
The Immune System in Atherosclerosis HANSSON Nat Immunol 2011
Upload
sholeh-rhazesCategory
view
217download
0
7/27/2019 Immunol 1995 Kennedy 203 9
http://slidepdf.com/reader/full/immunol-1995-kennedy-203-9 1/7
Molecular Cloning and Functional Characterization ofHuman Lymphotactin'
Jacqueline Kennedy,* Gregory S. Kelner,* Sarah Kleyensteuber,* Thomas J . Schall,*Mary C. Weiss,+ Hans Yssel,* Pat ricia V. Schneider,* Benjamin G. Cocks,* Kevin B. Bacon,*and Albert Zlot nik2*
Departmentsof *Immunology, Molecular Biology, and "Human Immunology, DNAX Research Institute of Molecular andCellular Biology, Palo Alto, CA94304
We describe the isolation of a cDNA that encodes human lymphotactin (Ltn), a new class of lymphocyte-specific
chemokine. Human Ltn shows similarity to some members of theC-C
chemokine family but has lost the first andthird cysteine residues that are characteristic of the C-C and C-X-C chemokines. Ltn is chemotactic for lymphocytesbut not for monocytes, a characteristic that makes it unique among chemokines. In addition, calcium flux desen-sitization studies indicate that Ltn uses a unique receptor. The human Ltn gene maps to a different chromosomethan do the C-C and C-X-C chemokine families. Taken together, these characteristics indicate that Ltn is the firstexample of a new class of human chemokines with preferential effects on lymphocytes. Th e Journal of Immu-nology, 1995, 155: 203-209.
Cemokines are small, secreted proteins that act as
imm une regulators through their ability to che-moattracteukocytes (1 , 2). Traditionally,he
chem okine superfam ily has been structurally subdividedinto two branches, C-C and C-X-C, depending on whetherthe first two cysteines of he mature molec ule are adjacentor separated by a single residue. Thesetwo branches seg-regate functionally as well,because C-X-C chemokinesact primarily on neutrophils, whereas C-C chemokines arelargely c hemo tactic ormon ocytes but not neutrophils.Chemokines have received increased attention in recentyears because of the mportantrole theyplay duringnorm al eukoc yte traffickingand dur ing inflammatoryand imm une respon ses. recent report that describes anIL-8R"- mouse generated through homolog ous recom-bination shows an exam ple of the imp ortance of C-X-C
chemokines in neutrophil function(3). However, althoughC-C and C-X-C chemokines are widely believed to be im-portant in the migration and function of neutrophils andmonocytes/macrophages,no clear counterpart for lympho-cytes has been identified. Although several C-C chemo-
Received for publication J anuary 27, 1995. Accepted for publication April 17,1995.
The costs of publication of this article were defrayed in part by the payment of
accordance with 18 U.S.C. Section 1734 solely to indicate this fact.page charges. This article must therefore be hereby marked advertisement in
' DNAX Research Institute is supported by Schering-P lough Corporation
' ddress correspondence and reprint requests to Dr . Albert Zlotnik, DNAXResearch Institute, 901 California Avenue, Palo Alto, CA 94304.
kines have been shown to chemoattract lym phocytes(2),this is unlikely to be their main function, because thesechemokines chemoattract other cell types as well.
The existence of chemo kines that specifically regulatelymphocyte migration has not been described untilre-cently. We have reported the cDNA cloning of a novelchemokine, designated lymphotactin (Ltn),3 from a mousepro-T cell library (4). This novel chem okine sa goodcandidate for such a ymphocyte-specific chemoattractant.How ever, there are several e xam ples of chemokines, suchas IL-8, C10, and eotaxin (1 ,5 ,6 ), for which a correspond-ing homologue either does not exist or has not been foun din other species. Therefore, it wa s important to establishwhether a human homo logue for L tn could be identified.
In this study, we describe the isolation of a cD NA thatencodes human Ltn. We present structural, functional, and
genetic data indicating that human Ltn is indeed the firstexample of ane wclass of human lymphocyte-specificchemokine, which we propose be designated the C class,that does not belong to either the C-C or the C-X-C fam-ilies. Furthermore, we show that human Ltn is an impor-tant and abundant product of several activated T cell sub-sets. All of these characteristics strongly suggest animportant role for Ltn during inflammatory and immuneresponses.
Abbreviations used in this paper: Ltn, lyrnphotactin; MCP , monocyte che-motactic protein.
Copyright 0 1995 by The American Association of Immunologists 0022-1 767/95/$02.00

7/27/2019 Immunol 1995 Kennedy 203 9
http://slidepdf.com/reader/full/immunol-1995-kennedy-203-9 2/7
204 CLONING OF HUMAN Ltn
Materials and Method s
Screening of a cD NA library
Construction of a cDNA library generated from the CD8' T cell cloneA10 has been described previously (7). The library was screened bystandard methods (8) using mouse Ltn cDNA as a probe.
DN A sequencing and analysisThe entire cDNA of human Ltn was sequenced by the dideoxynucleotidechain-termination method with T7 polymerase (United States Biochemi-cals, Cleveland, OH) using dsDNA as a template. Database searching andsequence analysis were performed using IntelliGenetics programs(Mountain View, CA).
