
Leishmania infection of macrophages: down regulation … · Leishmania infection of macrophages:...
23
Leishmania infection of macrophages: down regulation of 7SL RNA expression and impairment of vesicular protein transport pathways Smita Misra, Manish K. Tripathi 1 and Gautam Chaudhuri* Division of Microbial Pathogenesis and Immune Response, Department of Biomedical Sciences, Meharry Medical College, 1005 D. B. Todd Jr. Blvd., Nashville, TN 37208, USA; 1 Present address: Department of Molecular Physiology and Biophysics, Vanderbilt University Medical Center, Nashville, TN 37232. Running title: Manipulation of macrophage biology by Leishmania *Address correspondence to: Gautam Chaudhuri, Division of Microbial Pathogenesis and Immune Response, Department of Biomedical Sciences, Meharry Medical College, 1005 D. B. Todd Jr. Blvd., Nashville, TN 37208, USA, Telephone number: 615-327-6499; Fax number: 615-327-5559; e-mail: [email protected] The parasitic protozoan Leishmania specifically manipulates the expressions of the host macrophage genes during the initial interactions, as was revealed by the mRNA differential display RT-PCR and the cDNA microarray analyses. The genes that are down regulated in the mouse (J774G8) or the human (U937) macrophages upon the exposure to Leishmania include the small RNA transcripts from the short interspersed element (SINE) sequences. Among the SINE RNAs that are down regulated is the 7SL RNA, which is the RNA component of the signal recognition particle (SRP). Since the microbicidal functions of the macrophages profoundly count on the vesicular protein transport processes, down regulation of the 7SL RNA may be significant in the establishment of the infection by Leishmania to the macrophage phagolysosomes. In order to evaluate whether the down regulation of the 7SL RNA results in the inhibition of the SRP-mediated vesicular protein transport processes, we have tested and found that the targeting of proteins to the endoplasmic reticulum and the plasma membrane and the secretion of proteins from the macrophages are compromised in the Leishmania-infected J774G8 and U937 cells. Knocking down the 7SL RNA using siRNA mimicked the effect of Leishmania-exposure to the macrophages. The over expression of the 7SL RNA inside J774G8 or U937 cells made those cells resistant to the Leishmania infection, suggesting the possible biological significance of the down-regulation of the 7SL RNA synthesis in the establishment of infection by Leishmania. We conclude that Leishmania down regulates the 7SL RNA in the macrophages to manipulate the targeting of many proteins that use the vesicular transport pathway and thus, favors its successful establishment of infection in the macrophages. Pathogenic microbes often develop strategies to evade the host immune responses, learn to adapt to the host environment and/or manipulate the host system to make it more hospitable for the establishment of infection and propagation (1-3). Any microbe interacting with the mammalian immune system is subjected to ultimate phagocytosis by the blood mononuclear phagocytes or the tissue macrophages and then it is cytolyzed to death inside the macrophage phagolysosomes (2-4). The parasitic protozoan Leishmania has the uncanny ability to evade the immune reactions of the macrophages, is able to establish infection inside the macrophage phagolysosomes and propagate inside the tissue macrophages (1-3). To survive inside the macrophage and escape immunity, Leishmania has developed mechanisms that deactivate the macrophage immune functions, including the inhibition of the respiratory burst, inhibition of interleukin-12 and nitric oxide synthesis, and the down regulation of the major histocompatibility complex class II molecules, as well as promoting the synthesis of inhibitory cytokines like transforming growth factor β and IL-10 and the induction of the suppressor of the cytokine signaling (5,6). Cytokines, and especially γ- interferon, are the essential contributors to the JBC Papers in Press. Published on June 14, 2005 as Manuscript M504162200 Copyright 2005 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on September 24, 2018 http://www.jbc.org/ Downloaded from
Transcript of Leishmania infection of macrophages: down regulation … · Leishmania infection of macrophages:...

Leishmania infection of macrophages: down regulation of 7SL RNA expression and impairment of vesicular protein transport pathways
Smita Misra, Manish K. Tripathi1 and Gautam Chaudhuri* Division of Microbial Pathogenesis and Immune Response, Department of Biomedical Sciences,
Meharry Medical College, 1005 D. B. Todd Jr. Blvd., Nashville, TN 37208, USA; 1 Present address: Department of Molecular Physiology and Biophysics, Vanderbilt University Medical Center,
Nashville, TN 37232.
Running title: Manipulation of macrophage biology by Leishmania *Address correspondence to: Gautam Chaudhuri, Division of Microbial Pathogenesis and Immune Response, Department of Biomedical Sciences, Meharry Medical College, 1005 D. B. Todd Jr. Blvd., Nashville, TN 37208, USA, Telephone number: 615-327-6499; Fax number: 615-327-5559; e-mail: [email protected]
The parasitic protozoan Leishmania specifically manipulates the expressions of the host macrophage genes during the initial interactions, as was revealed by the mRNA differential display RT-PCR and the cDNA microarray analyses. The genes that are down regulated in the mouse (J774G8) or the human (U937) macrophages upon the exposure to Leishmania include the small RNA transcripts from the short interspersed element (SINE) sequences. Among the SINE RNAs that are down regulated is the 7SL RNA, which is the RNA component of the signal recognition particle (SRP). Since the microbicidal functions of the macrophages profoundly count on the vesicular protein transport processes, down regulation of the 7SL RNA may be significant in the establishment of the infection by Leishmania to the macrophage phagolysosomes. In order to evaluate whether the down regulation of the 7SL RNA results in the inhibition of the SRP-mediated vesicular protein transport processes, we have tested and found that the targeting of proteins to the endoplasmic reticulum and the plasma membrane and the secretion of proteins from the macrophages are compromised in the Leishmania-infected J774G8 and U937 cells. Knocking down the 7SL RNA using siRNA mimicked the effect of Leishmania-exposure to the macrophages. The over expression of the 7SL RNA inside J774G8 or U937 cells made those cells resistant to the Leishmania infection, suggesting the possible biological significance of the down-regulation of the 7SL RNA synthesis in the establishment of infection by Leishmania.
We conclude that Leishmania down regulates the 7SL RNA in the macrophages to manipulate the targeting of many proteins that use the vesicular transport pathway and thus, favors its successful establishment of infection in the macrophages.
Pathogenic microbes often develop strategies to evade the host immune responses, learn to adapt to the host environment and/or manipulate the host system to make it more hospitable for the establishment of infection and propagation (1-3). Any microbe interacting with the mammalian immune system is subjected to ultimate phagocytosis by the blood mononuclear phagocytes or the tissue macrophages and then it is cytolyzed to death inside the macrophage phagolysosomes (2-4). The parasitic protozoan Leishmania has the uncanny ability to evade the immune reactions of the macrophages, is able to establish infection inside the macrophage phagolysosomes and propagate inside the tissue macrophages (1-3). To survive inside the macrophage and escape immunity, Leishmania has developed mechanisms that deactivate the macrophage immune functions, including the inhibition of the respiratory burst, inhibition of interleukin-12 and nitric oxide synthesis, and the
down regulation of the major histocompatibility complex class II molecules, as well as promoting the synthesis of inhibitory cytokines like transforming growth factor β and IL-10 and the induction of the suppressor of the cytokine signaling (5,6). Cytokines, and especially γ-interferon, are the essential contributors to the
JBC Papers in Press. Published on June 14, 2005 as Manuscript M504162200
Copyright 2005 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
macrophage activation to promote the effective killing of the parasites (5,6). Many of the functions carried out by the macrophages, including the destruction of the intracellular parasites such as Leishmania, are enhanced by their activation by the extracellular factors. Proper targeting of the receptor molecules specific for those activating factors on the cell surface is thus very important for the optimal function of the macrophages. We hypothesized that Leishmania, in the early stages of the infection, manipulates the expression of the cell surface receptors of the macrophages, making the cells unable to respond to their external stimuli. It has become increasingly apparent that the macrophages, in addition to their phagocytic and immunomodulatory properties, have an extensive secretory capability that includes secretions of protein molecules such as lysozyme, lysosomal acid hydrolases, neutral proteases, lipases, arginase, protease inhibitors, phospholipase inhibitor, complement factors, blood coagulation factors and several cytokines (7,8). Macrophage secretion involves synthesis on the rough endoplasmic reticulum (ER), co-translational glycosylation and translocation into the lumen of the ER, transport to the Golgi and vesicular transport to the plasma membrane. The fusion of the secretory vesicle membrane and the plasma membrane then occurs with the re-establishment of the membrane bilayer structure, thereby maintaining the integrity of the cell while expelling vesicle contents to the external milieu (9). The secretion of many cytokines by macrophages is of particular importance because those cytokines in their turn interacts with macrophages and T- or B-cells and assist in the control and fine tuning of the immune response. Our proposal is that Leishmania successfully establishes infection in the macrophages by down regulating the biosynthesis of 7SL RNA, which in turn results in the inhibition of protein targeting into the ER lumen and the plasma membrane and the secretion of protein molecules from the macrophages. The signal recognition particle (SRP) is a ribonucleoprotein machine that delivers certain nascent polypeptides to specific recognition components on the cytoplasmic face of the ER membrane for translocation of secretory or membrane proteins (10,11). The RNA component
of the SRP (7SL RNA) contains two elements related to the human and rodent Alu families of interspersed repetitive DNA sequences connected by a unique sequence, the S domain (11). The 7SL RNA associates with six proteins termed, in mammalian cells, SRP72, SRP68, SRP54, SRP19, SRP14, and SRP9 (11).
