine Geology. Lafferty P and J Rowe (1994) Dictionary of Science. The World of science from Big Bang to Biodiversity, Brockhampton Press, London: 678p Lambshead PJD, Platt HM & Shaw KM (1983) The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. Journal of Natural History, 17: 859-874 Larson R, Morang A and Gorman L (1997) Monitoring the coastal environment 2: sediment sampling and geotechnical methods. Journal of Coastal Research 13 (2) 308-330

Le Danois E (1948) Les profondeurs de la mer. Payot Paris: 303p Ludwig JA & Reynolds JF (1988) Statistical Ecology, a primer on methods and
computing. John Wiley & sons. 85-103p
Mc Cune B & Mefford MJ (1999) PC-ORD. Multivariate Analysis of Ecological data, Version 4. MjM Softawer Design, Gleneden Beach, Oregon, USA. 237p Mortensen, PB, Hovland m, Brattegard T & Farestveit R (1995) Deep-water bioherms of the scleractinian coral Lophelia pertusa (L.) at 64°N on the Norwegian Shelf: structure and associated megafauna. Sarsia 80: 145-158 Mortensen, PB (2000) Lophelia pertusa (Scleractinia) in Norwegian waters – distribution, growth and associated fauna. Thesis submitted in partial fulfilment of the requirements for the degree of Dr. scient. Department of Fisheries and Marine Biology, University of Bergen. Mortensen, PB; Hovland M, Fossa JH and Furevik D (in press) Distribution, abundance and size of Lophelia pertusa coral reefs in mid-Norway in relation to seabed characteristics. Raes M. (2001) Biodiversiteit van meio-epifauna geassocieerd met koudwaterkoralen van de Porcupine Seabight. Licentiaatsthesis Biologie, University of Ghent . 160p Rogers AD (1999) The biology of Lophelia pertusa (Linnaeus, 1758) and other deep-
water reef-forming corals and impacts from human activities. Internat. Rev.
Hydrobiol. 84: 315-406
Ruppert EE & Barnes RD (1994) Invertebrate Zoology sixth edition. Saunders
College Publishing: 1056p
Soetaert K & Heip C. (1990) Sample-size dependence of diversity indices and the
determination of sufficient sample size in high-diversity deep-sea environment.
Marine Ecology Progress Series vol. 59: 305-307
Starmans A, Gutt J and Arntz WE (1999) Mega-epibenthic communities in Arctic and Antarctic shelf areas. Marine Biology 135: 269-280 Stetson TR, Squires DF and Pratt RM (1962) Coral banks occurring in deep water on the Blake Plateau. American Museum of Natural History. Number 2114 Strömgren T (1971) Vertical and horizontal distribution of Lophelia pertusa (Linné) in Trondheimsfjorden on the West Coast of Norway. Kongelige Norske Videnskabers Selskabs Skrifter 6: 1-9

Tasker M, Bett B, Fossa JH, Grehan A, Gordon J, Hall-Spencer J, Lorance P, Mortensen P and Wheeler A (2001) Initial report of the study group on cold water corals in relation to fishing (draft). SGCOR, 12p Teichert C (1958) Cold- and deep-water coral banks. Bull. Am. Ass. Petrol. Geol. 42 (5): 1064-1082 Van Gaever S (2001) Gemeenschapsanalyse van macrofauna geassocieerd met koudwaterkoraalriffen in de NO Atlantische Oceaan. Licentiaatsthesis biologie, University of Ghent. 110p Vetter EW & Dayton PK (1998) Macrofaunal communities within and adjacent to a detritus-rich submarine canyon system. Deep-Sea Research II 45 p 25-54 Vetter EW & Dayton PK (1999) Organic enrichment by macrophyte detritus, and abundance patterns of megafaunal populations in submarine canyons. Mar. Ecol. Prog. Ser. Vol. 186: 137-148 Vorberg R & van Bernem K-H (1998) Application of underwater video and imaging sonar in ecological investigations in the subtidal zone of the Wadden Sea. Arch. Fish. Mar. Res.46 (3), p 195-203 Wilson JB (1979) The first recorded specimens of the deep-water coral Lophelia pertusa (Linnaeus, 1758) from British waters. Bull. Br. Mus. Nat. Hist. (Zool.) 36 (4): 209-215 Wilson JB (1979) The distribution of the coral Lophelia pertusa (L.) (L. prolifera (Pallas)) in the North-East Atlantic. J. Mar. Biol. Ass. U.K. 59, 149-164

a
Appendix The appendix contains the 2 datasets constructed with the observations from the STATOIL film from the Haltenpipe coral reefs:
1) The dataset with samples, species and their abundances (raw data): pages a - e
2) The dataset with the estimations of the percentage seafloor coverage with debris plus sediment, living Lophelia and dead coral: page f

b
1:00-2:00 3:00-4:00 4:00-5:00 5:00-6:00 6:00-7:00 7:00-8:00 8:00-9:00 9:00-10:00 10:00-11:00 11:00-12:00
Mycale lingua (sp1) 5.000 12.500 7.692 25.000 13.333 17.647 7.692 33.333 11.538 8.333
Haliclona sp. (sp2) 0.000 2.083 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Axinella sp. (sp3) 5.000 0.000 30.769 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp4) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Geodiidae sp. (sp5) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp6) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Stellatta sp. (sp7) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp8 flat, white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Poecillastra compressa (sp9) 0.000 0.000 7.692 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Hydrozoa sp. 5.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Prothantea simplex 0.000 22.917 0.000 0.000 26.667 35.294 26.923 0.000 19.231 0.000 Bolocera tuedia 5.000 2.083 0.000 0.000 0.000 0.000 0.000 0.000 0.000 16.667 Unidentified anthozoan (white) 5.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified octocoral sp1(white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Primnoa resedaeformis & Paramuricea placomus 0.000 12.500 0.000 0.000 46.667 14.706 3.846 0.000 38.462 41.667 Paragorgia arborea 5.000 6.250 0.000 0.000 6.667 11.765 7.692 0.000 0.000 0.000 Sabellidae sp1(small, blue) 0.000 0.000 0.000 0.000 6.667 0.000 0.000 0.000 0.000 0.000 Sabella cf. pavonina 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Sabellidae sp2 (red) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified echiurid 0.000 0.000 7.692 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Acesta excavata 0.000 0.000 0.000 0.000 0.000 5.882 0.000 0.000 0.000 0.000 Cancer pagurus 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified crab 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified decapod 0.000 0.000 0.000 25.000 0.000 0.000 0.000 0.000 0.000 0.000 Munida sp. 0.000 2.083 23.077 0.000 0.000 0.000 3.846 0.000 0.000 0.000 Cidaris cidaris 10.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Hathrometra sarsi 20.000 33.333 0.000 0.000 0.000 8.824 15.385 0.000 0.000 0.000 Unidentified Asteroid sp1 35.000 6.250 23.077 50.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified Asteroid sp2 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified Ophiuroid 0.000 0.000 0.000 0.000 0.000 2.941 26.923 0.000 0.000 0.000 Sebastes viviparus 0.000 0.000 0.000 0.000 0.000 2.941 7.692 66.667 30.769 33.333 Brosme brosme 5.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Pollachius vi rens 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Micromesistius potassou 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified teleost 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000

