Interactions between Microsomal Triglyceride Transfer Protein and ...
10
Interactions between Microsomal Triglyceride Transfer Protein and Apolipoprotein B within the Endoplasmic Reticulum in a Heterologous Expression System* (Received for publication, December 8, 1995, and in revised form, May 15, 1996) Shailendra B. Patel‡§¶ and Scott M. Grundy‡§i** From the ‡Center for Human Nutrition, Departments of §Internal Medicine, iClinical Nutrition, and **Biochemistry, University of Texas Southwestern Medical Center, Dallas, Texas 75235-9052 When apolipoprotein B (apoB) is expressed in heter- ologous cells, it is not secreted but retained and de- graded within the endoplasmic reticulum (ER). We have previously characterized carboxyl-terminal truncated forms of apoB expressed in COS cells and have shown that these proteins were readily synthesized but re- tained within the ER and degraded, if the size of the truncated protein was larger than apoB 29. Below this size, the smaller the size of the apoB truncates, the greater the extent of secretion, although >50% of these smaller proteins were also degraded within the ER. In the present study, we demonstrate that this secretory defect can be overcome by coexpression with microso- mal triglyceride transfer protein (MTP); moreover, this complementation is inversely related to the size of apoB. Secretion of apoBs larger than B29 required the coex- pression of MTP and, in the presence of MTP, was oleate- responsive. MTP, in the presence or absence of oleate supplementation, had little or no effect on the secretion of the shorter truncates. We discovered, however, that MTP was physically associated with all forms of apoB intracellularly (B13–B41). The association of MTP with apoB 41 was stable to high salt washing, as well as to low pH, suggesting that these interactions may be hydro- phobic in nature. In addition to the interaction with MTP, apoB was also found to be associated with cal- nexin, confirming previous studies, and with proteins bearing the KDEL retention signal. However, studies on overexpression of human calnexin and tunicamycin in- hibition of glycosylation showed that interaction with calnexin was not necessary for the formation or secre- tion of apoB 41-containing lipoproteins; moreover, in the presence of MTP, the association of calnexin with apoB 41 was transient or absent. These data suggest that for apoB to attain a folded state sufficient to escape the quality control of the ER, it needs to obtain neutral lipid (supplied by MTP), as well as its ability to keep it pack- aged as a rudimentary lipoprotein, dependent on its size being larger than B29. The ER 1 plays a pivotal role in the fate of a nascent protein. All secretory and transmembrane proteins must, in general, attain some structural maturity to escape the “quality control” of the ER. Inability to attain “maturity” within the ER results in retention and degradation (1– 4). ApoB must undergo a se- ries of conformational changes that would allow it to proceed through the ER. This step appears to be rate-limiting, since up to 80% of newly synthesized apoB 100 is normally degraded (5– 8), a process that seems inefficient, given it takes approxi- mately 10 min to fully synthesize this 550-kDa apoB 100 pro- tein (9, 10) . For a complex secretory protein, such as apoB, maturation involves not only the constitutional machinery of the ER to impart a structure allowing it to proceed onto the Golgi but also specialized chaperones that are necessary for this progression. This requirement is evidenced by the autosomal recessive dis- orders of abetalipoproteinemia and chylomicron retention dis- ease (11); in both of these cases, apoB synthesis appears to be normal, but there is no secretion of apoB from the appropriate tissues. For abetalipoproteinemia, the genetic defect has been identified, cloned, and shown to involve a protein complex, consisting of the microsomal triglyceride transfer protein (MTP) and protein disulfide isomerase (PDI), that transfers neutral lipid into the core of the nascent lipoprotein particle (12, 13). For chylomicron retention disease, no chromosomal localization or candidate genes have so far been identified, although it is known that both the MTP and apoB genes are not the sites of defects (13–15) . A considerable amount has already been learned about the early biogenesis of apoB-containing lipoproteins, based on in vitro studies. A model of biogenesis of has emerged whereby apoB is cotranslationally inserted into the ER membrane, where it follows one of two fates; it is either inserted into the inner leaflet of the ER membrane, or it may begin to form a rudimentary lipoprotein particle cotranslationally and is trans- located directly into the ER lumen (6, 16 –18). This latter par- ticle is destined for secretion and may represent the “constitu- tive” pathway. apoB inserted into the inner ER membrane, in contrast, may be destined for ER degradation. It is unclear whether this pool of ApoB is diverted to the secretory pathway, at the expense of degradation, when the cells are supplemented with oleate to increase secretion (19). Very few of the specific ER proteins involved in controlling the fate of apoB in the ER have been characterized, although it is known that apoB is one of many de novo synthesized glycoprotein proteins that interact * This work was supported by unrestricted grants from Merck & Co., Inc., West Point, PA; Bristol-Myers Squibb, New Brunswick, NJ; the Southwestern Medical Foundation, Dallas, TX; and the Moss Heart Foundation, Dallas TX; and by National Institutes of Health Grant HL-29252. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. ¶ To whom correspondence should be addressed: Department of In- ternal Medicine, University of Texas Southwestern Medical Center, Y3.208, 5323 Harry Hines Blvd., Dallas, TX 75235-9052. Tel.: 214-648- 8734; Fax: 214-648-8734; E-mail: [email protected]. 1 The abbreviations used are: ER, endoplasmic reticulum; apoB, apo- lipoprotein B; MTP, microsomal triglyceride transfer protein; PDI, protein disulfide isomerase; CHAPS, 3-[(3-cholamidopropyl)dimethyl- ammonio]-1-propanesulfonic acid; PAGE, polyacrylamide gel electro- phoresis; BiP, binding protein; grp, glucose-regulated protein. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 271, No. 31, Issue of August 2, pp. 18686 –18694, 1996 © 1996 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. 18686 by guest on March 26, 2018 http://www.jbc.org/ Downloaded from
Transcript of Interactions between Microsomal Triglyceride Transfer Protein and ...

Interactions between Microsomal Triglyceride Transfer Protein andApolipoprotein B within the Endoplasmic Reticulum in aHeterologous Expression System*
(Received for publication, December 8, 1995, and in revised form, May 15, 1996)
Shailendra B. Patel‡§¶ and Scott M. Grundy‡§i**
From the ‡Center for Human Nutrition, Departments of §Internal Medicine, iClinical Nutrition, and **Biochemistry,University of Texas Southwestern Medical Center, Dallas, Texas 75235-9052
When apolipoprotein B (apoB) is expressed in heter-ologous cells, it is not secreted but retained and de-graded within the endoplasmic reticulum (ER). We havepreviously characterized carboxyl-terminal truncatedforms of apoB expressed in COS cells and have shownthat these proteins were readily synthesized but re-tained within the ER and degraded, if the size of thetruncated protein was larger than apoB 29. Below thissize, the smaller the size of the apoB truncates, thegreater the extent of secretion, although >50% of thesesmaller proteins were also degraded within the ER. Inthe present study, we demonstrate that this secretorydefect can be overcome by coexpression with microso-mal triglyceride transfer protein (MTP); moreover, thiscomplementation is inversely related to the size of apoB.Secretion of apoBs larger than B29 required the coex-pression ofMTP and, in the presence ofMTP, was oleate-responsive. MTP, in the presence or absence of oleatesupplementation, had little or no effect on the secretionof the shorter truncates. We discovered, however, thatMTP was physically associated with all forms of apoBintracellularly (B13–B41). The association of MTP withapoB 41 was stable to high salt washing, as well as to lowpH, suggesting that these interactions may be hydro-phobic in nature. In addition to the interaction withMTP, apoB was also found to be associated with cal-nexin, confirming previous studies, and with proteinsbearing the KDEL retention signal. However, studies onoverexpression of human calnexin and tunicamycin in-hibition of glycosylation showed that interaction withcalnexin was not necessary for the formation or secre-tion of apoB 41-containing lipoproteins; moreover, inthe presence of MTP, the association of calnexin withapoB 41 was transient or absent. These data suggest thatfor apoB to attain a folded state sufficient to escape thequality control of the ER, it needs to obtain neutral lipid(supplied by MTP), as well as its ability to keep it pack-aged as a rudimentary lipoprotein, dependent on its sizebeing larger than B29.