All proteins destined for the secretory pathway must first be targeted to the endoplasmic reticulum (ER). In the mammalian cells, this targeting reaction primarily occurs co-translationally via the SRP pathway. Proteins, targeted to the cell membrane for either secretion or integration, typically have an N-terminal signal peptide that directs them to their destinations. As the nascent polypeptide chain exits the ribosome, it is recognized and bound by the SRP. The SRP-ribosome complex then binds to the SRP receptor, which resides in the target membrane. Once bound, the paused translation machinery docks with a protein-translocating pore, or channel, and the SRP releases the signal peptide (10, 11). Translation resumes with the growing peptide integrating into or passing through the membrane. The SRP pathway is essential in all organisms examined to date except the yeast Saccharomyces cerevisiae (12). In order to substantiate our hypothesis that Leishmania actively manipulates the macrophages at the initial phases of its establishment of infection, we studied the differential expressions of genes in the cultured macrophages exposed to virulent Leishmania promastigotes (the insect form of the parasite) as compared to the un-exposed macrophages. Exposure of cultured mouse macrophages to virulent Leishmania promastigotes for 2-6 h revealed significant decrease in the levels of the small RNA transcripts from the short interspersed element (SINE) sequences in the cells (13). We report here our DD-RT-PCR as well as cDNA microarray analyses data that indicate differential expression of specific abundant mRNAs in the Leishmania-exposed macrophages. We hypothesize that the altered expressions of these genes may lead to the non-apoptotic growth arrest of the host cells favoring the parasitism of Leishmania. We further characterized the down regulation of one of the SINE RNAs, the 7SL RNA, in the Leishmania-exposed macrophages and presented evidence that this down regulation leads to the inhibition of
2
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
vesicular protein transport in the macrophages perhaps favoring leishmanial parasitism in the phagolysosomes of these cells. We have also shown that over-expression of 7SL RNA in macrophages confers resistance to the cells against Leishmania infection, at least in vitro.
Materials and Methods Leishmania, Trypanosoma and macrophages
The promastigotes and the amastigotes of L. amazonensis (LV78), L. major (Friedlin) and L. donovani (DD8) were used in this study. The promastigotes were grown at 25 °C in M199 medium with 10% heat-inactivated fetal bovine serum (13). Amastigotes were maintained in mouse tail-base lesions or in infected J774G8 cultures (14). Virulent promastigotes are those that are differentiated from amastigotes, freshly isolated from infected macrophages (14). Avirulent promastigotes are cloned laboratory stock of Leishmania that are maintained in axenic culture medium for more than 10 years and lost their infectivity to cultured macrophages or to mice (15). Trypanosoma brucei procyclics (16) were obtained from Dr. M. Chaudhuri. We used the mouse macrophages cell line J774G8 and the human monocytic leukemia cells U937 for our study. These cells were grown in tissue culture flasks in RPMI 1640 with 20% heat-inactivated (56 oC, 30 min) fetal bovine serum at 37 oC, as described (13, 14). U937 cells were differentiated into macrophages with phorbol-12-myristate-13-acetate (10 ng/ml) for 24 h, as described (17). The macrophages were incubated with the parasite cells at a parasite-to-macrophage ratio of 10:1 and incubated at 37 °C for 2-6 h for the exposure experiments (13-15, 18). DD-RT-PCR and microarray analyses Comparative gene expression analyses between unexposed and virulent L. amazonensis-exposed J774G8 cells were done by DD-RT-PCR as well as by mouse cDNA microarray analyses, similar to as described before (13,19,20). Total RNA was isolated from sub-confluent cultured cells using the Trizol reagents (Invitrogen). For the isolation of RNA from the unexposed macrophages, we added equivalent number of the parasite cells after addition of Trizol to the macrophages so that the contribution of
Leishmania RNA, if any, in the subsequent microarray hybridization is countered. The RNA quality was checked by formaldehyde-agarose gel electrophoresis. DD-RT-PCR analysis was performed as described previously (13). For cDNA microarray analysis, total RNAs (40 µg) from unexposed and exposed cells were labeled in reverse transcription reactions (Superscript II kit, Invitrogen) with dCTP-Cy3 and dCTP-Cy5, respectively (Amersham Biosciences, Arlington Heights, IL) (19, 20). Four microarrays were used for this study. In every second replicate experiment the fluorescent deoxyribonucleotides were swapped. Purified cDNA probes labeled with Cy3 and Cy5 were mixed per pair and hybridized
to mouse cDNA microarray chips (Mouse Research Genetics 5K) in the VMSR Microarray Core Facility at the Vanderbilt University. The slides were scanned with a GenePix 4000A microarray scanner (Axon Instruments, Union City, CA), and the images were analyzed using GenePix Pro 3 software. A uniform scale factor was applied to normalized signal intensities
between Cy5 and Cy3. Flagged spots and spots with an average intensity below 2.5-fold above the background were not retained for further analysis. The log2(Cy5/Cy3) ratio of the other spots was calculated for each slide. To compare the results from the different samples, data from each slide were normalized in log space to have a mean of 0 and a S.D. of 1 by using the Cluster program (21). Genes with significant changes in mRNA levels were identified using the Significant Analysis of Microarrays (SAM) procedure (22), a validated statistical technique for identifying differentially expressed genes across high-density microarrays. This procedure provides a list of "significant" genes and an estimate of the false discovery rate, which represents the percentage of genes that could be identified by chance (22). Primer extension and nuclear run-on analysis The levels of 7SL RNA in the control and parasite-exposed macrophages were determined by primer extension analysis using AMV reverse transcriptase (Promega, Madison, WI) according to the protocol supplied by Promega. β-Actin mRNA levels were evaluated as normalizing control. Primers used were as follows: 7SL RNA: 5′-ATGCCGAACTTAGTGCGG-3′; and, β-actin: 5′-TACACTGTAGCTGTCTTCAGACA-3′.