/solation of T cell subsets
CD4' and CD8+ thymocytes were isolated by cell sorting. In brief, asingle-cell suspension was prepared from human fetal thymus and la-beled with Leu-2-FITC and Leu-3-phycoerythrin (Becton Dickinson, SanJose, CA). Next, 1.2 X 10' single-positive CD4 and CD8 thymocyteswere purified by FACS sorting on a Quadra 650-CellQuest (Becton Dick-inson, Mountain View, CA). The purity of the sorted cells was >99.5%.Thymocyte populations were activated in the presence of calcium iono-phore A23187 (500 ng/ml) and PMA (1 ng/ml) for 5 h. CD4+ and CD8+PBL were prepared as follows: PBMC were isolated from buffy coats ofhealthy donors by density centrifugation over Ficoll Hypaque (SigmaChemical Co., St. Louis, MO), as described previously (9). CD4' andCD8+ T cells were purified from PBMC after adherence to plastic andafter positive selection using magnetic beads. In brief, 100 X 10' PBMCwere cultured for 1 h at 37°C in a 100-mm tissue culture dish (BectonDickinson, Lincoln Park, NJ) in Yssel's medium supplemented with 1%human AB+ serum (10). Then, nonadherent cells were incubated withanti-CD4- and anti-CD8-coated Dynabeads (Dynal Beads M450; DynalAS., Oslo, Norway), respectively, at a bead to cell ratio of 1 O : l . Themixture was incubated while gently rotating for 1 h at 4"C, and CD4+ orCD8' T cells were collected by using a magnetic particle concentrator,according to the manufacturer's recommendations. The purities of theCD4' and CD8+ T cell populations were >90% and >85%, respec-
tively. A portion of the CD4' and CD8+ T cells was activated in thepresence of calcium ionophore A23187 (500 ng/ml) and PMA (1 ng/ml)for 5 h .
Preparation of RNA and Northern blot analysis
Total RNA was prepared from the sorted and activated CD8+ and CD4+thymocytes and the activated and unactivated CD8+ and CD4+ PBLusing the RNAzol B method (TM Cinna Scientific Inc., Friendswood,TX). The RNA samples were fractionated on 0.85% denaturing agarosegels and transferred to BA-S nitrocellulose (Schleicher & Schuell, Keene,NH) as described (7). RNA from human spleen, thymus, prostate, testis,ovary, small intestine, colon, and PBL was purchased as a Northern blotfrom Clontech (Palo Alto, CA). Filters were hybridized as described (1 1)with a XrnrrI-PvuII fragment of pJFEhuLtn, which contains the Ltn cod-ing region.
Production o f recombinant human Ltn
A bacterial expression plasmid containing human Ltn cDNA was con-structed. The cDNA was mutagenized by using PCR to insert a Hind111restriction site proximal to the Val residue of the mature protein and anXhol site distal to the translation stop codon of the open reading frame sothat the resultant fragment could be subcloned into the pFLAG-I plasmid(International Biotechnologies, New Haven, CT). The expression plas-mid was transformed into the Topp5 Escherichia coli strain (Stratagene,La Jolla, CA), and recombinant protein was induced and purified as de-scribed previously (4). Fractions were collected and pooled. Two poolswere used in the following experiments: one deemed positive for Ltn andthe other deemed negative for Ltn on the basis of activity in the CaZ+ luxand chemotaxis assays.
Microchemotaxis assay
Chemotaxis assays were performed using a 48-well microchemotaxis ap-paratus as described previously (12). Cells for assay were prepared asfollows: CD4+ and CD8+ cells were purified from PBL after Ficoll treat-ment using OKMl anti-BL12 IgGl Abs (provided by Dr. K. Soo, Uni-versity College London, London, UK) for 30 min at 4°C followed byDynal bead separation to remove macrophages and B cells. CD4+ and
CD8+ cells were separated using anti-CD4 and anti-CD8 negative se-lection (Immunotech, Marseille, France) and Dynal beads. Purities of95% for CD4+ T cells and 93% for CD8+ T cells were determined byusing FACScan analysis. Monocytes were purified from PBL by Ficolltreatment and a two-step adherence on plastic petri dishes for 2 h fol-lowed by the removal of T and B cells by using negative selection. Then,each cell type was added to the top chamber, and recombinant Ltn wasadded to the bottom chamber of the microchemotaxis apparatus. Migra-tion was expressed as cell numberifive high power fields (X400); dupli-cate wells were counted fo r each of three experiments.
Ca2+ Nux assay
Human PBL were loaded in the presence of the calcium indicator indol-1AM (3 pM; Calbiochem, San Diego, CA) and then challenged with Ltn
or a combination of Ltn and IL-8. Fluorescence was measured on a Pho-ton Technology International spectrofluorometer at an excitation wave-length of 350 nm. Simultaneous emissions were recorded at 400 and 490nm, and ratios were calculated at two pointsis.
Chromosome mapping
A Southern blot containing EcoRI-digested humanihamster hybrid DNAwas purchased from BIOS Laboratories, Inc. (New Haven, CT) and hy-bridized to the XmnIiNotI cDNA fragment of pJFEhuLtn, as described(11). Discordance values for each chromosome were calculated by di-viding the sum of false positive and false negative events by 20 (thenumber of cell lines analyzed) an d multiplying by 100.
