


Identification, characterization, and molecular application of a ...
45
1 Identification, characterization, and molecular application of a virulence-associated autotransporter from a pathogenic Pseudomonas fluorescens strain Yong-hua Hu 1,2 , Chun-sheng Liu 1,2 , Jin-hui Hou 3 , and Li Sun 1 * Institute of Oceanology, Chinese Academy of Sciences, Qingdao, PR China 1 ; Graduate University of the Chinese Academy of Sciences, Beijing, PR China 2 ; and Xuzhou Institute of Technology, Xuzhou, PR China 3 *To whom correspondence should be addressed Mailing address: Institute of Oceanology Chinese Academy of Sciences 7 Nanhai Road Qingdao 266071 PR China Phone and Fax: 86-532-82898834 Email: [email protected] Running title: autotransporter of P. fluorescens Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Appl. Environ. Microbiol. doi:10.1128/AEM.00159-09 AEM Accepts, published online ahead of print on 15 May 2009 on April 5, 2018 by guest http://aem.asm.org/ Downloaded from
Transcript of Identification, characterization, and molecular application of a ...

1
Identification, characterization, and molecular application
of a virulence-associated autotransporter from a
pathogenic Pseudomonas fluorescens strain
Yong-hua Hu
1,2, Chun-sheng Liu
1,2, Jin-hui Hou
3, and Li Sun
1*
Institute of Oceanology, Chinese Academy of Sciences, Qingdao, PR China1; Graduate University of the
Chinese Academy of Sciences, Beijing, PR China2; and Xuzhou Institute of Technology, Xuzhou, PR
China3
*To whom correspondence should be addressed
Mailing address:
Institute of Oceanology
Chinese Academy of Sciences
7 Nanhai Road
Qingdao 266071
PR China
Phone and Fax: 86-532-82898834
Email: [email protected]
Running title: autotransporter of P. fluorescens
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.00159-09 AEM Accepts, published online ahead of print on 15 May 2009
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 1
2
A gene, pfa1, encoding an autotransporter was cloned from a pathogenic Pseudomonas fluorescens strain, 3
TSS, isolated from diseased fish. Expression of pfa1 is enhanced during infection and is regulated by 4
growth phase and growth conditions. Mutation of pfa1 significantly attenuates the overall bacterial 5
virulence of TSS and impairs the abilities of TSS in biofilm production, interaction with host cells, 6
modulation of host immune responses, and dissemination in host blood. The putative protein encoded by 7
pfa1 is 1242 amino acids in length and characterized by the presence of three functional domains that are 8
typical for autotransporters. The passenger domain of Pfa1 contains a putative serine protease (Pap) that 9
exhibits apparent proteolytic activity when expressed in and purified from E. coli as a recombinant protein. 10
Consistent with the important role played by Pfa1 in bacterial virulence, purified recombinant Pap has a 11
profound cytotoxic effect on cultured fish cells. Enzymatic analysis showed that recombinant Pap is 12
relatively heat-stable and has an optimal temperature and pH of 500C and pH 8.0. The domains of Pfa1 that 13
are essential to autotransporting activity were localized, and, on the basis of this, a Pfa1-based autodisplay 14
system (named AT1) was engineered to facilitate the insertion and transport of heterologous proteins. 15
When expressed in E. coli, AT1 was able to deliver an integrated Edwardsiella tarda immunogen (Et18) 16
onto the surface of bacterial cells. Compared to purified recombinant Et18, Et18 displayed by E. coli via 17
AT1 induced significantly enhanced immunoprotection. 18
19
Keywords: autotransporter, autodisplay, protease, Pseudomonas fluorescens, vaccine delivery, virulence 20
21
22
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
INTRODUCTION 23
24
Protein secretion plays important roles in bacterial life, as many of the secreted proteins are involved in 25
biological processes that are fundamental to the survival and environmental adaptations of the cells. 26
Gram-negative bacteria have evolved a number of secretion systems that utilize different secretion 27
apparatus and mechanisms (9). The classical autotransporter secretion pathway belongs to the type V 28
secretion system (8, 12). Compared to other types of secretion mechanisms, the autotransporter system is 29
unique in the feature that all the components that are required for protein translocation are contained within 30
a single polypeptide. Structurally, autotransporters are characterized by three domains: (i) an N-terminal 31
signal sequence that is recognized by the Sec translocon; (ii) a central passenger domain (or α domain) that 32
contains the effector molecule and is highly variable; (iii) a C-terminal translocation domain (or 33
β/autotransporter domain) that is conserved in length (250-300 amino acids) but varies in primary structure 34
(7, 12, 55). In most cases, the β-domain contains 12 antiparallel strands of 9 to 12 residues that, upon 35
integration into the outer membrane, form a β-barrel conformation (27, 57). Another conserved feature of 36
the β-domain is the presence at the C-terminus a sequence motif, (Y/V/I/F/W)-X-(F/W), that is 37
characterized by alternating hydrophobic and hydrophilic residues and ended with either a tryptophan or a 38
phenylalanine (12, 16, 27). The integrity of this end motif seems to be required for protein translocation, as 39
deletion of certain residues in this sequence impairs protein secretion. The secretion process of 40
autotransporter is initiated by the signal sequence, which directs the translocation of the protein precursor 41
