
Identification of two antibody-interaction sites on the surface of Panulirus interruptus hemocyanin
7
Eur. J. Biochem. 222, 155-161 (1994) 0 FEBS 1994 Identification of two antibody-interaction sites on the surface of Panulirus interruptus hemocyanin Felix de HAAS', Frank G. PERTON2, Jan F. L. van BREEMENl, Jan H. DIJKE?MA2, Jaap J. BEINTEMA' and Ernst F. J. van BRUGGEN' BIOSON Research Institute, Rijksuniversiteit Groningen, The Netherlands Biochemisch Laboratorium, Rijksuniversiteit Groningen, The Netherlands (Received September 2November 12, 1993) - EJB 93 1331/3 Negatively stained complexes of Panulirus interruptus (spiny lobster) hemocyanin with two different monoclonal antibodies, named E and J, were studied by electron microscopy and image processing. The attachment site of the antibodies to the hexameric hemocyanin molecule was de- duced from two perpendicular views of hernocyanidantibody complexes, in which either the three- fold axis or one of the twofold axes was oriented perpendicular to the supporting film. Images of complexes in these orientations were searched with reference images simulated from the known X-ray structure of P. interruptus hemocyanin. The two sites were further characterized by combining our results from electron microscopy with structural data obtained by X-ray diffraction and other methods. These two antibodies recognize different non-overlapping epitopes. The epitope for clone E is located on domain 3 at the surface of the p barrel and consists of certain loops, which form connections between /3-strand structures. The epitope for clone J is situated on domain 1 at the surface of an a-helical region and consists mainly of certain a-helices connecting loops. The orientation of the hemocyanin hexamers in the two complexes is very different, as is demon- strated most clearly when they form chains. Clone E forms complexes with the threefold axes perpendicular to the chain direction, while for clone J the threefold axes seem to be parallel to the main direction. The angle between the Fab part of an IgG molecule and the threefold axis of the hexamer is 60 5 5" for clone E and 35 5 7" for clone J. This observation is clearly related to the difference in orientation of the hexamers for the two complexes. Hemocyanin is an extracellular, copper containing pro- tein that occurs in high concentrations (10-100 mg/ml) in the blood of molluscs and certain arthropods belonging to the chelicerata (spiders, scorpions, horseshoe crabs) and crus- tacea (spiny lobsters, crabs; Van Holde and van Bruggen, 1971; Van Holde and Miller, 1982; Ellerton et al., 1983; Markl and Decker, 1991). It serves as an oxygen carrier, functionally similar to hemoglobin, but binds oxygen be- tween two copper ions. The architecture of arthropod hemo- cyanin multimers consists of discrete assemblies of hexam- eric building blocks (Mr approximately 450000). The three- dimensional X-ray structures of the hexameric hemocyanins from the spiny lobster (Panulirus interruptus ; Gaykema et al., 1984, 1986; Volbeda and Hol, 1989) and of subunit I1 from the horseshoe crab (Limulus polyphemus; Hazes et al., 1993) are known in detail. Roughly, six bean-shaped subunits (Mr approximately 75000) occupy the corners of a trigonal antiprism. Hemocyanin has powerful antigenic properties used in many medical tests. Our interest concerns the structural fea- tures which cause these strong immunogenic reactions. Correspondence to E. F. J. van Bruggen, BIOSON Research Institute, Rijksuniversiteit Groningen, Nijenborgh 4, NL-9747 AG Groningen, The Netherlands Fax: +31 50 634800. Therefore, monoclonal antibodies against P. interruptus he- mocyanin have been made for localization and characteriza- tion of the antigenic areas on its protein surface. P. interrup- tus hemocyanin was chosen because of our knowledge of its tertiary, secondary and primary structure. Immuno-electron microscopy was used for this localization. This method has supplied important information for other protein assemblies like the ribosome (Stoffler-Meilicke and Stoffler, 1987) or viruses (Wang et al., 1992). Electron microscopy of the multi-hexameric arthropod hemocyanins has yielded much insight relating to their quaternary structures (Lamy et al., 1980, 1981, 1982; Markl et al., 1981 ; Bijlholt et al., 1982; Boisset et al., 1990; de Haas et al., 1991) and the methods developed for these investigations can also be applied to studies of interactions of hemocyanins with antibodies. A study by Lamy et al. (1990) revealed four different epitopes by a combination of electron microscopy and classical immu- nological mapping for 13 different monoclonal antibodies binding to subunit Aa6 of scorpion (Androctonus australis) hemocyanin. One epitope was not accessible for antibody binding in the native four-hexameric molecule. The other three epitopes were identical to those found by immuno- electron microscopy (Boisset et al., 1988). A drawback of the latter study is the absence of sequence data and an X-ray structure of A. australis hemocyanin.
