
Identification of Functional Domains in Efb, a Fibrinogen Binding Protein ofStaphylococcus aureus
6
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 248, 690–695 (1998) ARTICLE NO. RC989028 Identification of Functional Domains in Efb, a Fibrinogen Binding Protein of Staphylococcus aureus David Wade,* ,1 Marco Palma,* Ingegerd Lo ¨fving-Arvholm,* Matti Sa ¨ llberg,² Jerzy Silberring,‡ and Jan-Ingmar Flock* Department of Immunology, Microbiology, Pathology, and Infectious Diseases, *F82 and ²F62, Huddinge Hospital, Karolinska Institute, S-141 86 Huddinge, Sweden; and ‡Faculty of Chemistry, Jagiellonian University, Ingardena 3, PL-30-060 Krakow, Poland Received June 11, 1998 AA segments in its amino (N-) terminal region (Fig. 1). Staphylococcus aureus produces and secretes a pro- Similar repeating regions have been found in another tein, Efb, that binds to fibrinogen, seems to be required S. aureus protein, coagulase, where 5 to 8 copies of a for virulence, and may benefit the microorganism by 27-residue repeating sequence are found in the carboxyl delaying wound healing. Interactions of Efb with fi- (C-) terminal region of the protein, and have been shown brinogen are influenced by divalent metal cations, in- to bind to Fg (4-5). A model for the three dimensional cluding Ca 2/ . Increasing concentrations of Ca 2/ in- (3D) structure of Efb predicts that the protein is globular creased the binding of fibrinogen to immobilized Efb, and contains a substantial amount of a-helix in its C- whereas binding of Efb to immobilized fibrinogen was terminal region, and the helix content has been verified decreased with increasing Ca 2/ concentration. Studies by circular dichroism (CD) analysis (6). with synthetic peptides showed that peptides from the Fg and Efb have opposite polarities [net charges at carboxyl terminal half of Efb bound to soluble fibrino- pH 7.3 and pI values are 020 (7) and 5.5 (8) for Fg, gen and enhanced the binding of fibrinogen to Efb. A and /16 and 10.6 for Efb (1)] which makes it highly peptide corresponding to a repeated sequence in the probable that electrostatic interactions could occur be- amino terminal half of the protein also bound fibrino- tween them. There is also a 21-fold difference in their gen and inhibited binding of fibrinogen to Efb. These results may provide clues to the biological function of sizes (340 kDa for Fg versus 15.9 kDa for Efb), but the Efb and aid in the rational design of agents to block stoichiometry of binding between the two proteins has the Efb fibrinogen interaction. q 1998 Academic Press been shown to be 1:1 (9), indicating a specific interac- tion. The interaction of Efb with Fg is influenced by the divalent metal cations, calcium (Ca 2/ ) and zinc (Zn 2/ ) (9), both of which are present in the plasma (10), Staphylococcus aureus produces several proteins and during an infection, Efb and Fg would be exposed that interact with proteins of host cells, including a to these metals. Fg is known to have both low and high 15.9 kDa, basic, protein, previously named Fib but now affinity binding sites for Ca 2/ (11-12), and potential designated Efb (extracellular fibrinogen binding pro- binding sites for both metals have also been located tein), that is secreted from the bacterial cell and binds within the Efb AA sequence (6). to fibrinogen (Fg) (1). Efb is produced constituitively, Early investigations showed that the whole Efb pro- and by all isolates of S. aureus tested (2). The protein tein (136 AA’s), and its N-terminal half (69 AA’s, con- has been shown to be important for virulence of S. taining the two 22 AA repeats), bound to both solid- aureus, and it has been hypothesized that it might ben- phase Fg (Fg-Sepharose) and soluble Fg, whereas the efit the bacterium by binding to Fg and preventing its C-terminal half (70 AA’s) bound only to solid-phase Fg conversion to fibrin, thereby delaying the healing pro- (1). More recent studies have confirmed that the bind- cess (3). However, it does not participate in bacterial ing of Efb to Fg is divalent, with one binding site in adherence to Fg. the N-terminus and the other in the C-terminus (9). Efb consists of 136 amino acids (AA’s), with no cys- However, the precise sequence of AA’s in Efb that binds teine residues, and it contains two nearly identical, 22 to Fg has not yet been determined, and a knowledge of the binding site residues could be useful in the design of inhibitors to block the Efb-Fg interaction and possibly 1 To whom correspondence should be addressed. Fax: /46-8-711- 3918; E-mail: [email protected]. reduce virulence. 0006-291X/98 $25.00 Copyright q 1998 by Academic Press All rights of reproduction in any form reserved. 690