across the inner membrane into the periplasmic space via the Sec system. Once inside the periplasm, the 42
β-domain inserts into the outer membrane and adopts the structure of a β-barrel, through which the 43
passenger domain is translocated to the cell surface where it may exist as a membrane-anchored protein 44
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
covalently linked to the β-domain or be cleaved from the β-domain as a result of proteolysis (7). 45
Since the discovery of the gonococcal immunoglobulin A1 protease (35), the first autotransporter, and 46
especially with the advent of genome sequencing technology, autotransporters have been identified in many 47
bacterial species (18, 19, 28, 33). Functions assigned to autotransporters are mostly associated with 48
bacterial pathogenicity, which include adhesion and invasion into host cells, biofilm formation, and 49
cytotoxicity (11, 54). In the present study, we identified and analyzed an autotransporter, Pfa1, from a 50
pathogenic Pseudomonas fluorescens strain isolated from diseased fish. We found that, like many of the 51
autotransporters identified in other pathogens, Pfa1 is a virulence factor that is involved in interactions with 52
host cells and modulation of host immune responses via a protease effector. In addition, we found that the 53
autotransporter property of Pfa1 could be exploited for the delivery and surface display of an 54
immunoprotective antigen. 55
. 56
MATERIALS AND METHODS 57
58
Bacterial strains and growth conditions. Bacillus sp. strain B187 (59), Edwardsiella tarda strain 59
TX1 (fish pathogen) (58), and Pseudomonas fluorescens strain TSS have been reported previously (52). 60
TSS was isolated from diseased Japanese flounder and genetically identified by 16S rDNA sequence 61
(Genbank accession no. FJ896114) analysis. The pathogenicity of TSS to Japanese flounder has been 62
described previously (52). Escherichia coli strain DH5α is a commercial strain from Takara (Dalian, 63
China). All strains were cultured in Luria-Bertani broth (LB) medium (39) at 370C (for E. coli) or 28
0C 64
(for all others). Cell cultures were maintained at 40C for short-term storage (1 to 3 days) and at -80
0C in 65
culturing medium containing 10 to 15% (v/v) glycerol for long-term storage. Appropriate antibiotics were 66
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
added at the following concentrations: ampicillin (Ap), 100 µg/ml; kanamycin (Kn), 50 µg/ml; tetracycline 67
(Tc), 15 µg/ml. 68
Plasmid construction. The plasmids and primers used in this study are listed in Table 1 and Table 2 69
respectively. pEPA1 was created by inserting the coding sequence of Pap (amplified by PCR with primers 70
F15/R13) into pET258 between the NdeI/XhoI sites. To construct pEPA2, overlapping PCR were 71
performed with the primer pairs F15/MR2 and MF2/R13, followed by a fusion PCR with the primers 72
F15/R13; the PCR products were inserted into pET258 between the NdeI/XhoI sites. To create pB328, the 73
ScaI site of pBR322 was mutated as follows: overlapping PCR were performed with the primer pairs 74
ApF8/ApMR1 and ApMF1/ApR7, followed by a fusion PCR with primers ApF8/ApR7; the PCR products 75
were digested with EcoRI/PstI, and the 750 bp DNA was inserted into pBR322 between the EcoRI/PstI 76
sites. To construct pAT1, the 657 bp DNA containing the 5’ region and upstream sequence of pfa1 was 77
amplified by PCR with primers F11/R9, and the PCR products were ligated into pB328 and pBS-T at the 78
EcoRV site, resulting in p328AT1 and pBSAT1 respectively; a 2 kb DNA was amplified by PCR from the 79
3’ region of pfa1 using primers F12/R12, and the PCR products were ligated into pBS-T at the EcoRV site, 80
resulting in pBSAT2, which was digested with ScaI, and the 2 kb DNA fragment was ligated into p328AT1 81
at the EcoRV site, yielding pAT1. To construct pAT2 and pAT3, pBSAT2 was digested with EcoRV/ScaI 82
and SmaI/ScaI respectively, and the resulting 1890 and 1420 bp DNA fragments were inserted into 83
p328AT1 at the EcoRV site. pAT1V, pAT2V, and pAT3V were constructed by inserting agaV (amplified by 84
PCR with primers UAF13/UAR14) into pAT1, pAT2, and pAT3 at the ScaI site. To construct pAT18, the 85
coding sequence of the processed Et18 preceded by an 8-glycine linker was amplified by PCR with 86
primers 18F9/18R11; the PCR products were inserted into pAT1 at the ScaI site. 87
Cloning of pfa1. pfa1 was cloned by using the in vivo-induced antigen technology (IVIAT) as 88
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
described by Kim et al. (20). Briefly, for the construction of a BL21(DE3)-based inducible expression 89
library, 0.5 to 3 kb Sau3A1 fragments of TSS genomic DNA were ligated into pET28abc (Novagen, USA), 90
and the ligation mixture was electroporated into electrocompetent DH5α. The transformants were used for 91
the preparation of a plasmid library, which was subsequently introduced into BL21(DE3) (Tiangen, Beijing, 92
China) by transformation. To prepare TSS antisera, adult rabbits were immunized with TSS, and antisera 93
were pooled and absorbed with TSS whole cells and cell lysate. As a negative control, sera from 94
PBS-immunized rabbits were pooled and similarly treated. For the screening of in vivo-expressed genes, 95
the genomic expression library was plated on LB agar plates, and, by using nitrocellulose disks, the 96
colonies appeared on the plates were replica plated onto LB agar plates supplemented with 97
isopropyl-β-D-thiogalactopyranoside (IPTG). After incubation at 370C, the colonies that emerged were 98
