
Identification of the TRiC/CCT Substrate Binding Sites Uncovers the ...
Upload
naoki-tanakaCategory
view
212download
0
Article No. jmbi.1999.3041 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 292, 173±180
Identification of Substrate Binding Site of GroELMinichaperone in Solution
Naoki Tanaka1 and Alan R. Fersht2*
1Department of PolymerScience and Engineering, KyotoInstitute of Technology, SakyoKyoto, 606-8585, Japan2Cambridge UniversityChemical Laboratory andCambridge Centre for ProteinEngineering, MRC Centre HillsRoad, Cambridge, CB2 2QHUK
E-mail address of the [email protected]
Abbreviations used: AEDANS, 5[2(acetyl)aminoethyl]aminonaphthaacetamido¯uorescein; DTT, dithioth5-[2 (iodoacetyl) aminoethyl]aminosulfonate; IAF, iodoacetamido¯uorlactalbumin, disul®de reduced a-lahistidine tag.
0022-2836/99/360173±8 $30.00/0
It is dif®cult to obtain high-resolution structural information on the sub-strate-binding site of intact GroEL. But minichaperones, domains contain-ing the peptide-binding site of GroEL, do constitute tractable systems fordetailed studies. A peptide-binding site was located in crystals of a mini-chaperone and proposed to constitute a model for substrate-binding. Wehave now located the substrate binding site of the minichaperoneGroEL(193-335) in solution by labelling it at various positions with a ¯u-orescent probe and detecting which positions are perturbed on binding adenatured substrate. The ¯uorescence of a probe attached to a cysteineresidue engineered at position 228 (N terminus of helix H8), 241 (helixH8), 261 (helix H9), or 267 (helix H9) was affected signi®cantly by bind-ing of substrate. But there was little change for a label at positions 193,212, 217 or 293. The dissociation constants between substrates and mini-chaperone were evaluated from ¯uorescence anisotropy assays. Theeffects of salt and temperature were the same as those with intact GroEL.These results indicate that the region around helices H8 and H9 is thesubstrate-binding site for the apical domain fragment. Intriguingly, thesame site is involved in the binding of GroES. Thus, an important func-tion of GroES in the regulation of the activity of GroEL for substrates isto displace the bound substrate by competing for its binding site.
# 1999 Academic Press
Keywords: protein; folding; ¯uorescence anisotropy; denatured; pepsin; a-lactalbumin
*Corresponding authorIntroduction
The molecular chaperone GroEL assists the fold-ing of certain proteins by preventing aggregationand by correcting misfolding (Lorimer, 1996; Ellis& Hartl, 1996). GroEL is a hollow cylinder of 14identical 57 kDa subunits arranged in two seven-membered rings (Braig et al., 1994, 1995). TheGroEL monomer is composed of three domains: anapical domain that is responsible for binding sub-strate (Fenton & Horwich, 1993), an equatorialdomain that contains the ATP-binding site, and anintermediate hinge domain that connects the apicaland equatorial domains. Fragments of the apical
ing author:
-lene-1-sulfonate; AF,reitol; IAEDANS,
naphthalene-1-escein; R-a-ctalbumin; sht, short
domain of GroEL, called minichaperones, cause theunfolding of barnase and help refold several pro-teins (Zahn et al., 1996; Altamirano et al., 1997;Chatellier et al., 1998). The importance of therefolding activity towards substrates that have ahigh spontaneous rate of refolding has been dis-puted (Wang et al., 1998; Weber et al., 1998), butexperiments on substrates that are very dif®cult tofold show unambiguously that minichaperones canhave very effective chaperoning activity(Altamirano et al, 1999). So far, no structure of acomplex between GroEL and a substrate has beensolved. But, minichaperones have been crystallisedand a high-resolution structure of shtGroEL(191-376) has led to a postulated model for the GroEL-substrate complex (Buckle et al., 1997). The nota-tion sht stands for a short histidine tag attached tothe N terminus of the domain via a 17 residue lin-ker sequence. Seven residues of the linker from onemolecule were observed to bind in the region ofthe next molecule that had been postulated to bethe substrate-binding site from mutagenesis exper-iments that altered activity (Fenton et al., 1994).However, we note that in the crystal structure of
# 1999 Academic Press

Figure 2. Emission spectra of AEDANS-GroEL(193-335), measured using an excitation wavelength of340 nm. AEDANS-GroEL(193-335) was at a concen-tration of 1 mM in 25 mM Tri-HCl (pH 7.8), 2 mM DTT,1 mM EDTA containing 0.2 M KCl.
Figure 1. The positions of the mutated residues ofminichaperone that were labelled with IAEDANS.Three-dimensional structure of GroEL(191-345) solved at2.5 AÊ (Zahn et al., 1996) is used in this Figure. Second-ary structure representation is drawn with MOLSCRIPT(Kraulis, 1991) and Raster3D (Merritt & Bacon, 1997).