3
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
Primers were end-labeled with γ-32P-ATP and T4 polynucleotide kinase (23). Nuclear run-on analysis was performed as described (23). RT-PCR evaluation of 7SL RNA levels
Total RNA was isolated from cells and analyzed for 7SL RNA or β-actin by RT-PCR analysis (Amersham Biosciences). β-Actin was used as a loading control. Total RNAs (5 µg) were treated with DNase I (Promega) and reverse transcribed with Superscript II (Invitrogen) and random hexanucleotides as primers. The resulting first-strand cDNAs were used as templates to amplify 7SL RNA (157 bp; 5′-GGAGTTCTG GGCTGTAGTGC-3′ and 5′-ATCAGCACGGG AGTTTTGAC-3′), and β-actin (353 bp; 5′-GCTC GTCGTCGACAACGGCTC-3′ and 5′-CAAACA TGATCTGGGTCATCTTCTC-3′). Knock down of 7SL RNA gene expression
The siRNAs against human 7SL RNA designed from the nucleotide sequence (GenBank Accession #X04248) using the software available at the Invitrogen web site (https://rnaidesigner.invi trogen.com/sirna) and custom synthesized from Invitrogen. The nucleotide sequences of the siRNA pair (SR) and its respective control (SRC) are as follows (the number indicates the location of the sequence in the 7SL RNA, see ref 24): SR110, 5′-UCCGCACUAAGUUCGGCAU-3′/5′-AUGCCG AACUUAGUGCGGA-3′; SRC110, 5′-UCCACUAAGUUCGGGCCAU-3′/5′-AUGGCC CGAACUUAGUGGA-3′. We used Trans-messenger reagent (Qiagen) and protocols for the transfection of the macrophages with the siRNAs. RNAs were purified from the siRNA-treated cells using Trizol reagent and were treated with DNase I before RT-PCR analysis (19, 20). Over expression of 7SL RNA in cultured macrophages
Human 7SL cDNA sequence (24; GenBank Accession #X04248) was amplified from cDNA (made from RNA isolated from U937 cells) using Bgl II site anchored forward primer (5′-GGAGATCTGCCGGGCGCGGTGGCGCGT GC-3′) and the Hind III site anchored reverse primer (5′-GGAAGCTTAGAGACGGGGTCTCG CTATG-3′). The sequence-verified, amplified cDNA (315 bp) was digested with Bgl II/Hind III
and was cloned behind the H1 RNA promoter in the plasmid pSUPER (19) at the Bgl II/Hind III sites to obtain the plasmid pSUPER-7SL. We mutated the 7SL RNA gene in pSUPER-7SL simultaneously at G193A, G194A and G198A to create a nonfunctional 7SL RNA. Those ‘G’ residues are highly conserved across the genera and are essential for the function of 7SL RNA (25,26). We used the Quick-change site-directed mutagenesis reagents and protocols (Stratagene, La Jolla, CA) (27) to accomplish the mutagenesis. Mutation was verified by nucleotide sequencing. We named the mutated plasmid pSUPER-7SL*. The plasmid pSUPER-7SL or pSUPER-7SL* and pcDNA3.0 were co-transfected at a ratio of 10:1 into cultured macrophages using Lipofectamine 2000 (Invitrogen), as described (27). Control cells were only transfected with pcDNA3.0, which contains the G418 resistance gene. Recombinant cells were selected with G418 (200 µg/ml). To over express 7SL RNA in the cells that are expressing either enhanced cyan fluorescent protein (ECFP) or secreted alkaline phosphatase (SEAP) and are resistant to G418, we transfected them with pSUPER-7SL or pSUPER-7SL* and pTK-Hyg (a selection vector which confers hygromycin resistance, BD Clontech) and selected for cells that are resistant to both G418 (200 µg/ml) and hygromycin (200 µg/ml). Control cells were only transfected with pTK-Hyg. Over expression of 7SL RNA in the transfected cells were evaluated by RT-PCR, as described above. Evaluation of protein targeting in the endoplasmic reticulum To evaluate the effect of Leishmania- or siRNA-induced knock down of 7SL RNA in the macrophages on the targeting of proteins into endoplasmic reticulum, we employed a commercially available (BD Clontech) plasmid construct designed to test this cellular process. The Living Color endoplasmic reticulum targeting vector pECFP-ER (BD Clontech) encodes a fusion protein consisting of enhanced cyan fluorescent protein (ECFP); the endoplasmic reticulum (ER) targeting sequence of calreticulin cloned at the 5' end; and the sequence encoding the ER retrieval sequence, KDEL, cloned at the 3'-end. ECFP-ER is a soluble protein that localizes in the lumen of the ER in transfected cells. We stably transfected macrophages with the pECFP-ER plasmid.
4
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
Cultured J774G8 or undifferentiated U937 cells (in 6-well tissue culture plates) were transfected with pECFP-ER plasmid (1 µg) using Lipofectamine 2000 (Invitrogen) (27). Stable transfectants were obtained after selection with G418 (200 µg/ml). U937 cells were differentiated before exposure to virulent Leishmania promastigotes, as described above. Virulent Leishmania-exposed or unexposed cells were washed two times with PBS and fixed with 4% paraformaldehyde. Cells were mounted onto slides with Pro-Long (Molecular Probes) mounting medium and visualized with a 100x objective lens using an Olympus BX60 epifluorescent microscope. Images were recorded with an Optronics CCD camera (DEI-750). To evaluate whether the disappearance of the ECFP-ER protein in the Leishmania-exposed cells is due to proteasome-mediated degradation, we used the potent, membrane-permeable proteasome inhibitor, MG132 (Sigma Chemical Co., St Louis, MO) (28). MG132 was reconstituted in DMSO to obtain a 10 µM stock solution. Macrophages were treated with 1 µM MG132 for 24 h. Evaluation of protein targeting to the plasma membrane To evaluate the effect of Leishmania- or siRNA-induced knock down of 7SL RNA in the macrophages on the concentration of a membrane receptor, we assayed the level of macrophage scavenger receptor AII (SRAII) molecules on the surface of these cells. SRAII is highly abundant on the surface of macrophages (29). We used maleylated BSA (MBSA), an artificial ligand for this receptor (29), to probe this molecule. BSA was maleylated (~55 maleyl groups/molecule) using maleic anhydride (29). MBSA was radiolabeled with 125I using the ICl catalyzed reaction (29). Binding of 125I-MBSA to macrophages was measured as follows: After an overnight culture, the medium was removed from the 24-well dishes, cells were washed twice with 1 ml of phosphate-buffered saline (PBS) and they were incubated for 3 h at 4 °C with 0–50 µg protein per ml of 125I-labelled MBSA in 125 µl of RPMI 1640 containing 4% fatty-acid-free BSA and 25 mM Hepes, pH 7.4, in a total volume of 250 µl (total binding). Nonspecific binding was determined by the addition of 1 mg unlabelled
MBSA per ml of the incubation mix. At the end of the incubation, the cell monolayer was washed twice with 1 ml of PBS containing 0.2% BSA then twice with 1 ml of PBS. The cells were solubilized in 1.5 ml of 1 M NaOH, assayed for protein content and the radioactivity were counted (29). The specific binding was calculated by subtracting the nonspecific binding of 125I-labelled MBSA
from the total binding. The curves generated by the specific binding data were transformed into plots of the ratio of cell-bound to free 125I-labelled MBSA vs. cell-bound 125I-labelled MBSA according to the method of Scatchard (30). The
ordinate of the Scatchard plots (bound/free) represents the amount of specifically bound ligand (fmols of MBSA per mg cellular protein) divided by the concentration of unbound ligand in the reaction mixture (fmols of MBSA per ml). The dissociation constant (Kd) was calculated from the slopes and the maximum binding capacity (Bmax) was obtained from the x-axis intercept (30). Evaluation of the rate of protein secretion from the macrophages To evaluate the effect of 7SL RNA knock down on the secretion of proteins from macrophages we stably transfected the cells with pSEAP2-Control plasmid (BD Clontech) and evaluated the level of alkaline phosphatase in the culture medium. pSEAP2-Control is a positive control vector expressing secreted alkaline phosphatase (SEAP) under the control of the SV40 early promoter and the SV40 enhancer. The plasmid pSEAP2-control and pcDNA3.0 were co-transfected at a ratio of 10:1 into cultured macrophages using Lipofectamine 2000 (Invitrogen), as recommended by the manufacturer. Control cells were only transfected with pcDNA3.0, which contains the neomycin resistance gene. Transfected cells were selected with G418 (200 µg/ml). SEAP activity in secretion media and cell extracts were quantitated with the Great Escape chemiluminescent assay in a Turner design 20/20 luminometer. Evaluation of leishmanial infectivity of macrophages We tested both the J774G8 and the U937 cells for this purpose. U937 cells were differentiated with PMA in antibiotic-free medium before infection. Monolayer of macrophages was
5
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
incubated with or without parasite cells in antibiotic-free growth medium, with a macrophage:parasite ratio of 1:10, continuously for 5 days. One hundred macrophages were examined for the number of infected macrophages and the total number of amastigotes, microscopically (14,18). Statistical analysis All experimental data were expressed as the mean ± SE. A one-way analysis of variance, a two-way repeated measure analysis of variance,
and Student's t test were used to determine the significance of the difference (31).
Results Specific alterations in the transcriptome are induced in macrophages by their interactions with the virulent Leishmania promastigotes Parasites often manipulate the biology of their host cells to make the host environment hospitable for their growth and development. Many researchers have documented this over the past years for the interactions between Leishmania and its host macrophage. We employed DD-RT-PCR and cDNA microarray techniques to evaluate whether at their early stages of exposure to their host cells Leishmania induces differential gene expressions in the mouse macrophage cell line J774G8. Our data (Table 1) suggest that indeed the exposure of the macrophages to Leishmania induces up and down regulations of many genes. These inductions are reproducible and specific, as these changes are not revealed when J774G8 cells were exposed to avirulent promastigotes of L. amazonensis or to T. brucei procyclics (data not shown). We positively identified and validated twenty-two differentially regulated genes in virulent L. amazonensis-exposed J774G8 cells by independent RNA evaluation techniques like RNase protection assay, real-time and end-point RT-PCR (Table 1). Majority of these genes are detected by both DD-RT-PCR and cDNA microarray analyses (Table 1). Some of them are either detected by cDNA microarray analysis or by DD-RT-PCR-analysis (Table 1). Details of the evaluations of all these differentially expressed genes and the significance of their differential expression individually or as a group in leishmanial parasitism of macrophages are under
study. We report here the significance of the down regulation of 7SL RNA in macrophages by Leishmania exposure.