lysed by exposure to chloroform vapor. The nitrocellulose disks were blotted with TSS antisera. Positive 99
colonies were immunoblotted with the control sera, and the resulting negative colonies were blotted again 100
with TSS antisera. Plasmids were extracted from positive colonies, and the TSS DNA inserts in the 101
plasmids were identified by DNA sequencing. One of the inserts contains pfa1 truncated at the 3’ end. The 102
complete gene of pfa1was obtained by genome walking as described previously (60). 103
Construction of a pfa1 null strain (TSM). To construct TSM, an internal 766 bp DNA (positions 104
1354 to 2119) of pfa1 was generated by PCR with primers F6/R14; the PCR products were inserted into 105
pBSAT1 at the EcoRV site, resulting in pBSAT3, which was digested with SwaI, and the 1423 bp DNA 106
fragment was inserted into p7TS at the SmaI site, resulting in p7TSP1. p7TSP1 was introduced into 107
S17-1λpir (Biomedal, Spain) by transformation. S17-1λpir/p7TSP1 was conjugated with TSS. The 108
transconjugants were selected first on LB plates supplemented with tetracycline and kanamycin and then on 109
LB plates supplemented with 5% sucrose and kanamycin. The colonies that were resistant to sucrose and 110
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
sensitive to tetracycline were analyzed by PCR; the PCR products were subsequently subjected to DNA 111
sequencing to confirm the deletion in pfa1. 112
Bacterial conjugation. This was performed as described previously (58). 113
Quantitative real-time reverse transcriptase PCR (qRT-PCR). qRT-PCR was carried out in an ABI 114
7300 Real-time Detection System (Applied Biosystems) by using the SYBR ExScript qRT-PCR Kit 115
(Takara) as described previously (59). Each assay was performed in triplicate with 16S rRNA as controls. 116
The primers used for qRT-PCR of pfa1 and 16S rRNA were RTF33/RTR10 (Table 2) and 933F/16SRTR1 117
(59), respectively. All data are given in terms of relative mRNA, expressed as means plus or minus 118
standard errors of the means (SE). 119
Bacterial cell number determination. Plate count was used to determine the number of viable TSS 120
corresponding to an OD600 of 1, which is ~109 CFU. The bacterial cell numbers indicated in all experiments 121
were estimated based on OD600 measurements. 122
Expression of pfa1 under different growth conditions. To examine pfa1 expression under in vivo 123
conditions, Japanese flounder were injected intraperitoneally (i.p.) with 107 CFU of TSS. The livers of the 124
fish (five) were taken at 24 h post-infection and used for total RNA preparation with the RNAprep 125
Tissue/Bacteria Kit (Tiangen, Beijing, China). To examine pfa1 expression under in vitro conditions and in 126
relation to growth phase, TSS was cultured in LB medium at 280C to an OD600 of ~1.8 and used for total 127
RNA extraction as described above. To examine pfa1 expression at different temperatures or pH, TSS was 128
cultured to an OD600 of ~1.8 in LB medium at various temperatures or in LB medium with different pH at 129
280C; the cells were then used for total RNA extraction. The same amounts of total RNA from each 130
preparation were used for qRT-PCR. 131
Biofilm assay. Cells were cultured in LB medium to exponential phase and diluted to 105 CFU/ml. 132
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
The diluted cells were transferred into a 96-well polystyrene plate (Nunc, Danmark) and incubated at 280C 133
for 24 h without agitation. After the incubation, the unattached cells were removed from the wells by using 134
a pipette, and the wells were washed 5× with PBS. The attached cells were treated with Bouin fixative for 1 135
h and stained with 1% crystal violet solution for 20 min. The unbound dye was removed by rinsing the 136
plate several times with running water. The plate was air dried. The bound dye was eluted in ethanol, and 137
the eluates were measured for absorbance at A570. 138
Serum bactericidal activity assay. This was performed as described previously (52). 139
Interaction of bacteria with cultured fish cells. Japanese flounder gill cells (FG cells) were cultured 140
and maintained as described previously (47). Examination of interactions between FG cells and TSS/TSM 141
was performed according to the method of Kolodziejek et al. (22). Briefly, FG cells were cultured in 142
96-well cell culturing plates to monolayer and mixed with TSS or TSM. After incubation at 300C for 1 h, 143
the plates were washed 5× with PBS. To determine the number of bacterial cells associated with the entire 144
FG cell, the washed FG cells were lysed with 200 µl 1% (v/v) Triton X-100 in PBS, and 50 µl of the lysate 145
were plated on LB agar plates supplemented with kanamycin. After incubation overnight at 300C, the 146
number of colonies appeared on the plates were counted. To determine the number of bacterial cells that 147
had penetrated into FG cells, the above washed FG cells were incubated with penicillin and streptomycin 148
(100 U) for 2 h at 300C to kill the extracellular bacteria. FG cells were lysed and plated as above. 149
Experimental infection and virulence determination. TSS and TSM were cultured to an OD600 of 150
0.7 in LB medium and resuspended in PBS to 3 × 108
CFU/ml. Japanese flounder were divided randomly 151
into three groups (20 fish/group), and each group was injected i.p. with 100 µl of TSS or TSM suspension 152
or PBS. The fish were monitored for mortality for 14 days, during which time the fish were maintained at 153
