
LECTIN-MEDIATED AGGLUTINATION OF AMPHIBIAN … · Fixation was carried out at 4 °C in agar-lined...
17
J. Cell Sci. 27, 227-243 (1977) 227 Printed in Great Britain © Company of Biologists Limited LECTIN-MEDIATED AGGLUTINATION OF AMPHIBIAN EMBRYONIC CELLS BROCK R. FRASER AND SARA E. ZALIK Department of Zoology, The University of Alberta, Edmonton, Alberta, Canada T6G 2E9 SUMMARY Dissociated blastula cells of Xenopus laevis are agglutinated with wheat germ agglutinin (WGA), Ricinus communis agglutinin (RCA), Concanavalin A (Con A) and, to a lesser extent with soya bean agglutinin (SBA). They are not agglutinated with fucose-binding protein. Neuraminidase treatment of cells enhances their agglutinability with RCA and SBA, but has no effect on Con A- and WGA-mediated agglutinability. Treatment of cells with procaine, or xylocaine, has no effect on the cells' agglutinability or on the extrusion of lobopodia. Treatment with colchicine or cytochalasin B either separately or simultaneously has no effect on lectin- mediated agglutinability. Cells treated with cytochalasin B or colchicine and cytochalasin B simultaneously lack lobopodial extensions, while colchicine alone has no effect on these structures. Phenothiazine tranquillizers inhibit agglutination mediated by all of the above mentioned lectins. Lobopodial extensions are absent in cells treated with these compounds. Glutaraldehyde fixation inhibits RCA and WGA mediated agglutinability and reduces the Con A-mediated agglutinability. Results suggest that in this system microtubules and micrcfilaments are not involved in lectin-mediated agglutination. INTRODUCTION During embryogenesis a series of complex migratory movements occurs as the embryo undergoes gastrulation. These movements are the result of precisely regulated cell interactions involving the formation and exchange of specific intercellular contacts which in turn reflect the capacity of cells to discriminate between each other. That specific cellular affinities exist in gastrulating amphibian embryos was demonstrated by Holtfreter (1939) and Townes & Holtfreter (1955). Evidence from other systems suggests that the cell surface plays an important role in cellular recognition and adhesion, and that these two phenomena may reflect the physical and chemical organization of the cell surface (Curtis, 1973; Moscona, 1974). Complex carbohydrates are important components of the cell surface and lectins have proved useful as investigative probes into the nature of the carbohydrate-bearing groups at the cell surface (Cook & Stoddart, 1973). In embryonic systems Krach, Green, Nicolson & Oppenheimer (1974) reported changes in the agglutinability of embryonic cells during development of the sea-urchin embryo. Also, in this system, differences in the agglutinability between cells derived from different germ layers have been reported (Roberson, Neri & Oppenheimer, 1975). In the chick embryo, work by several investigators (Zalik & Cook, 1974, 1976; Kleinschuster & Moscona, 1972; Moscona, 1974) has shown that early embryonic cells possess surface lectin receptors capable of participating in an agglutination reaction and that lectin-induced
Transcript of LECTIN-MEDIATED AGGLUTINATION OF AMPHIBIAN … · Fixation was carried out at 4 °C in agar-lined...

J. Cell Sci. 27, 227-243 (1977) 227Printed in Great Britain © Company of Biologists Limited
LECTIN-MEDIATED AGGLUTINATION OF
AMPHIBIAN EMBRYONIC CELLS
BROCK R. FRASER AND SARA E. ZALIKDepartment of Zoology, The University of Alberta,Edmonton, Alberta, Canada T6G 2E9
SUMMARY
Dissociated blastula cells of Xenopus laevis are agglutinated with wheat germ agglutinin(WGA), Ricinus communis agglutinin (RCA), Concanavalin A (Con A) and, to a lesser extentwith soya bean agglutinin (SBA). They are not agglutinated with fucose-binding protein.Neuraminidase treatment of cells enhances their agglutinability with RCA and SBA, but hasno effect on Con A- and WGA-mediated agglutinability. Treatment of cells with procaine, orxylocaine, has no effect on the cells' agglutinability or on the extrusion of lobopodia. Treatmentwith colchicine or cytochalasin B either separately or simultaneously has no effect on lectin-mediated agglutinability. Cells treated with cytochalasin B or colchicine and cytochalasin Bsimultaneously lack lobopodial extensions, while colchicine alone has no effect on these structures.Phenothiazine tranquillizers inhibit agglutination mediated by all of the above mentionedlectins. Lobopodial extensions are absent in cells treated with these compounds. Glutaraldehydefixation inhibits RCA and WGA mediated agglutinability and reduces the Con A-mediatedagglutinability. Results suggest that in this system microtubules and micrcfilaments are notinvolved in lectin-mediated agglutination.
INTRODUCTION
During embryogenesis a series of complex migratory movements occurs as theembryo undergoes gastrulation. These movements are the result of precisely regulatedcell interactions involving the formation and exchange of specific intercellular contactswhich in turn reflect the capacity of cells to discriminate between each other. Thatspecific cellular affinities exist in gastrulating amphibian embryos was demonstratedby Holtfreter (1939) and Townes & Holtfreter (1955). Evidence from other systemssuggests that the cell surface plays an important role in cellular recognition andadhesion, and that these two phenomena may reflect the physical and chemicalorganization of the cell surface (Curtis, 1973; Moscona, 1974).
Complex carbohydrates are important components of the cell surface and lectinshave proved useful as investigative probes into the nature of the carbohydrate-bearinggroups at the cell surface (Cook & Stoddart, 1973). In embryonic systems Krach,Green, Nicolson & Oppenheimer (1974) reported changes in the agglutinability ofembryonic cells during development of the sea-urchin embryo. Also, in this system,differences in the agglutinability between cells derived from different germ layershave been reported (Roberson, Neri & Oppenheimer, 1975). In the chick embryo,work by several investigators (Zalik & Cook, 1974, 1976; Kleinschuster & Moscona,1972; Moscona, 1974) has shown that early embryonic cells possess surface lectinreceptors capable of participating in an agglutination reaction and that lectin-induced

