
JOURNAL OF CHEMISTRY Vol. 256. No. 17, 10, 1981 in S ... · A Secretory Cholinesterase from...
8
THE JOURNAL OF BIOLOGICAL CHEMISTRY Printed in U. S. A. Vol. 256. No. 17, Issue of September 10, pp. 9316-9323, 1981 Cholinesterase in the Parasitic Nematode, Stephanurus dentatus CHARACTERIZATION AND SEX DEPENDENCE OF A SECRETORY CHOLINESTERASE* (Received for publication, February 20,1981) Marcia L. Rhoads From the Animal Parasitolom Institute, ApriculturalResearch, Science and Education Administration,United States Department of Agriculture, B&sville, Maryland 20705 An antigenic secretory protein with cholinesterase activity was isolated from the excretory gland cells of Stephanurus dentatus and was purified by gel filtration and ion exchange chromatography. The antigenicity of the cholinesterase was demonstrated by an esterase- active immunoprecipitate formed with s. dentatus an- tiserum and by the ability of the antiserum to protect the enzyme from heat inactivation. The enzyme was found to be secreted by the adult nematodes during in vitro cultivation. The level of cholinesterase activity and its release from the excretory gland cells of the parasite were 27-fold greater in the male than in the female. Ninety per cent of the enzyme activity was localized in the soluble fractionof the gland cells. The molecular weight of the enzyme, estimated by sucrose density gradient centrifugation, was 100,000. Two mo- lecular forms were separated by isoelectrofocusing, with isoelectric points of 7.0 and 6.9. At optimum sub- strate concentrations, the rate of hydrolysis of acetyl- thiocholine was 8 times greater than that of butyryl- thiocholine; the Michaelis constants were 560 PM and 81 PM foracetylthiocholine and butyrylthiocholine, re- spectively. The enzyme exhibited substrate inhibition at substrate concentrations greater than 10 mM and was inhibited by eserine sulfate, 1,5-bis(4-allyldimethy- lammoniumphenyl)-pentan-3-one dibromide, Tris, and acetone. The enzyme was highly unstable in dilute pro- tein solutions. Mammalian cholinesterases are classified according to their substrate specificity and inhibitor sensitivity as acetylcholin- esterase (acetylcholine hydrolase; EC 3.1.1.7) or nonspecific or pseudocholinesterase (acylcholine acylhydrolase; EC 3.1.1.8) (1-4). Cholinesterases are widely distributed among nervous and non-nervous tissues of vertebrates and inverte- brates (4). Pseudocholinesterases, in particular, are often as- sociated with secretory tissues and their secretions, for ex- ample, salivary glands (5, 6), sweat glands (7,8), and pancreas (9). Apart from the well defined role of acetylcholinesterase in the transmission of nerve impulses across the neuromuscular junction (lo), the physiological role of the non-neuronal ace- tylcholinesterases and pseudocholinesterases hasnot been established. Acetylcholinesterases have been demonstrated in nervous tissues of several nematodes, including the livestock parasites Ascaris suum (11, 12) and Cooperia pectinata (13). These enzymes are of interest as possible targets of anthelmintic drugs (14, 15). Of perhaps greater importance, in view of the * The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. unique association of host and parasite, is the presence of a high level of acetylcholinesterase in the secretory glands of a number of parasitic nematodes, such as Nippostrongylus bras- iliensis (16), Trichostrongylus colubriformis, and Oesopha- gostomum radiatum (13). Acetylcholinesterases are secreted by these nematodes during in vitro cultivation (13). In addi- tion, the acetylcholinesterases released by these nematodes during the course of infection stimulate the production of specific antiacetylcholinesterase antibodies in their hosts (17- 20). One of these nematodes (N. brasiliensis) modulates the concentration and isozyme pattern of the secreted enzyme in response to this immune reaction of its host (17, 21, 22). Despite the potential of enzyme secretions of parasites as important biological and immunological factors in determin- ing host-parasite specificity, the acetylcholinesterases from the secretory glands of parasitic nematodes have not been extensively characterized. Furthermore, the small sue of these particular nematodes has precluded the isolation of cholines- terases from individual worm tissues. In this investigation, the distribution of cholinesterase was examined in Stephanurus dentatus, a parasite infecting the kidneys of swine. This is the fiist report of a high concentra- tion of cholinesterase located in and secreted from the excre- tory gland cells of a tissue-dwelling nematode. The level of cholinesterase and its release from the excretory gland cells are significantly higher in male than in female adult worms. Thisstudy defines the biochemical, kinetic, and antigenic properties of the purified secretory cholinesterase from S. dentatus and examines the possible significance of this secre- tory activity in the host-parasite relationship. EXPERIMENTAL PROCEDURES’ RESULTS AND DISCUSSION Whole worm extracts of male and female S. dentatus hy- drolyze acetylthiocholine (Table I). The cholinesterase activ- ity is approximately 40 times higher in male than in female worms. For comparison, the specific activity of cholinesterase was determined in 0. radiatum, an intestinal nematode of cattle, and N. brasiliensis, an intestinal nematode of rats. Cholinesterase specific activities cited in the literature (13,20, 23) are included in the table for comparison. Only small variations in cholinesterase activity between male and female parasitic nematodes have been reported: male N. brasiliensis ’ Portions of this paper (including “Materials,” “Experimental Pro- cedures,” Figs. 3-10, Table V, and additional references) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Be- thesda, MD 20814. Request Document No. 81M-384, cite author, and include a check or money order for $7.20 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press. 9316
Transcript of JOURNAL OF CHEMISTRY Vol. 256. No. 17, 10, 1981 in S ... · A Secretory Cholinesterase from...

