
Intracytoplasmic Membrane Structures in Vibrio …MEMBRANESIN V. MARINUS V. marinus strain MP-1 has...
9
JOURNAL OF BACTERIOLOGY, May 1970, p. 552-560 Vol. 102, No. 2 Copyright a 1970 American Society for Microbiology Printed in U.S.A. Intracytoplasmic Membrane Structures in Vibrio marinus R. A. FELTER, S. F. KENNEDY, R. R. COLWELL, AND G. B. CHAPMAN Department of Biology, Georgetown University, Washington, D.C. 20007 Received for publication 2 February 1970 An electron microscope study of Vibrio marinus strains MP-1, an obligate psy- chrophile, and PS-207, a moderate psychrophile, revealed numerous intracellular membranous structures. The structures were found to occur more frequently in V. marinus strain MP-1 than in strain PS-207. The frequency of occurrence'and com- plexity of structure were related to age of the culture. In early logarithmic phase, cells revealed invaginations of the plasma membrane. More complex membrane forms, found in late logarithmic and stationary phase, were either myelin-like sheaths, for which the term "myelemma" is proposed, or membranes randomly arranged throughout the cells. The complex membrane forms were not observed to be di- rectly connected with the plasma membrane. However, they were often found in approximation to the plasma membrane or associated with vacuoles and circular membrane profiles. Individual membranes were of a tripartite structure and of dimensions similar to the cell wall and plasma membrane. Bacterial cell membranes have been the subject of a number of morphological studies. These studies have included the early notations of the existence of the structure (6, 8, 21), description of the behavior of the membrane during cell division (7), demonstration of the role of the membrane in spore formation (16), revelation of the relation- ship between the peripheral cytoplasmic mem- brane and the mesosome (16, 19), and notations (13, 27) and elaborate observations (5, 12, 28) of the ramifications of the membrane. Only the last three reports have addressed themselves to marine bacteria. None of the reports has presented the type of incisive analysis of the bacterial membrane which has been conducted by Robertson (29, 30) and by Korn (24) on the animal cell membrane. Our recent studies of marine bacteria (14, 22) and a preliminary report of continuing work (R. R. Colwell and G. B. Chapman, Bacteriol. Proc., p. 42, 1968) have led to the observation of a striking facility for the production of cell mem- branes by several strains of Vibrio marinus. This membrane production is apparently related to several conditions of culture, including age and temperature (R. R. Colwell and G. B. Chapman, Bacteriol. Proc., p. 42, 1968). The purpose of this report is to promulgate these observations in the hope that they will stimulate others, as they are stimulating us, to give further attention to the membranes of marine bacteria. A system which apparently possesses an exceptional facility for membrane production should prove a propitious field for the study of conditions in- fluencing membrane formation. This report, then, presents our observations of simple to complex membrane structures in V. marinus, occurring during the complete growth cycle, from lag phase through decline phase. MATERIALS AND METHODS Organisms. The strains used were V. marinus strain MP-1, a marine obligate psychrophile, growth temperature range 0 to 20C, and strain PS-207, a marine moderate psychrophile, growth temperature range 0 to 30 C (18). V. marinus strain MP-1 was isolated from a water sample raised from a depth of 1,200 m in the North Pacific Ocean (26). Isolation and description of V. marinus strain MP-1 have been published (11). V. marinus strain PS-207 was isolated from the skin of a Pacific cod caught off Port Orchard, Washington; sampling conditions and procedures for growth, etc., have been described elsewhere (9). V. marinus strain MP-1 cultures were grown at 15 C and V. marinus strain PS-207 cultures at 25 C, the optimal growth temperatures for these strains (18). Details of growth medium and procedures employed for the growth studies and electron microscopy have been published elsewhere (14). RESULTS An electron micrograph of an ultrathin section of a representative bacterial cell, grown in broth culture under aeration at 15 C, revealing the characteristic morphology of V. marinus strain PS-207, is shown in Fig. 1. The ultrastructure of 552 on April 22, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript of Intracytoplasmic Membrane Structures in Vibrio …MEMBRANESIN V. MARINUS V. marinus strain MP-1 has...

JOURNAL OF BACTERIOLOGY, May 1970, p. 552-560 Vol. 102, No. 2Copyright a 1970 American Society for Microbiology Printed in U.S.A.