220C in aerated seawater that was changed twice daily. Cumulative mortalities were calculated at the end of 154
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
the monitored period. 155
To examine bacterial dissemination in blood, Japanese flounder were challenged with TSS or TSM as 156
described above. Blood was taken from the fish at 6 h post-infection and, after serial dilution, plated in 157
triplicate on LB agar plates supplemented with kanamycin. After incubation at 280C for 48 h, the colonies 158
that appeared on the plates were enumerated. The genetic nature of these colonies was verified by PCR 159
analysis using primers specific to TSS and TSM. The PCR products were randomly selected for DNA 160
sequencing. 161
Respiratory burst assay. Two groups (five fish/group) of Japanese flounder were i.p. injected with 162
the same dose (107 CFU) of TSS or TSM. Blood was colleted from the caudal veins of the fish at different 163
time points after the infection and used for respiratory burst assay, which was performed exactly according 164
to the method of Kumar et al. (24). 165
Purification and reconstitution of recombinant proteins. For the purification of recombinant Pap 166
and PapM, BL21(DE3) harboring pEPA1 and pEPA2 were grown in LB medium to an OD600 of 0.7, 167
followed by adding to the culture 1 µM of IPTG. After an additional 5 h of growth, recombinant proteins 168
were purified under denaturing conditions by using nickel-nitrilotriacetic acid (Ni-NTA) columns (GE 169
Healthcare, USA) as recommended by the manufacturer. The proteins were then reconstituted as follows: 170
the proteins were incubated with Ni-NTA agarose (QIAGEN, USA) in the lysis buffer (100 mM NaH2PO4, 171
10 mM Tris.Cl, and 8 M urea; pH 8.0) for 30 min at 200C and then dialyzed against the reconstitution 172
buffer containing 50 mM Tris.Cl, 200 mM KCl, 10 mM MgCl2, 10 uM ZnCl2, 5 mM β-mercaptoethanol, 1 173
mM EDTA, 20% glycerol, and decreasing concentrations of urea (4, 2, 1, 0.5, 0.25, and 0 M). 174
Reconstituted proteins were eluted in elution buffer (50 mM NaH2PO4, 300 mM NaCl, and 250 mM 175
imidazole; pH 8.0) and dialyzed overnight against 2 liters of PBS. The proteins were concentrated using 176
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
Amicon Ultra Centrifugal Filter Devices (Millipore, USA). Purification of recombinant Et18 was described 177
previously (14). 178
Protease activity assay. The protease activity of purified recombinant Pap and PapM was analyzed as 179
described previously in the assay buffer with azocasein as the substrate (59). One unit of enzyme activity 180
was defined as an increase of 0.001 at A350 over the control (azocasein incubated with PBS). The effect of 181
Na+, Ca
2+, Mg
2+, and Zn
2+ was determined by adding each of the metals to the assay buffer at 1 and 10 mM. 182
Antisera. Antisera to recombinant Pap was prepared by subcutaneously injecting adult rats (~242 g) 183
with 60 µg of the purified recombinant protein. The animals were boosted, and the blood was collected as 184
described previously (46). The titer and specificity of the antisera were determined by enzyme-linked 185
immunosorbent assay (ELISA) and Western and immunoblotting analysis as described previously (45). 186
Western and immunoblotting analysis. Cells were grown in LB medium to an OD600 of 1. Proteins 187
of the extracellular, periplasm, cytoplasm, and outer membrane were prepared as described previously (5, 188
45, 59). The supernatant proteins were concentrated ~300 times by using Amicon Ultra Centrifugal Filter 189
Devices (Millipore). The proteins were analyzed by sodium dodecyl sulfate-polyacrylamide gel 190
electrophoresis (SDS-PAGE), and immunoblotting was performed as described previously (45, 59) using 191
anti-Pap or anti-His antibodies (Tiangen, Beijing, China). 192
Cytotoxic effect of Pap on FG cells. This was performed according to the method of Alamuri and 193
Mobley (2). In brief, FG cells were cultured to confluence in 96-well plates. Different concentrations of 194
Pap or PapM or PBS was added to the cells. After incubation at 20°C for various times, the cells were used 195
either for the determination of viability using the MTT Cell Proliferation and Cytotoxicity Assay Kit 196
(Beyotime, Beijing, China) or for microscopic observation. For the latter purpose, the cells were fixed with 197
4% paraformaldehyde for 10 min, washed with PBS, stained with Giemsa solution (Solarbio, Beijing, 198
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
China), and then destained with 10% methanol. The plate was dried and examined under an inverted 199
microscope. 200
Whole cell enzyme-linked immunosorbent assay (ELISA). Whole cell ELISA was performed as 201
described previously (45). Briefly, 107 CFU of TSS or TSM or PBS (the control) was added to a 96-well 202
ELISA plate. After blocking with BSA, the cells were treated with mouse anti-His antibody and then with 203
horse radish peroxidase-conjugated goat anti-mouse IgG (BIOS, Beijing, China). The plate was read at 450 204
nm, and positive readings were defined as at least twice of that of the control. 205
Vaccination. This was performed as described previously (59). Briefly, DH5α/pAT18, DH5α/pAT1, 206
and B187 were cultured in LB medium and resuspended in PBS to 2 x 108 CFU/ml. Purified recombinant 207
Et18 was suspended in B187 to 200 µg/ml (named Et18-B187 mix). Healthy Japanese flounder (~12 g) 208
were divided randomly into four groups (30 fish/group) designated A to D, which were injected i.p. with 209