The genes that are significantly down regulated in Leishmania-exposed J774G8 cells include DNA primase p49 (catalytic subunit); Replication protein A p14; Proliferating cell nuclear antigen (PCNA); p160 c-Myb binding protein; RAD50; CDK regulatory protein 1; p70S6 kinase; CDK2; and a protein of replication origin recognition complex (Table 1). Up-regulated genes include histone H1.1 and protein phosphatase 1A. AAppooppttoottiicc mmaarrkkeerr ggeenneess tthhaatt aarree ddoowwnn rreegguullaatteedd iinncclluuddee Pdcd8 (apoptosis inducing factor) and mNapor (an apoptosis inducing ELAV-type RNA binding protein) (Table 1). The profiling may indicate non-apoptotic growth arrest of the Leishmania-exposed J774G8 cells at the non-dividing stage. Significant inhibition of the expression of 7SL RNA in Leishmania-exposed macrophages
An important observation we have made during our previous differential gene expression analysis is the down regulation of the SINE RNAs induced in the J774G8 cells by short exposure (2-6 h) to Leishmania (13). One of the SINE RNAs in mammals is the 7SL RNA, which is the RNA component of the signal recognition particle (10,11). The nucleotide sequence of the 7SL RNA is highly conserved between mouse and human. Thus, we were able to use the same primers for the primer extension as well as RT-PCR analyses of 7SL RNA both in the J774G8 and U937 cells. We found that the 7SL RNA is down regulated in the mouse as well as in the human macrophages during the exposure to virulent Leishmania promastigotes (Fig. 1A). U937 cells were differentiated to macrophages before exposure to virulent L. amazonensis promastigotes. The down regulation of 7SL RNA in J774G8 cells by virulent L. amazonensis was at the transcriptional level, as was revealed by nuclear run-on analysis (Fig. 1B). We have made similar observations with differentiated U937 cells and other virulent Leishmania promastigotes (data not shown). RT-PCR analysis with the RNA isolated from exposed and unexposed J774G8 cells shows the knock down of 7SL transcript in virulent L. amazonensis, L. major and L. donovani-exposed cells but not in the avirulent L. amazonensis promastigotes-
6
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
exposed cells (Fig. 1C). Procyclics of T. brucei and avirulent stock of L. donovani promastigotes, maintained in the lab for more than 10 years in axenic culture medium, failed to knock down the 7SL RNA level in the J774G8 cells (data not shown). Similar observations were made with differentiated U937 cells and virulent or avirulent Leishmania promastigotes (data not shown). We evaluated β-actin mRNA level to use as a normalization control in all these experiments. The 7SL RNA was knocked down or over expressed in macrophages in attempts to mimic or alleviate, respectively, the action of Leishmania
To mimic the action of Leishmania in knocking down of the 7SL RNA level, we treated macrophages (J774G8 and differentiated U937 cells) with siRNA specific to 7SL RNA. When compared with the cells treated with control siRNA containing several mismatches, the siRNA specific to 7SL RNA was able to ablate the expression of this RNA in J774G8 cells (Fig. 2A). Similar results were obtained with differentiated U937 cells (data not shown). To evaluate whether the consequences of leishmanial knock down of 7SL RNA may be alleviated by the over expression of the 7SL RNA in the macrophages, we have developed stable J774G8 and undifferentiated U937 cell lines in which the 7SL RNA is over expressed from the H1 RNA promoter. RT-PCR data indicated that the J774G8 cells that are stably co-transfected with pSUPER-7SL and pcDNA3.0 have 2-3 fold higher 7SL RNA than the cells that are not transfected or those that are transfected with pcDNA3.0 alone (Fig 2B). We obtained similar results with undifferentiated U937 cells (data not shown). To over express a 7SL RNA that is not functional, we made three point mutations in the 7SL RNA gene in pSUPER-7SL and stably transfected the macrophage cell lines with this mutated plasmid along with pcDNA3.0. RT-PCR analysis suggests that the mutated 7SL RNA is expressed in the stably transfected cells in the amount comparable to that in the cells transfected with the wild type construct (Fig. 2B). Exposure of the 7SL siRNA-treated J774G8 cells to virulent L. amazonensis promastigotes for 2 h further reduced the levels of 7SL RNA (Fig. 2C). On the other hand, exposure of macrophages over expressing 7SL RNA from
H1 RNA promoter to virulent L. amazonensis promastigotes for 2 h decreased only the endogenous 7SL RNA synthesis and the level of 7SL RNA remains at least similar to the control, untransfected cells (Fig. 2D).
Leishmania-induced down regulation of 7SL RNA impairs the targeting of proteins to the endoplasmic reticulum in the macrophages
To understand the consequences of the down-regulation of 7SL RNA in J774G8 cells by Leishmania, we tested the targeting of proteins to endoplasmic reticulum of the cells. The plasmid pECFP-ER (BD Clontech) encodes a fusion protein consisting of enhanced cyan fluorescent protein (ECFP) containing the endoplasmic reticulum (ER) targeting sequence of calreticulin. J774G8 cells permanently expressing this protein were developed. These cells were exposed to virulent L. amazonensis promastigotes for 4 h. Expression of ECFP in the ER was monitored by fluorescence microscopy. Leishmania exposure appeared to inhibit (50-80%) the ER-targeting of ECFP to the ER (Figs. 3a′ and 3b′). Knock down of 7SL RNA with specific siRNA in the ECFP-ER-expressing cells similarly inhibited the expression of ECFP protein fluorescence (Fig. 3c′). On the other hand, over expression of 7SL RNA in these cells alleviated the effect of Leishmania exposure (Fig. 3d′). The alleviating effect of over expressed 7SL RNA depends upon the functionality of this RNA, as the over expression of mutated 7SL RNA instead, failed to alleviate the effect of Leishmania exposure on the targeting of ECFP in the ER (data not shown). The disappearance of ECFP in the Leishmania-exposed cells is probably due to the proteasomal degradation of the untargeted protein, as pre-treatment of the ECFP-ER expressing J774G8 cells with the proteasome inhibitor MG132 (1 µM) for 24 h before Leishmania exposure, prevented the disappearance of the fluorescence (data not shown). Leishmania-induced down regulation of 7SL RNA decreased the levels of receptor protein molecules on the macrophage cell surface
One of the major roles of SRP is to help in the targeting receptor proteins on the plasma membrane. To understand the effect on the level of a receptor on the cell surface, Leishmania-
7
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
exposed (4 h) and unexposed J774G8 cells were assayed for 125I-labeled maleylated BSA binding to the scavenger receptors at 4 oC for 5 h with different concentrations of the ligand. There were 2-3 fold reductions in the level of scavenger receptor (SRAII) on the surface of Leishmania-exposed J774G8 cells (Fig. 4A). Similar down regulation in the level of the scavenger receptor concentration was observed when the J774G8 cells were treated with 7SL siRNA (Fig. 4B). The effect of Leishmania-exposure on the lowering of the level of the scavenger receptor was alleviated when wild type, but not the mutated 7SL RNA, is over expressed in the J774G8 cells (Fig, 4C). The procyclics of T. brucei could not lower the scavenger receptor level on J774G8 cells (Fig. 4A). Similarly, avirulent promastigotes of L. amazonensis or L. donovani could not lower the scavenger receptor level on J774G8 cells or differentiated U937 cells (data not shown). Leishmania-induced knock down of 7SL RNA inhibited the secretion of proteins from macrophages
Another consequence of the down regulation of the 7SL RNA expression should be the impairment of the secretion of protein molecules from the macrophages. To evaluate the effect of Leishmania-induced reduction of 7SL RNA level on the secretion of proteins from J774G8 cells, we stably transfected J774G8 cells with pSEAP2-control plasmid (BD Clontech). These cells were exposed (4 h) to virulent L. amazonensis promastigotes, and the levels of the thermostable alkaline phosphatase (coded by the plasmid), secreted in the growth medium, were assayed and compared to that from the uninfected cells, using a fluorescent substrate (BD Clontech). There was 60-85% inhibition of the secretion of alkaline phosphatase due to Leishmania-exposure (Fig. 5A). Exposure of the transfected cells to T. brucei, on the other hand, did not bring any significant change in the alkaline phosphatase secretion (Fig. 5A). The inhibition of alkaline phosphatase secretion occurred only when the macrophages were exposed to virulent promastigotes of Leishmania and not when the corresponding avirulent stocks were used (Fig. 5A). The degree of the inhibition of alkaline phosphate secretion was dependent upon the time of exposure to Leishmania promastigotes (Fig. 5B)
and on the macrophage to parasite ratio (Fig. 5C). Knock down of 7SL RNA by siRNA also decreased the secretion of alkaline phosphatase whereas control, mismatched siRNA had no effect (Fig. 5D). Leishmania-induced inhibition of alkaline phosphatase secretion from J774G8 cells was alleviated in the cells that over expressed the wild type 7SL RNA but not in that expressed the mutated 7SL RNA (Fig. 5E). These experiments were repeated with U937 cells and we obtained similar results (data not shown). SiRNA-mediated knock down of 7SL RNA in macrophages transiently helped avirulent Leishmania promastigotes to establish infection Avirulent Leishmania promastigotes could not down regulate the expression of the 7SL RNA in macrophages. To understand whether the macrophages will be more hospitable to the avirulent promastigotes to establish infection if the 7SL RNA expression is artificially knocked down, we exposed the control siRNA or the 7SL siRNA-treated J774G8 or differentiated U937 cells with the virulent or the avirulent L. amazonensis promastigotes. We counted the amastigotes inside those macrophages to evaluate the number of infected cells as well as the number of amastigotes per 100 macrophages. When the macrophages were treated with the control siRNA, only the virulent promastigotes were able to establish infection to those macrophages (Figs. 6A and 6B). The avirulent promastigotes apparently could establish infection in the 7SL siRNA-treated macrophages but could not multiply, as compared to the infection by the virulent promastigotes (Figs. 6A and 6B). The establishment of the infection by the avirulent promastigotes was indicted by the increased number of infected macrophages in the population (Figs. 6A and 6B). On the other hand, their inability to grow in the macrophages was indicated by the lower number of amastigotes per 100 macrophages (Figs. 6A and 6B). These data, thus, indicate that transient ablation of the 7SL RNA expression may be beneficial for the establishment of infection by the promastigotes of Leishmania, but the ability of the transformed amastigotes to multiply inside the phagolysosomes of the infected macrophages may be determined by other phenotypes of the parasite.
