
Understanding intestinal lipopolysaccharide permeability ...

Isolation and structural characterization of an R-formlipopolysaccharide from Yersinia enterocolitica serotype O:8
Clemens Oertelt1, Buko Lindner2, Mikael Skurnik3 and Otto Holst1
1Division of Analytical Biochemistry, and 2Division of Biophysics, Center for Medicine and Biosciences, Borstel, Germany;3Department of Medical Biochemistry, Institute of Biomedicine, University of Turku, Finland
The lipopolysaccharide (LPS) of strain 8081-c-R2, a
spontaneous R-mutant of Yersinia enterocolitica serotype
O:8, was isolated using extraction with phenol/chloroform/
light petroleum. Its compositional analysis indicated the
presence of d-GlcN, d-Glc, l-glycero-d-manno- and d-
glycero-d-manno-heptose, 3-deoxy-d-manno-oct-2-uloso-
nic acid (Kdo) and phosphate. From deacylated LPS
obtained after successive treatment with hydrazine and
potassium hydroxide, three oligosaccharides (1±3) were
isolated using high-performance anion-exchange chromato-
graphy, the structures of which were determined by
compositional analysis and one- and two-dimensional
NMR spectroscopy as
in which all sugars are pyranoses, and R and R 0 represent
b-d-Glc (in 1 and 2) and b-d-GlcN (in 1 only), respectively.
d-a-d-Hep is d-glycero-a-d-manno-heptose, l-a-d-Hep is
l-glycero-a-d-manno-heptose, Kdo is 3-deoxy-d-manno-
oct-2-ulosonic acid, and P is phosphate. The liberated lipid
A was analyzed by compositional analyses and MALDI-
TOF MS. Its b-d-GlcN4P-(1!6)-a-d-GlcN-1!P back-
bone is mainly tetra-acylated with two amide- and one
ester-linked (at O3 of the reducing GlcN) (R)-3-hydroxy-
tetradecanoic acid residues, and one tetradecanoic acid that
is attached to the 3-OH group of the amide-linked (R)-3-
hydroxytetradecanoic acid of the nonreducing GlcN.
Additionally, small amounts of tri- and hexa-acylated
lipid A species occur.
Keywords: lipopolysaccharide; mass spectrometry; NMR
spectroscopy; structural analysis; Yersinia enterocolitica.
Lipopolysaccharide (LPS) is a major constituent of theouter leaflet in the outer membrane found in Gram-negativebacteria. In complete (S-form) LPS three major domainsconstitute this molecule: (a) lipid A [1], which is embeddedin the bacterial membrane, serves as anchor for thecarbohydrate moiety, and exhibits the endotoxic activities,(b) the core region [2], which is covalently linked to lipidA, always contains 3-deoxy-d-manno-oct-2-ulosonic acid(Kdo), which is essential for bacterial viability, and in manycases l-glycero-d-manno-heptopyranose (l,d-Hep), and (c)the O-specific polysaccharide (O-antigen) [3,4] which is in
many cases built up from 20 to 40 repeating units of 1±8monosaccharides and is highly immunogenic, and thusdetermines the bacterial O-serotype. Each region is clearlydistinguishable in its biochemical structure and function,and in its genetic organization and biosynthesis.
Yersinia enterocolitica is a common human Gram-negative enteropathogen causing gastroenteritis followedby severe diarrhea, mainly in children. In adults, typicalsymptoms include terminal ileitis and mesenteric lympha-denitis, mimicking appendicitis [5]. Furthermore, cases ofreactive arthritis have been reported [6,7]. Y. enterocoliticacan be acquired by the ingestion of contaminated food(mainly milk products [8]). The bacteria are able to crossthe intestinal epithelium through the M cells and formextracellular colonies in the infected tissue. From here theycan spread to other organs through the lymphatic system[9]. Pathogenic Yersinia produce a set of proteins encodedby a virulence plasmid (pYV) [10] that enables the bacteriato evade the host's innate defense mechanisms (e.g. lysis,mediated by complement, and phagocytosis by macro-phages or polymorphonuclear cells) [11]. Additionally,chromosomally encoded factors are also known to
Eur. J. Biochem. 268, 554±564 (2001) q FEBS 2001
Correspondence to O. Holst, Research Center Borstel, Parkallee 22,
D-23845 Borstel, Germany. Fax: 1 49 4537188419,
Tel.: 1 49 4537188472, E-mail: [email protected].
Abbreviations: d,d-Hep, d-glycero-d-manno-heptose; HPAEC,
high-performance anion-exchange chromatography; Kdo,
3-deoxy-d-manno-oct-2-ulosonic acid; l,d-Hep,
l-glycero-d-manno-heptose; LPS, lipopolysaccharide.
(Received 25 September 2000, revised 6 November 2000, accepted
10 November 2000)

contribute to yersinial virulence. Interestingly, little atten-tion has been paid to the role of LPS in the virulence ofYersinia. However, the virulence of different pathogenicserotypes varies, i.e. in a mouse model, serotype O:8 ismore virulent than O:3 or O:9. Only bacteria of serotypeO:8 kill mice while those possessing the other serotypes donot unless iron is made available to bacteria by treatment ofmice with desferroxamine or iron [12]. The role of differentO-antigen expression in this context is not clear to date.
For Y. enterocolitica over 70 serotypes can be distin-guished based on O-specific polysaccharides (O-antigens)[13]. Mainly serotypes O:3, O:9 (in Europe), and O:8 (inthe USA and Japan) are of clinical importance [14]. Formany serotypes the chemical structures of the O-antigensare known [3,4], however, the most comprehensiveknowledge on yersinial LPS structure and biosynthesishas been accumulated from studies on serotype O:3. Inaddition to the structure of the O-specific polysaccharide ofY. enterocolitica serotype O:3, the gene clusters encodingthe O-antigen and core region and the complete chemicalstructure of the core region have been elucidated [15±18].Structural analyses [19] and mass spectrometry studies [20]have been performed on the lipid A.
With regard to Y. enterocolitica serotype O:9 nothing isyet known about LPS genetics, however, the structures ofthe lipid A [20] and of the core region [21] were elucidated.In case of Y. enterocolitica serotype O:8, only the structureof the O-antigen (comprising a heteropolymer of branchedpentasaccharide repeating units of GalNAc, Gal, Man, Fuc,and 6d-Gul [22,23]) has been investigated and the genecluster coding for the biosynthesis has been cloned andcharacterized [24,25].
So far, no information has been available on the chemicalstructure and genetic organization of the core region andlipid A of Y. enterocolitica serotype O:8. Here, thestructural elucidation of the lipid A and the core regionfrom LPS of a rough mutant of Y. enterocolitica serotypeO:8 is reported.
M A T E R I A L S A N D M E T H O D S
Bacteria and bacterial LPS
A virulence plasmid cured derivative of Y. enterocoliticastrain 8081-R2 [25], 8081-c-R2, was isolated usingcalcium-restricted growth medium [26]. Strain 8081-c-R2was grown in a fermenter (10 L) in Luria±Bertani mediumat 37 8C. Cells were killed by addition of 5% (w/v) phenol,harvested by centrifugation, washed with acetone and dried.