c
12:00-13:00 13:00-14:00 14:00-15:00 15:00-16:00 16:00-17:00 17:00-18:00 18:00-19:00 19:00-20:00 20-00:21:00 21:00-22:00 22:00-23:00
Mycale lingua (sp1) 10.526 54.545 22.222 25.000 23.077 36.842 18.919 12.963 21.429 21.053 13.333
Haliclona sp. (sp2) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Axinella sp. (sp3) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp4) 0.000 0.000 0.000 0.000 5.128 2.632 2.703 0.000 0.000 2.632 0.000 Geodiidae sp. (sp5) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 2.381 2.632 0.000 unidentified sponge (sp6) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Stellatta sp. (sp7) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp8 flat, white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Poecillastra compressa (sp9) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Hydrozoa sp. 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Prothantea simplex 21.053 22.727 35.556 12.500 30.769 31.579 43.243 64.815 35.714 10.526 23.333 Bolocera tuedia 5.263 0.000 0.000 0.000 0.000 2.632 0.000 0.000 0.000 0.000 0.000 Unidentified anthozoan (white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified octocoral sp1(white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Primnoa resedaeformis & Paramuricea placomus 5.263 4.545 6.667 31.250 23.077 2.632 0.000 7.407 30.952 26.316 20.000 Paragorgia arborea 0.000 0.000 0.000 0.000 5.128 5.263 0.000 1.852 2.381 10.526 3.333 Sabellidae sp1(small, blue) 26.316 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Sabella cf. pavonina 0.000 0.000 0.000 6.250 0.000 7.895 5.405 0.000 0.000 0.000 0.000 Sabellidae sp2 (red) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified echiurid 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Acesta excavata 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Cancer pagurus 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified crab 0.000 0.000 0.000 0.000 0.000 0.000 2.703 0.000 0.000 0.000 0.000 unidentified decapod 0.000 0.000 0.000 0.000 0.000 2.632 0.000 0.000 4.762 0.000 0.000 Munida sp. 15.789 9.091 4.444 18.750 0.000 2.632 0.000 5.556 0.000 0.000 0.000 Cidaris cidaris 0.000 0.000 2.222 0.000 2.564 2.632 2.703 0.000 0.000 5.263 0.000 Hathrometra sarsi 0.000 0.000 2.222 0.000 0.000 0.000 0.000 0.000 0.000 0.000 26.667 Unidentified Asteroid sp1 0.000 0.000 4.444 0.000 0.000 0.000 2.703 1.852 0.000 0.000 0.000 Unidentified Asteroid sp2 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified Ophiuroid 10.526 0.000 20.000 6.250 0.000 2.632 18.919 3.704 0.000 2.632 0.000 Sebastes viviparus 5.263 4.545 2.222 0.000 2.564 0.000 0.000 0.000 0.000 2.632 3.333 Brosme brosme 0.000 0.000 0.000 0.000 2.564 0.000 2.703 1.852 2.381 0.000 0.000 Pollachius virens 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 5.263 10.000 Micromesistius potassou 0.000 4.545 0.000 0.000 0.000 0.000 0.000 0.000 0.000 2.632 0.000 unidentified teleost 0.000 0.000 0.000 0.000 5.128 0.000 0.000 0.000 0.000 7.895 0.000

d
23:00-24:00 24:00-25:00 25:00-26:00 26:00-27:00 27:00-28:00 28:00-29:00 29:00-30:00 30:00-31:00 31:00-32:00 32:00-33:00 33:00:34:00
Mycale lingua (sp1) 21.739 8.333 18.182 33.333 33.333 42.593 6.897 5.714 1.724 0.000 12.308
Haliclona sp. (sp2) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Axinella sp. (sp3) 4.348 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 3.077 unidentified sponge (sp4) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 5.556 0.000 Geodiidae sp. (sp5) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp6) 0.000 4.167 0.000 0.000 0.000 1.852 0.000 0.000 0.000 0.000 0.000 Stellatta sp. (sp7) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp8 flat, white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Poecillastra compressa (sp9) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Hydrozoa sp. 0.000 0.000 0.000 0.000 4.167 0.000 0.000 0.000 0.000 0.000 1.538 Prothantea simplex 13.043 8.333 22.727 5.556 4.167 0.000 0.000 0.000 0.000 0.000 10.769 Bolocera tuedia 0.000 0.000 0.000 0.000 0.000 1.852 0.000 0.000 0.000 0.000 0.000 Unidentified anthozoan (white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified octocoral sp1(white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Primnoa resedaeformis & Paramuricea placomus 4.348 12.500 9.091 11.111 0.000 3.704 0.000 0.000 0.000 0.000 13.846 Paragorgia arborea 0.000 0.000 0.000 11.111 0.000 3.704 1.724 2.857 1.724 5.556 3.077 Sabellidae sp1(small, blue) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Sabella cf. pavonina 0.000 33.333 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 3.077 Sabellidae sp2 (red) 0.000 0.000 4.545 5.556 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified echiurid 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Acesta excavata 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Cancer pagurus 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified crab 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified decapod 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Munida sp. 4.348 0.000 0.000 0.000 0.000 1.852 5.172 11.429 1.724 0.000 1.538 Cidaris cidaris 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Hathrometra sarsi 0.000 12.500 4.545 0.000 4.167 1.852 0.000 0.000 0.000 0.000 3.077 Unidentified Asteroid sp1 0.000 0.000 0.000 0.000 0.000 0.000 1.724 2.857 1.724 0.000 0.000 Unidentified Asteroid sp2 0.000 0.000 0.000 11.111 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified Ophiuroid 0.000 0.000 22.727 0.000 0.000 0.000 6.897 0.000 0.000 0.000 0.000 Sebastes viviparus 43.478 12.500 0.000 5.556 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Brosme brosme 4.348 0.000 0.000 0.000 4.167 0.000 0.000 0.000 3.448 0.000 0.000 Pollachius virens 0.000 8.333 18.182 16.667 50.000 42.593 75.862 74.286 89.655 88.889 47.692 Micromesistius potassou 0.000 0.000 0.000 0.000 0.000 0.000 1.724 2.857 0.000 0.000 0.000 unidentified teleost 4.348 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000