-
Upload
david-wade -
Category
Documents
-
view
213 -
download
0
Transcript of Identification of Functional Domains in Efb, a Fibrinogen Binding Protein ofStaphylococcus aureus

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 248, 690–695 (1998)ARTICLE NO. RC989028
Identification of Functional Domains in Efb, a FibrinogenBinding Protein of Staphylococcus aureus
David Wade,*,1 Marco Palma,* Ingegerd Lofving-Arvholm,* Matti Sallberg,†Jerzy Silberring,‡ and Jan-Ingmar Flock*Department of Immunology, Microbiology, Pathology, and Infectious Diseases, *F82 and †F62, Huddinge Hospital,Karolinska Institute, S-141 86 Huddinge, Sweden; and ‡Faculty of Chemistry, Jagiellonian University,Ingardena 3, PL-30-060 Krakow, Poland
Received June 11, 1998
AA segments in its amino (N-) terminal region (Fig. 1).Staphylococcus aureus produces and secretes a pro- Similar repeating regions have been found in another
tein, Efb, that binds to fibrinogen, seems to be required S. aureus protein, coagulase, where 5 to 8 copies of afor virulence, and may benefit the microorganism by 27-residue repeating sequence are found in the carboxyldelaying wound healing. Interactions of Efb with fi- (C-) terminal region of the protein, and have been shownbrinogen are influenced by divalent metal cations, in- to bind to Fg (4-5). A model for the three dimensionalcluding Ca2/. Increasing concentrations of Ca2/ in-
(3D) structure of Efb predicts that the protein is globularcreased the binding of fibrinogen to immobilized Efb,and contains a substantial amount of a-helix in its C-whereas binding of Efb to immobilized fibrinogen wasterminal region, and the helix content has been verifieddecreased with increasing Ca2/ concentration. Studiesby circular dichroism (CD) analysis (6).with synthetic peptides showed that peptides from the
Fg and Efb have opposite polarities [net charges atcarboxyl terminal half of Efb bound to soluble fibrino-pH 7.3 and pI values are 020 (7) and 5.5 (8) for Fg,gen and enhanced the binding of fibrinogen to Efb. Aand /16 and 10.6 for Efb (1)] which makes it highlypeptide corresponding to a repeated sequence in theprobable that electrostatic interactions could occur be-amino terminal half of the protein also bound fibrino-tween them. There is also a 21-fold difference in theirgen and inhibited binding of fibrinogen to Efb. These
results may provide clues to the biological function of sizes (340 kDa for Fg versus 15.9 kDa for Efb), but theEfb and aid in the rational design of agents to block stoichiometry of binding between the two proteins hasthe Efb fibrinogen interaction. q 1998 Academic Press been shown to be 1:1 (9), indicating a specific interac-
tion. The interaction of Efb with Fg is influenced bythe divalent metal cations, calcium (Ca2/) and zinc(Zn2/) (9), both of which are present in the plasma (10),Staphylococcus aureus produces several proteinsand during an infection, Efb and Fg would be exposedthat interact with proteins of host cells, including ato these metals. Fg is known to have both low and high15.9 kDa, basic, protein, previously named Fib but nowaffinity binding sites for Ca2/ (11-12), and potentialdesignated Efb (extracellular fibrinogen binding pro-binding sites for both metals have also been locatedtein), that is secreted from the bacterial cell and bindswithin the Efb AA sequence (6).to fibrinogen (Fg) (1). Efb is produced constituitively,
Early investigations showed that the whole Efb pro-and by all isolates of S. aureus tested (2). The proteintein (136 AA’s), and its N-terminal half (69 AA’s, con-has been shown to be important for virulence of S.taining the two 22 AA repeats), bound to both solid-aureus, and it has been hypothesized that it might ben-phase Fg (Fg-Sepharose) and soluble Fg, whereas theefit the bacterium by binding to Fg and preventing itsC-terminal half (70 AA’s) bound only to solid-phase Fgconversion to fibrin, thereby delaying the healing pro-(1). More recent studies have confirmed that the bind-cess (3). However, it does not participate in bacterialing of Efb to Fg is divalent, with one binding site inadherence to Fg. the N-terminus and the other in the C-terminus (9).Efb consists of 136 amino acids (AA’s), with no cys- However, the precise sequence of AA’s in Efb that bindsteine residues, and it contains two nearly identical, 22 to Fg has not yet been determined, and a knowledge ofthe binding site residues could be useful in the design ofinhibitors to block the Efb-Fg interaction and possibly1 To whom correspondence should be addressed. Fax: /46-8-711-
3918; E-mail: [email protected]. reduce virulence.
