
J Exp Med-2012-Joubert-1029-47
19
Article The Rockeeller University Press $30.00 J. Exp. Med. Vol. 209 No. 5 1029-1047 www.jem.org/cgi/doi/10.1084/jem.20110996 1029 Chikungunya virus (CHIKV) is the causative agent or Chikungunya ever, an arbovira dis- ease transmitted by mosquitoes. CHIKV was rst isoated in 1953 during an epidemic in Tanzania, East Arica (Mason and Haddow, 1957) and has recenty emerged in isands o the Indian Ocean in 2005 (Enserink, 2006). La Reunion, an isand in the Indian Ocean with a popuation o amost 785,000, was the most aected region, with an estimation o 300,000 cumuative cases in 2005–2006 (Schuenecker et a., 2006; Simon et a., 2006; Gérardin et a., 2008). The epidemic invoved India, where estimates approach six miion inected peope (Watson, 2007). It has aso emerged in Itay, southern France, and Austraia, and ongoing in- ections exist in Southeast Asia (Ng et a., 2009; Manimunda et a., 2011). CHIKV is a member o the Togaviridae amiy, genus Alphav irus , which are characterized by being enveoped, singe-stranded positive poarity RNA viruses (Inoue et a., 2003; Sourisseau et a., 2007; Schwartz and Abert, 2010). In human, CHIKV typicay induces symptoms 2–7 d ater inection and is characterized by a rapid onset o ever, severe arthragia, and myagia, oowed by constitutiona symptoms (headache, photopho- bia, nausea, and abdomina pain) and a rash (Bodenmann et a., 2007; Borgherini et a., 2007). In vivo, CHIKV mainy targets the con- nective tissue (i.e., primariy brobast ces), but inection has aso been reported in the iver, musce, and brain in human and mice (Ozden et a., 2007). In vitro, CHIKV is abe to inect a wide range o adherent ces, incuding CORRESPONDENCE Matthew Albert: albertm@pasteur.r Abbreviations used: BDI, bright detai intensity; CHIKV, Chikungunya virus; CPE, cytopathic eect; MOI, mutipicity o inection; PE, phosphatidyethanoamine; RNS, reactive nitrogen species; ROS, reactive oxygen species. Chikungunya virus–induced autophagy delays caspase-dependent cell death Pierre-Emmanuel Joubert, 1,5 Scott W. Werneke, 6 Claire de la Calle, 2 Florence Guivel-Benhassine, 4 Alessandra Giodini, 1,5 Lucie Peduto, 3 Beth Levine, 7,8 Olivier Schwartz, 4 Deborah J. Lenschow, 6 and Matthew L. Albert 1,2,5 1 Unité Immunobiologie des Cellules Dendritiques, 2 Centre d’Immunologie Humaine, and 3 Lymphoid Tissue Development Unit, Department o Immunology, and 4 Unité de recherche Virus et Immunité, Institut Pasteur, 75724 Paris, Cedex 15, France 5 INSERM U818, 75724 Paris, France 6 Department o Pathology and Immunology, Department o Medicine, Washington Univers ity School o Medicine, St. Louis, MO 63110 7 Department o Internal Medicine and 8 Howard Hughes Medical Institute, University o Texas S outhwestern Medical Center, Dallas, TX 75390 Autophagy is an important survival pathway and can participate in the host response to inec- tion. Studying Chikungunya virus (CHIKV), the causative agent o a major epidemic in India, Southeast Asia, and southern Europe, we reveal a novel mechanism by which autophagy limits cell death and mortality ater inection. We use biochemical studies and single cell multispectral assays to demonstrate that direct inection triggers both apoptosis and autophagy. CHIKV- induced autophagy is mediated by the independent induction o endoplasmic reticulum and oxidative stress pathways. These cellular responses delay apoptotic cell death by inducing the IRE1–XBP-1 pathway in conjunction with ROS-mediated mTOR inhibition. Silencing o autoph- agy genes resulted in enhanced intrinsic and extrinsic apoptosis, avoring viral propagation in cultured cells. Providing in vivo evidence or the relevance o our fndings, Atg16L HM mice, which display reduced levels o autophagy, exhibited increased lethality and showed a higher sensitivity to CHIKV-induced apoptosis. Based on kinetic studies and the observation that ea- tures o apoptosis and autophagy were mutually exclusive, we conclude that autophagy inhibits caspase-dependent cell death but is ultimately overwhelmed by viral replication. Our study suggests that inducers o autophagy may limit the pathogenesis o acute Chikungunya disease. © 2012 Joubert et al. This article is distributed under the terms o an Attribution– Noncommercial–Share Alike–No Mirror Sites license or the rst six months ater the publication date (see http://www.rupress.org/terms). Ater six months it is available under a Creative Commons License (Attribution–Noncommercial– Share Alike 3.0 Unported license, as described at http://creativecommons.org/ licenses/by-nc-sa/3.0/). T h e J o u r n a l o f E x p e r i m e n t a l M e d i c i n e o n M a y 2 8 , 2 0 1 2 j e m . r u p r e s s . o r g D o w n l o a d e d f r o m Published April 16, 2012
-
Upload
maitrayee-chakraborty -
Category
Documents
-
view
216 -
download
0
Transcript of J Exp Med-2012-Joubert-1029-47

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 1/19
Article
The Rockeeller University Press $30.00
J. Exp. Med. Vol. 209 No. 5 1029-1047
www.jem.org/cgi/doi/10.1084/jem.20110996
1029
Chikungunya virus (CHIKV) is the causative
agent or Chikungunya ever, an arbovira dis-
ease transmitted by mosquitoes. CHIKV was
rst isoated in 1953 during an epidemic in
Tanzania, East Arica (Mason and Haddow,
1957) and has recenty emerged in isands o
the Indian Ocean in 2005 (Enserink, 2006).
La Reunion, an isand in the Indian Ocean with
a popuation o amost 785,000, was the most
aected region, with an estimation o 300,000cumuative cases in 2005–2006 (Schuenecker
et a., 2006; Simon et a., 2006; Gérardin et a.,
2008). The epidemic invoved India, where
estimates approach six miion inected peope
(Watson, 2007). It has aso emerged in Itay,
southern France, and Austraia, and ongoing in-
ections exist in Southeast Asia (Ng et a., 2009;
Manimunda et a., 2011). CHIKV is a member
o the Togaviridae amiy, genus Alphavirus,
which are characterized by being enveoped,
singe-stranded positive poarity RNA viruses
(Inoue et a., 2003; Sourisseau et a., 2007;
Schwartz and Abert, 2010). In human, CHIKV
typicay induces symptoms 2–7 d ater inection
and is characterized by a rapid onset o ever,
severe arthragia, and myagia, oowed by
constitutiona symptoms (headache, photopho-
bia, nausea, and abdomina pain) and a rash
(Bodenmann et a., 2007; Borgherini et a.,
2007). In vivo, CHIKV mainy targets the con-nective tissue (i.e., primariy brobast ces),
but inection has aso been reported in the
iver, musce, and brain in human and mice
(Ozden et a., 2007). In vitro, CHIKV is abe to
inect a wide range o adherent ces, incuding
CORRESPONDENCEMatthew Albert:[email protected]
Abbreviations used: BDI,
bright detai intensity; CHIKV,
Chikungunya virus; CPE,
cytopathic eect; MOI,
mutipicity o inection; PE,
phosphatidyethanoamine;
RNS, reactive nitrogen species;
ROS, reactive oxygen species.
Chikungunya virus–induced autophagy
delays caspase-dependent cell death
Pierre-Emmanuel Joubert,1,5
Scott W. Werneke,6
Claire de la Calle,2
Florence Guivel-Benhassine,4 Alessandra Giodini,1,5 Lucie Peduto,3 Beth Levine,7,8 Olivier Schwartz,4 Deborah J. Lenschow,6 and Matthew L. Albert1,2,5
1Unité Immunobiologie des Cellules Dendritiques, 2Centre d’Immunologie Humaine, and 3Lymphoid Tissue Development Unit,Department o Immunology, and 4Unité de recherche Virus et Immunité, Institut Pasteur, 75724 Paris, Cedex 15, France
5INSERM U818, 75724 Paris, France6Department o Pathology and Immunology, Department o Medicine, Washington University School o Medicine, St. Louis, MO 631107Department o Internal Medicine and 8Howard Hughes Medical Institute, University o Texas Southwestern Medical Center,Dallas, TX 75390
Autophagy is an important survival pathway and can participate in the host response to inec-
tion. Studying Chikungunya virus (CHIKV), the causative agent o a major epidemic in India,Southeast Asia, and southern Europe, we reveal a novel mechanism by which autophagy limitscell death and mortality ater inection. We use biochemical studies and single cell multispectralassays to demonstrate that direct inection triggers both apoptosis and autophagy. CHIKV-
induced autophagy is mediated by the independent induction o endoplasmic reticulum andoxidative stress pathways. These cellular responses delay apoptotic cell death by inducing the
IRE1–XBP-1 pathway in conjunction with ROS-mediated mTOR inhibition. Silencing o autoph-agy genes resulted in enhanced intrinsic and extrinsic apoptosis, avoring viral propagation
in cultured cells. Providing in vivo evidence or the relevance o our fndings, Atg16L HM mice,which display reduced levels o autophagy, exhibited increased lethality and showed a highersensitivity to CHIKV-induced apoptosis. Based on kinetic studies and the observation that ea-
tures o apoptosis and autophagy were mutually exclusive, we conclude that autophagy inhibitscaspase-dependent cell death but is ultimately overwhelmed by viral replication. Our study
suggests that inducers o autophagy may limit the pathogenesis o acute Chikungunya disease.
© 2012 Joubert et al. This article is distributed under the terms o an Attribution–Noncommercial–Share Alike–No Mirror Sites license or the rst six monthsater the publication date (see http://www.rupress.org/terms). Ater six monthsit is available under a Creative Commons License (Attribution–Noncommercial–Share Alike 3.0 Unported license, as described at http://creativecommons.org/licenses/by-nc-sa/3.0/).
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 2/19
1030 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
autophagic processes to enhance their repication or transmis-
sion. Exampes incude Shigella exnerii and Listeria monocyto-
genes, which express viruence actors capabe o inhibiting
autophagosome ormation, herpes simpex 1, which expresses
ICP-34.5 or inhibition o initiation step in the autophagic
process, and inuenza A, which encodes M2 or inhibition o
autophagosome/ysosome usion (Deretic and Levine, 2009).
Some viruses manage to be even more subversive, using
mechanisms o autophagy or enhanced repication and vira
reease, incuding hepatitis C virus, poiovirus, and HIV-1
(Deretic and Levine, 2009; Banchet et a., 2010).
Recent data suggest cross tak between autophagic and
apoptotic pathways; or exampe, evidence supports inter-
actions between Bc-2 and Becin-1 (aso known as Atg6), and
both may be engaged ater induction o simiar ce stress path-
ways (e.g., ER or oxidative stress; Thorburn, 2008; Djavaheri-
Mergny et a., 2010; Kang et a., 2011). Herein, we characterized
the roe o apoptosis and autophagy in the context o CHIKV
pathogenesis and report that host mechanisms, incuding ER
and oxidative stress, imit the cytopathic eect (CPE) o
CHIKV through the induction o autophagy. Moreover, in
the absence o autophagic genes, we observe higher eves o
apoptosis, both in vitro and in vivo. With respect to the atter,
Atg16LHM mice were more sensitive to CHIKV-mediated
ethaity. These resuts dene autophagy as a host deense
mechanism that imits CHIKV pathogenesis.
RESULTSCHIKV inection induces autophagosome ormationin inected cellsDuring autophagy, Becin-1–PI3K-III compex activation
resuts in isoation membrane ormation that surrounds its
substrates to generate an autophagic vesice characterized bya doube membrane, which is caed an autophagosome
(Mehrpour et a., 2010). Two ubiquitin-ike systems have
been shown to be essentia or autophagosome ormation. In
the rst, Atg12 (autophagy-reated gene 12) is conjugated to
Atg5, together orming a compex with Atg16L1, which
decorates the outer membrane o the isoation membrane.
Microtubue-associated protein 1 LC3 (ight chain 3, aso
known as Atg8) constitutes the second ubiquitin-ike system:
LC3 conjugates phosphatidyethanoamine (PE) at the outer
and inner autophagosoma membrane. Unike the Atg12–
Atg5–Atg16L1 compex that is recyced, the LC3-PE
(reerred to as LC3-II) remains associated with the inner
membrane o autophagosome, making it a useu marker o autophagosomes (Kroemer et a., 2010; Mehrpour et a.,
2010). To dene the reationship between CHIKV inection
and autophagy, we rst examined autophagosome ormation.