e
34:00-35:00 35:00-36:00 36:00-37:00 37:00-38:00 38:00-39:00 39:00-40:00 40:00-41:00 41:00-42:00 42:00-43:00 43:00-44:00 44:00-45:00
Mycale lingua (sp1) 18.605 28.000 9.091 19.355 15.625 24.324 1.695 12.500 50.000 23.810 8.333
Haliclona sp. (sp2) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Axinella sp. (sp3) 0.000 0.000 0.000 3.226 0.000 2.703 0.000 0.000 0.000 0.000 4.167 unidentified sponge (sp4) 2.326 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Geodiidae sp. (sp5) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp6) 0.000 0.000 0.000 3.226 0.000 0.000 0.000 0.000 0.000 0.000 4.167 Stellatta sp. (sp7) 0.000 0.000 0.000 0.000 0.000 5.405 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp8 flat, white) 0.000 0.000 0.000 0.000 0.000 0.000 1.695 6.250 0.000 0.000 0.000 Poecillastra compressa (sp9) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Hydrozoa sp. 0.000 0.000 3.030 0.000 0.000 0.000 0.000 0.000 0.000 0.000 4.167 Prothantea simplex 16.279 6.000 24.242 29.032 18.750 5.405 13.559 6.250 18.750 23.810 29.167 Bolocera tuedia 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 4.762 0.000 Unidentified anthozoan (white) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified octocoral sp1(white) 0.000 0.000 0.000 0.000 3.125 0.000 0.000 0.000 0.000 0.000 0.000 Primnoa resedaeformis & Paramuricea placomus 27.907 34.000 48.485 0.000 0.000 13.514 0.000 0.000 0.000 0.000 0.000 Paragorgia arborea 4.651 4.000 3.030 0.000 0.000 2.703 0.000 0.000 0.000 4.762 8.333 Sabellidae sp1(small, blue) 0.000 0.000 0.000 0.000 6.250 0.000 0.000 0.000 0.000 0.000 0.000 Sabella cf. pavonina 6.977 0.000 0.000 3.226 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Sabellidae sp2 (red) 0.000 0.000 0.000 0.000 0.000 0.000 0.000 6.250 0.000 0.000 4.167 unidentified echiurid 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Acesta excavata 2.326 2.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Cancer pagurus 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified crab 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified decapod 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Munida sp. 0.000 0.000 0.000 12.903 9.375 0.000 0.000 18.750 6.250 0.000 0.000 Cidaris cidaris 0.000 0.000 0.000 0.000 0.000 2.703 0.000 6.250 0.000 0.000 0.000 Hathrometra sarsi 2.326 0.000 0.000 0.000 0.000 0.000 0.000 18.750 6.250 0.000 4.167 Unidentified Asteroid sp1 0.000 0.000 0.000 0.000 0.000 0.000 1.695 0.000 0.000 0.000 0.000 Unidentified Asteroid sp2 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified Ophiuroid 0.000 0.000 0.000 0.000 43.750 0.000 1.695 18.750 0.000 0.000 0.000 Sebastes viviparus 0.000 2.000 9.091 3.226 3.125 5.405 0.000 6.250 6.250 0.000 12.500 Brosme brosme 0.000 4.000 3.030 6.452 0.000 2.703 0.000 0.000 0.000 0.000 0.000 Pollachius virens 18.605 20.000 0.000 19.355 0.000 35.135 79.661 0.000 12.500 42.857 20.833 Micromesistius potassou 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 unidentified teleost 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
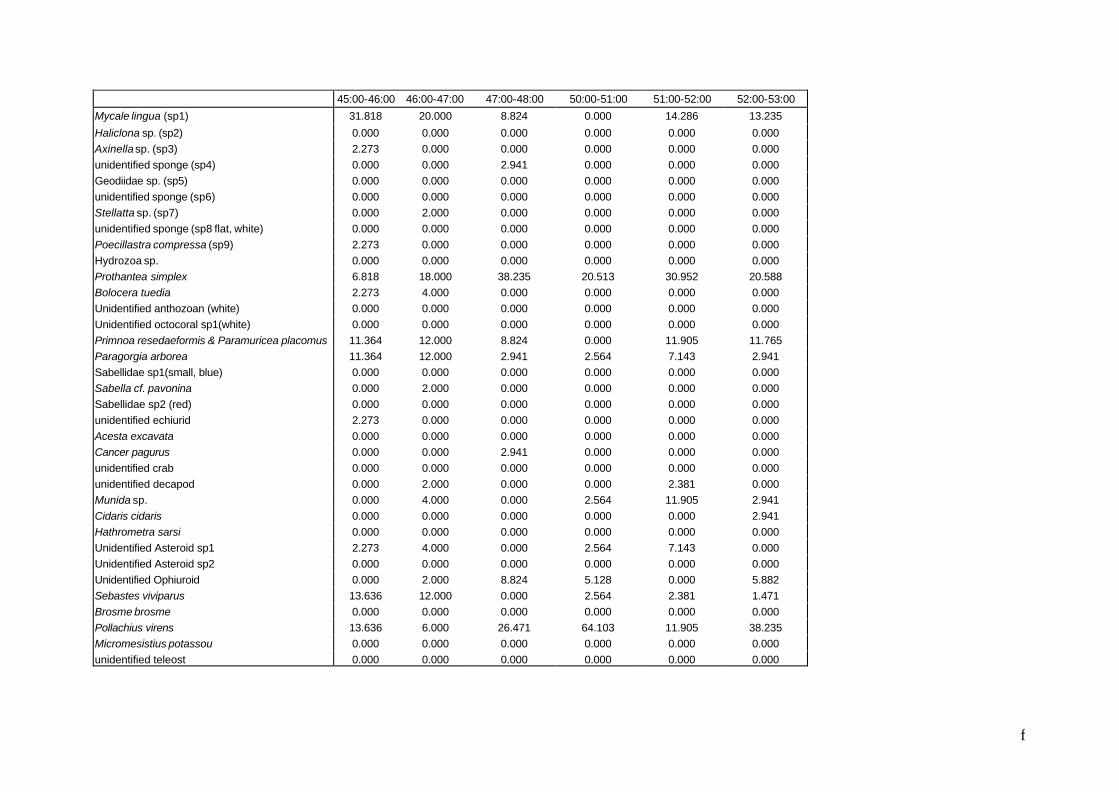
f
45:00-46:00 46:00-47:00 47:00-48:00 50:00-51:00 51:00-52:00 52:00-53:00
Mycale lingua (sp1) 31.818 20.000 8.824 0.000 14.286 13.235