Results
Isolation and characterization of a cD NA encodinghuman Ltn
Human cDNAclonesencoding L m were identified byscreening a library generated from the CD8+T cell cloneA 10 (7 ) with the use of mouse Ltn cDNA asa probe. FourcD NA clones or Ltn were isolated, representing 730-,625-,562-, and 520-bp species. Th e 730-,562-, and520-bp cDNA clones contained identical open readingframes of 114 amino acids but differed in the lengths oftheir 3' untranslated regions. The open reading frame forthe 625-bp speciescontained conservative aminoacid sub-stitutions at positions 7 (Asp-H is) and8 (Ly s -kg ) , p rob-ably as a result of polymorphism (Fig. 1). Th e 562 - and520-bp clones were the most predominant in the library.Only two clones were represented by the 625-bp species,and only one clone was represented by the 730-bp species.Figure 1 shows the nucleotide and deduced amino acidsequence of the 562-bp cDN A clone. The3' untranslatedregion of the cDN A contains two polyadenylation signalsites. Th e 625- and 520-bp clones use thefirst polyade-nylation signal site, wherea s the 730-bp species uses thesecond but contains a 180-bp insertion upstream of thefirst that contains two more polyadenylation sign al sites.

7/27/2019 Immunol 1995 Kennedy 203 9
http://slidepdf.com/reader/full/immunol-1995-kennedy-203-9 3/7
The J ournal of Immunology 205
1 GCGGCACCTCAGCCTGCA CTTTCTCTGCCTCTT GG C ATCGC TCTTCCT 59-2 1 ICt A r p Leu Le u I la Leu A l a Leu Leu G l y Ile Cys Ser Leu Th r
60 GCA TI C ATT GTG W GGTGTA GGG ACTGAAGTCCAATAGGG AC C TGT GTG AGC 11 6-6 A l aTy r I le V al Gl u Gl y Val Gl ySe r Gl u V al Ser Asp LysAr gTh rCy r V al Ser
’1 17 CTC CT K C A G CGA CTG CCG Gn AGCCATCAG AC C TIC ACCTCCG GApl GGC 17314 Le u Th rTh rGl nArg Leu Pro V a l Ser Ar g I le Ly sTh rTy rTh r Il e Th r Gl u Gl y
174 TCCTG GA GCA GTA ATT T lT A l l A CC AA A CGT GGC CTAAA GTC TGT GCT CAT CA 23 033 Ser Leu A r gA l a Val Ile Phe Il e Th r Ly s Ar gGly Leu Lys Va l Cys Ala Asp Pro
231 CAACC AC G TGGGTGCAACGTGGTCGG AG C ATGACGG AAA TCCACCCGA 2875 2 G l n A l aTh r Tr p Val Ar g Asp Val Va l Ar g Ser Met Asp ArgLys Ser Asn ThrAr g
28 8 AAT AC TG TCCAE CC AAG CCACAGGACCAG C M TCG CC AT CAGCTGTG 3447 1 AsnAsn Me t Ile Gln Th rL y rPr o Thr Gl yTh rGl n Gln Ser Thr Asn ThrA la Val
345 ACCTGCTGCAG T A G T C T C T G G C A C C C T G T C C G T C T C C A G C C A G C C A G C T C C A C41296 Th r Le u Th rGl y ***
413 C C T C A T G G A C T G A G ~ A T A C T C A C C ~ A T G A A A G C A C T G C A T G ~ T T A ~ ~ C ~ G T A T ~ A C ~87
48 8 T T A A A T G T C ~C T G T A T T C A C TT A T A T G ~ CT M ~ ~ T TA T T T A ~ A ~ A A G A p lT A A A A A A A A A A A A p l A A62
FIGURE 1. Nucleotide and redicted amino cid se-quences of human Ltn. The deduced amino acid sequence is
shown below the nucleotide sequence. The polyadenylationsignal sites ( A ATA A A )are underlined. The nucleotide se-quence of human Ltn has been submitted to GenBank underthe accession number U23772.
Comparison of Ltn wi th the C-C chemokines
Comparison of the aminoacid sequence deduced from hehuman cDN A clone with mouse Ltn evealed that theyshare 60% sequence identity(Fig.2). Wh en theproteinsequence of human Ltn was compared with the Swiss Pro-tein Database, the C-C chemokinefamily members mono-cyte hemotactic rotein-2MCP -2) and macrophageinflammatory pro tei n-l a we re dentified as sharing signif-icanthomology to Ltn (Fig. 2). Therefore, human Ltnshare s the highest amin o acid identity with the C -C che-
mokines, especially in the region that correspondsto theputative hird exon. Hum an Ltn lacks the first and hirdcysteine residues that would pairto form one of two di-sulfide bridges characteristic of the C -C and C-X -C che-mokine families (13). It is interesting to note that there isahigh degre e of homology in the tail region betw eenmo use and human Ltn. Although a similar C-terminal ex-tension can be found in mouse MCP-1 orJE (14), this tailregion has been lost in human MCP-1 (15-17); therefore,there are no known human C-C chem okineswith extendedC termini. However, the human C-X-C chemokine mono-kine nduced by IFN -y (mig -y) featur es a similar C-ter-minal extension (18).