Intracytoplasmic Membrane Structures inVibrio marinus
R. A. FELTER, S. F. KENNEDY, R. R. COLWELL, AND G. B. CHAPMANDepartment of Biology, Georgetown University, Washington, D.C. 20007
Received for publication 2 February 1970
An electron microscope study of Vibrio marinus strains MP-1, an obligate psy-chrophile, and PS-207, a moderate psychrophile, revealed numerous intracellularmembranous structures. The structures were found to occur more frequently in V.marinus strain MP-1 than in strain PS-207. The frequency of occurrence'and com-plexity of structure were related to age of the culture. In early logarithmic phase, cellsrevealed invaginations of the plasma membrane. More complex membrane forms,found in late logarithmic and stationary phase, were either myelin-like sheaths, forwhich the term "myelemma" is proposed, or membranes randomly arrangedthroughout the cells. The complex membrane forms were not observed to be di-rectly connected with the plasma membrane. However, they were often found inapproximation to the plasma membrane or associated with vacuoles and circularmembrane profiles. Individual membranes were of a tripartite structure and ofdimensions similar to the cell wall and plasma membrane.
Bacterial cell membranes have been the subjectof a number of morphological studies. Thesestudies have included the early notations of theexistence of the structure (6, 8, 21), description ofthe behavior of the membrane during cell division(7), demonstration of the role of the membranein spore formation (16), revelation of the relation-ship between the peripheral cytoplasmic mem-brane and the mesosome (16, 19), and notations(13, 27) and elaborate observations (5, 12, 28) ofthe ramifications of the membrane. Only the lastthree reports have addressed themselves to marinebacteria. None of the reports has presented thetype of incisive analysis of the bacterial membranewhich has been conducted by Robertson (29, 30)and by Korn (24) on the animal cell membrane.Our recent studies of marine bacteria (14, 22)
and a preliminary report of continuing work (R.R. Colwell and G. B. Chapman, Bacteriol. Proc.,p. 42, 1968) have led to the observation of astriking facility for the production of cell mem-branes by several strains of Vibrio marinus.This membrane production is apparently relatedto several conditions of culture, including ageand temperature (R. R. Colwell and G. B.Chapman, Bacteriol. Proc., p. 42, 1968). Thepurpose of this report is to promulgate theseobservations in the hope that they will stimulateothers, as they are stimulating us, to give furtherattention to the membranes of marine bacteria. Asystem which apparently possesses an exceptionalfacility for membrane production should prove a
propitious field for the study of conditions in-fluencing membrane formation.
This report, then, presents our observations ofsimple to complex membrane structures in V.marinus, occurring during the complete growthcycle, from lag phase through decline phase.
MATERIALS AND METHODSOrganisms. The strains used were V. marinus
strain MP-1, a marine obligate psychrophile, growthtemperature range 0 to 20C, and strain PS-207, amarine moderate psychrophile, growth temperaturerange 0 to 30 C (18). V. marinus strain MP-1 wasisolated from a water sample raised from a depth of1,200 m in the North Pacific Ocean (26). Isolation anddescription of V. marinus strain MP-1 have beenpublished (11). V. marinus strain PS-207 was isolatedfrom the skin of a Pacific cod caught off Port Orchard,Washington; sampling conditions and procedures forgrowth, etc., have been described elsewhere (9).V. marinus strain MP-1 cultures were grown at 15 Cand V. marinus strain PS-207 cultures at 25 C, theoptimal growth temperatures for these strains (18).Details of growth medium and procedures employedfor the growth studies and electron microscopy havebeen published elsewhere (14).