The LPS was extracted using the phenol/chloroform/lightpetroleum extraction (2 : 5 : 8, by volume) [27]. Remain-ing polynucleotides were digested with Benzonasew(Boehringer Mannheim) according to the manufacturer'sspecifications, and proteins were digested with Proteinase Ktreatment (Boehringer Mannheim) at 60 8C for 4 h. Theremaining suspension was dialyzed against water(5 � 10 L, 4 8C) and lyophilized. Such purified LPS wasobtained in a yield of 1.3% of bacterial dry mass.
Isolation of oligosaccharides and of lipid A
The LPS (394 mg) was de-O-acylated using anhydroushydrazine (Kodak, New York; yield: 303 mg, 77% of the
LPS) and subsequently de-N-acylated under strong alkalineconditions (4 m KOH, 120 8C, 16 h) [28]. After neutraliza-tion, extraction of the fatty acids, and desalting usinggel-permeation chromatography on Sephadex G-10 (Phar-macia), 297 mg of sample (75% of the LPS) was obtained,of which 150 mg was fractionated by semipreparative high-performance anion-exchange chromatography (HPAEC) atpH 6.0, followed by desalting. Three fractions wereisolated (7 mg, 23 mg, and 10 mg, representing 3.5%,11.6%, and 5%, respectively, of the LPS), of which 5 mg,11 mg, and 7 mg, respectively, were rechromatographed byHPAEC under alkaline conditions, then neutralized anddesalted. Thus, pure oligosaccharides 1±3 were obtained(yields: 1, 1 mg; 2, 5 mg; 3, 2 mg [0.7%, 5.3%, and 1.5%,respectively, of the LPS]).
For the preparation of lipid A, LPS (52 mg) washydrolysed (100 8C, 1 h) in 1% aqueous acetic acid(5 mL) and centrifuged (118 000 g, 4 8C, 3 h). Thesediment was washed with water (118 000 g, 4 8C, 18 h)and lyophilized (lipid A, yield: 25 mg, 48% of the LPS).
General and analytical methods
Compositional analyses, the determination of the absoluteconfiguration of GlcN and Glc, methylation analysis ofoligosaccharide 1, and SDS/PAGE (18% acrylamide)following visualization of LPS with silver nitrate wereperformed as described previously [21]. Methylation ofoligosaccharide 1 was carried out according to Hakomori[29]. Measurements of protein and polynucleotide contentswere performed on a Uvicon 931 spectrophotometer at190±400 nm on sample solutions of 0.25 mg´mL21. TLCof LPS was performed on precoated silica gel 60 plates(Merck) with chloroform/methanol/aqueous ammonia (7 m)as mobile phase (6.5:2.5:0.4, by volume). Spots werevisualized by charring with 10% (v/v) ethanolic sulfuricacid. Analyses of amide- and ester-bound fatty acids wereperformed according to Wollenweber and Rietschel [30].The absolute configuration of hydroxy fatty acids wasdetermined by GLC of their corresponding 2(R) and 2(S)-phenyl ethyl amides [31]. Proteins were quantified accord-ing to Bradford [32]. GLC and GLC-MS were carried out asdescribed [33]. The temperature programs for sugarderivatives were 150 8C for 3 min, then 3 8C´min21 to300 8C (for GLC-MS: 5 8C´min21 to 330 8C). The follow-ing temperature programs were used for the analyses offatty acid derivatives on GLC and GLC-MS: 120 8C for3 min, then 5 8C´min21 to 260 8C (or to 320 8C for theanalyses of phenyl ethyl amides).
High-performance anion-exchange chromatography
Analytical HPAEC was performed on a Dionex DX 300chromatography system either equipped with a CarboPacPA1 column (4 � 250 mm, Dionex) and eluted at1 mL´min21 using a gradient of aqueous sodium acetate(1 m, pH 6.0) linearly from 1% to 8% over 25 min, andthen from 8% to 15% over 40 min, or equipped with aCarboPac PA-100 column (4 � 50 mm, Dionex) andeluted with sodium acetate (1 m) in aqueous sodiumhydroxide (0.1 m) linearly from 20% to 40% over65 min. Elution of the oligosaccharides was monitoredwith pulsed amperometric detection, including (at pH 6.0)
q FEBS 2001 Lipopolysaccharide from Y. enterocolitica O:8 (Eur. J. Biochem. 268) 555

postcolumn derivatization using 1.5 m sodium hydroxide.Semipreparative HPAEC was performed with a CarboPacPA1 column (9 � 250 mm, Dionex) at 4 mL´min21 elutedwith the same gradients as above. Appropriate fractionswere neutralized and desalted using gel-permeation chro-matography on a column (2.5 � 57 cm) of Sephadex G-10(Pharmacia) eluted with 10 mm aqueous NH4HCO3.
MALDI-TOF MS
MALDI-TOF MS analyses of LPS and lipid A wereperformed with the two-stage reflectron time-of-flight massanalyzer Bruker-Reflex III (Bruker-Franzen Analytik) inlinear TOF configuration at an acceleration voltage of20 kV. Details of the applied methods are given inLindner [34]. In general the compounds were dissolved indistilled water at a concentration of 10 mg´mL21 andtreated with ion exchanger (Amberlite IR-120, Merck) toremove contaminating cations. A 1-mL aliquot of thesample was then mixed with 0.5 m 2,5-dihydroxybenzoicacid (gentisic acid, Aldrich) in methanol and aliquots of0.5 mL were deposited on a metallic sample holder. Massscale calibration was performed externally with similarcompounds of known chemical structure.
NMR spectroscopy
For structural assignments, NMR spectra were recorded onsolutions (0.5 mL) of 1, 2, and 3 (1.3 mg, 5.4 mg, and1.8 mg, respectively) in 2H2O with Bruker AMX 600(operating frequencies 600.1 MHz for 1H NMR,150.9 MHz for 13C NMR, and 242.9 MHz for 31P NMR)and DPX360 spectrometers (operating frequency 90.6 MHzfor 13C NMR) at 27 8C. The resonances were measuredrelative to internal acetone (dH 2.225; dC 31.45) and to 85%phosphoric acid (dP 0), respectively. Coupling constantswere determined on a first order basis (accuracy ^ 1 Hz)from two-dimensional phase-sensitive double-quantumfiltered (DQF)-1H-1H-COSY spectra using the Brukerprogram cosydfprtp [35,36] with 0.170 s acquisitiontime and 2048 data points in the F2 dimension. Thedata matrix was zero-filled in the F1 dimension to givea matrix of 2048 � 1028 data points and was resolutionenhanced in both dimensions by a shifted sine-bellfunction before Fourier transformation. Total correlationspectroscopy (TOCSY) experiments were performedusing the Bruker standard program mlevtp [37], witha spinlock time of 100 ms. Heteronuclear multiplequantum coherence (HMQC) experiments were per-formed using invbtp [38], rotating frame nuclearOverhauser effect spectroscopy (ROESY) experimentswere carried out using roesytp.2 [39,40], with a mixingtime of 250 ms. In order to assign heptose resonances,HMQC-TOCSY experiments were performed using theBruker program inv4mlprtp [41] with a TOCSY mixingtime of 80 ms. In F2 2048 data points were recordedand zero filling was applied in F1. The spectra wereassigned on a Silicon Graphics workstation using Brukerxwinnmr software.