228 B. R. Fraser and S. E. Zalik
agglutinability changes as differentiation proceeds. In the amphibian embryo, Con-canavalin A has been reported to have an effect on development (Moran, 1974) and thebinding of fluorescent-labelled Con A has been studied in Xenopus laevis (O'Dell,Tencer, Monroy & Brachet, 1974), and recently in several species of Rana (Johnson &Smith, 1976).
In the present study a variety of lectins were used to explore the nature of some ofthe cell surface carbohydrates of mechanically dissociated cells from embryos ofX. laevis. The agglutinability of cells induced by 5 different lectins was determined.In addition, the effects of cell fixation, treatment of cells with neuraminidase, cyto-chalasin B, colchicine, and local anaesthetics and phenothiazine tranquillizers on theabove-mentioned process were studied. Some of these results have been presentedrecently in preliminary form (Fraser & Zalik, 1976).
MATERIALS AND METHODS
Chemicals
Steinberg's physiological saline (S.S.) was prepared according to Hamburger (1962) andused in the agglutination tests. Calcium-free Steinberg's or Calcium-Magnesium-free Stein-berg's were used in cell dissociation while a modified Calcium-Magnesium-free Steinberg'swas used in experiments with local anaesthetics and phenothiazine tranquillizers, in the lattercase the concentration of the Trizma buffer was increased to 20 mM. Wheat germ agglutinin(WGA) in a 1-35 mg/ml aqueous solution (lotWGA 15) and fucose-binding protein (FBP) in a3-3 mg/ml solution in phosphate buffer (lot 7), were purchased from Miles-Yeda (Israel).Concanavalin A (Con A) was purchased from Sigma as a highly purified, salt-free, lyophilizedpowder (Grade IV, lot 123C-5500). Ricinus communis agglutinin (RCA), a gift from Dr G. M. WCook of the Strangeways Laboratory, Cambridge, U.K., was prepared from castor beansaccording to Nicolson & Blaustein (1972). The larger molecular weight fraction (RCA 120;mol. wt. 120000) was used. Soya bean agglutinin (SBA), also a gift from Dr Cook, was preparedby the method of Lis, Sharon & Katchalski (1966). Lectins were dissolved or diluted in Calcium-Magnesium-free Steinberg's to obtain stock solutions of appropriate concentration, and storedat —20 °C. Haptens were made up as o-i M stock solutions, dissolved in Calcium-Magnesium-free Steinberg's, and were stored at —20 °C until used. a-methyl-D-glucoside (lot 82C-1010),a-methyl-D-mannoside (lot 52C-3870), and N-acetyl-D-glucosamine (lot 31C-3000) wereobtained from Sigma. y?-lactose was obtained from Nutritional Biochemical Corp. (U.S.A.),and iV-acetyl-D-galactosamine (lot X3738) from Schwarz/Mann (U.S.A.). The local anaes-thetics procaine hydrochloride (Matheson, Coleman and Bell, U.S.A.) and xylocaine hydro-chloride (Astra Pharmaceuticals Ltd, Canada) were obtained as crystalline powders. Thephenothiazine derivatives chlorpromazine hydrochloride and prochlorperazine (methanesulphonate) were obtained in crystalline form from Poulenc Pharmaceuticals Ltd, Canada.Trifluoperazine was donated by Mowatt and Moore Ltd, Canada (trade name ' Clinazine').Stock solutions of local anaesthetics and phenothiazine tranquillizers were made in Calcium-Magnesium-free Steinberg's or modified Calcium-Magnesium-free Steinberg's respectively andwere stored at room temperature in the dark. Colchicine (Sigma, lot 77B-1580) was prepared inCalcium-Magnesium-free Steinberg's and cytochalasin B (Aldrich Chemical Co, U.S.A.,lot PH/2435/60E) was dissolved in dimethyl sulphoxide (DMSO) (Baker Chemical Co., U.S A.).Glutaraldehyde (a 70 % aqueous solution from Ladd Research Industries, U.S.A.) was dilutedto 7 % with Calcium-free Steinberg's and adjusted to pH 7-8. Special Agar-Noble (DifcoLaboratories, U.S.A.) was dissolved in heated Calcium-Magnesium-free Steinberg's to a 2 %(w/v) concentration. It was cooled in dialysis tubing and dialysed for 24 h at 4 °C againstCalcium-Magnesium-free Steinberg's. The agar was then melted and used to line some of theagglutination dishes.