Expression and distribution of Ltn mRNA
The expression of human Ltn mR NA is very abundant insom e activated T cells. Activated CD8+ thymocytes ex-press Ltn mRN A, but activated CD 4+ thymocy tes do notexpressdetectable evels of Ltn (Fig. 3A). Ltn isalsoabundantly expressed in activated peripheral CD8+T cellsisolated from PBL and is expressed at lower levels in ac-tivated peripheral CD4+T cells (Fig. 3 B ) . In addition, LtnmR NA w as detected in resting spleen, thymus, small in-testine, and PBL at high lev els and in prostate a nd ovaryat lower levels (Fig. 3C ).
Chemotactic properties of Ltn
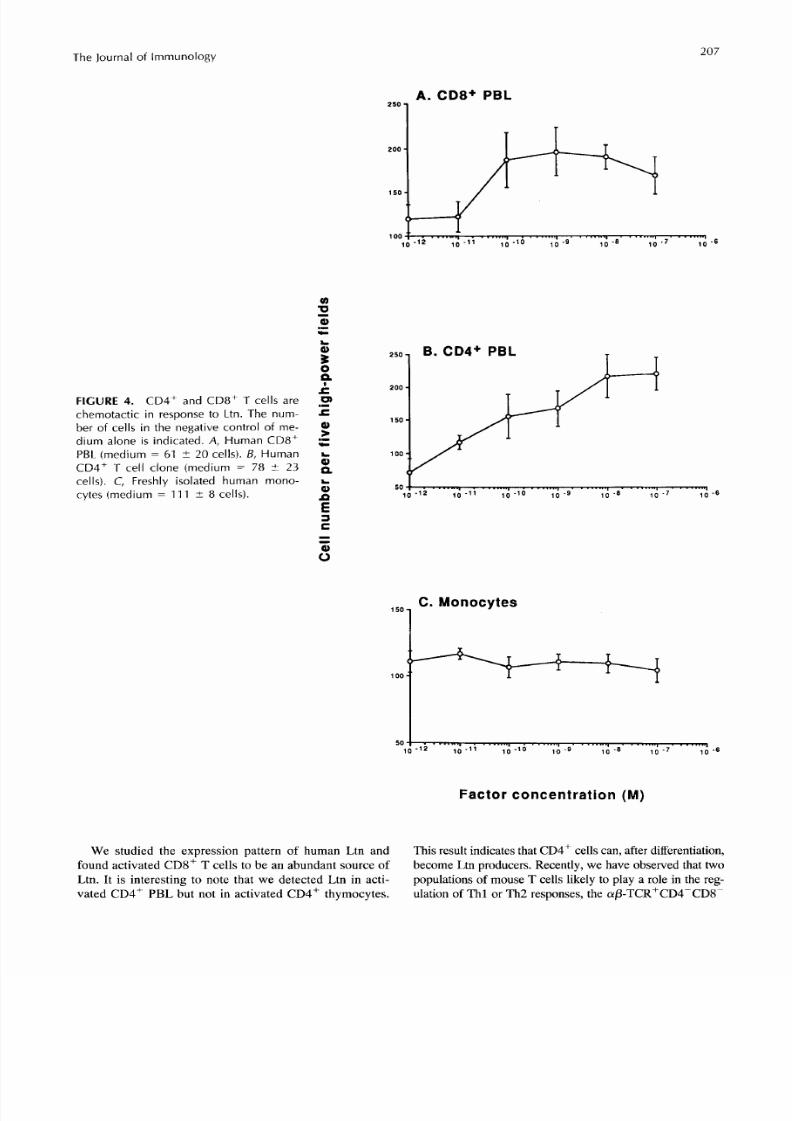
Che mok ines are defined by their ability to induce migra-tion of various leukocytepopulations (2). The chem otacticactivity of Ltn was m easured by dem onstra ting that Ltncan induce the migration of various lymp hocy te popula-tions. Human Ltn nduced chemotaxis inhuman CD8+and CD4+ PBL (Fig. 4,A and B ) . CD8+ Tcells wereoptimally responsive at a concentration of10”” M andCD4+ T cells at 10” MLtn.Human monocytes, how-ever, are not responsive to Ltn (Fig. 4C).