The ER1 plays a pivotal role in the fate of a nascent protein.All secretory and transmembrane proteins must, in general,attain some structural maturity to escape the “quality control”of the ER. Inability to attain “maturity” within the ER resultsin retention and degradation (1–4). ApoB must undergo a se-ries of conformational changes that would allow it to proceedthrough the ER. This step appears to be rate-limiting, since upto 80% of newly synthesized apoB 100 is normally degraded(5–8), a process that seems inefficient, given it takes approxi-mately 10 min to fully synthesize this 550-kDa apoB 100 pro-tein (9, 10) .For a complex secretory protein, such as apoB, maturation
involves not only the constitutional machinery of the ER toimpart a structure allowing it to proceed onto the Golgi but alsospecialized chaperones that are necessary for this progression.This requirement is evidenced by the autosomal recessive dis-orders of abetalipoproteinemia and chylomicron retention dis-ease (11); in both of these cases, apoB synthesis appears to benormal, but there is no secretion of apoB from the appropriatetissues. For abetalipoproteinemia, the genetic defect has beenidentified, cloned, and shown to involve a protein complex,consisting of the microsomal triglyceride transfer protein(MTP) and protein disulfide isomerase (PDI), that transfersneutral lipid into the core of the nascent lipoprotein particle(12, 13). For chylomicron retention disease, no chromosomallocalization or candidate genes have so far been identified,although it is known that both the MTP and apoB genes are notthe sites of defects (13–15) .A considerable amount has already been learned about the
early biogenesis of apoB-containing lipoproteins, based on invitro studies. A model of biogenesis of has emerged wherebyapoB is cotranslationally inserted into the ER membrane,where it follows one of two fates; it is either inserted into theinner leaflet of the ER membrane, or it may begin to form arudimentary lipoprotein particle cotranslationally and is trans-located directly into the ER lumen (6, 16–18). This latter par-ticle is destined for secretion and may represent the “constitu-tive” pathway. apoB inserted into the inner ER membrane, incontrast, may be destined for ER degradation. It is unclearwhether this pool of ApoB is diverted to the secretory pathway,at the expense of degradation, when the cells are supplementedwith oleate to increase secretion (19). Very few of the specificER proteins involved in controlling the fate of apoB in the ERhave been characterized, although it is known that apoB is oneof many de novo synthesized glycoprotein proteins that interact
* This work was supported by unrestricted grants from Merck & Co.,Inc., West Point, PA; Bristol-Myers Squibb, New Brunswick, NJ; theSouthwestern Medical Foundation, Dallas, TX; and the Moss HeartFoundation, Dallas TX; and by National Institutes of Health GrantHL-29252. The costs of publication of this article were defrayed in partby the payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
¶ To whom correspondence should be addressed: Department of In-ternal Medicine, University of Texas Southwestern Medical Center,Y3.208, 5323 Harry Hines Blvd., Dallas, TX 75235-9052. Tel.: 214-648-8734; Fax: 214-648-8734; E-mail: [email protected].
1 The abbreviations used are: ER, endoplasmic reticulum; apoB, apo-lipoprotein B; MTP, microsomal triglyceride transfer protein; PDI,protein disulfide isomerase; CHAPS, 3-[(3-cholamidopropyl)dimethyl-ammonio]-1-propanesulfonic acid; PAGE, polyacrylamide gel electro-phoresis; BiP, binding protein; grp, glucose-regulated protein.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 271, No. 31, Issue of August 2, pp. 18686–18694, 1996© 1996 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
18686
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

with calnexin (20) .The identification of MTP has been a majoradvance in this early process; Gordon et al. (22) have shownthat MTP expression can overcome the secretory block on apoB53 (using the centile numbering (21), full-length is apoB 100,550 kDa) secretion from HeLa cells, and preliminary findingsby Leiper et al. (23) have shown that coexpression of apoB 41and MTP in COS cells can also lead to apoB 41 secretion (23).To investigate the early steps involved in apoB secretion, we
have characterized heterologous expression of carboxyl-termi-nal truncated apoBs in COS cells. Previously, we showed thatwhen apoB 41 is expressed in COS cells, it was fully synthe-sized, N-glycosylated, but was then degraded in the ER or anER-related compartment (24). Expression of apoBs that wereprogressively truncated led to some secretion of the smallertruncates. Others have reported similar findings in various cellsystems; for example, expression of apoB truncates in COScells led to their intracellular degradation (7), apoB 17 wassecreted from mouse mammary tumor line C-127 (25), andexpression of apoB 53 in CHO cells resulted in a rapid degra-dation in the ER, a process that could be inhibited by a proteaseinhibitor, calpain I (26). Furthermore, coexpression of apoBand MTP in HeLa (22) or COS cells (23) abrogates this ERretention and degradation, resulting in secretion of a lipopro-tein particle in the high density liproprotein range. These stud-ies suggest that the ability of non-hepatic/non-intestinal cellsto secrete apoB can be conferred solely by the coexpression ofMTP. We have utilized this approach to characterize this sys-tem further and suggest that heterologous expression may beuseful in defining some of the early events in apoB-containinglipoproteins.In the present study, we have investigated the interaction of
MTP with the various sizes of apoB carboxyl-truncates to de-termine whether expression of MTP can affect secretion ofsmall apoB proteins that are incapable of forming lipoproteins(27, 28). We have characterized the nature of interaction ofMTP with these truncates. Finally, we have studied what rolethat resident ER proteins, such as calnexin, may play in thebiogenesis of apoB-containing lipoproteins.