0006-291X/98 $25.00Copyright q 1998 by Academic PressAll rights of reproduction in any form reserved.
690
AID BBRC 9028 / 6959$$$861 07-15-98 21:36:10 bbrcg AP: BBRC

Vol. 248, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
human Fg (3 mg/ml; 0.4 nmoles in assay; Fraction I, Type I fromMATERIALS AND METHODShuman plasma, Sigma Chemical Co., St. Louis, MO) in a total volumeof 50 ml. After 2 hours, Fg-bound and free peptides were separatedProtein purification. Efb was purified as described earlier (13).by centrifugation of the solution in Amicon Microcon 30 filters [Ami-Briefly, culture supernate was chromatographed through Fg-Sepha-con Inc., Beverly, MA; molecular weight cutoff (MWCO) Å 30 kDa;rose and the bound material was further purified by ion exchangeFg MW Å 340 kDa] for 15 minutes at 7,000 1 g. Aliquots of filtratechromatography (FPLC). Protein purity was assessed by electropho-were subjected to LSC, all assays were done in duplicate, and theresis on sodium dodecyl sulfate, polyacrylamide gel (SDS-PAGE).amount of peptide cpm that bound to Fg was calculated as a percent-
Peptide design, synthesis, and purification. Peptides were de- age of the free peptide cpm. Nonspecific binding of peptide to thesigned to be 20 AA’s in length, and to cover the entire AA sequence filter membrane was determined in the absence of Fg.of Efb, with 10 residue overlaps, starting from the N-terminal end
Effect of unlabelled Efb peptides on the binding of Fg to Efb.of the protein (Fig. 1). In addition, two peptides of 21 AA’s lengthELISA plate (Costar Corp., Cambridge, Massachusetts) wells werewere synthesized, representing the two repeat regions within the N-coated with 100 ml of Efb (3 mg/ml). After coating with Efb overnightterminal half of Efb. All peptides were synthesized as C-terminalat room temp., nonspecific binding sites were blocked by addition ofamides. The advantage of blocking the C-terminus is to eliminate100 ml of 2% bovine serum albumin (BSA) for 1 hr at 377C. Afteran additional negative charge that, except for the C-terminal peptidewashing in PBS, serial dilutions of peptides were added to the wells.{Efb [121-136]}, does not occur in the native protein, and which couldImmediately thereafter, 100 ml of Fg [1 mg/ml in PBS Tween (PBST)]destabilize secondary structures (e.g., a-helices) (14).was added to the wells and incubated for 1 hour at 377C. After wash-One set of peptides were radiolabelled at their N-termini withing, anti-Fg Ab-HRP conjugate (DAKO A/S, Glostrup, Denmark)[3H3]-acetic anhydride (3H-Ac2O) (15). This modification also elimi-(1:1,000 diluted) was added to the wells, and incubated for 1 hournated a positive charge that, except for the N-terminal peptide {Efbat 377C. After washing, OPD substrate (DAKO) was added to the[1-20]}, does not occur in the native protein, and resulted in a set ofwells, and the color reaction was monitored at 492 nm in a microplateN-terminal, [3H3]-acetylated, and C-terminal amidated peptides.reader.The peptides were synthesized by Fmoc methodology (16) on a
SyRo Multipler Synthese-Roboter (MultiSynTech, Bochum, Ger- Effect of Ca2/ on the interactions between Efb and Fg. Efb andmany) using a solid phase of Rink Amide MBHA (4-methyl-benzhy- Fg were dialysed separately against PBS containing 10 mM EDTA,drylamine) Resin, 0.5 mmol/g (Calbiochem-Novabiochem Corp., La and then the EDTA was removed by repetitive dialysis against sev-Jolla, California). Sources of AA’s were Millipore (Bedford, MA, eral changes of PBS. For all subsequent procedures, the proteinsUSA), PerSeptive Biosystems GmbH (Hamburg, Germany), and were diluted in Tris buffer (20 mM, 0.15 M NaCl, pH 7.3) to avoidAlexis Corporation (Laufelfingen, Switzerland). the formation and precipitation of calcium phosphate salts. ELISA