-
Upload
felix-de-haas -
Category
Documents
-
view
212 -
download
0
Transcript of Identification of two antibody-interaction sites on the surface of Panulirus interruptus hemocyanin

Eur. J. Biochem. 222, 155-161 (1994) 0 FEBS 1994
Identification of two antibody-interaction sites on the surface of Panulirus interruptus hemocyanin Felix de HAAS', Frank G. PERTON2, Jan F. L. van BREEMENl, Jan H. DIJKE?MA2, Jaap J. BEINTEMA' and Ernst F. J. van BRUGGEN'
BIOSON Research Institute, Rijksuniversiteit Groningen, The Netherlands Biochemisch Laboratorium, Rijksuniversiteit Groningen, The Netherlands
(Received September 2November 12, 1993) - EJB 93 1331/3
Negatively stained complexes of Panulirus interruptus (spiny lobster) hemocyanin with two different monoclonal antibodies, named E and J, were studied by electron microscopy and image processing. The attachment site of the antibodies to the hexameric hemocyanin molecule was de- duced from two perpendicular views of hernocyanidantibody complexes, in which either the three- fold axis or one of the twofold axes was oriented perpendicular to the supporting film. Images of complexes in these orientations were searched with reference images simulated from the known X-ray structure of P. interruptus hemocyanin.
The two sites were further characterized by combining our results from electron microscopy with structural data obtained by X-ray diffraction and other methods. These two antibodies recognize different non-overlapping epitopes. The epitope for clone E is located on domain 3 at the surface of the p barrel and consists of certain loops, which form connections between /3-strand structures. The epitope for clone J is situated on domain 1 at the surface of an a-helical region and consists mainly of certain a-helices connecting loops.
The orientation of the hemocyanin hexamers in the two complexes is very different, as is demon- strated most clearly when they form chains. Clone E forms complexes with the threefold axes perpendicular to the chain direction, while for clone J the threefold axes seem to be parallel to the main direction. The angle between the Fab part of an IgG molecule and the threefold axis of the hexamer is 60 5 5" for clone E and 35 5 7" for clone J. This observation is clearly related to the difference in orientation of the hexamers for the two complexes.
Hemocyanin is an extracellular, copper containing pro- tein that occurs in high concentrations (10-100 mg/ml) in the blood of molluscs and certain arthropods belonging to the chelicerata (spiders, scorpions, horseshoe crabs) and crus- tacea (spiny lobsters, crabs; Van Holde and van Bruggen, 1971; Van Holde and Miller, 1982; Ellerton et al., 1983; Markl and Decker, 1991). It serves as an oxygen carrier, functionally similar to hemoglobin, but binds oxygen be- tween two copper ions. The architecture of arthropod hemo- cyanin multimers consists of discrete assemblies of hexam- eric building blocks (Mr approximately 450000). The three- dimensional X-ray structures of the hexameric hemocyanins from the spiny lobster (Panulirus interruptus ; Gaykema et al., 1984, 1986; Volbeda and Hol, 1989) and of subunit I1 from the horseshoe crab (Limulus polyphemus; Hazes et al., 1993) are known in detail. Roughly, six bean-shaped subunits (Mr approximately 75000) occupy the corners of a trigonal antiprism.
Hemocyanin has powerful antigenic properties used in many medical tests. Our interest concerns the structural fea- tures which cause these strong immunogenic reactions.
Correspondence to E. F. J. van Bruggen, BIOSON Research Institute, Rijksuniversiteit Groningen, Nijenborgh 4, NL-9747 AG Groningen, The Netherlands
Fax: +31 50 634800.
Therefore, monoclonal antibodies against P. interruptus he- mocyanin have been made for localization and characteriza- tion of the antigenic areas on its protein surface. P. interrup- tus hemocyanin was chosen because of our knowledge of its tertiary, secondary and primary structure. Immuno-electron microscopy was used for this localization. This method has supplied important information for other protein assemblies like the ribosome (Stoffler-Meilicke and Stoffler, 1987) or viruses (Wang et al., 1992). Electron microscopy of the multi-hexameric arthropod hemocyanins has yielded much insight relating to their quaternary structures (Lamy et al., 1980, 1981, 1982; Markl et al., 1981 ; Bijlholt et al., 1982; Boisset et al., 1990; de Haas et al., 1991) and the methods developed for these investigations can also be applied to studies of interactions of hemocyanins with antibodies. A study by Lamy et al. (1990) revealed four different epitopes by a combination of electron microscopy and classical immu- nological mapping for 13 different monoclonal antibodies binding to subunit Aa6 of scorpion (Androctonus australis) hemocyanin. One epitope was not accessible for antibody binding in the native four-hexameric molecule. The other three epitopes were identical to those found by immuno- electron microscopy (Boisset et al., 1988). A drawback of the latter study is the absence of sequence data and an X-ray structure of A. australis hemocyanin.

156
MATERIALS AND METHODS Isolation and immunization
Panulirus hemocyanin was isolated as described by van Eerd and Folkerts (1981) and by Neuteboom et al. (1992). Monoclonal antibodies were raised by the standard lympho- cyte hybridoma technique (Perton et al., 1990). Spleen cells of BALBlc mice immunized intraperitoneally with intact he- mocyanin in complete Freund's adjuvant were fused with P,X6,Ags6, myeloma cells in the presence of poly(ethy1ene) glycol. The hybridomas were selected in hypoxanthine, aminopterin, thymidine medium. Positive clones were iden- tified by the ELISA procedure in which the hemocyanin was attached to the microtiter plate. Only hybridomas secreting IgG were selected because the conjugate in the ELISA was IgG specific. IgG was purified from ascitic fluid by affinity chromatography with protein-A- Sepharose according to the method described by the supplier (Pharmacia).