EXPERIMENTAL PROCEDURES
Reagents—Tissue culture medium, sheep antibodies to apoB, andreagents were obtained as described previously (24). Rabbit antiserumto MTP (13) was a gift from Dr. John Wetterau (Bristol-Myers Squibb)and mouse antibody, A139, to human calnexin was a gift from Dr.Michael Brenner (29). Although the rabbit antiserum to MTP has beenwell characterized for Western blotting to detect MTP, we have foundthat it can also immunoprecipitate MTP, although this may not bequantitative. Rabbit antisera to canine calnexin and mouse monoclonalantibody to peptide KSEKDEL (clone 10C3, recognizes BiP/grp 78, grp94, and HSP 40) (30) were obtained from StressGen BiotechnologiesCorp. (Victoria, British Columbia, Canada). Mouse monoclonal anti-body against human apoB, C1.4 (31), was a gift from Dr. GustavSchonfeld. All other chemicals were of reagent grade or better.DNA Constructs—The pSV7D-based expression plasmids containing
carboxyl-truncated apoB fragments are as described previously (24, 32).The expression clones for MTP (under agreement from Bristol-MyersSquibb, NJ) and calnexin (gift from Dr. Brenner) are as describedpreviously (29), except the MTP was driven by the Rous sarcoma viruspromoter in the vector pRSV.Neo (Invitrogen Corp.).Cell Culture, Transfection, Metabolic Labeling, and Immunoprecipi-
tation—COS-1 cells, maintained in DMEM supplemented with 10%FCS, were transfected, and quantitative pulse-chase analyses werecarried out as described previously (24), except 10 mg of each DNA forcotransfection was used unless otherwise stated. In all experimentswhere treatment with inhibitors or oleate was performed, the controlpool of cells was always derived from the same pool of cells as thetreated cells, maintaining identical transfection efficiencies. Briefly,cell lysis was performed in 150 mM NaCl, 50 mM Tris, pH 7.5, 5 mM
EDTA, and 1% Triton X-100. The cell lysates were cleared of cell debrisby centrifugation at 13,000 3 g for 15 min at 4 °C; total protein con-centrations were determined using a dye-binding assay (Bio-Rad) and
immunoprecipitated with the appropriate antibodies, collected by pro-tein A-Sepharose beads and analyzed as described previously (24).Sodium phosphate buffers were used for the investigation of the effectsof pH on MTPzapoB complexes at a final concentration of 100 mM.Immunoprecipitates were incubated at room temperature for 5 minbetween each wash. For calnexin immunoprecipitations, 1% CHAPSwas substituted for Triton X-100. In the presence of Triton X-100,calnexin-associated proteins appeared to be less stably bound (data notshown).2
For tunicamycin inhibition, 24 h after transfection, the cells weretreated with 0.02% DMSO or 0.02% DMSO and 4 mg/ml tunicamycin(final concentrations). This treatment resulted in greater than 90%inhibition of labeled glucosamine incorporation into total cell acid-precipitable material, as well as into immunoprecipitated apoB 41 (datareviewed but not shown). Immunoprecipitation and SDS-PAGE wasperformed essentially as described previously. Quantitation was per-formed by exposure of the dried gels to PhosphorImager screens andanalysis using a PhosphorImager (Molecular Dynamics, Sunnyvale,CA).For pulse labeling with [3H]oleate (specific activity, 60 Ci/mmol;
Moravek Biochemicals, Brea, CA), transfected cells were incubated for18 h in labeled oleate at 83.3 mCi/ml, the medium was cleared free ofcellular debris, and subjected to cesium chloride density centrifugationand fractionated as described below. The density of each fraction wasdetermined as described previously, and total lipids were extracted bythe Folch method (33), concentrated under a nitrogen stream, loadedonto thin layer silica gel plates, and chromatographed in hexane:eth-anol:acetic acid mixture (80:20:1); then the radiolabeled lipid specieswas detected by autoradiography, after spraying the plates with En-hance spray (DuPont NEN). Specific bands, identified by comparison tostandards, were scraped off the plates and quantitated by liquid scin-tillation counting.Cesium Chloride Density Gradient Centrifugation—Density gradient
ultracentrifugation was performed as described previously (24). ApoBwas either immunoprecipitated when radiolabeled protein was present,as above, or the samples were desalted and concentrated in Centricon30 filters (Amicon, Beverly, MA) separated by SDS-PAGE, and ana-lyzed by Western blotting, using C1.4 primary antibody.Western Blotting—Immunoprecipitated proteins from equal amounts
of cell lysates were separated by SDS-PAGE and electroblotted in 25mM Tris, pH 8.3, and 192 mM glycine with 20% methanol onto polyvi-nylidene difluoride membranes (Millipore Corp., Bedford, MA). Themembranes were blocked in 5% non-fat milk, incubated with primaryantibodies at a dilution of 1:1000, washed extensively, incubated withsecondary antibodies conjugated with horseradish peroxidase (Bio-Rad)at a dilution of 1:10,000, and washed; then the antibody conjugateswere detected by chemiluminescence using the Renaissance kit fromDuPont NEN as per kit instructions.
RESULTS
Effects of Coexpression of MTP and apoB on Secretion andOleate Responsiveness—This study was carried out to deter-mine whether the “secretory block” and lack of oleate respon-siveness reported on carboxyl-terminal truncated apoBs largerthan B29 (24) could be overcome by coexpression with MTP.Coexpression of apoB with MTP in COS cells resulted incomplementation of the secretory block for apoB truncates thatare larger than B29, and the rates of secretion were thenresponsive to oleate supplementation (Fig. 1). Coexpression ofMTP with apoB truncates smaller than B29 did not result inincreased secretion of these proteins (data not shown); further-more, oleate supplementation also had a small or no effect onthe secretory rates (Fig. 1). In contrast, supplementation ofoleate, in the presence of MTP, had the greatest effect on apoB41 (Fig. 1). Although there is variability due to transfectionefficiencies on expression of both cDNAs in individual cells, wehave found that oleate supplementation consistently increasedapoB 41 secretion from 10 6 2.4% (S.E.) to 30.0 6 6.1% (n 5 4),for apoB 29 from 14.7 6 2.4% to 22.6 6 3.5% (n 5 6), and forapoB 23 from 31.9 6 4.0% to 39.8 6 4.5% (n 5 7) during the 5-hchase period. Oleate and MTP had little or no effect on the
2 S. Rajagopalan, personal communication.
MTP and ApoB Interactions in the ER 18687
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

shorter truncates, apoB 13 (32.6 6 7.4% versus 33.1 6 3.1%,n 5 3) and apoB 17 (30.8 6 4.4% versus 28.7 6 3.8%, n 5 5).Although the data shown for apoB 23 (Fig. 1) would suggestthat oleate could have subtle effects, such as a reduction in therate of degradation of intracellular apoBs, this finding was notconsistent, and in the majority of the experiments, no signifi-cant differences were observed for the effects of oleate supple-mentation on the kinetics of degradation.Examination of the buoyant densities of apoBs secreted in
the presence of MTP showed that apoBs smaller than apoB 29were relatively lipid poor (Fig. 2), whereas newly secreted apoB41 was present at a density of around 1.15 g/ml. Secreted apoB13 was only found at a density of about 1.24 g/ml, suggesting itdid not contain any lipid.Buoyant Density and Lipid Profile of Secreted ApoB 41 Li-
poprotein—To examine the buoyant density of secreted apoB 41further, COS cells cotransfected with MTP and apoB 41, pulse-labeled and chased in medium containing no serum (Fig. 3, Aand B), 10% fetal calf serum (Fig. 3C), or 10% fetal calf serumsupplemented with 0.8 mM oleate (Fig. 3D) were studied. Fig.3B shows the effect of incubating secreted apoB 41, in theabsence of serum, in 10% bovine serum albuminz0.8 mM oleatecomplex for 2 h at 37 °C prior to density gradient centrifuga-tion, and fractionation. In the absence of oleate supplementa-tion of the cells, apoB 41 was secreted with a buoyant densityof around 1.15 g/ml, and secreted apoB 41 did not appear toobtain lipid directly from the medium, as shown by the mixingexperiment (Fig. 3B). Oleate supplementation led to the secre-
tion of apoB 41 particles of densities as low as 1.09 g/ml. Toexamine the lipid content of the secreted apoB 41 particles,cotransfected cells were pulse-labeled with [3H]oleic acid, andthe medium was subjected to density gradient centrifugationand fractionated. Lipid from each fraction was extracted by theFolch method (33) and analyzed by thin layer chromatography,as described under “Experimental Procedures.” Fig. 4A showsthe chromatograms of the labeled lipid fraction at a density ofabout 1.14 g/ml secreted from apoB 41/MTP or apoB 41 alonetransfected cells. The chromatograms shown are for fractions 4shown in Fig. 4B. In the presence of MTP, a large amount oftriglyceride was detected at buoyant densities where secretedapoB 41 is expected, but very little was seen in the controlmedium. Fig. 4B shows quantitation of the triglyceride profiles
FIG. 1. Effect of oleate supplementation on the kinetics ofsecretion of carboxyl-terminal truncates of apoB in the pres-ence of MTP expression. ApoB 13, 17, 23, 29, and 41 cDNAs werecotransfected with MTP into COS cells, pulse-labeled, and chased in thepresence (broken lines) or absence (continuous lines) of 0.8 mM oleate;apoB was immunoprecipitated quantitatively and analyzed by SDS-PAGE, as described under “Experimental Procedures.” The solid sym-bols represent apoB present in cell pellets, and the open symbols, thatin the media. MTP had little or no significant effect on the secretion ofapoBs ,B29. For apoB 41, in the presence of MTP, oleate supplemen-tation increased apoB 41 secretion by more than 3-fold (see text fordiscussion).