Mouse embryonic brobasts (MEFs) were inected and LC3
puncta or LC3-II ormation was measured by immunouor-
escence or Western bot, respectivey. Compared with unin-
ected MEFs, CHIKV-inected ces showed increased
numbers o LC3 puncta (Fig. 1, A–C) and stronger staining
or LC3-II by Western botting (Fig. 1 D). Autophagosome
ormation coud be rst observed ater 5 h and peaked at 9 h
macrophages, brobasts, and HeLa ces (Sourisseau et a., 2007;
Schwartz and Abert, 2010). As observed or other aphaviruses
(e.g., SINV and Ross River), CHIKV inection has the capac-
ity to induce apoptosis, which has been suggested to be associ-
ated with pathogenesis. Notaby, apoptotic bebs have been
shown to contain CHIKV, which may represent a mechanism
o ce-to-ce vira spread (Krejbich-Trotot et a., 2011a).
Apoptosis (or programmed ce death type I) is an energy-
dependent process that is reguated by a preormed cascade
o proteases caed caspases (Dania and Korsmeyer, 2004;
Kurokawa and Kornbuth, 2009). Two main pathways are
invoved in apoptosis: (1) the intrinsic pathway, in which ac-
tivation o Bak and Bax resuts in mitochondria outer mem-
brane permeabiization inducing apoptosome ormation and
activation o caspase-9 (Kroemer et a., 2007); and (2) the ex-
trinsic pathway that is initiated by death receptor oigomer-
ization and ceavage o caspase-8 (or caspase-10; Wison
et a., 2009; Kurokawa and Kornbuth, 2010). Both pathways
resut in the activation o executioner caspases (e.g., caspase-3,
-6, and -7), eading to the morphoogica and biochemica mod-
ications recognized as apoptosis (Kurokawa and Kornbuth,
2009). Apoptosis is considered a rst ine o deense against
vira inection, where engagement o intraceuar stress path-
ways may trigger ce death, serving to imit vira repication
(Grifn and Hardwick, 1997; Everett and McFadden, 1999; Li
and Stoar, 2004). Many viruses have evoved strategies to
escape or deay apoptosis, urther pointing toward the roe o
ce death as an antivira mechanism (Teodoro and Branton,
1997; Kepp et a., 2009).
The autophagic pathway is a buk degradation system,
which contros the cearance and recycing o intraceuar
constituents or the maintenance o ceuar surviva (Deretic
and Levine, 2009). Autophagy consists o at east three path-ways: microautophagy, macroautophagy, and chaperone-
mediated autophagy (Crotzer and Bum, 2010; Li et a., 2011).
O those, macroautophagy is best characterized and has been
impicated in both innate and adaptive immunity (Deretic and
Levine, 2009). For the purpose o this study we wi reer to
macroautophagy as autophagy. Athough the autophagic path-
way was originay identied as a process induced by ceuar
starvation, there has been a strong interest in characterizing
the roe o autophagy as a mechanism o host deense (Deretic
and Levine, 2009; Crotzer and Bum, 2010; Joubert et a.,
2011). In vitro, autophagy is abe to isoate and degrade Strep-
tococcus pyogenes ater it enters in the cytoso (Nakagawa et a.,
2004; Joubert et a., 2009). Other in vitro and in vivo exam-pes incude the contro o Mycobacterium tuberculosis and Listeria
monocytogenes (Deretic and Levine, 2009). Some vira proteins
are aso targeted by autophagy (e.g., Sindbis or Tobacco
Mosaic virus; Deretic and Levine, 2009; Orvedah et a., 2010,
2011). Other roes or autophagy in the host response incudes
the enhancement o type I IFNs, or the processing and pre-
sentation o antigen or MHC I or MHC II presentation and
T ce priming (Dengje et a., 2005; Engish et a., 2009; Uh
et a., 2009; Crotzer and Bum, 2010). There aso exist exampes
o microbes that are capabe o abrogating and/or expoiting
Published April 16, 2012
7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 3/19
JEM Vol. 209, No. 5
Article
1031
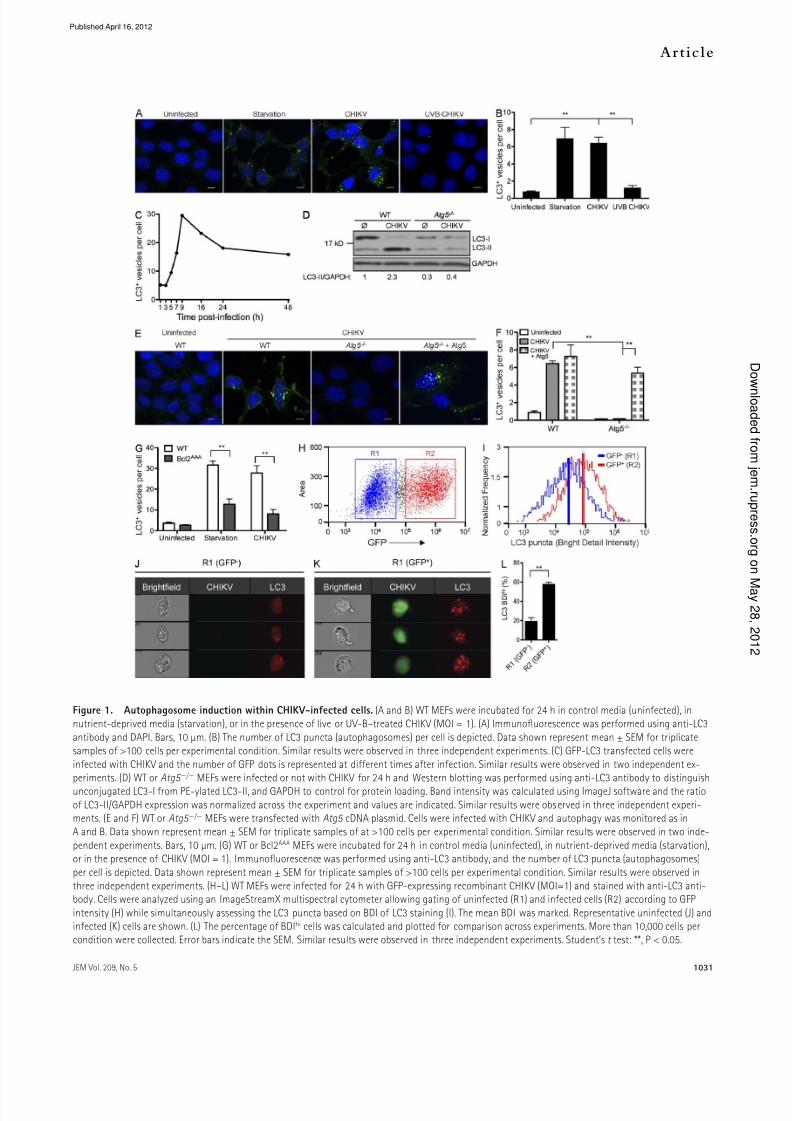
Figure 1. Autophagosome induction within CHIKV-inected cells. (A and B) WT MEFs were incubated or 24 h in control media (uninected), innutrient-deprived media (starvation), or in the presence o live or UV-B–treated CHIKV (MOI = 1). (A) Immunofuorescence was perormed using anti-LC3
antibody and DAPI. Bars, 10 µm. (B) The number o LC3 puncta (autophagosomes) per cell is depicted. Data shown represent mean ± SEM or triplicatesamples o >100 cells per experimental condition. Similar results were observed in three independent experiments. (C) GFP-LC3 transected cells were
inected with CHIKV and the number o GFP dots is represented at dierent times ater inection. Similar results were observed in two independent ex-periments. (D) WT or Atg5 / MEFs were inected or not with CHIKV or 24 h and Western blotting was perormed using anti-LC3 antibody to distinguishunconjugated LC3-I rom PE-ylated LC3-II, and GAPDH to control or protein loading. Band intensity was calculated using ImageJ sotware and the ratio
o LC3-II/GAPDH expression was normalized across the experiment and values are indicated. Similar results were observed in three independent experi-ments. (E and F) WT or Atg5 / MEFs were transected with Atg5 cDNA plasmid. Cells were inected with CHIKV and autophagy was monitored as inA and B. Data shown represent mean ± SEM or triplicate samples o at >100 cells per experimental condition. Similar results were observed in two inde-
pendent experiments. Bars, 10 µm. (G) WT or Bcl2AAA MEFs were incubated or 24 h in control media (uninected), in nutrient-deprived media (starvation),or in the presence o CHIKV (MOI = 1). Immunofuorescence was perormed using anti-LC3 antibody, and the number o LC3 puncta (autophagosomes)
per cell is depicted. Data shown represent mean ± SEM or triplicate samples o >100 cells per experimental condition. Similar results were observed inthree independent experiments. (H–L) WT MEFs were inected or 24 h with GFP-expressing recombinant CHIKV (MOI=1) and stained with anti-LC3 anti-body. Cells were analyzed using an ImageStreamX multispectral cytometer allowing gating o uninected (R1) and inected cells (R2) according to GFP
intensity (H) while simultaneously assessing the LC3 puncta based on BDI o LC3 staining (I). The mean BDI was marked. Representative uninected (J) andinected (K) cells are shown. (L) The percentage o BDIhi cells was calculated and plotted or comparison across experiments. More than 10,000 cells percondition were collected. Error bars indicate the SEM. Similar results were observed in three independent experiments. Student’s t test: **, P < 0.05.
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 4/19
1032 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
ater inection (Fig. 1 C), suggesting the requirement or
CHIKV repication as the trigger or autophagy induction.
Repication-deective CHIKV (achieved by UV-B irradia-
tion) aied to induce autophagosome ormation, thus sup-
porting the requirement or vira repication (Fig. 1, A and B).
Serum starvation served as a positive contro or autophagy
induction (Fig. 1, A and B; and not depicted).
To evauate i the induction o autophagosomes was oc-
curring via a cassica macroautophagy pathway, we evauated
CHIKV inection using MEFs decient in the key autophagy
gene Atg5 . As predicted, Atg5 / MEFs showed no evidence
o LC3-II conversion upon CHIKV inection (Fig. 1 D).
Simiary, immunouorescence studies demonstrated that
LC3 puncta observed during CHIKV inection were depen-
dent on Atg5 expression (Fig. 1, E and F). Restoration o
Atg5 expression, achieved by cDNA transection, rescued the
ces’ abiity to induce autophagy (Fig. 1, E and F). We ob-
served simiar resuts using ces decient or Atg7 , another
key autophagy gene (data not depicted). Furthermore, we
investigated the impication o Becin-1 in CHIKV-induced
autophagy using MEFs that express a mutant orm o Bc-2
(MEF-Bc2AAA). Notaby, Bc-2 directy reguates the acti-
vation o Becin-1 and aanine substitution o the three
phosphoryation sites (T69A/S70A/S87A) in Bc-2 prevents
dissociation o the Bc-2–Becin-1 compex, seectivey in-
hibiting autophagy induction by Becin-1 without aecting
the antiapoptotic roe o Bc-2 (Wei et a., 2008; He et a.,
2012). As expected, MEF-Bc2AAA ces were not abe to in-
duce autophagy under starvation conditions (Fig. 1 G). Simi-
ary, CHIKV-induced autophagy was abrogated in Bc-2AAA
MEFs, indicating that CHIKV-induced autophagy occurs via
a Becin-1–dependent mechanism. To conrm that CHIKV
inection induced autophagosome ormation in other cetypes, we aso investigated the appearance o autophagosomes
in HeLa ces and human oreskin brobasts. Siencing o
Atg5 or Atg7 genes using sma intererence RNA (siRNA)
conrmed data shown using MEFs (unpubished data).
To anayze whether autophagosome ormation was de-
pendent on direct vira inection, we marked active repication
using GFP-expressing recombinant CHIKV (Vanandingham
et a., 2005) and anayzed LC3 puncta using ImageStreamX.
In brie, mutispectra cytometric anaysis enabes the capture
o high-resoution images o ces in ow (up to 500 ces/s)
and permits anaysis o LC3 puncta (de a Cae et a., 2011).
24 h ater inection, GFP-expressing ces were gated (Fig. 1 H,
R2), and LC3 bright detai intensity (BDI) was integrated or each ce as a measure o autophagosome ormation. For
comparison, GFP-negative ces were gated (Fig. 1, H [R1]
and I [bue ine]), and histogram pots representing LC3
puncta indicate that CHIKV-inected ces (Fig. 1 I, red ine)
have higher LC3 BDI. Representative ImageStreamX images
with median intensity eves o BDI are shown or CHIKV-
inected and uninected ce popuations (Fig. 1, J and K),
conrming that LC3 puncta (scored based on high BDI) cor-
reated with the presence o both vira-encoded GFP and
robust autophagosome accumuation. Using this method, we
quantied the percentage o LC3-positive ces (BDIhi) when
buk cutures are segregated or CHIKV inection (Fig. 1 L,
R2 vs. R1, P < 0.05). Starved ces were used as positive
contro or autophagy induction (unpubished data). Based
on these data, we concude that autophagy induction occurs
via a Becin-1–dependent mechanism in a ce-intrinsic man-
ner; in other words, vira repication within the ce, as op-
posed to secreted actors produced by neighboring inected
ce, is the stimuus or autophagy induction.