Ability of Ltn to induce a Ca2’ flux
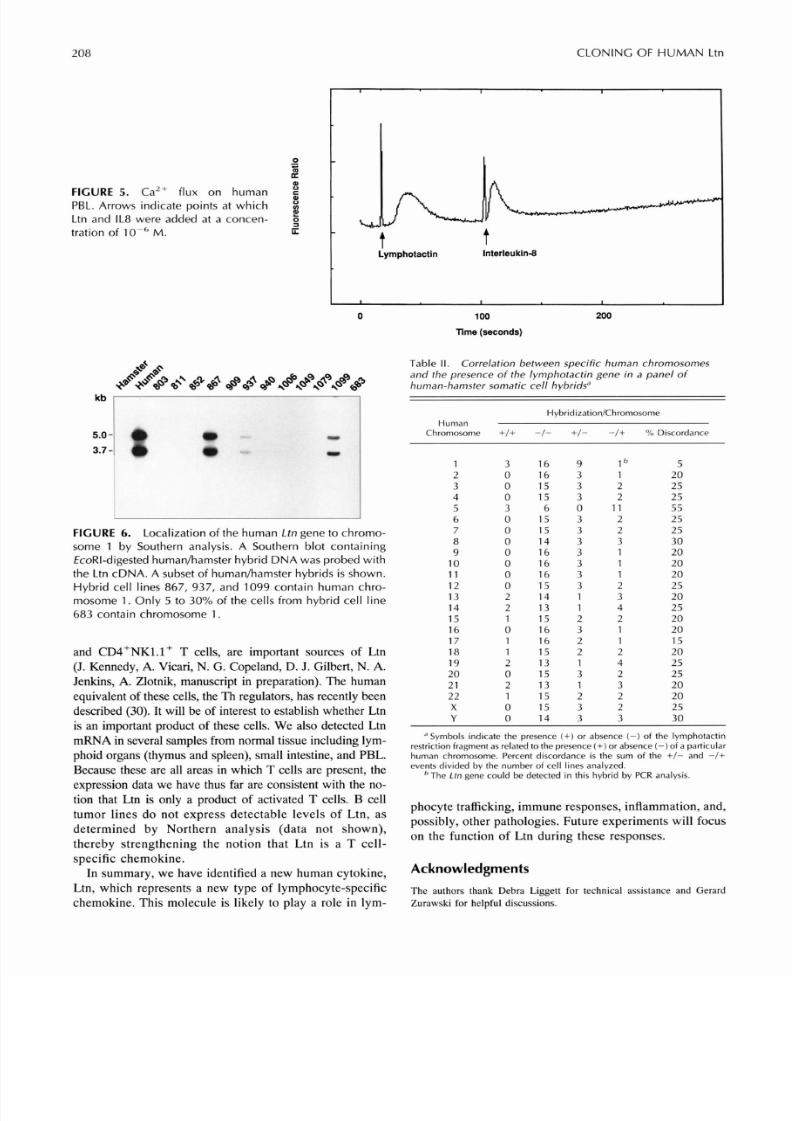
Th e measurement of an intracellular C a2+ flux is generallyregarded as an indicator of leukocyte receptor binding bychemokines, and it can be usedto predict whether variouschem okine s signal through th e sa me or different receptors(19). When recombinant Ltn wa s usedto measure ntra-cellular Ca 2+ fluxes in hum an PB L, a clear rise in intra-cellular C a2 + concentration w as observed (Fig.5) . In ad-dition, a second calcium lux was observed when IL-8 wasadded after Ltn treatment (Fig.5), which s ugge sts that Ltndoes not share the same signalingathway as IL-8. Similarresults w ere obtained when C-C chemokine s were addedafter Ltn treatment (K. Bacon, unpublished observations).
Chromosomal location of the Ltn gene
To determine the chrom osomal location of the humanLtngene, a Southern blot containing EcnRI-digested human/hamster hybrid DN A was probed w ith the Ltn cDNA . TheLtnprobedetectedapproximately 5- and 3.7-kbEcoRIband s and did not cross-hybridize with hamster DN A (Fig.6). Each som atic hybrid contains a known subset of humanchromosomes. On the basis of discordance values calcu-lated for each chromosom e (Table I), we can unambigu-ously assign the humanLtn gene to human chromosom e 1,although the DNA from hybrid 683, which contains hu-man chromosome 1 at a reduced frequency, did not hy-bridize to the Ltn cDN A. How ever, we onfirmed the pres-ence of the Ltn gen e in this hybrid by PCR analy sis (datanot shown).
Discussion
Previously, we have reported on the cloning and charac-terization of mouse Ltn and concluded hat this chemokinerepresented a new type of chemokine that did not f i t intoeither the C-C or the C-X-C chem okine family. The func-tional characteristics of Ltn, specifically, its abilityto che-moattract ymphocytes both T and B cells)but notmacrophages or monocytes, suggested thatt is a lympho-cyte-specific chemoattractant. Clearly, Ltn has the poten-tial of being an important molecule in the regulation oflymphocyte trafficking and inflammation. Therefore, itwa s very mportant to identifyahumanhomologue forLtn. Taking adv antage of the observation that mouse Ltn

7/27/2019 Immunol 1995 Kennedy 203 9
http://slidepdf.com/reader/full/immunol-1995-kennedy-203-9 4/7
206 CLONING OF HUMAN Ltn
FIGURE 2. Comparison of humanLtn protein to mouse Ltn and the hu-man C-C chemokine proteins MCP-2,macrophage inflammatory protein-1 a,
and RANTES. Boxed residues are ho-mologous between mouse and humanLtn.Black boxed residues epresentamino acids that are diagnostic of theC-C chemokine family. Shaded boxesrepresent esidues hatare homolo-gous between all sequences.