174 Substrate-Binding Site of GroEL Fragment
the GroEL-GroES complex (Xu et al., 1997), a loopof GroES binds in the postulated substrate-bindingsite of GroEL. If this is so, then it is of mechanisticimportance and it is necessary to con®rm indepen-dently the location of the substrate binding site.
In the present study, we have investigated thesite of the interaction of the apical domain frag-ment GroEL(193-335) (Chatellier et al., 1998) witha-lactalbumin and denatured pepsin using a ¯uor-escence method. We have chemically attached a¯uorescence probe to single cysteine mutants ofGroEL(193-335) site-selectively on its surface. Thechange of ¯uorescence upon adding substrateindicates if the substrate covers the site on binding.
The molten globule state of a-lactalbumin hasbeen well characterised (Kuwajima et al., 1992).a-Lactalbumin without bound Ca2� (apo-a-lact-albumin) assumes a stable molten globule state atlow ionic strength, and this state resembles anintermediate in the refolding kinetics of disul®de-intact a-lactalbumin from the fully unfolded state.The reduced form, R-a-lactalbumin, is moreunfolded than the molten globule state, although ithas some secondary structures (Okazaki et al.,1994). The mechanism of recognition of a-lactalbu-min with GroEL has been investigated in detail(Hayer-Hartl et al., 1994; Robinson et al., 1994;Okazaki et al., 1994; Katsumata et al. 1996a,b; Aokiet al., 1997). Pepsin is a gastric aspartic proteinasewith a pH optimum for catalytic activity at pH < 2,and irreversibly loses its conformation when thepH is raised to above pH 7 (Aoki et al., 1997; Linet al., 1993). We evaluated the dissociation con-stants between GroEL(193-335) and substrates by¯uorescence anisotropy assays.
Results
The effect of substrate binding onfluorescence of AEDANS-GroEL(193-335)
Eight different single cysteine mutants of mini-chaperone were prepared by site-directed muta-genesis of GroEL(193-335) (Figure 1). A241C islocated in helix H8, T261C and M267C are in helixH9, and S228C is in the N terminus of helix H8.M193C, A212C, S217C and A293C are not locatedin the vicinity of these helices. CD spectra of nearand far-UV regions for each mutant were identicalwith those of wild-type, and the native secondaryand tertiary structures are maintained for themutants. The thiol-reactive ¯uorescence probe IAE-DANS was attached to a cysteine residue to pre-pare AEDANS-GroEL(193-335). IAEDANS is anenvironment-sensitive ¯uorophore whose emissionmaximum varies with polarity of the environment.Figure 2 shows the ¯uorescence spectra ofAEDANS attached to the single cysteine residue ofthe different GroEL(193-335) mutants. The wave-length of emission maximum of AEDANS-A241Cis blue-shifted relative to the spectra of otherAEDANS-GroEL(193-335) mutants. This may bebecause AEDANS attached at A241C is buried
between hydrophobic residues such as V273 andL237, which are located adjacent to A241.
The effect of substrate binding on the ¯uor-escence spectra of AEDANS-GroEL(193-335) wasexamined. Figure 3(a) shows the ¯uorescence spec-

Figure 3. The effect of the bind-ing of a-lactalbumin on the emis-sion spectra of AEDANS-GroEL(193-335). The concentrationof IAEDANS-GroEL(193-335) was1 mM in 25 mM Tri-HCl (pH 7.8),2 mM DTT, 1 mM EDTA, 0.2 MKCl. Each Figure shows the spec-trum without substrate and onaddition of 243 mM a-lactalbumin.(a) AEDANS-S228C; (b) AEDANS-A241C; (c) AEDANS-S217C.
Substrate-Binding Site of GroEL Fragment 175
tra of AEDANS-S228C in the presence of excess ofapo-a-lactalbumin and R-a-lactalbumin. Themaximum wavelength of ¯uorescence ofAEDANS-S228C was little affected in the presenceof apo-a-lactalbumin, but it was blue-shifted in thepresence of R-a-lactalbumin. It has been shownthat R-a-lactalbumin binds tightly to GroEL,whereas the af®nity of apo-a-lactalbumin to GroELis weak (Okazaki et al., 1994; Katsumata et al.,1996a,b). Thus, the results in Figure 3(a) indicatethat R-a-lactalbumin binds to GroEL(193-335) andthe binding causes the environment of AEDANS tobe more hydrophobic. Figure 3(b) shows the ¯uor-escence spectra of AEDANS-A241C in the presenceof apo-a-lactalbumin and R-a-lactalbumin. Themaximum wavelength of ¯uorescence ofAEDANS-A241C was red-shifted in the presenceof R-a-lactalbumin. It has been shown that thebinding of amphipathic peptides to GroEL isaccompanied by an increase in hydrophobic sur-face exposure (Brazil et al., 1997). Thus, it seemslikely that the substrate binding induces localstructural rearrangement in the binding site ofGroEL(193-335), and so a ¯uorophore buried in thehydrophobic surface is exposed to a polar side-chain of substrate or solvent. L237, which is closeto A241, is involved in binding of polypeptide sub-strates to GroEL (Fenton et al., 1994). The change ofthe location of L237 induced by substrate bindingmay be responsible for the red-shift of ¯uor-escence. On the other hand, little change in ¯uor-escence was observed for AEDANS-S217C onadding R-a-lactalbumin (Figure 3(c)). This indicatesthat R-a-lactalbumin does not bind to the regionaround position 217C. The shift of average emis-sion wavelength (hli) of ¯uorescence of AEDANS-GroEL(193-335) induced by substrate binding weresummarised in Table 1. The average emissionwavelength (hli) was calculated using theequation:
hli �PlN
i�li�Fili�PlN
i�li Fi
�1�
where F is the ¯uorescence intensity, and l is thewavelength.