THE JOURNAL OF BIOLOGICAL CHEMISTRY
Printed in U. S. A. Vol. 256. No. 17, Issue of September 10, pp. 9316-9323, 1981
Cholinesterase in the Parasitic Nematode, Stephanurus dentatus CHARACTERIZATION AND SEX DEPENDENCE OF A SECRETORY CHOLINESTERASE*
(Received for publication, February 20,1981)
Marcia L. Rhoads From the Animal Parasitolom Institute, Apricultural Research, Science and Education Administration, United States Department of Agriculture, B&sville, Maryland 20705
An antigenic secretory protein with cholinesterase activity was isolated from the excretory gland cells of Stephanurus dentatus and was purified by gel filtration and ion exchange chromatography. The antigenicity of the cholinesterase was demonstrated by an esterase- active immunoprecipitate formed with s. dentatus an- tiserum and by the ability of the antiserum to protect the enzyme from heat inactivation. The enzyme was found to be secreted by the adult nematodes during in vitro cultivation. The level of cholinesterase activity and its release from the excretory gland cells of the parasite were 27-fold greater in the male than in the female. Ninety per cent of the enzyme activity was localized in the soluble fraction of the gland cells. The molecular weight of the enzyme, estimated by sucrose density gradient centrifugation, was 100,000. Two mo- lecular forms were separated by isoelectrofocusing, with isoelectric points of 7.0 and 6.9. At optimum sub- strate concentrations, the rate of hydrolysis of acetyl- thiocholine was 8 times greater than that of butyryl- thiocholine; the Michaelis constants were 560 PM and 81 PM for acetylthiocholine and butyrylthiocholine, re- spectively. The enzyme exhibited substrate inhibition at substrate concentrations greater than 10 mM and was inhibited by eserine sulfate, 1,5-bis(4-allyldimethy- lammoniumphenyl)-pentan-3-one dibromide, Tris, and acetone. The enzyme was highly unstable in dilute pro- tein solutions.
Mammalian cholinesterases are classified according to their substrate specificity and inhibitor sensitivity as acetylcholin- esterase (acetylcholine hydrolase; EC 3.1.1.7) or nonspecific or pseudocholinesterase (acylcholine acylhydrolase; EC 3.1.1.8) (1-4). Cholinesterases are widely distributed among nervous and non-nervous tissues of vertebrates and inverte- brates (4). Pseudocholinesterases, in particular, are often as- sociated with secretory tissues and their secretions, for ex- ample, salivary glands (5, 6), sweat glands (7 ,8) , and pancreas (9). Apart from the well defined role of acetylcholinesterase in the transmission of nerve impulses across the neuromuscular junction (lo), the physiological role of the non-neuronal ace- tylcholinesterases and pseudocholinesterases has not been established.
Acetylcholinesterases have been demonstrated in nervous tissues of several nematodes, including the livestock parasites Ascaris suum (11, 12) and Cooperia pectinata (13). These enzymes are of interest as possible targets of anthelmintic drugs (14, 15). Of perhaps greater importance, in view of the
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
unique association of host and parasite, is the presence of a high level of acetylcholinesterase in the secretory glands of a number of parasitic nematodes, such as Nippostrongylus bras- iliensis (16), Trichostrongylus colubriformis, and Oesopha- gostomum radiatum (13). Acetylcholinesterases are secreted by these nematodes during in vitro cultivation (13). In addi- tion, the acetylcholinesterases released by these nematodes during the course of infection stimulate the production of specific antiacetylcholinesterase antibodies in their hosts (17- 20). One of these nematodes (N. brasiliensis) modulates the concentration and isozyme pattern of the secreted enzyme in response to this immune reaction of its host (17, 21, 22).
Despite the potential of enzyme secretions of parasites as important biological and immunological factors in determin- ing host-parasite specificity, the acetylcholinesterases from the secretory glands of parasitic nematodes have not been extensively characterized. Furthermore, the small sue of these particular nematodes has precluded the isolation of cholines- terases from individual worm tissues.
In this investigation, the distribution of cholinesterase was examined in Stephanurus dentatus, a parasite infecting the kidneys of swine. This is the fiist report of a high concentra- tion of cholinesterase located in and secreted from the excre- tory gland cells of a tissue-dwelling nematode. The level of cholinesterase and its release from the excretory gland cells are significantly higher in male than in female adult worms. This study defines the biochemical, kinetic, and antigenic properties of the purified secretory cholinesterase from S. dentatus and examines the possible significance of this secre- tory activity in the host-parasite relationship.