A+ +
u uZ
j i l iLtn
B
TiLtn
mLtnhLtnhMCP-2hMIP-lahRANTES
mLtnh l t nhMCP-2hMIP-lahRANTES
mLtnhLtn
C
p-actin
FIGURE 3. Northern analysis of Ltn mRNA expressed inlymphocyte subsets and various tissues. A , Total RNA from1 X lo6 sorted and activated CD4+ and CD8' thymocytes.B, Total RNA from unactivated and activated CD4+ andCD8' PBL. C, mRNA from resting spleen, thymus, prostate,testis, ovary, small ntestine, colon, and PBL.
is abundantly expressed by CD8+ T cells, we screened ahuman CD8+ T cell clone cDNA library and isolated sev-
eral clones. Nucleic acid and amino acid analyses con-
Q ~ K ~ T [ Q ~ N ~ V ~E T V P T G A Q R S T S T A I T L T G
firmed that these clones encoded human Ltn. Both humanand mouse Ltn, which hare 60% amino acid identity, lackthe first and third cysteine residues that would pair to formone of two disulfide bridges that define the C-C and C-X-Cchemokine structure (13).
In addition, the functional activities of human Ltn sup-
port the notion that it is a unique member of the chemo-kine superfamily. Human Ltn seems to be a specific che-moattractant for lymphocytes, or at least for T cells. It isimportant to note that, although Ltn shares some homol-ogy with C-C chemokines, it has lost the ability to che-moattract monocytes. Because he latter is an importantfunctional characteristic of C-C chemokines, this featuredistinguishes Ltn from this family functionally. The Ca2+flux studies provide an explanation for the functionaluniqueness of Ltn. Previous studies have established thatCa2+ fluxes provide a rapid readout of the potential ofchemokines to induce responses in target cells (19). After
such a response, cells become desensitized for a certainperiod of time. This effect provides an assay to determinethespecificity of chemokine-receptor interaction. Usingthis approach, we concluded that Ltn does not share thesame signaling pathway as IL-8 or the C-C chemokinestested. This finding suggests that Ltn uses a different re-ceptor than do the C-C and C-X-C chemokines.
Chromosomal mapping data provide additional evi-dence for the identification of a new class of chemokine.All the genes for the C-X-C chemokines that have beenreported havebeen localized to human chromosome 4(20-23). Likewise, all of the C-C chemokine genes that
have been described thus far have been found to be clus-tered on chromosome 17 (24-29). We have localized theLrn gene to human chromosome 1. It was not possible touse fluorescence in situ hybridization to map the region onchromosome 1, where the Lrn gene resides, because of thesmall size of the cDNA probe. However, mouse Ltn waslocalized by interspecific backcross analysis to a region onchromosome 1 that is syntenic with human l q (4), whichstrongly suggests that the human Lrn gene resides in thelq23-q25 region of human chromosome 1. The chromo-somal location of the Lrn gene further supports the notionthat Ltn does not belong o either the C-C or C-X-C
family.
7/27/2019 Immunol 1995 Kennedy 203 9
http://slidepdf.com/reader/full/immunol-1995-kennedy-203-9 5/7
The J ournal of immunology207
FIGURE 4. CD4+ and CD8+ T cells arechemotactic in response to Ltn. The num-ber of cells in the negative control of me-dium alone is indicated. A, Human C D8+PBL (medium = 61 i 20 cells). B, HumanCD4' T cell clone medium = 78 I 3cells). C, Freshly isolated human mono-cytes (medium = 111 i 8 cells).
L
L
Q)nL
Q)B
E
2501B -CD4+
p B L T
150, C. Monocytes
Factor concentration (M)
We studied he expression pattern of humanLtnand This result indicates thatCD4+ cells can, after differentiation,found activated CD8+ T cells to be an abundant sourceof become Ltn producers. Recently, we have observed thattwoLtn. It is interesting to note that we detected Ltn in acti- populations of mouse T cells likely to playa role in the reg-vated CD4+ PBL but not in activated CD4+ thymocytes. ulationof Thl or Th2 responses, the a/3-TCR+CD4-CDSP
7/27/2019 Immunol 1995 Kennedy 203 9
http://slidepdf.com/reader/full/immunol-1995-kennedy-203-9 6/7
20 8
FIGURE 5 . Ca2+ flux on humanPBL. Arrows indicate points at whichLtn and IL8 were added at a concen-tration of lo-' M.
CLONING OF HUMAN Ltn
-
2
-0
C0 )
M
0E
lz
t tLymphotactin Interleukina
I I
0
: a 0
0
FIGURE 6. Localization of the human Ltn gene to chromo-some 1 by Southern analysis. A Southern blot containingEcoRI-digested humanhamster hybrid DNA was probed withthe Ltn cDNA. A subset of humanhamster hybrids is shown.Hybrid cell lines 867, 937, and 1099 contain human chro-mosome 1. Only 5 to 30% of the cells from hybrid cell line683 contain chromosome 1 .
and CD4+NK1.1+ T cells, are mportant sources of Ltn(J. Kennedy, A. Vicari, N. G . Copeland, D. J. Gilbert, N. A.Jenkins, A. Zlotnik, manuscript in preparation). The humanequivalent of these cells, he Th regulators, has recently beendescribed (30). It will be of interest to establish whether Ltnis an important product of these cells. We also detected LtnmRNA in several samples from normal tissue including lym-phoid organs (thymus and spleen), small intestine, and PBL.Because these are all areas in which T cells are present, theexpression data we have thus far are consistent with the no-tion that Ltn is only a product of activated T cells. B celltumor lines do not express detectable levels of Ltn, asdetermined by Northern analysis data not shown),thereby strengthening the notion that Ltn is a T cell-specific chemokine.