Results summarised in Table 1 indicate that R-a-lactalbumin binds to the local region aroundhelices H8 and H9 of the minichaperone. Similarresults were obtained for the effects of binding ofdenatured pepsin on the ¯uorescence of AEDANS-GroEL(193-335) shown in Figure 4(a)-(c) andTable 1.
Fluorescence anisotropy assays of binding ofsubstrate proteins to apical domain fragment
We estimated the dissociation constant of thebinding of GroEL(193-335) to R-a-lactalbumin anddenatured pepsin using ¯uorescence anisotropyassays. IAF is used as an extrinsic ¯uorescenceprobe because of its high degree of sensitivity. Ifthe ¯uorophore attached to minichaperone isdirectly in contact with substrate, the ¯uorescencelifetime will change, so that the anisotropy value isnot related to the rotational correlation time of¯uorophore. The ¯uorescence anisotropy is relatedto the correlation time (tc) of the ¯uorophorethrough the Perrin equation (Perrin, 1926):
A0
Aÿ 1 � t
tc�2�
where A0 is the limiting anisotropy of the ¯uoro-phore and t is the ¯uorescence lifetime. Anincrease in the measured anisotropy as a functionof macromolecules is a measure of macromoleculebinding.
S217C was labelled with IAF and used for aniso-tropy measurement because we found that the ¯u-orescence of AEDANS-S217C is not affected bysubstrate binding. An equilibrium binding iso-therm was constructed by titrating a ®xed concen-tration of AF-S217C with increasing concentrationsof a-lactalbumin (Figure 5(a)). The anisotropy ofAF-S217C was little affected on addition of apo-a-lactalbumin, but it was increased with increasingconcentration of R-a-lactalbumin, re¯ecting thebinding of R-a-lactalbumin to minichaperone. The
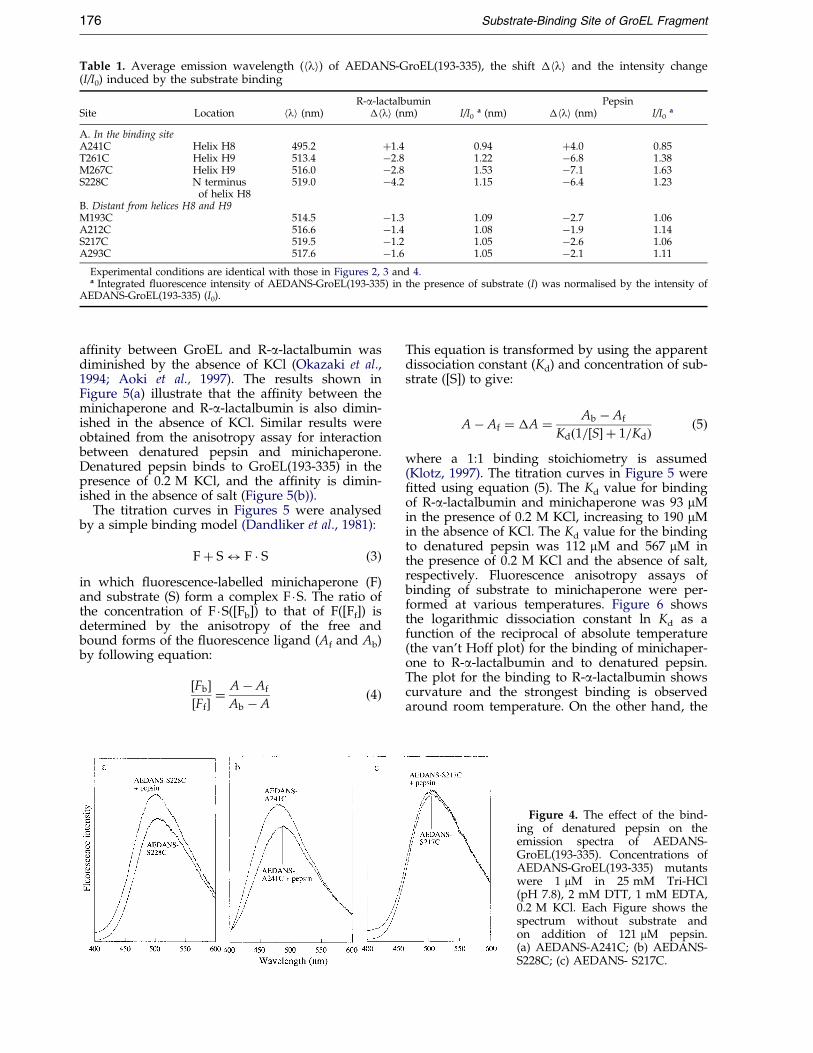
Table 1. Average emission wavelength (hli) of AEDANS-GroEL(193-335), the shift �hli and the intensity change(I/I0) induced by the substrate binding
R-a-lactalbumin PepsinSite Location hli (nm) �hli (nm) I/I0
a (nm) �hli (nm) I/I0a
A. In the binding siteA241C Helix H8 495.2 �1.4 0.94 �4.0 0.85T261C Helix H9 513.4 ÿ2.8 1.22 ÿ6.8 1.38M267C Helix H9 516.0 ÿ2.8 1.53 ÿ7.1 1.63S228C N terminus 519.0 ÿ4.2 1.15 ÿ6.4 1.23
of helix H8B. Distant from helices H8 and H9M193C 514.5 ÿ1.3 1.09 ÿ2.7 1.06A212C 516.6 ÿ1.4 1.08 ÿ1.9 1.14S217C 519.5 ÿ1.2 1.05 ÿ2.6 1.06A293C 517.6 ÿ1.6 1.05 ÿ2.1 1.11
Experimental conditions are identical with those in Figures 2, 3 and 4.a Integrated ¯uorescence intensity of AEDANS-GroEL(193-335) in the presence of substrate (I) was normalised by the intensity of
AEDANS-GroEL(193-335) (I0).
176 Substrate-Binding Site of GroEL Fragment