R E S U L T S A N D D I S C U S S I O N
Isolation and characterization of the LPS fromY. enterocolitica strain 8081-c-R2
Extraction of dry bacterial cells of Y. enterocolitica strain8081-c-R2 using the phenol/chloroform/light petroleum
Table 1. Composition of LPS from Y. enterocolitica strain 8081-
c-R2 (serotype O:8). Phosphateorg, organic phosphate.
Component nmol´mg21 Relative amount a
d-GlcN 676 2�.0
Phosphateorg 639 1�.9
Kdo 605 1�.8
d-Glc 536 1�.6
l,d-Hep 832 2�.5
d,d-Hep
(R)-C12:0(3OH)
(R)-C14:0(3OH)
280
traces
815
0�.8
2.4
C12:0 76 0�.2
C14:0 210 0�.6
a Amount relative to 2 mol of GlcN.
Fig. 1. HPAE chromatogram of
deacylated LPS of Y. enterocolitica
serotype O:8 (strain 8081-c-R2). The
analysis was performed on an analytical
CarboPac PA1 column (4 � 250 mm,
Dionex Corp.) at 1 mL´min21, using a
gradient program of 1% 1 m aqueous sodium
acetate linearly to 8% over 25 min, then
linearly to 15% over 40 min. Bold numbers
correspond to the oligosaccharides isolated
by preparative HPAEC.
556 C. Oertelt et al. (Eur. J. Biochem. 268) q FEBS 2001
method gave LPS in moderate yield. After treatment of theLPS with Benzonasew and proteinase K, the polynucleo-tide and protein contents were below the threshold in UVdetection and protein assay. No glycerophospholipidcontamination was detected in TLC. SDS/PAGEanalysis revealed the presence of a typical R-form LPSwhich was not contaminated with any S-form LPS [24].
This finding was confirmed by MALDI-TOF MS (data notshown).
Chemical analyses identified d-GlcN, Kdo, phosphate,d-Glc, d-glycero-d-manno-heptose (d,d-Hep), and l-gly-cero-d-manno-heptose (l,d-Hep), as well as the fattyacids (R)-3-hydroxytetradecanoic acid (C14:0[3OH]),tetradecanoic acid (C14:0), dodecanoic acid (C12:0) and
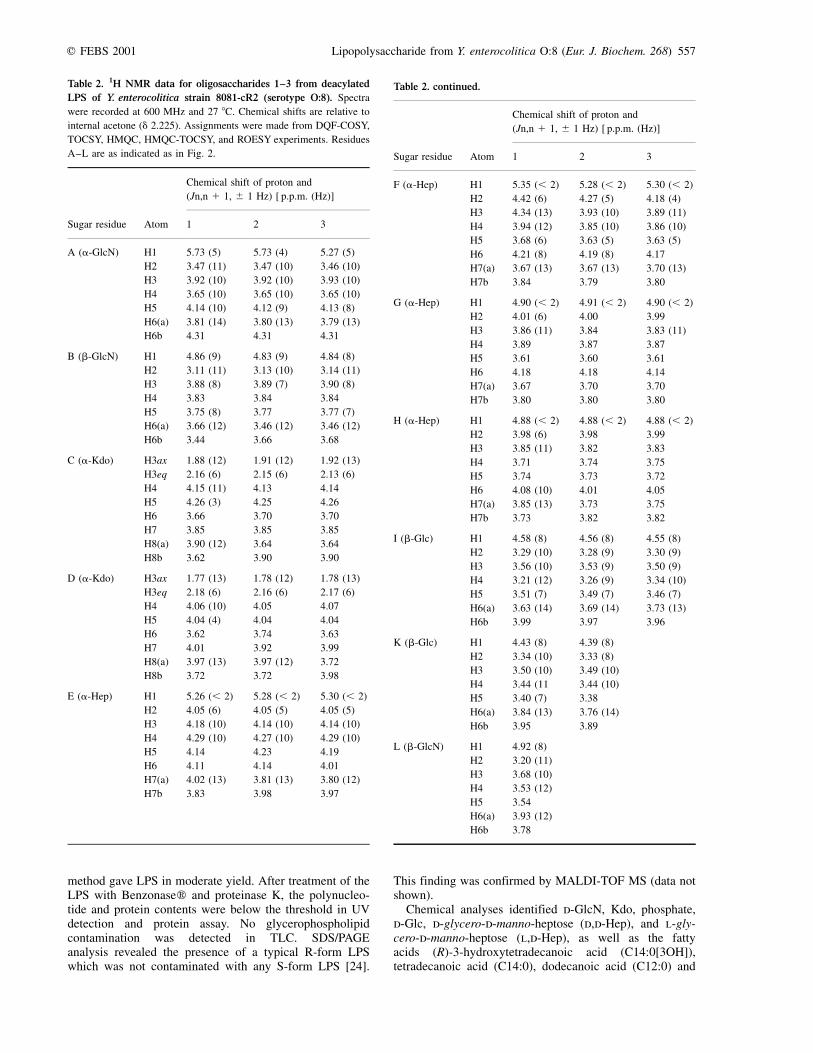
Table 2. 1H NMR data for oligosaccharides 1±3 from deacylated
LPS of Y. enterocolitica strain 8081-cR2 (serotype O:8). Spectra
were recorded at 600 MHz and 27 8C. Chemical shifts are relative to
internal acetone (d 2.225). Assignments were made from DQF-COSY,
TOCSY, HMQC, HMQC-TOCSY, and ROESY experiments. Residues
A±L are as indicated as in Fig. 2.