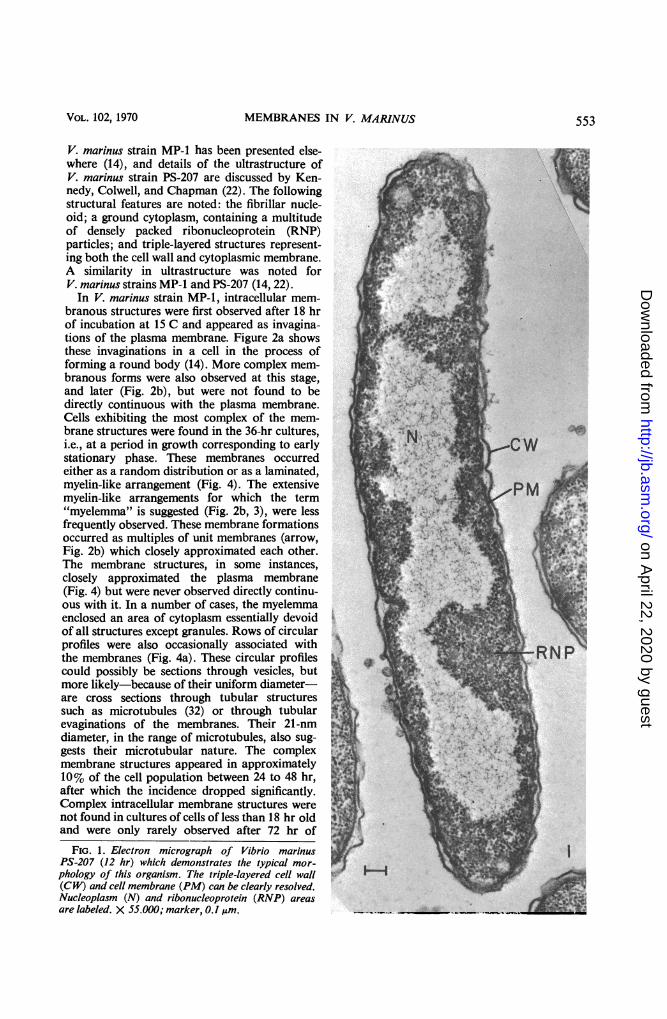
RESULTSAn electron micrograph of an ultrathin section
of a representative bacterial cell, grown in brothculture under aeration at 15 C, revealing thecharacteristic morphology of V. marinus strainPS-207, is shown in Fig. 1. The ultrastructure of
552
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
MEMBRANES IN V. MARINUS
V. marinus strain MP-1 has been presented else-where (14), and details of the ultrastructure ofV. marinus strain PS-207 are discussed by Ken-nedy, Colwell, and Chapman (22). The followingstructural features are noted: the fibrillar nucle-oid; a ground cytoplasm, containing a multitudeof densely packed ribonucleoprotein (RNP)particles; and triple-layered structures represent-ing both the cell wall and cytoplasmic membrane.A similarity in ultrastructure was noted forV. marinus strains MP-1 and PS-207 (14, 22).In V. marinus strain MP-1, intracellular mem-
branous structures were first observed after 18 hrof incubation at 15 C and appeared as invagina-tions of the plasma membrane. Figure 2a showsthese invaginations in a cell in the process offorming a round body (14). More complex mem-branous forms were also observed at this stage,and later (Fig. 2b), but were not found to bedirectly continuous with the plasma membrane.Cells exhibiting the most complex of the mem- *brane structures were found in the 36-hr cultures,i.e., at a period in growth corresponding to early * N wstationary phase. These membranes occurredeither as a random distribution or as a laminated,myelin-like arrangement (Fig. 4). The extensivemyelin-like arrangements for which the term"myelemma" is suggested (Fig. 2b, 3), were lessfrequently observed. These membrane formationsoccurred as multiples of unit membranes (arrow,Fig. 2b) which closely approximated each other.The membrane structures, in some instances, - f1closely approximated the plasma membrane(Fig. 4) but were never observed directly continu-ous with it. In a number of cases, the myelemmaenclosed an area of cytoplasm essentially devoidof all structures except granules. Rows of circularprofiles were also occasionally associated withthe membranes (Fig. 4a). These circular profiles R Ncould possibly be sections through vesicles, butmore likely-because of their uniform diameter-are cross sections through tubular structuressuch as microtubules (32) or through tubularevaginations of the membranes. Their 21-nmdiameter, in the range of microtubules, also sug- . .gests their microtubular nature. The complex :.?X*membrane structures appeared in approximately10% of the cell population between 24 to 48 hr,after which the incidence dropped significantly.Complex intracellular membrane structures werenot found in cultures of cells of less than 18 hr old i'and were only rarely observed after 72 hr of
FIG. 1. Electron micrograph of Vibrio marinusPS-207 (12 hr) which demonstrates the typical mor-phology of this organism. The triple-layered cell wall(CW) and cell membrane (PM) can be clearly resolved.Nucleoplasm (N) and ribonucleoprotein (RNP) areasare labeled. X 55.000; marker, 0.! IAm.