af®nity between GroEL and R-a-lactalbumin wasdiminished by the absence of KCl (Okazaki et al.,1994; Aoki et al., 1997). The results shown inFigure 5(a) illustrate that the af®nity between theminichaperone and R-a-lactalbumin is also dimin-ished in the absence of KCl. Similar results wereobtained from the anisotropy assay for interactionbetween denatured pepsin and minichaperone.Denatured pepsin binds to GroEL(193-335) in thepresence of 0.2 M KCl, and the af®nity is dimin-ished in the absence of salt (Figure 5(b)).
The titration curves in Figures 5 were analysedby a simple binding model (Dandliker et al., 1981):
F� S$ F � S �3�in which ¯uorescence-labelled minichaperone (F)and substrate (S) form a complex F �S. The ratio ofthe concentration of F �S([Fb]) to that of F([Ff]) isdetermined by the anisotropy of the free andbound forms of the ¯uorescence ligand (Af and Ab)by following equation:
�Fb��Ff� �
Aÿ Af
Ab ÿ A�4�
This equation is transformed by using the apparentdissociation constant (Kd) and concentration of sub-strate ([S]) to give:
Aÿ Af � �A � Ab ÿ Af
Kd�1=�S� � 1=Kd� �5�
where a 1:1 binding stoichiometry is assumed(Klotz, 1997). The titration curves in Figure 5 were®tted using equation (5). The Kd value for bindingof R-a-lactalbumin and minichaperone was 93 mMin the presence of 0.2 M KCl, increasing to 190 mMin the absence of KCl. The Kd value for the bindingto denatured pepsin was 112 mM and 567 mM inthe presence of 0.2 M KCl and the absence of salt,respectively. Fluorescence anisotropy assays ofbinding of substrate to minichaperone were per-formed at various temperatures. Figure 6 showsthe logarithmic dissociation constant ln Kd as afunction of the reciprocal of absolute temperature(the van't Hoff plot) for the binding of minichaper-one to R-a-lactalbumin and to denatured pepsin.The plot for the binding to R-a-lactalbumin showscurvature and the strongest binding is observedaround room temperature. On the other hand, the
Figure 4. The effect of the bind-ing of denatured pepsin on theemission spectra of AEDANS-GroEL(193-335). Concentrations ofAEDANS-GroEL(193-335) mutantswere 1 mM in 25 mM Tri-HCl(pH 7.8), 2 mM DTT, 1 mM EDTA,0.2 M KCl. Each Figure shows thespectrum without substrate andon addition of 121 mM pepsin.(a) AEDANS-A241C; (b) AEDANS-S228C; (c) AEDANS- S217C.

Figure 5. Titration of AF-S217C with substrate pro-teins. The measured increase (�Anisotropy) from theinitial anisotropy value is plotted on the y-axis. Concen-tration of AF-S217C was 0.25 mM. Lines through thedata points are the results of the ®t with a simple bind-ing model described in equation (5). The ®tting wasdone using the non-linear least-squares method withKaleidaGraph2. (a) Titration with R-a-lactalbumin: (*),R-a-lactalbumin in 25 mM Tris (pH 7.8), 1 mM EDTA,2 mM DTT, 0.2 M KCl; (*), in 25 mM Tris (pH 7.8),1 mM EDTA, 2 mM DTT; (&), apo-a-lactalbumin in25 mM Tris (pH 7.8), 1 mM EDTA. (b) The titrationwith denatured pepsin: (*), pepsin in 25 mM Tris(pH 7.8), 1 mM EDTA, 2 mM DTT, 0.2 M KCl; (*),pepsin in 25 mM Tris (pH 7.8), 1 mM EDTA, 2 mMDTT.