Crude peptides were analyzed for purity by reversed phase, high plates were coated with EDTA-treated Efb (0.5 mg/ml) or Fg (3 mg/performance liquid chromatography (RP-HPLC; Gilson Medical Elec- ml) as described above. Aqueous solutions of Ca2/ (0-50 mM) weretronics Inc., Middleton, Wisconsin) on a C2/C18 column of SuperPac added to wells of the plate, followed immediately by the addition ofPep-S, 5 mm, 41 250 mm (Pharmacia LKB, Uppsala, Sweden). Semi- EDTA-treated Fg (1 mg/ml) or Efb (10 mg/ml), respectively. The bind-preparative RP-HPLC purification was done on a C2/C18 column of ing of Fg to Efb was determined by anti Fg horse radish peroxidaseSuperPac Pep-S, 15 mm, 9.31 250 mm (Pharmacia Biotech, Uppsala, (HRP) conjugated antibodies. The binding of Efb to Fg was deter-Sweden). Both analytical and semi-preparative RP-HPLC were done mined by rabbit anti Efb antibodies followed by swine anti rabbitwith 2%/min gradients of water/0.1% TFA to 60% acetonitrile HRP conjugated antibodies. In both cases, the binding of HRP conju-(CH3CN)/0.1% TFA, and flow rates of 1ml/min (analytical) or 3 ml/ gated antibodies was detected by measuring the absorbance at 492min (semi-preparative). nm on an ELISA plate reader.
Radiolabelling of peptides. A 3H-acetylating reagent (1.5 mCi 3H- Test of the ability of Efb peptides to bind Ca2/. EquivalentAc2O/ml) was prepared to contain 5 mmoles each of Ac2O and trieth- amounts of radiolabelled Efb [1–20] (not predicted to bind Ca2/) andylamine (TEA) in 15 ml of DMF (15). After synthesis of the entire Efb [48-68], (second repeat region and predicted to bind Ca2/) werepeptide, the peptide resin was treated with 20% piperidine in DMF incubated with 0-100 mM CaCl2 in water at pH 6, as described aboveto remove the Na-Fmoc protecting group from the N-terminal AA. (see Binding of radiolabelled Efb peptides to soluble Fg), and thenThe peptide resin was then treated for 30 minutes with 5-10 equiva- subjected to centrifugal filtration with Amicon Micron 30 membranelents of acetylating agent per equivalent of peptide. filters. Radiolabelled peptide that passed through the filter was
Specific radioactivity was estimated by determining the peptide quantitiated by LSC, and the difference between the amount of radio-concentration of an aqueous solution of purified radiolabelled peptide activity added to the assay and the amount that passed through theby UV spectroscopy (17), and then measuring the radioactivity of a filter was equal to the amount of peptide bound to the filter. Nonspe-diluted aliquot of the same sample by liquid scintillation counting cific binding of peptide to the filters was determined in the absence(LSC). The mean specific radioactivity was estimated to be 14,000 of CaCl2.cpm/nmole of peptide.
Electrospray mass spectrometry (ESI MS). Mass spectra of Efb RESULTSprotein and Efb peptides were measured on a Finnigan MAT 95S(Finnigan MAT, Bremen, Germany), double focussing mass spec- Binding of peptides to soluble Fg. Fig. 1 shows thetrometer. The instrument was equipped with an electrospray source results of assays to determine the binding of radiola-and operated essentially as described previously (18). The samples
belled peptides to soluble Fg. In general, peptides corre-were diluted with a solution consisting of 30% CH3CN and 0.1%sponding to the C-terminal part of Efb and to the seg-formic acid in water and then injected onto the ESI source at a flow
rate of 30 ml/min. Spectra were taken in a positive-ion mode and ments near the second repeat region, bound soluble Fgfurther evaluated using deconvolution software supplied by the man- to the greatest extent. Five peptides bound soluble Fgufacturer. All synthetic peptides, except for the nonradiolabelled ver- to the extent of 10% or more [(Efb segment, average %sion of Efb [81-100], were found to have the expected molecular
bound): 31-50, 10%; 48-68, 10%; 91-110, 45%; 111-130,weights.12%, 121-136, 10%], which corresponded to an esti-Binding of radiolabelled Efb peptides to soluble Fg. Approxi-mated 0.2-1.1 moles of peptide bound per mole of solu-mately 10,000 cpm (1 nmole) of radiolabelled peptide was incubated,
in water at pH 6 and 4 7C, with a physiological concentration of ble Fg.