Haliclona sp. (sp2) 0.000 0.000 0.000 0.000 0.000 0.000 Axinella sp. (sp3) 2.273 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp4) 0.000 0.000 2.941 0.000 0.000 0.000 Geodiidae sp. (sp5) 0.000 0.000 0.000 0.000 0.000 0.000 unidentified sponge (sp6) 0.000 0.000 0.000 0.000 0.000 0.000 Stellatta sp. (sp7) 0.000 2.000 0.000 0.000 0.000 0.000 unidentified sponge (sp8 flat, white) 0.000 0.000 0.000 0.000 0.000 0.000 Poecillastra compressa (sp9) 2.273 0.000 0.000 0.000 0.000 0.000 Hydrozoa sp. 0.000 0.000 0.000 0.000 0.000 0.000 Prothantea simplex 6.818 18.000 38.235 20.513 30.952 20.588 Bolocera tuedia 2.273 4.000 0.000 0.000 0.000 0.000 Unidentified anthozoan (white) 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified octocoral sp1(white) 0.000 0.000 0.000 0.000 0.000 0.000 Primnoa resedaeformis & Paramuricea placomus 11.364 12.000 8.824 0.000 11.905 11.765 Paragorgia arborea 11.364 12.000 2.941 2.564 7.143 2.941 Sabellidae sp1(small, blue) 0.000 0.000 0.000 0.000 0.000 0.000 Sabella cf. pavonina 0.000 2.000 0.000 0.000 0.000 0.000 Sabellidae sp2 (red) 0.000 0.000 0.000 0.000 0.000 0.000 unidentified echiurid 2.273 0.000 0.000 0.000 0.000 0.000 Acesta excavata 0.000 0.000 0.000 0.000 0.000 0.000 Cancer pagurus 0.000 0.000 2.941 0.000 0.000 0.000 unidentified crab 0.000 0.000 0.000 0.000 0.000 0.000 unidentified decapod 0.000 2.000 0.000 0.000 2.381 0.000 Munida sp. 0.000 4.000 0.000 2.564 11.905 2.941 Cidaris cidaris 0.000 0.000 0.000 0.000 0.000 2.941 Hathrometra sarsi 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified Asteroid sp1 2.273 4.000 0.000 2.564 7.143 0.000 Unidentified Asteroid sp2 0.000 0.000 0.000 0.000 0.000 0.000 Unidentified Ophiuroid 0.000 2.000 8.824 5.128 0.000 5.882 Sebastes viviparus 13.636 12.000 0.000 2.564 2.381 1.471 Brosme brosme 0.000 0.000 0.000 0.000 0.000 0.000 Pollachius virens 13.636 6.000 26.471 64.103 11.905 38.235 Micromesistius potassou 0.000 0.000 0.000 0.000 0.000 0.000 unidentified teleost 0.000 0.000 0.000 0.000 0.000 0.000
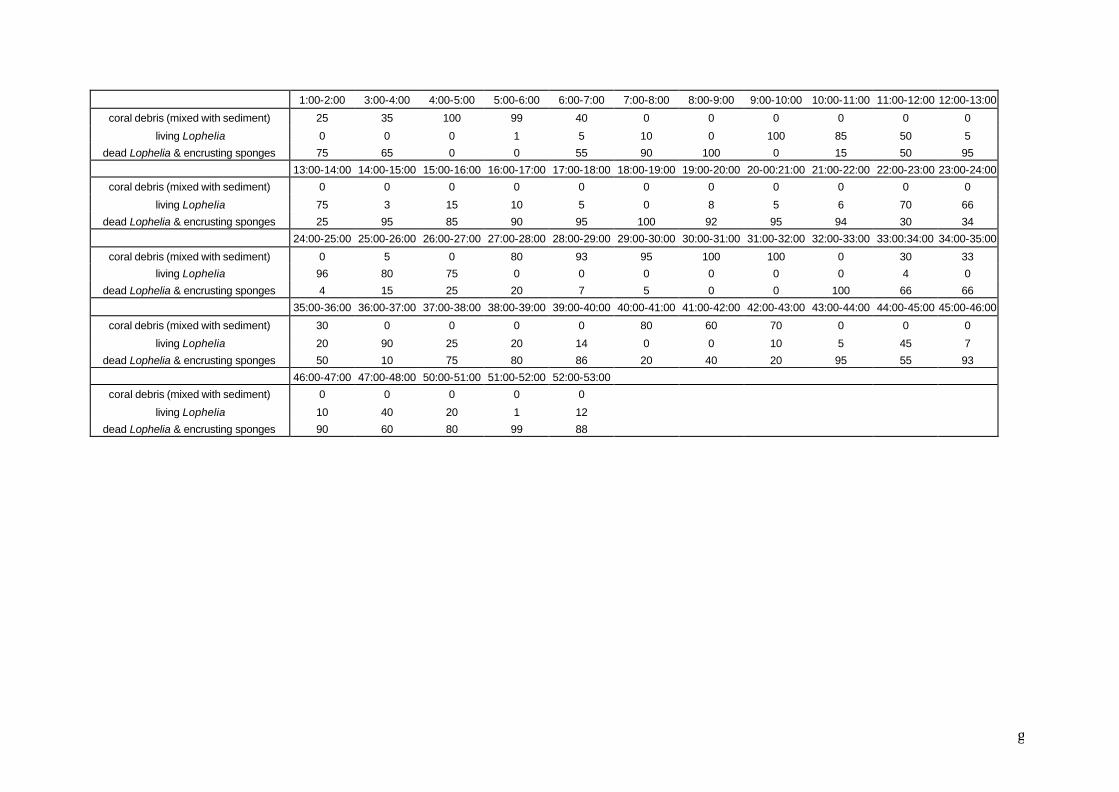
g
1:00-2:00 3:00-4:00 4:00-5:00 5:00-6:00 6:00-7:00 7:00-8:00 8:00-9:00 9:00-10:00 10:00-11:00 11:00-12:00 12:00-13:00
coral debris (mixed with sediment) 25 35 100 99 40 0 0 0 0 0 0
living Lophelia 0 0 0 1 5 10 0 100 85 50 5
dead Lophelia & encrusting sponges 75 65 0 0 55 90 100 0 15 50 95
13:00-14:00 14:00-15:00 15:00-16:00 16:00-17:00 17:00-18:00 18:00-19:00 19:00-20:00 20-00:21:00 21:00-22:00 22:00-23:00 23:00-24:00
coral debris (mixed with sediment) 0 0 0 0 0 0 0 0 0 0 0
living Lophelia 75 3 15 10 5 0 8 5 6 70 66
dead Lophelia & encrusting sponges 25 95 85 90 95 100 92 95 94 30 34
24:00-25:00 25:00-26:00 26:00-27:00 27:00-28:00 28:00-29:00 29:00-30:00 30:00-31:00 31:00-32:00 32:00-33:00 33:00:34:00 34:00-35:00
coral debris (mixed with sediment) 0 5 0 80 93 95 100 100 0 30 33
living Lophelia 96 80 75 0 0 0 0 0 0 4 0
dead Lophelia & encrusting sponges 4 15 25 20 7 5 0 0 100 66 66
35:00-36:00 36:00-37:00 37:00-38:00 38:00-39:00 39:00-40:00 40:00-41:00 41:00-42:00 42:00-43:00 43:00-44:00 44:00-45:00 45:00-46:00
coral debris (mixed with sediment) 30 0 0 0 0 80 60 70 0 0 0
living Lophelia 20 90 25 20 14 0 0 10 5 45 7
dead Lophelia & encrusting sponges 50 10 75 80 86 20 40 20 95 55 93
46:00-47:00 47:00-48:00 50:00-51:00 51:00-52:00 52:00-53:00
coral debris (mixed with sediment) 0 0 0 0 0
living Lophelia 10 40 20 1 12
dead Lophelia & encrusting sponges 90 60 80 99 88