100 µl of DH5α/pAT18, DH5α/pAT1, Et18-B187 mix, and PBS, respectively. At the 20th
day 210
post-immunization, groups A, B, and D were boosted with the same amount of bacterial cells or PBS used 211
in the initial immunization; group C was boosted with 20 µg of Et18 suspended in PBS without B187. The 212
fish were challenged via i.p. injection with 5 × 106 CFU of E. tarda TX1 at the 14
th day post-boost and 213
monitored for mortality for 14 days. Relative percent of survival (RPS) of the fish was calculated according 214
to the method of Amend (3): RPS = {1 – (% mortality in vaccinated fish/% mortality in control fish)} × 215
100. 216
Statistical analysis. All statistical analyses were performed using SPSS 15.0 software (SPSS Inc., 217
USA). Differences in the cytotoxic effect of Pap and the interaction between Pap and FG cells were 218
analyzed using one-way analysis of variance (ANOVA); the vaccination data were analyzed using 219
chi-square test. All other statistical analyses were performed using Student t test. In all cases, the 220
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
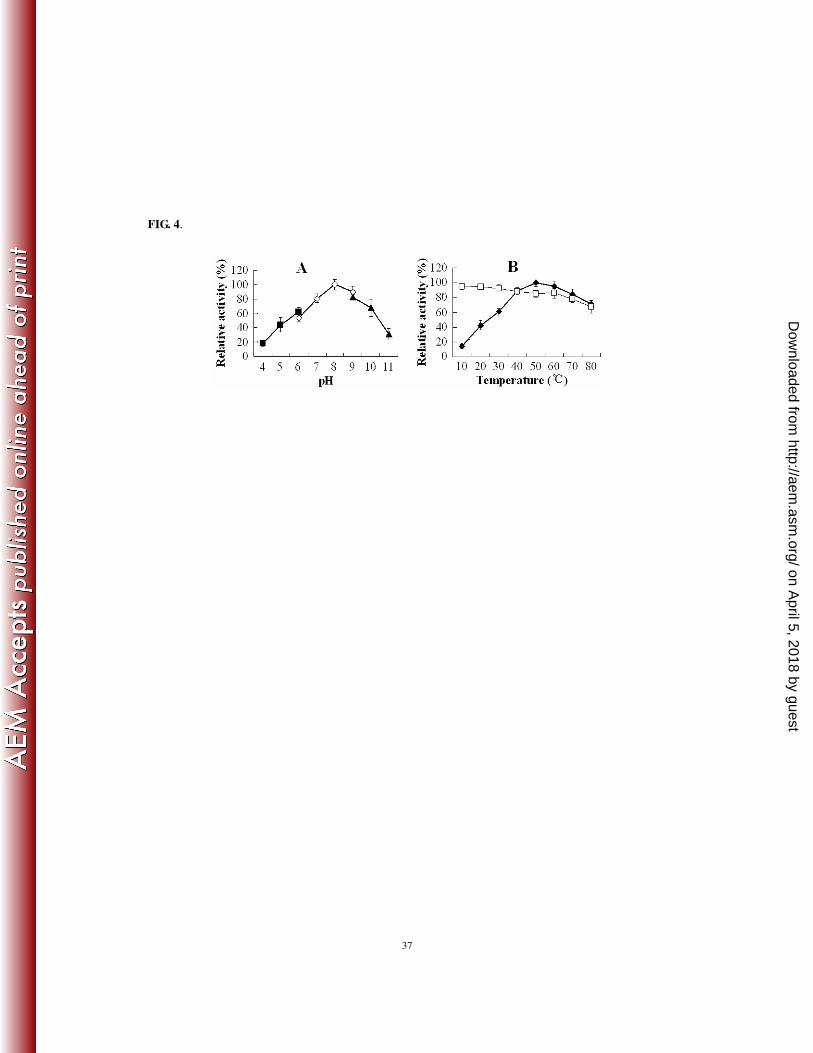
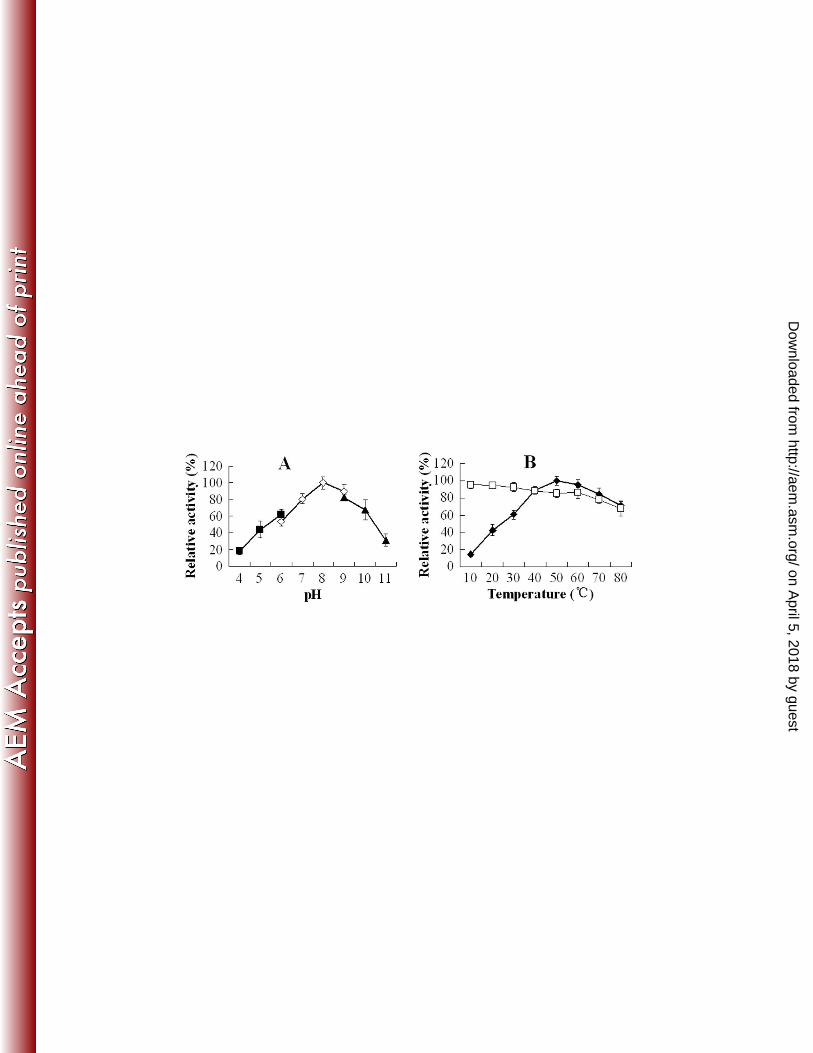
significance level was defined as P < 0.05. 221
Database search and in silico analysis. Database search was conducted using the BLAST programs 222
at the NCBI. Structural analysis was performed using the MOTIF, SMART, and NCBI Conserved Domain 223
Search servers. The secondary structure of the autotransporter domain of Pfa1 was predicted by using 224
AnthePro 5. Signal peptide search was performed by using SignalP 3.0. 225
Nucleotide sequence accession numbers. The nucleotide sequence of pfa1 has been deposited in the 226
GenBank database under the accession number FJ539022. 227
228
RESULTS 229
230
Sequence characterization of Pfa1. Pfa1 codes for a putative protein of 1242 amino acids. Sequence 231
BLAST analyses indicated that the closest homologues of Pfa1 are two outer membrane autotransporters 232
from the P. fluorescens strain Pf0-1 and the Serratia proteamaculans strain 568 (GenBank accession nos. 233
ABA74424 and ABV43830, respectively), which share approximately 37% overall sequence identity with 234
Pfa1. Bioinformatic analyses using SignalP 3.0, SMART sequence analysis tools, and NCBI Conserved 235
Domain Search server identified three domains in Pfa1: a Sec-dependent signal peptide formed by the 236
N-terminal 43 residues, a passenger domain (residues 209 to 553), and an autotransporter beta domain 237
(residues 970 to 1234). The passenger domain contains a putative protease (named Pap, for ‘Pfa1 protease’) 238
of the MEROPS peptidase family S8. Pap possesses a serine active site (residues 231 to 242) and an 239
aspartic acid active site (residues 505 to 515) typical for proteases of subtilase family. Structural analysis of 240
the beta domain using the AnthePro 5 software revealed the existence of ten membrane-spanning 241
antiparallel β-sheets corresponding to those that form the outer membrane β barrels of autotransporters. 242
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
The beta domain of Pfa1 terminates with the sequence of NAGFTFLF, which conforms to the consensus 243
sequence motif, (Y/V/I/F/W)-X-(F/W), discovered in most autotransporters. Taken together, these 244
structural features identify Pfa1 as likely a novel protein of the autotransporter family. 245
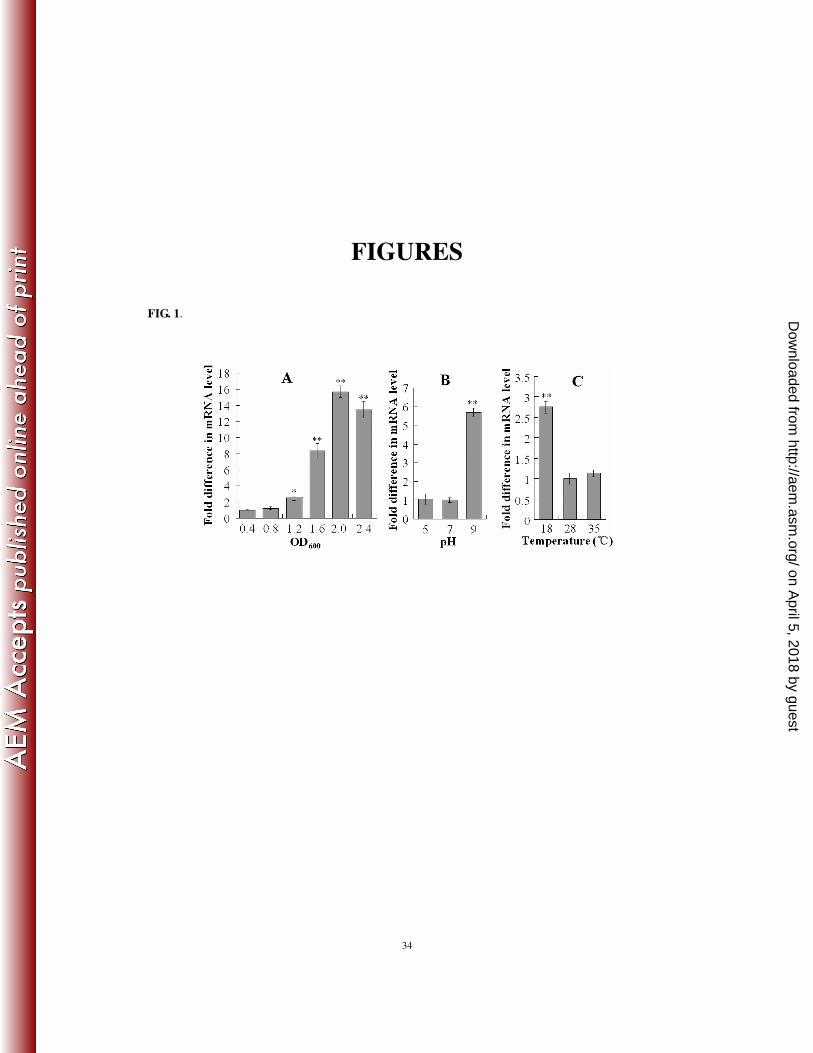
Expression of pfa1 is modulated by growth phase and growth conditions. Since Pfa1 appeared 246
among the antigenic proteins whose productions were heightened during infection, we determined the 247
expression levels of pfa1 under in vivo (i.e., during infection) and in vitro (i.e., cultured in LB medium at 248
280C) conditions. For this purpose, Japanese flounder were infected with TSS, and pfa1 expression in the 249
liver of the infected fish was determined by qRT-PCR. The results showed that pfa1 expression in vivo was 250
13-fold higher than that in vitro. We next examined the potential effects of other growth conditions, i.e., pH 251
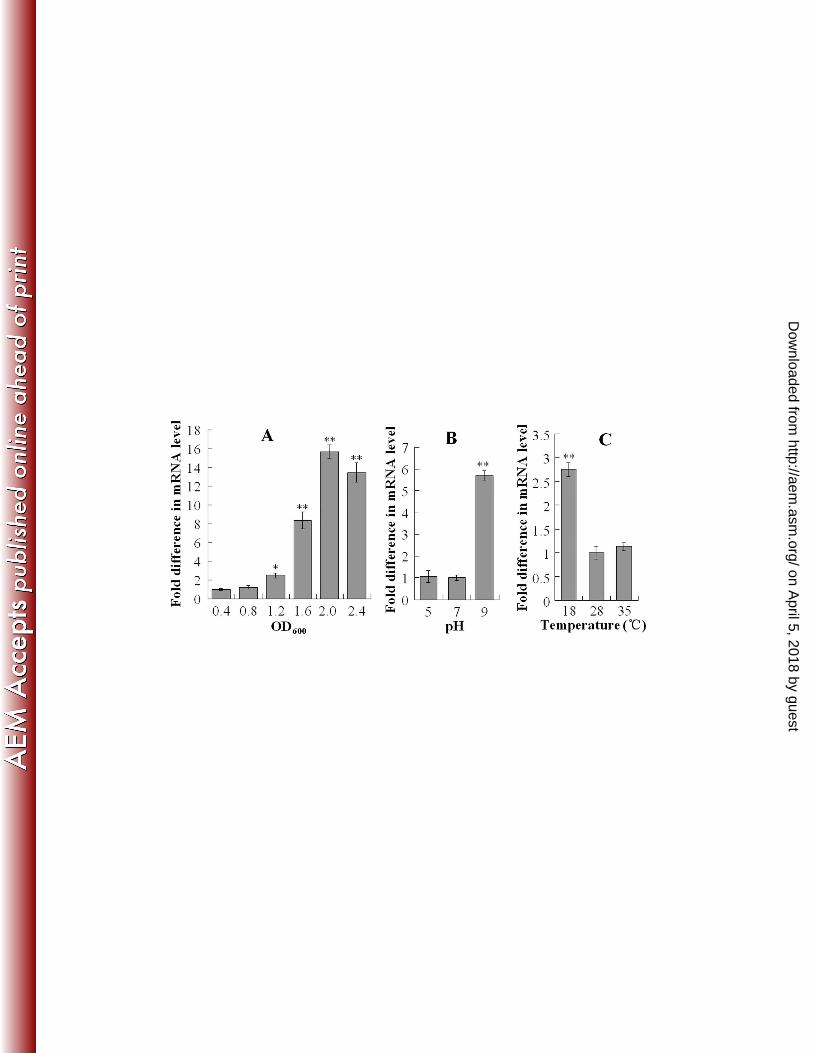
and temperature, and growth phase on pfa1 expression by qRT-PCR. The results showed that pfa1 252
expression increased with cell density and reached maximum at an OD600 of 2 (Fig. 1A). High pH (pH 9) 253
increased the expression of pfa1 to the level of 5.7-fold of that of the pfa1 expression at pH 7, whereas low 254
pH (pH 5) had no effect on pfa1 expression (Fig. 1B). Compared to pfa1 expression at 280C, low 255
temperature (180C) significantly enhanced pfa1 expression, whereas sub-lethal high temperature (35
0C) 256
had no significant effect on pfa1 expression (Fig. 1C). 257
Construction and characterization of a pfa1 null mutant. To investigate the functional importance 258
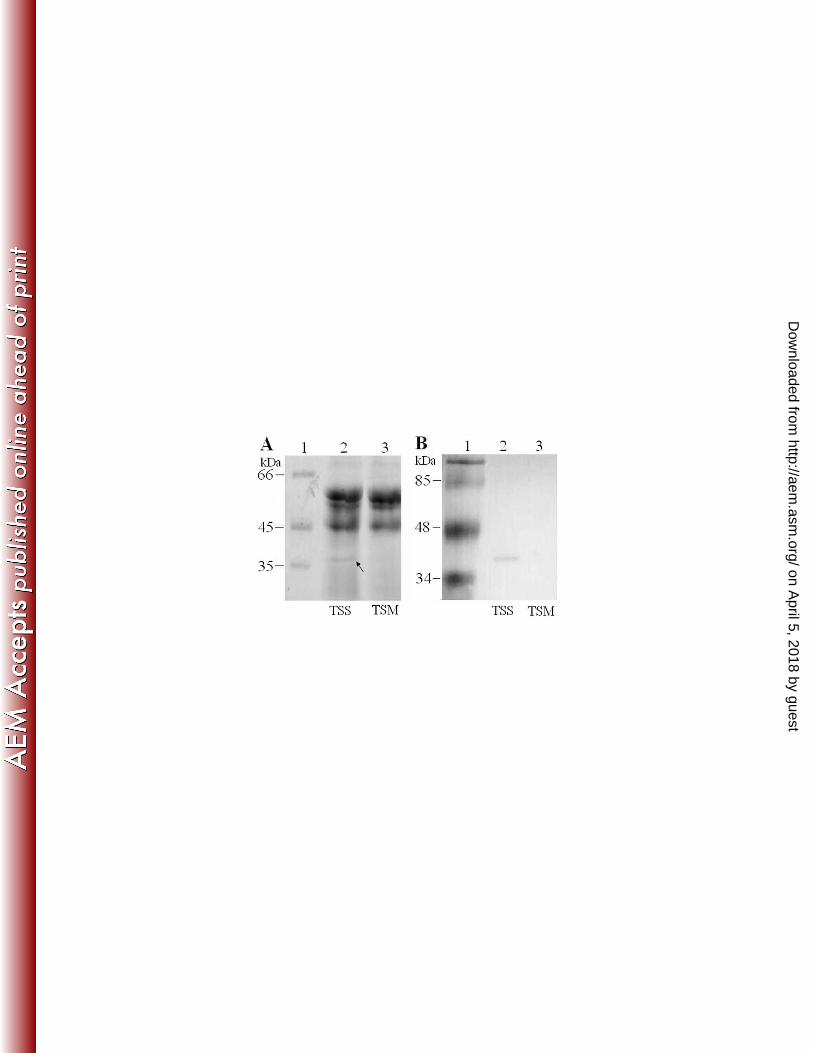
of pfa1, the gene was mutated by markerless in-frame deletion. SDS-PAGE analysis of the supernatant 259
proteins of TSS and the pfa1 null mutant, TSM, showed that they differed mainly in one protein, which 260
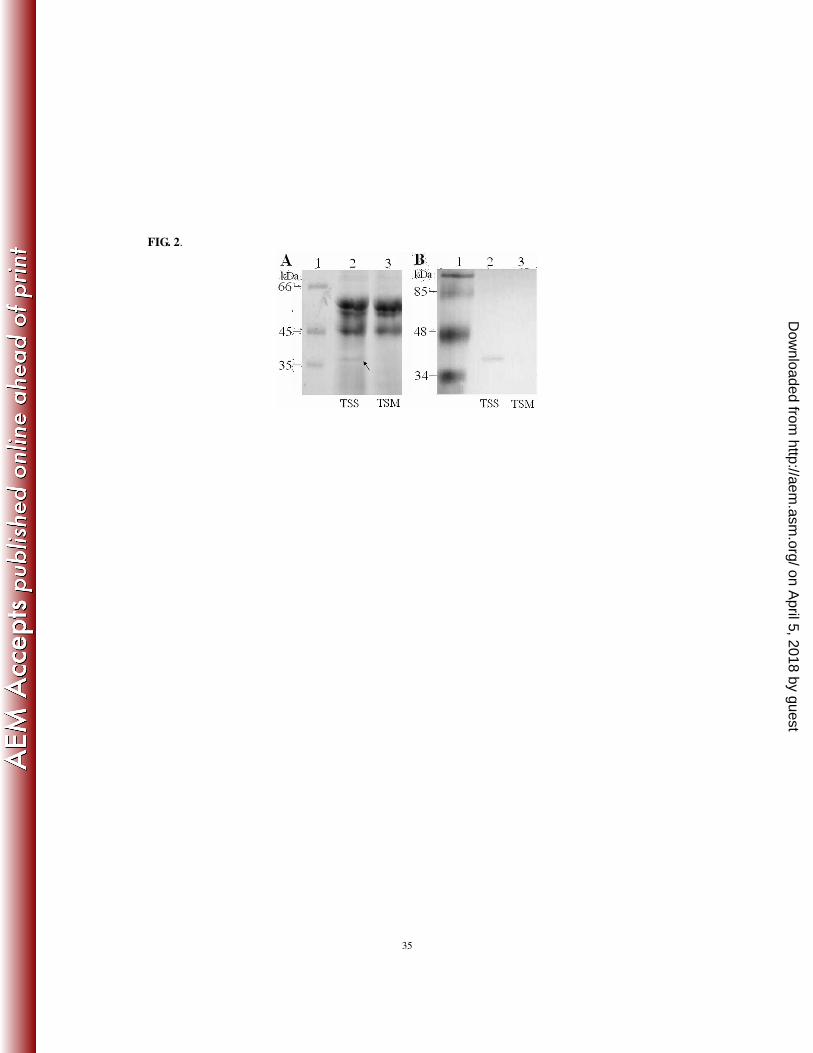
appeared in the supernatant of TSS but was absent in that of TSM (Fig. 2A). This TSS-unique protein 261
exhibits a molecular weight that matches the predicted molecular weight of Pap (37.6 kDa). Western 262
immunoblotting analysis showed that the 37.6 kDa protein could react with anti-Pap antibodies but not 263
with the preimmune serum (Fig. 2B and data not shown). Taken together, these results suggest that pfa1 264
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