Lectin-mediated agglutination 229
Cells and agglutination assays
Embryos of Xenopus laevis obtained by hormonal stimulation, were allowed to develop to thelate blastula stage (stage 9, Nieuwkoop & Faber, 1967). Embryos were transferred to Steinberg's,where jelly coats and vitelline layers were removed manually. They were transferred toCalcium-free Steinberg's, pre-chilled to the temperature of melting ice, and incubated for5 min, after which the saline was replaced by fresh cold Calcium-free Steinberg's. Embryos weredissociated by gently pipetting with a wide mouth pipette, and the cell suspension was centri-fuged for 5 min at 19 g. Cells were then resuspended in fresh, cold Calcium-free Steinberg's,pipetted further if necessary, re-centrifuged, and resuspended in ice-cold Calcium-freeSteinberg's. Since the presence of calcium in the medium results in rapid aggregation of cells,a process that interferes with the assessment of agglutination, freshly dissociated cells suspendedin ice-chilled Calcium-free Steinberg's were used in agglutination tests. Tests were performedeither within porcelain rings mounted on glass slides, or in agar-lined wells of ' Linbro multi-dish disposotrays'. Agglutination assays were performed in total volumes of 50 or 60 ji\. Haptenspecificity was determined by addition of specific sugars to the agglutination mixture to give afinal concentration of 0-03 M. Cells were always added last to the agglutination mixture. Afterthe addition of cells, agglutination plates were gently rotated by hand to bring the cells intocontact with each other and agglutination was scored visually under a dissecting microscope.For evaluation of agglutination, plates were vigorously shaken by reciprocal motion to dispersenon-agglutinated cells, and scored according to Inbar & Sachs (1969) as follows: o, the degree ofclustered cells in the cell suspension in the absence of lectin - this base line takes into accountoccasional cells aggregates sometimes present in cell suspension; + , up to 30% of the cellsagglutinated over the base line; + + , 30—60 % of the cells agglutinated; + + + , 60-90 % of thecells agglutinated into small clusters of 10 cells or less; + + + + , more than 90% of the cellsagglutinated into clusters of more than 10 cells. This assay does provide a basis for assessing theextent of agglutination induced by lectins. Cell suspensions were also examined under the phase-contrast microscope.
Neuraminidase, drugs, and cell fixation
For neuraminidase treatment, cells were resuspended in Magnesium-free saline, with 0-005 MCaCl2 and neuraminidase from Vibrio cholerae (Hoecht Pharmaceuticals) was added to a finalenzyme concentration of 60 units/ml. This enzyme is stated by the suppliers to be free of protease,aldolase, and lecithinase C activities. Controls consisted of cells suspended in the above-mentioned saline without enzyme. Since higher temperatures are deleterious to the cells,suspensions were incubated at room temperature, and quickly rinsed twice in ice-chilledCalcium-free Steinberg's. Cells were then suspended and tested for lectin-induced agglurin-ability.
Local anaesthetics were used at a concentration of 7-5 x io~3 to 1-4 x io"1 M for procaine andat I - I x io~2 to 11 x io"1 M forxylocaine and dissolved in Calcium-free Steinberg's, or modifiedCalcium-free Steinberg's. Control and experimental cells were incubated for 1 h at roomtemperature, in the absence or presence of drugs, after which agglutination assays were per-formed. Cells were examined at high magnification under phase-contrast microscopy for generalmorphological features. The phenothiazine derivatives chlorpromazine, prochlorperazine andtrifluoperazine were added to experimental cell suspensions at concentrations varying from2 x io~' to 2 x io~* M, and dissolved in modified Calcium-free Steinberg's. Control and drug-treated cells were incubated at room temperature for 30-60 min, and tested for lectin-inducedagglutinability. Cells were added to the agglutination plates while suspended in the incubationmedium. In some experiments the reversibility ofthe drug-induced effect was investigated.Control and drug-treated cells were washed up to 3 times with either modified Calcium-freeSteinberg's or Calcium-free Steinberg's, and cells were tested again for agglutinability. In someexperiments washed, drug-treated cells were incubated in drug-free saline for up to 2 h andagglutination assays were performed. Colchicine was used at concentrations ranging from 30to 120/4M while cytochalasin B was used at 20 fig I ml. Controls for cytochalasin B containedDMSO at equivalent concentrations. In some experiments, cells were incubated in colchicine
23° B. R. Fraser and S. E. Zalik
(60 /(M) and cytochalasin B (20 fig/ml) simultaneously. Cells were incubated with or withoutdrugs for 1 h at room temperature and tested for RCA- and WGA- mediated agglutinability.
Fixation was carried out at 4 °C in agar-lined Petri dishes. Cell suspensions in Calcium-freeSteinberg's were added to the dishes and placed in a rotary shaker at 80 rev/min. An equalvolume of 7% glutaraldehyde was then added drop-wise. Cells were fixed for 24 h in rotation;they were subsequently washed 3 times with ice-cooled Calcium-free Steinberg's and testedfor lectin-mediated agglutinability.
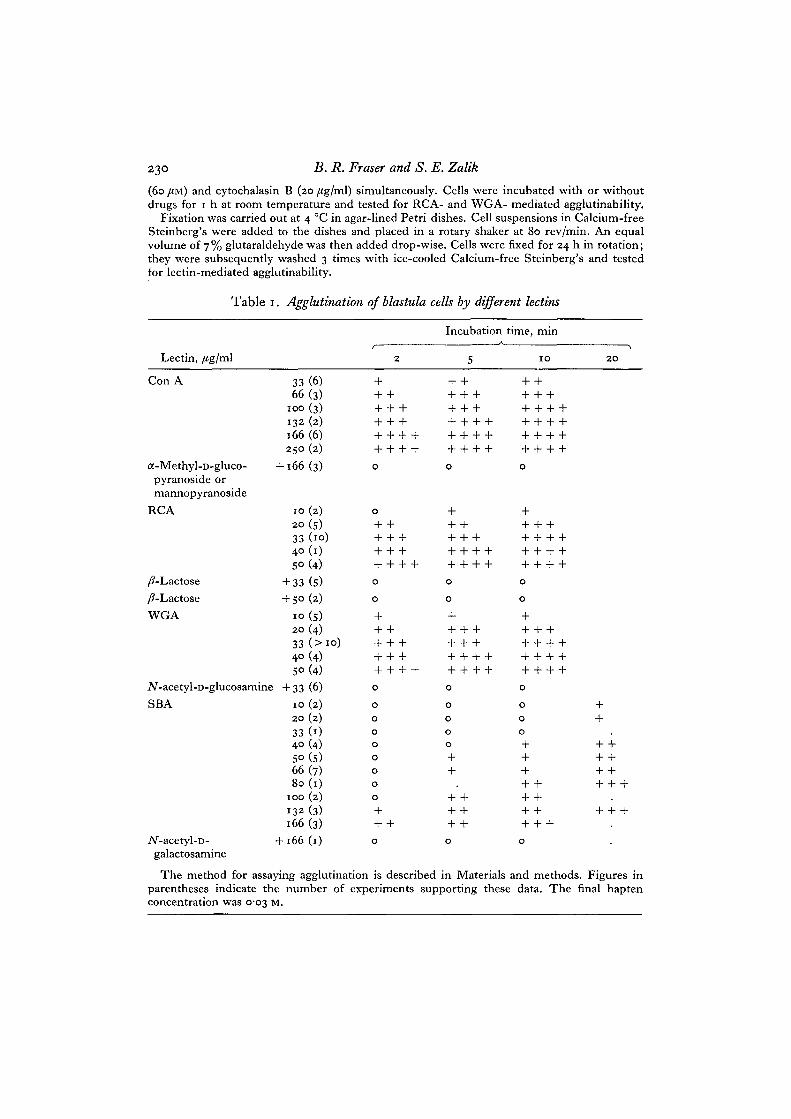
Lectin,
Table i
/tg/ml
. Agglutination of blastula
2
cells by different lectins
Incubation time, minA
5 IO 2O
Con A
a-Methyl-D-gluco-pyranoside ormannopyranoside
RCA
33(6)66(3)
100 (3)
132 (2)
166 (6)250 (2)
+ 166(3)
10(2)
20 (s)
33 (10)
40 (1)
S°(4)/?-Lactose +33 (5)/?-Lactose +50 (2)WGA 10 (5)
20 (4)33 ( > i <4° (4)5° (4)
iV-acetyl-D-glucosamine + 33 (6)SBA
iV-acetyl-D-galactosamine
10(2)
20 (2)
33(04° (4)SO(5)66(7)80(1)
100 (2)
132 (3)
166 (3)+ 166(1)
o
o
o
o
oooo
oooo
The method for assaying agglutination is described in Materials and methods. Figures inparentheses indicate the number of experiments supporting these data. The final haptenconcentration was 0-03 M.