Preparation of Fab fragments and determination of hemocyanidantibody complex dissociation constants
Fab fragments were prepared from IgG molecules after cleavage with papain (Boehringer) according to Goding (1986). Fab fragments were separated from Fc fragments and non-digested immunoglobulins by protein-A-Sepharose affinity chromatography.
Dissociation constants for antibodylantigen complexes were determined by the method of Friguet et al. (1985).
Immuno-electron microscopy Complexes of immunoglobulins or Fab fragments with
hemocyanin were isolated after incubating at 37 "C for 30 min at several ratios of antibodylantigen. The complexes were purified over a 160 mmX4.5 mm column of Sepharose CL-6B (Pharmacia). The complexes were prepared for electron microscopy by the droplet technique with uranyl acetate as the negative stain. Micrographs were taken with Philips CM12 and Jeol 1200EX electron microscopes at a magnification of 60000 for hemocyanin-IgG (E) and hemo- cyanin-Fab (J), but at a magnification of 45000 for hemo- cyanin-IgG (J). Micrographs were recorded on an Agfa Scientia EM film (23D56 P3 AH) and developed in Kodak HRP I11 developer.
Image processing and image analysis Electron micrographs showing unambiguously identifi-
able complexes were digitized using a Kodak EIKONIX camera. The sampling step size was 25 pm corresponding to a step size of approximately 0.42 nm (clone E) and 0.56 nm (clone J) at the specimen level. The dimensions (in pixels) of the images were large enough to contain a hemocyanin molecule with all bound Fab fragments. Fc fragments were disregarded in the case of complexes with IgG. Image analy- sis was performed using the IMAGIC software package orig- inally developed by van Heel and Keegstra (1981). The se- lected images were subjected to a multiple-reference align- ment procedure (Harauz et al., 1988; Boekema and Bottcher, 1992). Twelve reference images were calculated from the l? interruptus hemocyanin X-ray structure following the pro- cedure of de Haas et al. (1991). These images contain projec- tions of the hexamer in various orientations. The calculated reference images were treated with a band-pass filter which
Fig. 1. Electron micrographs of hemocyanidantibody complexes stained with 1 % uranyl acetate. (A) Network of hemocyanin mol- ecules with IgG molecules from clone E with views mainly along the threefold (hexagons) and twofold (rectangles) axes. (B) A ladder structure showing rectangular hemocyanin profiles connected with IgG molecules from clone E. (C) Complexes of hemocyanin mol- ecules with complete IgG molecules from clone J. (D) A gallery of images of complexes of hernocyanin molecules with Fab fragments from clone J. The bar indicates 25 nm.
allows details to pass within the range 1-25 nm. This is in accordance with details present in the electron micrographs. Before multiple-reference alignment, the images containing the complexes of hemocyanin with complete IgG or single Fab fragments were treated with a circular mask in such a way that the antibody parts are temporarily removed and only the hemocyanin molecule is involved in the alignment. The densities in the images were normalized to an average of zero and to a variance of ten. A band-pass filter was applied by which spatial frequencies with wavelengths above 25 nm and below 1 nm were suppressed. In this way, the effects caused by noise and uneven staining are reduced without loss of relevant structural information. Once the images are aligned, they are grouped according to their corresponding reference image and an averaged image is calculated for each group. Before averaging, the mask is removed so that the hemocyanin molecule plus the attached antibody parts be- come visible again.
RESULTS AND DISCUSSION Electron microscopy
Sixteen stable clones that produced antibodies recogniz- ing €? interruptus hemocyanin were isolated. Almost all the clones had the highest affinity with the a chain of this hemo- cyanin. To check whether the antibodies reacted with native hemocyanin in the hexameric form, complexes were investi- gated by electron microscopy. Only for the two clones, named E and J, were sufficient numbers of images of unam- biguously identifiable complexes obtained which were suita- ble for image analysis. Fig. 1 shows projections of complexes

157
of hemocyanin with IgG from clones E and J. The character- istic views of the negatively stained P. interruptus hemocya- nin hexamer are visible as hexagonal and rectangular projec- tions. Fig. 1A shows for clone E a small network of these views. Another typical 'ladder' or 'chain' structure, also formed by hemocyanin and clone E molecules, is observed in Fig. 1 B. For electron micrographs of complexes with clone J (Fig. 1 C), these large network structures were not observed. In these electron micrographs, the most abundant characteris- tic structure is a complex of two hemocyanins with two IgG molecules in between the hemocyanins. It was difficult to find hexagonal views in complexes with clone J. Those com- plexes showed in most cases the rectangular view or a com- plex representing a structure in-between a hexagonal and rec- tangular view. The complexes for clone E and for clone J show a difference in the orientation of the threefold axis of the hemocyanin hexamer. In the chain-like structures of com- plexes with clone E, the threefold axes are perpendicular to the direction of the chain. The complexes with clone J repre- sent a parallel arrangement of the threefold axis with respect to their main axis.