Autophagosome/lysosome usion remains intactduring CHIKV inectionUpon maturation, autophagosomes use with ate endosomes
and ysosomes, which resuts in the ormation o a degradative
compartment reerred to as autoysosomes (Deretic and Levine,
2009). Some viruses encode inhibitors o this event (e.g., inu-
enza virus), and as a resut enhanced numbers o LC3 puncta
coud be a reection o basa autophagy accumuation and not
de novo autophagosome ormation (Gannagé et a., 2009). To
discriminate between these two possibiities, we anayzed auto-
phagy in the presence o ysosoma inhibitors eupeptin and
E64D in transected GFP-LC3 HeLa ces (Fig. 2, A and B). As
expected, inhibition o autophagosome/ysosome usion re-
suted in an increased number o LC3 puncta in contro (basa
autophagy ux) and starved (induced autophagy) HeLa ces.
The presence o eupeptin and E64D aso enhanced the num-
ber o autophagosomes in inected ces, aowing us to con-
cude that CHIKV inection induces de novo autophagosome
ormation (Fig. 2 B). To conrm this nding, we directy
assessed coocaization o LC3 and the ysosoma-associated
membrane protein LAMP-1 (Fig. 2 C). Optica sectioning o
ces was perormed and LC3+, LAMP-1+, and doube-abeed
vesices were enumerated on a per ce basis. Approximatey45% o the LC3+ puncta coocaized with LAMP-1+ ysosomes
in CHIKV-inected ces (Fig. 2 D). Line scan anaysis con-
rmed coocaization and indicated that the ysosoma struc-
tures were discrete vesices within the ce (Fig. 2 E).
As a na measure o autophagic ux, we transected MEF
ces with a construct encoding an RFP-GFP-LC3 poypro-
tein. In this way, it is possibe to distinguish autophagosomes
(RFP+ GFP+ puncta) and autoysosomes (RFP+ GFP vesi-
ces), with the atter being GFP negative as a resut o the
quenching o signa by the acidic microenvironment o the
ysosome. CHIKV inection increased the number o both
autophagosomes and autoysosomes compared with unin-
ected ces (Fig. 2, F and G). Moreover, an equivaent ratioo autophagosomes and autoysosomes was measured in in-
ected ces and starved ces (Fig. 2 G). Together, the data
presented in Figs. 1 and 2 provide evidence or CHIKV in-
ection inducing de novo autophagosome ormation without
inhibition o autophagosome maturation.
ER stress serves as a trigger or autophagyduring CHIKV inectionThe ER serves as an important sensor o ceuar stress. It de-
tects changes in ce homeostasis and responds by triggering
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 5/19
JEM Vol. 209, No. 5
Article
1033
Figure 2. CHIKV induces autophagic fux. (A and B) GFP-LC3–expressing HeLa cells were starved (starvation) or inected with CHIKV (MOI = 1) alone
or in the presence o leupeptin and E64D (L+E) or 24 h. The numbers o GFP+ vesicles per cell were enumerated. Data shown represent mean ± SEM ortriplicate samples o at least 100 cells per sample. Similar results were observed in our independent experiments. Bars, 5 µm. (C–E) WT MEFs were in-ected with CHIKV or 24 h. Immunofuorescence was perormed using anti-LC3, anti-Lamp-1, and DAPI to assess autophagosome and autolysosome
ormation. The number o LC3 puncta (green), Lamp-1+ vesicles (red), or costained vesicles (yellow) are depicted (C and D). Data represent mean ± SEM ortriplicate samples o at least 100 cells per sample. Similar results were observed in two independent experiments. Bars, 10 µm. (E) Line scale bar analysis
o a representative cell indicated in C is depicted. LC3 proles (green curve), Lamp-1 proles (red curve), and DAPI staining (blue curve) are shown. Theblack vertical line indicates colocalization o LC3 and Lamp-1 staining. (F and G) WT MEF cells were transected with a plasmid encoding a dual-labeledLC3 probe, RFP-GFP-LC3. Cells were again starved (starvation) or inected with CHIKV or 24 h. Representative images are shown and the number o total
autophagic vacuoles (GFP+RFP+ + GFP-RFP+), autophagosomes (GFP+RFP+), and autolysosomes (GFPRFP+) were enumerated. The inset permits visualiza-tion o yellow autophagosome (yellow arrow) and red autolysosomes (red arrow). Data represent mean ± SEM or three independents experiments o atleast 100 cells per sample. Similar results were observed in three independent experiments. Bars, 5 µm. Student’s test: **, P < 0.05.
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 6/19
1034 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
pathways reerred to as the unoded protein response (UPR;
Lee et a., 2003; McGuckin et a., 2010). Vira inection has
been shown to activate UPR as a resut o the accumuation
o vira proteins. At east three dierent pathways may be acti-
vated during ER stress, which are reguated by the signaing
moecues eIF2, IRE1, and ATF6, respectivey (McGuckin
et a., 2010). We screened a three pathways (unpubished
data) and identied a critica roe or IRE1. A roe or IRE1
activation during CHIKV inection was rst demonstrated
by anayzing the phosphoryation o IRE1 (p-IRE-1) at
Figure 3. CHIKV-induced autophagy is regu-lated by ER and oxidative stress. (A–C) WT MEFswere inected with CHIKV at indicated time pointsand Western blotting was perormed to detect phos-
phorylation o IRE1 (p-IRE1) and JNK (p-JNK) aswell as the ormation o spliced orm o XBP1 (XBP1s)
and the conjugation o LC3 (LC3-II). (A) IRE1 andGAPDH were also ollowed to control protein expres-sion and loading. Similar results were observed in two
independent experiments. (B) Immunofuorescencewas perormed using anti-pIRE1 and anti-E3 anti-body. Bars, 15 µm. (C) The number o cells positive or
pIRE1 in the E3+ (inected cells) and E3 (uninectedcells) populations is depicted. Data shown representmean ± SEM or triplicate samples o >100 cells per
experimental condition. Similar results were observedin three independent experiments. (D and E) WT MEFs
were pretreated with control siRNA or siRNA againstIRE1 or 3 d ollowed by inection with CHIKV or24 h at MOI 1. The number o LC3 punctas per cell
and the amount o LC3-II are depicted. Data shown
represent mean ± SEM or triplicate samples o >100cells per experimental condition. Similar results were
observed in three independent experiments. (F and G)WT MEFs were incubated or 24 h in control media (ø)
or in the presence o CHIKV (MOI = 1). (F) Immuno-fuorescence was perormed using an ROS/RNS de-tection kit that specically stains oxygen species and
ree NO. As positive controls or ROS or RNS induc-tion, WT MEFs were incubated or 5 h with pyco-
cyanin and l-arginine, respectively. Bars, 10 µm.(G) Percentage o cells containing ROS or NO amonginected with CHIKV and/or pretreated with specic
inhibitor o ROS and RNS as indicated is depicted.Data shown represent mean ± SEM or triplicate
samples o >100 cells per experimental condition.Similar results were observed in two independentexperiments. (H) WT MEFs or cells pretreated with
siRNA against IRE1 or 3 d were inected with CHIKV or 24 h in presence o a ROS inhibitor. The number o LC3 punctas per cell and the amount o LC3-II are
depicted. Data shown represent mean ± SEM ortriplicate samples o >100 cells per experimentalcondition. Similar results were observed in three inde-
pendent experiments. (I) WT MEFs were inected withCHIKV at indicated time points and Western blotting
was perormed to detect phosphorylation o mTOR(p-mTOR), S6KI (p-S6K1), and AMPK (p-AMPK). mTOR,S6K1, AMPK, and GAPDH were also ollowed to con-
trol protein expression and loading. As control to ROS
implication, similar experiments were perormed incells pretreated with ROS inducer and/or ROS inhibi-
tor. Black lines indicate that intervening lanes werespliced out. Similar results were observed in three
independent experiments. Student’s test: **, P < 0.05.
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 7/19
JEM Vol. 209, No. 5
Article
1035
RNS scavenger cross-inhibited ROS production, highighting
the interconnectivity between the ROS and RNS pathways.
This phenomenon coud be expained by NO reacting with
O2 to orm the oxidant peroxynitrite (ONOO-; Djavaheri-
Mergny et a., 2007; Novo and Paroa, 2008).
To conrm the impication o ROS/RNS production in
CHIKV-induced autophagy, we investigated LC3+ staining and
the conversion o LC3-I to LC3-II in inected MEFs pretreated
with N -acety-l-cysteine (Fig. 3 H). A signicant decrease in
LC3 puncta was observed, demonstrating the importance o
ROS production in CHIKV-induced autophagy. We urther
evauated potentia overap with the ER stress pathway by assess-
ing autophagy in ces sienced or IRE1 mRNA and treated
with N -acety-l-cysteine. Strikingy, we observed an additive
inhibitory eect that reduced the number o autophagosomes
per ce to near baseine eves (Fig. 3 H). This data suggests that
ER stress and oxidative stress act via independent mechanisms to
induce autophagy during CHIKV inection.
Athough oxidative stress is known to induce autophagy
upon microbia inection, the precise mechanism remains
poory documented. Based on recenty estabished inks be-
tween ROS and mTORC1 inhibition, which appears to be
dependent on TSC2 (tuberous scerosis compex 2), itse regu-
ated by the AMP-activated protein kinase (AMPK; Aexander
et a., 2010a,b), we investigate the reguation o this compex
during CHIKV inection. Importanty, phosphoryated mTOR
can be integrated into two dierent compexes, caed mTORC1
and mTORC2, depending on its interactions with Raptor and
Rictor, respectivey (Zoncu et a., 2011). Athough both com-
pexes are impicated in protein synthesis, ony mTORC1 is
inked to autophagy (Thomson et a., 2009). To discriminate
the activation o mTORC1 rom mTORC2, we anayzed
both mTOR phosphoryation and the induction o p-S6K1, aspecic substrate o mTORC1. As shown, we observed a
diminished eve o both p-mTOR and p-S6K1 24 h ater in-
ection (Fig. 3 I). This inhibition was transient, and 2–3 d ater
inection, a strong induction o the mTOR–S6K1 pathway
coud be detected. The kinetics o mTOR–S6K1 inhibition
correated with conversion o LC3-I to LC3-II, suggesting a
roe or mTORC1 as a mediator o CHIKV-induced auto-
phagy. To dene how CHIKV-induced ROS is capabe o in-
hibiting mTORC1, we next evauated the activation o AMPK.
Strikingy, the active orm o AMPK was detected 24 h ater
inection, coincident with the inhibition o mTORC1 (Fig. 3 I).
Moreover, the impication o the AMPK pathway in ROS-
mediated inhibition o mTORC1 coud be conrmed usingN -acety-l-cysteine (Fig. 3 I). Together, these data provide a
mechanistic understanding o CHIKV-induced autophagy.
Autophagy is a prosurvival mechanismthat limits CHIKV-induced cell deathIn addition to autophagy, other orms o ce stress may be
triggered as a resut o vira inection, incuding activation o
ce death pathways. Increasing evidence suggests that ce
stress pathways intersect and in some instances cross-inhibit
each other (Thorburn, 2008). As CHIKV inection triggers a
dierent time points ater inection. Western botting and
immunouorescence indicated that p-IRE1 was observed
during the eary phase o inection (Fig. 3, A and B) and coud
be detected ony in inected ces (as evauated based on E3
coocaization; Fig. 3, B and C). These data suggested that
CHIKV eads to an intrinsic activation o ER stress. Impor-
tanty, p-IRE1 was no onger detected 3 d ater inection.
This shutdown seems to be the resut o a decreased eve o
IRE1 protein, as indicated by Western bot anaysis (Fig. 3 A).
Remarkaby, the kinetics o IRE1 phosphoryation correated
with the conversion o LC3-I to LC3-II (Fig. 3 A).
To dene the moecuar events triggered by p-IRE1, we
investigated the activation o XBP1 and c-Jun amino-termina
kinases (JNK; McGuckin et a., 2010). Activation o XBP1 is
reguated by a dierentia spice variant o XBP1 mRNA,
which may be evauated based on the expression o a protein
o higher moecuar weight and is reerred to as XBP1s (or
spiced XBP1; Yoshida et a., 2001). This pathway has been
shown to avor ce surviva (McGuckin et a., 2010). In con-
trast, IRE1-induced phosphoryation o JNK is considered a
ink between ce stress and apoptosis (Urano et a., 2000).
During CHIKV inection, we observed an induced expres-
sion o XBP1s, but did not detect enhanced phosphoryation
o JNK (p-JNK; Fig. 3 A). To examine the unctiona ink
between IRE1 in CHIKV-induced autophagy, we sienced
expression o IRE1 using siRNA and anayzed CHIKV-
induced LC3 puncta as we as LC3-II conversion (Fig. 3,
D and E). Reduced IRE1 gene expression was conrmed by
Western bot, and shown to resut in ewer CHIKV-induced
autophagosomes. These data dene a roe or CHIKV activa-
tion o ER stress, which induces autophagy via an IRE1-
and XBP1s-mediated signaing pathway.