EXPERIMENTAL PROCEDURES’
RESULTS AND DISCUSSION
Whole worm extracts of male and female S. dentatus hy- drolyze acetylthiocholine (Table I). The cholinesterase activ- ity is approximately 40 times higher in male than in female worms. For comparison, the specific activity of cholinesterase was determined in 0. radiatum, an intestinal nematode of cattle, and N. brasiliensis, an intestinal nematode of rats. Cholinesterase specific activities cited in the literature (13,20, 23) are included in the table for comparison. Only small variations in cholinesterase activity between male and female parasitic nematodes have been reported: male N. brasiliensis
’ Portions of this paper (including “Materials,” “Experimental Pro- cedures,” Figs. 3-10, Table V, and additional references) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Be- thesda, MD 20814. Request Document No. 81M-384, cite author, and include a check or money order for $7.20 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
9316

A Secretory Cholinesterase from Stephanurus dentatus 9317
TABLE I Comparison of cholinesterase activity (measured with
acetylthiocholine as substrate) in whole worm extracts of s. dentatus, 0. radiatum, N. brasiliensis, and 0. venulosum
Nematode Sex Activity Activity prnol/rnin/rng pmal /h /g wet
protem weight S. dentatus Male 0.65" 2000" S. dentatus Female 0.015" 50" 0. radiatum Mixed 0.24" 97@ N. brasiliensis Mixed 0.18" 956" N. brasiliensis Male 1038' N. brasiliensis Female 948' 0. venulosum Mixed 1288'' 0. venulosum Male 1080'' 0. venulosum Female 1117"
a Values determined in this study. Value from Ref. 13.
' Values from Ref. 23. '' Values from Ref. 20.
homogenate has a slightly higher specific activity than the female homogenate (23); the reverse is found for male and female Oesophagostomum venulosum, an intestinal parasite of sheep (20). The cholinesterase activity in male S. dentatus is in the high value range reported for other nematode species; in comparison, S. dentatus females have an exceptionally low level of cholinesterase activity.
Tissue Localization of Cholinesterase Activity The cholinesterase activity in individual tissues of S. den-
tatus male and female adult worms is shown in Table 11. All male worm tissues had a higher cholinesterase specific activity than did the corresponding female tissue. (The esophageal glands from male and female worms were not compared because of the small amount of tissue available.) The male excretory gland cells contained by far the highest enzyme specific activity, followed by the male reproductive system and the female excretory gland cells. All other tissues had low levels of cholinesterase activity. The largest sex-related differ- ence in specific activity of cholinesterase was found in the excretory gland cells where the activity in male excretory gland cells was 27 times higher than the activity in female excretory gland cells.
A high level of cholinesterase activity was demonstrated in the male reproductive system. In mammalian species, pseu- docholinesterase has been demonstrated in the reproductive tract of male guinea pigs (5) and acetylcholinesterase in the spermatozoa of a number of species (24-26). Cholinesterase has been demonstrated histochemically in the vas deferens of T. colubriformis and Trichostrongylus axei, gastrointestinal parasites of sheep (13, 18). The specific activity of cholines- terase in an extract of testes from A. suum, an intestinal parasite of swine, was reported at 0.0583 pmol of acetylcholine hydrolyzed per min per mg of protein (27); this is one-half the cholinesterase specific activity found in this study in the male reproductive tissue from S. dentatus. Functions proposed for the cholinesterases in male reproductive tissues include in- volvement in muscle contraction for ejaculation and in the propulsion mechanism of spermatozoa (28).
Properties of the Enzymes In all adult tissues tested, acetylthiocholine was hydrolyzed
approximately 4 times faster than butyrylthiochohe. The hydrolysis of acetylthiocholine in all adult tissues tested was inhibited approximately 93% by 10 p~ eserine sulfate and 78% by 10 p~ BW284C5L2 The cholinesterase in both male excre-
* The abbreviations used are: BW284C51, 1,5-bis (4-allyldimeth-
tory gland cells and male reproductive system was 90% soluble; the remaining activity could be solubilized by 0.1% Triton X- 100.
A low level of cholinesterase activity was found in the infective larval stage of S. dentatus (Table 11). In contrast to the cholinesterase activity found in adult tissues, the activity of this enzyme hydrolyzed acetylthiocholine 19 times faster than butyrylthiocholine and 68% of the enzyme was in the soluble fraction, The specificity of the enzyme is, therefore, either modified during development or a new enzyme species is synthesized during a later developmental phase.
Secretion of Cholinesterase The level of cholinesterase activity in maintenance medium
after 20 h incubation with adult male and female worms is shown in Table 111. Although individual cultures showed wide variation, the average cholinesterase activity released per male worm in a 20-h period was 29 times that released per female worm. This is the same difference in enzyme activity as found between extracts of male and female excretory gland cells (Table 11) and indicates comparable rates of secretion from these gland cells by both sexes under the described in vitro incubation conditions; 45% of the total cholinesterase activity is released from the excretory gland cells of both male and female worms in a 20-h period. Protease inhibitors which are also secreted from the excretory gland cells of adult S. dentatus are present in approximately the same concentra- tions and are released in nearly identical concentrations by both male and female worms (29). The enzyme activity re- leased was higher for both male and female worms cultured in API-16 medium as compared to the enzyme activity released in Earle's balanced salt solution. In 24 h, adults of N . brasi- liensis and Necator americanus (a human hookworm) syn- thesize and secrete 4 times the amount initially present in these worms (30). The lower rate of secretion by S. dentatus may reflect the instability of the enzyme at low protein concentrations.