691
AID BBRC 9028 / 6959$$$861 07-15-98 21:36:10 bbrcg AP: BBRC

Vol. 248, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. The AA sequence of Efb, synthetic Efb peptides, and the binding of radiolabelled Efb peptides to soluble human Fg. The AAsequence of Efb, with single letter abbreviations for the amino acids and underlined repeating regions, is shown at the top of the figure.Sequences of synthetic Efb peptides are shown under the parent sequence. The results of binding assays, with N-terminal radiolabelledand purified Efb peptides, are shown in the bar graph to the right. The results are means or averages of two to three determinations. RR1,repeat region 1 [Efb (17–38)]; RR2, repeat region 2 [Efb (48–69)]; ND, not determined. The bars show that five of the peptides [Efb (31–50), Efb (48–68), Efb (91–110), Efb (111–130), and Efb (121–136)] bound Fg to the extent of 10% or greater of added peptide, with one[Efb (91–110)] binding an average of 45%.
Effect of Efb peptides on the binding of Fg to Efb.Fig. 2 shows the effect of pure, nonradiolabelled Efbpeptides on the binding of Fg to Efb. Peptides corre-sponding to segments of the N-terminal region of Efb(Efb [1-20] and the repeating regions, Efb [17-37] andEfb [48-68]) inhibited the binding of Fg to Efb that hadbeen immobilized on plastic microtitre (ELISA) plates.In contrast, peptides corresponding to segments of theC-terminal half of Efb (Efb [91-100], Efb [111-130], andEfb [121-136]) enhanced the binding of Fg to Efb. Twoadditional experiments were done, giving qualitativelysimilar results (data not shown).
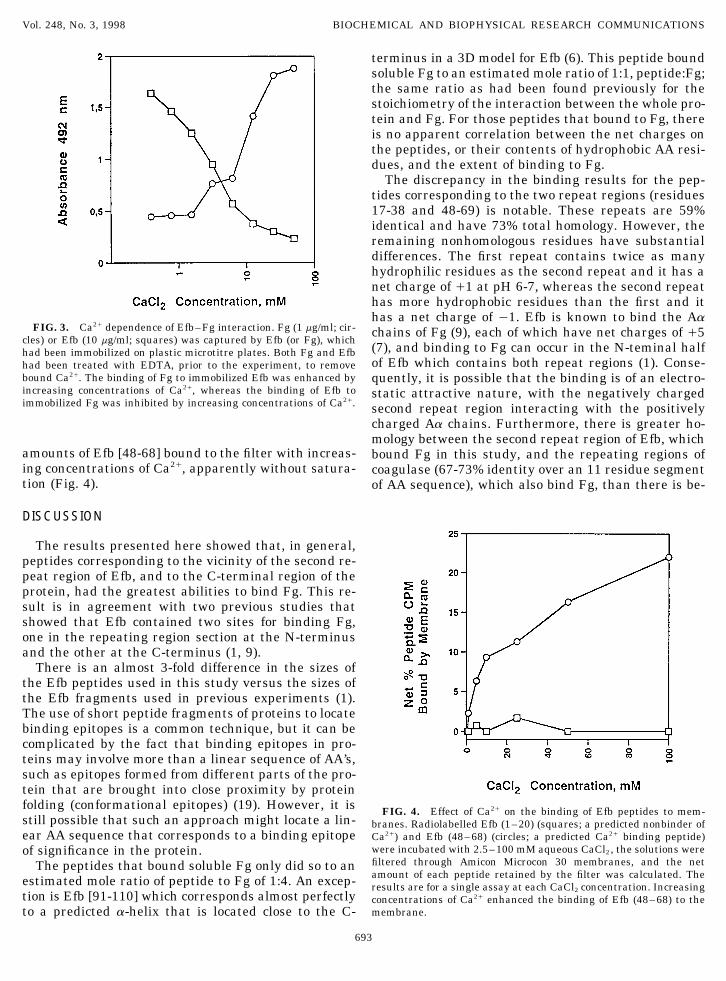
Effect of Ca2/ on the interactions between Efb andFg. Fig. 3 shows the results of experiments to deter-mine the effect of Ca2/ on the binding of Fg to Efb andvice versa. The results show that Ca2/ concentrationsabove 1 mM, and in the range of plasma Ca2/ concen-trations, caused two different types of effects whichdepended on the protein initially immobilized on themicrotitre plate. When Fg was the immobilized protein,
FIG. 2. The effect of Efb peptides on the binding of Fg to Efb.an increase in the Ca2/ concentration caused a decreaseThe results of one experiment are shown, and the results of twoin the binding of Efb to Fg. When Efb was the immobi- others were qualitatively similar. Pure, nonradiolabelled Efb pep-
lized protein, an increase in Ca2/ concentration caused tides, from the N-terminal region of Efb [Efb (1–20), squares] andthe two repeating regions [Efb (17–37), squares with cross, and Efban increase in the binding of Fg to Efb.(48–68), diamonds with cross], inhibited binding of Fg to immobilizedTest of the ability of Efb peptides to bind Ca2/. Radio- Efb. Peptides from the carboxyl terminal region of Efb [Efb (91–
labelled Efb [1-20] was not retained by the filter mem- 110), open diamonds, Efb (111–130), circles, and Efb (121–136),triangles] enhanced binding.brane at any concentration of Ca2/, whereas increasing
692
AID BBRC 9028 / 6959$$$861 07-15-98 21:36:10 bbrcg AP: BBRC
Vol. 248, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
terminus in a 3D model for Efb (6). This peptide boundsoluble Fg to an estimated mole ratio of 1:1, peptide:Fg;the same ratio as had been found previously for thestoichiometry of the interaction between the whole pro-tein and Fg. For those peptides that bound to Fg, thereis no apparent correlation between the net charges onthe peptides, or their contents of hydrophobic AA resi-dues, and the extent of binding to Fg.
