
International Journal of Pharma and Bio Sciences ISSN 0975 ... · PDF fileAmity Institute of...
12
Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343 This article can be downloaded from www.ijpbs.net B - 333 Research Article Microbiology International Journal of Pharma and Bio Sciences ISSN 0975-6299 PLANT PROMOTING ACTIVITIES OF FUNGAL ENDOPHYTES ASSOCIATED WITH TOMATO ROOTS FROM CENTRAL HIMALAYA, INDIA AND THEIR INTERACTION WITH PIRIFORMOSPORA INDICA NEHA CHADHA, RAM PRASAD AND AJIT VARMA * Amity Institute of Microbial Technology, Amity University of Uttar Pradesh, Sector 125, Noida- 201303, India ABSTRACT Endophytic fungi promote plant growth through the production of IAA, siderophore and phosphate solubilization. Out of thirty fungal endophytes isolated from tomato roots grown in a stressed, cold climate of Central Himalayan hills of district Pithoragarh, Uttarakhand, India twelve showed distinct morphological and study was undertaken, to understand the plant growth promoting properties possessed by these endophytes residing under stress environment. All the fungi showed the zone of phosphate solubilization, seven were positive for siderophore, four for HCN and three-showed ammonia production. IAA production was found maximum in Fusarium fusarioides. All the endophytic fungi showed remarkable phosphate solubilizing activity up to eight days with a decrease on the 10 th day with maximum reporting from Trichoderma peudokoningii. All the isolates showed maximum siderophore production at 21 th day after the inoculation. These isolated root endophytic fungi possess plant growth promoting properties and thus have a potential to be used for biofertilizer. KEYWORDS: Root endophytic fungi, Central Himalayas, Phosphate solubilisation, IAA, siderophore, Piriformospora indica *Corresponding author AJIT VARMA Amity Institute of Microbial Technology, Amity University of Uttar Pradesh, Sector 125, Noida- 201303, India
Transcript of International Journal of Pharma and Bio Sciences ISSN 0975 ... · PDF fileAmity Institute of...

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 333
Research Article Microbiology
International Journal of Pharma and Bio Sciences ISSN
0975-6299
PLANT PROMOTING ACTIVITIES OF FUNGAL ENDOPHYTES ASSOCIATED
WITH TOMATO ROOTS FROM CENTRAL HIMALAYA, INDIA AND THEIR
INTERACTION WITH PIRIFORMOSPORA INDICA
NEHA CHADHA, RAM PRASAD AND AJIT VARMA*
Amity Institute of Microbial Technology, Amity University of Uttar Pradesh,
Sector 125, Noida- 201303, India
ABSTRACT
Endophytic fungi promote plant growth through the production of IAA, siderophore and phosphate solubilization. Out of thirty fungal endophytes isolated from tomato roots grown in a stressed, cold climate of Central Himalayan hills of district Pithoragarh, Uttarakhand, India twelve showed distinct morphological and study was undertaken, to understand the plant growth promoting properties possessed by these endophytes residing under stress environment. All the fungi showed the zone of phosphate solubilization, seven were positive for siderophore, four for HCN and three-showed ammonia production. IAA production was found maximum in Fusarium fusarioides. All the endophytic fungi showed remarkable phosphate solubilizing activity up to eight days with a decrease on the 10th day with maximum reporting from Trichoderma peudokoningii. All the isolates showed maximum siderophore production at 21th day after the inoculation. These isolated root endophytic fungi possess plant growth promoting properties and thus have a potential to be used for biofertilizer. KEYWORDS: Root endophytic fungi, Central Himalayas, Phosphate solubilisation, IAA, siderophore, Piriformospora indica
*Corresponding author
AJIT VARMA
Amity Institute of Microbial Technology, Amity University of Uttar Pradesh,
Sector 125, Noida- 201303, India

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 334
INTRODUCTION