Characterization of the Cholinesterase from the Excretory Gland Cells of Male Worms
Electrophoretic Analysis-The protein-staining patterns in polyacrylamide gels after electrophoresis of the fractions ob- tained at the different stages of cholinesterase purification (Table V, miniprint supplement) are shown in Fig. 1 (gels A, B, and C). The pattern of enzyme activity in the polyacryl- amide gels after electrophoresis of the pooled fraction from the DEAE-Sephacel column is also shown. The identical band is stained for both nonspecific esterase activity, using a-naph- thy1 acetate as substrate (gel D), and for cholinesterase activ- ity, using either acetylthiocholine (gel E ) or butyrylthiocho- line (gel F) as substrate. The same pattern of enzyme activity in polyacrylamide gels is found after electrophoresis of the crude extract of male excretory gland cells and of the pooled fraction from the Sephacryl column (not shown). Thus, the esterase activity present in the male excretory gland cells appears to be exclusively cholinesterase. Furthermore, by the criteria of polyacrylamide gel electrophoresis, there appears to be one molecular form of the male excretory gland cell cholinesterase and this enzyme hydrolyzes both acetylthio- choline and butyrylthiochoIine. This conclusion is supported by fractionation studies in which the same distribution of enzyme activity is obtained with either acetylthiocholine or butyrylthiocholine as substrate (data not shown).
Kinetic Studies-The kinetic parameters (K, and V,,,)
ylammoniumphenyl)-pentan-3-one dibromide; DTNB, 5,5'-dithiobis- 2-nitrobenzoic acid; PBS, phosphate-buffered saline.

9318 A Secretory Cholinesterase from Stephanurus dentatus
TARLE I1 Cholinesterase activity in individual tissues of 5'. dentatus
Relative rate of Per cent inhibi- Per cent inhibi- Tissue and sex Per cent activity IO' X Specific activity hydrolysis of tion by eserine tion by
soluble (Mean k S. E.)". butyrylthiocho- sulfate RW2MC51 line' ( 1 0 /" ( 1 0 pM)
Excretory glands Male 83-91 1900 f 120 24.4 95.0 76.0 Female nd" 71 * 7.4 21.1 94.4 82.2
Reproductive system Male Female
Intestines Male Female
Body walls Male Female
90 110 f 13 27.8 90.0 77.0 nd 4.6 f 0.35" nd nd nd
nd 40 f 8.1 23.5 93.2 75.6 nd 2.50 f 0.40 nd nd nd
nd 25 f 6.3 28.6 nd nd nd 4.9 f 0.21 nd nd nd
Esophageal glands, male and female nd 11.0 20 nd nd
Infective larvae 68 16.0 5.3 94.7 84.2 " Specific activity is defined as micromoles of acetylthiocholine hydrolyzed per min per mg of protein.
Results are expressed as the mean of three separate tissue extracts; values for esophageal glands and infective larvae are from one extract. 100 X rate of hydrolysis of but.v.vlthiocholine + rate of hvdrolvsis of acetylthiocholine. ' Not determined. '' Activity is not linear with time.
. . "
TARLE 111 Release of cholinesterase into culture medium API-16 by adult S.
dentatus I O ' x Cholin-
Experiment No, esterase units released per
worm in 20 h" Sex Range
Male 1 (16)* 370 nd" 2 (10) 350 20-960 3 (10) 200 80-540 4 (lo)'! 110 nd
Female 1 (18) 21 nd 2 (6) 2.9 nd 3 (8) 8.6 nd 4 (lor' 1.4 nd
I' In experiments 2 and 3 (male), the results are expressed as the mean activity of the individual cultures. In all other experiments, the cultures were pooled and then assayed for cholinesterase activity.
hValues in parentheses are the total number of worms in the experiment.
' Not determined. " In experiment 4, worms were maintained in Earle's balanced salt
solution with antibiotics; all other experiments are in AH-16 culture medium.
were determined graphically according to the method of Hof- stee (31). The Michaelis constants for the reactions of acetyl- thiocholine and butyrylthiocholine with male excretory gland cell extract are 560 pM and 81 pM, respectively (Fig. 5, mini- print supplement). The maximum velocities of the reactions with acetylthiocholine and butyrylthiocholine are 5.3 pmol/ min/mg of protein and 0.62 pmol/min/mg of protein, respec- tively. A t optimum substrate concentrations, the male excre- tory gland cell cholinesterase hydrolyzed acetylthiocholine 8 times faster than butyrylthiocholine. This substantial rate of hydrolysis of butyrylthiocholine and the cytosolic localization of the cholinesterase from the male excretory gland cells (Table 11) are characteristics not in common with mammalian acetylcholinesterases which show little hydrolysis of butyryl- thiocholine and are membrane bound (2-4). EDTA (1 mM), MgC12 (2 mM), and CaC12 (0.5 and 1.0 mM) had no effect on
A B C D E F
+ FIG. 1. Polyacrylamide gel electrophoresis of the fractions
obtained a t each stage of the purification of the male excretory gland cell cholinesterase. A, initial male excretory gland cell ex- tract stained for protein (124 pg of protein); E , pooled fraction from the Sephacryl S-200 column stained for protein (143 pg of protein); C, pooled fraction from the DEAE-Sephacel column stained for protein (28 pg of protein); D, pooled fraction from the DEAE-Sephacel column stained for nonspecific esterase (28 pg of protein); E , pooled fraction from the DEAE-Sephacel column stained for cholinesterase with acetylthiocholine as substrate (28 pg of protein); and F, pooled fraction from the DEAE-Sephacel column stained for cholinesterase with butyrylthiocholine as substrate (57 pg of protein).
the rate of hydrolysis of acetylthiocholine by male excretory gland cell cholinesterase.