FIG. 2. Density gradient centrifugation of secreted apoB trun-cates. ApoB 13, 17, 23, 29, and 41 were cotransfected with MTP; thenmedium from cells was collected after 48 h, cleared of cellular debris,subjected to density gradient centrifugation, and fractionated. Thepresence of apoB detected by Western blotting. The various apoB spe-cies are as indicated. ApoB 41 attained a buoyant density of about 1.15g/ml (track 12), compared with 1.19 g/ml for apoB 29 (tracks 7–8) and anaverage density of 1.22 g/ml for apoBs 17 and 23 (tracks 4–5). ApoB 13blotted poorly on the result shown but remained at a density of 1.25g/ml in other experiments (data not shown).
FIG. 3. Density gradient centrifugation of secreted apoB 41.Cells cotransfected with apoB 41 and MTP were pulse-labeled andchased in medium containing no serum (A and B), 10% fetal calf serum(C), or 10% fetal calf serum and 0.8 mM oleate. Medium containinglabeled protein was subjected to cesium chloride density centrifugation,fractionated, apoB immunoprecipitated, and analyzed by SDS-PAGE asdescribed under “Experimental Procedures.” B, the effects of incubatingapoB 41, secreted in the absence of serum, with 10% bovine serumalbumin/0.8 mM oleate prior to density gradient centrifugation. As canbe seen, in the absence or presence of serum, apoB 41 had a peakbuoyancy in the high density lipoprotein (HDL1) range (1.125–1.210g/ml) of 1.15 g/ml (A, B, and C). Oleate supplementation of the cellsresulted in a more widespread spectrum of densities, with a significantamount of apoB now present in the high density lipoprotein (HDL2)range (1.063–1.1.125 g/ml), panel D.
MTP and ApoB Interactions in the ER18688
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

across the gradients. Fraction 1 is from the bottom of thedensity gradient centrifugation and contains some cellular de-bris not removed by the pre-spin of the medium. As can be seen,
there is a significant amount of triglyceride associated withfractions 3–6, in the density range 1.18–1.12 g/ml, where se-creted apoB 41 would be expected to float (see Figs. 2 and 3 forcomparison). Quantitation of the radioactivity migrating at thecholesterol ester position across the gradients did not show asignificant difference when compared to the control profile(data not shown).Hydrophobic Interactions between MTP and ApoB—In the
course of our pulse-chase studies, it was noted that, when MTPwas cotransfected with any of the apoBs (B13–B41), a proteinspecies coprecipitated with apoB; this was absent when MTPwas omitted (Fig. 5A). To exclude the possibility that the anti-bodies could cross-react with MTP or apoB, COS cells trans-fected with either apoB 41 or MTP alone were radiolabeled andimmunoprecipitated with the apoB and MTP antisera (Fig.5B). As can be seen, each of these antisera react only with their
FIG. 4. Analysis of the lipids secreted by COS cells in the pres-ence and absence of MTP. COS cells transfected with apoB 41 andMTP or apoB 41 alone (control) were incubated with [3H]oleate for 18 h,and medium was collected and subjected to density gradient centrifu-gation and fractionated. Total lipids were extracted from each fractionand analyzed by thin layer chromatography. The presence of labeledlipids was detected by fluorography, and specific lipid species werequantitated by liquid scintillation counting. A, the qualitative fluoro-graphic thin layer chromatography pattern for fractions 4 from apoB41/MTP and control medium. B, the quantitative triglyceride radioac-tivity profiles across all of the gradient fractions together with thedensity gradient profiles. Fraction 1 may contain some cellular debrisnot cleared by the initial pre-spin of the medium. The position ofmigration of the various lipid species on thin layer chromatography wasdetermined by comparison to known standards. As can be seen, there isa considerable amount of triacylglycerol that is present at the densitywhere apoB 41 is expected to float in the density range of 1.15–1.1 g/ml(B: fractions 4, 5, and 6). No difference on the patterns of the otherlabeled lipid species was noted, and in particular, quantitation of theradioactivity at the level of cholesterol ester migration did not show anydifferences between the two samples (data not shown).
FIG. 5. Coprecipitation of MTP with apoBs in COS cells. COScells were transfected with either apoB (A: tracks 2, 4, 6, and 8) or apoBand MTP (A: tracks 1, 3, 5, and 7) as indicated in the legend above thefigure, pulse-labeled for 10 min, chased for 1 h, and immunoprecipi-tated for apoB as described under “Experimental Procedures.” Equalamounts of radioactivity for tracks 1–8 were loaded on the gel. In thepresence of cotransfection with MTP (tracks 1, 3, 5, and 7), a radiola-beled band of the molecular weight of MTP coprecipitated with all of theapoB species. The migration of the apoB truncates is indicated by thearrowheads. The dye front, containing unresolved proteins at the bot-tom of the gel, is indicated by the presence of the radiolabeled material.A similar band of unresolved proteins at the dye front is noted insubsequent figures. Note that the coprecipitating ;98 kDa band islarger than apoB 13 or 17, excluding the possibility that this is adegradation product of apoB. To exclude the possibility that the anti-sera cross-react, COS cells were transfected with apoB 41 alone (B:tracks 1 and 2) or MTP alone (B: tracks 3 and 4) and immunoprecipi-tated with anti-apoB and anti-MTP antibodies. Anti-MTP antibodydoes not immunoprecipitate apoB (B: track 2), and apoB antiserum doesnot immunoprecipitate MTP (B: track 3).
MTP and ApoB Interactions in the ER 18689
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

appropriate antigen and do not show considerable cross-reac-tivity to other proteins.To uncover the identity of the protein coprecipitating with
apoB, immunoprecipitation from labeled cells as well as West-ern blotting was used (Fig. 6). Cells were cotransfected withapoB 41 and MTP, pulse-labeled, lysed, and immunoprecipi-tated for apoB (Fig. 6A, track 1) or MTP (Fig. 6B, track 2). Aband comigrating with MTP was present when apoB 41 wasimmunoprecipitated. A faint band comigrating with apoB 41was also present when MTP was specifically immunoprecipi-tated (Fig. 6A, track 2). The protein band above apoB 41 (Fig.6A, track 2) was also present when nonimmune serum wasused. Although a considerable amount of labeled MTP copre-cipitated with apoB (Fig. 6A, track 1), very little apoB copre-cipitated when MTP was immunoprecipitated (Fig. 6A, track2). In sequential immunoprecipitation studies, precipitationwith apoB antiserum appears to immunoprecipitate more MTPthan the rabbit antiserum against MTP (data not shown, butsee Fig. 6). The anti-MTP antibody has been used for Westernblotting previously but not for immunoprecipitation. Our stud-ies would suggest that this reagent can immunoprecipitate
MTP, but this may, therefore, not be used for quantitativepurposes. To further prove that MTP coprecipitated with apoBand vice versa, cell lysates from unlabeled cotransfected cellswere immunoprecipitated with anti-MTP antiserum (Fig. 6B,track 1) or with anti-apoB antiserum (Fig. 6B, track 2) andWestern blotted for the corresponding protein. ApoB 41 couldbe detected when MTP was immunoprecipitated, and MTPcould detected when apoB 41 was immunoprecipitated. In con-trol experiments where MTP was omitted or when nonimmunesera were used, no such coprecipitation was detected.To examine the nature of the association of MTP with apoB
41, immunoprecipitation with anti-apoB antiserum was per-formed, and the immunoprecipitates were washed in increas-ing concentration of salt. Fig. 7 shows a salt wash profile, whenimmunoprecipitation was performed in 150 mM NaCl (Fig. 7,track 1) and washed at various salt concentrations as indicated.The association of MTP with apoB 41 appeared to be unaffectedby salt; this finding was reproducible, even when initial lysiswas performed at the various salt concentrations indicated(data not shown). We have previously reported coprecipitationof an unidentified protein (;80 kDa) with apoB 41 from COScells (24). This protein is indicated by the asterisk (Fig. 7). Italso was resistant to high salt washing. The interaction of MTPand apoB 41 was also stable at pH 8.0–pH 5.8 (Fig. 8). LabeledapoB 41 and MTP coprecipitates (using anti-apoB antiserum)were isolated as described under “Experimental Procedures” in150 mMNaCl at pH 7.5 and washed in buffers at the various pHconditions shown. To account for the dissociation of apoB 41from the antibody, the apoB 41 and MTP bands were quanti-tated and expressed as a ratio of apoB 41 to MTP (arbitraryunits). Preferential removal of MTP should result in a dramaticrise in this ratio. Lowering the pH of the wash buffers resultedin some loss of apoB 41, presumably via antibody-antigen dis-sociation (compare apoB 41 intensity, tracks 1 with 5 and 6,Fig. 8). However, the proportion of MTP associated with apoB41 under these conditions remains comparable (ratio of apoB41 to MTP is 2.3 at pH 8.0 compared with 1.9 at pH 5.8). Thus,this interaction was also relatively insensitive to pH changes.Note that the unidentified protein that coprecipitates also withapoB 41 appears to dissociate at a pH less than 6.8 (Fig. 8,track 4).Association of Calnexin and KDEL-bearing ER Proteins with
ApoB—ApoB is likely to interact with a variety of other pro-teins in the ER as it undergoes a complex folding process to
FIG. 6. Coprecipitation of apoB 41 and MTP in cotransfectedCOS cells. COS cells were cotransfected with apoB 41 and MTP, thecells were pulse-labeled, and apoB (A: track 1) or MTP (A: track 2) wasimmunoprecipitated and analyzed by SDS-PAGE. The migration ofapoB 41 and MTP are as indicated by the arrowheads. A band with themobility of MTP coprecipitates when apoB is immunoprecipitated, anda faint band with the mobility of apoB 41 is seen when MTP is immu-noprecipitated. To further confirm this association, cell lysates fromunlabeled cotransfected cells were immunoprecipitated with MTP,Western blotted, and probed for the presence of apoB (B: track 1) orimmunoprecipitated for apoB, Western blotted, and probed for thepresence of MTP. Immunoreactive bands are seen in both cases. Theimmunoreactive bands at around 50 kDa are the heavy chains from theimmunoprecipitating antibodies (sheep anti-apoB and rabbit anti-MTP)that cross-react with the secondary anti-mouse immunoglobulinantibody.