Lectin-mediated agglutination 231
RESULTS
The data presented here represent results pooled from several experiments, eachusing embryos from different clutches. Variations in the degree of agglutinability ofcells derived from different clutches were observed; however, in spite of this varia-bility, certain surface properties of cells in this system were consistently demonstrated.The effects of WG A, RCA, Con A, and SB A on cells of Xenopus embryos are presentedin Tables 1 and 2. Rapid and massive agglutination is observed when blastula cells are
Table 2. Specificity of lectin-mediated cell agglutination
WGA»,Hapten, 003 M 33 fig/ml
None + + +A^-Acetyl-D-glucosamine 0iV-Acetyl-D-galactosamine + +/?-Lactose + + +a-Methyl-D-glucoside + + +a-Methyl-D-mannoside + + +
Lectin
RCA»,33 /ig/ml
+ + ++ + +-|- +
0
+ + ++ + +
used
Con A*166 /ig/ml
+ + + ++ ++ + + ++ + + +++
The method for assaying agglutination is described in Materials and• Agglutination scored at 2 min, -f" at 10 min.
SBAf,166 fig/m\
+ + ++ + +
0
0
+ + ++ + +
methods.
placed in the presence of WGA, Con A or RCA, at relatively low concentrations.The agglutination response to these lectins is near maximal within 2 min at roomtemperature, although at lower lectin concentrations, agglutination increases withincubation time. At 33 y«g/ml WGA and RCA, and 100 /tg/ml Con A, further incuba-tion for 8 min more does not appreciably increase the extent of agglutination, andthese are denned in this paper as optimal lectin concentrations (Figs. 1, 3, 5). Subopti-mal lectin concentrations represent the amount of lectin necessary to induce 30-60%agglutination after a 2-min incubation period, and correspond to io/tg/ml, 16/ig/mland 33 /Jg/ml for WGA, RCA and Con A respectively (Table 1). In contrast to theresults with WGA, RCA and Con A, a lower agglutinability was observed with SBA(Table 1). Under the conditions of these experiments, the agglutination with thislectin is a slower process; it is near maximal at 10 min of incubation and is completedafter 20 min. Also, a higher lectin concentration (i66/ig/ml) is necessary to inducenear maximal agglutination at 10 min (Fig. 6), and for the purposes of subsequentexperiments this was taken as the optimal lectin concentration, the suboptimal SBAconcentration being 66 /ig/ml. No agglutination was observed when FBP was added atconcentrations up to 167 /Jg/ml. The hapten specificity of the lectin mediated agglutina-tion is illustrated in Tables 1 and 2, and Figs. 2 and 4; lectins were used at optimal ornear optimal concentrations. WGA-mediated agglutination is inhibited completely byiV-acetyl-D-glucosamine, and slight inhibition is obtained in the presence of iV-acetyl-D-galactosamine. RCA-mediated agglutination is completely inhibited by /?-lactose,

232 B. R. Fraser and S. E. Zalik

Lectin-mediated agglutination 233
while iV-acetyl-D-galactosamine is slightly inhibitory. In the majority of the ex-periments agglutination by Con A is completely inhibited by either a-methyl-D-glucoside or a-methyl-D-mannoside, and some inhibition was observed withiV-acetyl-D-glucosamine. Complete inhibition of SBA-induced agglutination isobservedin the presence of either 7V-acetyl-D-galactosamine or /Mactose (Tables 1 and 2).
Amphibian embryos at the late blastula stage are composed of presumptive ectoder-mal cells located at the animal pole, cells of the marginal zone which will give rise tothe mesoderm, and presumptive endodermal cells located at the vegetal pole of theembryo. The degree of pigmentation decreases from the animal pole toward thevegetal pole, while the number and size of yolk granules, as well as cell size, increase inthe same direction. The criteria of cell size, abundance of yolk, and degree of pigmen-tation were used in order to determine if any of the lectins studied preferentiallyagglutinated a particular cell type. Upon close microscopical examination of theagglutinated and non-agglutinated cells, no selective cell agglutination is observed forany of the lectins at the concentrations used in these experiments. In all cases,agglutinated cell masses appear to be composed of indiscriminate mixtures of differentcell types. The relative proportion of cells of any one type present in agglutinationmasses reflects the relative proportion of cells of that type present in the cell suspension.
The effects of neuraminidase treatment on lectin-mediated agglutination wereinvestigated. Using any of the 4 lectins at optimal concentration, no reduction inagglutinability was found when enzyme-treated cell suspensions were compared tocontrols. The results of similar experiments using suboptimal lectin concentrationsare summarized in Table 3. Neuraminidase treatment has no effect on Con A- orWGA-mediated agglutination; however, treatment with this enzyme greatly enhancesthe agglutinability of cells in response to RCA. This enhanced response at low RCAconcentrations is rapid and comparable in time to that obtained at higher lectinconcentrations. A similar although less dramatic enhancement of agglutination bySBA (66 /tg/ml)) occurs after neuraminidase treatment. The enhanced SBA-inducedagglutinability shows the same temporal characteristics as described for control,non-enzyme-treated, cells; it is a relatively slow response. Neuraminidase treatmentdoes not induce agglutination in the presence of FBP (166 fig/ml).
In other cell systems, local anaesthetics and phenothiazine tranquillizers have beenfound to influence some cell surface-mediated phenomena such as lectin-inducedagglutination and cell capping (Poste, Papahadjopoulos & Nicolson, 1975 a; Poste,
Fig. 1. Dissociated cells in the presence of WGA (33 /tg/ml), 2 min after addition ofcells, x 40Fig. 2. Cells from Xenopus blastulae in the presence of WGA (33 /tg/ml) and 0-03M iV-acetyl-D-glucosamine, 5 min after addition of cells, x 40.Fig. 3. Amphibian blastula cells in the presence of RCA (33 /tg/ml), 2 min afteraddition of cells, x 40.Fig. 4. Blastula cells incubated with RCA (33 /tg/ml) and 0-03 M /Mactose, 5 min afteraddition of cells x 40.Fig. 5. Dissociated cells in the presence of Con A (100/tg/ml), 2 min after additionof cells, x 40.Fig. 6. Agglutination induced by SBA (166 /tg/ml), 8 min after addition of cells, x 40.

234 B. R. Fraser and S. E. Zalik
Table 3. Effect of newaminidase treatment on lectin-induced agglutination
Lectin
Incubation time,min
No. of exps.
ControlNANase
The method forand methods.
Con A, WGA,30/ig/ml io/tg/ml
A A
2 8 2 5 2I 2
+ + + + + + 0
assaying agglutination and the procedure
RCA,16/ig/ml
A
84
for enzyme
SBA,66 /ig/ml
A
4 io
5
0 +
treatment are described
FBP,166 /ig/ml
2 10
2
0 0
0 0
in Materials
Papahadjopoulos, Jacobson & Vail, 19756; Ryan, Unanue & Karnovsky, 1974). Inorder to investigate further the mechanisms involved in the lectin-mediated agglutina-tion of embryonic Xenopus cells, the effects of these drugs upon this process werestudied. Cells were treated with either procaine or xylocaine, and the effect on WGA-mediated agglutination was studied. At low concentrations of procaine and xylocaine,and using optimal and suboptimal lectin concentrations, agglutinability is neitherenhanced nor diminished by drug treatment. At higher anaesthetic concentrationssome cell lysis occurred during incubation. However, a sufficient number of non-lysedcells remained for use in the agglutination assays. Cell lysis increased further withaddition of suboptimal concentrations of WGA, concentrations which rarely causecytolysis in anaesthetic-free cells within the time limits of the experiment. Sincelysed cells tend to clump into amorphous masses, this makes the extent of agglutina-tion difficult to evaluate. Moreover, the presence of the appropriate hapten inhibitedthe lectin-mediated cytotoxicity as well as the agglutination. Agglutinated clumps werecomposed of packed cells with individual cell boundaries being distinguishable,whereas cell peripheries could not be distinguished within masses of lysed cells; celllysis also being a slower process. When these criteria are considered in the evaluationof the lectin-induced agglutination of cells treated with high concentrations of procaineor xylocaine, no change in the WGA-induced agglutinability of cells in this system wasobserved (Table 6, p. 236). However, these results are not conclusive due to thefactors mentioned previously.
Preliminary results indicated that chlorpromazine at a 2 x icr3 M concentrationinhibited lectin-mediated agglutination. Subsequent experiments were conducted toestablish the concentration of the phenothiazine tranquillizers which would affect theagglutination mediated by optimal concentrations of WGA and RCA. The results ofthese experiments are summarized in Table 4. The data indicate a similar inhibitingconcentration for all 3 drugs. At 2 x io~* M, chlorpromazine, prochlorperazine andtrifluoperazine do not inhibit agglutination. At 4 x io~* M, agglutination is occasionallyinhibited, while at 8 x io"4 M higher, this inhibition is consistently observed. Itappears that the minimum concentration of any one of these 3 phenothiazine deriva-tives which is effective in inhibiting the agglutination response lies between 4X io"4