The discrepancy in the binding results for the pep-tides corresponding to the two repeat regions (residues17-38 and 48-69) is notable. These repeats are 59%identical and have 73% total homology. However, theremaining nonhomologous residues have substantialdifferences. The first repeat contains twice as manyhydrophilic residues as the second repeat and it has anet charge of /1 at pH 6-7, whereas the second repeathas more hydrophobic residues than the first and ithas a net charge of 01. Efb is known to bind the Aa
FIG. 3. Ca2/ dependence of Efb–Fg interaction. Fg (1 mg/ml; cir- chains of Fg (9), each of which have net charges of /5cles) or Efb (10 mg/ml; squares) was captured by Efb (or Fg), which
(7), and binding to Fg can occur in the N-teminal halfhad been immobilized on plastic microtitre plates. Both Fg and Efbof Efb which contains both repeat regions (1). Conse-had been treated with EDTA, prior to the experiment, to remove
bound Ca2/. The binding of Fg to immobilized Efb was enhanced by quently, it is possible that the binding is of an electro-increasing concentrations of Ca2/, whereas the binding of Efb to static attractive nature, with the negatively chargedimmobilized Fg was inhibited by increasing concentrations of Ca2/. second repeat region interacting with the positively
charged Aa chains. Furthermore, there is greater ho-mology between the second repeat region of Efb, which
amounts of Efb [48-68] bound to the filter with increas- bound Fg in this study, and the repeating regions ofing concentrations of Ca2/, apparently without satura- coagulase (67-73% identity over an 11 residue segmenttion (Fig. 4). of AA sequence), which also bind Fg, than there is be-
DISCUSSION
The results presented here showed that, in general,peptides corresponding to the vicinity of the second re-peat region of Efb, and to the C-terminal region of theprotein, had the greatest abilities to bind Fg. This re-sult is in agreement with two previous studies thatshowed that Efb contained two sites for binding Fg,one in the repeating region section at the N-terminusand the other at the C-terminus (1, 9).
There is an almost 3-fold difference in the sizes ofthe Efb peptides used in this study versus the sizes ofthe Efb fragments used in previous experiments (1).The use of short peptide fragments of proteins to locatebinding epitopes is a common technique, but it can becomplicated by the fact that binding epitopes in pro-teins may involve more than a linear sequence of AA’s,such as epitopes formed from different parts of the pro-tein that are brought into close proximity by proteinfolding (conformational epitopes) (19). However, it is FIG. 4. Effect of Ca2/ on the binding of Efb peptides to mem-still possible that such an approach might locate a lin- branes. Radiolabelled Efb (1–20) (squares; a predicted nonbinder ofear AA sequence that corresponds to a binding epitope Ca2/) and Efb (48–68) (circles; a predicted Ca2/ binding peptide)
were incubated with 2.5–100 mM aqueous CaCl2, the solutions wereof significance in the protein.filtered through Amicon Microcon 30 membranes, and the netThe peptides that bound soluble Fg only did so to anamount of each peptide retained by the filter was calculated. Theestimated mole ratio of peptide to Fg of 1:4. An excep- results are for a single assay at each CaCl2 concentration. Increasing
tion is Efb [91-110] which corresponds almost perfectly concentrations of Ca2/ enhanced the binding of Efb (48–68) to themembrane.to a predicted a-helix that is located close to the C-
693
AID BBRC 9028 / 6959$$$861 07-15-98 21:36:10 bbrcg AP: BBRC

Vol. 248, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
tween the first repeat region of Efb, which does not ity of soluble Efb for Ca2/ than for Fg. This hypothesisis partially supported by the results of the test of thebind Fg, and the repeat regions of coagulase.