Exercise 1
Below, you will find some objectives of ecological investigations concerning deep-sea
ecosystems. The aim of this exercise is that you try to find the most appropriate
sampling technique.
A. You have found a site that used to be covered with Lophelia pertusa and its
associated fauna but now it is heavily damaged due to intensive trawling. You would
like to monitor the changes in megafauna on this particular site to determine whether
the ecosystem can recover from this damage or not. What sampling method or
device would you use?
1. Dredge sampler
2. ROV with video equipment
3. Bathysnap
B. You would like to study the meiobenthos (animals between 38 µm and 1mm of
size, living in the sediment) in the sediment around Lophelia reefs. What sampling
method or device would you use?
1. Video recordings
2. Multicorer
3. Grab sampler
C. Your research team has discovered a mound-like feature during a side-scan sonar
survey of the seafloor. You suspect that there is some Lophelia on top of it, but of
course you have to verify that by taking a seafloor sample. What sampling method or
device would you use?
1. Dredge
2. TV-grab
3. ROV with video equipment

Solutions exercise 1
A. 1. No, you are wrong! A dredge is just as damaging as a trawl so it is
certainly not appropriate to use this device to study the possible recovery of a
damaged site.
2. No, you are wrong! Monitoring is a long-term project that requires regular
observations of the site. An ROV is very expensive and it would cost you a lot
of money to lower the ROV every month for only a few hours.
3. Yes, you are right! The Bathysnap device can be put on the seafloor for a
long period of time and it takes pictures at regular intervals. It is less
expensive than an ROV.
B. 1. No, you are wrong! Video material can only be used to observe large
animals living on top of the sediment, the so-called mega-epibenthos.
2. Yes, you are right! The multicorer can take up to nine small sediment
samples at the same time. These samples are virtually undisturbed and have
an intact upper layer, which is an important habitat for meiobenthic fauna.
3. No, you are wrong! A grab sampler takes large sediment samples, which is
unnecessary for meiobenthic studies. The samples are more disturbed and
you can only take one sample at the same time. It would save you a lot of
time if you used a multicorer.
C. 1. No, you are wrong! A dredge has been used frequently for this purpose in
the past, but this device is very damaging. As a good ecologist with a lot of
respect for the wonders of nature, you will choose an alternative sampling
method.
2. Yes, you are right! A TV-grab allows you to trace the corals first, after
which you can take a sample.
3. No, you are wrong! An ROV would be more suited if you would want to
map the coral reef and investigate zonation patterns.





Exercise 1
Imagine 2 samples, each containing 10 species and 100 individuals. In sample 1
(blue), every species occurred with 10 individuals while there was one species with
91 individuals in sample 2 (pink). The other 9 species occurred with only one
individual.
Sp.1 Sp.2 Sp.3 Sp.4 Sp.5 Sp.6 Sp.7 Sp.8 Sp.9 Sp.10
Sample 1 10 10 10 10 10 10 10 10 10 10
Sample 2 91 1 1 1 1 1 1 1 1 1
table 1. Two samples, each are containing 10 species (sp.), with indication of the
abundance of the species in each sample.
SR.1 SR.2 SR.3 SR.4 SR.5 SR.6 SR.7 SR.8 SR.9 SR.10
Sample 1 10 20 30 40 50 60 70 80 90 100
Sample 2 91 92 93 94 95 96 97 98 99 100
table 2. The same two samples, each containing 100 individuals. The species are
ranked according to their abundance and the cumulative abundance of all individuals
within each sample is calculated.
Question:
What sample has the highest diversity?
A. Sample 1
B. Sample 2
Figure 4.4. represents the graph of the cumulative frequencies (%) of the species in two samples.
number of species and evenness
0
20
40
60
80
100
120
1 2 3 4 5 6 7 8 9 10 11
species rank
cum
ula
tive
fre
qu
ency