Chemical shift of proton and
(Jn,n 1 1, ^ 1 Hz) [ p.p.m. (Hz)]
Sugar residue Atom 1 2 3
A (a-GlcN) H1 5�.73 (5) 5�.73 (4) 5�.27 (5)
H2 3.47 (11) 3�.47 (10) 3�.46 (10)
H3 3.92 (10) 3�.92 (10) 3�.93 (10)
H4 3.65 (10) 3�.65 (10) 3�.65 (10)
H5 4.14 (10) 4�.12 (9) 4�.13 (8)
H6(a) 3.81 (14) 3�.80 (13) 3�.79 (13)
H6b 4.31 4�.31 4�.31
B (b-GlcN) H1 4�.86 (9) 4�.83 (9) 4�.84 (8)
H2 3.11 (11) 3�.13 (10) 3�.14 (11)
H3 3.88 (8) 3�.89 (7) 3�.90 (8)
H4 3.83 3�.84 3�.84
H5 3.75 (8) 3�.77 3�.77 (7)
H6(a) 3.66 (12) 3�.46 (12) 3�.46 (12)
H6b 3.44 3�.66 3�.68
C (a-Kdo) H3ax 1�.88 (12) 1�.91 (12) 1�.92 (13)
H3eq 2.16 (6) 2�.15 (6) 2�.13 (6)
H4 4.15 (11) 4�.13 4�.14
H5 4.26 (3) 4�.25 4�.26
H6 3.66 3�.70 3�.70
H7 3.85 3�.85 3�.85
H8(a) 3.90 (12) 3�.64 3�.64
H8b 3.62 3�.90 3�.90
D (a-Kdo) H3ax 1�.77 (13) 1�.78 (12) 1�.78 (13)
H3eq 2.18 (6) 2�.16 (6) 2�.17 (6)
H4 4.06 (10) 4�.05 4�.07
H5 4.04 (4) 4�.04 4�.04
H6 3.62 3�.74 3�.63
H7 4.01 3�.92 3�.99
H8(a) 3.97 (13) 3�.97 (12) 3�.72
H8b 3.72 3�.72 3�.98
E (a-Hep) H1 5�.26 (, 2) 5�.28 (, 2) 5�.30 (, 2)
H2 4.05 (6) 4�.05 (5) 4�.05 (5)
H3 4.18 (10) 4�.14 (10) 4�.14 (10)
H4 4.29 (10) 4�.27 (10) 4�.29 (10)
H5 4.14 4�.23 4�.19
H6 4.11 4�.14 4�.01
H7(a) 4.02 (13) 3�.81 (13) 3�.80 (12)
H7b 3.83 3�.98 3�.97
Table 2. continued.
Chemical shift of proton and
(Jn,n 1 1, ^ 1 Hz) [ p.p.m. (Hz)]
Sugar residue Atom 1 2 3
F (a-Hep) H1 5�.35 (, 2) 5�.28 (, 2) 5�.30 (, 2)
H2 4.42 (6) 4�.27 (5) 4�.18 (4)
H3 4.34 (13) 3�.93 (10) 3�.89 (11)
H4 3.94 (12) 3�.85 (10) 3�.86 (10)
H5 3.68 (6) 3�.63 (5) 3�.63 (5)
H6 4.21 (8) 4�.19 (8) 4�.17
H7(a) 3.67 (13) 3�.67 (13) 3�.70 (13)
H7b 3.84 3�.79 3�.80
G (a-Hep) H1 4�.90 (, 2) 4�.91 (, 2) 4�.90 (, 2)
H2 4.01 (6) 4�.00 3�.99
H3 3.86 (11) 3�.84 3�.83 (11)
H4 3.89 3�.87 3�.87
H5 3.61 3�.60 3�.61
H6 4.18 4�.18 4�.14
H7(a) 3.67 3�.70 3�.70
H7b 3.80 3�.80 3�.80
H (a-Hep) H1 4�.88 (, 2) 4�.88 (, 2) 4�.88 (, 2)
H2 3.98 (6) 3�.98 3�.99
H3 3.85 (11) 3�.82 3�.83
H4 3.71 3�.74 3�.75
H5 3.74 3�.73 3�.72
H6 4.08 (10) 4�.01 4�.05
H7(a) 3.85 (13) 3�.73 3�.75
H7b 3.73 3�.82 3�.82
I (b-Glc) H1 4�.58 (8) 4�.56 (8) 4�.55 (8)
H2 3.29 (10) 3�.28 (9) 3�.30 (9)
H3 3.56 (10) 3�.53 (9) 3�.50 (9)
H4 3.21 (12) 3�.26 (9) 3�.34 (10)
H5 3.51 (7) 3�.49 (7) 3�.46 (7)
H6(a) 3.63 (14) 3�.69 (14) 3�.73 (13)
H6b 3.99 3�.97 3�.96
K (b-Glc) H1 4�.43 (8) 4�.39 (8)
H2 3.34 (10) 3�.33 (8)
H3 3.50 (10) 3�.49 (10)
H4 3.44 (11 3�.44 (10)
H5 3.40 (7) 3�.38
H6(a) 3.84 (13) 3�.76 (14)
H6b 3.95 3�.89
L (b-GlcN) H1 4�.92 (8)
H2 3.20 (11)
H3 3.68 (10)
H4 3.53 (12)
H5 3.54
H6(a) 3.93 (12)
H6b 3.78
q FEBS 2001 Lipopolysaccharide from Y. enterocolitica O:8 (Eur. J. Biochem. 268) 557

traces of 3-hydroxydodecanoic acid (C12:0[3OH])(Table 1).
Isolation and structural analysis of oligosaccharides 1±3
The LPS was deacylated in order to elucidate the structureof its carbohydrate backbone. The elution profile obtainedin HPAEC at pH 6.0 of deacylated LPS is shown in Fig. 1.Further purification by HPAEC at pH 12.0 resulted in theisolation of the three oligosaccharides 1, 2, and 3 (Fig. 2).
Other fractions obtained were shown to represent artefactsof these molecules, including those ones lacking thebranching Kdo residue (D in Fig. 2), and were notinvestigated further.
The structures of oligosaccharides 1±3 (Fig. 2) werecharacterized by NMR spectroscopy (Fig. 3). Assignmentsof the signals was accomplished with DQF-COSY, TOCSY,HMQC, P,H-COSY, and HMQC-TOCSY experiments.Chemical shift data are presented in Tables 2 and 3. Inall three oligosaccharides, the signals of four anomeric
Fig. 2. Structures of oligosaccharides 1, 2,
and 3 isolated by preparative HPAEC from
deacylated LPS of Y. enterocolitica
serotype O:8 strain 8081-c-R2.
Fig. 3. 1H NMR spectrum of 1 and the anomeric regions of 1, 2, and 3. Residues A to L are as in Fig. 2.
558 C. Oertelt et al. (Eur. J. Biochem. 268) q FEBS 2001

protons were identified (1: E, d 5.26; F, d 5.35; G, d 4.90; H,d 4.88; 2: E, d 5.28; F, d 5.28; G, d 4.91; H, d 4.88; 3: E,d 5.30; F, d 5.30; G, d 4.90; H, d 4.88; for designation ofresidues see Fig. 2) accounting to manno-configuredheptose residues as established by small 3JH1,H2 couplingconstants (# 2 Hz) and by the coupling constants of ringprotons [21]. Furthermore, in oligosaccharide 1 and 2 four
signals of anomeric protons (1: A [a-GlcN, d 5.73, 3JH1,H2
5 Hz], B [b-GlcN, d 4.86, 3JH1,H2 9 Hz], I [b2Glc, d 4.58,3JH1,H2 8 Hz], K [b-Glc, d 4.43, 3JH1,H2 8 Hz]; 2: A [d 5.73,3JH1,H2 4 Hz], B [d 4.83, 3JH1,H2 9 Hz], I [d 4.56, 3JH1,H2
8 Hz], K [d 4.39, 3JH1,H2 8 Hz]) and in 3, three such signals(A [d 5.27, 3JH1,H2 5 Hz], B [d 4.84, 3JH1,H2 8 Hz], I(d 4.55, 3JH1,H2 8 Hz]) were identified and attributed tosugars possessing the gluco configuration. Residue L in 1was established as b-linked GlcN (H1: d 4.92, 3JH1,H2
8 Hz), by characteristic high-field shifts of H2 (d 3.20) andC2 (d 56.95). Residues C and D were identified as a-linkedKdo residues (1: C, H3ax, d 1.88, H3eq, d 2.16; D, H3ax,d 1.77, H3eq, d 2.18; 2: C, H3ax, d 1.91, H3eq, d 2.15; D,H3ax, d 1.78, H3eq, d 2.16; 3: C, H3ax, d 1.92, H3eq,d 2.13; D, H3ax, d 1.78, H3eq, d 2.17).