The results obtained in assays of the effect of unla- ability of Ca2/ to enhance the binding of Efb peptidesto Amicon Microcon 30 filters, in which it was foundbelled Efb peptides on the binding of Fg to Efb were as
might be expected from the results obtained in the ear- that Efb [48-68] (which includes the predicted Ca2/
binding site at residues 51-58), but not Efb [1-20],lier experiments (1), where it was shown that residues1-69 of Efb bound to both solid phase and soluble Fg. bound to the filter in increasing amounts as the Ca2/
concentration was increased. Amicon Microcon 30 fil-As might be expected, peptides from this region (Efb [1-20] and both repeat regions) were capable of competing ters are composed of cellulose, which absorbs water and
could coordinate Ca2/ through its own hydroxyl groupswith Efb for interaction with Fg. The enhancement ofFg binding to Efb by peptides from the C-terminal re- and those of bound water. Both of the test peptides
had the same length (20 AA’s), a similar number ofgion of Efb was a surprising finding, and possiblecauses for this result are currently under investigation. hydrophobic AA’s (7-9 for Efb [1-20] and 9-10 for Efb
[48-68]), and an equal number of acidic AA’s (3 perMany proteins are known to bind metals, includinga few divalent metal binding proteins that are secreted peptide) which could serve as Ca2/ binding sites. How-
ever, they differed in their net charges at assay pHby Staphylococci (20). Also, it has been shown that highconcentrations (25-200 mM) of Ca2/ and Zn2/, but not (Efb [1-20] Å /1 and Efb [48-68] Å 01), and also in the
distribution of acidic AA’s within the peptides (widelyMg2/ (all chloride salts), stimulated the precipitationof 1:1 complexes of Efb and Fg from equimolar mixtures distributed in Efb [1-20] and clustered in Efb [48-68]).
The results obtained in the presence of Ca2/, no addi-of these two proteins (9). Ca2/ is involved in many ofthe steps of the sequence of reactions comprising blood tional binding by positively charged Efb [1-20] and
greatly enhanced binding of negatively charged Efbclotting, and it is present in human plasma at a concen-tration of 2.5 mM (10), of which 1.2 mM is in a free [48-68], might be due to electrostatic repulsion in
the first situation and electrostatic attraction in theform. Therefore, during a S. aureus infection, the bacte-rium, the Efb that it secretes, and Fg would all be second.
In summary, the second repeat region of Efb, repre-exposed to plasma Ca2/ at millimolar levels. It seemedpossible that Efb protein, or peptides corresponding to sented by peptide Efb [48-68], has the dual ability to
bind both to Fg and Ca2/. We have also confirmed previ-portions of the Efb AA sequence, might also bind Ca2/
and/or other divalent metal ions, such as Zn2/. Such a ous notions that an interaction between Fg and Efbtakes place at the C-terminal portion of Efb.potential Ca2/ binding site can be predicted at residues
51-58 (i.e., E-F-N-D-G-T-F-E), within peptide Efb[48-68]. The results presented here (Fig. 4) support that ACKNOWLEDGMENTSprediction.