Root endophytic fungi live asymptomatically within the roots of many plant species without causing any harm to the host. Every plant harbor at least one or more endophytic fungi in the universe 1, 2. Endophytic fungi are ubiquitous and highly diverse group in their geographical location. It has been documented that endophytes appear more in tropical than temperate zone 3, 4. They are also found in xerophytic 5 and aquatic environment 6. Fungal endophytic diversity in extreme environmental conditions is of great interest and little attention has been given to these microbes because of their slow growth rate and strenuous handling. Pandey and Palni (2007) have reported that the microbial population decreases with an increase in the altitude 7. Endophytic fungi interact mutualistically with the host plant by producing metabolites that induces resistance and protection against various pathogens. They protect plant in drought 8 as well as in cold climatic conditions. Endophytes are considered as an important source of bioactive components 9. Plant growth promoting fungi (PGPF) promotes plant growth through the production of enzymes, phosphate solubilization10, 11, siderophore production 12 and antagonism to phytopathogens 13. The identification of endophytic fungi through their ability to produce IAA, phosphate, siderophore may be useful in increasing the production of crop plants 14 or in suppression of diseases by pathogen or pests. The Uttarakhand Himalayas, India is extremely rich in flora and fauna. Pithoragarh, situated in the middle hill region of Central Himalaya’s hills has an average altitude of about 5,000 feet. Annual rainfall is 1200 mm approximately and temperature ranges from maximum to minimum i.e. 35°C (summer) to -2°C (winter). Tomato (family Solanaceae) is the most widely cultivated vegetable crop next to potato; they are rich source of nutrition and play an important role in scavenging free radicals from the body. Lycopene present in tomato has an excellent antioxidant activity to fight against anti-aging, cancer and heart diseases 15. Plant growth promoting fungi (PGPF) are non- pathogenic saprophytes and
act as a biocontrol agent against pest and soil pathogens 13. Root colonized with PGPF can lead to systemic resistance in distal parts of the plant 16, 17. PGPF are crop specific and their significance on plant is limited because of variation in climate and soil inconsistency 18,
19. The endophytic fungi residing in cold conditions, where soil is deficient in organic matter and low moisture content possessed plant growth promoting activities and these fungi can be used in increased production of plant in the same environment 20, 21, 22, 23. The present study revealed the plant growth promoting properties of fungal isolates from tomato roots residing under cold stress condition of the central Himalayas of India.
MATERIALS AND METHODS (i) Sample collection and Isolation Tomato plants with the roots were randomly collected from Defence Institute of Bio-Energy Resources (DIBER) field station Pithoragarh district of Uttrakhand India (29°35’N 80°13’E) in the month of September 2012. The whole plant was kept in sterile polythene bags and transported to laboratory for further study. The roots were surface sterilized with 70% ethanol followed by 4% sodium hypochlorite with subsequent washing with distilled water. The roots were crushed with phosphate buffer saline solution pH 7 in mortar pestel and spread on Potato dextrose agar (PDA) plates. Distinct colonies were picked from the mother plate for isolation and were sub cultured on fresh PDA plates. (ii) Plant growth promoting characteristics
of fungal root endophytes The fungal root endophytes were analyzed for their plant growth promoting characteristics viz., production of IAA, ammonia, siderophores, HCN and their ability to solublize phosphates. (iii)Qualitative estimation of phosphate
solubilization on solid medium 24 The phosphate solubilizing activity of fungal isolates were tested on Pikovskaya’s solid agar medium with following composition; (Glucose 10g, Ca3(PO4)2 5g, (NH4)2SO4 0.5g,