The effect of substrate concentration (0.01-100 mM) on the rate of hydrolysis by male excretory gland cell cholinesterase of acetylthiocholine and butyrylthiocholine is shown in Fig. 6, miniprint supplement. Substrate concentrations greater than 10 mM inhibit the reaction; the inhibition by butyrylthiocho- line is greater than the inhibition by acetylthiocholine. The cholinesterase from N . brasiliensis was inhibited when the concentration of acetylthiocholine exceeded 20 mM (23). The cholinesterase from Metastrongylus a p i , a lungworm of swine, exhibited substrate inhibition at concentrations of ace- tylthiocholine and butyrylthiocholine greater than 10 mM (32). These values are similar to that reported for spermatozoa acetylcholinesterase which is inhibited by acetylthiocholine

A Secretory Cholinesterase from Stephanurus dentatus 9319
concentrations greater than 10 mM (33), but are significantly higher than the values reported for the acetylcholinesterases of most other tissues (2.5-3.0 mM) (4). Nonspecific cholines- terases are reported not to exhibit excess substrate inhibition
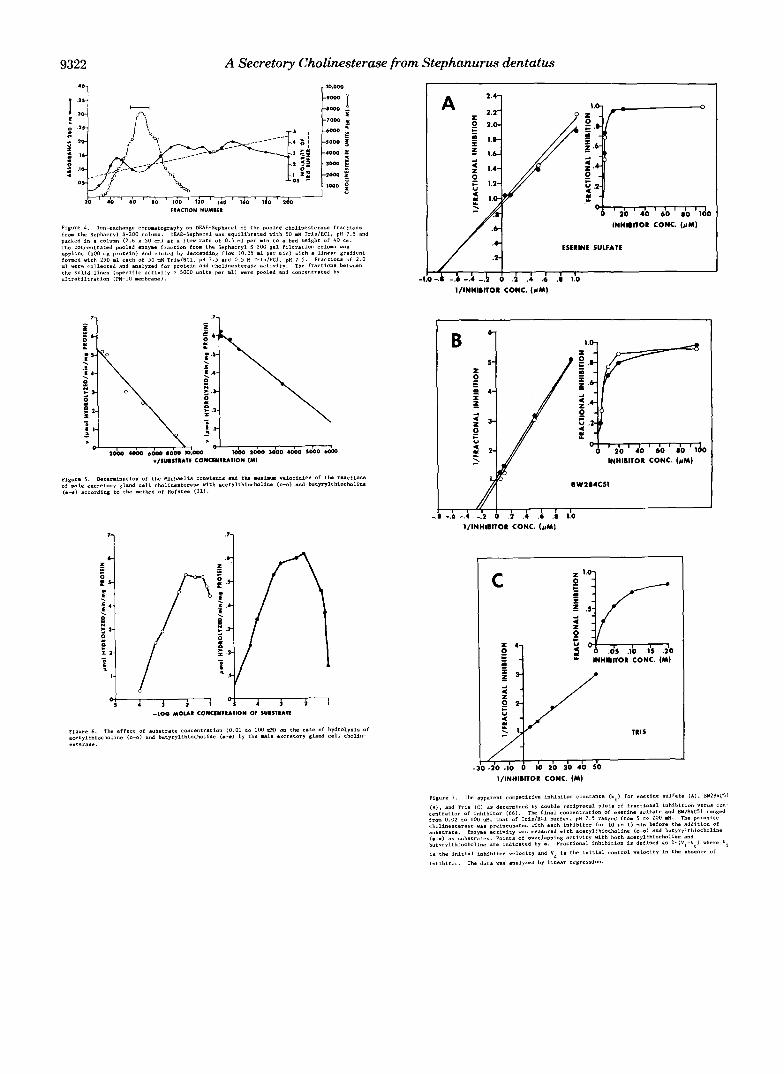
Inhibitor Studies-The effects of the specific cholinesterase competitive inhibitor eserine sulfate and of the specific ace- tylcholinesterase competitive inhibitor BW284C51 on the rate of hydrolysis of acetylthiocholine and butyrylthiocholine by male excretory gland cell extract are shown in Fig. 7, A and B, miniprint supplement. Under the specific assay conditions, the per cent inhibition by eserine sulfate and by BW284C51 of the male excretory gland cell cholinesterase was essentially the same using either substrate (insets of Fig. 7, A and B ) . The inhibitor constants for eserine sulfate were calculated at 0.53 VM and 0.13 VM for acetylthiocholine and butyrylthio- choline, respectively. BW284C51 was a less efficient inhibitor of male excretory gland cell cholinesterase, as indicated by higher Ki values, and this inhibition was slightly less when measured with acetylthiocholine, Ki = 1.7 p ~ , as compared with butyrylthiocholine, K, = 0.52 VM.