For a detailed characterization of the attachment site of the antibodies on the surface of hemocyanin, the complete structure of a hemocyanidantibody complex is required. The position of the Fab parts on the hemocyanin surface would be deducable from two mutually perpendicular views. For this reason, we looked only for views along the threefold axis (hexagons) and along a twofold axis (rectangles). Aiming for more views along the threefold axis, additional studies were performed for clone J with Fab fragments obtained by cleav- age of IgG with papain. Such complexes were expected to have more rotational freedom.
Image analysis
Complexes with clone E
Images of 560 complexes were selected from six electron micrographs. The images were made large enough to contain one hemocyanin molecule (in the center) with the Fab parts of the IgG molecules attached to it. These images were pre- viously treated as described earlier and aligned to the twelve reference images (Fig. 2) calculated from the l? interruptus hemocyanin X-ray structure. This resulted in a distribution into twelve groups classified by the value of the correlation coefficient with respect to each of the twelve reference images. For these groups averaged images were calculated by using the images in which the IgG parts were added again (Fig. 3 A -L). For further processing, those averaged images were selected which showed the following properties: (a) hemocyanin in hexagonal or rectangular view; (b) IgG com- ponents distinguishable from the background.
The averaged images of Fig. 3A and B show indeed a clear hexagonal view, but the antibody parts can hardly be detected against the background. This can be caused by the use of a hexagon as the only reference image for the align- ment, as shown in Fig. 2A. Therefore, we re-aligned these two groups together using as reference an image from the group itself consisting of a complex of a hexagon and an Fab part. The resulting averaged image (Fig. 5A), clearly shows a hexagon with an antibody part attached to a so-called pointed protrusion corresponding with the projection of two superim- posed domains 3 (Wichertjes et al., 1989). The reference de- pendence in these cases may be considerable, but the preser- vation of threefold symmetry in the hexamer and also in the
Fig. 2. Calculated projections. Twelve projections calculated from the f! interruptus hemocyanin X-ray structure which served as refer- ences for the alignments. Projections have been band-pass filtered with a filter allowing details to pass within the range 1-25 nm. The X-axis runs vertically and the Y-axis runs horizontally in the plane of the paper through the center of the hexamer. (A-F) Hexamer rotated around the X-axis in steps of 12" starting from an orientation with the threefold axis perpendicular to the XY plane and one two- fold axis coinciding with the X-axis (A = Oo, B = 1 2 O , C = 24O, D = 36", E = 48°F = 60"). (G-L) Hexamer rotated around the X- axis in steps of 12" but starting from an orientation with a twofold axis perpendicular to the XY-plane and the threefold axis coinciding with the X-axis (G = 0", H = 12", I = 24", J = 36", K = 48", L = 60").
attachment of the antibody shows us that this dependency is not severe. Two other similar contact areas, related by three- fold symmetry, are observed though with less contrast.
Fig. 3L shows a rectangular view observed (average of 167 images) along a twofold axis with IgG parts attached to all four comers. Looking at the number of molecules in this class, this seems to be a preferential orientation for these complexes. At the comers of this rectangular projection both domains 1 and 3 overlap.
Figs 5A and B summarize both mutually perpendicular views from which the attachment sites of the antibody can be deduced. Fig. 5B is identical to Fig. 3L.
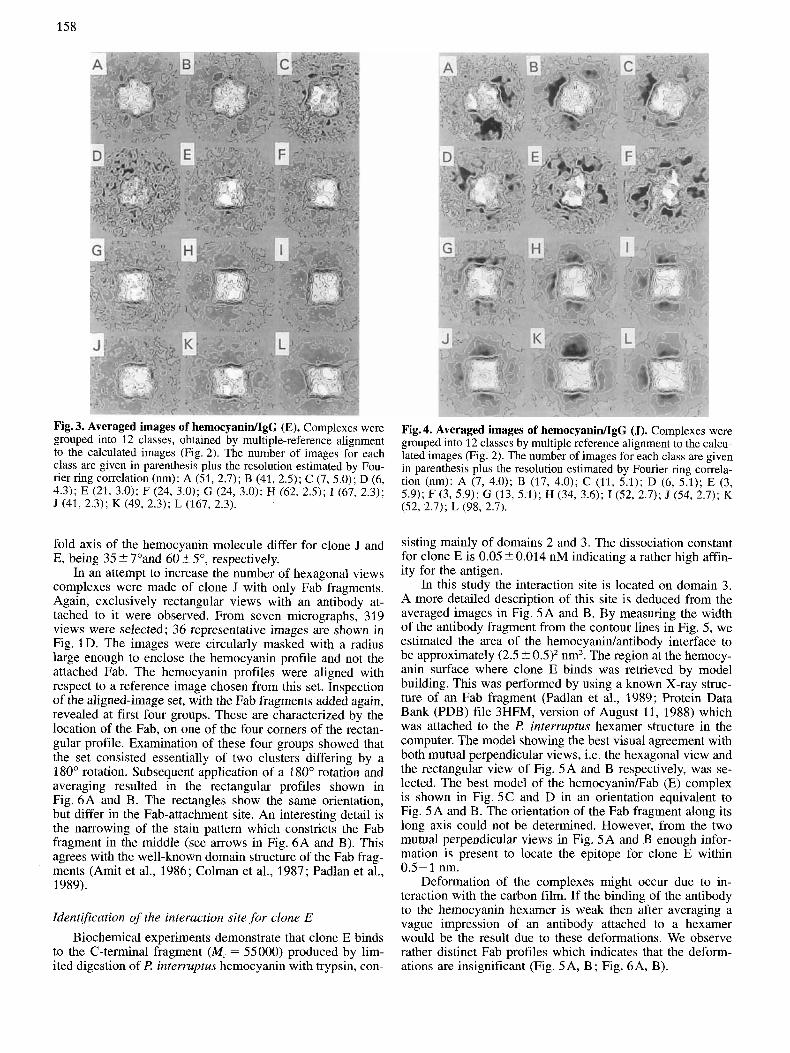
Complexes with clone J Images of 350 complexes with clone J were selected from