Carbon chemical shifts were assigned by HMQC andHMQC-TOCSY experiments. For undecasaccharide 1, 11signals of anomeric carbons were identified (Table 3). Inboth, oligosaccharides 2 and 3 nine anomeric signals were
Table 3. 13C NMR data for oligosaccharides 1±3 from deacylated
LPS of Y. enterocolitica strain 8081-c-R2 (serotype O:8). Spectra
were recorded at 90.6 MHz and 27 8C. Chemical shifts are relative to
internal acetone (d 31.45). Assignments were made from 135-DEPT
and HMQC experiments. Residues A±K are as indicated as in Fig. 2.
Chemical shift of carbon (p.p.m.)
Sugar residue Atom 1 2 3
A (a-GlcN) C1 92�.76 a 92�.34 a 91�.51 a
C2 55.37b 55�.07b 54�.24 b
C3 70.51 70�.42 69�.32
C4 70.53 70�.18 69�.46
C5 73.32 73�.51 72�.36
C6 70.75 70�.21 69�.78
B (b-GlcN) C1 100�.4 100�.07 99�.27
C2 56.75 56�.4 55�.56
C3 72.85 72�.53 71�.69
C4 75.56 c 75�.27 e 74�.73 e
C5 75.04 d 74�.72 f 73�.92 f
C6 63.58 63�.21 62�.37
C (a-Kdo) C1 175�.81 175�.53 175�.91
C2 100.76 100�.45 99�.69
C3 35.03 35�.17 34�.26
C4 71.95 71�.35 70�.30
C5 68.78 69�.71 69�.26
C6 72.93 72�.85±73.30 g 72�.0±72.3 g
C7 71.05 70�.23 70�.54
C8 64.88 64�.52 63�.63
D (a-Kdo) C1 175�.36 175�.06 174�.62
C2 102.88 102�.28 101�.04
C3 35.08 35�.41 34�.32
C4 67.51 67�.14 66�.17
C5 67.83 67�.42 66�.45
C6 73.10±73.50 g 72�.85±73.30 g 72�.0±72.3 g
C7 71.77 70�.87 70�.44
C8 64.28 64�.00 63�.08
E (a-Hep) C1 99�.78 99�.78 99�.15
C2 72.01 72�.30 71�.61
C3 75.62 76�.43 74�.57
C4 74.08 73�.51 72�.53
C5 73.39 73�.36 72�.31
C6 72.01 69�.71 68�.69
C7 65.24 64�.91 63�.94
F (a-Hep) C1 100�.7 101�.28 101�.66
C2 73.77 79�.12 69�.82
C3 74.46 70�.54±70.61 g 70�.34
C4 65.45 67�.51 66�.24
C5 73.6 72�.85±73.30 g 72�.0±72.3 g
C6 69.02 68�.49 67�.39
C7 73.05 71�.98 71�.43
Table 3. continued.
Chemical shift of carbon (p.p.m.)
Sugar residue Atom 1 2 3
G (a-Hep) C1 103�.55 102�.36 101�.57
C2 71.18 71�.29 70�.06
C3 71.53 71�.33 70�.38
C4 67.22 66�.95 66�.03
C5 73.10±73.50 g 72�.85±73.30 g 72�.0±72.3 g
C6 69.02 68�.47 68�.20
C7 71.18 70�.54±70.61 g 69�.59
H (a-Hep) C1 101�.56 101�.28 100�.47
C2 70.69 70�.54±70.61 g 70�.04
C3 72.01 71�.68 71�.14
C4 68.53 68�.22 67�.31
C5 74.33 74�.00 73�.28
C6 72.74 71�.65 70�.83
C7 62.73 62�.46 61�.50
I (b-Glc) C1 103�.31 103�.03 102�.19
C2 74.77 74�.5 73�.62
C3 76.71 76�.31 75�.50
C4 72.14 71�.68 69�.98
C5 77.97 77�.4 76�.22
C6 62.73 62�.46 61�.48
K (b-Glc) C1 103�.89 103�.47
C2 73.39 73�.44
C3 73.79 76�.05
C4 70.33 70�.13
C5 77.03 77�.05
C6 60.55 61�.48
L (b-GlcN) C1 94�.79
C2 56.95
C3 73.35
C4 70.89
C5 77.27
C6 61.43
a JC1, P 4 Hz. b JC2, P 9 Hz. c JC4 0 , P 3 Hz. d JC5 0 , P 6 Hz. e JC4 0 , P 4 Hz.f JC5 0 , P 5 Hz. g Nonresolved multiplet.
q FEBS 2001 Lipopolysaccharide from Y. enterocolitica O:8 (Eur. J. Biochem. 268) 559

Table 4. ROESY data for oligosaccharides 1±3. Residues A±K are as indicated as in Fig. 2. Bold numbers in parentheses designate the
oligosaccharides in which the particular NOE contact was found. In all other cases, the NOE contacts were identified in all three oligosaccharides.
NOE signal
Sugar From Intra-unit Interunit
A A1 A2
B B1 B2 (2,3), B3 (2,3), B4 (2), B5 (2) A6a
C
D
C3ax
C3eq
D3eq
C4
C4
D4
E2
D6
E5 (2,3)
E E1 E2 C5, C7
F
G
H
F1
F2
G1
G6 (1)
H1
F2
G2, G3 (1,2)
G7
H2
E2, E3, I2 (1,2), K1 (2)
E1 (1), I2 (1), I4 (1), I6 (1,2)
F6, F7a,b (2,3)
G6, G7a,b
I I1 I2, I3, I5 E3 (2), E4, E5 (2), E6, K5 (2)
K (1,2) K1 K2, K3, K4, K5 F2 (2)
L (1) L1 L2, L3, L4 F2, F3, F4
Fig. 4. Part of the (A) ROESY and (B) TOCSY spectrum of 1. Residues A to L are designated as in Fig. 2.