very likely encodes an active autotransporter that can deliver its effector protease, Pap, out of the cell as an 265
extracellular protein. 266
Mutation of pfa1 has multiple effects - (i) effect on growth and biofilm production. Since TSS is a 267
fish pathogen, we compared TSS and TSM for the capacities that are known to be associated with 268
pathogenicity, which include biofilm formation, serum resistance, interaction with host cells, and 269
dissemination in host blood. Growth studies showed that the growth profile of TSM in nutrient medium 270
(LB) was similar to that of TSS, but the biofilm growth of TSM on polystyrene surface was significantly 271
lower (2.1-fold less) than that of TSS. 272
(ii) effect on serum resistance. For many pathogens, resistance against host serum killing constitutes 273
part of the virulence mechanism. TSS exhibits apparent serum resistance, as 69% of the cells survived after 274
incubation with Japanese flounder serum. In contrast, only 17% of TSM survived after the same treatment, 275
which is significantly lower than that of TSS. The presence of Mg2+
EGTA, which inhibits the classical 276
pathway of complement activation by removing Ca2+
, increased the survival rates of TSS and TSM to 84.8 277
and 72.5%, respectively. These results suggest that Pfa1 is required for blocking the activation of the 278
classical complement pathway. 279
(iii) effect on interaction with cultured host cells. To examine whether Pfa1 played any role in 280
interaction with host cells, cultured FG cells were incubated with TSS or TSM, and the bacterial cells 281
associated with the host cells were enumerated. The results showed that 47.4 and 12.5% of TSS and TSM, 282
respectively, were recovered from the entire (i.e., from the surface and the intracellular) FG cells, while 283
16.2 and 2% of TSS and TSM, respectively, were recovered from the intracellular of FG cells. Hence, TSM 284
is significantly impaired in the ability to adhere to and invade into FG cells. 285
(iv) effect on dissemination in blood and overall bacterial virulence. Blood dissemination analysis 286
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
showed that, following i.p. injection, the number of TSM recovered from the blood was 15-fold lower than 287
that of TSS. Consistently, i.p. injection of 3 x 107 CFU of TSS into Japanese flounder caused 90% 288
cumulative mortality in a period of two weeks, whereas i.p. injection of the same dose of TSM led to 30% 289
cumulative mortality, which is significantly (P < 0.01) lower than that caused by TSS injection. Injection 290
of PBS caused no mortality. These results demonstrate that TSM is significantly attenuated in overall 291
bacterial virulence. 292
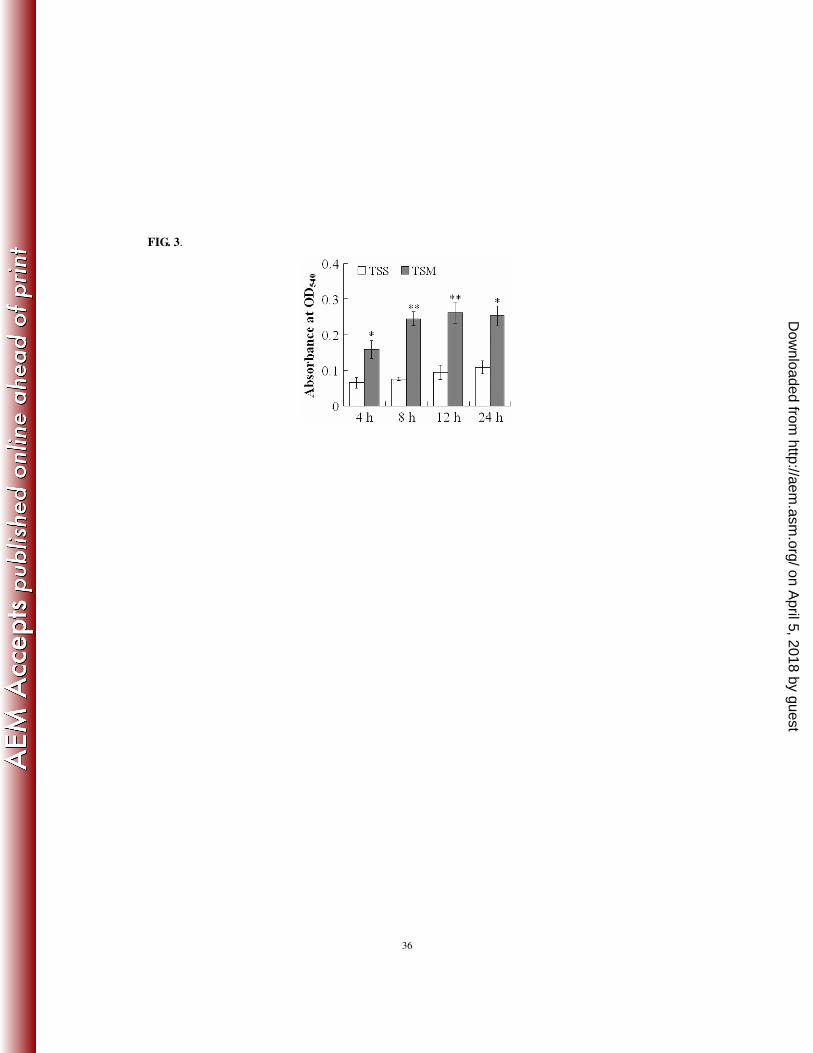
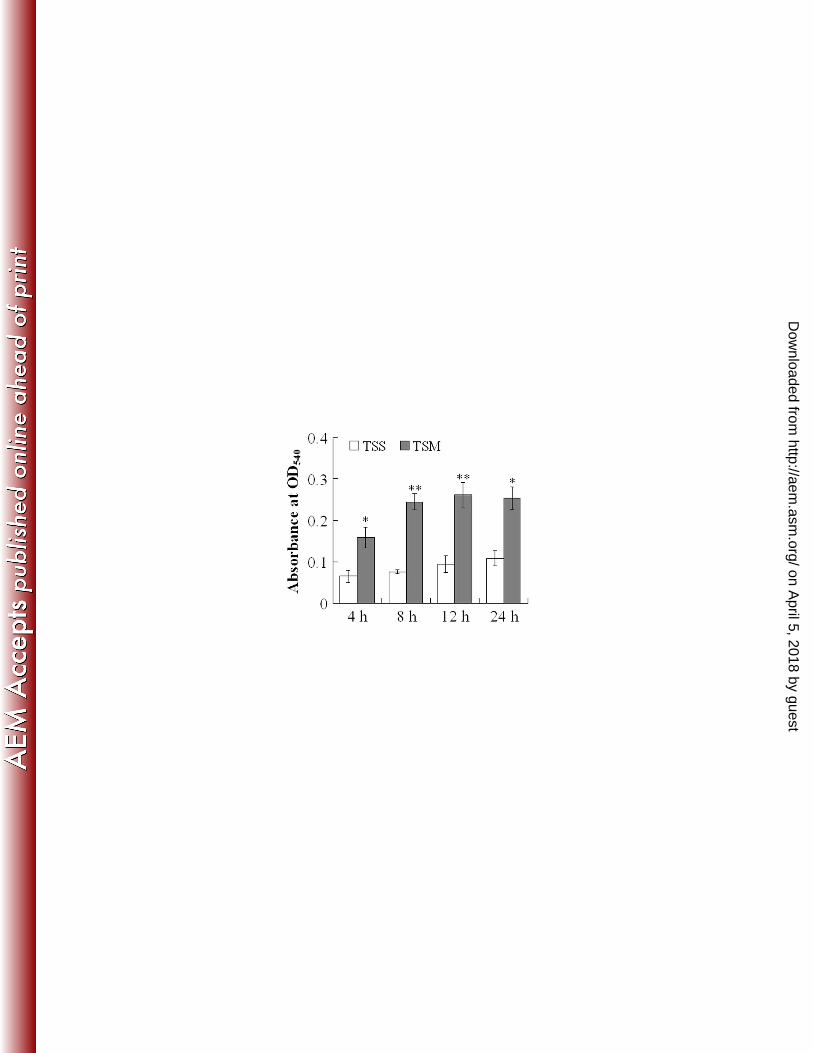
(v) effect on respiratory burst activity of the host phagocytes. Since, as demonstrated above, 293
TSM exhibited impaired infectivity compared to TSS, we examined whether TSM and TSS induced 294
different host immune response during infection. For this purpose, Japanese flounder were infected 295
separately with the same dose of TSS and TSM, and the respiratory burst activities of the neutrophils of the 296
infected fish were determined at 4, 8, 12, and 24 h post-infection. The results showed that, at all the 297
examined time points, the respiratory burst activities of the TSM-infected fish were significantly higher 298
than those of the TSS-infected fish (Fig. 3). 299
Analysis of the protease activity of the purified recombinant Pap. To examine the potential 300
protease activity of Pap, the coding sequences of the wild type and the mutant Pap (named PapM) bearing 301
an in-frame deletion of part of the serine active site (S507 to S514) were expressed in E. coli. Recombinant 302
Pap and PapM were purified and appeared as a single band of the expected molecular weight (~37 kDa) 303
following SDS-PAGE analysis (Supplemental data Figure S1). Protease activity analyses based on A350 304
absorbance showed that incubation of azocasein with 1 µl of Pap and PapM caused an absorbance increase 305
of 0.025 and 0.0004 respectively, over the control, which correspond to protease activities of 25 and 0.4 306
U/µl respectively. The protease activity of Pap was abolished in the presence of the serine protease inhibitor 307
phenylmethanesulfonyl fluoride. These results suggest that Pap is a serine protease whose activity requires 308
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
the integrity of the serine active site. 309
Effect of pH, temperature, and metal ions on the protease activity of recombinant Pap. 310
Enzymatic analyses showed that the optimal pH and temperature of recombinant Pap were 8.0 and 500C, 311
respectively. The protein exhibits more than 60% of the maximum activity over the pH range of 6 to 10 and 312
the temperature range of 30 to 800C (Fig. 4). Thermostability analysis showed that recombinant Pap was 313
stable over the temperature range of 10 to 800C and retained 76% of the maximum activity after incubation 314
at 800C for 1 h. The presence of Ca
2+ (10 mM) caused a 21% increase in the activity of Pap. Na
+, Mg
2+, and 315
Zn2+
had no apparent effect on the protease activity of Pap. 316
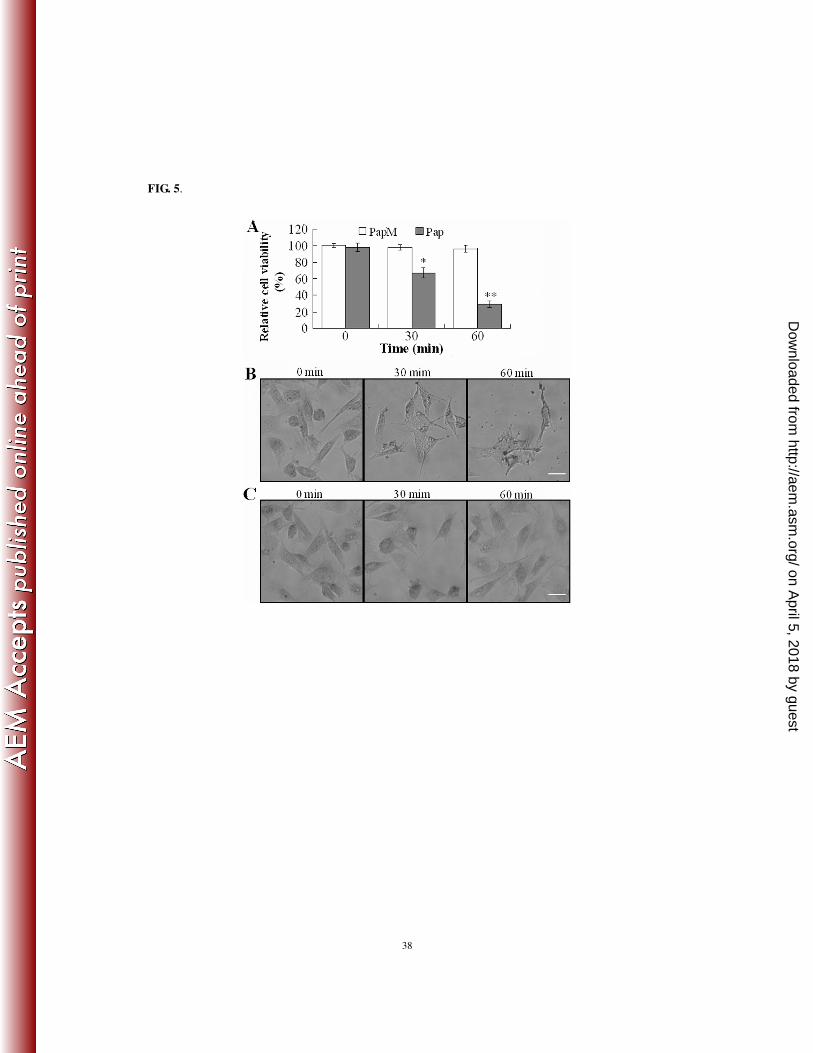
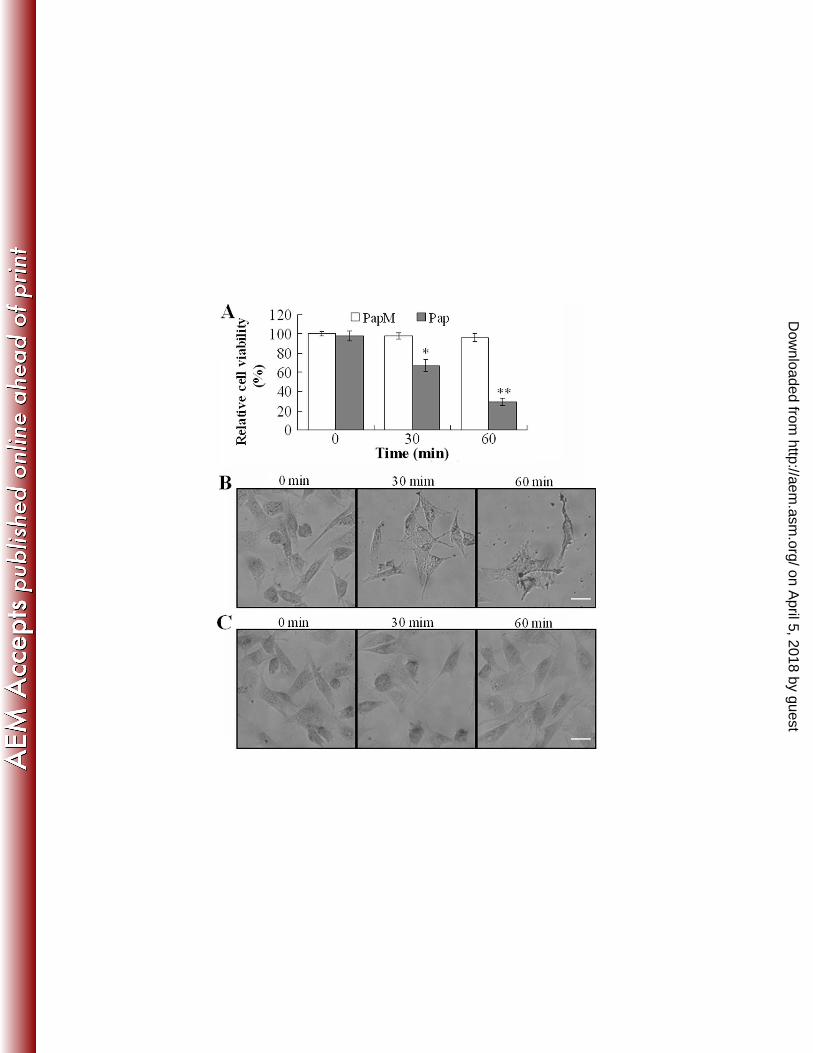
Cytotoxicity analysis of the purified recombinant Pap. Cytotoxicity analyses showed that the 317
presence of Pap significantly reduced the viability of FG cells in a time- and dose-dependent fashion, 318