eight electron micrographs. Multi-reference alignment to the twelve (calculated) reference images (Fig. 2) resulted after addition of the IgG parts in the averaged images of Fig. 4A- L. No clearly interpretable hexagonal view of the complex is visible and the group sizes are small. The averages shown in Fig. 4A and B, calculated from 7 and 17 images, respec- tively, give the impression of a tilted complex. Fig. 4A, if compared to Fig. 2A, might indicate an attachment of an an- tibody part at a so-called broad protrusion, the projection of two overlaping domains 1. The rectangular view (Fig. 4L) shows the attachment of antibody parts at all four comers. The angles between the direction of attachment and the three-
158
Fig. 3. Averaged images of hemocyaniflgG (El. Complexes were grouped into 12 classes, obtained by multiple-reference alignment to the calculated images (Fig. 2) . The number of images for each class are given in parenthesis plus the resolution estimated by Fou- rier ring correlation (nm): A (51, 2.7); B (41, 2.5); C (7,5.0); D (6, 4.3); E (21, 3.0); F (24, 3.0); G (24, 3.0); H (62, 2.5); I (67, 2.3); J (41, 2.3); K (49, 2 .3 ) ; L (167, 2.3).
Fig. 4. Averaged images of hemocyanidIgG (J). Complexes were grouped into 12 classes by multiple reference alignment to the calcu- lated images (Fig. 2). The number of images for each class are given in parenthesis plus the resolution estimated by Fourier ring comela- tion (nm): A (7, 4.0); B (17, 4.0); C (11, 5.1); D (6, 5.1); E (3, 5.9); F (3, 5.9); G (13, 5.1); H (34, 3.6); I(52, 2.7); J (54, 2.7); K (52, 2.7); L (98, 2.7).
fold axis of the hemocyanin molecule differ for clone J and E, being 35 ? 7"and 60 2 5", respectively.
In an attempt to increase the number of hexagonal views complexes were made of clone J with only Fab fragments. Again, exclusively rectangular views with an antibody at- tached to it were observed. From seven micrographs, 319 views were selected; 36 representative images are shown in Fig. 1D. The images were circularly masked with a radius large enough to enclose the hemocyanin profile and not the attached Fab. The hemocyanin profiles were aligned with respect to a reference image chosen from this set. Inspection of the aligned-image set, with the Fab fragments added again, revealed at first four groups. These are characterized by the location of the Fab, on one of the four corners of the rectan- gular profile. Examination of these four groups showed that the set consisted essentially of two clusters differing by a 180" rotation. Subsequent application of a 180" rotation and averaging resulted in the rectangular profiles shown in Fig. 6A and B. The rectangles show the same orientation, but differ in the Fab-attachment site. An interesting detail is the narrowing of the stain pattern which constricts the Fab fragment in the middle (see arrows in Fig. 6A and B). This agrees with the well-known domain structure of the Fab frag- ments (Amit et al., 1986; Colman et al., 1987; Padlan et al., 1989).
Identification of the interaction site for clone E Biochemical experiments demonstrate that clone E binds
to the C-terminal fragment (Mr = 55000) produced by lim- ited digestion of l? interruptus hemocyanin with trypsin, con-
sisting mainly of domains 2 and 3. The dissociation constant for clone E is 0.05 ? 0.014 nM indicating a rather high affin- ity for the antigen.
In this study the interaction site is located on domain 3. A more detailed description of this site is deduced from the averaged images in Fig. 5A and B. By measuring the width of the antibody fragment from the contour lines in Fig. 5, we estimated the area of the hemocyanidantibody interface to be approximately (2.5 -+_ 0.5)' nm'. The region at the hemocy- anin surface where clone E binds was retrieved by model building. This was performed by using a known X-ray struc- ture of an Fab fragment (Padlan et al., 1989; Protein Data Bank (PDB) file 3HFM, version of August 11, 1988) which was attached to the l? interruptus hexamer structure in the computer. The model showing the best visual agreement with both mutual perpendicular views, i.e. the hexagonal view and the rectangular view of Fig. 5A and B respectively, was se- lected. The best model of the hemocyanin/Fab (E) complex is shown in Fig. 5C and D in an orientation equivalent to Fig. 5A and B. The orientation of the Fab fragment along its long axis could not be determined. However, from the two mutual perpendicular views in Fig. 5A and B enough infor- mation is present to locate the epitope for clone E within 0.5-1 nm.