Solution:
A. That is correct: Although both samples are characterized by the same
number of species, they display a different distribution of the individuals over
the species. Species 1 in sample 2 is clearly dominant, while all individuals in
sample 1 are equally spread over the species: high evenness.
B. That is not correct. The cumulative frequency curve indicates that sample 2 is
clearly dominated by one or more species, indicating a lower evenness than
sample 1.
Exercise 2
As already mentioned, photographs and video images were used to build up this
handbook. These images are not standardised, so every picture covers a different
surface on the seafloor (e.g. figure 4.5).
Question:
When you would want to compare the diversity between the species assemblages in
the pictures, what diversity indices do you think would be appropriate?
a) Shannon-Wiener index
b) Hill’s diversity numbers
c) Rarefaction
Answer:
a) You are partly right. Although the Shannon-Wiener index is sample size-
dependent, it can be a useful index if you want to compare with literature
concerning Lophelia and its associated fauna.
Figure 4.5 Pictures from Theresa Mound, representing different surfaces of the seafloor.
Copyright IFREMER Copyright IFREMER

b) You are partly right. The dominance index N∞ can be used in the case of
variable sample sizes but this is not true for N0 and N1
c) You are right. Rarefaction allows comparing samples in terms of number of
species; it is not correlated with sample size.
Exercise 3
The following picture was taken in one of the Haltenpipe coral reefs in Norway. The
lower part of the picture shows coral rubble, while the upper part shows dead coral
framework. The aim of this exercise is to calculate the species richness, diversity
(Shannon-Wiener index) and evenness.
The following table contains the species recognised in this picture and the number of
individuals per species. You can try to find these species back in the picture (if you
cannot see them on this picture, try examining the digital picture on the CD-rom using
a zoom function). Disregard the encrusting yellow and blue sponges; it is impossible
to quantify them because individual sponges are hard to discern. They can be
regarded as part of the substrate (! this means you will underestimate diversity!). For
this exercise you will need a calculator or a spreadsheet!
STATOIL

Notations:
Pi = the number of individuals of a given species divided by the total number of
organisms
S = the number of species in the picture
Shannon-Wiener index: H’ = -sum(Piln[Pi])
Evennes: J = H’/ ln S
Now calculate the values in the columns and make the sum of each column. The
negative value of the sum of the Pi ln Pi values will give you the Shannon-Wiener
index. The quotient of the Shannon-Wiener index and ln S will give you the
evenness.
Species Name # ind. Pi Pi ln Pi
Munida sp. 3 Prothantea simplex 2
Ophiuroidea sp. 5 Sabella pavonina 2
Asteroidea sp. 1 Fish (probably Sebastes
viviparus) 1
∑
The final result should look like this:
Species Name # ind. Pi Pi ln Pi
Munida sp. 3 0.214286 -0.3301 Prothantea simplex 2 0.142857 -0.27799
Ophiuroidea sp. 5 0.357143 -0.36772 Sabella pavonina 2 0.142857 -0.27799
Asteroidea sp. 1 0.071429 -0.1885 Fish (probably Sebastes
viviparus) 1
0.071429 -0.1885 ∑ 14 1 -1.6308
Note the high evenness, which is an important contributor to the high biodiversity
found associated with cold-water coral reefs! The evenness is high because the
individuals are evenly spread over the species; there is no clearly dominant species.
S = Ln S = H’ = J =
S = 6 Ln S = 1.79 H’ = 1.63 J = 0.91

Exercise 4 The idea of this exercise is to make the concept of sample-size dependency clear to you.
The exercise is based on a photo taken on Theresa Mound, Ireland, during the CARACOLE
cruise of the summer of 2001. During this campaign, images were made with cameras
mounted on ROV Victor, from which you can see a part on the right side of the complete
photo (3).
Below you can see three pictures: (1) a small part of the photo, (2) a larger part of the photo
and (3) the complete photo. Calculate the Shannon-Wiener diversity index for each picture
using the method demonstrated in the previous exercise. You will have to make a matrix with
species and abundances per picture. Disregard encrusting sponges; colonies of octocorals
can be regarded as one individual. Compare the indices you have calculated and answer the
following question: is the Shannon-Wiener index influenced by sample-size?
i
Species Name # individuals Pi pilnpi
? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? S ? ? ?
1
Copyright IFREMER
Shannon-Wiener index picture 1,2 and 3 = ?
a
b
c
d

3
Copyright IFREMER
2
Copyright IFREMER
e
f
g
h
i

Solution Picture 1
Species Name # individuals Anemone sp. 2 Munida sp. 1 Sponge sp. (yellow) 1 Aphrocallistes bocasei 1
Picture 2
Species Name # individuals Anemone sp. 2 Munida sp. 1 Sponge sp. (yellow) 2 Aphrocallistes bocasei 4 Echinus sp. 1 Octocoral sp1 (orange) 3 Octocoral sp2 (yellow) 1 Fish sp. 1 Hydrozoa sp. 1
Picture 3
Species Name # individuals Anemone sp. (a) 2 Munida sp.(b) 1 Sponge sp. (yellow: c) 4 Aphrocallistes bocasei (d) 8 Echinus sp. (e) 1 Octocoral sp1 (orange: f) 4 Octocoral sp2 (yellow: g) 1 Fish sp. (h) 2 Hydrozoa sp. (i) 1
Is the Shannon-Wiener index influenced by sample-size?
Yes. The difference is very clear between de indices of picture 1 and 2, where 5 additional
species were found. The indices are almost the same between picture 2 and 3 because no
new species were found, only additional individuals of the already known species.
Remark
This exercise also tells you something about the best sample size for biodiversity studies
about this coral reef. Picture 1 only sampled one patch or microhabitat (see 4.2.1) of the
coral reef, which contains a limited number of species. However, when studying the diversity
of the complete coral reef, you have to sample all the different patches to get a complete
view of the biodiversity. That is why more small samples or bigger samples have to be taken.
In this case, a good sample size is represented by picture 2: the sample is large enough to
sample all species. Larger samples will only yield more individuals of the same species.
Shannon-Wiener index = 1.33
Shannon-Wiener index = 2.04
Shannon-Wiener index = 1.91

Exercise 5 On this picture from Theresa Mound, Ireland, you can recognize the following species:
species abundance Cidaris cidaris (a) 2 Echinus sp. (b) 2 Octocorallia sp. (c) 1 Aphrocallistes bocasei (d) 24 Yellow sponge (dead A. bocasei??) (e) 3
The aim of this exercise is to calculate the Hill-numbers for this sample (the photo). The
principle is the same as in exercise 3.
Species Name # ind. Pi ln Pi Pi ln P i Pi * Pi
Cidaris cidaris 2 Echinus sp. 2
Octocorallia sp. 1 Aphrocallistes bocasei 24 Yellow sponge (dead
A. bocasei??) 3
∑
With these values you can calculate the Hill-numbers as:
N0 = the number of species in the sample
Copyright IFREMER
d
b
a
c
e