Lectin-mediated agglutination 235
and 8 x io~* M. A series of experiments was designed to determine if the phenothiazinederivatives affect differentially the agglutination mediated by WGA, RCA, Con A, andSB A. Cells were incubated in 2 x io"3 M chlorpromazine or prochlorperazine, or in1 x io"3 M trifluoperazine after which they were tested for agglutinability at optimallectin concentrations. The results of these experiments are reported in Table 5. These
Table 4. Concentration effect of phenothiazine tranquillizers onWGA- and RCA-mediated agglutination
Chlorpromazine Prochlorperazine TrifluoperazineConcentration , * > , K 1 , * (
M WGA RCA WGA RCA WGA RCA
2xio-« OOO O OOO O OOO O4XIO-1 o » o om o 00 o8xio-> • • • • • • • • • O« O« • • •IOXIO-3 . . . • •
1-2x10-* • • • • • .
I-SXIO-3 • • • . . .
16X10-3 • . • •2-OXIO"3 • « • • • • • • • •
Cells were incubated in the presence of the drug for 1 h at room temperature. The finalconcentration of WGA and RCA waa 33 fig/ml and agglutination was scored after 2 min.The method for assaying agglutination is described in Materials and methods. O . o n e experimentcells agglutinate; 0 , one experiment, inhibition of agglutination.
Table 5. Effect of phenothiazine tranquillizers on lectin-induced agglutination
WGA RCA Con A SBA
Control + + + + + + + + + + + +Chlorpromazine-HCl 0 0 0 oProchlorperazine 0 0 0 oTrifluoperazine-HCl 0 0 0 oCells were incubated in the presence of the drug for 1 h. Chlorpromazine and prochlorpera-
zine were used at a concentration of 2 x io"3 M, trifluoperazine at 1 x io~' M. WGA and RCAwere used at a concentration of 33 /4g/ml and Con A and SBA at 166 /tg/ml. Agglutination wasscored at 4 min.
data indicate a similar effect of phenothiazine derivatives on the agglutination mediatedby any of the above-mentioned lectins. While some inhibition of agglutination isevident in drug-treated cells after 45 min, a 60-min drug incubation period is necessaryfor complete inhibition of the agglutination response (Figs. 7 and 8). The reversibilityof the inhibition of agglutination brought about by phenothiazine tranquillizers wasinvestigated. Drug-treated cells were rinsed by up to 3 times with fresh drug-freesaline and, in some cases, reincubated in the absence of drug for 2 h. Cells weretreated again for lectin-induced agglutinability. Under these experimental conditionsthe inhibition of lectin-induced agglutination by phenothiazine tranquillizers was notreversible.
After incubation with colchicine (30-120 /iu) and/or cytochalasin B (20/tg/ml),cells were tested for agglutinability with RCA and WGA at optimal and suboptimal

236 B. R. Fraser and S. E. Zalik
lectin concentrations. Agglutination appeared to be neither enhanced nor diminishedby either colchicine or cytochalasin B, or by both drugs in combination (Table 6).However, as was the case with cells treated with local anaesthetics, colchicine- andcytochalasin B-treated cells undergo lysis in the presence of these lectins more readilythan untreated cells; this was also observed in control DMSO-treated cells. Theseeffects complicated the evaluation of cell agglutinability as was the case for cellstreated with local anaesthetics.
Table 6. Effect of drugs on WGA- and RCA-mediated agglutination
WGA, 16 /zg/ml WGA, 33 /tg/ml RCA, 16 /zg/ml RCA, 33 /tg/mla n u r — * f • • — — — - , t *. f —
concen- Experi- Experi- Experi- Experi-tration, M Control mental Control mental Control mental Control mental
Procaine1 x io-1 4- + + + + +1-4 x io-1 . . + + + + + + + + • + + + + + •
Xylocainei-i x io- ' 4- + + + + + + +I-I X IO"1 + + + + + + + +
Colchicine3 x 10-5 4- + 4-4-4- 4-4-4-
I-2XI0-4 + + + + + + + +
Cytochalasin B,20 /zg/ml + + + 4- 4-4-
Colchicine andcytochalasin 4- 4- • . . 4 - 4 -
Cells were incubated in the presence of the drug for 1 h at room temperature. Agglutination was scored 4min after addition of lectin. •, severe cell lysis.
Dissociated blastula cells were observed under the phase-contrast microscope;cell size varies from 20 to 110/im in diameter, the majority of the pigmented cellsmeasuring approximately 70 fim. On many of the presumptive ectodermal and meso-dermal cells large rounded lobopodia were present (Fig. 9) similar to those recentlydescribed by Satoh, Kageyama & Sirakami (1976). These structures, usually trans-parent and devoid of cytoplasmic inclusions, changed position by ploughing aroundthe cell's periphery. Treatment of the cells with local anaesthetic did not prevent theextrusion of lobopodia or alter the general cellular outlines. Those concentrations ofphcnothiazine derivatives that failed to inhibit lectin-mediated agglutination did notappear to affect the surface morphology of cells or the formation of these processes.However, chlorpromazine, prochlorperazine, or trifluoperazine at concentrationswhich inhibited agglutination also inhibited the formation of lobopodia. In these cases,the morphology of the peripheral regions of the cells was altered. The transparentcytoplasm at the cell periphery disappeared and the plasma membrane appeared to bein close apposition to the cellular inclusions. This gave the cell a contracted appearancewith the yolk platelets appearing to protrude from the cell periphery (Fig. 10).