Deformation of the complexes might occur due to in- teraction with the carbon film. If the binding of the antibody to the hemocyanin hexamer is weak then after averaging a vague impression of an antibody attached to a hexamer would be the result due to these deformations. We observe rather distinct Fab profiles which indicates that the deform- ations are insignificant (Fig. 5A, B; Fig. 6A, B).

159
Fig. 5. Averaged images of the two mutually perpendicular pro- jections of the hemocyanidIgG (E) complex. (A) Antibodies at- tached to a hexagonal profile. The resolution determined by Fourier ring correlation is 2.7 nm. (B) Antibodies attached to a rectangular profile. This figure is identical to Fig. 3L with a resolution by Fou- rier ring correlation of 2.3 nm. (C) An C,-atom backbone model of the deduced hemocyanin/Fab complex in a view along the threefold axis of the hexamer with the Fab fragment attached to domain 3 . (D) The same model as in C but viewed along a twofold axis of the hexamer.
The site region is visualized in more detail in Fig. 7. One subunit of P. interruptus hemocyanin is shown in C,-atom backbone representation. Loops at the surface of the interac- tion site (Fig. 7) are indicated. From the studies of Geysen et al. (1987) and Getzoff et al. (1987), convex-shaped protein surfaces seem to be amongst the favorite sites for antibody recognition. Therefore, the convex shape of the p barrel in domain 3 may be important. The loops between p3. D and p3. E, between p3. G and p3. H, and a part of the loop between (13.1 and p3. A are clearly candidates for contact with the antibody. After inspection of the type of amino acids present in these regions, the loop p3. DID. E seems to con- tain a higher fraction of charged residues. Other high-resolu- tion X-ray studies concerning antigen/antibody complexes (Amit et al., 1986; Colman et al., 1987) showed that the van der Waals interactions with hydrogen bonds are important for the interaction.
A similar loop between p3. D and p3. E is involved in the interdodecameric interaction in the four-hexameric mole- cules of Eurypelma californicum (tarantula) hemocyanin (de Haas, 1993) and is part of a region in the polypeptide chain of arthropod hemocyanins with considerable variability in amino acid sequence (Linzen et al., 1985; Hazes et al., 1993).
Identification of the interaction site for clone J
Biochemical evidence demonstrated that clone J interacts with the N-terminal fragment (domain 1, M, = 18000) of R interruptus hemocyanin. Clone J has a high affinity for the antigen; the dissociation constant is 0.12 -+ 0.02 nM. To be able to build a model, as for clone E, we used the views
Fig. 6. Averaged images of the hernocyaniflab (J) complexes. (A) Fab fragment attached at the upper-left comer. The resolution as determined by Fourier ring correlation is 3.8 nm. (B) Fab fragment attached at the upper-right comer. The resolution by Fourier ring correlation is 3.8 nm. (C) A modelled hemocyanin/Fab structure in C,-atom backbone representation viewed along the threefold axis with the Fab attached to domain 1. (D) The same model as in C but viewed along a twofold axis. The arrows point to the space between variable and constant domains of the fragment.
depicted in Fig. 4A and L, Fig. 6A and B, plus the additional biochemical information, as mentioned above. The final de- duced model is shown in Fig.6C and D. The model in Fig. 6C compares with the view in Fig. 4A. A better agreement is noticed between Fig. 6D and Fig. 6A. Rotation of the complex in Fig. 6D through 120" along the threefold axis of the hexamer gives a view (data not shown) compara- ble to Fig. 6B. Fig. 4L can be considered to be a superposi- tion of views from Fig. 6D and its rotational equivalents. We conclude that domain 1 is the candidate for the clone-J site. The loops at the flat surface of this domain, indicated in Fig. 7, are possibly involved in antibody binding. These are the N-terminus going into al.1, a loop connecting a1.2 to al.3, the loops between al.4 and al.5, and between al.6 and pl. A, and the C-terminal part of a3.3. Of these loops, a1.4/ a1.5 contains more charged residues and is in the center of the epitope.
Conclusions The sites of interaction of two monoclonal antibodies,
named E and J, were localized on the surface of P. interrup- tus hemocyanin by combining results from electron micro- scopy, X-ray diffraction and protein chemistry. These anti- bodies recognize different non-overlapping epitopes. The epitope for clone E is located on domain 3 at the surface of the p barrel and consists of loops, which form connections between certain /3-strand structures. The epitope for clone J is situated on domain 1 at the surface of an a-helical region and consists of loops connecting some of these a helices. In the center of the epitopes, loops are located with a higher

160
Fig. 7. Antibody-binding sites. Stereo drawing showing the epitopes for clones E and J on the surface of a monomer of the P. interruptus hemocyanin X-ray structure. Only the backbone of the C,-carbon atoms is drawn. Stretches in the polypeptide chain supposed to be in contact with the antibodies are in white. The monomer is viewed parallel to the threefold axis of the hexamer.
fraction of charged amino acid residues as compared to the other loops involved with the antibody.