CHIKV-induced oxidative stress avors autophagosomeproduction through the inhibition o mTORC1Oxidative stress, primariy caused by increased eves o reac-
tive oxygen species (ROS) and reactive nitrogen species
(RNS), is a eature o the host response to vira inections
(Catadi, 2010). O2 and NO are considered to be the most
important mediators among ROS and RNS, respectivey. Free
oxidative agents are known to induce autophagy and can aso
ead to ce death during strong and proonged stimuation
(Djavaheri-Mergny et a., 2007; Fiomeni et a., 2010; Guo
et a., 2010). To assess the impact o ROS/RNS production in
CHIKV-induced autophagy, we rst investigated whether
CHIKV inection induces ROS and/or RNS production. Weinected WT MEF or 24 h and monitored the presence o
oxygen species and ree NO. As positive contro, we used pyco-
cyanin and l-arginine, inducers o ROS and NO, respectivey
(Fig. 3 F). As expected, pycocyanin increased the percentage o
ces that produced ROS, in the absence o NO production,
whereas l-arginine induced ree NO but not ROS. Interest-
ingy, we observed that CHIKV inection ed to increased pro-
duction o both ROS and NO (Fig. 3 F), which coud be
inhibited using the ROS inhibitor N -acety-l-cysteine or the
RNS scavenger c-PTIO (Fig. 3 G). O note, exposure to a
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 8/19
1036 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
was most prominent at ow mutipicity o inection
(MOI), suggesting a ink between ce death and
vira propagation throughout the cutured MEFs.
Simiar resuts were obtained by siencing expression
o autophagy genes in WT MEFs (Fig. 4, E and G).
To expore aternative ce death pathways, autoph-
agy genes were sienced in the Bax/ Bak/ MEFs.
Importanty, the absence o both autophagy andapoptosis pathways did not urther sensitize the ces
to aternative orms o ce death (Fig. 4 F). Finay,
we tested the importance o vira repication or ce
death induction. WT MEFs were inected with ive
or UVB-inactivated CHIKV, and ce death was
evauated. As shown, repication deective CHIKV
ais to induce ce death (Fig. 4 H).
To dene the orm o programmed ce death re-
sponsibe or the CPE in WT and Atg5 / MEFs, we
used pharmacoogica inhibitors o apoptosis (z-VAD,
a broad spectrum caspase inhibitor) or necroptosis
(necrostatin-1, an inhibitor o RIPK1). Inected ces
treated with necrostatin-1 exhibited a simiar eve o ce deathas compared with inected contro ces, whereas z-VAD rescued
both the WT and Atg5 / MEFs rom CHIKV-induced CPE.
(Fig. 4, I and J). These data, as we as the absence o CPE in
Bax/ Bak/ MEFs, demonstrate that the principe orm o
ce death induced by CHIKV is caspase-mediated apoptosis.
Autophagy delays both intrinsicand extrinsic apoptosis pathwaysTo dene the interaction between autophagy and apoptosis at
the singe ce eve, WT and Atg5 / or Bax/ Bak/ ces
Figure 4. Enhanced CHIKV-induced cell death in theabsence o autophagy. (A–D) WT, Bax / Bak / , or Atg5 / MEFs were inected with CHIKV at indicated doses and timepoints. Percentage o cell death was measured using a mem-
brane-permeable fuorescent probe and assessed by cytometry.Death curves over the 3 d ater inection are shown or a rep-
resentative experiment (A and C). The induction o cell deathwas also evaluated as a unction o the MOI (B and D). In allconditions ≥10,000 cells were acquired. Similar results were
observed in ve independent experiments. (E and F) WT orBax / Bak / MEFs were pretreated with the indicated siRNAor 3 d, ollowed by inection with CHIKV or 24 h at the indi-
cated doses. Cells death is analyzed as described or A–D.Similar results were observed in three independent experi-ments. (G) WT MEFs were treated with si-Atg5 or si-Atg7, and
Western blotting was perormed using anti-Atg5 or anti-Atg7antibodies. Similar results were observed in two independent
experiments. (H) WT MEFs were inected with CHIKV or UVB-treated CHIKV at indicated doses. Percentage o cell death wasmeasured using a membrane permeable fuorescent probe and
assessed by cytometry. Similar results were observed in two
independent experiments. (I and J) WT or Atg5 / MEFs werepretreated with z-VAD or necrostatin-1 (Nec-1) beore inec-tion by CHIKV. Percentage o cell death is depicted. Error barsindicate mean values ± SD rom three independent experi-
ments. Student’s test: **, P < 0.05.
pronounced CPE (Sourisseau et a., 2007), it was important toinvestigate the unction o autophagy on CHIKV-induced
ce death. Atg5 / MEFs and ces unabe to engage the
intrinsic apoptosis pathway (Bax/ Bak/ MEFs) were
inected with CHIKV and oss o membrane integrity was
anayzed. Whereas CHIKV inection triggered ce death in
WT ces in a time- and dose-dependant manner, Bax/
Bak/ MEFs remained reractory, showing ony minima ev-
idence o CPE at day 3 (Fig. 4, A and B). In contrast, Atg5 /
MEFs showed a dramatic increase in ce death compared with
its WT contro (Fig. 4, C and D). Notaby, the enhanced CPE
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 9/19
JEM Vol. 209, No. 5
Article
1037
Data are represented in Fig. 5 A with each dot indicating a
singe ce. Regions were estabished as detaied previousy
(de a Cae et a., 2011), and the percentage o autophagic
ces (LC3 BDIhi, ceaved caspase-3o; dened by R1), apoptotic
were inected with CHIKV. As previousy, LC3 BDI was
used as a measure or autophagy induction, and apoptosis ac-
tivity was characterized by abeing with antibodies specic or
the active, ceaved orm o caspase-3 (de a Cae et a., 2011).
Figure 5. Autophagy limits CHIKV-induced apoptosis. (A–D) WT, Bax / Bak / , or Atg5 / MEFs were inected with CHIKV (MOI = 1) or 24 h and
were stained or LC3 and activated caspase-3. (A) Representative ImageStreamX dot plots rom WT, Bax / Bak / , or Atg5 / inected MEFs are depictedand the gating strategy is indicated. (B–D) The relative percentage o autophagic cells (LC3 BDIhi, cleaved caspase-3lo; dened by R1), apoptotic cells (LC3
BDIlo, cleaved caspase-3hi; dened by R2), or cells with evidence or both processes (LC3 BDIhi, cleaved caspase-3hi; dened by R3) is shown. Error barsindicate mean ± SD o three independent experiments. (E and F) WT, Bax / Bak / , or Atg5 / MEFs were inected with CHIKV (MOI = 1) at the indicated
time points, and activated caspase-3 (a-CASP3), activated caspase-9 (a-CASP9), or activated caspase-8 (a-CASP8) were stained rom parallel cultures.Representative microscopic images are shown (E), and the percentage o positive cells were determined or >100 cells per condition (F). Error bars showmean ± SD o three independent experiments. Bars, 25 µm. (G and H) WT, Bax / Bak / , or Atg5 / MEFs were inected with GFP-expressing recombi-nant CHIKV (MOI = 1) and stained with anti–active capase-3, -8, and -9 antibody. Cells were gated as uninected (R1) and inected (R2) cells according to
GFP intensity (G) while simultaneously assessing the active capase-3, -8, and -9 staining (H). Similar results were observed in three independent experi-ments. (I) HFFs were inected with CHIKV (MOI = 1) or 24 h, and the percentage o activated caspase-3 cells was determined or >100 cells per condition.
Error bars show mean ± SD o three independent experiments.
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 10/19
1038 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
uninected ces. These data suggest that CHIKV inection
induces apoptosis through both intra- and extraceuar ac-
tors (Fig. 5 H) and that autophagy, acting in a ce-intrinsic
manner, preerentiay protects inected ces rom apoptosis.
CHIKV-induced apoptosis is independento ER and oxidative stressAs introduced in the previous sections, both ER and oxida-
tive stress may trigger proapoptotic pathways (McGuckin
et a., 2010). To determine the reative contribution o stress
pathway induction on autophagy versus apoptosis, we again
used Atg5 / MEFs. We rst conrmed the antiapoptotic
eect o autophagy by investigating the ceavage o pro–
caspase-3 in WT and Atg5 / MEFs (Fig. 6, A and B).
Whereas pro–capase-3 was ceaved ony ater 48 h o inec-
tion in WT ces, the active orm o caspase-3 was detected
24 h ater inection in Atg5 / MEFs. Moreover, the ratio o
active caspase-3/GAPDH was increased in Atg5 / ce dur-
ing a time points investigated (Fig. 6 B).
We next investigated the activation o IRE1 and inhibi-
tion o mTOR in Atg5 / MEFs. Interestingy, both path-
ways were simiary reguated by CHIKV inection as
compared with WT MEFs (Fig. 6 C compared with Fig. 3,
A and I). To ascertain the impact o these pathways on apop-
tosis, we evauated by immunouorescence the percentage o
active caspase-3–positive ces in WT or Atg5 / MEFs in
which ER and oxidative stress had been inhibited (Fig. 6 D).
IRE1 and/or ROS inhibition increased the percentage o
WT ces with active caspase-3. Importanty, no change in
active caspase-3 expression was observed in Atg5 / MEFs.
These data suggest that ER and oxidative stress occur beore
the roe o Atg5 in aciitating autophagy and are not critica
or the induction o CHIKV-induced apoptosis.Together, the data presented in Figs. 1–6 demonstrate
that during the eary phase o CHIKV inection, autophagy is
induced via the activation o ER stress and the inhibition o
mTOR by ROS production. By triggering autophagy,
CHIKV-induced ce stress indirecty imits apoptotic ce
death. Nevertheess, ater 48 h o inection, ER stress is
bunted, in part secondary to IRE1 degradation, and mTOR
becomes hyperphosphoryated. These events resut in de-
creased autophagic tone and, via a sti undened mechanism,
apoptosis pathways dominate, in turn resuting in pronounced
CHIKV-induced CPE.
Regulation o apoptosis and in vitro CHIKV propagationBoth autophagy and apoptosis have been reated to the regu-
ation o vira repication and/or propagation. Athough au-
tophagy can resut in the degradation o vira proteins (e.g.,
Sindbis) without signicanty aecting vira inection, it has
aso been reported to enhance vira repication (e.g., HCV;
Dreux and Chisari, 2009). Simiary, apoptosis has been im-
picated in both pro- and antivira responses (Li and Stoar,
2004). As both pathways are engaged by CHIKV inection, it
was important to evauate the eect o autophagy and apop-
tosis on vira propagation. To achieve this, we measured vira
ces (LC3 BDIo, ceaved caspase-3hi; dened by R2), or ces
with evidence or both processes (LC3 BDIhi, ceaved caspase-
3hi; dened by R3) were enumerated and represented graphi-
cay (Fig. 5, B–D). Supporting immunouorescence resuts
(Fig. 1), CHIKV inection triggered an increase in LC3
puncta in WT and Bax/ Bak/ MEFs, but not in Atg5 /
MEFs (Fig. 5 B). Strikingy, the number o ces exhibiting
active caspase-3 was increased upon CHIKV inection as
compared with uninected ces, shown at 0 h (Fig. 5 C). The
number o LC3+ ces was dramaticay reduced in WT ces 3
d ater inection. This observation correated with high eve
o apoptosis and suggests that autophagy and apoptosis are
mutuay excusive processes (Fig. 5, compare B with C). This
observation was urther supported by the absence o doube-
positive ces (region R3; Fig. 5 D).
Two important observations emerged rom the study o
the knockout MEFs. The rst intriguing nding concerned a
marked increase in active caspase-3+ ces when Atg5 is absent
(Fig. 5 C). These data suggested an important roe or
autophagy in the reguation o CHIKV-induced apoptosis.
The second discovery concerned the possibiity o the Bax/
Bak/ ces to activate caspase-3; the timing o apoptosis on-
set showed interexperimenta variation with ces showing
detectabe eves o ceaved caspase-3 between 48 and 72 h
(Fig. 5, C, E, and F). Evidence or caspase-3 activation in
Bax/ Bak/ ces indicated activation o the extrinsic
apoptosis pathway, which may be induced independenty o
mitochondria outer membrane permeabiization.
We then investigated whether autophagy is abe to regu-
ate apoptotic ce death in human brobasts ces (HFF),
which are known ce targets o CHIKV inection (Sourisseau
et a., 2007). We down-reguated the expression o Atg5 and
Atg7 genes in HFF by an siRNA strategy and anayzed theactivation o caspase-3 ater 24 h o inection with dierent
vira inputs (Fig. 5 I). Inhibition o both Atg5 and Atg7 dra-
maticay increased the percentage o ceaved caspase-3–positive
ces according to vira doses, demonstrating that the antiapop-
totic unction o autophagy is aso observed in human ces.