VOL. 102, 1970 553
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

. p
FIG. 2. Plasma and intracytoplasmic membranes. (a) Numerous triple-layered infoldings of the plasma mem-brane (PM) of V. marinus MP-J (18 hr). X43,320; insert, X 131,575. (b) Intracytoplasmic membrane structure(myelemma) of V. marinus MP-I (36 hr). X47,475; markers, 0.1 ,um.
554
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

MEMBRANES IN V. MARINUS
j
'.
. C4
FIG. 3. Enlargement of the myelemma seen in Fig. 2b; V. marinus MP-J (36 hr). X200,825; marker, 0.1 pm.
growth. The intracellular membrane structureswere persistent, however, appearing in necrotic,but apparently intact, cells. Since the number ofcells showing these structures was high and thegrowth studies (14, 22) did not show a markeddrop in viability, even at 72 hr, the complexmembrane structures would appear not to repre-sent only nonviable, degenerating cells.The internal membrane structures observed in
V. niarinus strain PS-207 were, in general, nearlyidentical to those of V. marinus strain MP-1, butsome differences were detected. Intracellularmembranes were first observed at 12 hr and there-after noted at 18, 24, and 36 hr. Figure 5a showsthe distinct intracytoplasmic membrane formationof V. marinus strain PS-207 near the polar regionof the cell, closely approximating the cell mem-brane. In Fig. 5b, a lamination of the membraneis demonstrated, at a site suggesting, but probablynot representing, a presumptive plane of celldivision. Dense, myelin-like formations wereoccasionally observed to enclose an area of cyto-plasm without ribosomal material as previouslynoted in V. marinus strain MP-1 (Fig. 5c). Inboth strains of V. marinus, formation of complexmembranes characteristically was at or near thepolar regions of the cell. The membranes arranged
as lamina were discernible, lying immediately inapproximation to the plasma membrane (Fig. 5c,arrows). The complex myelin-like forms were notas numerous in cells of V. marinus strain PS-207,and the relationship between appearance of com-plex membrane structures with age was not asclear-cut as in V. marinus MP-1. Nevertheless,concentrically ringed membrane systems wereobserved in V. marinus PS-207. Figure 6 showsone such structure in the area between the cellwall and the plasma membrane.
DISCUSSIONDetails of the ultrastructure of V. marinus
have been presented elsewhere for strains MP-1(14) and PS-207 (22). During studies of the ultra-structure of these marine psychrophilic strains ofV. marinus, the occurrence of intracytoplasmicmembranes at various stages of the growth cyclewas noted. Further examination cast appreciablelight on the unusual nature of the membranes.Although intracytoplasmic membranes and ar-rangements of membranes were first observed inheat-treated cells of V. marinus strain MP-1 (10;R. R. Colwell and G. B. Chapman, Bacteriol.Proc., p. 42, 1968), the present study indicatesthat they may also arise under growth conditions
VOL. 102, 1970 555
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

4
4a
FIG. 4. Intracytoplasmic membrane of V. marinus MP-J (36 hr). (a) Polarly located intracytoplasmic mem-brane with concentric configuration. Delineated area containing circular profiles thought to represent cross sec-tions through microtubules or tubular evaginations of the membrane. X 57,000. (b) Laterally located intracyto-plasmic membrane. The membrane is associated with what may be a mesosome-like body cut very superficially ortangentially. X 66,300; markers, 0.1 ,Am.
556
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

N',;44K - *-- 11%- Sb5bFIG. 5a-c. Electron micrographs of V. marinus strain PS-207. (a) Polarly located concentric membrane figure
(18-hr culture); X 62,600. (b) Laminar arrangement ofplasma membrane presumably at the site of cell division(12-hr culture); X 48,000. (c) Myelin-likefigure at polar region (24-hr culture); arrows indicateflattened, lamellararrangement of mesosome-like bodies peripherally located along the plasma membrane; (X 70,200; markers,0.1 IAM.
557
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

FELTER ET AL.