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 335
NaCl 0.2g, MgSO4.7H20 0.1g, KCl 0.2g, Yeast extract 0.5g, MnSO4.H2O 0.002g, FeSO4.7H2O 0.002g and Agar 15g in one litre of medium) with bromo phenol blue at a concentration of 0.003%. After incubation of seven days at 28°C, the formation of clear zone around the fungal hyphae indicates the ability of fungus to solubilize inorganic phosphorous. (iv)Quantitative estimation of phosphate
solubilization in liquid medium 25 Fungal disc was inoculated in Pikovskaya’s broth and culture was filter out using whatman filter paper 1 after two, four, six, eight and ten day after inoculation. 1ml supernatant was taken and 10 ml chloromolybdic acid to 40 ml distilled water was added. After this, 1ml chlorostanous acid was added to the above solution and distilled water was added to make up the volume to 50ml. The absorbance of the resulting blue color was measured at 600nm. The concentration of soluble phosphate was calculated by the standard curve of KH2PO4. (v) Quantitative determination of Indole
Acetic Acid (IAA) production 26 For indole acetic acid production, isolates were grown in 25ml Czapek medium (pH 6.5) with and without tryptophan. After 7 days of incubation, 1ml culture filtrate, 2ml Salkowski reagent (0.5M FeCl3 in 35% perchloric acid) was added and incubated for 20 minutes in dark. The reading was taken at 530 nm using UV spectrophotometer (UV 1800 Schimadzu). The concentration of indol was determined using IAA standard curve. (vi)HCN production 27 Fungal isolates were placed on potato dextrose agar medium amended with 4.4g glycine/l. Whatman filter paper no.1 soaked in 2% sodium carbonate in 0.5% picric acid solution was placed at the top of the plate. Plates were sealed with parafilm and incubated for HCN production for 7 days at
28ºC. Discoloration of the filter paper from yellow to orange brown after incubation was considered as positive result for cyanide. (vii) Qualitative estimation of Siderophore
production 28 Detection of siderophore production was carried on CAS medium with some modification. The oranges zone around the fungal colony was an indicative for siderophore production. (viii) Quantitative estimation of
Siderophore production 29 For Quantitative estimation, FeCl3 test was done on culture filtrate. Fungal endophytes were grown on Fries basal medium. The mycelium was separated by whatmann filter paper 1. The culture filtrate was harvested at 4-day interval for a period of 28 days. 1-5 ml of ferric chloride solution (3 %) was added to 1 ml of culture filtrate and absorbance was taken at 260nm. ` (ix)Ammonia production 30 Fungal disc were inoculated in 10 ml potato dextrose broth and incubated at 30°C for 7 days in a rotatory shaker. After incubation, 0.5 ml Nessler’s reagent was added to each tube. The development of yellow to brown color indicated a positive reaction for ammonia production (x) Interaction of root endophytic fungi
with Piriformospora indica 31 The effect of root endophytic fungi on Piriformospora indica was performed on Yeast Maltose Peptone (YMP) agar plates by the dual culture method. Disc of P. indica and fungal isolates (5 mm diameter) were placed on the same YMP agar media, 6 cm from each other. The plates were incubated at 25°C for 7 days. Radius of P. indica and other fungal isolates were recorded. All the experiments were done thrice with three replicates.

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 336
Table 1 Plant growth promoting attributes of endophytic fungi isolated from tomato roots
Root endophytic fungi Phosphate
solubilization Siderophore HCN Ammonia
Chaetomium globosum ++ ++ + - Fusarium oxysporum + + - - Fusarium semitectum + - - - Fusarium solani + - - - Fusarium fusarioides ++ - + - Fusarium moniliforme + - - + Mucor sp + + + - Aspergillus niger gr. ++ + - + Aspergillus sp + - - - Aspergillus versicolor ++ ++ + - Mucor hiemalis + + - + Trichoderma peudokoningii rifai