Lectin-mediated agglutination 237
Fig. 7. Agglutinated cells incubated for 1 h in CMFS.S. These are controls for thephenothiazine experiments and were treated with WGA. Conditions are the same asfor Fig. 1. x 40.Fig. 8. Cells pre-incubated with 8 x io~* M chlorpromazine, in the presence of WGA,5 min after addition of cells, x 40. Compare with Fig. 7.Fig. 9. Amphibian blastula cells under phase-contrast microscopy. Arrows indicateperipheral lobopodial extensions, x 400.Fig. 10. Cells incubated for 1 h in the presence of trifluoperazine (8 x io"1 M) underphase-contrast microscopy, x 400.
CEL 27

238 B. R. Fraser and S. E. Zalik
Colchicine-treated cells maintained smooth outlines with many typical lobopodialextensions similar to those of controls. On the other hand, cytochalasin B preventedthe extrusion of lobopodia and cells treated with this drug had the same appearance asthose treated with concentrations of phenothiazine tranquillizers that inhibit Iectin-induced agglutination. Cells treated simultaneously with both colchicine and cyto-chalasin B exhibited the same surface characteristics as cells treated with cytochalasinB alone.
Glutaraldehyde fixation of embryonic Xenopus cells for 24 h renders them non-agglutinable with either RCA or WGA (33 /ig/ml). Fixed cells agglutinated to acertain extent in the presence of Con A (166/tg/ml); however, this agglutinationresponse Was drastically reduced from that observed with unfixed cells. Maximalagglutination of fixed cells with Con A (166 /tg/ml) corresponds to a score of approxi-mately + + on the test scale, as opposed to a score of + + + + for unfixed cells.Under the phase-contrast microscope agglutinated clusters of fixed cells appear to becomposed of cells as well as yolk platelets. Yolk platelets released from ruptured cellsare found in all cell suspensions, fixed and unfixed; however, only in experimentsusing fixed cells has clustering of yolk with Con A been observed.
DISCUSSION
The evidence presented above suggests that cells of the late blastula of Xenopuslaevis possess surface macromolecules which are capable of combining with certainlectins and of participating in an agglutination reaction. The hapten specificities forthese lectins suggest that the exposed surface carbohydrates probably contain N-acetyl-D-glucosamine-like residues (WGA), D-galactose-like residues (RCA and SBA),iV-acetyl-D-galactosamine-like residues (SBA) and a-methyl-D-glucose or D-mannose-like residues (Con A) (Lis & Sharon, 1973). These experiments do not rule out thepossibility that blastula cells may possess FBP receptors; that cells may bind lectin,yet remain non-agglutinable is well documented (Sharon & Lis, 1975). The presence ofa variety of surface receptors displayed in an agglutinable condition seems to be acharacteristic of early embryonic cells (Krach et al. 1974; Zalik & Cook, 1976), cells atinitial stages of tissue differentiation (Moscona, 1974; Kleinschuster & Moscona, 1972)and many cell lines which have undergone malignant transformation (Sachs, 1974).
A strictly objective and efficient method for the quantification of the extent of cellagglutination has yet to be developed, especially for use with cells as large as those ofamphibian blastulae. Although the technique used in the present study for assessingcell agglutinability is rather subjective, it is useful and it is practical. However, adifficulty in these experiments is the variability in the cell numbers present in cellsuspensions in different agglutination experiments. The number of cells obviouslyinfluences the number of available lectin receptors, and the ratio of the number oflectin receptors to available lectin molecules affects the extent of the agglutinationresponse (Nicolson, 1974). However, in spite of this variability, the results reportedhere are valid as being indicative of general characteristics of agglutinability. For eachdose-response experiment, aliquots of cells from the same cell suspension were used in

Lectin-mediated agglutination 239
each of the agglutination tests. Thus the number of cells was comparable at all thelectin concentrations within each experiment, and the results of separate dose-responseexperiments in most cases compared favourably with each other.
Preferential agglutinability of distinct populations of cells was not observed amongthe cells derived from whole Xenopus blastulae. However, these results do not provethat such differences in agglutinability do not exist. Distinction between different celltypes in this study was based only on general characteristics of presumptive endoder-mal, ectodermal and mesodermal cells such as cell size, the degree of pigmentation,and the yolk content of the cells. Such features, although useful, are not definitive cellmarkers among embryonic amphibian cells. In addition, an important factor to consideris the possibility that cells in close proximity may influence the surface-related pro-perties of each other, among these, agglutinability. That the attachment of cells tolectin-derivatized beads elicits a general cell surface change, which may in turn inducealterations in the surface properties of neighbouring cells resulting in cell to celladhesion, has been suggested by Whitehead & Marcus (1975). Similarly, Chipowsky,Lee & Roseman (1973) studying cell attachment to galactose derivatized beads,described this process as ' . . . autocatalytic, in that the cells that are bound to the beadsseem to become more sticky, facilitating the binding of new cells to their surfaces..."The possible implications of such induced cooperativity have been largely overlookedin the investigations on lectin-mediated agglutinability. In the present study, such acooperative effect may have masked subtle differences in the agglutinability of cellsfrom different presumptive germ layers. In experiments using mixed cell populations,the response of a more agglutinable cell type may enhance or induce a response in cellswhich would otherwise not be as agglutinable. To investigate this point further, cellsfrom each of the 3 presumptive germ layers ought to be separately tested.
Neuraminidase treatment of embryonic Xenopus cells enhances their agglutinabilitywith RCA and SBA, but not with Con A or WGA. This suggests a change at the cellsurface resulting from removal of exposed terminal sialic acid residues which preferen-tially affects receptors for RCA and SBA. Since both of these lectins bind D-galactosyl-and iV-acetyl-D-galactosyl residues with high affinity, and since the most commonsugars subterminal to sialic acid residues in membrane glycoproteins are D-galactoseand A^-acetyl-D-galactosamine (Spiro, 1973), one of the most likely explanations for theenhanced agglutinability is that neuraminidase, by removing terminal sialic acidresidues, is exposing previously inaccessible receptors to the lectins. Such an explana-tion has been proposed to account for the appearance of cell agglutinability with RCAand/or SBA after neuraminidase treatment of malignant fibroblasts (Lloyd & Cook,1974), and chick blastoderm cells (Zalik & Cook, 1976). That neuraminidase treatmentof cells exposes SBA and RCA receptors that are otherwise not accessible to the lectinmolecules has been shown in other systems by lectin-binding studies (Vlodovosky &Sachs, 1975; Chan & Oliver, 1976). In the present study, enhanced agglutinability wasobserved in neuraminidase-treated blastula cells at suboptimal concentrations of theselectins. Under these conditions the factor limiting agglutinability would appear to bethe decreased lectin concentration and not the number of receptors available to thelectin. A mere increase in RCA and SBA receptors cannot adequately account for the
16-2