To distinguish the roe o autophagy in imiting the dis-
tinct apoptosis pathways, we evauated ceaved caspase-9
(a marker o the intrinsic pathway) and ceaved caspase-8
(indicative o activation o the extrinsic pathway) during
CHIKV inection (Fig. 5, E and F). Eary during the kinetics
o vira inection (16 h), caspase-9 activation was evident in
WT ces in the absence o detectabe eves o ceaved
caspase-8. In comparison, Atg5 / MEFs dispayed a twoodincrease in the percentage o active caspase-9–positive ces, as
we as eary evidence or ceaved caspase-8. By 40 h ater in-
ection, both the intrinsic and extrinsic pathways were engaged
in the WT ces and, again, Atg5 / ces demonstrated higher
eves o activation or both ce death pathways (Fig. 5 F).
Using GFP-expressing CHIKV, we next evauated the
reationship between inection and activation o caspase-3,
-9, and -8 (Fig. 5, G and H). Whereas the intrinsic apoptotic
pathway was engaged primariy in CHIKV+ ces, the extrin-
sic pathway was detectabe in both inected and bystander
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 11/19
JEM Vol. 209, No. 5
Article
1039
decient ces expressed simiar expression intensity to
CHIKV E3 proteins as compared with its WT contro, indi-
cating that autophagy had minima impact on viray encoded
GFP expression within the ce (Fig. 7 D). These data were
urther conrmed in a secondary cuture–based assay; ce
supernatants rom the respective ce ines were exposed to
uninected WT MEFs and GFP expression was scored ater
an additiona 24-h incubation, indicating higher eves o in-
ectious virus in the absence o autophagic ux and ower
eves in apoptosis-decient ces (unpubished data). To-
gether, these data suggested that autophagy coud reguate
vira propagation by imiting the reease o virus induced by
apoptotic ce death.
To support this observation, we estabished an image
anaysis script using the ImageStreamX, which integrated ce
area (unpubished data), thereby permitting quantication o
ces in the na stages o apoptosis based on their being sma
and pyknotic. Importanty, anaysis o the pyknotic ces indi-
cated that 80% expressed vira capsid protein as compared
with arger, ess dead ces, o which 45% stained positive
or E3 (Fig. 7, E and F), suggesting that ingestion o apoptotic
bodies coud contribute to inection o phagocytic neighbor-
ing ces (Krejbich-Trotot et a., 2011a). Apoptotic body ac-
cumuation upon CHIKV inection was next determined in
WT or Atg5 / MEFs (Fig. 7 G, R1). We aso conrmed
that accumuation o sma area events was dependent on an
apoptotic process, as use o z-VAD eiminated this popuation
oad in the supernatant o WT, Atg5 / , or Bax/ Bak/
MEFs. CHIKV titers seemed enhanced in supernatant o
Atg5 / ces, suggesting that autophagy coud restrict vira
reease (Fig. 7 A). However, modest vira titers were ower in
Bax/ Bak/ MEFs as compared with WT contros at day 1
ater inection, thereby suggesting a roe or apoptotic ce
death in vira reease during eary phase inection (Fig. 7 B).
Simiar resuts were obtained in HeLa ces using siRNA spe-
cic or Atg5 or Atg7 (unpubished data).
To urther anayze the unction o autophagy and apop-
tosis in vira inection, we assessed the percentage o inected
ces ater 24 h o inection by using GFP-expressing recom-
binant CHIKV as a marker o active inection (Fig. 7 C). At
ow vira dose (MOI = 0.1), Atg5 / MEFs showed greater
inection than WT contros. This dierence is a resut o en-
hanced inection and not dierentia vira entry, as suggested
by a kinetic study o inected ces (data not depicted). Resuts
or Bax/ Bak/ MEFs were even more striking as ony a
minority o ces were GFP+ ces, supporting that apoptotic
ce death inuences CHIKV propagation (Fig. 7 C). Cyto-
metric assessment o the Bax/ Bak/ MEFs indicated that
athough ewer ces were inected, those ces that were in-
ected showed higher expression o CHIKV E3 proteins as
compared with its WT contro (Fig. 7 D). This suggested that
a deay in ce death aowed or greater per ce vira repica-
tion, but aiure to undergo rapid ce death resuted in ewer
inected ces at the popuation eve. In contrast, autophagy-
Figure 6. ER and oxidative stress don’t enhance in CHIKV-induced apoptosis. (A and B) WT or Atg5/ MEFs were inected with CHIKV at theindicated time points. (A) Western blotting was perormed using anti-LC3 and anti–caspase-3 (Casp3) to discriminate pro–caspase-3 and cleavedcaspase-3, and GAPDH to control or protein loading. Black lines indicate that intervening lanes were spliced out. (B) Band intensity was calculated using
ImageJ sotware and the ratio o pro–caspase-3/GAPDH and cleaved-caspase-3/GAPDH was represented by graphs. Similar results were observed inthree independent experiments. (C) Atg5/ MEFs were inected with CHIKV at indicated time points and Western blotting was perormed to detectedphosphorylation o IRE1 (p-IRE1) and mTOR (p-mTOR). IRE1, mTOR, and GAPDH were ollowed to control protein expression and loading. Similar
results were observed in two independent experiments. (D) WT and Atg5/ MEFs were pretreated or not with siRNA against IRE1 or 3 d and were theninected by CHIKV or 48 h in control media or in the presence o ROS inhibitor. Activated caspase-3 was stained and the percentage o positive cells was
determined. Error bars show mean ± SD o three independent experiments. Student’s test: **, P < 0.05.
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 12/19
1040 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
Figure 7. Regulation o apoptotic cell death limits CHIKV propagation in vitro. (A and B) WT, Bax / Bak / , or Atg5 / MEFs were inected withCHIKV at indicated MOI and extracellular viral titers were determined during the 3 d ater inection. Results were expressed as TCID50/ml. Error bars indi-
cate mean viral titer ± SD o our independent experiments. (C) WT, Bax / Bak / , or Atg5 / MEFs were inected with GFP-expressing recombinantCHIKV (MOI = 0.1). ImageStreamX dot plots based on GFP intensity are shown. Similar results were observed in three independent experiments. (D) WT,
Atg5 / , or Bax / Bak / MEFs were inected at MOI = 1 and viral proteins were stained using anti-E3 Ab and analyzed by cytometry. Similar results were
observed in three independent experiments. (E and F) WT MEFs were inected with GFP-CHIKV (MOI = 0.1) or 48 h and arealo events were gated (R1 in G).
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 13/19

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 14/19
1042 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
propagation. In vivo data in Atg16LHM mice support an anti-
apoptotic eect or autophagy and indicate that in the ab-
sence o autophagic genes, there is an increased susceptibiity
to severe orms o Chikungunya disease.The roe or autophagy in host deense has been docu-
mented or bacteria and severa viruses (Deretic and Levine,
2009); however, the resuts rom our current study identiy a
previousy uncharacterized roe or autophagy as a mecha-
nism o imiting disease pathogenesis. Importanty, we dem-
onstrate that autophagy is triggered in a ce-intrinsic manner
by direct CHIKV inection, which eads to the induction o
both ER and oxidative stress (Figs. 1 and 3). Specicay,
among the ER stress pathways that are induced during vira
inection, ony IRE1/XBP1s is invoved in autophagic
DISCUSSIONThe recent CHIKV outbreak has exposed how itte we un-
derstand about the pathogenesis o this virus or the abiity to
harness host responses to enhance the contro o arbovirainection. Immunoogica studies have suggested that eary
events o vira–host ce interactions determine whether
CHIKV achieves disseminated inection, or i repication is
imited and controed by innate deense mechanisms. In our
prior studies, we evauated the roe o pattern recognition re-
ceptors and type I IFNs as mediators o vira contro (Schite
et a., 2010). Here, we examined an aternative stress response
pathway that is activated upon inection, making the exciting
discovery that engagement o the autophagy pathway imits
vira-induced ce death as a mechanism or sowing vira
Figure 8. Autophagy limits apoptotic induction in CHIKV inected tissues and delays lethality o mice. (A–F) WT (n = 43) and Atg16LHM mice(n = 34) were inected at 9 d o age with 4 × 105 PFU CHIKV subcutaneously. (A) Mice were monitored or lethality or 21 d with data displayed asKaplan-Meier curves. (B) Skin, muscle, and serum were collected ater days 1 (n = 3), 2 (n = 3), 5 (n = 4), and 9 (n = 5) o inection, homogenized, and viral
titers were determined by standard plaque assay. Median values or WT (black bars) or Atg16LHM (blue bars) mice are depicted. (C) Muscle, skin, liver, brain,BM, and spleen were collected rom WT inected mice at day 5. Tissues were xed in PFA, rozen in OCT blocks, and stained or cleaved capsase-3 (green)and E3 (red). Similar results were observed in two independent experiments. Bars: (muscle) 50 µm; (other tissues) 20 µm. (D–F) Muscle rom inected WT
or Atg16LHM mice was collected at days 1, 2, and 5 ater inection and stained or cleaved caspase-3, E3, or DAPI or nucleus staining (D). The percentageo cleaved capsase-3–positive cells was numerated in all population (E) or in E3+ and E3 cells (F). Error bars mean ± SD o three independent experi-
ments. Bars, 60 µm. Student’s test: **, P < 0.05.
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 15/19
JEM Vol. 209, No. 5
Article
1043
pathogens (Deretic and Levine, 2009). Physica degradation
o the microbe or microbia components may be achieved
through their isoation in autophagic vesices that in turn use
with ysosomes, which is reerred to as xenophagy (Orvedah
and Levine, 2008). A seective mechanism that eads to their
capture is reguated, in part, by the poy-ubiquitin binding
adaptor proteins p62 and NDP52. Components o the SinV
are aso sequestered by autophagy in a p62-dependant man-
ner (Orvedah et a., 2010). In vivo, genetic inactivation o
Atg5 in viray inected mouse neurons resuted in deayed
cearance o SinV capsid, the accumuation o p62 aggregates,
increased neurona apoptosis, and rapid mortaity, a in the
apparent absence diering vira oad.
A second mechanism by which autophagy may partici-
pate in the host response to inection reates to the cross tak
between autophagy genes and pattern recognition receptors.
Lee et a. (2007) demonstrated in a semina study that
autophagy sequesters cytosoic vira genome and/or host
RNA during VSV inection. This aowed igand to access
the umina domain o TLR7 in pDCs and enhanced produc-
tion o IFN-/ and other proinammatory cytokines. A-
though this pathway is unikey to contribute to the contro
o CHIKV during inection o brobast or epitheia ces, as
a resut o their ack o TLR7 expression, it is possibe that
autophagy my impact RIG-I signaing, athough as discussed
previousy, the intersection between Atg5 and RIG-I resuts
in a bunting o the antivira response (Jounai et a., 2007).
However, we did not observe atered IFN- production in
ces acking Atg5 or atered IFN- induction in mice hypo-
morphic or Atg16L (unpubished data), suggesting that this
mechanism o reguation did not account or autophagy-
mediated suppression o CHIKV repication in brobasts
and epitheia ces.Our study provides mechanistic inormation or another
strategy by which autophagy participates in the host response
to imit ce mortaity ater vira inection, thus controing
the reease o inectious vira partices secondary to apoptotic
ce death. We demonstrate that autophagy promotes ce sur-
viva by imiting apoptotic ce death. Importanty, autophagy
induction is dependent on CHIKV repication; consequenty,
the deay in apoptosis occurs in a ce-intrinsic manner. In
other words, neighboring uninected ces remain sensitive
to putative extraceuar inducers o the extrinsic pathway
(Fig. 5 H). The mechanism accounting or autophagy induc-
tion is inked to the activation o host stress responses during
acute inection, arguing that autophagy is induced by ces toreguate these stress responses. Regarding the oxidative stress,
autophagy has been described to restrict ROS production,
or exampe by eiminating damaged mitochondria. Indeed,
this is occurring in the context o CHIKV inection, as indi-
cated by their being higher eves o ROS production in
inected Atg5 / ces (unpubished data). Nonetheess, au-
tophagy did not succeed to competey contro the high eves
o oxidative stress induced by vira inection and repication.
For both ER and oxidative stress, it has been suggested
that their induction may resut in autophagy and apoptosis,
reguation (Fig. 3, A–D; and not depicted). Oxygen species
produced during CHIKV inection are abe to independenty
enhance autophagy via the inhibition o mTORC1 (Fig. 3,
E–I). The autophagy pathway induced is dependent on
Becin-1 and proceeds unimpeded, with utimate usion be-
tween autophagosomes and degradative ysosomes (Figs. 1 G
and 2). Using nove, singe ce assays that permitted simuta-
neous assessment o autophagy and apoptosis, we were abe
to demonstrate that at a time points studied, the two bio-
ogica processes are mutuay excusive (Fig. 5, A–D). That
said, kinetic studies, as we as assessment o vira protein ex-
pression in dying ces, suggest that as vira inection pro-
gresses, the autophagy pathway is overwhemed and ces
began to undergo apoptosis (Fig. 5).