FIG. 7. Stability of the MTP/apoB 41 coprecipitates to saltwashing. ApoB 41/MTP complexes were immunoprecipitated from co-transfected cells with anti-apoB antiserum and washed at various saltconcentrations as indicated. Despite increasing the salt concentrationfrom 0.15 mM (track 1) to 0.5 mM (track 4), the amount of MTP associ-ated with apoB 41 was unchanged. An unidentified protein, indicatedby the asterisk (;80 kDa), was also co-associated and resistant to saltwashing (compare the effect of pH, Fig. 8).
MTP and ApoB Interactions in the ER18690
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from
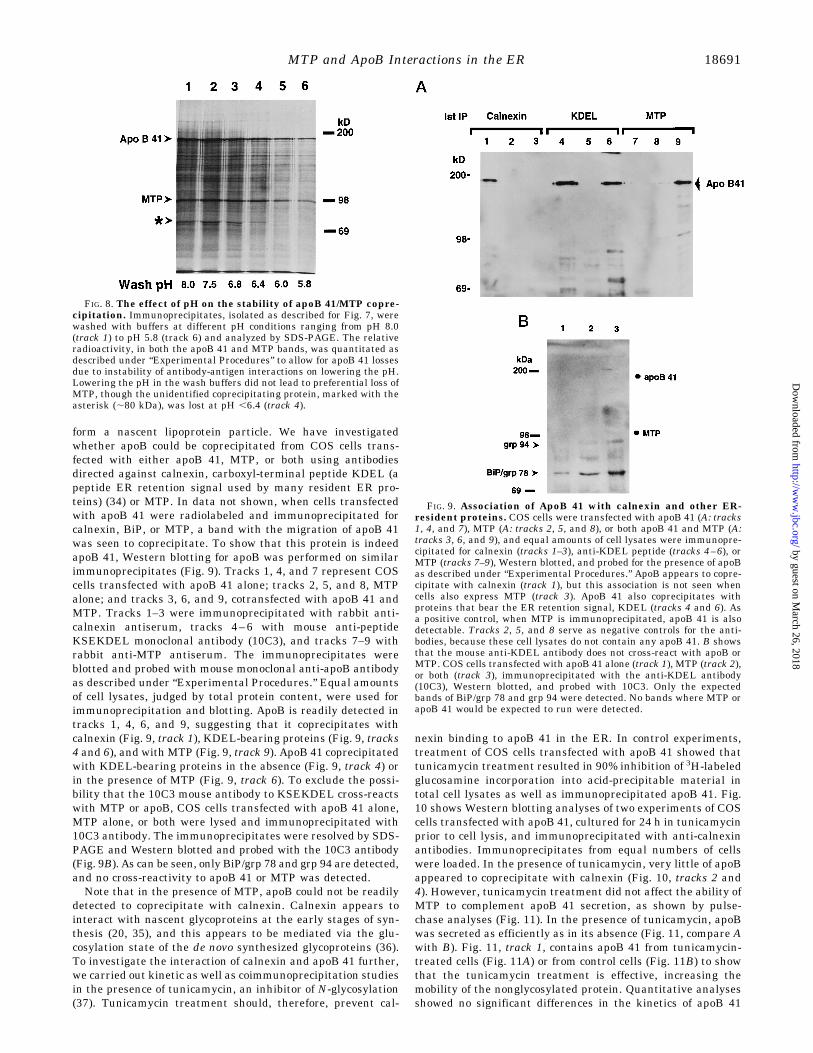
form a nascent lipoprotein particle. We have investigatedwhether apoB could be coprecipitated from COS cells trans-fected with either apoB 41, MTP, or both using antibodiesdirected against calnexin, carboxyl-terminal peptide KDEL (apeptide ER retention signal used by many resident ER pro-teins) (34) or MTP. In data not shown, when cells transfectedwith apoB 41 were radiolabeled and immunoprecipitated forcalnexin, BiP, or MTP, a band with the migration of apoB 41was seen to coprecipitate. To show that this protein is indeedapoB 41, Western blotting for apoB was performed on similarimmunoprecipitates (Fig. 9). Tracks 1, 4, and 7 represent COScells transfected with apoB 41 alone; tracks 2, 5, and 8, MTPalone; and tracks 3, 6, and 9, cotransfected with apoB 41 andMTP. Tracks 1–3 were immunoprecipitated with rabbit anti-calnexin antiserum, tracks 4–6 with mouse anti-peptideKSEKDEL monoclonal antibody (10C3), and tracks 7–9 withrabbit anti-MTP antiserum. The immunoprecipitates wereblotted and probed with mouse monoclonal anti-apoB antibodyas described under “Experimental Procedures.” Equal amountsof cell lysates, judged by total protein content, were used forimmunoprecipitation and blotting. ApoB is readily detected intracks 1, 4, 6, and 9, suggesting that it coprecipitates withcalnexin (Fig. 9, track 1), KDEL-bearing proteins (Fig. 9, tracks4 and 6), and with MTP (Fig. 9, track 9). ApoB 41 coprecipitatedwith KDEL-bearing proteins in the absence (Fig. 9, track 4) orin the presence of MTP (Fig. 9, track 6). To exclude the possi-bility that the 10C3 mouse antibody to KSEKDEL cross-reactswith MTP or apoB, COS cells transfected with apoB 41 alone,MTP alone, or both were lysed and immunoprecipitated with10C3 antibody. The immunoprecipitates were resolved by SDS-PAGE and Western blotted and probed with the 10C3 antibody(Fig. 9B). As can be seen, only BiP/grp 78 and grp 94 are detected,and no cross-reactivity to apoB 41 or MTP was detected.Note that in the presence of MTP, apoB could not be readily
detected to coprecipitate with calnexin. Calnexin appears tointeract with nascent glycoproteins at the early stages of syn-thesis (20, 35), and this appears to be mediated via the glu-cosylation state of the de novo synthesized glycoproteins (36).To investigate the interaction of calnexin and apoB 41 further,we carried out kinetic as well as coimmunoprecipitation studiesin the presence of tunicamycin, an inhibitor of N-glycosylation(37). Tunicamycin treatment should, therefore, prevent cal-
nexin binding to apoB 41 in the ER. In control experiments,treatment of COS cells transfected with apoB 41 showed thattunicamycin treatment resulted in 90% inhibition of 3H-labeledglucosamine incorporation into acid-precipitable material intotal cell lysates as well as immunoprecipitated apoB 41. Fig.10 shows Western blotting analyses of two experiments of COScells transfected with apoB 41, cultured for 24 h in tunicamycinprior to cell lysis, and immunoprecipitated with anti-calnexinantibodies. Immunoprecipitates from equal numbers of cellswere loaded. In the presence of tunicamycin, very little of apoBappeared to coprecipitate with calnexin (Fig. 10, tracks 2 and4). However, tunicamycin treatment did not affect the ability ofMTP to complement apoB 41 secretion, as shown by pulse-chase analyses (Fig. 11). In the presence of tunicamycin, apoBwas secreted as efficiently as in its absence (Fig. 11, compare Awith B). Fig. 11, track 1, contains apoB 41 from tunicamycin-treated cells (Fig. 11A) or from control cells (Fig. 11B) to showthat the tunicamycin treatment is effective, increasing themobility of the nonglycosylated protein. Quantitative analysesshowed no significant differences in the kinetics of apoB 41
FIG. 8. The effect of pH on the stability of apoB 41/MTP copre-cipitation. Immunoprecipitates, isolated as described for Fig. 7, werewashed with buffers at different pH conditions ranging from pH 8.0(track 1) to pH 5.8 (track 6) and analyzed by SDS-PAGE. The relativeradioactivity, in both the apoB 41 and MTP bands, was quantitated asdescribed under “Experimental Procedures” to allow for apoB 41 lossesdue to instability of antibody-antigen interactions on lowering the pH.Lowering the pH in the wash buffers did not lead to preferential loss ofMTP, though the unidentified coprecipitating protein, marked with theasterisk (;80 kDa), was lost at pH ,6.4 (track 4).