Figure 6. The van't Hoff plot for the dissociation con-stant between GroEL(193-335) and substrate proteins((*), R-a-lactalbumin in 25 mM Tris (pH 7.8), 1 mMEDTA, 2 mM DTT, 0.2 M KCl; (*), pepsin in 25 mMTris (pH 7.8), 1 mM EDTA, 2 mM DTT, 0.2MKCl).
Substrate-Binding Site of GroEL Fragment 177
van't Hoff plot for the binding to denatured pepsinis almost linear.
Discussion
A polypeptide binding site for GroEL was pro-posed to be located in its apical domain involvinghelices H8 and H9 and the 296-320 loop betweenhelix H10 and strand S12, based on the sites ofmutations that inactivated GroEL (Braig et al.,1994). A crystallographic study of GroEL(191-376)with a 17-residue N-terminal tag showed that theN-terminal tag of one molecule binds to a neigh-bouring molecule in the unit cell (Buckle et al.,1997). Residues ÿ7 to ÿ1 (sequence GLVPRGS) ofthe N-terminal tag were located within a shallow
cleft between helices H8 and H9, and in an adja-cent surface that is formed by the packing betweenhelix H9 and a neighbouring loop. This form ofbinding was proposed to mimic the binding ofsubstrate molecules, and the region around helicesH8 and H9 is the substrate binding site forGroEL(191-376). This region is also responsible forthe binding of GroES to GroEL (Xu et al. , 1997). Inthe accompanying paper, it is shown that minicha-perone-bound peptides exhibit a non-sequentiallinear binding motif with similar dimensions andchemical characteristics to the GroEL-boundmobile loop of GroES (Chatellier et al., 1999).
The study of the effects of substrate on the ¯uor-escence of AEDANS-GroEL(193-335) indicates thatthe local region around helices H8 and H9 is thesubstrate binding site of minichaperone in solution.The extrinsic ¯uorescence probe, 5-[2 (iodoacetyl)aminoethyl]aminonaphthalene-1-sulfonate wasattached to mutants of GroEL(193-335) via a uniquecysteine residue on the surface of the proteinengineered at eight different positions: A241C(helix H8); T261C (helix H9), M267C (helix H9);S228C (N terminus of helix H8); M193C; A212C;S217C; and A293C. The ¯uorescence of AEDANSlabelled at positions 241, 261, 267 and 228 wasaffected by the substrate binding, but little changewas observed for the ¯uorescence of AEDANSlabelled at 193, 212, 217 and 293. These resultsindicate that the site around helices H8 and H9,which is involved in the binding of GroES, is alsothe substrate-binding site of the apical domainfragment. Thus, GroES competes directly with sub-strates for the binding site on GroEL.
Comparison of binding to GroELand minichaperone
The af®nity of substrates for the apical domainshould be less than that for intact GroEL because

178 Substrate-Binding Site of GroEL Fragment
of the presence of multiple sites in its heptamericring. However, relative effects of changes in sub-strates and conditions are comparable. Okazakiet al. (1994) investigated the interaction of a-lactal-bumin with GroEL by size-exclusion chromatog-raphy. A strong af®nity of R-a-lactalbumin toGroEL was detected in the presence of 0.2 M KCl,and the dissociation constant between GroEL andR-a-lactalbumin was estimated to be of the orderof 10ÿ7 M. Binding of apo-a-lactalbumin to GroELwas too weak to be detected by this method.Katsumata et al. (1996a) have evaluated the bind-ing constant between GroEL and a-lactalbumin inthe molten globule state by measuring the refold-ing kinetics of apo-a-lactalbumin in the absenceand presence of GroEL. The dissociation constantwas estimated to be of the order of 10 mM which islarger than that between GroEL and R-a-lactalbu-min by two orders of magnitude. Consistent withthis, the result shown in Figure 5(a) indicates thatthe Kd between minichaperone and apo-a-lactalbu-min is larger than that between minichaperone andR-a-lactalbumin by at least two orders of magni-tude.
It has been shown that electrostatic interactionsare important for the binding of GroEL to severalproteins. Point mutation studies revealed that posi-tive charges on chymotrypsin inhibitor 2 increasebinding to GroEL, while negative charges weakenbinding (Itzhaki et al., 1995). The association ofGroEL with barnase (Perrett et al., 1997) and sta-phylococcal nuclease (Tsurupa et al., 1998) wasdiminished at high ionic strength. On the otherhand, GroEL binds to R-a-lactalbumin and pepsintightly in the presence of a high concentration ofsalt, indicating that there is electrostatic repulsionbetween GroEL and these proteins. GroEL has anegative charge at neutral pH and so a positivecharge on the substrate promotes the interactionwith GroEL. Conversely, a negative chargediminishes the interaction. Residues of GroELwhich are responsible for the electrostatic inter-action to substrate are not identi®ed, and it may bepossible that electrostatic interaction betweenGroEL and substrate protein is from non-speci®cinteractions with the negatively charged surface ofGroEL as a whole. However, the present resultsindicate that af®nity of minichaperone with R-a-lactalbumin and pepsin is weakened at low ionicstrength, suggesting that the electrostatic inter-action between GroEL and substrate originatesfrom the local charge of the apical domain ofGroEL.