#4" -
t a, :'?to
FIG. 6. Complex membrane formation (myelemma) between plasma membrane and cell wall of V. marinusPS-207 (12 hr). X 67,500; marker, 0.1 "M.
considered optimal for these strains (14), withoutspecific treatment or special culture conditionmodifications.The most common intracellular membranes
observed in both strains of V. marinus, MP-1 andPS-207, arose from invaginations of the plasmamembrane and occurred at > 18 hr (Fig. 2).These invaginations were similar to the invagina-tion of plasma membrane, not associated withmesosomes, occurring near the cell nucleus as
reported by Ryter and Jacob (31) for Escherichiacoli and considered by them to be the site of at-tachment of the bacterial chromosome to thecell membrane. Mesosomes have not been ob-served by us to occur in the strains of V. marinusstudied. Also, the unusual mode of cell divisionin V. marinus [i.e., lack of cell wall participation inthe initial stages of division (14, 22)] suggests aninterpretation of the function of the invaginatedplasma membrane (Fig. 2a), namely, a role inchromosome distribution, consistent with theobservations stated by Ryter and Jacob (31). Theclose approximation of the invaginated membraneto the nuclear material in V. marinus may involvean attachment site, or sites, within the cell, ofthe fibrillar nuclear material to the membrane.
However, this interpretation requires much moreexhaustive study, including analysis of serialsections, before concrete statements can be madewith respect to function.The intracellular membrane structures, shown
in Fig. 2b are similar to those reported for E. coli(4, 15, 31). Fischman and Weinbaum (15) havesuggested that E. coli B, grown in enriched me-dium, synthesizes both an increased amount ofintracellular membrane and an increased amountof membrane-associated enzymes. For purposesof comparison, V. marinus in its normal habitat,i.e., seawater containing milligram quantities oforganic nutrient per liter, may be considered tobe in a minimal medium. Under laboratory cul-tural conditions, V. marinus may be consideredto be in an "ultrarich" growth medium. It wouldnot, therefore, be surprising if "extra" mem-branes were to be synthesized by V. marinus inlaboratory culture. Although many species ofmarine bacteria have been grown in laboratoryculture without producing complex intracellularmembranes, no studies of their ultrastructureover the entire growth cycle have been reported.Relationship of presence of complex membranestructures to age of culture should be examined in
J. BACTERIOL.558
;,I av-
Ix91 I f
4
09
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

MEMBRANES IN V. MARINUS
other strains of bacteria, both marine and non-marine. Comparisons of ultrastructure of V.marinus grown under nutrient conditions approxi-mating the in situ environment and in laboratorymedia are in progress.The complex lamellar membrane system,
termed myelemma, with alternating major denseand intraperiod lines, shown in Fig. 2b, and inenlargement in Fig. 3, is similar in organizationto the myelin-like lamellar structures of lecithinand equal molar portions of lecithin and choles-terol as presented by Bangham and Horne (3) intheir study of the bimolecular leaflet arrangementof lipids. Without appropriate biochemicalanalyses, it is impossible to describe the chemicalnature of these bacterial membranes.