560 C. Oertelt et al. (Eur. J. Biochem. 268) q FEBS 2001

found, however, 2 turned out to be a decasaccharide as thesignal at d 101.28 consisted of two unresolved resonances.Low-field shifted signals indicated the substitution of O6 ofresidues A and B (1: C6 at d 70.75 and d 63.58, respec-tively; 2: d 70.21 and d 63.21; 3: d 69.78 and d 62.37; in
general, the substitution of Kdo to O6 of GlcN B inducesonly a weak a-effect [42]), O4 and O5 of residue C (1: C4at d 71.95 and C5 at d 68.78; 2: d 71.35 and d 69.71; 3:d 70.30 and d 69.26), O3 and O4 of residue E (1: C3 atd 75.62 and C4 at d 74.08; 2: d 76.43 and at d 73.51; 3:
Fig. 5. Negative and positive ion MALDI-TOF spectra of lipid A from Y. enterocolitica serotype O:8. [LAhexa], [LAtetra] and [LAtri], hexa-,
tetra- and tri-acylated lipid A moieties, respectively. Oxonium ions 1±3 (O1±O3) indicate fragments resulting from the cleavage of the glycosidic
GlcN-GlcN bond in the lipid A. GlcN A is as in Fig. 2.
q FEBS 2001 Lipopolysaccharide from Y. enterocolitica O:8 (Eur. J. Biochem. 268) 561

d 74.57 and d 72.53), and O7 of residue G (1: C7 at d 71.18,2: d 70.54±70.61; 3: d 69.59). Significant differences werefound in the chemical shifts of residue F: in undecasaccharide1 the signals for C2 (d 73.77), C3 (d 74.46), and C7 (d 73.05),are shifted downfield, indicating the substitution at O2, O3,and O7. However, in decasaccharide 2 only the chemical shiftsof C2 (d 79.12) and C7 (d 71.98) and in nonasaccharide 3only the one of C7 (d 71.43) are shifted downfield,indicating the substitution at O2 and O7, and at O7 alone,respectively. Residues D, I, K, and L are terminal residues.
The sequence of the monosaccharides in oligosacchar-ides 1±3 was established by ROESY experiments (Table 4,Fig. 4). The following interresidual contacts were identifiedin all three molecules: from B1 to A6a, from E1 to C5 andC7, from F1 to E2 and E3, from G1 to F6, from H1 to G6and G7a,b, and from I1 to E4 and E6. Additionally, NOEcontacts in 1 were detected from L1 to F2, F3 and F4. ForK1, an interresidual contact was not detectable, due tosignal overlap with F2. However, the linkage betweenresidues K and F was established from oligosaccharide 2,for which an interresidual contact between K1 and F2 wasobserved. The linkage between residues C and D wasproven by an NOE contact between C3eq and D6,indicating an a-(2±4)-linked disaccharide. Summarizingthe data, oligosaccharides 1±3 share the same structuralmotif, in which B is linked to O6 of residue A, D to O4 ofC, E to O5 of C, F to O3 of E, G to O7 of F, H to O7 of Gand I to O4 of E. Therefore, residue C must be linked to O6of B. In addition, residue K is linked to O2 of F in 1 and 2and residue L to O3 of F in 1.
1H and 13C NMR spectra of the core oligosaccharidemixture obtained after mild acetic acid hydrolysis revealed
inter alia a signal of an N-acetyl group (1H, d2.06; 13C,d2.39) and corresponding signals of H2 (d3.81) and C2(d56.64) which showed L to be a GlcNAc residue in theLPS.
The 31P NMR spectra gave two signals (1: d21.21 andd 0.38; 2: d21.02 and d 0.28; 3: d20.82 and d 0.32) whichcould be assigned to monophosphate groups attached to O1of residue A (a-GlcN) and O4 of residue B (b-GlcN),respectively.
Methylation analysis of 1 yielded the products ofterminal Glcp (residues I and K), terminal d,d-Hepp (H),terminal GlcpN (L), 3,4-disubstituted l,d-Hepp (E),7-substituted l,d-Hepp (G 0), and 2,3,7-trisubstitutedl,d-Hepp (F).
MALDI-TOF MS experiments
Lipid A was obtained after acetic acid hydrolysis of LPSand was subjected to MALDI-TOF MS analyses (Fig. 5).The negative ion MALDI-TOF mass spectrum reveals threequasimolecular ions [M±H]2. One major molecular ion atm/z 1388.9 and two minor molecular ions at m/z 1797.6 and1178.5 were found. The first molecular ion (at m/z 1797.6)corresponds to a hexa-acylated lipid A species commonlyfound in enterobacteria comprising the GlcN disaccharidesubstituted by two monophosphate groups and fourmolecules of C14:0(3OH), and one each of C14:0 andC12:0. The most abundant molecular ion at m/z 1388.9corresponds to a tetra-acylated lipid A species comprisingthe GlcN disaccharide substituted by two monophosphategroups and three molecules of C14:0(3OH) and one C14:0.The molecular ion at m/z 1178.5 corresponds to a triacyl
Fig. 6. Chemical structure of the LPS from
Y. enterocolitica strain 8081-c-R2 (serotype
O:8).
562 C. Oertelt et al. (Eur. J. Biochem. 268) q FEBS 2001

lipid A comprising three C14:0(3OH) residues. For allthree molecular species, analogues containing only onemonophosphate residue were identified (at m/z 1717.5, m/z1308.3 and m/z 1098.6, respectively). As cleavage of thephosphate in glycosidic linkage may occur during acidichydrolysis, milder acidic conditions were used to liberatelipid A (acetate buffered hydrolysis with pH 4.4 at varyingreaction times), however, no differences in terms ofphosphate substitution were observed (data not shown).Thus, the smaller amounts of monophosphoryl lipid Aspecies may be due to natural heterogeneity.
In order to establish the distribution of fatty acids, apositive ion MALDI-TOF mass spectrum of lipid A wasrecorded (Fig. 5, bottom). Besides the three quasimolecularions [M 1 Na]1 at m/z 1821.8 and 1413.2, severalfragment ions are recorded originating from the cleavageof the phosphate group linked to C1 [M 2 (Na 1 P)]2, m/z1701.5, 1292.8, and 1081.5 (from triacyl lipid A,quasimolecular ion was not detectable). Furthermore, thespectrum comprises oxonium ions at m/z 1087.7, 859.6, and679.0 which represent nonreducing GlcN moieties inducedby the cleavage of the b-(1!6)-bond of the GlcNdisaccharide during the desorption/ionization process. Theoxonium ion at m/z 1087.7 is related to the hexaacyl lipid A(one GlcN substituted by one monophosphate group, twomolecules of C14:0(3OH), one C14:0 and one C12:0,calculated mass 1087.5 Da). The fragment with m/z 859.6resulted from b-elimination of C14:0 [one GlcN substitutedby a monophosphate group, one (D2)C14:1, and one C12:0,calculated mass 859.1 Da]. The most abundant oxoniumion (m/z 679.0) corresponds to the nonreducing GlcNsubstituted by one monophosphate residue, oneC14:0(3OH) and one C14:0 (calculated mass 678.9 Da).No oxonium ion was identified which contained C12:0instead of C14:0, proving that C14:0 is attached to the3-hydroxy group of the amide-linked C14:0(3OH) at GlcNB. The ion at m/z 735.1 corresponds to a reducingmonophosphorylated GlcN A substituted by twoC14:(3-OH), and the ion at m/z 615 a fragment inducedby the cleavage of the glycosidically linked phosphate.These species prove that only two 3-hydroxy tetradecanoicacid residues are attached to this residue.