N1 = the reciprocal of the Shannon-Wiener index = exp (-sum pilnpi)
N2 = the reciprocal of Simpson’s diversity index = 1/ (sum pi*pi)
N∞ = 1/pi of the most dominant species
The result should look like this:
Species Name # ind. Pi ln Pi Pi ln P i Pi * Pi
Cidaris cidaris 2 0.0625 -2.77259 -0.17329 0.003906 Echinus sp. 2 0.0625 -2.77259 -0.17329 0.003906 Octocorallia sp. 1 0.03125 -3.46574 -0.1083 0.000977 Aphrocallistes bocasei 24 0.75 -0.28768 -0.21576 0.5625 Yellow sponge (dead A. bocasei??) 3 0.09375 -2.36712 -0.22192 0.008789 ∑ 32 1 -11.6657 -0.89256 0.580078
With N0 = 5 N1 = 2.44 N2 = 1.72 N∞ = 1.33

Exercise 1 Suppose that you are working on a comparison between the species assemblages of cold-
water coral reefs from different sites in the Northeast-Atlantic. You have at your disposal data
of species abundances obtained from boxcore-samples at each site on these maps:
You could apply multivariate analysis in order to define groups of sites with similar species
assemblages. But what kind of multivariate analysis would you use?
a. classification
b. ordination
c. both
Solution:
c. It is recommended to use both techniques. The data will probably be discontinuous as
samples are taken from different sites with different environmental variables. There can
however, be gradients you are unaware of and those will not appear on a classification
dendrogram. It will be clear from both dendrogram and ordination if your data are
discontinuous. However, a gradient will only be visible on an ordination graph.
54°
56°
58°
60°
62°
64°
66°
68°
70°
5 °
5 °
10°
10°
15°
15°
20°
20°
25°
25°
Kosterfjord
Sula Ridge
35°
40°
45°
50°
-30°
-30°
-25°
-25°
-20°
-20°
-15°
-15°
-10°
-10°
-5°
-5°
0°
0°
Rockall Trough
Porcupine Slope
Galicia Bank
Figure 5.18 Five important cold-water coral areas in the Atlantic Ocean.

Exercise 2
The aim of this exercise is to make your own cluster dendrogram, starting from a simple data
matrix. For a cluster analysis, the following steps need to be followed (check every step with
the solutions before you proceed to the next!):
1. Create your data matrix with samples (first row), species (first column) and their
abundances (matrix entries)
29:00-30:00
9:00-10:00
18:00-19:00
20:00-21:00
24:00-25:00
30:00-31:00
Mycale lingua (sp1) 4 3 7 9 2 2 unidentified sponge (sp4) 0 0 1 0 0 0 Geodiidae sp. (sp5) 0 0 0 1 0 0 unidentified sponge (sp6) 0 0 0 0 1 0 Hymedesmia sp (sp10) 1 0 0 0 0 1 Prothantea simplex 0 0 16 15 2 0 Primnoa resedaeformis & Paramuricea placomus 0 0 0 13 3 0 Paragorgia arborea 1 0 0 1 0 1 Sabella cf. pavonina 0 0 2 0 8 0 unidentified crab 0 0 1 0 0 0 unidentified decapod 0 0 0 2 0 0 Munida sp. 3 0 0 0 0 4 Cidaris cidaris 0 0 1 0 0 0 Hathrometra sarsi 0 0 0 0 3 0 Unidentified Asteroid sp1 1 0 1 0 0 1 Unidentified Ophiuroid 4 0 7 0 0 0 Sebastes viviparus 0 6 0 0 3 0 Brosme brosme 0 0 1 1 0 0 Pollachius virens 44 0 0 0 2 26 Micromesistius potassou 1 0 0 0 0 1
number of species 8 2 9 7 8 7
Table 1. This matrix contains 6 samples from the Statoil film recorded on the Haltenpipe A reef.
It will be the basis of your cluster dendrogram.
2. Calculate the distance between all samples. Store the results in a distance matrix
The distance between samples can be expressed as a similarity index. For this
exercise, you can use the Sørensen index:
cda
Ss += with
The principle of the Sørensen index is as follows: the index only takes into account
presence or absence of species in the samples (so you don’t really need the
a = the number of common species in sample 1 and 2 b = the total number of species in sample 1 c = the total number of species in sample 2

abundances: you can replace the numbers in the matrix by zeros and ones if you
want). The more absences and presences of species two samples have in common,
the higher the index will become.
Example: similarity between the samples 29:00-30:00 and 9:00-10:00
cd
aSs +
= = 1/(8+2) = 0.1 now fill out the rest of the distance matrix
29:00-30:00 9:00-10:00 18:00-19:00 20:00-21:00 24:00-25:00 30:00-31:00
29:00-30:00 / 0.1 ? ? ? ? 9:00-10:00 / / ? ? ? ? 18:00-19:00 / / / ? ? ? 20:00-21:00 / / / / ? ? 24:00-25:00 / / / / / ? 30:00-31:00 / / / / / /
Table 2. Distance matrix with Sørensen indices
3. Search through the distance matrix and find the two most similar samples. Fuse these
two samples to produce a cluster.
ð These 2 samples will form your first cluster
ð
4. Calculate the distances between this new cluster and all other clusters/samples
These distances can be calculated using the Flexible Sorting method:
ijhjjhiihk dddd βαα ++=
with h, i and j: groups (samples) with nh, ni and nj elements (species)
dhi, dhj and dij: similarity indices between samples
k: cluster formed with samples i and j
dhk: similarity between new cluster and sample h
a = 0.625 and ß = -0.25
Try this yourself. If you don’t know how to do it, there is an example in the solutions.
5. Repeat until all samples are in one cluster
In the case of 2 identical distances: select the samples with the least similarity with
other samples/clusters.
Sample 1
Sample 2