The effect of Tris on the rate of hydrolysis of acetylthio- choline is shown in Fig. 7C, miniprint supplement. Tris in- hibits the hydrolysis of acetylthiocholine by male excretory gland cell cholinesterase, with a K , calculated a t 22 mM. The inhibitory effect of Tris on the pseudocholinesterase from horse plasma and the acetylcholinesterase from the electric organ of Torpedo marmorata, using both acetylthiocholine and butyrylthiocholine as substrates, has been reported (K t = 13 and 14 mM for the respective enzymes with either substrate) (34). I t was further noted in the cited study that when M e or Ca2+ was not included in the reaction, an activation rather than an inhibition by Tris resulted. The inhibition by Tris of the male excretory gland cell cholines- terase occurred in the absence of Mg2' and Ca'+. The activity of the cholinesterase from Haemonchus contortus, an abom- asal (stomach) parasite of sheep, was unaffected by Tris, but was inhibited by borate (35).
The activity of the cholinesterase from the male excretory gland cells was inhibited by acetone. This inhibition was demonstrated during staining procedures for esterase activity after polyacrylamide gel electrophoresis. No activity could be detected in the gels when a-naphthyl acetate, a general ester- ase substrate, was dissolved in acetone (2% final concentra- tion). Cholinesterases have previously been reported to be inhibited by acetone (4).
Apparent Molecular Weight-The apparent molecular weight of the male excretory gland cell cholinesterase was estimated from the sedimentation coefficient determined by sucrose density gradient centrifugation. The sedimentation coefficient was estimated at 5.8 and corresponds to an average molecular weight 2 S. E. of 100,000 * 4,000 (Fig. 8, miniprint supplement). The apparent molecular weight of the enzyme as estimated by polyacrylamide gel electrophoresis was 110,000 (Fig. 9, miniprint supplement). These values are in close agreement with one another but are higher than the molecular weight estimated by gel filtration (80,000 * 7,000 S. E.). This discrepancy may reflect the glycoprotein nature of the enzyme resulting in adsorption by Sephacryl and thus a lower molecular weight estimate by gel filtration.
The molecular weights of the cholinesterases from N . bras- iliensis and T. colubriformis were estimated by gel filtration a t 70,000 (36). N. americanus cholinesterase has an estimated molecular weight by gel filtration of 380,000-400,OOO (37). This enzyme is membrane bound (10% of the activity is in the soluble fraction) (38). It is, therefore, surprising that this enzyme has been reported to be secreted at a high rate during
(2-4).
in uitro cultivation (30, 38). Isoelectric Point-The isoelectric point of the cholinester-
ase from the male excretory gland cells, estimated by isoelec- trofocusing (pH gradient of 3-10) in polyacrylamide cylindri- cal gels, was 7.0 k 0.042 S. E. Two molecular forms of the enzyme were separated by electrofocusing in a 6-8 pH gra- dient; each band had both nonspecific esterase (Fig. 1OA, miniprint supplement) and cholinesterase activity (Fig. 10B, miniprint supplement). The isoelectric points for the cholin- esterase isozymes were calculated at 7.0 and 6.9.
Immunochemical Studies-The presence of antibodies in the serum of swine infected with S. dentatus, having specific- ity for the parasite cholinesterase, was demonstrated by spe- cific enzyme staining of the precipitin bands obtained after immunodiffusion in agarose (Fig. 2). The cholinesterase frac- tion from the DEAE-Sephacel column (in the center wells) was reacted by double diffusion with S. dentatus antiserum (in the top wells) and with control serum (in the bottom wells). Precipitin reactions which were stained with Coomassie blue occurred after incubation overnight a t 4 "C (Fig. 2B). Esterase activity was demonstrated in the immunoprecipitate (Fig. 2 A ) ; a single precipitin band was stained. There were no precipitin reactions nor enzyme staining reactions with control serum. All control slides for the enzyme staining reaction were negative.
The effect of the globulin fractions of S. dentatus antiserum and of control serum on the heat inactivation of the cholin- esterase activity from the excretory gland cells of male worms is shown in Table IV. After incubation of the male excretory gland cell cholinesterase with either S. dentatus antiserum or control serum and with PBS, 100% of the cholinesterase activity remains in the supernatant fractions after centrifu- gation. Heating these supernatants at 60 "C for 5 min or longer inactivates 100% of this activity in the enzyme samples incu- bated with control serum or PBS. Stability of the parasite cholinesterase to heat inactivation occurred only with S. den- tatus antiserum. A ratio of 101 (globulins/extract) protected an average of 5.3% of the enzyme activity from heat inactiva- tion after 5-20 min; a ratio of 50:l increased this protection to 54%.
To determine whether the activity found in the supernatant fraction of the enzyme sample incubated with S. dentatus antiserum is indeed complexed with antibodies, goat anti- swine serum was added to each of the supernatants. No activity was precipitated by goat anti-swine serum from the enzyme sample incubated with either control serum or with PBS. Forty-two per cent of the activity was precipitated by goat anti-swine serum from the supernatant fraction of the enzyme sample incubated with S. dentatus antiserum; this is approximately the same percentage of activity which was found to be heat stable. Therefore, it does appear that soluble
w " "
A
FIG. 2. Ouchterlony immunodiffusion. Top wells contained S. dentatus antiserum (20 p l ) ; center wells contained pooled fraction from the DEAE-Sephacel column (20 pl, 11 pg of protein); bottom wells contained control swine serum (20 pl). Slide A was stained for nonspecific esterase. Slide B was stained with Coomassie blue.