whereas the presence of the proteolytically defective PapM had no apparent effect on FG cells (Fig. 5A and 319
data not shown). The number of viable FG cells decreased linearly after treatment with 0.5 to 4 µg of Pap. 320
Consistently, microscopic observation showed that incubation of FG cells with Pap had a profound 321
damaging effect on the cells, which occurred in a time-dependent manner, beginning with disruption of 322
cellular structures at 30 min after Pap treatment and progressing to complete cell lysis at 60 min after the 323
treatment (Fig. 5B). In contrast, FG cells treated with PapM or PBS exhibited no morphological changes 324
during the entire incubation period (Fig. 5C). Together these results demonstrate that purified recombinant 325
Pap can act as a cytotoxin on FG cells. 326
Identification of Pfa1 domains that are essential to autotransporting activity. To determine the 327
autotransporter regions that are required for translocator function in Pfa1, three Pfa1 derivatives, AT1, AT2, 328
and AT3, were constructed, each consisting of two segments of Pfa1: (i) the N-terminal 1-208 region; (ii) 329
the C-terminal 534-1242 (AT1) or 614-1242 (AT2) or 771-1242 (AT3) region. To analyze the potential 330
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
autotransporter activity of AT1, AT2, and AT3, we utilized a previously identified extracellular β agarase, 331
AgaV (60). Owing to its ability to degrade agar in LB medium, AgaV can cause the formation of visible 332
craters around AgaV-secreting bacterial colonies on LB agar plates. With this feature, AgaV is an ideal 333
reporter for the study of translocation properties of carrier proteins with secretion capacity. In this study, 334
AgaV devoid of the secretion domain was inserted, as a passenger, into AT1, AT2, and AT3 at immediately 335
downstream of the N-terminal 1-208 region (i.e., at the position where Pap occupies in Pfa1). The plasmids 336
pAT1V, pAT2V, and pAT3V, which express the coding elements of AgaV fused in-frame to AT1, AT2, and 337
AT3, respectively, were introduced into DH5α by transformation. The transformants were plated on LB 338
agar plates and examined for crater formation. Deep craters were formed around colonies of DH5α/pAT1V, 339
whereas no craters were formed around the colonies of DH5α/pAT2V or DH5α/pAT3V (Supplemental data 340
Figure S2). These results suggest that AT1, but not AT2 or AT3, can act as an effective autotransporter and 341
deliver the heterologous passenger AgaV out of the cell. 342
Application of AT1 as an antigen carrier. Since AT1 is an effective transporter of AgaV, we 343
wondered whether it (i.e., AT1) could be used in the transport and surface display of bacterial antigens. We 344
have previously identified an immunoprotective antigen, Et18 (17.8 kDa), from a virulent strain of 345
Edwardsiella tarda (14), which is an important aquaculture pathogen that can infect many cultured marine 346
species. Et18 is a periplasmic protein that, when used as a vaccine in the form of purified recombinant 347
protein, can afford certain protection (RPS, 61%) upon fish against E. tarda infection (14). To examine 348
whether Et18 could be transported as a passenger by AT1, the mature and C-terminally His-tagged Et18 349
was inserted in-frame into AT1 between the N-terminal 1-208 region and the C-terminal autotransporter 350
domain. The plasmid pAT18, which expresses the coding element of the AT1-Et18 chimera, was 351
introduced into DH5α by transformation. The subcellular localization of AT1-Et18 in DH5α/pΑΤ18 was 352
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
determined by Western immunoblotting, which showed that the chimeric protein was detected in the 353
outermembrane (Fig. 6). Consistently, whole cell ELISA showed that DH5α/pΑΤ18 could react with 354
anti-His antibody (A450 absorbance was 4.5-fold higher than that of the control), whereas DH5α harboring 355
the control plasmid pAT1 could not (A450 absorbance was comparable to that of the control). 356
To examine the immunoprotective effect of DH5α/pAT18, Japanese flounder were vaccinated 357
separately with DH5α/pAT18, DH5α/pAT1, purified recombinant Et18 protein, and PBS. The fish were 358
challenged with the pathogenic E. tarda strain TX1 and monitored for mortality over a period of two weeks. 359
The results showed that the accumulated mortalities of the fish vaccinated with DH5α/pAT18, Et18, 360
DH5α/pAT1, and PBS were 16.7, 33.3, 86.7, and 90%, respectively. Hence, the protection efficacy, in 361
terms of RPS, of DH5α/pAT18 was 81%, which was significantly higher than that of the recombinant Et18 362
(63%). 363
364
DISCUSSION 365
366
As mentioned earlier, most of the autotransporters with known functions are found to be implicated in 367
bacterial pathogenicity (14). Since the effector molecules of autotransporters are either surface-exposed on 368
the outer membrane or secreted into the extracellular milieu, autotransporters contribute to virulence 369
mainly by direct interaction with host cells and host defense systems (2, 6, 10, 25, 42, 49, 53). In our study, 370
we found that Pfa1 is required for interactions with cultured host cells. Since Pap appears as a soluble 371
protein in TSS and recombinant Pap displayed clear cytotoxic effect on FG cells, Pap probably acts as a 372
cytotoxin, rather than an adhesin, during the course of TSS infection. This hypothesis agrees with the 373
observation that mutation of pfa1 had a more profound effect on invasion into than attachment to FG cells. 374