FIG. 9. Association of ApoB 41 with calnexin and other ER-resident proteins. COS cells were transfected with apoB 41 (A: tracks1, 4, and 7), MTP (A: tracks 2, 5, and 8), or both apoB 41 and MTP (A:tracks 3, 6, and 9), and equal amounts of cell lysates were immunopre-cipitated for calnexin (tracks 1–3), anti-KDEL peptide (tracks 4–6), orMTP (tracks 7–9), Western blotted, and probed for the presence of apoBas described under “Experimental Procedures.” ApoB appears to copre-cipitate with calnexin (track 1), but this association is not seen whencells also express MTP (track 3). ApoB 41 also coprecipitates withproteins that bear the ER retention signal, KDEL (tracks 4 and 6). Asa positive control, when MTP is immunoprecipitated, apoB 41 is alsodetectable. Tracks 2, 5, and 8 serve as negative controls for the anti-bodies, because these cell lysates do not contain any apoB 41. B showsthat the mouse anti-KDEL antibody does not cross-react with apoB orMTP. COS cells transfected with apoB 41 alone (track 1), MTP (track 2),or both (track 3), immunoprecipitated with the anti-KDEL antibody(10C3), Western blotted, and probed with 10C3. Only the expectedbands of BiP/grp 78 and grp 94 were detected. No bands where MTP orapoB 41 would be expected to run were detected.
MTP and ApoB Interactions in the ER 18691
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

secretion. Note also that tunicamycin treatment did not affectthe coprecipitation of MTP with apoB 41. Although these re-sults would suggest that calnexin may not be necessary forapoB secretion, it may play a role in the enhancement of folding
and, hence, secretion of apoB. We cotransfected human cal-nexin cDNA and apoB 41 to investigate whether overexpres-sion of calnexin would lead to secretion of apoB 41. No secretionof apoB 41 was seen under these conditions, although coimmu-noprecipitation of apoB and human calnexin was also demon-strable (data not shown). Similarly, transfection of apoB 41,human calnexin, and MTP together did not result in enhance-ment of apoB 41 secretion when compared to apoB 41 and MTPalone (data not shown).
DISCUSSION
MTP Effects on ApoB Secretion and Oleate Responsiveness—When MTP is coexpressed with apoB truncates, apoBs largerthan B29 are secreted; moreover, secretion becomes responsiveto oleate supplementation and secreted apoB particles containnewly synthesized triglyceride. MTP coexpression does not sig-nificantly alter the metabolism of apoBs smaller than B29, andthe secretion of these species, in the presence of MTP, is notresponsive to oleate supplementation. To place these results incontext, studies in human and rat hepatoma cell lines usingcarboxyl-terminal apoB cDNAs have shown that there is alinear relationship between the size of apoB and its buoyantdensity and diameter (7, 27, 28, 32). Hence, the larger theapoB, the greater the amount of lipid it is secreted with, andthe greater its lipoprotein diameter. However, this relationshipappears to breakdown around truncates in the B26–B29 range.Below this size, the apoB proteins do not bind sufficient lipidand have densities of more than 1.2 g/ml (38). In our system, wecan now demonstrate that MTP affects the secretion of apoBslarger than B29 and that the oleate responsiveness is also onlyevident for apoBs larger than B29. Oleate responsiveness hasbeen shown previously to be mediated at the posttranslationallevel, primarily at the expense of apoB degradation. Our stud-ies demonstrate that MTP mediates oleate responsiveness,since only in the presence of MTP is the oleate effect seen. Wecannot distinguish whether this effect results from increasedproduction of cotranslationally formed lipoproteins (and de-creased membrane-inserted apoB) or whether MTP attaches tothe membrane-inserted apoB and converts it into lumenal ru-dimentary lipoprotein. However, since apoB interaction withcalnexin is less pronounced in the presence of MTP, we favorthe latter mechanism (see below).Our experiments show that apoB 41, secreted in the presence
of MTP, contains a significant amount of newly synthesizedtriglyceride. Incubation with labeled oleate showed that muchof this fatty acid was incorporated as triglyceride into thesecreted lipoprotein, and very little was present as cholesterolester. Although we could not detect any changes in cholesterolester levels in our experiments, we cannot exclude the possi-bility that some cholesterol ester is necessary for apoB secre-tion. Previous studies have shown that newly synthesized tri-glycerides are preferentially incorporated into the lipoproteinparticles, and our studies in a non-hepatic line corroborate this.What is not clear is the source of this pool. MTP is not anenzyme but appears to facilitate the transfer of neutral lipidfrom one compartment to another to equilibrium. MTP would,therefore, need to be in contact with both the triglyceride donorpool (from an active metabolic site) and the nascent apoBparticle. Although the specific site of the MTPzPDI complexwithin the ER has not been localized, it is likely that thiscomplex is lumenal, since neither proteins have transmem-brane domains. Hence, this complex must be in contact with ametabolically active neutral lipid pool, as well as apoB to facil-itate transfer of lipid into the nascent lipoprotein particle.MTP and ApoB Interactions—MTP associated with apoB
intracellularly, and this association was noted with all apoBs,from B13 to B41. Qualitatively, less MTP coimmunoprecipi-
FIG. 10. Tunicamycin inhibits the coprecipitation of apoBwith calnexin. COS cells were transfected with apoB 41 and incubatedin tunicamycin at 5 mg/ml for 24 h; then cell lysates were prepared,immunoprecipitated with anti-calnexin antibodies, andWestern blottedfor the presence of apoB 41. The results of two such experiments areshown. In the absence of tunicamycin (tracks 1 and 3), apoB 41 (arrow-head) is readily detectable, but in its presence, very little of apoB 41 wasdetectable (tracks 2 and 4).