8
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
Macrophages over expressing 7SL RNA are resistant to Leishmania infection
The ability to down regulate the expression of the 7SL RNA in the macrophages seems to be associated with the virulent phenotype of Leishmania. If the parasite cells could not down regulate the 7SL RNA level, they could not establish infection to the macrophages. We tested whether the macrophage cells, which are over expressing the 7SL RNA, are resistant to the infection by the virulent Leishmania promastigotes. Our data show that the virulent L. amazonensis or L. major promastigotes cannot establish infection either in the J774G8 cells (Fig. 7A) or in the differentiated U937 cells (Fig. 7B) when these cells are over expressing the functional 7SL RNA but not the mutated, nonfunctional 7SL RNA.
Discussion
The molecular events involved in the host-
parasite interactions between the Leishmania and the macrophages have captivated scientists for the last half-century. The mechanism of the successful establishment of infection by Leishmania in the phagolysosomes of the human or the rodent macrophages is far from clear. It is apparent from the studies of several scientists that Leishmania takes active part in manipulating the cell biology of the host macrophages to make these cells more hospitable to them (32). Our study, reported here, indicates that the virulent Leishmania promastigotes indeed specifically alter the transcriptome profile of the macrophages at the early stages of their encounter to the host cells. The ability of the promastigotes to manipulate the macrophage gene expression in such a way may determine whether it will be able to establish the infection to the host cells. The reproducible changes in the transcriptome profile, as was induced by short (2 h) exposure to virulent Leishmania promastigotes, indicated non-apoptotic growth arrest of the cells at the non-dividing stage. Detailed analysis of the individual regulated genes is needed before we will be able to precisely determine their roles in the Leishmania-macrophage interactions. We do not know at present the molecule or molecules in the virulent Leishmania promastigotes responsible for the induction of the changes in the transcriptome of
the macrophages. Abundant parasite surface molecules, such as the major surface proteinase gp63 or lipophospho glycan (33) individually did not bring these changes (Misra, S. and Chaudhuri, G., unpublished data). It is possible that a combinatorial display of several molecules on the parasite surface and their simultaneous and/or sequential/progressive interactions with the macrophage surface molecules may bring those changes.
We are intrigued by the significant decrease in the level of 7SL RNA in the macrophages exposed to the virulent, but not to the avirulent, Leishmania promastigotes. The mechanism of this regulation remains elusive at present. We know from our nuclear run-on experiment that the regulation is at the transcriptional level. 7SL RNA gene is transcribed by RNA polymerase III (34-36). Our preliminary experiments suggested that the gene-internal promoters of RNA polymerase III, such as that of the tRNAs or the gene-external promoter of RNA polymerase III such as that of the H1 RNA, are not affected by Leishmania-exposure (Misra, S. and Chaudhuri, G., unpublished). Mammalian 7SL RNA gene has transcriptional elements in the outside as well as in the inside of the gene (34-36). Since Leishmania specifically inhibits the transcription of 7SL RNA gene, it may not be inhibiting the RNA polymerase III transcription in general. The specific molecular target in this RNA polymerase III transcription pathway, which is manipulated by the Leishmania-exposure of the macrophages to down regulate the 7SL RNA gene promoter, is yet to be identified.
The signal recognition particle (SRP) is a soluble ribonucleoprotein complex that was originally identified as an important intermediary in the transport of proteins into the secretory pathway in the mammalian cells (37). During translation, the 54-kDa polypeptide subunit of SRP (SRP54) binds to hydrophobic targeting signals that are found in both the pre-secretory and the integral membrane proteins (38). The targeting signals are generally either amino-terminal signal sequences (39) or, in the case of many membrane proteins that lack discrete signal peptides, the first transmembrane segment (40). Subsequently, SRP targets the ribosome-nascent chain complexes to the endoplasmic reticulum (ER), where an interaction between SRP54 and a heterodimeric
9
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
SRP receptor (SR) catalyzes the release of the nascent polypeptides and their insertion into a translocation channel or "translocon" (37-41). In the final step of the targeting cycle SRP dissociates from the ER membrane. In mammalian cells, the entry of the vast majority of proteins into the secretory pathway is completely dependent on the SRP targeting pathway. Partial and gradual inhibition of 7SL RNA biosynthesis will reduce the steady supply of the SRP particles from the nucleus and thus the vesicular transport of the proteins, extremely important for macrophage immune functions, will be paralyzed. Conceivably, this should make the macrophages more hospitable to Leishmania than before.
We have evaluated possible consequences of the down regulation of the 7SL RNA by Leishmania-exposure to macrophages. We have measured the targeting of a microsomal protein to the endoplasmic reticulum, the targeting of a receptor protein on the surface of the macrophages and the secretion of a protein from the cells. We have determined that the decrease in the efficiency of the protein secretory pathway in macrophages is indeed a consequence to the ablation of 7SL RNA expression. The knock down of this RNA by RNAi produced results very comparable to that with the virulent Leishmania-exposure. We have also shown that the over expression of the wild type, but not the mutated 7SL RNA, abrogated the negative effects of Leishmania exposure on the protein secretory pathway in macrophages. Earlier studies by others have shown that Leishmania down regulates the expression of both the class I and the class II major histocompatibility complexes on the macrophage cell surface and also leads to less production of IL-1 (32). The ability of the parasite to inhibit both the major histocompatibility complex expression and IL-1 production should be beneficial to the parasite, since the macrophages would thus be less capable of activating the parasite-specific T cell immunity. Our findings offer an explanation to these decreases of protein expression and/or secretion.