Other authors have described similar myelin-like configurations in microorganisms (2, 17).Kaye and Chapman (20) described membranestructures, also myelin-like, in the initial stagesof antibiosis. Myelin-like configurations havebeen described by Anderson and Roels (2) in thealga, Ochromonas malhamensis. The fine structureof the Ochromonas myelin body was described asa series of concentric membranes folding backupon themselves or appearing to fuse together.Interestingly, Anderson and Roels (2) also pro-duced a lamellar structure in phospholipids (egglecithin) hydrated in water dispersion, electronmicrographs of which were very similar to thoseof the myelin body preparations.From the studies of V. marinus, the more com-
plex membranes were occasionally closely as-sociated with, and, possibly, derived from, smallcircular profiles of membrane (Fig. 4). Thesecircular profiles may represent cross sectionsthrough tubular evaginations of the membraneor through microtubules. The uniform diameterof the profiles makes it seem highly unlikely thatthey represent spherical vesicles. The sections ofthermolabile mutant CR 341 Til of E. coli K-12(23) show vesicular elements but approximationto, or relationship with intracellular membraneswas not clear from the included micrographs.Glauert and Hopwood (17) noted extensiveintracellular membrane systems in Streptomycescoelicolor, in which the membranes were observedto be continuous with the plasma membrane,with invaginations of the plasma membrane form-ing vesicles and lamellar structures of varyingcomplexity.A trilaminar composition and a dimension
similar to the plasma membrane appear to becharacteristics of the intracellular membranes ofthe several types described for V. marinus. Asnoted by Abram (1) for Bacillus stearothermo-philus, the various intracytoplasmic membranouselements in V. marinus possess a similarity in
structural detail to the limiting membranes ofother cells and organelles, namely, they appearas a "unit membrane" (29, 30).The complexity of some of the intracellular
membranes (i.e., the myelemma) of V. marinusstrain MP-1, a marine obligate psychrophile, maybear some physiological relationship to its natu-ral environment. Remsen et al. (28) postulatedthat the intracellular membranes found in thenitrifying bacteria originally arose from theplasma membrane. The "membranous organelle"of Nitrosocystis oceanus, in their view, probablyrepresented a degree of specialization not foundin other bacteria, since in most other bacteriathe cells apparently synthesize the membranes"on demand." Hence, obviously, cytomembraneswhich are not always present are not permanentorganelles. Cytomembranes and myelemma wereobserved in V. marinus throughout its variousgrowth phases. On the basis of the evidence ob-tained to date, it appears that the cytomembranesof V. marinus are membrane systems which arisein the course of growth, probably in response tofactors such as temperature, age, and nutrientsupply. At present, there is no evidence of theirrelationship to the peripheral cytoplasmic mem-brane, other than that of structural similarity.Further study will be required to define the roleof the complex intracellular membranes in V.marinus.
ACKNOWLEDGMENTS
The excellent technical assistance of Sandra Zane and BenjaminElliott is gratefully acknowledged.
This investigation was supported by contract NONR 4810between the Office of Naval Research and Georgetown Univer-sity, and by Public Health Service training grant 5TOlGM01268-05from the Division of General Medical Sciences.
LITERATURE CITED
1. Abram, D. 1965. Electron microscope observations on intactcells, protoplasts, and the cytoplasmic membrane of Bacillusstearothermophilus. J. Bacteriol. 89:855-873.
2. Anderson, 0. R., and 0. A. Roels. 1967. Myelin-like configur-ations in Ochromonas malhamensis. J. Ultrastruct. Res. 20:127-139.
3. Bangham, A. D., and R. W. Horne. 1964. Negative staining ofphospholipids and their structural modification by surface-active agents as observed in the electron microscope. J. Mol.Biol. 8:660-668.
4. Birdsell, D. C., and E. H. Cota-Robles. 1967. Production andultrastructure of lysozyme and ethylenediaminetetraacetate-lysozyme spheroplasts of Escherichia coli. J. Bacteriol. 93:427-437.
5. Brown, A. D., D. G. Drummond, and R. J. North. 1962. Theperipheral structure of gram-negative bacteria. It. Mem-branes of bacilli and spheroplasts ofa marine pseudomonad.Biochim. Biophys. Acta 58:514-531.
6. Chapman, G. B. 1959. Electron microscopy of ultra-thin sec-
tions of bacteria. 111. Cell wall, cytoplasmic membrane andnuclear material. J. Bacteriol. 78:96-104.
7. Chapman, G. B. 1959. Electron microscope observations on
the behavior of the bacterial cytoplasmic membrane duringcellular division. J. Biophys. Biochem. Cytol. 6:221-224.
VOL. 102, 1970 559
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

FELTER ET AL.
8. Chapman, G. B., and A. J. Kroll. 1957. Electron microscopyof ultra-thin sections of Spirillum serpens. J. Bacteriol. 73:63-71.
9. Colwell, R. R. 1962. The bacterial flora of Puget Sound fish.J. Appl. Bacteriol. 25:147-158.
10. Colwell, R. R. 1968. In Marine biology IV, C. H. Oppenheimer(ed.). Unresolved problems in marine microbiology. TheNew York Academy of Sciences Interdisciplinary Com-munications Program, New York.
11. Colwell, R. R., and R. Y. Morita. 1964. Reisolation and emen-dation of description of Vibrio marinus. (Russell) Ford. J.Bacteriol. 88:831-837.