Role o apoptosis in acute viral inectionApoptosis ater vira inection is considered to be an important
mechanism o host deense, as autodestruction may imit vira
repication (Everett and McFadden, 1999). RNA viruses re-
sponsibe or acute inection may have a distinct agenda, and
growing evidence suggests that a subset o agents may have
evoved to activey trigger programmed ce death (Yatim and
Abert, 2011). Members o the aphavirus amiy have been
shown to activey induce programmed ce death. For exam-
pe, Sindbis virus (SinV) may trigger both intrinsic and extrin-
sic apoptosis (Li and Stoar, 2004). Athough transection o
SinV nsp2 or the E1, pE2, and E3 structura proteins may
trigger ce death, the precise mechanism remains unknown
(Zhang et a., 1997; Li and Stoar, 2004). Mathematica mod-
es argued that a virus wi evove toward increasing cytopath-
icity when the mean ietime o a ce is high and the rate o
vira budding is ow (Krakauer and Payne, 1997). Indeed,
CHIKV inects primariy brobasts and other ong-ived stro-ma ces (Powers and Logue, 2007). In accordance with this
prediction, it was recenty demonstrated by Krejbich-Trotot
et a. (2011a) that apoptotic ce death is required or efcient
CHIKV propagation. Our ndings support this observation,
as mutispectra anaysis o CHIKV-inected ces indicated the
eary engagement o the intrinsic ce death pathway (Fig. 5 F).
Simiar to SinV, CHIKV inection aso resuted in caspase-8
activation (Fig. 5 F), suggesting a possibe roe or TNFSF re-
ceptor engagement (e.g., TRAIL-R). Interestingy, in con-
trast to caspase-9 activation, there was evidence or uninected
ces with active caspase-8 (Fig. 5 H).
Using z-VAD or gene siencing o caspase-3, we urther
showed that inhibition o apoptosis signicanty inhibitedCHIKV propagation (Fig. 7, B, C, and I; and not depicted).
Simiar resuts were observed in ces decient or Bax/
Bak/ , abeit with a ess dramatic eect as a resut o the act
that the extrinsic pathway remains intact in these ces (Fig. 7).
Thus, we suggest that both the intrinsic and extrinsic path-
ways participate in enhancing CHIKV propagation.
Autophagy as an antiviral mechanismThe autophagy pathway and/or autophagy proteins have been
recenty shown to participate in the host response to intraceuar
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 16/19
1044 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
or deaying apoptosis and protecting the host rom acute
aphavira inections. Integration o these data in the context
o ER and oxidative stress may revea nove mechanisms by
which viruses moduate host pathways. Our data characteriz-
ing ate phases o CHIKV inection (e.g., 48–72 h) identied
mTOR overexpression and IRE1 degradation as possibe
cues or how these stress pathways may be perturbed.
In sum, we have provided evidence or a nove mecha-
nism by which autophagy participates in host deense. In the
context o acute inection by CHIKV, apoptotic ce death is
important or vira propagation, and autophagy, in a ce-
intrinsic manner, protects ces rom apoptosis and may imit
vira reease. These data suggest that inducers o autophagy
may be useu therapeutics or imiting the pathogenesis o
acute Chikungunya disease or protecting neonates rom
mother-to-chid transmission.
MATERIALS AND METHODS
Cells and mice. WT and Bax/ Bak/ MEF ces rom the Korsmeyer
aboratory (Dana-Farber Cancer Institute, Boston, MA) were derived romC57BL/6 mice (H-2b). Atg5 / ces rom the Kroemer Laboratory
(NSERM U848, Institut Gustave Roussy, Viejui, France) were derived
rom C57BL/6 mice. GFP-LC3-HeLa ces were obtained ater transection
with peGFP-LC3 pasmid (peGFP-LC3, Addgene; rom the Yoshimori
aboratory, Okazaki, Japan) using Lipoectamine 2000 reagent (Invitrogen).
A stabe GFP-LC3-HeLa cone was estabished by imited diution in
0.5 mM geneticin-seective medium. A ce ines were maintained at 37°C
in humidied atmosphere containing 5% CO2 in medium suppemented
with 10% heat inactivated eta ca serum, 100 µg/m peniciin (Invitrogen),
100 U/m streptomycin (Invitrogen), and 1× MEM nonessentia amino acid
(Invitrogen). HFFs were obtained rom American Type Cuture Coection.
WT mice were obtained rom Chares River or The Jackson Laboratory.
Atg16LHM mice on the C57BL/6 background, uy backcrossed by speed
congenics, were generated by H. Virgin (Washington University Schoo o
Medicine, St. Louis, MO) and have been previousy described (Cadwe
et a., 2008). These mice were received into an enhanced barrier aciity with
speciaized eatures as previousy described (Cadwe et a., 2010). Mouse
studies at the Institut Pasteur and at Washington University were perormed
in strict accordance with the recommendations in the Guide or the Care
and Use o Laboratory Animas o the Nationa Institutes o Heath and
according to the Internationa Guiding Principes or Biomedica Research
Invoving Animas. The protocos were approved by the Anima Studies
Committee at Washington University (#20090287) and the Institutiona
Committees on Anima Weare o the Institut Pasteur (OLAW assurance
# A5476-01). A eorts were made to minimize suering.
siRNA treatment. SMARTpoo siRNA targeting Atg5 , Atg7 , or caspase-3
and contro siRNA were obtained rom Thermo Fisher Scientic. 0.106 HeLa
or MEF ces were cutured in 6-we pates or 1 d in OptiMEM (Invitrogen)
containing 10% FCS and transected with 30 nM o indicated siRNA using
Lipoectamine RNAiMAX (Invitrogen). In some experiments, Atg5 or Atg7 cDNA was transected ater 2 d o siRNA treatment. For a experiments,
CHIKV inection was perormed ater 3 d o siRNA incubation.
CHIKV inection and UV inactivation. The preparation o CHIKV
rom cinica sampes has been previousy described (Schuenecker et a.,
2006). CHIKV-21 strain was propagated in C6/36 ces and supernatants
were harvested and rozen at 80°C beore titration and urther use. MEF or
HeLa ces (pate at50% conuence in 24- or 48-wes pates) were exposed
to the indicated viruses or 1–2 h at 37°C, extensivey washed with PBS, and
cutivated or various periods o time beore urther anaysis. The MOI was
dened as the amount o CHIKV inectious units (cacuated on BHK ces as
PFU) per target ce. 3CHIKV-GFP was generated using a u-ength inection
depending on the signa intensity or duration o the stress in-
ducer. In the case o IRE1, we demonstrate that during the
eary phase o CHIKV inection, IRE1 avors XBP1 mRNA
spicing and autophagy, yet does not ead to proapoptotic
JNK phosphoryation. Future studies wi be required to as-
certain i this signaing event is dictated by viray encoded
proteins or a reection o the concentration o unoded pro-
teins during CHIKV repication. It wi aso be vauabe to
estabish a better understanding o how autophagy is termi-
nated, with an absoute switch to an apoptotic phenotype as
suggested by our singe ce anaysis (Fig. 5 A).
Cross talk between autophagy and apoptosisOur study provides an interesting ink between autophagy,
apoptosis, and vira propagation, aso highighting the active
investigation into cross tak between ce stress and ce death.
Severa mechanisms may account or autophagy imiting
apoptotic ce death. For exampe, degradation o protein ag-
gregates and damaged mitochondria that resut rom vira in-
ection may deay induction o apoptosis (Kroemer et a.,
2010). Additionay, it has been suggested that reease o bc-2
and FLIP rom Atg3 protein compexes may activey inhibit
the intrinsic and extrinsic pathways o apoptosis (Thorburn,
2008; Kroemer et a., 2010). Conversey, apoptosis may in-
hibit autophagy as capain has been shown to ceave Atg5 and
caspase-3 ceaves Atg4D, an enzyme impicated in the deipi-
dation o Atg protein (Kang et a., 2011). Many moecuar
detais must sti be dened, yet the avaiabe resuts have ed
to the concept that autophagy and apoptosis are antagonistic
events that are cross inhibitory. That said, prior studies evau-
ating these antagonistic mechanisms used buk assays such as
Western botting and thus coud not discern activation o
both pathways in a singe ce.One o the unique aspects presented here is the use o mu-
tispectra images cytometer, which permitted unbiased me-
dium throughput and simutaneous assessment o vira protein
expression, LC3 puncta, and caspase activation at the singe
ce eve. Consistent with the prevaiing dogma, but dierent
rom other vira inections such as inuenza A (de a Cae
et a., 2011), we demonstrate the absence o ces showing
eatures o both autophagy and apoptosis. Even in situations
where both pathways are active in the buk popuation o
ces, the two pathways were mutuay excusive. This resut
suggests that vira inection may trigger either autophagy or
apoptosis and that the moecuar decision is a proxima event.
One obvious candidate is Becin-1 (Atg6), which interactswith bc-2, reguating both pathways. In resting ces, Becin-
1–bc-2 compexes imit autophagy without atering the anti-
apoptotic eects o bc-2. Nutrient starvation, ER stress, or
oxidative damage may disrupt these interactions, avoring the
association o Becin-1 with PI3K-III and the induction o
autophagy (Kang et a., 2011). I, however, apoptosis pre-
cedes autophagy, caspase ceavage o Becin-1 wi prevent its
induction (Kang et a., 2011). Interestingy, Becin-1 has
been shown to imit SinV propagation (Liang et a., 1998),
suggesting a more genera roe or autophagy as a mechanism
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 17/19
JEM Vol. 209, No. 5
Article
1045
with Cytox/Cytoperm beore abeing with anti-capsid (rom the Schwartz
aboratory). To detect inected ces, MEFs were inected with CHIKV-GFP
and GFP intensity was anayzed according to ImageStreamX procedure.
In vivo inection, titration, and staining. WT ittermate contros were
used or a experiments (C57BL/6). 9-d-od C57BL/6 WT (n = 43) and
Atg16LHM mice (n = 34) were inected with 4 × 105 PFU CHIKV s.c. in the
upper chest and were oowed or ethaity or 25 d ater inection. Litters
were weight matched at the start o the experiments. For vira titration
(n = 3), tissues were coected, homogenized, and vira sampes were titrated
by standard paque assay on BHK ces. For histoogy bocks, skin (n = 3) and
musce (n = 3) rom the site o inection was xed in PFA and rozen in OCT
bocks. Tissues were sectioned and stained or activated caspase-3 and CHIKV
E3 protein and image anaysis was perormed as previousy described.
The authors would like to acknowledge Herbert Virgin III or providing Atg16LHM
mice, and the Centre d’Immunologie Humaine at Institut Pasteur or support
o experimental study. We also thank F. Rey, M. Lecuit, and members o the
Immunobiologie des Cellules Dendritiques laboratory or their helpul advice and
review o the paper.
This work was supported by the Institut Pasteur, Institut National de la Santé et
de la Recherche Médicale, Agence Nationale de Recherches sur le Sida et les hépatites
virales, the European Research Council, and the French National Research Agency (ANR).None o the authors have competing nancial interests.
Submitted: 17 May 2011
Accepted: 19 March 2012
REFERENCESAexander, A., S.L. Cai, J. Kim, A. Nanez, M. Sahin, K.H. MacLean,
K. Inoki, K.L. Guan, J. Shen, M.D. Person, et a. 2010a. ATM sig-
nas to TSC2 in the cytopasm to reguate mTORC1 in response to
ROS. Proc. Natl. Acad. Sci. USA. 107:4153–4158. http://dx.doi.org/
10.1073/pnas.0913860107
Aexander, A., J. Kim, and C.L. Waker. 2010b. ATM engages the TSC2/
mTORC1 signaing node to reguate autophagy. Autophagy. 6:672–673.
http://dx.doi.org/10.4161/auto.6.5.12509
Banchet, F.P., A. Moris, D.S. Nikoic, M. Lehmann, S. Cardinaud, R.
Stader, E. Garcia, C. Dinkins, F. Leuba, L. Wu, et a. 2010. Humanimmunodeciency virus-1 inhibition o immunoamphisomes in den-
dritic ces impairs eary innate and adaptive immune responses. Immunity.
32:654–669. http://dx.doi.org/10.1016/j.immuni.2010.04.011
Bodenmann, P., F. Athaus, B. Burnand, P. Vaucher, A. Pécoud, and B.
Genton. 2007. Medica care o asyum seekers: a descriptive study o
the appropriateness o nurse practitioners’ care compared to traditiona
physician-based care in a gatekeeping system. BMC Public Health. 7:310.
http://dx.doi.org/10.1186/1471-2458-7-310
Borgherini, G., P. Poubeau, F. Staikowsky, M. Lory, N. Le Mouec, J.P.