Aoki et al. (1997) have analysed the binding ofGroEL to R-a-lactalbumin and denatured pepsinby isothermal titration calorimetry at various tem-peratures. The �H of pepsin-GroEL binding showsan almost constant value from 11.7 to 30.1 �C. Onthe other hand, the �H of R-a-lactalbumin-GroELis strongly dependent on the temperature. Positive�H values at low temperatures decrease withincreasing temperature, and the value is zero atroom temperature. As shown in Figure 6, the van't
Hoff plot for the binding of pepsin with minicha-perone is linear, indicating that �H is not stronglytemperature dependent. On the other hand, thevan't Hoff plot for the binding of R-a-lactalbuminwith minichaperone is a concave curve, and �H iszero at room temperature. These results show thatthe minichaperone binds the denatured proteinswith the same thermodynamic properties as doesthe intact GroEL. In the accompanying paper, it isshown that minichaperone recognizes a-helicalconformation (Kobayashi et al., 1999) as shown forthe recognition by GroEL (Landry & Gierasch,1991; Brazil et al., 1997).
Mechanistic implications
The af®nity of GroEL for denatured states isvery high (Gray & Fersht,1993; Katsumata et al.,1996a), and is ampli®ed by the existence of mul-tiple binding sites. Such tight binding slows downthe folding of denatured states. One of the func-tions of the binding of ATP and GroES to GroEL isto weaken the binding and allow folding to takeplace more readily. It can be seen in electronmicroscopy studies that the binding sites of GroELare displaced on the binding of GroES (Rosemanet al., 1996). It has also been noted from the crystalstructure that the hydrophobic surfaces of GroELthat are exposed to substrate become more polar inthe GroEL-GroES complex (Xu et al., 1997). Ourresults here and the accompanying analysis(Chatellier et al., 1999) show that GroES anddenatured substrates compete directly for the samesites and so the binding of GroES directly displacesbound substrates, either into the cavity or into sol-ution.
Materials and Methods
Materials
Bovine apo-a-lactalbumin (type III, Ca(II)-depleted)and porcine pepsin were purchased from Sigma (St.Louis, MO) and puri®ed by size-exclusion FPLC. R-a-lac-talbumin was prepared by dissolving bovine apo-a-lac-talbumin in 25 mM Tris-HCl (pH 7.8), 2 mM DTT, 1 mMEDTA. IAEDANS and IAF were obtained from Molecu-lar Probe, Inc. (Eugene, OR). Other reagents were of thehighest grade commercially available and were usedwithout further puri®cation.
The GroEL(193-335) with the short N-terminal histi-dine tag (sht, MRGSHHHHHHGLVPRGS), which con-tains an engineered thrombin cleavage site, was clonedinto a pRSETA vector (Invitrogen (Carlsbad, CA))(Chatellier et al., 1998). A Quickchange2 site-directedmutagenesis kit (Strategene (La Jolla, CA)) was used inorder to introduce the desired mutation into the targetgene (using a pair of completely complementary pri-mers) and to amplify the full-length plasmid. Sht-GroEL(193-335) and its mutants were expressed inEscherichia coli C41(DE3), a mutant of BL21(DE3)(Miroux & Walker, 1996), puri®ed by af®nity chromatog-raphy on a Ni-NTA-agarose column and cleaved fromthe histidine tail by thrombin digestion, essentially asdescribed by Chatellier et al. (1998). The cleaved protein

Substrate-Binding Site of GroEL Fragment 179
was puri®ed further by size-exclusion FPLC (Superdex75, Pharmacia) in 50 mM Tris (pH 8.2), 150 mM NaCl,2 mM DTT.
The modi®cation of single cysteine mutant GroEL(193-335) with a thiol-reactive ¯uorescence probe was carriedout as follows. IAEDANS (®nal concentration 5 mM) orIAF (®nal concentration 1 mM) was added to solution ofGroEL(193-335) mutant in 50 mM Tris (pH 8.2), 150 mMNaCl, containing 2 mM DTT with stirring in the dark at25 �C. The reactions were allowed to proceed for 50 min-utes (for IAEDANS) or ten minutes (for IAF), and thereaction mixture was applied to a Sephadex G-25 col-umn. The protein fraction thus eluted was modi®edGroEL(193-335) mutant whose single cysteine residuewas blocked with a ¯uorescence group. The ®nal proteinconcentration was determined by BCA protein assay andthe concentration of AEDANS and AF were determinedby absorbance at 337 nm ("337 � 6100 Mÿ1 cmÿ1) and at490 nm ("490 � 42,000 Mÿ1 cmÿ1). The labelling ratio forthe ¯uorescent derivatives of GroEL(193-335) mutantswere con®rmed to be 1.0.