+ ++ - -
(++ maximum intensity; + minimum intensity; - no observation)
Figure 1 Fungal endophytes shows clear zone on (a) Pikovskaya’s medium with bromophenol blue,
(b) siderophore production and (c) HCN production test
Figure 2 Interaction of Piriformospra indica with a) Fusarium oxysporum, b) Aspergillus niger, c) Mucor sp., d) Chaetomium globosum, e) Aspergillus versicolor, f) Mucor hiemalis, g) Trichoderma peudokoningii rifai. One disc of each root endophytic fungi was placed on left side and P. indica disc on right side of YMP medium.
a) b) c)
Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 337
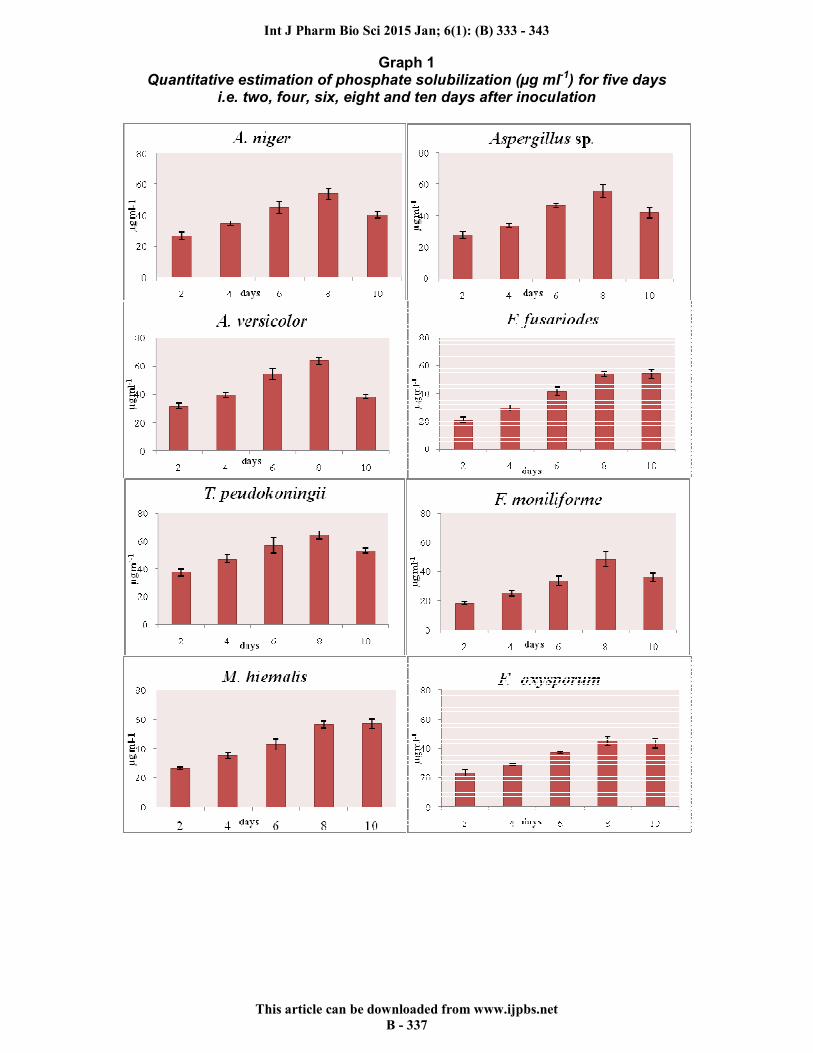
Graph 1 Quantitative estimation of phosphate solubilization (µg ml-1) for five days
i.e. two, four, six, eight and ten days after inoculation

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 338
Graph 2 Comparison of indole acetic acid (µg ml-1) by root fungal endophytes
Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 339
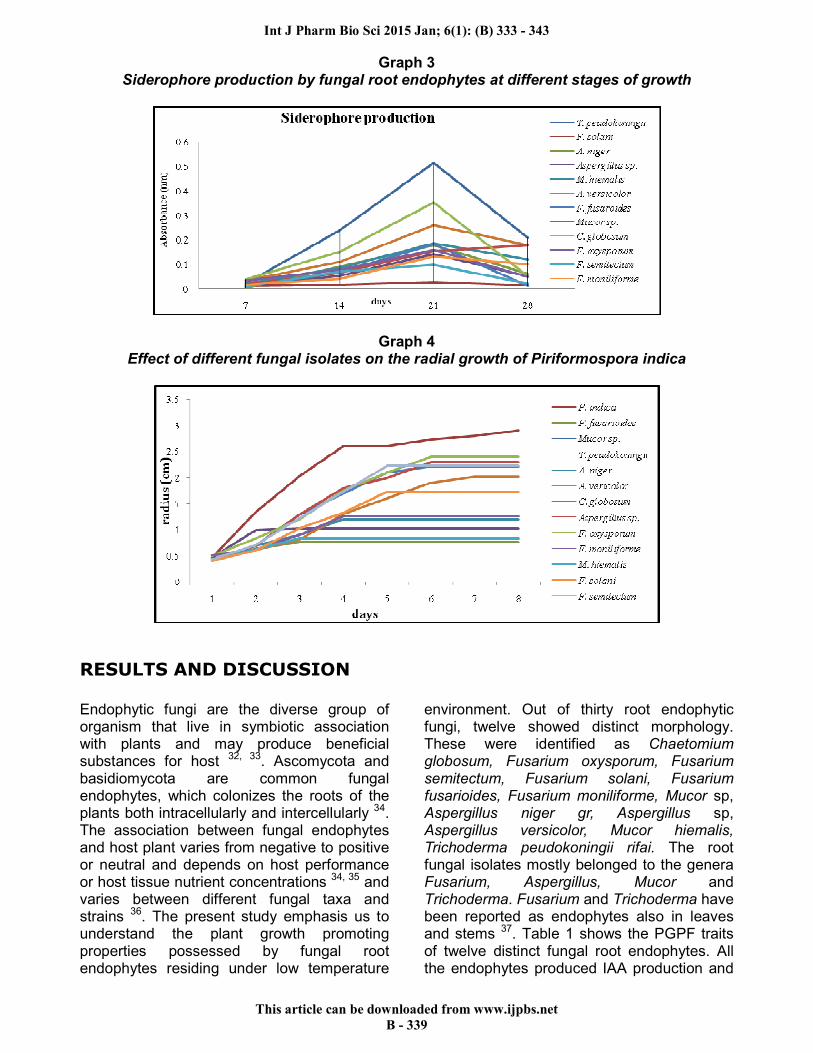
Graph 3 Siderophore production by fungal root endophytes at different stages of growth
Graph 4 Effect of different fungal isolates on the radial growth of Piriformospora indica
RESULTS AND DISCUSSION
Endophytic fungi are the diverse group of organism that live in symbiotic association with plants and may produce beneficial substances for host 32, 33. Ascomycota and basidiomycota are common fungal endophytes, which colonizes the roots of the plants both intracellularly and intercellularly 34. The association between fungal endophytes and host plant varies from negative to positive or neutral and depends on host performance or host tissue nutrient concentrations 34, 35 and varies between different fungal taxa and strains 36. The present study emphasis us to understand the plant growth promoting properties possessed by fungal root endophytes residing under low temperature
environment. Out of thirty root endophytic fungi, twelve showed distinct morphology. These were identified as Chaetomium globosum, Fusarium oxysporum, Fusarium semitectum, Fusarium solani, Fusarium fusarioides, Fusarium moniliforme, Mucor sp, Aspergillus niger gr, Aspergillus sp, Aspergillus versicolor, Mucor hiemalis, Trichoderma peudokoningii rifai. The root fungal isolates mostly belonged to the genera Fusarium, Aspergillus, Mucor and Trichoderma. Fusarium and Trichoderma have been reported as endophytes also in leaves and stems 37. Table 1 shows the PGPF traits of twelve distinct fungal root endophytes. All the endophytes produced IAA production and

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 340
solubilize phosphate. Seven were found positive for siderophore, four for HCN and three for ammonia production. Nahas (1996) reported that among the microbial group, fungi are more efficient in solubilizing phosphate than bacteria 38. All the endophytic fungi exhibited clear zone of phosphate solubilization around the fungal growth in Pikovskaya’s medium supplemented with tri calcium phosphate (Fig 1a). The quantitative estimation of phosphate solubilization for 5 days revealed a similar trend in all the endophytic fungi. The phosphate solubilization increased from day 2 to day 8 and then decreases slowly. The highest range was observed in T. peudokoningii (37.45±2.78 to 64.32±2.87) µg/ml followed by C. globosum (33.62±5.92 to 69.32±3.21) µg/ml, F. semitectum (32.64±1.89 to 57.63±2.11) µg/ml, A. versicolor (31.63±2.02 to 63.72±2.36) µg/ml (Graph 1). A similar finding has been reported on Pseudomonas striata 39 and on two different Penicillium sp 40, where they observed a gradual increase in the available P level in the medium up to a certain period and then available P level drops off. The mechanism behind the solubilization of phosphate could be the production of organic acids and a drop in pH 41, 42,43. P solubilization occurs when the media is supplemented with a simple