12. Costerton, J. W., C. Forsberg, T. I. Matula, F. L. A. Buckmire,and R. A. Mac Leod. 1967. Nutrition and metabolism ofmarine bacteria. XVI. Formation of protoplasts, sphero-plasts, and related forms from a gram-negative marine bac-terium. J. Bacteriol. 94:1764-1777.
13. de Petris, S. 1967. Ultrastructure of the cell wall of Escherichiacoil and chemical nature of its constituent layers. J. Ultra-struct. Res. 19:45-83.
14. Felter, R. A., R. R. Colwell, and G. B. Chapman. 1969.Morphology and round body formation in Vibrio marinus.J. Bacteriol. 99:326-335.
15. Fischman, D. A., and G. Weinbaum. 1967. The formation ofmultiple layers of membrane-like structures in Escherichiacoli B. J. Cell Biol. 32:524-528.
16. Fitz-James, P. C. 1960. Participation of the cytoplasmic mem-brane in the growth and spore formation of bacilli. J. Bio-phys. Biochem. Cytol. 8:507-528.
17. Glauert, A. M., and D. A. Hopwood. 1959. A membranouscomponent of the cytoplasm in Streptomyces coelicolor.J. Biophys. Biochem. Cytol. 6:515-516.
18. Hanus, F. J., and R. Y. Morita. 1968. Significance of thetemperature characteristic of growth. J. Bacteriol. 95:736-737.
19. Imaeda, T., and M. Ogura. 1963. Formation of intracyto-plasmic membrane system of mycobacteria related to celldivision. J. Bacteriol. 85:150-163.
20. Kaye, J. J., and G. B. Chapman. 1963. Cytological aspects ofantimicrobial antibiosis. III. Cytologically distinguishable
stages in antibiotic action of Colistin sulfate on Escherichiacoli. J. Bacteriol. 86:536-543.
21. Kellenberger, E., and A. Ryter. 1958. Cell wall and cytoplas-mic membrane of Escherichia coli. J. Biophys. Biochem.Cytol. 4:323-325.
22. Kennedy, S. F., R. R. Colwell, and G. B. Chapman. 1969.Ultrastructure of a psychrophilic marine vibria. Can. J.Microbiol., in press.
23. Kohiyama, M., D. Cousin, A. Ryter, and F. Jacob. 1966.Mutants, thermosensibles d'Escherichia coli K12. I. Isole-ment et characterisation rapide. Ann. Inst. Past. 110:465-486.
24. Korn, E. D. 1966. Structure of biological membranes. Science153:1491-1498.
25. Moore, R. T., and G. B. Chapman. 1959. Observations on thefine structure and modes of growth of a streptomycete. J.Bacteriol. 78:878-885.
26. Morita, R. Y., and R. D. Haight. 1964. Temperature effectson the growth of an obligate psychrophilic marine bac-terium. Limnol. Oceanogr. 9:103-106.
27. Murray, R. G. E., P. Steed, and H. E. Elson. 1965. The loca-tion of the mucopeptide in sections of the cell wall of Esch-erichia coli and other gram-negative bacteria. Can. J.Microbiol. 11:547-560.
28. Remsen, C. C., F. W. Valois, and S. W. Watson. 1967. Finestructure of the cytomembranes of Nitrosocystis oceanus.
J. Bacteriol. 94:422-433.29. Robertson, J. D. 1959. The ultrastructure of cell membranes
and their derivatives. Biochem. Soc. Symp. (Cambridge)16:3-43.
30. Robertson, J. D. 1965. Current problems of unit membranestructure and substructure, p. 379-433. In S. Seno and E. V.Cowdry (ed.), Intracellular membrane structure. JapanSociety for Cell Biology, Tokyo.
31. Ryter, A., and F. Jacob. 1966. Etude morphologique de laliaison du noyau a la membrane chez E. coli et chez lesprotoplastes de B. subtilis. Ann. Inst. Pasteur. 110:801-812.
32. Van Iterson, W., J. F. M. Hoeniger, and E. N. Van Zanten.1967. A "microtubule" in a bacterium. J. Cell Biol. 32:1-10.
560 J. BACrERIOL.
on April 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from


