Methods
Fluorescence measurements were made with anAMINCO-Bowman spectro¯uorimeter. Fluorescence ani-sotropy measurements for AF-GroEL(193-335) were per-formed with an excitation wavelength of 490 nm and anemission wavelength of 520 nm. Anisotropy wasmeasured ten times at each titration point with an inte-gration time of eight seconds for each intensity measure-ment, and the resulting anisotropy values wereaveraged. R-a-lactalbumin tends to aggregate when it isleft for a long while. Anisotropy values of AF-GroEL(193-335) were measured repeatedly at ®ve minuteintervals in the presence of denatured protein. Stable ani-sotropy values were obtained within about 30 minutes.Thus, the effect of aggregation on the anisotropy value isnegligible under our experimental conditions. CDmeasurements were made with a Jasco-720 spectropolari-meter using a 1 mm path-length cell for far-UV spectraand a 10 mm path-length cell for near-UV spectra. Spec-troscopic measurements used a buffer containing 25 mMTris-HCl (pH 7.8), 2 mM DTT, 1 mM EDTA, and 0.2 MKCl. The KCl was omitted where indicated.
Acknowledgements
Dr J. Chatellier is acknowledged for providing plas-mid pRSETA-sht-GroEL(193-335) and advice in proteinexpression and puri®cation. We thank Dr A.M. Bucklefor preparing Figure 1. We also thank Drs T. Ikura, N.Kobayashi, C. Johnson and L. S. Itzhaki, Q. Wang andU. Binne for discussion and technical assistance.
References
Altamirano, M. M., Golbik, R., Zahn, R., Buckle, A. M.& Fersht, A. R. (1997). Refolding chromatographywith immobilized mini-chaperones. Proc. Natl Acad.Sci. USA, 94, 3576-3578.
Altamirano, M. M., Garcia, C., Possani, L. D. & Fersht,A. R. (1999). Oxidative refolding chromatography:folding of the scorpion toxin Cn5. Nature Biotechnol.17, 187-191.
Aoki, K., Taguchi, H., Shindo, Y., Yoshida, M.,Ogasawara, K., Yutani, K. & Tanaka, N. (1997).Calorimetric observation of a GroEL-protein bind-ing reaction with little contribution of hydrophobicinteraction. J. Biol. Chem. 272, 32158-32162.
Braig, K., Otwinowski, Z., Hedge, R., Biosvert, D. C.,Joachimiak, A., Horwich, A. L. & Sigler, P. B.(1994). The crystal structure of the bacterial chaper-onin GroEL at 2.8 AÊ . Nature, 371, 578-586.
Braig, K., Adams, P. D. & Brunger, A. T. (1995). Confor-mational variability in the re®ned structure of thechaperonin GroEL at 2.8 AÊ resolution. Nature Struct.Biol. 2, 1083-1094.
Brazil, B. T., Cleland, J. L., McDowell, R. S., Skelton,N. J., Paris, K. & Horowitz, P. M. (1997). Modelpeptide studies demonstrate that amphipathic sec-ondary structure can be recognized by the chapero-nin GroEL (Cpn60). J. Biol. Chem. 272, 5105-5111.
Buckle, A. M., Zahn, R. & Fersht, A. R. (1997). A struc-tural model for GroEL-polypeptide recognition.Proc. Natl Acad. Sci. USA, 94, 3571-3575.
Chatellier, J., Hill, F., Lund, P. A. & Fersht, A. R. (1998).In vivo activities of GroEL minichaperones. Proc.Natl Acad. Sci. USA, 95, 9861-9866.
Chatellier, J., Buckle, A. M. & Fersht, A. R. (1999).GroEL recognises sequential and non-sequentiallinear structural motifs compatible with extendedb-strands and a-helices. J. Mol. Biol. 292, 163-172.
Dandliker, W. B., Hus, M., Levin, J. & Rao, R. (1981).Equilibrium and kinetic inhibition assays basedupon ¯uorescence polarization. Methods Enzymol.74, 3-29.
Ellis, R. J. & Hartl, F. U. (1996). Protein folding in thecell: competing models of chaperonin function.FASEB J. 10, 20-26.
Fenton, W. A. & Horwich, A. L. (1993). GroEL-mediatedprotein folding. Protein Sci. 6, 1379-1385.
Fenton, W. A., Kashi, Y., Furtak, K. & Horwich, A. L.(1994). Residues in chaperonin GroEL required forpolypeptide binding and release. Nature, 371, 614-619.
Gray, T. E. & Fersht, A. R. (1993). Refolding of barnasein the presence of GroE. J. Mol. Biol. 232, 1197-1207.