carbon source along with phosphate solubilizing fungi. This also results in the production of various organic acids and other substances 44. All twelve endophytic fungi were screened for IAA production with tryptophan (5mg/ml) and the intensity of red color varied among all the fungal endophytes (Graph 2). F. fusarioides (49.21±3.5)µg/ml showed maximum IAA production followed by Aspergillus sp. (35.08±2.34) µg/ml and M. hiemalis (28.52±1.83) µg/ml. The other showed IAA comparatively in lesser quantity – C. globosum (26.78±1.11) µg/ml, T. peudokoningii (24.12±1.6) µg/ml, A. versicolor (20.85±1.43) µg/ml and F. moniliforme (20.54±1.1) µg/ml. C. globosum and other fungi produce IAA production through the IAA biosynthesis pathway. All the fungal endophytes were subjected to siderophore production. Seven showed siderophore production on on solid CAS agar media (Fig 1b). It was seen that the maximum intensity of siderophore production was shown by C.
globosum, A. vericolor and T. peudokoningii followed by F. oxysporum, Mucor sp, A. niger and M. hiemalis. However, F. semitectum, F. solani, F. fusarioides, F. moniliforme and Aspergillus sp did not produce siderophores. The quantitative estimation of siderophore production through FeCl3 test in Fries basal medium revealed that all the isolates showed maximum siderophore production at 21th day of the inoculation and the concentration decreases afterwards (Graph 3). The maximum concentration was seen in T. peudokoningii followed by C. globosum and A. versicolor. However, the siderophore production was found almost negligible in all Fusarium species. Similar findings were also reported in F. oxysporum 45. The rhizospheric organisms produce siderophores through several factors, one of which can be high iron concentrations in the soil 46. A. niger does not produce siderophore but is a good producer of citric and other organic acids that are thought to play a role in the reaction with CAS reagent 47. Siderophore production by A. niger and A. flavus in both solid and broth media is well supported by many workers. All the isolates were checked for HCN and ammonia production. Four showed postive results for HCN production and three for ammonia production. C. globosum, F. fusarioides, Mucor sp. and A. versicolor were found positive for HCN production and resulted in the filter paper color change from yellow to brown after incubation (Fig 1c). M. hiemalis, A. niger and F. moniliforme showed positive results for ammonia production and resulted in the development of a yellow to brown color onto reaction with Nesselers reagent. (Table 1) Chaetomium is a large genera belonging to family Chaetomiaceae (Ascomycota). It is a source of important bioactive secondary metabolites. More than 200 compounds have been reported from this genus 48. T. harzianum t – 22 and its cell-free culture filtrates solubilizes insoluble rock phosphate containing calcium phosphate and micronutrients like Mn and Zn49. For this reason, it has been hypothesized that the fungus can be applied as growth enhancement on various crops. In field plots, the beneficial action of F. oxysporum on AM fungi resulted in an increase in the growth and nutrient concentration during the early stage of

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 341
root colonization in pea plants. This has been hypothesized by many workers 50, 51. Microscopic examinations of dual culture assay between fungal endophytes and P. indica shows an alteration in the mycelium of P. indica where it was in contact with the fungal endophytes (Fig 2). The interaction of fungal root endophytes with a known root endophyte, P. indica revealed that none of the isolated endophyte showed mutual interaction and the radial growth of P. indica incorporated with endophytes was drastically reduced (Graph 4). The experiment indicated that P. indica interacts with a diverse group of soil fungi and its interaction varied from the negative to positive association.