FIG. 11. The effect of tunicamycin on the kinetics of apoB 41secretion in the presence of MTP. COS cells were transfected withMTP and apoB 41 as described under “Experimental Procedures,” in-cubated in the presence or absence of tunicamycin for 24 h, pulse-labeled, chased, apoB immunoprecipitated, and analyzed by SDS-PAGE. A shows the pulse-chase analysis in the absence of tunicamycin,and B, in its presence. Track 1 in A contains apoB 41 immunoprecipitedin the presence of tunicamycin, and track 1 in B contains apoB 41immunoprecipited from control cells to indicate that the effects of tu-nicamycin do lead to an increased mobility because of inhibition ofglycosylation. In both cases, about 15% of apoB 41 was secreted in 5 h.Note that MTP coprecipitates in the presence of tunicamycin, suggest-ing that the apoB glycosylation state does not play a role incoprecipitation.
MTP and ApoB Interactions in the ER18692
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

tated with the smaller truncates (,B23) than with the largerones. The interaction with apoB 41 was resistant to high saltwashing and was also resistant to lowering of pH buffers as lowas 5.8. These results suggest that this interaction may bepredominantly hydrophobic in nature. Clearly, MTP must dis-associate from apoB at some point, because no MTPzPDI com-plexes are secreted under normal circumstances. Since there isevidence that the lipoprotein particle continues to obtain lipidfollowing its exit from the ER, this complex may disassociate inthe cis-Golgi, where a more acidic environment is present, in amanner analogous to the disassociation of receptor-associatedprotein from newly synthesized low density lipoprotein recep-tor-related protein (39), although our results suggest thatchanges in pH alone may not suffice for MTP to disassociatefrom apoB. The KDEL retrieval signal on PDI could then shut-tle the MTP complex back to the ER (40). The smaller apoBs,such as B17, are secreted with very little lipid but can obtainfurther lipid from serum to attain densities in the high densitylipoprotein range (24, 25). However, MTP coexpression did notaugment secretion of these smaller truncates, in the absence orpresence of oleate supplementation, despite being able to bindto them. MTP binding to these smaller apoBs may not be asrobust as to the larger apoBs. Cotransfection experimentsmake it difficult to measure this, but stable heterologous ex-pression of MTP may allow us to study this hypothesis, as wellas to better define the domains of apoB that interact with MTP.Calnexin and ApoB Interactions—Only one other ER protein
has been shown to interact with apoB, i.e. calnexin (20). One ofthe proposed roles of calnexin is to recognize incompletelyfolded newly synthesized glycoproteins, and that it may do byrecognition of the glucosylation state. The binding to calnexinis thought to engage a round of folding, perhaps by recruitingother ER proteins to help in this process (36, 41). In the process,glucose residues are cleaved from the N-glycosylated moieties,and the protein is released. If the protein is incompletelyfolded, it is recognized by UDP-glucosyltransferase and glu-cosylated (42, 43), making it a substrate for calnexin binding,and a further round of folding ensues. Once correctly folded, theprotein can then leave this quality control onto to its finaldestination. The binding of calnexin to the newly synthesizedglycoproteins is transient, but in cases where cell lines fail toexpress one chain of a multichain complex, such as the T-cellreceptor, the binding of calnexin to the residual chains is pro-longed (35). These authors suggested that calnexin may, there-fore, play a role in ER degradation. However, it has beendemonstrated previously that tunicamycin treatment of cells,which inhibits N-glycosylation, does not affect apoB secretionfrom cultured hepatocytes (44), and some secreted apoB 100from human hepatoma cell lines may also not be fully glycosyl-ated (45). We have investigated the interaction of calnexin andapoB in COS cells. Overexpression of human calnexin in COScells with apoB did not lead to attainment of a “folded” state forapoB such that it could leave the ER, nor were the degradationkinetics altered. Indeed, although apoB could be readily dem-onstrated to coprecipitate with endogenous calnexin, in thepresence of MTP this interaction was absent or very transient.Tunicamycin treatment of COS cells inhibited N-glycosylation,and thus calnexin binding, but did not alter the secretionkinetics and lipoprotein formation of apoB in the presence ofMTP, confirming previous findings that N-glycosylation wasnot necessary for apoB secretion. Calnexin has been shown tobind to nonglycosylated proteins (46, 47), but our Westernblotting data (Fig. 9) suggest that the binding of calnexin toapoB is likely to be mediated via the glycosylation interactions.In the presence of overexpressed MTP, the interaction withcalnexin is diminished in COS cells, but in hepatoma cells,
calnexin can be readily seen to associate with apoB (20). Oneexplanation for this may be that MTP may be rate-limiting inhepatoma cells. This would allow for the relative excess of apoBsynthesized to interact with calnexin, even though this apoB isultimately destined for degradation. The possibility that thepool of apoB that interacts with MTP in hepatoma cells is thelumenal fraction compared with the membrane-inserted frac-tion that interacts with calnexin should be amenable to testing.Experiments to address the partition of MTP may allow us todistinguish whether this model is correct.ApoB Interactions with KDEL Proteins—We have attempted
to identify some of the other proteins that may be involved inapoB folding within the ER. Using antibodies directed againstthe carboxyl-terminal peptide KSEKDEL, apoB coprecipitatedwith proteins recognized by this antibody. This interaction waspresent in the absence of MTP (which is associated with theKDEL signal of PDI), suggesting that other ER proteins areinvolved. One obvious candidate is BiP/grp 78, a ubiquitousand relatively abundantly expressed ER chaperone known tobind to misfolded proteins. BiP has a preference for hydropho-bic sequences (48), a large number of which are present onapoB. There is a protein of about 80 kDa that coprecipitateswith apoB, irrespective of MTP expression, and which appearsto disassociate from the immunoprecipitates at a pH of 6.0 butis stable to high salt (see Figs. 7 and 8). In data not shown, thisprotein band has a half-life of more than 5 h, is not secreted, ispresent at short and long chase time points, and absent whennonimmune antiserum is used, suggesting that it may be anER-resident protein. Our antibodies at present do not allow usto specifically identify simian BiP, although we strongly believethat the coprecipitating protein is BiP.In this study, we show that the “secretory block” imposed on
heterologously expressed apoB can be overcome by coexpres-sion with MTP. This coexpression also leads to responsivenessto oleate supplementation, and the secreted lipoprotein parti-cles contain newly synthesized triglyceride. In addition, oleatesupplementation is most effective for truncates larger thanB29, and the secretion of the smaller truncates is not aug-mented by either oleate and/or MTP coexpression. MTP wasfound to be physically associated with intracellular apoB 41,and this interaction is hydrophobic in nature. Other ER pro-teins were also found to interact with apoB, i.e. calnexin and anunidentified protein of 80 kDa. Interaction with calnexin didnot appear to be necessary for lipoprotein secretion, and inter-estingly, in the presence of MTP, this interaction may be tran-sient or absent. Although the identification of the 80-kDa pro-tein is not known at this stage, it may interact with apoB viahydrophobic bonding, and it dissociates from apoB at pH con-ditions present in the Golgi. These studies suggest that MTPplays role in the folding of apoB by providing it with neutrallipid; this neutral lipid is necessary for the formation of alipoprotein but also for allowing apoB to attain a conformationsufficient for it to escape the quality control of the ER.
Acknowledgments—We gratefully acknowledge Drs. Michael Bren-ner, David Gordon, John Wetterau, Richard Gregg, and Gustav Schon-feld for their gifts of reagents and clones, to Dr. Michael Roth for criticalreading of the manuscript, and to Loyce Rutledge for excellent technicalassistance.