Hayer-Hartl, M. K., Ewbank, J. J., Creighton, T. E. &Hartl, F. U. (1994). Conformational speci®city of thechaperonin GroEL for the compact folding inter-mediates of a-lactalbumin. EMBO J. 13, 3192-3202.
Itzhaki, L. S., Otzen, D. E. & Fersht, A. R. (1995). Natureand consequences of GroEL-protein interactions.Biochemistry, 34, 14581-14587.
Katsumata, K., Okazaki, A. & Kuwajima, K. (1996a).Effect of GroEL on the re-folding kinetics of a-lac-talbumin. J. Mol. Biol. 258, 827-838.
Katsumata, K., Okazaki, A., Tsurupa, G. P. &Kuwajima, K. (1996b). Dominant forces in the rec-ognition of a transient folding intermediate of a-lac-talbumin by GroEL. J. Mol. Biol. 264, 643-649.
Klotz, I. M. (1997). Ligand-Receptor Energetics, John Wiley& Sons, Inc., New York.
Kobayashi, N., Freund, S. M. V., Chatellier, J. & Fersht,A. R. (1999). NMR analysis of the binding of a rho-danese peptide to a minichaperone in solution. J.Mol. Biol. 292, 181-190.
Kraulis, P. J. (1991). MOLSCRIPT: a program to produceboth detailed and schematic plots of protein struc-tures. J. Appl. Crystallog. 24, 946-950.
Kuwajima, K. (1992). Protein folding in vitro. Curr. Opin.Biotechnol. 3, 462-467.

180 Substrate-Binding Site of GroEL Fragment
Landry, S. J. & Gierasch, L. M. (1991). The chaperoninGroEL binds a polypeptide in an alpha-helical con-formation. Biochemistry 30, 7359-7362.
Lin, X., Loy, J. A., Sussman, F. & Tang, J. (1993). Confor-mational instability of the N- and C-terminal lobesof porcine pepsin in neutral and alkaline solutions.Protein Sci. 2, 1383-1390.
Lorimer, G. H. (1996). A quantitative assessment of therole of chaperonin proteins in protein foldingin vivo. FASEB J. 10, 5-9.
Merritt, E. A. & Bacon, D. J. (1997). Raster3D photorea-listics molecular graphics. Methods Enzymol. 277,505-524.
Miroux, B. & Walker, J. E. (1996). Over-production ofprotein in Escherichia coli: mutant hosts that allowsynthesis of some membrane proteins and globularproteins at high level. J. Mol. Biol. 260, 289-298.
Okazaki, A., Ikura, T., Nikaido, K. & Kuwajima, K.(1994). The chaperonin GroEL does not recognizeapo-a-lactalbumin in the molten globule state.Nature Struct. Biol. 1, 439-446.
Perrett, S., Zahn, R., Stenberg, G. & Fersht, A. R. (1997).Importance of electrostatic interactions in the rapidbinding of polypeptides to GroEL. J. Mol. Biol. 269,892-901.
Perrin, F. (1926). Polarisation de la lumieÁre de ¯uor-escence vie moyenne des molecules dans l'eÃtatexciteÂ. J. Phys. Radium. 7, 390-401.
Robinson, C. V., Gros, M., Eyles, S. J., Ewbank, J. J.,Mayhew, M., Hartl, F. U., Dobson, C. M. &
Radford, S. E. (1994). Conformation for GroEL-bound a-lactalbumin probed by mass spectrometry.Nature, 372, 646-651.
Roseman, A. M., Chen, S., White, H., Braig, K. & Saibil,H. R. (1996). The chaperonin ATPase cycle:mechan-ism of allosteric switching and movements of sub-unit. Cell, 87, 241-51.
Tsurupa, G. P., Ikura, T., Makio, T. & Kuwajima, K.(1998). Refolding kinetics of staphylococcal nucleaseand its mutants in the presence of the chaperoninGroEL. J. Mol. Biol. 277, 733-745.
Wang, J. D., Michelitsch, M. D. & Weissman, J. S. (1998).GroEL-GroES-mediated protein folding requires anintact central cavity. Proc. Natl Acad. Sci. USA, 95,12163-12168.
Weber, F., Keppel, F., Georgopoulos, C., Hayer-Hartl,M. K. & Hartl, F. U. (1998). The oligomeric structureof GroEL/GroES is required for biologically signi®-cant chaperonin function in protein folding. NatureStruct. Biol. 11, 977-985.
Xu, Z., Horwich, A. L. & Sigler, P. B. (1997). The crystalstructure of the asymmetric GroEL-GroES-(ADP)7chaperonin complex. Nature, 388, 741-750.
Zahn, R., Buckle, A. M., Perrett, S., Johnson, C. M.,Corrales, F. J., Golbik, R. & Fersht, A. R. (1996).Chaperone activity and structure of monomericpolypeptide binding domains of GroEL. Proc. NatlAcad. Sci. USA, 93, 15024-15029.
Edited by J. Karn
(Received 20 May 1999; received in revised form 16 July 1999; accepted 18 July 1999)


