CONCLUSION Plant growth promoting fungi (PGPF) are closely associated with plant roots and have been reported in increasing the biomass and yield as well as modulate the nutrient uptake and protect the host plant from pathogen attack. The growth activity of PGPF is due to its colonization with the roots and concerting the inorganic minerals into organic forms that are rapidly taken up by the plants. This study
sheds light on the potential ability of some root endophytic fungi isolated from extreme cold conditions of Central Himalayas as plant growth promoters. However, further studies are required to establish the exact contribution by root endophytic fungi as PGPF organisms and as growth promoters. The geographical location represents a unique ecological niche for the isolation of root endophytic fungi from the tomato roots habituating that particular area. The beneficial microbes along with the crop evolve and adapts to the prevailing climatic conditions. The analysis of endophytic fungi of plants roots is an important step in the selection of new isolates and their survival under stress conditions.
ACKNOWLEDGEMENT
The work was supported by the Defence Institute of Bioenergy Resources, DRDO, Haldwani. We are also thankful to Dr. S.K. Singh from Agarkar Research Institute, Fungal Identification Service, Mycology and Plant Pathology Group Agharkar Research Institute, G, G Agarkar Road, Pune for identifying the fungal isolates.
REFERENCES
1. Petrini O, Sieber TH, Toti L, Viret O,
Ecology, metabolite production, and substrate utilization in endophytic fungi. Natural Toxins, 1: 185 – 196, (1992).
2. Gaylord ES, Preszler RW, Boecklen WJ, Interactions between host plants, endophytic fungi, and a phytophagous in- sect in an oak (Quercus grisea Quercus gambelii) hydrid zone. Oecologia, 105: 336 – 342, (1996).
3. Fisher PJ, Petrini LE, Sutton BC, Petrini O, A study of fungal endophytes in leaves, stem and roots of Gynoxis oleifolia Muchler (Compositae) from Ecuador. Nova Hedwigia, 60 (4): 589 - 594, (1995).
4. Arnold AE, Lutzoni F, Diversity and host range of foliar fungal endophytes: are tropical trees biodiversity hot spots? Ecol, 88: 541 – 549, (2007).
5. Suryanarayanan TS, Thirunavukkarasu N, Hariharan GN, Balaji P, Occurrence of non-obligate micro fungi inside lichen thalli. Sydowia 57: 120 – 130, (2005).
6. Šraj-Kržič N, Pongrac P, Klemenc M, Kladnik A, Regvar M, Gaberščik A, Mycorrhizal colonisation in plants from intermittent aquatic habitats. Aquat Bot, 85: 331 – 336, (2006).
7. Pandey A, Palni LMS, The rhizosphere effect in trees of the Indian Central Himalaya with special reference to altitude. App Ecol and Env Res, 5(1): 93 - 102, (2007).
8. Clay K, Schardl CL, Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. American Nat, 160: S99 – S127, (2002).
9. Chadha N, Mishra M, Prasad R, Varma A, Root endophytic fungi: Research update. Journal of Biology and Life Science, USA, 5 (2): 135 - 158,(2014). doi:10.5296/jblsjbls.v5i2.59605960
10. Malla R, Prasad R, Giang PH, Pokharel U, Oelmueller R, Varma A, Characteristic features of symbiotic fungus Piriformospora indica. Endocytobiosis and Cell Research,

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 342
15: 579 - 600, (2004). 11. Wakelin SA, Warren RA, Harvey PR, Ryder
MH, Phosphate solubilization by Penicillium spp. closely associated with wheat roots. Biology and Fertility of Soils, 40: 36 - 43, (2004).