Becquart, C. Wenging, A. Michaut, and F. Paganin. 2007. Outbreak
o chikungunya on Reunion Isand: eary cinica and aboratory eatures
in 157 adut patients. Clin. Inect. Dis. 44:1401–1407. http://dx.doi.org/
10.1086/517537
Cadwe, K., J.Y. Liu, S.L. Brown, H. Miyoshi, J. Loh, J.K. Lennerz, C.
Kishi, W. Kc, J.A. Carrero, S. Hunt, et a. 2008. A key roe or autophagy
and the autophagy gene Atg161 in mouse and human intestina Panethces. Nature . 456:259–263. http://dx.doi.org/10.1038/nature07416
Cadwe, K., K.K. Pate, N.S. Maoney, T.C. Liu, A.C. Ng, C.E. Storer,
R.D. Head, R. Xavier, T.S. Stappenbeck, and H.W. Virgin. 2010.
Virus-pus-susceptibiity gene interaction determines Crohn’s disease
gene Atg16L1 phenotypes in intestine. Cell . 141:1135–1145. http://
dx.doi.org/10.1016/j.ce.2010.05.009
Catadi, A. 2010. Ce responses to oxidative stressors. Curr. Pharm. Des.
16:1387–1395. http://dx.doi.org/10.2174/138161210791033969
Couderc, T., F. Chrétien, C. Schite, O. Disson, M. Brigitte, F. Guive-
Benhassine, Y. Touret, G. Barau, N. Cayet, I. Schuenecker, et a.
2008. A mouse mode or Chikungunya: young age and inefcient type-
I intereron signaing are risk actors or severe disease. PLoS Pathog.
4:e29. http://dx.doi.org/10.1371/journa.ppat.0040029
cDNA cone provided by S. Higgs (Vanandingham et a., 2005). Inectious
virus was obtained by transection o BHK21 ces with RNA-derived cDNA,
as described. UV inactivation o the virus was perormed as described pre-
viousy (Jan and Grifn, 1999; Krejbich-Trotot et a., 2011a) by exposure o
the vira suspension to UV ight (2 J over 10 min at room temperature) rom
UV transiuminator (Spectra; The Daavin Company).
Immunofuorescent study. MEFs were xed with 4% paraormadehyde
(PFA) or 20 min and stained or LC3 (anti-LC3 Ab, cone 4E12; MBL),
activated caspase-3 (anticeaved caspase-3; Ce Signaing Technoogy), acti-
vated caspase-8 (anticeaved caspase-8; Ce Signaing Technoogy), activated
caspase-9 (anticeaved caspase-9; Ce Signaing Technoogy), p-IRE1 (rab-
bit poycona; Abcam), or Lamp-1 (anti–Lamp-1; Ce Signaing Techno-
ogy) according to the experiments. The nuceus was stained with 300 nM
DAPI (Invitrogen) or 5 min at 37°C in cuture media. For some experi-
ments, ces were treated with 1 µg/m eupeptin hemisuate (Invitrogen)
and 1 µg/m E64D (Invitrogen) 15 min beore CHIKV inection or starva-
tion. Ces were observed using an AxioCam MRm version 3 (Car Zeiss)
with a magnication o 63 (oi immersion) and anayzed using AxioVision
sotware (Car Zeiss). The number o vesices per ce was anayzed rom one
singe optica section per ce using the ApoTome (Car Zeiss) and numer-
ated using ImageJ sotware (Nationa Institutes o Heath). The RFP-GFP-LC3
construct (ptLC3, Addgene, rom the Yoshimori aboratory) was transienty
expressed in HeLa ces 48 h beore starvation or CHIKV inection. For a
experiments, the numbers o vesices were determined orm at east 100 ces
per sampes.
Western blot analysis. Lysates were prepared in 1× Dubecco’s Phosphate
Buer Saine (DPBS; Invitrogen) containing 1% Nonidet P 40 substitute
(NP40; Sigma-Adrich) and protease inhibitor cocktai (Roche). Tota protein
was determined by Lowry’s method and 25 mg was oaded on a 4–12% gradi-
ent SDS–PAGE (Invitrogen). Proteins were transerred to PVDF membrane
and botted with anti-LC3 (mouse monocona; Ce Signaing Technoogy),
anti-Atg5 (mouse monocona; Ce Signaing Technoogy), anti-ATG7
(mouse monocona; Ce Signaing Technoogy), anti-IRE1 (rabbit poy-
cona; Abcam), anti-pIRE1 (rabbit poycona; Abcam), anti-pJNK (rabbit
poycona; Abcam), anti-XBP1 (rabbit poycona; Abcam), anti-pmTOR
(rabbit poycona; Abcam), anti-mTOR (rabbit poycona; Abcam), anti-pS6K1 (rabbit poycona; Abcam), anti-pAMPK (rabbit poycona; Abcam),
or anti-GAPDH (rabbit poycona; Ce Signaing Technoogy). Secondary
HRP-couped Abs were detected using ECL Pus (GE Heathcare).
Cell viability assays and fow cytometry. MEFs or HeLA ces were in-
ected with CHIKV at the indicated time and xed with 4% PFA or 20 min.
Ater xation, ces were stained with ive/dead xabe vioet ce stain kit
(405 nm excitation; Invitrogen) and the percentage o ce death was measured
by ow cytometry using FACSCanto (BD) and FowJo sotware (Tree Star).
For detection o intraceuar CHIKV proteins, ces were permeabiized with
Cytox/Cytoperm (BD) beore abeing with anti-E3 (rom the Lecuit abora-
tory, Microorganismes et barrières de ’hôte, Institue Pasteur, France) or
anti-capsid (rom the Schwartz aboratory, viruses et immunité, URA 3015
Centre Nationa de a Recherche Scientique, Institue Pasteur, France).
CHIKV titers in cell lines. MEFs or HeLa ces were inected with
CHIKV-21 and supernatants were recovered at indicated time points. Vira
sampes were titrated as TCID50 endpoint on Vero ces using a standard
procedure. Seria 10-od diutions (100 µg/iter) o supernatants were added
in six repicates in 96-we pates seeded with 104 Vero ces. The CPE was
scored 5 d ater inection and the titers were cacuated by determining the
ast diution, giving 50% o wes with ces dispaying a CPE. Resuts were
expressed as TCID50/m.
ImageStreamX procedure. Pease reer to de a Cae et a. (2011) or detais
on the methods o sampe preparation, ImageStreamX acquisition, and anay-
sis. For detection o intraceuar CHIKV proteins, ces were permeabiized
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 18/19
1046 Autophagy limits CHIKV-induced apoptosis | Joubert et al.
Jan, J.T., and D.E. Grifn. 1999. Induction o apoptosis by Sindbis virus
occurs at ce entry and does not require virus repication. J. Virol.
73:10296–10302.
Joubert, P.E., G. Meiren, I.P. Grégoire, G. Pontini, C. Richetta, M.
Facher, O. Azocar, P.O. Vidaain, M. Vida, V. Lotteau, et a. 2009.
Autophagy induction by the pathogen receptor CD46. Cell Host Microbe .
6:354–366. http://dx.doi.org/10.1016/j.chom.2009.09.006
Joubert, P.E., I. Pombo Grégoire, G. Meiren, C. Rabourdin-Combe, and M.Faure. 2011. [Autophagy and pathogens: “Bon appétit Messieurs!”].Med.
Sci. (Paris). 27:41–47. http://dx.doi.org/10.1051/medsci/201127141
Jounai, N., F. Takeshita, K. Kobiyama, A. Sawano, A. Miyawaki, K.Q. Xin,
K.J. Ishii, T. Kawai, S. Akira, K. Suzuki, and K. Okuda. 2007. The Atg5
Atg12 conjugate associates with innate antivira immune responses. Proc.
Natl. Acad. Sci. USA. 104:14050–14055. http://dx.doi.org/10.1073/
pnas.0704014104
Kang, R., H.J. Zeh, M.T. Lotze, and D. Tang. 2011. The Becin 1 net-
work reguates autophagy and apoptosis. Cell Death Difer. 18:571–580.
http://dx.doi.org/10.1038/cdd.2010.191
Kepp, O., L. Senovia, L. Gauzzi, T. Panaretakis, A. Tesniere, F.
Schemmer, F. Madeo, L. Zitvoge, and G. Kroemer. 2009. Vira
subversion o immunogenic ce death. Cell Cycle . 8:860–869. http://
dx.doi.org/10.4161/cc.8.6.7939
Krakauer, D.C., and R.J. Payne. 1997. The evoution o virus-induced apoptosis.
Proc. Biol. Sci. 264:1757–1762.http://dx.doi.org/10.1098/rspb.1997.0243Krejbich-Trotot, P., M. Denizot, J.J. Hoarau, M.C. Jaar-Bandjee, T. Das,
and P. Gasque. 2011a. Chikungunya virus mobiizes the apoptotic
machinery to invade host ce deenses. FASEB J. 25:314–325. http://
dx.doi.org/10.1096/j.10-164178
Krejbich-Trotot, P., B. Gay, G. Li-Pat-Yuen, J.J. Hoarau, M.C. Jaar-
Bandjee, L. Briant, P. Gasque, and M. Denizot. 2011b. Chikungunya
triggers an autophagic process which promotes vira repication. Virol. J.
8:432. http://dx.doi.org/10.1186/1743-422X-8-432
Kroemer, G., L. Gauzzi, and C. Brenner. 2007. Mitochondria membrane
permeabiization in ce death. Physiol. Rev. 87:99–163. http://dx.doi
.org/10.1152/physrev.00013.2006
Kroemer, G., G. Mariño, and B. Levine. 2010. Autophagy and the integrated
stress response. Mol. Cell . 40:280–293. http://dx.doi.org/10.1016/j
.moce.2010.09.023
Kurokawa, M., and S. Kornbuth. 2009. Caspases and kinases in a death grip.
Cell . 138:838–854. http://dx.doi.org/10.1016/j.ce.2009.08.021
Kurokawa, M., and S. Kornbuth. 2010. Staing in mitosis and reeas-
ing the apoptotic brake. EMBO J. 29:2255–2257. http://dx.doi.org/
10.1038/emboj.2010.150
Lee, A.H., N.N. Iwakoshi, and L.H. Gimcher. 2003. XBP-1 reguates a sub-
set o endopasmic reticuum resident chaperone genes in the unoded
protein response. Mol. Cell. Biol. 23:7448–7459. http://dx.doi.org/10
.1128/MCB.23.21.7448-7459.2003
Lee, H.K., J.M. Lund, B. Ramanathan, N. Mizushima, and A. Iwasaki. 2007.
Autophagy-dependent vira recognition by pasmacytoid dendritic ces.
Science . 315:1398–1401. http://dx.doi.org/10.1126/science.1136880
Li, M.L., and V. Stoar. 2004. Aphaviruses and apoptosis. Int. Rev. Immunol.
23:7–24. http://dx.doi.org/10.1080/08830180490265529
Li, W., Q. Yang, and Z. Mao. 2011. Chaperone-mediated autophagy:
machinery, reguation and bioogica consequences. Cell. Mol. Lie Sci.
68:749–763. http://dx.doi.org/10.1007/s00018-010-0565-6
Liang, X.H., L.K. Keeman, H.H. Jiang, G. Gordon, J.E. Godman, G.
Berry, B. Herman, and B. Levine. 1998. Protection against ata Sindbis
virus encephaitis by becin, a nove Bc-2-interacting protein. J. Virol.
72:8586–8596.
Manimunda, S.P., D. Mavaankar, T. Bandyopadhyay, and A.P. Sugunan.
2011. Chikungunya epidemic-reated mortaity. Epidemiol. Inect.
139:1410–1412. http://dx.doi.org/10.1017/S0950268810002542
Mason, P.J., and A.J. Haddow. 1957. An epidemic o virus disease in Southern
Province, Tanganyika Territory, in 1952-53; an additiona note on Chikun-
gunya virus isoations and serum antibodies. Trans. R. Soc. Trop. Med. Hyg.
51:238–240. http://dx.doi.org/10.1016/0035-9203(57)90022-6
McGuckin, M.A., R.D. Eri, I. Das, R. Lourie, and T.H. Forin. 2010.
ER stress and the unoded protein response in intestina inammation.
Crotzer, V.L., and J.S. Bum. 2010. Autophagy and adaptive immunity.
Immunology. 131:9–17. http://dx.doi.org/10.1111/j.1365-2567.2010
.03321.x
Dania, N.N., and S.J. Korsmeyer. 2004. Ce death: critica contro points.
Cell . 116:205–219. http://dx.doi.org/10.1016/S0092-8674(04)00046-7
de a Cae, C., P.E. Joubert, H.K. Law, M. Hasan, and M.L. Abert. 2011.