REFERENCES
1. Lodish, H. F. (1988) J. Biol. Chem. 263, 2107–21102. Rose, J. K., and Doms, R. W. (1988) Annu. Rev. Cell Biol. 4, 257–2883. Klausner, R. D. (1989) New Biol. 1, 3–84. Hurtley, S. M., and Helenius, A. (1989) Annu. Rev. Cell Biol. 5, 277–3075. Sato, R., Imanaka, T., Takatsuki, A., and Takano, T. (1990) J. Biol. Chem. 265,
11880–118846. Dixon, J. L., Furukawa, S., and Ginsberg, H. N. (1991) J. Biol. Chem. 266,
5080–50867. Yao, Z., Blackhart, B. D., Linton, M. F., Taylor, S. M., Young, S. G., and
McCarthy, B. J. (1991) J. Biol. Chem. 266, 3300–3308
MTP and ApoB Interactions in the ER 18693
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8. White, A. L., Graham, D. L., LeGros, J., Pease, R. J., and Scott, J. (1992)J. Biol. Chem. 267, 15657–15664
9. Bostrom, K., Wettesten, M., Boren, J., Bondjers, G., Wiklund, O., and Olofsson,S. O. (1986) J. Biol. Chem. 261, 13800–13806
10. Dixon, J. L., and Ginsberg, H. N. (1993) J. Lipid Res. 34, 167–17911. Kane, J. P., and Havel, R. J. (1989) in The Metabolic Basis of Inherited Disease,
pp. 1139–1164, McGraw-Hill Inc., New York12. Sharp, D., Blinderman, L., Combs, K. A., Kienzle, B., Ricci, B., Wager-Smith,
K., Gil, C. M., Turck, C. W., Bouma, M. E., Rader, D. J., Aggerbeck, L. P.,Gregg, R. E., Gordon, D. A., and Wetterau, J. R. (1993) Nature 365, 65–69
13. Wetterau, J. R., Aggerbeck, L. P., Bouma, M. E., Eisenberg, C., Munck, A.,Hermier, M., Schmitz, J., Gay, G., Rader, D. J., and Gregg, R. E. (1992)Science 258, 999-1001
14. Patel, S., Pessah, M., Beucler, I., Navarro, J., and Infante, R. (1994) Athero-sclerosis 108, 201–207
15. Pessah, M., Benlian, P., Beucler, I., Loux, N., Schmitz, J., Junien, C., andInfante, R. (1991) J. Clin. Invest. 87, 367–370
16. Davis, R. A., Thrift, R. N., Wu, C. C., and Howell, K. E. (1990) J. Biol. Chem.265, 10005–10011
17. Boren, J., Wettesten, M., Sjoberg, A., Thorlin, T., Bondjers, G., Wiklund, O.,and Olofsson, S. O. (1990) J. Biol. Chem. 265, 10556–10564
18. Boren, J., Graham, L., Wettesten, M., Scott, J., White, A., and Olofsson, S. O.(1992) J. Biol. Chem. 267, 9858–9867
19. Sakata, N., Wu, X., Dixon, J. L., and Ginsberg, H. N. (1993) J. Biol. Chem. 268,22967–22970
20. Ou, W. J., Cameron, P. H., Thomas, D. Y., and Bergeron, J. J. (1993) Nature364, 771–776
21. Kane, J. P., Hardman, D. A., and Paulus, H. E. (1980) Proc. Natl. Acad. Sci.U. S. A. 77, 2465–2469
22. Gordon, D. A., Jamil, H., Sharp, D., Mullaney, D., Yao, Z., Gregg, R. e., andWetterau, J. (1994) Proc. Natl. Acad. Sci. U. S. A. 91, 7628–7632
23. Leiper, J. M., Bayliss, J. D., Pease, R. J., Brett, D. J., Scott, J., and Shoulders,C. C. (1994) J. Biol. Chem. 269, 21951–21954
24. Patel, S. B., and Grundy, S. M. (1995) J. Lipid Res. 36, 2090–210325. Herscovitz, H., Hadzopoulou-Cladaras, M., Walsh, M. T., Cladaras, C., Zannis,
V. I., and Small, D. M. (1991) Proc. Natl. Acad. Sci. U. S. A. 88, 7313–731726. Thrift, R. N., Drisko, J., Dueland, S., Trawick, J. D., and Davis, R. A. (1992)
Proc. Natl. Acad. Sci. U. S. A. 89, 9161–9165
27. Spring, D. J., Chen-Liu, L. W., Chatterton, J. E., Elovson, J., and Schumaker,V. N. (1992) J. Biol. Chem. 267, 14839–14845
28. McLeod, R. S., Zhao, Y., Selby, S. L., Westerlund, J., and Yao, Z. (1994) J. Biol.Chem. 269, 2852–2862
29. Hochstenbach, F., David, V., Watkins, S., and Brenner, M. B. (1992) Proc.Natl. Acad. Sci. U. S. A. 89, 4734–4738
30. Huovila, A. P., Eder, A. M., and Fuller, S. D. (1992) J. Cell Biol. 118,1305–1320
31. Krul, E. S., Kleinman, Y., Kinoshita, M., Pfleger, B., Oida, K., Law, A., Scott,J., Pease, R., and Schonfeld, G. (1988) J. Lipid Res. 29, 937–947
32. Graham, D. L., Knott, T. J., Jones, T. C., Pease, R. J., Pullinger, C. R., andScott, J. (1991) Biochemistry 30, 5616–5621
33. Folch, J., Lees, M., and Stanley, G. H. S. (1957) J. Biol. Chem. 226, 497–50934. Pelham, H. R. (1990) Trends Biochem. Sci. 15, 483–48635. David, V., Hochstenbach, F., Rajagopalan, S., and Brenner, M. B. (1993)
J. Biol. Chem. 268, 9585–959236. Hammond, C., Braakman, I., and Helenius, A. (1994) Proc. Natl. Acad. Sci.
U. S. A. 91, 913–91737. Elbein, A. D. (1987) Annu. Rev. Biochem. 56, 497–53438. Schumaker, V. N., Phillips, M. L., and Chatterton, J. E. (1994) Adv. Prot.
Chem. 45, 205–24839. Bu, G., Geuze, H. J., Strous, G. J., and Schwartz, A. L. (1995) EMBO J., 14,
2269–228040. Munro, S., and Pelham, H. R. (1987) Cell 48, 899–90741. Hammond, C., and Helenius, A. (1994) J. Cell Biol. 126, 41–5242. Trombetta, S. E., Bosch, M., and Parodi, A. J. (1989) Biochemistry 28,
8108–811643. Sousa, M. C., Ferrero-Garcia, M. A., and Parodi, A. J. (1992) Biochemistry 31,
97–10544. Siuta-Mangano, P., Howard, S. C., Lennarz, W. J., and Lane, M. D. (1982)
J. Biol. Chem. 257, 4292–430045. La Belle, M., McCall, M. R., Krauss, R. M., and Forte, T. M. (1990) Biochim.
Biophys. Acta 1046, 288–29346. Margolese, L., Waneck, G. L., Suzuki, C. K., Degen, E., Flavell, R. A., and
Williams, D. B. (1993) J. Biol. Chem. 268, 17959–1796647. Rajagopalan, S., Xu, Y., and Brenner, M. B. (1994) Science 263, 387–39048. Fourie, A. M., Sambrook, J. F., and Gething, M. J. (1994) J. Biol. Chem. 269,
30470–30478
MTP and ApoB Interactions in the ER18694
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Shailendra B. Patel and Scott M. Grundywithin the Endoplasmic Reticulum in a Heterologous Expression System
Interactions between Microsomal Triglyceride Transfer Protein and Apolipoprotein B
doi: 10.1074/jbc.271.31.186861996, 271:18686-18694.J. Biol. Chem.
http://www.jbc.org/content/271/31/18686Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/271/31/18686.full.html#ref-list-1
This article cites 47 references, 31 of which can be accessed free at
by guest on March 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from