In summary, we have identified a new human cytokine,Ltn, which represents a new type of lymphocyte-specific
chemokine. This molecule is likely to play a role in lym-
100 200
Time (seconds)
Table II. Correlation between specific human chromosomesand the presence of the lymphotactin gene in a panel ofhuman-hamster somatic cell hybrids"
Hybridization/Chromosome
Chromosome +/+ -I - +/- -/+ OO DiscordanceHuman
1 32 03 04 056 07 0
8 09 0
10 01 1 012 01314 215 116 017 118 11920 021 222 1X 0Y 0
16161515
61515
1416161615141315161615131513151514
9333033
33333112322131233
lb 51 202525
1 1 552525
301 201 201 20253 204 25201 201 152045253 202 20253 30
restriction fragment as related to the presence + I or absence (- )of a particular"Symbols indicate the presence (+) or absence (-) of the lymphotactin
human chromosome. Percent discordance is the sum of the +/- and -/+events divided by the number of cell lines analyzed.
'The L f n gene could be detected in this hybrid by PCR analysis.
phocyte trafficking, mmune responses, inflammation, and,possibly, other pathologies. Future experiments will focuson the function of Ltn during these responses.
Acknowledgments
The authors thank Debra Liggett for technical assistance and Gerard
Zurawski for helpful discussions.

7/27/2019 Immunol 1995 Kennedy 203 9
http://slidepdf.com/reader/full/immunol-1995-kennedy-203-9 7/7
The J ournal of Immunology 209
References
1. Oppenheim, J. J. , C. 0. Zachariae, N. Mukaida, and K. Matsushima.1991. Properties of the novel proinflammatory supergene “inter-crine” cytokine family.Annu. Rev. Immunol.9:617.
2. Schall,T. 994.The chemokines. In The Cytokine Handbook.A. Tho mson, ed. Academ ic Press Limited, New York, p. 419-460.
3. Cacalano, G., J. Lee, K. Kikly, A. Ryan,S. Pitts-Meek, B. Hultgren,W. Wood, and M. M oore. 199 4. Neutrophil and B cell expansion inmice that lack the murine IL-8 receptor homolog.Science 265:682.
4. Kelner, G. S., J. Kennedy, K.B. Bacon, S . Kleyensteuber, D. A.Largaespada, N.A. Jenkins, N. G. Cope land, J. F. Bazan ,K. W.Moore, T. J. Schall, and A. Zlotnik. 1994. Lymphotactin: a novelcytokine which represents a new class of chemokine.Science. 266:1395.
5. Orlofsky, A., M. S. Berger, and M. B. Prystowsky. 1991 . Novelexpression pattern of a new member of the MIP-1 family of cyto-kine-like genes. Cell Regul. 2:403.
6. Jose, P., D. Griffiths-Johnson, P. Collins, D. Walsh, R. Moqbel,N. Totty, 0 . Truong, J . Hsuan, and T. Williams. 1994. Eotaxin: apotent eosinophil chemoattractant cyto kine detected in a guinea pigmodel of allergic airways inflammation.J . Exp. Med. I79:881.
7. Cocks, B. G., R. de Waal Malefyt, J. P. Galizzi,J . E. de Vries, and
G. Aversa. 1993. IL-13 induces proliferation and differentiation ofhuman B cells activated by the CD40 ligand.Int. Immunol. 5:657.
8. Maniatis, T., E. F. Fritsch, and J . Sambrook. 1982.Molecular Clon-ing: A Laboratory Manual. Cold Spring Harbor LaboratoryPress,Cold Spring Harbor, NY.
9. Boyum, A . 1968 . Separation of leukocytes from blood and bonemarrow. Scand. J. Clin. Lab. Invest. 2I(Suppl. 97):77.
10. Yssel, H., J . E. de Vries, M. Koken, W. Van Blitterswijk, andH. Spits. 1984. Serum-free medium for the generation and propaga-tion of functional human cytotoxic and helperT cell clones.J. Immunol. Methods 72:219.
11. Godfrey, D., J. Kennedy, T. Suda, and A. Zlotnik. 1993. A devel-opm ental pathway involving four phenotypically and functionallydistinct subsets of CD3-C D4C CD8 - triple-negative adult mousethymocytes defined by CD44 and CD25 expression.J . Immunol.150t4244.
12. Bacon, K. B., R. D. Camp, F. M. Cunningham, and P. M. Woollard.1988. Contrasting in vitro lymphocyte chemotactic activity of thehydroxl enantiomers of 12-hydroxy-5,8,10,14-eicosatetraenoic cid.Br . J. Pharmacol. 95t966.
13. Lodi, P., D. Garrett, J. Kuszewski, M. Tsang, J. Weatherhee,W. Leonard, A. Gronenborn, and G. Clore. 1994. High-resolutionsolution structure of the beta chem okine hMIP-1 beta by multidi-mensional NMR. Science 263:1762.