9320 A Secretory Cholinesterase from Stephanurus dentatus
TABLE IV Effect of the globulin fractions of serum from S. dentatus-infected pigs and from helminth-free pigs on the heat inactivation of the cholinesterase from the excretory gland cells of male S. dentatus
Per cent activity remaining Per cent
(v/v) of activity Ratio Per cent after heating at 60 “C for: activity
precipi- globu- remain- tated
or, LI
PBS) to superna- 5 min iyn mln m,n lins (or ing in with goat
extract tant swine se- ‘5 20 anti-
S. dentatus 1O:l 100 antiserum 50:l 100
Control 1O:l 1 0 0 serum 50:l 100
PBS 1O:l 1 0 0 50:l 100
‘* Not determined.
rum
5.3 7.9 5.3 2.6 nd“ 72 42 50 50 42 O O O O n d 0 0 0 0 0 O O O O n d 0 0 0 0 0
antigen-antibody complexes are formed with the parasite cho- linesterase and that this complexing confers stability to de- naturation by heat to the enzyme. Furthermore, this complex of cholinesterase and antibody retains enzymatic activity. This is true of the enzyme-antibody complex of other parasitic nematodes (19, 39) as well as the bovine erythrocyte acetyl- cholinesterase-antibody complex (40).
Concluding Discussion Cholinesterase activity was present in all tissues of male
and female adult S. dentatus. The highest activity was not associated with nervous or m~sc le tissues, as might be ex- pected, but was found in the excretory gland cells of male worms. The cholinesterase from the male excretory gland cells manifests properties which resemble mammalian acetylcho- linesterases; however, a definite classification of the enzyme is difficult because not all properties follow those of the classical acetylcholinesterase enzyme. This difficulty in defin- ing a clear cut division between the properties of acetylcholin- esterase and pseudocholinesterase for the invertebrate en- zymes has been discussed by Chadwick (41).
The interpretation of the sex dependence of the level of cholinesterase activity in the excretory gland cells of S. den- tatus remains unclear, as does the enzyme’s physiological function. One suggested function for the enzyme in gastroin- testinal nematodes is as a “biological holdfast” enabling the worms to resist expulsion by reducing the peristaltic action of the host’s intestinal tract (16,42). However, a number of gut- dwelling nematodes, for example, Trichinella spiralis (43) and Ancylostoma tubaeforme, the cat hookworm (43), do not secrete cholinesterase. In addition, acetylcholinesterase is found in the secretory glands of non gut-dwelling filarial worms, such as Dipetalonema viteae (44) and Dictyocaulus filaria (15), although there is no evidence that the enzyme is secreted by these worms (43). Adult S. dentatus are located in thick walled cysts along the ureter of their host and, therefore, the anesthetic action of the released enzyme would be of little benefit.
The functional significance of worm secretions may be their ability to modulate the host’s immune response. It was pre- viously speculated that the protease inhibitors secreted from the excretory gland cells of male and female adult S. dentutus may protect the parasite from the lytic enzymes of the host’s white blood cells (46). These inhibitors may have an immu- noregulatory role similar to that proposed for the protease inhibitors found in serum namely, an-macroglobulin (47) and al-antitrypsin (48). The ability of cholinergic agents such as acetylcholine and carbamylcholine to enhance the release of chemical mediators of inflammation from mast cells, poly-
morphonuclear leukocytes, and lymphocytes (49) suggests a role for the parasite cholinesterase in the regulation of the inflammatory response of its host.
Acknowledgments-I would like to thank Dr. Harry Herlich, Dr. Joseph Urban, and Dr. Frank W. Douvres (Beltsville Agricultural Research Center, Animal Parasitology Institute, Beltsville, MD) for the materials they kindly contributed and the Smithfield Packing Co., Smithfield, VA, and Briggs and Co., Landover, MD for supplying the swine kidneys. I would also like to thank Gary Peck for preparing the illustrations and to acknowledge the technical assistance of A. W. Jones.
1.
2.
3. 4.
5.
6. 7. 8. 9.
10. 11. 12. 13.
14.
15. 16. 17. 18.
19.
20.
21.
22.
23. 24. 25.
26.
27.
28. 29.
30. 31. 32.
33.
3 4 . 35.
36.
37.
38.
REFERENCES
Rothenberg, M. A., and Nachmansohn, D. (1947) J. Biol. Chem.
Augustinsson, K. B., and Nachmansohn, D. (1949) Science 110,
Augustinsson, K. B. (1971) Bull. World Health Org. 44,81-89 Silver, A. (1974) The Biology of Cholinesterases. Frontiers of
Biology (Neuberger, A., and Tatum, E. L., eds) Vol. 36, pp. 1- 50,411-448, Elsevier, New York
Gerebtzoff, M. A. (1959) Cholinesterases. A Histochemical Con- tribution to the Solution of Some Functional Problems, Per- gamon Press, London
168,223-231
98-99
Snell, R. S. (1959) Z. Zellforsch. Mikroskop. Anat. 49,330-334 Aoki, T. (1964) Nature 202, 1124-1126 Aoki, T. (1966) Nature 211,886-887 Hebb, C., and Hill, K. J. (1955) Quart. J. Exp. Physiol. 40, 168-
Nachmansohn, D. (1970) Science 168, 1059-1066 Lee, D. L. (1962) Parasitology 52, 241-260 Lui, A., Cork, D., and Krvavica, S. (1964) Vet. Arh. 34,84-86 Ogilvie, B. M., Rothwell, T. L. W., Bremner, K. C., Schnitzerling,
H. J., Nolan, J., and Keith, R. K. (1973) Znt. J. Parasitol. 3,
Knowles, C. O., and Casida, J. E. (1966) J. Agric. Food Chem. 14,
Hart, R. J., and Lee, R. M. (1966) Exp. Parasitol. 18,332-337 Lee, D. L. (1970) Tissue and Cell 2, 225-231 Jones, V. E., and Ogilvie, B. M. (1972) Zmmunology 22,119-129 Rothwell, T. L. W., Ogilvie, B. M., and Love, R. J . (1973) Znt. J .