In summary, the data show that the main lipid A speciesfound in LPS of Y. enterocolitica serotype O:8 is a tetra-acylated and bisphosphorylated glucosamine disaccharide.The reducing GlcN is substituted by two C14:0(3OH)residues at positions 2 and 3. This finding is in accordancewith earlier reports [20] and undermines the conservednature of this structural element. The nonreducing GlcNresidue is substituted with only one (amide-linked)C14:0(3OH) and one C14:0. The positions of these twofatty acid residues were deduced from the oxonium ion atm/z 1087.7 in which the amide linked 3-acyloxyacyl moietyconstitutes C14:0(3-O(C14:0).
Taken together, the complete structure of the LPS of arough mutant from Y. enterocolitica serotype O:8 (strain8081-c-R2) was characterized as shown in Fig. 6. The lipidA part consists mainly of a tetra-acylated and bisphos-phorylated glucosamine disaccharide, harboring twoC14:(3OH) residues on the reducing GlcN, and oneamide-linked acyloxyacyl (C14:0[3-O-C14:0]) residue onthe nonreducing GlcN. The carbohydrate backbone of themajor portion of the core region of this LPS (decasaccharide
2 in Fig. 2) is identical to that of LPS from Y. enterocoliticaserotypes O:3 (strains 75S and 75R) [15,16] and O:9 (strainRuokola/71-c-FR1±37) [21]. However, in serotype O:3 theterminal d,d-Hep is substituted nonstoichiometrically withphosphate at O7 which is not the case in LPS of serotypesO:8 and O:9. Apparently, the Y. enterocolitica corestructures are basically conserved among different sero-types. The structure of an outer core region has beendescribed for the LPS of serotype O:3, but moleculargenetic studies suggest that this hexasaccharide mayrepresent one repeating unit of an ancestral O-antigen,because the organization and localization of the genecluster show similarities to O-antigen gene clusters of otherYersiniae [13]. This outer core region is required for fullvirulence of Y. enterocolitica O:3 in a mouse model [43].Noteworthy, the O-antigen expressed in Y. enterocoliticaserotype O:3 is not attached to the outer core but instead tothe inner core region [17].
Interestingly, in the serotype O:8 strain an additionalbranching b-linked GlcNAc was found linked to O3 of l,d-Hep residue F (undecasaccharide 1, Fig. 2). This structuralvariant has not been described before in LPS of Yersinia.Since the outer core in serotype O:3 is attached at thesame position, it is feasible to speculate that this branchingsugar might represent a rudimentary outer core orO-antigen residue. Studies are underway to clarify thisquestion.
A C K N O W L E D G E M E N T S
The authors thank H. Moll and R. Engel for help with GC-MS, H.-P.
Cordes for recording the NMR spectra, H. LuÈthje for help with
MALDI-TOF MS, R. Venho for construction of 8081-c-R2, and
V. Susott and A. MuÈller for expert technical assistance. This work was
funded in part by the Deutscher Akademischer Austauschdienst (to
C. O. and O. H.) and by the Academy of Finland (to M.S.).
R E F E R E N C E S
1. ZaÈhringer, U., Lindner, B. & Rietschel, E.T. (1999) Chemical
structure of lipid A: Recent advances in structural analysis of
biologically active molecules. In Endotoxin in Health and Disease
(Brade, H., Opal, S.M., Vogel, S.N. & Morrison, D.C., eds),
pp. 93±113. Marcel Dekker, New York, USA.
2. Holst, O. (1999) Chemical structures of the core region of
lipopolysaccharides. In Endotoxin in Health and Disease (Brade,
H., Opal, S.M., Vogel, S.N. & Morrison, D.C., eds), pp. 115±154.
Marcel Dekker, New York, USA.
3. Knirel, Y.A. & Kochetkov, N.K. (1994) The structure of
lipopolysaccharides of Gram-negative bacteria. III. The structure
of O-antigens: a review. Biochemistry (Moscow) 59, 1325±1383.
4. Jansson, P.E. (1999) The chemistry of O-polysaccharide chains in
bacterial lipopolysaccharides. In Endotoxin in Health and Disease
(Brade, H., Opal, S.M., Vogel, S.N. & Morrison, D.C., eds),
pp. 155±178. Marcel Dekker, New York, USA.
5. Chandler, N.D. & Parisi, M.T. (1994) Yersinia enterocolitica
masquerading as appendicitis. Arch. Pediatr. Adolesc. Med. 148,
527±528.
6. Simonet, M.L. (1999) Enterobacteria in reactive arthritis: Yersinia,
Shigella, and Salmonella. Rev. Rheum. Engl. 66, 14S±18S.
7. Gaston, J.S., Cox, C. & Granfors, K. (1999) Clinical and
experimental evidence for persistent Yersinia infection in reactive
arthritis. Arthritis Rheum. 42, 2239±2242.
8. Ackers, M.L., Schoenfeld, S., Markman, J., Smith, M.G.,
Nicholson, M.A., DeWitt, W., Cameron, D.N., Griffin, P.M. &
q FEBS 2001 Lipopolysaccharide from Y. enterocolitica O:8 (Eur. J. Biochem. 268) 563

Slutsker, L. (2000) An outbreak of Yersinia enterocolitica O:8
infections associated with pasteurized milk. J. Infect. Dis. 181,
1834±1837.
9. Bottone, E.J. (1997) Yersinia enterocolitica: the charisma
continues. Clin. Microbiol. Rev. 10, 257±276.
10. Cornelis, G.R., Boland, A., Boyd, A.P., Geuijen, C., Iriarte, M.,
Neyt, C., Sory, M.P. & Stainier, I. (1998) The virulence plasmid of
Yersinia, an antihost genome. Microbiol. Mol. Biol. Rev. 62,
1315±1352.
11. Cornelis, G.R. (1998) The Yersinia deadly kiss. J. Bacteriol. 180,
5495±5504.
12. Lee, W.H., Smith, R.E., DamareÂ, J.M., Harris, M.E. & Johnston,
R.W. (1981) Evaluation of virulence test procedures for Yersinia
enterocolitica recovered from foods. J. Appl. Bacteriol. 50,
529±539.
13. Skurnik, M. (1999) Molecular genetics of Yersinia lipopolysac-
charide. In Genetics of Bacterial Polysaccharides (Goldberg, J.B.,
ed.), pp. 23±51. CRC Press, Boca Raton FL, USA.
14. Bottone, E.J. (1999) Yersinia enterocolitica: overview and
epidemiologic correlates. Microbes Infect. 1, 323±333.
15. Radziejewska-Lebrecht, J., Shashkov, A.S., Stroobant, V., Warten-
berg, K., Warth, C. & Mayer, H. (1994) The inner core region of
Yersinia enterocolitica Ye75R (0:3) lipopolysaccharide. Eur. J.
Biochem. 221, 343±351.
16. Shashkov, A.S., Radziejewska-Lebrecht, J., Kochanowski, H. &
Mayer, H. (1995) The chemical structure of the outer core region
of the Yersinia enterocolitica O:3 lipopolysaccharide. In 8th
European Carbohydrate Symposium, BO-17. Seville, Spain.