Our study also revealed that the ability of Leishmania to down regulate the expression of the
7SL RNA is critical for the ability of the parasite to establish infection in the macrophage phagolysosomes. Knock down of 7SL expression by siRNA in the macrophages made those macrophages amenable to infection by even otherwise attenuated Leishmania promastigotes. Although the parasites were able to establish infection in the 7SL RNA-ablated macrophages, they were unable to propagate therein. It is thus possible that down regulation 7SL RNA alone is not sufficient for Leishmania to parasitize the macrophages. On the other hand, if virulent Leishmania promastigotes are barred from down regulating the level of the 7SL RNA in the macrophages by artificial over expression of this RNA in these cells, they could not establish infection. The macrophages that have elevated levels of the 7SL RNA are apparently resistant to Leishmania infection. The over expressed 7SL must be biologically active to confer such resistance, as macrophages expressing the mutated 7SL RNA are not resistant to Leishmania infection. Chemical manipulation of macrophages to overproduce 7SL RNA may thus well be a mean to combat Leishmania infection of mammalian macrophages. Understanding the mechanism of the regulation of the 7SL RNA biosynthesis in detail may help us achieve this goal.
The down-regulation of the 7SL RNA synthesis in the macrophages may not be the primary event of Leishmania-macrophage interactions; nor is it the only molecular event happening as a result of the parasite-host encounter. However, this could be a pivotal event that translates the hostile behavior of the macrophages towards Leishmania into moderate, hospitable interactions. It is also possible that several secondary and tertiary biochemical changes may happen in the inside and the outside of the infected macrophages as a direct or indirect result of the down-regulation of the 7SL RNA biosynthesis, which may in turn make the macrophages more vulnerable towards leishmanial parasitism.
References 1. Kane, M. M., and Mosser, D. M. (2000) Curr. Opin. Hematol. 7, 26-31 2. Handman, E., and Bullen, D. V. (2002) Trends Parasitol. 18, 332-334 3. Denkers, E. Y., and Butcher, B. A. (2005) Trends Parasitol. 21, 35-41.
10
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
4. Solbach, W. and Laskay, T. (2000) Adv. Immunol. 74, 275-317 5. Bogdan, C., Gessner, A., Solbach, W., and Röllinghoff, M. (1996) Curr. Opin. Immunol. 8, 517-
525 6. Bertholet, S., Dickensheets, H. L., Sheikh, F., Gam, A. A., Donnelly, R. P., and Kenney, R. T.
(2003) Infect. Immun. 71, 2095-2101 7. Hamilton, T. A., Ohmori, Y., Tebo, J. M., and Kishore, R. (1999) Pathobiology 67, 241-244 8. Hamilton, T. A, Ohmori, Y., and Tebo, J. (2002) Immunol. Res. 25, 229-245 9. Sollner, T. H. (2004) Curr. Opin. Cell Biol. 16, 429-435 10. Koch, H. G., Moser, M., and Muller, M. (2003) Rev. Physiol. Biochem. Pharmacol. 146, 55-94 11. Doudna, J. A., and Batey, R. T. (2004) Annu. Rev. Biochem. 73, 539-557 12. Hann, B. C., and Walter, P. (1991) Cell 67, 131-144 13. Ueda, Y. and Chaudhuri, G. (2000) J. Biol. Chem. 275, 19428-19432 14. Mishra, M., Porter-Kelley, J., Singh, P. K., Bennett, J. R., and Chaudhuri, G. (2001) Biochem.
Pharmacol. 62, 569-580 15. Seay, M. B., Heard, P. L., and Chaudhuri, G. (1996) Infect. Immun. 64, 5129-5137 16. Walker, R. Jr., Saha, L., Hill, G. C., and Chaudhuri, M. (2005) Mol. Biochem. Parasitol. 139, 153-
162 17. Guizani-Tabbane, L., Ben-Aissa, K., Belghith, M., Sassi, A., and Dellagi, K. (2004) Infect. Immun.
72, 2582-2589 18. Mishra, M., Bennett, J. R., and Chaudhuri, G. (2001) Biochem. Pharmacol. 61, 465-474 19. Tripathi, M. K., and Chaudhuri, G. (2005) Biochem. Biophys. Res. Commun. 328, 43-48 20. Tripathi, M. K., Misra, S., and Chaudhuri, G. (2005) Biochem. Biophys. Res. Commun. 329, 508-
515 21. Eisen, M. B., Spellman, P. T., Brown, P. O., and Botstein, D. (1998) Proc. Natl. Acad. Sci. USA.
95, 14863-14868 22. Tusher, G. V., Tibshirani, R., and Chu, G. (2001) Proc. Natl. Acad. Sci. USA. 98, 5116-5121 23. Sambrook, J., and Russell, D. W. (2001) Molecular Cloning: A Laboratory Manual. 3rd Ed. Cold
Spring Harbor Laboratory Press, Cold Spring Harbor, NY 24. Ullu, E., and Weiner, A. M. (1984) EMBO J. 3, 3303-3310 25. Liao, X., Selinger, D., Althoff, S., Chiang, A., Hamilton, D., Ma, M., and Wise, J. A. (1992)
Nucleic Acids Res. 20,1607-1615 26. Kuglstatter, A., Oubridge, C., and Nagai, K. (2002) Nat. Struct. Biol. 9, 740-744 27. Tripathi, M. K., Misra, S., Khedkar, S. V., Hamilton, N., Irvin-Wilson,, C., Sharan, C., Sealy, L.,
and Chaudhuri, G. (2005) J. Biol. Chem. (In press ONLINE) 28. Lee, D. H., and Goldberg, A. L. (1998) Trends Cell Biol. 8, 397-403 29. Chaudhuri, G. (1997) Biochem. Pharmacol. 53, 385-391 30. Sarret, P., Esdaile, M. J., McPherson, P. S., Schonbrunn, A., Kreienkamp, H. J., and Beaudet, A.
(2004) J. Biol. Chem. 279, 8029-8037 31. Campbell, M. J., and Machin, D. (1994) Medical statistics: a commonsense approach, 2nd ed. John
Wiley, New York 32. Titus, R. G., Theodos, C. M., Shankar, A. H., and Hall, L. R. (1994) In: Macrophage-pathogen
interactions. Zwilling, B. S., and Eisenstein, T. K., eds. Marcel Dekker, Inc. New York, pp. 437-459
33. Chang, K. P., Chaudhuri, G., and Fong, D. (1990) Annu. Rev. Microbiol. 44, 499-529 34. Bredow, S., Kleinert, H., and Benecke, B. J. (1990) Gene 86, 217-225 35. Bredow, S., Surig, D., Muller, J., Kleinert, H., and Benecke, B. J. (1990) Nucleic Acids Res. 18,
6779-6784 36. Emde, G., Frontzek, A., and Benecke, B. J. (1997) RNA 3, 538-549 37. Halic, M., and Beckmann, R. (2005) Curr. Opin. Struct. Biol. 15, 116-125 38. Shan, S. O., and Walter, P. (2005) FEBS Lett. 579, 921-926
11
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
39. Zwieb, C., van Nues, R. W., Rosenblad, M. A., Brown, J. D., and Samuelsson, T. (2005) RNA 11, 7-13
40. Luirink, J., and Sinning, I. (2004) Biochim. Biophys. Acta 1694, 17-35 41. Wild, K., Halic, M., Sinning, I., and Beckmann, R. (2004) Nat. Struct. Mol. Biol. 11, 1049-1053
FOOTNOTES
*We thank Dr. Minu Chaudhuri, Department of Microbiology, Meharry Medical College, Nashville, TN, for Trypanosoma brucei procyclics. We also thank Mr. F. M. Faseehuddin and Dr. Y. Ueda for their contributions at the initial stages of this study. Supported by National Institute of Health, USA grants 5-R01AI42327-03, 2S06GM08037-24, and 3SO6GM008037-33S1 to GC. **The abbreviations used are: SRP, signal recognition particle; 7SL RNA, the RNA component of SRP; DD-RT-PCR, differential display reverse transcription PCR; ECFP, enhanced cyan fluorescent protein, SINE, short interspersed element; MBSA, maleylated bovine serum albumin; SRAII, scavenger receptor AII, SEAP, secreted alkaline phosphatase.