Portions of this work were presented at the Fifth InternationalIt is obvious from Fig. 3 that the presence of Ca2/ atSymposium on Solid Phase Synthesis and Combinatorial Chemicalplasma concentrations also had effects on the interac-Libraries, Imperial College, London, England, September 2–6, 1997.tions between Efb and Fg. The binding of soluble Efb D.W. thanks Dr. Z.-X. Zhang for technical advice on the peptide
to immobilized Fg was greater than half maximal at syntheses, Dr. Kurt D. Berndt for CD analyses of Efb, and also thethese concentrations of Ca2/, but as the concentration Swedish Medical Research Council and the Karolinska Institutet for
fellowship support during 1996–1998. Grants were received fromof Ca2/ was increased, interaction was inhibited. Thethe Swedish Medical Research Council (K97-16X-12218-01A).decrease in affinity of Efb for immobilized Fg, that ac-
companies an increase in the Ca2/ concentration (Fig.REFERENCES3), is not due to changes in the structure of Efb because
CD experiments with purified Efb have shown no1. Boden, M. K., and Flock, J.-I. (1994) Mol. Microbiol. 12, 599–change in the structure of Efb over the range of 0-50 606.
mM CaCl2 (data not shown). At plasma Ca2/ concentra- 2. Boden, M. K., and Flock, J.-I. (1995) J. Clin. Microbiol. 33, 2347–tions, the binding of soluble Fg to immobilized Efb was 2352.only about 25% of maximal, but binding was enhanced 3. Palma, M., Nozohoor, S., Schennings, T., Heimdahl, A., andas the Ca2/ concentration was increased. Fg is known Flock, J.-I. (1996) Infect. Immun. 64, 5284–5289.to contain both high and low affinity binding sites for 4. Phonimdaeng, P., O’Reilly, M., Nowlan, P., Bramley, A. J., and
Foster, T. J. (1990) Mol. Microbiol. 4, 393–404.Ca2/ (11-12; human Fg contains 3 high affinity sites5. Kaida, S., Miyata, T., Yoshizawa, Y., Kawabata, S., Morita, T.,with Kd Å 19 mM), and both types of sites would be
Igarashi, H., and Iwanaga, S. (1987) J. Biochem. 102, 1177–saturated at plasma Ca2/ concentrations. Conse-1186.quently, the effects obtained by varying the Ca2/ con-
6. Wade, D., Palma, M., Flock, J.-I., Berndt, K., Silberring, J., andcentration were probably not due to changes in the Galaktionov, S. G. (1998) Protein Peptide Letts., submitted forbinding of Ca2/ by Fg. However, the inhibition of solu- publication.ble Efb binding to immobilized Fg, with increasing Ca2/
7. Doolittle, R. F., Watt, K. W. K., Cottrell, B. A., Strong, D. D., andRiley, M. (1979) Nature 280, 464–468.concentration, may have been due to the greater affin-
694
AID BBRC 9028 / 6959$$$861 07-15-98 21:36:10 bbrcg AP: BBRC

Vol. 248, No. 3, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
8. Mihalyi, E. (1970) Biochemistry 9, 804–816. 15. Stewart, J. M., and Young, J. D. (1984) Solid Phase Synthesis,2nd ed., Pierce Chemical Company, Rockford, IL.9. Palma, M., Wade, D., Flock, M., and Flock, J.-I. (1998) J. Biol.
16. Atherton, E., and Sheppard, R. C. (1989) Solid Phase PeptideChem., in press.Synthesis, A Practical Approach, IRL Press, Eynsham, Oxford,10. Ganong, W. F. (1997) Review of Medical Physiology, 18th ed.,England.Appleton & Lange, Stamford, CT.
17. Segel, I. H. (1976) Biochemical Calculations, 2nd ed., John Wiley11. Marguerie, G., Chagniel, G., and Suscillon, M. (1977) Biochim. and Sons, New York, NY.
Biophys. Acta 490, 94–103.18. Nylander, I., Tan-No, K., Winter, A., and Silberring, J. (1995)
12. Nieuwenhuizen, W., van Ruijven, A. M., Nooijen, W. J., Ver- Life Sci. 57, 123–129.mond, A., and Haverkate, F. (1981) Thromb. Res. 22, 653–657. 19. Kaumaya, P. T. P., Kobs-Conrad, S., and DiGeorge, A. M. (1994)
13. Boden, M. K., and Flock, J.-I. (1992) Microbial Pathogenesis 12, in Innovation and Perspectives in Solid Phase Synthesis: Pep-289–298. tides, Proteins and Nucleic Acids (Epton, R., Ed.), Mayflower
Worldwide Limited, Birmingham, England.14. Shoemaker, K. R., Kim, P. S., York, E. J., Stewart, J. M., andBaldwin, R. L. (1987) Nature 326, 563–567. 20. Ayora, S., and Gotz, F. (1994) Mol. Gen. Genetics 242, 421–430.
695
AID BBRC 9028 / 6959$$$861 07-15-98 21:36:10 bbrcg AP: BBRC


