The direction of the threefold axes of the hemocyanin hexamers in the hemocyanidIgG complexes for clone E, as compared to complexes with clone J, are different, as is de- monstrated most clearly in the hemocyanidIgG chains (Fig. 1). Clone E forms complexes with the threefold axes perpendicular to the chain direction, while for clone J the threefold axes are in a parallel arrangement to the direction of the main axis of the complexes.
The angle between the Fab part of an IgG molecule and the threefold axis of the hexamer is 60 5 5" for clone E and 35 2 7" for clone J. This observation is clearly related to the difference in orientation of the hexamer in the images of the two complexes.
We thank Prof. Dr W. G. J. Hol and Prof. Dr H. J. C. Berendsen for critically reading the manuscript and Dr B. Hazes for making the X-ray data available to us. We are grateful to Dr W. Keegstra who we thank for his assistance with the use of IMAGIC. This study was supported by the Netherlands Organization for Chemical Research (SON) with financial aid from the Netherlands Organiza- tion for Scientific Research (NWO).
REFERENCES Amit, A. G., Mariuzza, R. A., Phillips, S. E. V. & Poljak, R. J.
(1986) Thr5e-dimensional structure of an antigen-antibody com- plex at 2.8A resolution, Science 233, 747-753.
Bijlholt, M. M. C., van Heel, M. G. & van Bruggen, E. F. J. (1982) Comparison of the 4 X 6-meric hemocyanins from three different arthropods using computer alignment and correspondence analy- sis, J. Mol. Biol. 161, 139-153.
Boekema, E. J. & Bottcher, B. (1992) The structure of ATP synthase from chloroplasts. Conformational changes of CF1 studied by electron microscopy, Biochim. Biophys. Acta 1098, 131 -143.
Boisset, N., Frank, J., Taveau, J. C., Billiald, P., Motta, G., Lamy, J. N., Sizaret, P. Y. & Lamy, J. (1988) Intramolecular localization of epitopes within an oligomeric protein by IMEM and image processing, Proteins Struct. Funct. Genet. 3, 161 -183.
Boisset, N., Taveau, J. C., Lamy, J. N., Wagenknecht, T., Rader- macher, M. & Frank, J. (1990) Three-dimensional reconstruction of native Androctonus australis hemocyanin, J. Mol. Biol. 216, 743 -760.
Colman, P. M., Laver, W. G., Varghese, J. N., Baker, A. T., Tulloch, P. A., Air, G. M. & Webster, R. G. (1987) Three-dimensional structure of a complex of antibody with influenza virus neuraminidase, Nature 326, 358-363.
de Haas, F., Bijlholt, M. M. C. & van Bruggen, E. F. J. (1991) An electron microscopic study of two-hexameric hemocyanins from the crab Cancer pagurus and the tarantula Eurypelma califomi- cum: Determination of their quaternary structure using image processing and simulation models based on X-ray diffraction data, J. Struct. Biol. 107, 86-94.
de Haas, F. (1993) Electron microscopic studies on structure and function of arthropod hemocyanins, Ph.D. thesis, Rijksuniversi- teit Groningen.
Ellerton, D. H., Ellerton, N. F. & Robinson, H. A. (1983) Hemocya- nin: a current perspective, Prog. Biophys. Mol. Biol. 41, 143- 248.
Friguet, B., Chaffotte, A. F., Djavadi-Ohaniance, L. & Goldberg, M. E. (1985) Measurements of the true affinity constant in solution of antigen-antibody complexes by enzyme-linked immunosor- bent assay, J. lmmunol. Methods 77, 305-319.
Gaykema, W. P. J., Hol, W. G. J., Vereijkcn, J. M., Soeter, N. M., Bak, H. J. & Beintema, J. J. (1984) 3.2A structure of the copper- containing, oxygen-carrying protein Panulirus interruptus hemo- cyanin, Nature 309, 23 - 29.
Gaykema, W. P. J., Volbeda, A. & Hol, W. G. J. (1986) Structur? determination of Panulirus interruptus hemocyanin at 3.2A resolution, J. Mol. Biol. 187, 255-275.
Getzoff, E. D., Geysen, H. M., Rodda, S. J., Alexander, H., Tainer, J. A. & Lerner, R. A. (1987) Mechanisms of antibody binding to a protein, Science 235, 1191 -1196.
Geysen, H. M., Tainer, J. A., Rodda, S. J., Mason, T. J., Alexander, H., Getzoff, E. D. & Lerner, R. A. (1987) Chemistry of antibody binding to a protein, Science 235, 11 84- 11 90.
Goding, J. W. (1986) Monoclonal antibodies, principles and prac- tice, 2nd edn, Academic Press Harcourt Brace Janovich pub- lisher, London.
Harauz, G., Boekema, E. J. & van Heel, M. (1988) Statistical image analysis of electron micrographs of ribosomal subunits, Methods Enzymol. 164, 35-49.