14. Rollins, B. J., E. D. Morrison, and C. D. Stiles. 1988. Cloning andexpression of JE , a gene inducible by platelet-derived growth factorand whose product has cytokine-like properties.Procy Natl. Acad.Sci. USA 85:3738.
15. Chang, H. C., F. Hsu, G. J. Freeman, J . D. Griffin, and E. L. R ein-herz. 1989. Cloning and expressionof a y-interferon-inducible ge nein monocytes: a new member of a cytokine gene family.Int. Immu-nol. 1:388.
16. Furutani, Y., H. Nomura, M. Notake, Y. Oyamada, T. Fukui,M. Yamada, C. G. Larsen, J. J. Oppenheim, andK. Matsushima.1989 . Cloning and sequencing of the cDN A for human m onocyte
chemotactic and activating factor (MCAF).Biochem. Biophys. Res.Commun. 159t249.
17. Yoshimura, T., N. Yuhki, S. K. Moore, E. Appella, M. I. Lerman,and E. J . Leonard. 1989. Human mo nocyte chemoattractant protein-1(MCP-1): full-length cDNA cloning, expression in mitogen-stimu-lated blood mononuclear leukocytes, and sequence similarity tomouse competent geneJE . FEBS Lett. 244:487.
18. Farber, J. M. 1993.HuMIG: a new hum an member of the chemokinefamily of cytokines. Biochem. Biophys. Res. Commun.192:223.
19. Neote, K., G. D. Di, J. Y. Mak, R. Horuk, and T. J. Schall. 1993.Molecular cloning, functional expression and signaling characteris-tics of a C-C chemokine receptor.Cell 72t415.
20. Griffin, C. A.,B. S. Emanuel, P. LaRocco, E. Schwartz, andM. Ponez. 1987. Human platelet factor 4 gene is mapped to 4q12-q21. Cytogenet. Cell Genet.45:67.
21. Luster, A,,S . Jhanwar, R. Chaganti,J. Kersey, and J. Ravetch. 1987.Interferon inducible gene maps to a chromosome band associatedwith a (4;11) translocation in acute leukemia cells.Proc. Nafl .Acad.Sci. USA84:2868.
22. Richmond, A,, E. Balentien, H.G. Thomas, G. Flaggs, D. E. Barton,J. Spiess, R. Bordoni, U. Francke, and R. Derynck. 1988. Molecularcharacterization and chromosomal mapping of melanoma growthstimulatory activity, a grow th factor structurally related to P-throm-boglobulin. EMBO J. 7:2025.
23. Modi, W. S., M. Dean, H.N . Seuanez, N. Mukaida, K. Matsushima,and S. J. O’Brien. 1 990. Monocyte-derived neutrophil chemotacticfactor (MDNCF/IL-8) resides in a gene cluster along with severalother members of the platelet factor 4 gene superfamily.Hum. Gene?.84: 185.
24. Irving, S . G., P. F. Zipfel, J. Balke, 0. W. McBride, C. C. Morton,P. R. Bud, U. Siehenlist, and K. Kelly. 1990 . Two inflammatorymediator cytokine gen es are closely linked and variably amplified onchromosome 17q.Nucleic Acids Res. 18:326I.
25. Miller, M. D., S . D. Wilson, M. E. Dorf, H.N. Seuanez, S. J.O’Brien, and M. S. Krangel. 1990. Sequence and chromosomallo-cation of the I-309 gene: relationship to genes encoding a family ofinflammatory cytokines.J. Immunol. 145:2737.
26. Rollins, B.J., C. C. Morton, D. H. Ledbetter, R. L.J. Eddy, and T. B.Shows. 1991. Assignmentof the human small inducible cytokine A2gene, SCYA2 (encoding JE or M C P - I ) , to 17q11.2-12: evolutionaryrelatedness of cytokines clustered at the same locus.Genomics 10 :489.
27. Donlon, T. A., A. M. Krensky, M.R. Wallace, F. S . Collins,M. Lovett, and C. Clayberger. 1990 . Localization of a human T-cell-specific gene,RANTES(D17S136E), to chromosome 17q11.2-ql2.Genomics 6:548.
28. Napolitano, M., W. S. Modi, S. J. Cevario, J . R. Gnarra, H. N.Seuanez, andW. J. Lenoard. 19 91. The gene encoding the Act-2cytokine: genomic structure, HTLV-1max responsiveness of5’ up -
266: 753 .stream sequences, and chromosomal localization.J. Biol. Chem.
29. Hirashima, M., T . Ono, M. Nakao, H. Nishi, A. Kimura,H. Nomiyama, F. Hamada, M. C. Yoshida, and K. Shimada. 1992.Nucleotide sequence of the third cytokine LD78 and mapping of allthree LD78 gene loci to human chromosome 17.DNA Seq.3:203.
30. Lantz, O. , and A. Bendelac. 1994.An invariant T cell receptor achain is used by a unique subsetof major histocompatibility complexclass I-specific CD 4+ and CD 4-8- T cells in mice and humans.J. Exp. Med. 180:1097.


