Rothwell, T. L. W., and Merritt, G. C. (1974) Int. J. Parasitol. 4,
Bremner, K. C., Ogilvie, B. M., Keith, R. K., and Berrie, D. A.
Edwards, A. J., Burt, J. S., and Ogdvie, B. M. (1971) Parasitology
Sanderson, B. E., Jenkins, D. C., and Phillipson, R. F. (1972) Znt.
Sanderson, B. E. (1969) Comp. Biochem. Physiol. 29, 1207-1213 Sekine, T. (1951) J. Biochem. (Tokyo) 38, 171-179 Nelson, L. (1970) in Comparative Spermatology (Baccetti, B., ed)
Mann, T. (1964) The Biochemistry of Semen and of the Male
Hutchinson, G. W., and Probert, A. J . (1972) Exp. Parasitol. 32,
Nelson, L. (1972) Exp. Cell Res. 74, 269-274 Rhoads. M. L.. and Romanowski. R. D. (1974) Exp. Parasitol.
175
589-597
566-572
Parasitol. 3,599-608
63-71
(1973) Znt. J. Parasitol. 3,609-618
62,339-347
J. Parasitol. 2,227-232
pp. 465-474, Academic Press, New York
Reproductive Tract, p. 209, Methuen, London
109-116
35,363-368 ’
Burt, J. S., and Ogilvie, B. M. (1975) Exp. Parasitol. 38, 75-82 Hofstee, B. H. (1952) Science 116, 329-331 Reiner, E., Skrinjaric-Spoljar, M., Kralj, M., and Krvavica, S.
Stewart, T. A,, and Forrester, I. T. (1978) Biol. Reprod. 19, 271-
PavliE, M. (1967) Biochzm. Biophys. Acta 139, 133-137 Lee, R. M., and Hodsden, M. R. (1963) Biochem. Pharmacol. 12,
Hogarth-Scott, R. S., Watt, B. J., Ogilvie, B. M., and Rothwell, T. L. W. (1973) Znt. J. Parasitol. 3, 735-741
Yeates, R. A., and Ogilvie, B. M. (1976) in Biochemistry of Parasites and Host Parasite Relationships (Van den Bossche, H., ed) pp. 307-310, Elsevier/North Holland, Amsterdam
McLaren, D. J., Burt, J. S., and Ogilvie, B. M. (1974) Znt. J.
(1978) Comp. Biochem. Physiol. 60C, 155-157
279
1241-1252

A Secretory Cholinesterase from Stephanurus dentatus 9321
Parasitol. 4,39-46 39. Rothwell, T. L. W., Anderson, N., Bremner, K. C., Dash, K. M.,
LeJambre, L. F., Merritt, G. C., and Ng, B. K. Y. (1976) Vet. Parasitol. 1, 221-230
40. Michaeli, D., Pinto, J. D., Benjamini, E., and de Buren, F. P. (1969) Immunochemistry 6,101-109
41. Chadwick, L. E. (1963) in Cholinesterases and Anticholinesterase Agents (Koelle, G. B., ed) pp. 741-798, Springer, Berlin
42. Ogdvie, B. M., and Jones, V. E. (1971) Enp. Parasitol. 29, 138- 177
43. McLaren, D. J.(1976) in The Organization of Nematodes (Croll,
N. A., ed) pp. 139-161, Academic Press, New York 44. McLaren, D. J. (1972) Parasitology 65,507-524 45. Deleted in proof 46. Rhoads, M. L., Romanowski, R. D., Doherty, R. F., and Stewart,
K. K. (1978) J. Biol. Chem. 253, 1639-1642 47. Chase, P. S. (1972) Cell. Zmmunol. 5,544-554 48. Arora, P. K., Miller, H. C., and Aronson, L. D. (1978) Nature 274,
49. Kaliner, M., and Austen, K. F. (1974) Biochem. Pharrnacol. 23,
Additional references are found on p. 9323.
589-590
763-771
6 1 . 5 3 . 0
9322 A Secretory Cholinesterase from Stephanurus dentatus 'D1 I
A
INHIBIlOR CONC. (cM)
I / ';I ESERINE SULFATE
/ I -1.0 -A -.k -.i -3 6 .A .; .i .i IlO
I/lNHIBITO~ CONC. (#MI
B 6-
INMlBllOR CONC. (rM1
BW284CSI
, , I -.I -.6 -.4 -.I2 0 .h .I .6 .I 1.0
IIINHI~ITOR CONC. ( I "
7 1 "1
I/INHIBIlOR CONC. (MI

A Secretory Cholinesterase from Stephanurus dentatus 9323
A B
SO.
51.
52.
53.
5 4 .
5 5 . 56.
18. 57.
60.
59.
61.
62.
61.
64.
b5. bb.
67,
68.


