Figure Legends Fig. 1. Evaluation of the levels of the 7SL RNA in the unexposed and the Leishmania-exposed macrophages. (A) Primer extension analysis of the 7SL RNA in the J774G8 or the differentiated U937 cells, which are either unexposed or exposed to the virulent L. amazonensis promastigotes for 2 h before RNA isolation. β-Actin was used as a loading control. (B) Nuclear run-on analysis of the 7SL RNA in the J774G8 cells unexposed or exposed to the virulent L. amazonensis promastigotes, showing apparent transcriptional regulation of the 7SL RNA gene in the Leishmania-exposed macrophages. β-Actin was used as a control. (C) RT-PCR analysis of the levels of the 7SL RNA in the J774G8 cells unexposed or exposed (2 h) to the promastigotes of the virulent L. amazonensis, L. major, L. donovani or to the avirulent promastigotes of L. amazonensis. β-Actin was used as a normalization control. Fig. 2. RT-PCR analysis of the levels of the 7SL RNA in the 7SL RNA knocked-down or over-expressed J774G8 cells. (A) SiRNA-mediated knock down of the 7SL RNA as compared to the cells that were treated with the mismatched control siRNA. (B) Over expression of the 7SL RNA in the J774G8 cells stably transfected with the pSUPER-7SL plasmid. β-Actin was used as a normalization control. (C) Effect of exposure to the virulent L. amazonensis on the 7SL siRNA treated J774G8 cells. Mismatched siRNA was used as a negative control. Densitometric scanning and normalization against β-actin of several RT-PCR photographs are shown. (D) Effect of the exposure to the virulent L. amazonensis promastigotes on the 7SL RNA over expressing J774G8 cells. Cells with mutated 7SL RNA over expression were used as control. Results shown in (C) and (D) are mean + SE (n=12). The differences in the 7SL RNA levels between the cells treated with the control siRNA or the 7SL siRNA (as in C) or between the cells having no over expression or the over expression of either the wild type or the mutated 7SL RNA (as in D), were statistically significant (p<0.001). Fig. 3. Inhibition of the targeting of proteins to the endoplasmic reticulum in the J774G8 cells exposed (4 h) to the virulent L. amazonensis promastigotes. Recombinant J774G8 cells, stably expressing ECFP-ER, were tested. (a,a′) Control recombinant J774G8 cells without any exposure; (b, b′) effect of exposure to the virulent L. amazonensis promastigotes; (c, c′) effect of the treatment with the 7SL siRNA; (d, d′) effect of the exposure to the virulent L. amazonensis promastigotes on the J774G8 cells over expressing the 7SL RNA. Left panels (a-d) show the phase contrast photographs of the
12
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
macrophages and the right panels (a′-d′) show the corresponding fluoromicrographs. Stably transfected G418/hygromycin-resistant cells were used for the experiments in d and d′. Fig. 4. Down regulation of the levels of the scavenger receptor II on the J774G8 cells exposed (4 h) to the virulent L. amazonensis promastigotes. (A) Effects of the exposure (4 h) to the virulent L. amazonensis promastigotes or to the T. brucei procyclics; (B) effect of the knock down of the 7SL RNA with the siRNA without any exposure to the Leishmania promastigotes; (C) effect of the over expression of the wild type (wt) or the mutated (Mt) 7SL RNA followed by exposure (4 h) to the virulent L. amazonensis promastigotes. The Bmax for the binding of 125I-labeled maleylated BSA to the scavenger receptor on the cells surface was determined by the Scatchard analysis. Fig. 5. Inhibition of the secretion of alkaline phosphatase from the J774G8 cells exposed (4 h) to the virulent L. amazonensis promastigotes. (A) Effects of the exposure to different parasite cells. Stably transfected J774G8 cells were exposed to equivalent numbers of the avirulent, the virulent (vLam), the heat-killed (100 oC for 10 min) virulent promastigotes of L. amazonensis (Lam) or the T. brucei procyclics for 4 h before the alkaline phosphatase assay with the cell-free supernatant. (B) Effect of the length of exposure of the recombinant J774G8 cells to the avirulent or the virulent L. amazonensis promastigotes on the secretion of the alkaline phosphatase; (C) Effect of the number of the avirulent or the virulent L. amazonensis promastigotes per recombinant J774G8 cell on the secretion of the alkaline phosphatase; (D) Effect of the knock down of the 7SL RNA by the siRNA on the secretion of the recombinant alkaline phosphatase; (E) Effect of the exposure of the recombinant J774G8 cells over expressing either the wild type or the mutated 7SL RNA to the virulent L. amazonensis promastigotes on the secretion of the alkaline phosphatase. Data are expressed as the percent of the data obtained from the unexposed or the untreated J774G8 cells. Results are mean (n=12) + SE. The differences in the levels of the alkaline phosphatase activities between the virulent and the avirulent L. amazonensis-exposed cells (A), between the 7SL siRNA-treated and the control siRNA-treated cells (D), and between the wild type and the mutated 7SL RNA over expressing cells (E) were statistically significant (p<0.001). Fig. 6. The establishment of the infection by the avirulent L. amazonensis promastigotes in the 7SL siRNA-treated macrophages. (A) Infection of the 7SL siRNA-treated J774G8 cells by the virulent or the avirulent L. amazonensis promastigotes: (i) effect on the number of the infected cells per 100 macrophages checked; (ii) effect on the total number of the amastigotes per 100 macrophages (infected or uninfected). (B) Infection of the 7SL siRNA-treated, differentiated U937 cells by the virulent or the avirulent L. amazonensis promastigotes: (i) effect on the number of infected cells per 100 macrophages checked; (ii) effect on the total number of the amastigotes per 100 macrophages (infected or uninfected). Results are expressed as percent of macrophages that are not treated with any siRNA and are infected with the virulent L. amazonensis promastigotes. In these control cells typically 80-85% of the macrophages are infected and there are 485-515 amastigotes per 100 macrophages. Results are mean (n=12) + SE. The differences in the number of macrophages infected with the avirulent L. amazonensis between those that are treated with the control siRNA and those that are treated with the 7SL siRNA were statistically significant (p<0.001). Fig. 7. Development of resistance against the virulent L. amazonensis promastigotes in the macrophages over expressing functional 7SL RNA. Effects of the over expression of the wild-type (wt) or the mutant 7SL RNA in the J774G8 (A) and the differentiated U937 (B) cells on their infection by the virulent L. amazonensis or L. major promastigotes, respectively. The J774G8 and the undifferentiated U937 cells were stably transfected with the pSUPER-7SL or pSUPER-7SL* plasmid construct and were maintained in the culture medium. The recombinant U937 cells were differentiated with PMA before infection with the parasite cells. The cells were not treated with G418 during their incubation with the parasite cells. We randomly counted 100 macrophages and determined the number of the macrophages that are infected by the parasite (i) as well as the total number of the amastigotes per 100 cells (ii) 5 days
13
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Manipulation of macrophage biology by Leishmania
post-infection. In the control non-recombinant cells typically 80-85% of the macrophages are infected and there are 485-515 amastigotes per 100 macrophages. Results are mean (n=12) + SE. The differences in the numbers of infected macrophages and the number of amastigotes per 100 macrophages between the cells over expressing the mutant and the wild-type 7SL RNA were statistically significant (p<0.001).
14
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Table 1
Mouse genes up- or down-regulated in J774G8 cells after 2 h exposure to virulent L. amazonensis promastigotes
Regulated gene Fold changes
(increase or decrease) UP-REGULATED
Histone H1.1a 10-11 Protein Phosphatase 1Aa 3-5 Transcription elongation factor SIIa 3-4 Branched chain ketoacid dehydrogenase E1, alpha polypeptideb
5-6
Syndecan-3 precursor proteina 3-4 Fibrillin 2b 6-7
DOWN-REGULATED DNA Primase p49a 3-5 Replication protein A p14b 4-5 Proliferating cell nuclear antigen (PCNA)a
8-11
P160 c-Myb binding proteinb 4-6 RAD50a 3-5 CDK regulatory protein 1b 4-5 P70S6 kinasea 5-7 CDK2b Replication origin recognition proteinb 7-8 Pdcd8 (programmed cell death 8 or AIF)a
5-6
mNapor (mouse neuroblastoma apoptosis-related RNA binding protein)a
4-5
Glycine transporter 1b 6-7 Ceruloplasminb 6-8 T-complex protein 1, zeta subunita 5-6 Glvr1 mRNAb 6-7 7SL RNAc 15-18
aDetected by both microarray and DD-RT-PCR analyses. bDetected only by cDNA microarray analysis.
cDetected only in DD-RT-PCR analysis.