Simutaneous assessment o autophagy and apoptosis using mutispec-
tra imaging cytometry. Autophagy. 7:1045–1051. http://dx.doi.org/ 10.4161/auto.7.9.16252
Dengje, J., O. Schoor, R. Fischer, M. Reich, M. Kraus, M. Müer, K.
Kreymborg, F. Atenberend, J. Brandenburg, H. Kabacher, et a. 2005.
Autophagy promotes MHC cass II presentation o peptides rom intra-
ceuar source proteins. Proc. Natl. Acad. Sci. USA. 102:7922–7927.
http://dx.doi.org/10.1073/pnas.0501190102
Deretic, V., and B. Levine. 2009. Autophagy, immunity, and microbia
adaptations. Cell Host Microbe . 5:527–549. http://dx.doi.org/10.1016/
j.chom.2009.05.016
Djavaheri-Mergny, M., M. Ameotti, J. Mathieu, F. Besançon, C. Bauvy,
and P. Codogno. 2007. Reguation o autophagy by NFkappaB tran-
scription actor and reactives oxygen species. Autophagy. 3:390–392.
Djavaheri-Mergny, M., M.C. Maiuri, and G. Kroemer. 2010. Cross tak
between apoptosis and autophagy by caspase-mediated ceavage o
Becin 1. Oncogene . 29:1717–1719. http://dx.doi.org/10.1038/onc
.2009.519Dreux, M., and F.V. Chisari. 2009. Autophagy proteins promote hepati-
tis C virus repication. Autophagy. 5:1224–1225. http://dx.doi.org/10
.4161/auto.5.8.10219
Engish, L., M. Chemai, J. Duron, C. Rondeau, A. Lapante, D. Gingras,
D. Aexander, D. Leib, C. Norbury, R. Lippé, and M. Desjardins. 2009.
Autophagy enhances the presentation o endogenous vira antigens on
MHC cass I moecues during HSV-1 inection. Nat. Immunol. 10:480–
487. http://dx.doi.org/10.1038/ni.1720
Enserink, M. 2006. Inectious diseases. Massive outbreak draws resh atten-
tion to itte-known virus. Science . 311:1085. http://dx.doi.org/10.1126/
science.311.5764.1085a
Everett, H., and G. McFadden. 1999. Apoptosis: an innate immune response
to virus inection. Trends Microbiol. 7:160–165. http://dx.doi.org/
10.1016/S0966-842X(99)01487-0
Fiomeni, G., E. Desideri, S. Cardaci, G. Rotiio, and M.R. Cirioo. 2010.
Under the ROS…thio network is the principa suspect or autophagy
commitment. Autophagy. 6:999–1005.http://dx.doi.org/10.4161/auto.6
.7.12754
Gannagé, M., D. Dormann, R. Abrecht, J. Dengje, T. Torossi, P.C.
Rämer, M. Lee, T. Strowig, F. Arrey, G. Coneneo, et a. 2009.
Matrix protein 2 o inuenza A virus bocks autophagosome usion
with ysosomes. Cell Host Microbe . 6:367–380. http://dx.doi.org/10
.1016/j.chom.2009.09.005
Gérardin, P., V. Guernier, J. Perrau, A. Fianu, K. Le Roux, P. Grivard, A.
Michaut, X. de Lambaerie, A. Fahaut, and F. Favier. 2008. Estimating
Chikungunya prevaence in La Réunion Isand outbreak by serosurveys:
two methods or two critica times o the epidemic. BMC Inect. Dis.
8:99. http://dx.doi.org/10.1186/1471-2334-8-99
Grifn, D.E., and J.M. Hardwick. 1997. Reguators o apoptosis on the road
to persistent aphavirus inection. Annu. Rev. Microbiol. 51:565–592.
http://dx.doi.org/10.1146/annurev.micro.51.1.565
Guo, W.J., S.S. Ye, N. Cao, J. Huang, J. Gao, and Q.Y. Chen. 2010. ROS-
mediated autophagy was invoved in cancer ce death induced by nove
copper(II) compex. Exp. Toxicol. Pathol. 62:577–582. http://dx.doi.org/
10.1016/j.etp.2009.08.001
He, C., M.C. Bassik, V. Moresi, K. Sun, Y. Wei, Z. Zou, Z. An, J. Loh,
J. Fisher, Q. Sun, et a. 2012. Exercise-induced BCL2-reguated
autophagy is required or musce gucose homeostasis. Nature . 481:511–
515. http://dx.doi.org/10.1038/nature10758
Inoue, S., K. Morita, R.R. Matias, J.V. Tupano, R.R. Resueo, J.R.
Candeario, D.J. Cruz, C.A. Mapua, F. Hasebe, A. Igarashi, and F.F.
Natividad. 2003. Distribution o three arbovirus antibodies among mon-
keys (Macaca ascicuaris) in the Phiippines. J. Med. Primatol. 32:89–94.
http://dx.doi.org/10.1034/j.1600-0684.2003.00015.x
Published April 16, 2012

7/31/2019 J Exp Med-2012-Joubert-1029-47
http://slidepdf.com/reader/full/j-exp-med-2012-joubert-1029-47 19/19
Article
Am. J. Physiol. Gastrointest. Liver Physiol. 298:G820–G832. http://
dx.doi.org/10.1152/ajpgi.00063.2010
Mehrpour, M., A. Escatine, I. Beau, and P. Codogno. 2010. Overview o
macroautophagy reguation in mammaian ces. Cell Res. 20:748–762.
http://dx.doi.org/10.1038/cr.2010.82
Nakagawa, I., A. Amano, N. Mizushima, A. Yamamoto, H. Yamaguchi,
T. Kamimoto, A. Nara, J. Funao, M. Nakata, K. Tsuda, et a. 2004.
Autophagy deends ces against invading group A Streptococcus. Science .306:1037–1040. http://dx.doi.org/10.1126/science.1103966
Ng, L.F., A. Chow, Y.J. Sun, D.J. Kwek, P.L. Lim, F. Dimatatac, L.C.
Ng, E.E. Ooi, K.H. Choo, Z. Her, et a. 2009. IL-1beta, IL-6, and
RANTES as biomarkers o Chikungunya severity. PLoS ONE . 4:e4261.
http://dx.doi.org/10.1371/journa.pone.0004261
Novo, E., and M. Paroa. 2008. Redox mechanisms in hepatic chronic
wound heaing and brogenesis. Fibrogenesis Tissue Repair . 1:5. http://
dx.doi.org/10.1186/1755-1536-1-5
Orvedah, A., and B. Levine. 2008. Vira evasion o autophagy. Autophagy.
4:280–285.
Orvedah, A., S. MacPherson, R. Sumpter Jr., Z. Taóczy, Z. Zou, and
B. Levine. 2010. Autophagy protects against Sindbis virus inection o
the centra nervous system. Cell Host Microbe . 7:115–127. http://dx.doi
.org/10.1016/j.chom.2010.01.007
Orvedah, A., R. Sumpter Jr., G. Xiao, A. Ng, Z. Zou, Y. Tang, M.
Narimatsu, C. Gipin, Q. Sun, M. Roth, et a. 2011. Image-basedgenome-wide siRNA screen identies seective autophagy actors.
Nature . 480:113–117. http://dx.doi.org/10.1038/nature10546
Ozden, S., M. Huerre, J.P. Riviere, L.L. Coey, P.V. Aonso, V. Mouy,
J. de Monredon, J.C. Roger, M. E Amrani, J.L. Yvin, et a. 2007.
Human musce sateite ces as targets o Chikungunya virus inection.
PLoS ONE . 2:e527. http://dx.doi.org/10.1371/journa.pone.0000527
Powers, A.M., and C.H. Logue. 2007. Changing patterns o chikungunya
virus: re-emergence o a zoonotic arbovirus. J. Gen. Virol. 88:2363–
2377. http://dx.doi.org/10.1099/vir.0.82858-0
Schite, C., T. Couderc, F. Chretien, M. Sourisseau, N. Gangneux, F.
Guive-Benhassine, A. Kraxner, J. Tschopp, S. Higgs, A. Michaut,
et a. 2010. Type I IFN contros chikungunya virus via its action on
nonhematopoietic ces. J. Exp. Med. 207:429–442. http://dx.doi.org/
10.1084/jem.20090851
Schuenecker, I., I. Iteman, A. Michaut, S. Murri, L. Frangeu, M.C. Vaney,
R. Lavenir, N. Pardigon, J.M. Reynes, F. Pettinei, et a. 2006. Genomemicroevoution o chikungunya viruses causing the Indian Ocean outbreak.
PLoS Med. 3:e263.http://dx.doi.org/10.1371/journa.pmed.0030263
Schwartz, O., and M.L. Abert. 2010. Bioogy and pathogenesis o chi-
kungunya virus. Nat. Rev. Microbiol. 8:491–500. http://dx.doi.org/10
.1038/nrmicro2368
Simon, F., H. Toou, and P. Jeande. 2006. [The unexpected Chikungunya
outbreak].Rev. Med. Interne . 27:437–441 (The unexpected Chikungunya
outbreak). http://dx.doi.org/10.1016/j.revmed.2006.03.028
Sourisseau, M., C. Schite, N. Casartei, C. Trouiet, F. Guive-Benhassine,
D. Rudnicka, N. So-Fouon, K. Le Roux, M.C. Prevost, H. Fsihi,
et a. 2007. Characterization o reemerging chikungunya virus. PLoS
Pathog. 3:e89. http://dx.doi.org/10.1371/journa.ppat.0030089
Teodoro, J.G., and P.E. Branton. 1997. Reguation o apoptosis by vira
gene products. J. Virol. 71:1739–1746.
Thomson, A.W., H.R. Turnquist, and G. Raimondi. 2009. Immunoreguatory
unctions o mTOR inhibition. Nat. Rev. Immunol. 9:324–337. http://dx.doi.org/10.1038/nri2546
Thorburn, A. 2008. Apoptosis and autophagy: reguatory connections be-
tween two supposedy dierent processes. Apoptosis. 13:1–9. http://
dx.doi.org/10.1007/s10495-007-0154-9
Uh, M., O. Kepp, H. Jusorgues-Sakani, J.M. Vicencio, G. Kroemer,
and M.L. Abert. 2009. Autophagy within the antigen donor ce
aciitates efcient antigen cross-priming o virus-specic CD8+
T ces.Cell Death Difer.16:991–1005.http://dx.doi.org/10.1038/cdd
.2009.8
Urano, F., X. Wang, A. Bertootti, Y. Zhang, P. Chung, H.P. Harding, and
D. Ron. 2000. Couping o stress in the ER to activation o JNK pro-
tein kinases by transmembrane protein kinase IRE1. Science . 287:664–
666. http://dx.doi.org/10.1126/science.287.5453.664
Vanandingham, D.L., K. Tsetsarkin, C. Hong, K. Kinger, K.L. McEroy,
M.J. Lehane, and S. Higgs. 2005. Deveopment and characterization
o a doube subgenomic chikungunya virus inectious cone to expressheteroogous genes in Aedes aegypti mosquitoes. Insect Biochem. Mol. Biol.
35:1162–1170. http://dx.doi.org/10.1016/j.ibmb.2005.05.008
Watson, R. 2007. Europe witnesses rst oca transmission o chikungunya
ever in Itay. BMJ . 335:532–533. http://dx.doi.org/10.1136/bmj
.39332.708738.DB
Wei, Y., S. Pattingre, S. Sinha, M. Bassik, and B. Levine. 2008. JNK1-mediated
phosphoryation o Bc-2 reguates starvation-induced autophagy. Mol.
Cell . 30:678–688. http://dx.doi.org/10.1016/j.moce.2008.06.001
Wison, N.S., V. Dixit, and A. Ashkenazi. 2009. Death receptor signa
transducers: nodes o coordination in immune signaing networks. Nat.
Immunol. 10:348–355. http://dx.doi.org/10.1038/ni.1714
Yatim, N., and M.L. Abert. 2011. Dying to repicate: the orchestration
o the vira ie cyce, ce death pathways, and immunity. Immunity.
35:478–490. http://dx.doi.org/10.1016/j.immuni.2011.10.010
Yoshida, H., T. Matsui, A. Yamamoto, T. Okada, and K. Mori. 2001. XBP1
mRNA is induced by ATF6 and spiced by IRE1 in response to ERstress to produce a highy active transcription actor. Cell . 107:881–891.
http://dx.doi.org/10.1016/S0092-8674(01)00611-0
Zhang, X., J. Qu, M. Ding, X. Ren, and Y. Qiu. 1997. [The mutipication
o Sindbis virus and host BHK-21 ce apoptosis]. Wei Sheng Wu Xue
Bao. 37:165–170.
Zoncu, R., A. Eeyan, and D.M. Sabatini. 2011. mTOR: rom growth sig-
na integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol.
12:21–35. http://dx.doi.org/10.1038/nrm3025
Published April 16, 2012


