240 B. R. Fraser and S. E. Zalik
increased agglutinability observed after neuraminidase treatment at suboptimalconcentrations. Other factors such as the interference of charged sialic acid residueswith the mobility of membrane-associated RCA and SBA receptors, and the effect ofsialic acid on the interactions between lectin-bound receptors on adjacent cells maybe involved in determining the agglutinability of cells (Nicolson, 1974; Sachs, 1974).Neuraminidase could, by removing these sialic acid residues, enhance the agglutin-ability of cells without altering the number of exposed lectin receptors.
The mobility of lectin receptors, as manifested in the redistribution of lectin-receptorcomplexes at the cell surface, has been implicated in the agglutination of some types ofcells (Nicholson, 1974; Poste et al. 1975 a). It has been suggested that a peripheralcytoskeleton may modulate receptor mobility and thereby regulate the agglutinabilityof these cells (Edelman, 1976). For this reason, the possible role of colchicine andcytochalasin B-sensitive structures in the lectin-mediated agglutination of embryonicXenopus cells was investigated. Our results indicate that these compounds used eitherseparately or together do not affect the agglutinability of embryonic Xenopus cellswith WGA at optimal or suboptimal lectin concentrations. Transformed 3T3 cellsare rendered non-agglutinable by pre-treatment of the cells with colchicine (Posteet al. 19756); this is not the case for embryonic Xenopus cells. This difference in re-sponse to colchicine treatment may be indicative of a fundamental difference in surfaceproperties between embryonic and transformed cells, since agglutinability of the formerdoes not appear to be regulated by colchicine-sensitive or cytochalasin B-sensitiveelements. Other factors such as lipid flow in the cell membrane (Bretscher, 1976) mayinfluence lectin-mediated agglutinability of early embryonic cells. It is also of interestto note that cytochalasin B inhibits the extrusion of peripheral cytoplasmic lobopodialextensions, suggesting that this process may be controlled by contractile cytoplasmicelements which are sensitive to this drug. That colchicine does not inhibit the extensionof lobopodia suggests that microtubular functions are not essential to this process.
Local anaesthetics may effect surface-related phenomena such as cell capping(Ryan et al. 1974) and agglutination (Poste et al. 19756), the latter via a mechanisminvolving the disruption of structures sensitive to colchicine and cytochalasin B. It hasbeen postulated that the effect of local anaesthetics on cells mimics the combinedeffects of cytochalasin B and colchicine (Poste et al. 1975 a, b). Treatment of embryonicXenopus cells with either of the anaesthetics procaine or xylocaine does not appear toalter their agglutinability with RCA or WGA. This is consistent with the abovesuggestion in so far as agglutinability is concerned, since incubation of these cells withcytochalasin B plus colchicine also does not affect agglutinability. However, localanaesthetics do not change the morphology of dissociated cells or limit the extrusion oflobopodia, whereas the peripheral processes are absent in cytochalasin B-treated cells.This indicates that local anaesthetics and cytochalasin B may affect cells via differentmechanisms.
Phenothiazine tranquillizers share many membrane-modifying properties with localanaesthetics (Seeman, 1972). At low concentrations they protect cells from hypotoniclysis, whereas at higher concentrations they induce haemolysis (Ahtee, 1966; Seeman,1972). These compounds have been reported to inhibit virus-induced cell fusion

Lectin-mediated agglutination 241
(Poste & Reeve, 1972), the blastogenic response induced by Con A in mouse lympho-cytes (Ferguson, Schmidtke & Simmons, 1975), and the capping of Con A and anti-Igreceptor complexes (Ryan et al. 1974). The agglutination induced by WGA, RCA,SBA and Con A is similarly inhibited by each of the 3 phenothiazines chlorpromazine,prochlorperazine, and trifluoperazine. This inhibition occurs at a drug concentrationof 8XIO~*M and, under these experimental conditions, an incubation of 1 h isrequired to completely abolish cell agglutinability. There are many possible mechanismswhich could account for abolition of cell agglutinability. Chlorpromazine has beenreported to bind preferentially to specific cell surface molecules such as the humanHLA antigen Al, and beta-adrenergic receptors (Smeraldi & Scorza-Smeraldi, 1976);phenothiazines could be inhibiting the agglutination of embryonic Xenopus cells byinterfering directly with interactions between lectins and cell surface lectin receptors.Alternatively these compounds could affect the mobility of membrane components(Seeman, 1972) or promote differential expansion in the bilayer of the membranephospholipid (Sheetz, Painter & Singer, 1976). Chlorpromazine has also been reportedto preferentially solubilize specific proteins of human erythrocyte ghosts (Boivin &Galand, 1975). Thus, the phenothiazines could be inhibiting cell agglutination bymasking or removing lectin receptors from the plasma membrane or by altering theirdistribution at the cell surface.
Fixation of cells from Xenopus embryos inhibits their agglutination with RCA andWGA and severely restricts their agglutinability with Con A. These results are inagreement with those of others (Inbar et al. 1973; Sachs, 1974). This effect has beensuggested to be due to a decreased mobility of membrane proteins in a less-fluidmembrane (Inbar et al. 1973). In general, even fixed cells can be rendered agglutinableif the lectin concentration is increased above that required to agglutinate unfixed cells(Nicolson, 1974). It was observed in these experiments that the fixed yolk plateletsagglutinate. Whether or not this is due to the presence of Con A receptors in the yolkplatelet membrane, or to possible cross-linking effects of the fixative itself remains tobe established.
This paper represents part of the thesis submitted by B.R.F. as part of the requirements foran M.Sc. degree at the University of Alberta. We thank the National Cancer Institute ofCanada for their continuous support. Portions of this work were also supported by the NationalResearch Council of Canada.
REFERENCES
AHTEE, L. (1966). 5-hydroxytryptamine release from blood platelets and haemolysis of redblood cells of rabbit induced by phenothiazines and related compounds. Anns Med. exp. Biol.Fertn. 44, 431-452.
BOIVIN, P. & GALAND, C. (1975). Prot6ines de la membrane erythrocytaire. I I : solubilisation desprote'ines de la membrane d'e'rythrocytes humains par la chlorpromazine. Nouv. Revue jr.Hbnat. 15, 589-596.
BRETSCHER, M. S. (1976). Directed lipid flow in cell membrane. Nature, Lond. 260, 21.CHAN, L. L. & OLIVER, J. M. (1976). Changes in number, mobility, and topographical distribu-
tion of lectin receptors during maturation of chick erythroid cells. J. Cell Biol. 69, 647—658.CHIPOWSKY, S., LEE, Y. C. & ROSEMAN, S. (1973). Adhesion of cultured fibroblasts to insoluble
analogues of cell-surface carbohydrates. Proc. natn. Acad. Sci. U.S.A. 70, 2309-2312.