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE 1
J774G8 cells
J774G8 U937
A. Primer Extension Analysis
Leishmania: - + - +
β-Actin RNA
7SL RNA
B. Nuclear Run-On Analysis
C. RT-PCR with RNA from J774G8 cells
L. amazonensis L. major L. donovani Avirulent La
by guest on Shttp://w
ww
.jbc.org/D
ownloaded from
Parasite: - + - + - + - +
eptemb
Leishmania: - +
7SL RNA er 24, 2018
β-Actin

FIGURE 2 C
ontr
ol
siR
NA
7S
L si
RN
A
A. 7SL RNA knock down B. 7SL RNA over-expression
7SL RNA
pSUPER-7SL: - - + +*pcDNA3.0: - + + +
C. Eexp
Dow
nloaded f
Contro0123456789
101112131415161718192021
7SL
RN
A le
vel
(% n
oLe
ishm
ania
exp
osur
e co
ntro
l)
ffect of Leosure on s
β-Actin
l siRNA 7SL siRNA
ishmania iRNA-treated cells
0
10
20
30
40
50
60
70
80
90
100
7SL
RN
A le
vel
(% n
oLe
ishm
ania
exp
osur
e co
ntro
l)
D. Efon 7S
fect of Leishmania exposure L RNA over expressing cells
by guest on Sehttp://w
ww
.jbc.org/rom
No over expression Wt 7SL RNA Mutant 7SL RNA
ptember 24, 2018

FIGURE 3
Control recombinant J774G8 cells
a′a
b b′
L. amazonensis-exposed (4 h) recombinant J774G8 cells
c c′
Recombinant J774G8 cells
treated for 48 h with 7SL siRNA
d′d
Phase contrast ECFP Fluorescence
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
L. amazonensis-exposed
recombinant J774G8 cells over expressing 7SL
RNA

FIGURE 4
A. Effect of exposure to parasite cells
B. Effect of exposure to 7SL siRNA
hD
ownloaded from
C. Effect of over expression of 7SL RNA
0 10 20 30 40 50 60 70 800
5
10
15
20
25 Unexposed
7SL siRNAControl siRNA
Bound (fmols/mg cell protein)
Bou
nd/F
ree
by guest on September 24, 2018
ttp://ww
w.jbc.org/
0 10 20 30 40 50 60 70 800
5
10
15
20 Unexposed ControlWt 7SL RNA/exposedMt 7SL RNA/exposed
Bound (fmols/mg cell protein)
Bou
nd/F
ree

FIGURE 5
1
234
56789
10
11
Alk
alin
e Ph
osph
atas
e A
ctiv
ity(%
No
siR
NA
con
trol
)
0102030405060708090
100110120
Alk
alin
e Ph
osph
atas
e A
ctiv
ity(%
Une
xpos
ed c
ontr
ol)
A. Effect of exposure to different parasite cells
E. Effect of parasite exposure on 7SL RNA
0 1 2 3 4 5 6
VirulentAvirulent
Length of exposure (h)
e
Avirulent Lam Virulent Lam Heat-killed vLam T. brucei0
10
2030
40
5060
70
80
90
100
110
Alk
alin
e Ph
osph
atas
e A
ctiv
ity(%
Une
xpos
ed c
ontr
ol)
0 1 2 3 4 5 6 7 8 9 100
102030405060708090
100110120
AvirulentVirulent
Parasite/Macrophage ratio
Alk
alin
e Ph
osph
atas
e A
ctiv
ity(%
Une
xpos
ed c
ontr
ol)
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
D. Effect of 7SL siRNA
B. Effect of the length of exposur
7SL siRNA Control siRNA00
000
000000
0
C. Effect of the parasite/macrophage ratio
over expressed cells
Wild-type 7SL Mutated 7SL0
25
50
75
100
Alk
alin
e Ph
osph
atas
e A
ctiv
ity(%
une
xpos
ed c
ontr
ol)

FIGURE 6
(
(
A. Infection of 7SL siRNA-treated J774G8 cells
Control siRNA 7SL siRNA0
10
20
30
40
50
60
70
80
90
100
110AvirulentVirulent
Num
ber
of in
fect
ed c
ells
(% N
o si
RN
A/v
irul
ent c
ontr
ol)
120 Avirulent(ii)
i)
Control siRNA 7SL siRNA0
10
20
30
40
50
60
70
80
90
100
110AvirulentVirulent
Num
ber
of in
fect
ed c
ells
(% N
o si
RN
A/v
irul
ent c
ontr
ol)
i)
Control siRNA 7SL siRNA0
10
20
30
40
50
60
70
80
90
100
110 Virulent
Num
ber
of a
mas
tigot
es/1
00 m
acro
phag
es(%
No
siR
NA
/vir
ulen
t con
trol
)Avirulent
( )
http://ww
Dow
nloaded from
ii
B. Infection of 7SL siRNA-treated U937 cellsC ontrol siR N A 7SL siR N A0
10
20
30
40
50
60
70
80
90
100 Virulent
Num
ber
of a
mas
tigot
es/1
00 m
acro
phag
es(%
No
siR
NA
/vir
ulen
t con
trol
)
by guest on September 24, 2018
w.jbc.org/
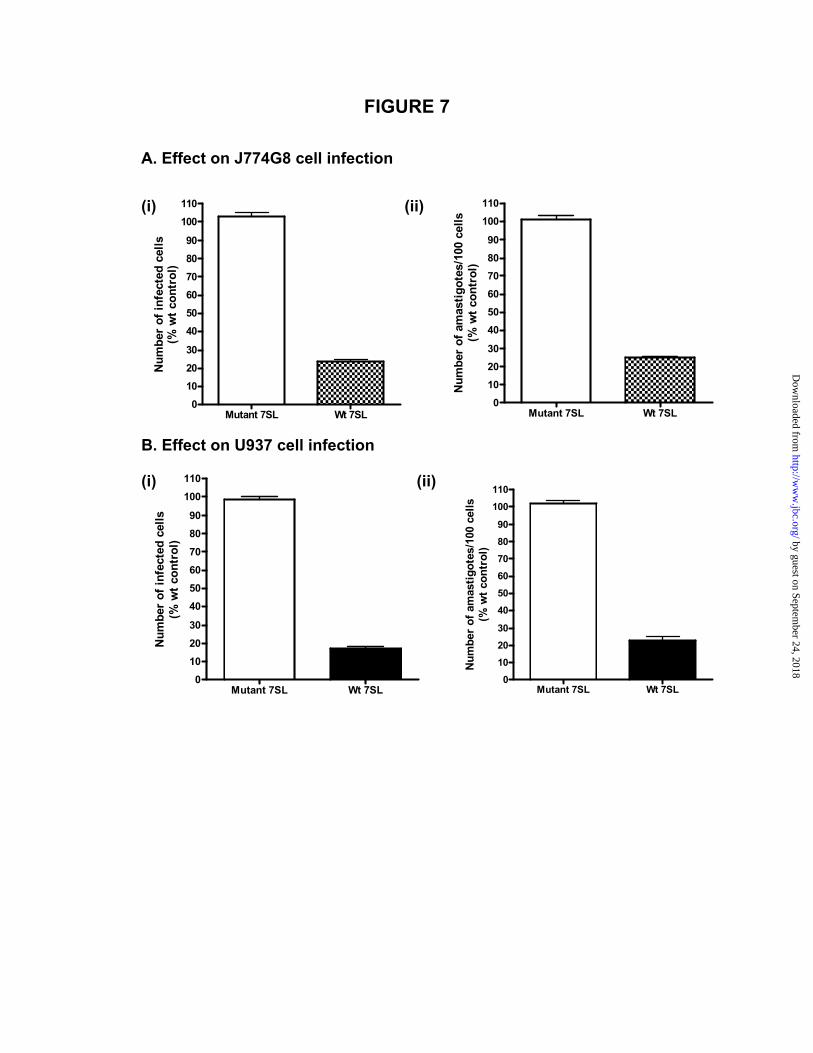
FIGURE 7
A. Effect on J774G8 cell infection
Mutant 7SL Wt 7SL0
10
2030
4050
60
7080
90
100
110
Num
ber
of a
mas
tigot
es/1
00 c
ells
(% w
t con
trol
)
Mutant 7SL Wt 7SL0
1020
30
40506070
8090
100110
Num
ber
of in
fect
ed c
ells
(% w
t con
trol
)
(ii) (i)
B. Effect on U937 cell infection http://wD
ownloaded from
Mutant 7SL Wt 7SL0
10
20
30
40
50
60
70
80
90
100
110
Num
ber
of in
fect
ed c
ells
(% w
t con
trol
)
110(ii) (i) w
Mutant 7SL Wt 7SL0
10
20
30
40
50
60
70
80
90
100
Num
ber
of a
mas
tigot
es/1
00 c
ells
(% w
t con
trol
)
by guest on September 24, 2018
w.jbc.org/

Smita Misra, Manish K. Tripathi and Gautam Chaudhuriimpairment of vesicular protein transport pathways
Leishmania infection of macrophages: Down regulation of 7SL RNA expression and
published online June 14, 2005J. Biol. Chem.
10.1074/jbc.M504162200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on September 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