Hazes, B., Magnus, K. A., Bonaventura, C., Bonaventura, J., Dauter, Z., Kalk, K. H. & Hol, W. G. J. (1993) Crystal structure of deoxygenated Limulus polyphemus subunit I1 hemocyanin at 2.18A resolution: clues for a mechanism for allosteric regula- tion, Protein Sci. 2, 597-619.
Lamy, J., Lamy, J., Bonaventura, J. & Bonaventura, C. (1980) Struc- ture, function and assembly in the hemocyanin system of the scorpion Androctonus australis, Biochemistry 19, 3033 -3039.

161
Lamy, J., Bijlholt, M. M. C., Sizaret, P. Y., Lamy, J. N. & van Bruggen, E. F. J. (1981) Quaternary structure of scorpion (An- droctonus australis) hemocyanin. Localization of subunits with immunological methods and electron microscopy, Biochemistry 20, 1849-1856.
Lamy, J., Sizaret, P. Y., Frank, J., Verschoor, A., Feldmann, R. & Bonaventura, J. (1 982) Architecture of Limulus polyphemus he- mocyanin, Biochemistry 21, 6825-6833.
Lamy, J., Billiald, P., Taveau, J. C., Boisset, N., Motta, G. & Lamy, J. N. (1990) Topological mapping of 13 epitopes on a subunit of Androctonus australis hemocyanin, J. Struct. Biol. 103, 64 -74.
Linzen, B., Soeter, M. N., Riggs, A. F., Schneider, H. J., Schartau, W., Moore, M. D., Yokata, E., Behrens, P. Q., Nakashima, H., Takagi, T., Nemoto, T., Vereijken, J. M., Bak, H. J., Beintema, J. J., Volbeda, A., Gaykema, W. P. J. & Hol, W. G. J. (1985) The structure of arthropod hemocyanins, Science 229, 519-524.
Markl, J., Kempter, B., Linzen, B., Bijlholt, M. M. C. & van Brug- gen, E. F. J. (1981) Hemocyanins in spiders, XVI. Subunit to- pography and a model of the quaternary structure of Eurypelma hemocyanin, Hoppe-Seyler’s Z. Physiol. Chem. 362, 1631 - 1641.
Markl, J. & Decker, H. (1991) Molecular structure of arthropod hemocyanins, in Advances in comparative and environmental physiology (Mangum, C., ed.) vol. 13, pp. 325-376, Springer- Verlag, Berlin.
Neuteboom, B., Jekel, P. A. & Beintema, J. J. (1992) Primary struc- ture of hemocyanin subunit c from Panulirus interruptus, Eul: J. Biochem. 206, 243-249.
Padlan, E. A., Silverton, E. W., Sheriff, S., Cohen, G. H., Smith- Gill, s. J. & Davies, D. R. (1989) Structure of an antibody- antigen complex: Crystal structure of the HyHEL-10 Fab-lyso- zyme complex, Proc. Natl Acad. Sci. USA 86, 5938-5942.
Perton, F. G., van Breemen, J. F. L., Wolters, G. K. & Beintema, J. J. (1990) Monoclonal antibodies against hemocyanin from Pa- nulirus interruptus, in Invertebrate dioxygen carriers (PrCaux, G. & Lontie, R., eds) pp. 339-342. Leuven University Press.
Stoffler-Meilicke, M. & Stoffler, G. (1987) The topography of ribo- somal proteins on the surface of the 30s subunit of Escherichia coli, Biochimie (Paris) 69, 1049- 1064.
van Eerd, J. P. & Folkerts, A. (1981) Isolation and characterization of five subunits of the hemocyanin from the spiny lobster Panuli- rus interruptus, in Invertebrate oxygen-binding proteins: struc- ture active site and function (Lamy, J. & Lamy, J. N., eds) pp. 139-149, Dekker, New York.
van Heel, M. & Keegstra, W. (1981) IMAGIC: a fast, friendly and flexible image analysis software system, Ultramicroscopy 7, 11 3 - 130.
Van Holde, K. E. & van Bruggen, E. F. J. (1971) The hemocyanins, in Biological macromolecules (Timasheff, S . N . & Fasman, G. D., eds) vol. 5, pp. 1-55, Decker, New York.
Van Holde, K. E. & Miller, K. J. (1982) Haemocyanins, Q. Rev. Biophys. 15, 1-129.
Volbeda, A. & Hol, W. G. J. (1989) Crystal structure of hpxameric hemocyanin from Panulirus interruptus refined at 3.2A resolu- tion, J. Mol. Biol. 209, 249-279.
Wang, G., Porta, C., Chen, Z., Baker, T. S. & Johnson, J. E. (1992) Identification of a Fab interaction footprint site on an icosahedral virus by cryo-electron microscopy and X-ray crystallography, Nature 355, 275-278.
Wichertjes, T., Keegstra, W., Neuteboom, B., Hazes, B., Beintema, J. J. & van Bruggen, E. F. J. (1989) Crystallization properties and structure of Panulirus interruptus hemocyanin, Eul: J. Bio- chem. 184, 287-296.


