12. Costa JM, Loper JE, Characterization of siderophore production by the biological control agent Enterobacter cloacae. Molecular Plant-Microbe Interaction, 7: 440 – 448, (1994).
13. Ramamoorthy V, Viswanathan R, Raguchander T, Prakasam V, Samiyappan R, Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Protection, 20: 1 - 11, (2001).
14. J.W. Kloepper, R.M. Zablotowicz, E.M. Tipping, R. Lifshitz. Plant growth promotion mediated by bacterial rhizosphere colonizers. In: D.L. Keister, and P.B. Cregan (eds.), The rhizosphere and plant growth, Kluwer Academic Publishers, Dordrecht, The Netherlands,pp.315-326,1991.
15. Bhowmik D, Kumar KPS, Shravan Paswan S, Srivastava S, Tomato-A Natural Medicine and Its Health Benefits. J of Pharmacognosy and Phytochem, 1 (1): 33 - 43, (2012).
16. Meera MS, Shivanna MB, Kageyama K, Hyakumachi M, Persistance of induced systemic resistance in cucumber inrelation to root colonization by plant growth promoting fungal isolates. Crop Protection, 14: 123 – 130, (1995).
17. Munoz Z, Moret A, Garces S, The use of Verticillium dahilae and Diplodia scrobiculata to induce resistance in Pinus halepensis against Diplodia pinea infection. Eur J Pl Pathol, 120: 331 – 337, (2008).
18. Khalid A, Arshad M, Zahir ZA, Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J Appl Microbiol, 96 (3): 473 – 480, (2004).
19. Wu SC, Caob ZH, Lib ZG, Cheunga KC, Wong MH, Effects of biofertilizer containing N-fixer, P and K solubilizers and AM fungi on maize growth: a greenhouse trial. Geoderma, 125: 155 – 166, (2005).
20. Pandey A, Durgapal A, Joshi M, Palni LMS, Influence of Pseudomonas corrugata inoculation on root colonization and growth promotion of two important hill crops. Microbiol Res, 154(3): 259 - 266, (1999).
21. P. Trivedi, B. Kumar, A. Pandey, and L.M.S.
Palni, Growth promotion of rice by phosphate solubilizing bioinoculants in a Himalayan location, in Plant and Soil, Developments in Plant and Soil Sciences. In: E. Velazqez, and C. Rodriguez-Barrueco (eds.), First International Meeting on Microbial Phosphate Solubilization, Springer, pp.291-299,2007.
22. P. Trivedi, A. Pandey, and L.M.S. Palni, Bacterial inoculants for field applications under Mountain Ecosystem: present initiatives and future prospects, in Bacteria in Agrobiology. In: D.K. Maheshwari (ed.), Plant Probiotics, Springer, pp.15-44, 2012.
23. Kumar V, Singh P, Jorquera MA et al, Isolation of phytase-producing bacteria from Himalayan soils and their effect on growth and phosphorus uptake of Indian mustard (Brassica juncea),” World Journal of Microbiol and Biotechnol, 29 (8): 1361 – 1369, (2013).
24. AC Gour, Ed. Phosphate solubilizing microorganisms as biofertilizers. Omega scientific publishers, New Delhi: 16 - 72, (1990).
25. Jackson ML, Ed. Soil Chemical Analysis, Prentice Hall of India Pvt Ltd, New Delhi: (1973).
26. Brick JM, Bostock RM, Silverstone SE, Rapid in situ assay for indole acetic acid production by bacteria immobilized on nitrocellulose membrane. Appl Environ Microbial, 57: 535 – 538, (2004).
27. Lorck H, Production of hydrocyanic acid by bacteria. Plant Physiol, 1: 142 – 146, (2004).
28. Schwyn B, Neilands JB, Universal Chemical Assay for the Detection and Determination of Siderophores. Anal Biochem, 160: 47 - 56, (1987).
29. Letendre ED, Gibbons WA, Isolation and purification of canadaphore, a siderophore produced by Helminthosporium carbonum. Biochemical and Biophysical Research Communications, 129: 262 - 276, (1991).
30. JC Cappuccino and N Sherman, Ed. In: Microbiology: A Laboratory Manual, 3rd Edn, Benjamin/Cummings publisher, New York: 125 – 179, (1992).
31. Fokkema NJ, Fungal antagonism in the phylosphere. Ann Appl Biol, 89: 115 - 117, (1978).
32. Weber J, A natural control of Dutch elm disease. Nature, London, 292: 449 – 451, (1981).

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 343
33. Shiomi HF, Silva HSA, De Melo IS, Nunes FV, Bettiol W, Bioprospecting endophytic bacteria for biological control of coffee leaf rust, Sci Agric, 63 (1): 32 - 39, (2006).