6. What can you conclude from the cluster?
ð Can you discern communities? If so, how many and can you discuss them if
you know that: 9:00-10:00 = 100% living coral 29:00-30:00 = 95% debris and sediment
24:00-25:00 = 96% living coral 20-00:21:00 = 95% dead coral
30:00-31:00 = 100% debris and sediment 18:00-19:00 = 100% dead coral
Solution of exercise 1
Calculate the distance between all samples. Store the results in a distance matrix:
29:00-30:00 9:00-10:00 18:00-19:00 20:00-21:00 24:00-25:00 30:00-31:00
29:00-30:00 / 0.1 0.18 0.13 0.125 0.47 9:00-10:00 / / 0.09 0.11 0.2 0.11 18:00-19:00 / / / 0.19 0.18 0.125 20:00-21:00 / / / / 0.2 0.14 24:00-25:00 / / / / / 0.13 30:00-31:00 / / / / / /
Search through the distance matrix and find the two most similar samples. Fuse these two
samples to produce a cluster.
The two most similar samples are the samples 29:00-30:00 and 30:00-31:00, so they will be
grouped in the first cluster.
Calculate the distances between this new cluster and all other clusters/samples
Example: distance between cluster 1 and sample 9:00-10:00
d = 0.625*0.1+0.625*0.11-0.25*0.4 = 0.001 A. 1 cluster
Cluster1 9:00-10:00 18:00-19:00 20:00-21:00 24:00-25:00
Cluster 1 / 0.01 0.06 0.048 0.039 9:00-10:00 / / 0.09 0.11 0.2 18:00-19:00 / / / 0.19 0.18 20:00-21:00 / / / / 0.2 24:00-25:00 / / / / /
29:00-30:00
30:00-31:00
1
Distance 29:00-30-00 and 9:00-10:00
Distance 30:00-31-00 and 9:00-10:00
Distance 29:00-30:00 and 30:00-31:00

There are 2 identical indices, but only one can be selected. Sample 9:00-10:00 shows
low similarities with other samples than 24:00-25:00. Sample 20:00-21:00 however, also
shows a high similarity with sample 18:00-19:00.
So the new cluster is
B. 2 clusters
Cluster 1 Cluster 2 18:00-19:00 20:00-21:00
Cluster 1 / 0.003 0.06 0.048 Cluster 2 / / 0.15 0.17
18:00-19:00 / / / 0.19 20:00-21:00 / / / /
The new cluster is
C. 3 clusters
Cluster 1 Cluster 2 Cluster 3 Cluster 1 / 0.003 0.02 Cluster 2 / / 0.15 Cluster 3 / / / The new cluster is
D. Final result
24:00-25:00
9:00-10:00
2
20:00-21:00
18:00-19:00
3
20:00-21:00
18:00-19:00
24:00-25:00
9:00-10:00
20:00-21:00
18:00-19:00
24:00-25:00
9:00-10:00
29:00-30:00
30:00-31:00 Debris & sediment
Dead coral
Living coral

E. Conclusion We can discern three communities depending on the substrate of the sample sites (dead
coral, living coral and debris & sediment). The groups of living and dead coral have more
species in common and therefore seem to be more similar to each other than to the
groups with a substrate of sediment and debris.
REMARK: The results from this classification are much clearer than the cluster analysis
in section 5.2.4 because the data are not continuous: there is no gradient from a low to a
high percentage in seafloor coverage.

Exercise 3 A picture from the Haltenpipe coral reef, Norway revealed the following species and
abundances (encrusting yellow and blue sponges not taken into account):
species abundance
Pollachius virens (a) 8 Mycale lingua (sponge)? (b) 2
Unidentified shrimp (reflecting eyes: c) 2 Sabellidae sp 2 (d) 1
Based on the analysis in section 5.4.3, you should be able to deduce the type of substrate
visible on the photograph using the table above with information on the species assemblage.
Question: What is the substrate type?
1. living Lophelia
2. dead Lophelia
3. coral debris and sediment
Answer: The substrate on the photograph consists of coral debris and sediment. This can be
concluded from the dominance of Pollachius virens, which is an indicator for this type of
substrate on the Haltenpipe reef.
STATOIL
a
b
c
d

Exercise 4
A picture from the Haltenpipe coral reef, Norway revealed the following species and
abundances (encrusting sponges not taken into account):
species abundance Sebastes viviparus (a) 4
Primnoa resedaeformis (b) 1 Sabella cf. pavonina (c) 1
Based on the analysis in section 5.4.3, you should be able to deduce the type of substrate
visible on the photograph using the table above with information on the species assemblage.
Question: What is the substrate type?
1. living Lophelia
2. dead Lophelia
3. coral debris and sediment
Answer: The photograph shows a cauliflower-shaped colony of living Lophelia. This could be
deduced from the presence of the redfish Sebastes viviparus (4 individuals) and the low
abundance of other species, which are all sessile suspension feeders.
STATOIL
a
a
a
b
a
c

Exercise 5
A picture from the Haltenpipe coral reef, Norway revealed the following species and
abundances. You can check this yourself using the picture on the CD and a zoom function
on your computer:
species abundance
Prothantea simplex (a) 9 Munida sp. (b) 2
Unidentified shrimp (reflecting eyes: c) 8 Ophiuroidea (d) 25
Based on the analysis in section 5.4.3, you should be able to deduce the type of substrate
visible on the photograph using the table above with information on the species assemblage.
Question: What is the substrate type?
1. living Lophelia
2. dead Lophelia
3. coral debris and sediment
Aswer: The substrate mainly consists of dead coral framework. You could have concluded
this from the high abundance of echinoderms (ophiuroids) and crustaceans (shrimps and
Munida sp.).
STATOIL
a
b
d
c

Exercise 6 A picture from the Haltenpipe coral reef, Norway revealed the following species and
abundances. You can check this yourself using the picture on the CD and a zoom function
on your computer:
species abundance
Primnoa resedaeformis (a) 4 Mycale lingua (b) 10
Sebastes viviparus (c) 2 Unidentified asteroid (d) 1
Munida sp. (e) 3 Unidentified shrimp (reflecting eyes: f) 22
Unidentified octocoral (g) 1 Based on the analysis in section 5.4.3, you should be able to deduce the type of substrate
visible on the photograph using the table above with information on the species assemblage.
Question: What is the substrate type?
1. living Lophelia
2. dead Lophelia
3. coral debris and sediment
Answer: The substrate mainly consists of dead coral framework. You could have concluded
this from the high abundance of crustaceans (shrimps and Munida sp.). The presence of the
indicator species Sebastes viviparus only indicates a possible presence of living Lophelia,
but the presence and abundance of other species have to be taken into account as well.
STATOIL
a
b
c
d
e f
g










![SYMMETRICALLY COMPLETE ORDERED SETS, ABELIAN … · of ordered sets; in particular, this gives an alternative approach to the construction of symmetrically complete elds in [12].](https://static.fdocuments.in/doc/165x107/5f13876c980f787f44061f70/symmetrically-complete-ordered-sets-abelian-of-ordered-sets-in-particular-this.jpg)