242 B. R. Fraser and S. E. Zalik
COOK, G. M. W. & STODDART, R. W. (1973). Surface Carbohydrates of the Eukaryotic Cell.London & New York: Academic Press.
CURTIS, A. S. G. (1973). Cell adhesion. In Progress in Biophysics and Molecular Biology, vol. 27(ed A. J. Butler & D. Noble), p. 315. Oxford: Pergamon.
EDELMAN, G. M. (1976). Surface modulation in cell recognition and cell growth. Science, N.Y.193, 218-226.
FERGUSON, R. M., SCHIMDTKE, J. R. & SIMMONS, R. L. (1975). Concurrent inhibition bychlorpromazine of concanavalin A-induced lymphocyte aggregation and mitogenesis. Nature,Lond. 256, 744-745-
FRASER, B. R. & ZALIK, S. E. (1976). Surface properties of embryonic amphibian cells: lectinmediated agglutination and the effects of selected drugs. Proc. Can. Fed. biol. Soc. 19, 2.
HAMBURGER, V. (1962). A Manual of Experimental Embryology. University of Chicago Press.HOLTFRETER, J. (i939)- Tissue affinity, a means of embryonic morphogenesis. In Foundations of
Experimental Embryology (ed. B. H. Willier & J. M. Oppenheimer), pp. 186-225. London:Prentice-Hall.
INBAR, M., HUET, C, OSEROFF, A. R., BEN-BASSAT, H. & SACHS, L. (1973). Inhibition of lectinagglutinability by fixation of the cell surface membrane. Biochim. biophys. Acta 311, 594-599.
INBAR, M. & SACHS, L. (1969). Structural difference in sites on the surface membrane of normaland transformed cells. Nature, Lond. 223, 710-712.
JOHNSON, K. E. & SMITH, E. P. (1976). The binding of Concanavalin A to dissociated embryonicamphibian cells. Expl Cell Res. 101, 63-70.
KLEINSCHUSTER, S. J. & MOSCONA, A. (1972). Interactions of embryonic and fetal neural retinacells with carbohydrate-binding phytoagglutinins: Cell surface changes with differentiation.Expl Cell Res. 70, 397-410.
KRACH, S. W., GREEN, A., NICOLSON, G. L. & OPPENHEIMER, S. B. (1974). Cell surface changesoccurring during sea urchin embryonic develoment monitored by quantitative agglutinationwith plant lectins. Expl Cell Res. 84, 191-198.
Lis, H., SHARON, N. & KATCHALSKI, E. (1969). Soybean haemagglutinin, a plant glycoprotein.J. biol. Chem. 241, 684-689.
Lis, H. & SHARON, N. (1973). The biochemistry of plant lectins. A. Rev. Biochem. 43, 541-574.LLOYD, C. W. & COOK, G. M. W. (1974). On the mechanism of increased aggregation by
neuraminidase of 16C malignant rat dermal fibroblasts in vitro. J. Cell Set. 15, 575-590.MORAN, D. (1974). Inhibitory effects of Concanavalin A on the development of the amphibian
embryos. J. exp. Zool. 188, 361-365.MOSCONA, A. (1974). Surface specifications of embryonic cells: lectin receptors, cell recognition,
and specific cell ligands. In The Cell Surface in Development (ed. A. A. Moscona), pp. 67-98.New York: John Wiley.
NICOLSON, G. L. (1974). The interaction of lectins with animal cell surfaces Int. Rev. Cytol.39. 89-190.
NICOLSON, G. L. & BLAUSTEIN, J. (1972). The interaction of Ricimis communis agglutinin withnormal and tumor cell surfaces. Biochim. biophys. Acta 226, 543-547.
NIEUWKOOP, P. D. & FABER, P. A. (1967). Normal Table of Xenopus laevis (Daudin). Amsterdam:North Holland Publishing.
O' DELL, D. S., TENCER, R., MONROY, A. & BRACHET, J. (1974). The pattern of ConcanavalinA-binding sites during the early development of Xenopus laevis. Cell Differentiation 3, 193-198.
POSTE, G. & REEVE, P. (1972). Inhibition of virus-induced cell fusion by local anaesthetics andphenothiazine tranquillizers. ,7. gen. Virology 16, 21-28.
POSTE, G., PAPAHADJOPOULOS, D. & NICOLSON, G. (1975 a). Local anaesthetics affect transmem-brane cytoskeletal control of mobility and distribution of cell surface receptors. Proc. natn.Acad. Set. U.S.A. 72, 4430-4434.
POSTE, G., PAPAHADJOPOULOS, D., JACOBSON, K. & VAIL, W. J. (19756). Effects of localanaesthetics on membrane properties: enhancement of the susceptibility of mammalian cellsto agglutination by plant lectins. Biochim. biophys. Acta 394, 520—539.
ROBERSON, M., NERI, A. & OPPENHEIMER, S. B. (1975). Distribution of concanavalin A receptorsites on specific populations of embryonic cells. Science, N. Y. 189, 639—640.
RYAN, G. B., UNANUE, E. R. & KARNOVSKY, M. J. (1974). Inhibition of surface capping ofmacromolecules by local anaesthetics and tranquillizers. Nature, Lond. 250, 56-57.

Lectin-mediated agglutination 243
SACHS, L. (1974). Lectins as probes for changes in membrane dynamics in malignancy and celldifferentiation. In The Cell Surface in Development (ed. A. A. Moscona), pp. 127-139.New York: John Wiley.
SATOH, N., KAGEYAMA, T. & SIRAKAMI, K-I. (1976). Motility of dissociated embryonic cells inXenopus laevis: its significance to morphogenetic movements. Dev. Growth Differ. 18, 55-67.
SEEMAN, P. (1972). The membrane actions of anaesthetics and tranquillizers. Pharmac. Rev. 24,583-655-
SHARON, N . & L I S , H. (1975). Useof lectins for the study of membranes. In Methods in MembraneBiology, vol. 3 (ed. E. D. Korn), pp. 147-200. New York: Plenum Press.
SHEETZ. M. P., PAINTER, R. G. & SINGER, S. J. (1976). Biological membranes as bilayer couples.III. Compensatory shape changes induced in membranes. J. Cell Biol. 70, 193-203.
SMERALDI, E. & SCORZA-SMERALDI, R. (1976). Interference between anti-HLA antibodies andchlorpromazine. Nature, Lond. 260, 532-533.
SPIRO, R. G. (1973). Glycoproteins. Adv. Protein Chem. 3, 349-467.TOWNES, P. L. & HOLTFRETER, J. (1955). Directed movements and selective adhesion of
embryonic amphibian cells. J. exp. Zool. ia8, 53-120.VLODOVSKY, I. & SACHS, L. (1975). Lectin receptors on the cell surface membrane and the
kinetics of lectin-induced cell agglutination. Expl Cell Res. 93, m-119.WHITEHEAD, J. S. & MARCUS, F. S. (1975). Cell surface: transfer of cellular adhesive properties
from cell to cell by induced membrane alterations. Proc. Soc. exp. Biol. Med. 148, 160—163.ZALIK, S. E. & COOK, G. M. W. (1974). Characterization of surface components of cells from
early chick blastoderms. J. Cell Biol. 63, 385A.ZALIK, S. E. & COOK, G. M. W. (1976). Comparisons of early embryonic and differentiating
cell surfaces: interactions of lectins with plasma membrane components. Biochim. biophys.Acta 419, 119-136.
{Received 18 January 1977)


