34. Jumpponen A, Dark septate endophytes: are they mycorrhizal? Mycorrhiza, 11: 207 – 211, (2001).
35. Declerck S, Strullu DG, Fortin JA, Eds. In Vitro Culture of Mycorrhizas. Springer-Verlag, Berlin Heidelberg: pp. 388, (2005).
36. Jumpponen A, Trappe JM, Dark-septate root endophytes: a review with special reference to facultative biotrophic symbiosis. New Phytologist, 140: 295 – 310, (1998).
37. Shoresh M, Harman GE, Mastouri F Induced systemic resistance and plant responses to fungal biocontrol agents. Annu Rev Phytopathol, 48: 1 – 23, (2010).
38. E. Nahas, Factors determining rock phosphate solubilization by microorganism isolated from soil. World J Microbiol Biotechnol, 12: 567 – 572, (1996).
39. Gaur AC, Phosphate solubilizing microorganism as biofertilizer, Omega Scientific Publications, New Delhi, India, (1990).
40. Nath R, Sharma GD, Barooah M, Efficiency of Tricalcium Phosphate Solubilization by Two Different Endophytic Penicillium sp. Isolated from Tea (Camellia sinensis L.). Euro J of Exper Biol, 2 (4): 1354 - 1358, (2012).
41. Illmer P, Schineer F, Solubilization of inorganic calcium phosphate solubilization mechanisms. Soil Biol Biochem, 27: 257 – 263, (1995).
42. Liu TS, Lee LY, Tai CY, Hung CH, Chang YS, Wolfram JH, Rogers R, Goldstein AH, Cloning of an Erwinia carotovora gene necessary for gluconic acid production and enhanced mineral phosphate solubilization in Escherichia coli HB101: Nucleotide sequence and probable involvement in biosynthesis of the coenzyme pyrroloquinoline quinone. J Bacteriol, 174: 5814 – 5819, (1992).
43. Kim KY, Jordan D, Krishnan HB, Rahnella aqualitis, bacterium isolated from soybean
rhizosphere, can solubilize hydroxyapatite. FEMS Microbiol Lett, 153: 273 – 277, (1997).
44. Johri BN, Sharma A, Virdi JS, Rhizobacterial diversity in India and Its Influence on Soil and Plant Health. Adv Biochem Engin/Biotechnol, 84: 49 – 89, (2003).
45. Wojesundera RLC, Amarasekera AS, Yapa DND, Production of siderophore by the fungus Fusarium oxysporum. J Natn Sci Coun Sri Lanka, 23 (3): 97 – 100, (1995).
46. Machuca A, Milagres AMF, Use of CAS-agar plate modified to study the effect of different variables on the siderophore production by Aspergillus, Letters in Appl Microbiol, 36: 177–181, (2003).
47. M. Roehr, C.P. Kubicek, J. Kominek. Industrial acids and other small molecules. In: J.B. Bennett, and Klich (eds.), In Aspergillus. Biology and Industrial Applications, Stoneham, USA: Butterworth-Heinemann,pp.91–153,1992.
48. Li HQ, Li XJ, Wang YL, Zhang Q, Zhang AL, Gao JM, Zhang XC, Antifungal metabolites from Chaetomium globosum, an endophytic fungus in Ginkgo biloba. Biochem Sys Eco, 39: 876 - 879, (2011).
49. Altomare C, Norvell WA, Bjorkman T, Harman GA, Solubilization of phosphates and micronutrients by the plant-growth promoting and biocontrol fungus Trichoderma harziamum Rifai 1295-22, Appl Env Microbiol, 65: 2926 - 2933, (1999).
50. Fracchia S, Mujica MT, García-Romera I, García-Garrido JM, Martín J, Ocampo JA, Godeas A, Interactions between Glomus mosseae and arbuscular mycorrhizal sporocarp- associated saprophytic fungi. Plant Soil, 200: 131 – 137, (1998).
51. García-Romera I, García-Garrido JM, Martín J, Fracchia S, Mujica MT, Godeas A, Ocampo JA, Interaction between saprophytic Fusarium strain and arbuscular mycorrhizas of soyabean plants. Symbiosis, 24: 235 – 246, (1998).

Int J Pharm Bio Sci 2015 Jan; 6(1): (B) 333 - 343
This article can be downloaded from www.ijpbs.net
B - 344


