17. Skurnik, M., Venho, R., Toivanen, P. & Al-Hendy, A. (1995) A
novel locus of Yersinia enterocolitica serotype O:3 involved in
lipopolysaccharide outer core biosynthesis. Mol. Microbiol. 17,
575±594.
18. Zhang, L., Al Hendy, A., Toivanen, P. & Skurnik, M. (1993)
Genetic organization and sequence of the rfb gene cluster of
Yersinia enterocolitica serotype O:3: similarities to the
dTDP-l-rhamnose biosynthesis pathway of Salmonella and to
the bacterial polysaccharide transport systems. Mol. Microbiol. 9,
309±321.
19. Jensen, M. (1980) Strukturanalyse der Lipoid A Komponente von
Lipopolysacchariden aus Yersinia enterocolitica, PhD Thesis,
University of Freiburg i. Br., Germany.
20. Aussel, L., TheÂrisod, H., Karibian, D., Perry, M.B., Bruneteau, M.
& Caroff, M. (2000) Novel variation of lipid A structures in strains
of different Yersinia species. FEBS Lett. 465, 87±92.
21. MuÈller-Loennies, S., Rund, S., ErvelaÈ, E., Skurnik, M. &
Holst, O. (1999) The structure of the carbohydrate backbone of
the core-lipid A region of the lipopolysaccharide from a
clinical isolate of Yersinia enterocolitica O:9. Eur. J. Biochem.
261, 19±24.
22. Tomshich, S.V., Gorshkova, R.P. & Ovodov, Y.S. (1987) Structural
investigations of the lipopolysaccharide of Yersinia enterocolitica
serovar O:8. Khimia Prirodnykh Soedeneii 5, 657±664.
23. Tomshich, S.V., Gorshkova, R.P. & Ovodov, Y.S. (1988) Structural
investigations of the lipopolysaccharide of Yersinia enterocolitica
serovar O:8. In Chemistry of Natural Compounds (Shakhidoyatov,
Kh. M., ed.), pp. 545±551. Plenum Publishing Corp., New York,
USA.
24. Zhang, L., Toivanen, P. & Skurnik, M. (1996) The gene cluster
directing O-antigen biosynthesis in Yersinia enterocolitica
serotype O:8: identification of the genes for mannose and
galactose biosynthesis and the gene for the O-antigen polymerase.
Microbiology 142, 277±288.
25. Zhang, L., Radziejewska-Lebrecht, J., Krajewska-Pietrasik, D.,
Toivanen, P. & Skurnik, M. (1997) Molecular and chemical
characterization of the lipopolysaccharide O-antigen and its role
in the virulence of Yersinia enterocolitica serotype O:8.
Mol. Microbiol. 23, 63±76.
26. Higuchi, K. & Smith, J.L. (1961) Studies on the nutrition and
physiology of Pasteurella pestis. VI. A differential plating medium
for the estimation of the mutation rate to avirulence. J. Bacteriol.
81, 605±608.
27. Galanos, C., LuÈderitz, O. & Westphal, O. (1969) A new method for
the extraction of R lipopolysaccharides. Eur. J. Biochem. 9,
245±249.
28. Holst, O. (2000) Deacylation of lipopolysaccharides and isolation
of oligosaccharide phosphates. In Bacterial Toxins Methods and
Protocols (Holst, O., ed.), pp. 345±353. Humana Press, Totowa
NJ, USA.
29. Hakomori, S.-I. (1964) A rapid permethylation of glycolipid and
polysaccharide catalyzed by methylsulfinyl carbanion in dimethyl
sulfoxide. J. Biochem. 51, 205±208.
30. Wollenweber, H.W. & Rietschel, E.T. (1990) Analysis of
lipopolysaccharide (lipid A) fatty acids. J. Microbiol. Meth. 11,
195±211.
31. Rietschel, E.T. (1976) Absolute configuration of 3-hydroxy fatty
acids present in lipopolysaccharides from various bacterial groups.
Eur. J. Biochem. 64, 423±428.
32. Bradford, M.M. (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal. Biochem. 72, 248±254.
33. Vinogradov, E.V., Holst, O., Thomas-Oates, J.E., Broady, K. &
Brade, H. (1992) The structure of the O-antigenic polysaccharide
from lipopolysaccharide of Vibrio cholerae strain H11 (non-O1).
Eur. J. Biochem. 210, 491±498.
34. Lindner, B. (2000) Matrix-assisted laser desorption/ionization
time-of-flight mass spectrometry of lipopolysaccharides. In
Bacterial Toxins Methods and Protocols (Holst, O., ed.),
pp. 311±325. Humana Press, Totowa NJ, USA.
35. Derome, A. & Williamson, M. (1990) Rapid-pulsing artifacts in
double-quantum-filtered COSY. J. Magn. Reson. 88, 177±185.
36. Rance, M., SoÈrensen, O.W., Bodenhausen, G., Wagner, G., Ernst,
R.R. & WuÈthrich, K. (1983) Improved spectral resolution in COSY
1H NMR spectra of proteins via double quantum filtering.
Biochem. Biophys. Res. Commun. 117, 479±485.
37. Bax, A. & Davis, D.G. (1985) MLEV-17-based two-dimensional
homonuclear magnetization transfer spectroscopy. J. Magn. Reson.
65, 355±360.
38. Bax, A. & Subramanian, S. (1986) Sensitivity-enhanced two-
dimensional heteronuclear shift correlation NMR spectroscopy.
J. Magn. Reson. 67, 565±569.
39. Bax, A. & Davis, D.G. (1985) Practical aspects of two-
dimensional transverse NOE spectroscopy. J. Magn. Reson. 63,
207±213.
40. Hwang, T.L. & Shaka, A.J. (1992) Cross relaxation without
TOCSY: Transverse rotating frame Overhauser effect spectro-
scopy. J. Am. Chem. Soc. 114, 3157±3159.
41. Lerner, L. & Bax, A. (1986) Sensitivity-enhanced two-dimensional
heteronuclear relayed coherence transfer NMR spectroscopy.
J. Magn. Reson. 69, 375±380.
42. Bock, K., Thomsen, J.U., Kosma, P., Christian, R., Holst, O. &
Brade, H. (1992) A nuclear magnetic resonance spectroscopic
investigation of Kdo- containing oligosaccharides related to the
genus-specific epitope of Chlamydia lipopolysaccharides.
Carbohydr. Res. 229, 213±224.
43. Skurnik, M., Venho, R., Bengoechea, J.A. & MoriyoÂn, I. (1999)
The lipopolysaccharide outer core of Yersinia enterocolitica
serotype O:3 is required for virulence and plays a role in outer
membrane integrity. Mol. Microbiol. 31, 1443±1462.
564 C. Oertelt et al. (Eur. J. Biochem. 268) q FEBS 2001





![Processing Sticky Cotton: Implication of Trehalulose in ... · separated on the columns (CarboPac PA1 anion-exchange guard column [Dionex Corporation, Sunnyvale, CA] and two CarboPac](https://static.fdocuments.in/doc/165x107/5e82f460e3e76f2f7d74fce0/processing-sticky-cotton-implication-of-trehalulose-in-separated-on-the-columns.jpg)