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

19
It is possible that Pap, being a protease, may function in the degradation of certain surface proteins of the 375
host cell or extracellular matrix components and thus facilitate bacterial invasion. Likewise, resistance 376
against serum bactericidal activity, which is an important innate immune response against bacterial 377
infection, mediated by Pfa1 could be due to proteolysis of some complement factors by Pap, though it is 378
also possible that Pap may inactivate the complement elements by negative interaction, as it is found in the 379
cases of some adhesin autotransporters (1, 26, 31, 37, 41). Like serum bactericidal activity, respiratory 380
burst is another key innate defense mechanism by which phagocytes destroy invaded pathogens. The 381
observation that Pfa1 is implicated in modulation of these immune responses and that mutation of pfa1 382
significantly reduced the infectivity of TSS indicates that Pfa1 is an essential virulence factor of TSS. In 383
line with this conclusion, pfa1 expression was augmented during infection and when the temperature was 384
at 180C, which approximates the in vivo temperature of the host animals (Japanese flounder) that are 385
maintained at 18 to 220C in most farms in north China. 386
In the autotransporter secretion system, the fate of the passenger, once being exported out of the cell, 387
varies among different autotransporters. In some cases, the secreted passengers undergo no further 388
processing, while in others the passengers are cleaved from the rest of the autotransporter domains as part 389
of the maturation process (7). In the case where the passenger is a protease, the cleavage can be catalyzed 390
by the passenger protease itself or by external peptidases of the host, as is found in the serine protease 391
autotransporters of Enterobacteriaceae (11, 34, 40, 44, 46). The cleaved passenger molecule may either 392
remain attached to the cell surface via noncovalent interactions or be released into the environment as a 393
soluble protein. In our study, we found that Pap, the passenger protease of Pfa1, is secreted into the culture 394
supernatant of TSS, which suggests that Pap must have gone through certain processing steps involving 395
proteolysis. Considering the facts that most autotransporter passengers with autocatalytic properties belong 396
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

20
to MEROPS peptidase families S6/S8 and that Pap is classified, by its structural features, as an S8 family 397
protease, it is possible that Pap may catalyze its own maturation with or without the facilitation of host cell 398
factors. In support of this hypothesis, Et18 delivered by AT1, which lacks Pap, remained surface-anchored. 399
Recent studies have indicated that protein secretion via the autotransporter pathway is not as 400
self-sufficient as it had been initially thought and that other cellular components, such as the outer 401
membrane protein Omp85 (30, 32, 43, 51, 57) and the periplasmic chaperone Skp (38, 51), that are 402
external to the autotransporter polypeptide are involved in the translocation process of many 403
autotransporters. Nevertheless, autotransporter is still the simplest form of protein secretion system and 404
perhaps the least host-specific. The simplicity of the autotransporter secretion system, together with the 405
modular structure of the protein, renders it serviceable in several ways, one of which is in the development 406
of surface display systems, in which the passenger domain of the autotransporter is replaced by a 407
heterologous protein that, as a surrogate of the native passenger, is translocated to cell surface by the 408
β-domain of the autotransporter. Autotransporter-based surface display, or autodisplay, has been applied to 409
the presentation on bacterial surface a wide range of molecules, including enzymes, antibody fragments, 410
peptides, and antigens (reviewed in 4, 15, 17). In this study, we found that, like the E. coli autotransporter 411
AIDA-I, which has been shown to be able to display various antigenic determinants (21, 23, 29, 36, 49), 412
the Pfa1 derivative, AT1, could translocate the heterologous antigen Et18, in the place of Pap, to cell 413
surface. Compared to Et18 in the form of purified recombinant protein, Et18 displayed by AT1 is a more 414
effective vaccine, which could be due to the possibilities that (i) compared to the soluble protein, Et18 415
covalently linked to the membrane-anchored AT1 is more stable; (ii) Et18, when presented on the cell 416
surface, may assume a higher-order structure that is more readily recognized by the host immune system; 417
(iii) the live bacterial carrier may function as an adjuvant and facilitate the induction or/and heighten the 418
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

21
intensity of the specific immune response induced by Et18. 419
420
421
ACKNOWLEDGEMENTS 422
423
This work was supported by the National Natural Science Foundation of China (NSFC) grant 424
40576071, the National Basic Research Program of China grant 2006CB101807, and the 863 High 425
Technology Project grant 2008AA092501. 426
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

22
REFERENCES
1. Ackermann, N., M. Tiller, G. Anding, A. Roggenkamp, and J. Heesemann. 2008. Contribution of
trimeric autotransporter C-terminal domains of oligomeric coiled-coil adhesin (Oca) family members
YadA, UspA1, EibA, and Hia to translocation of the YadA passenger domain and virulence of Yersinia
enterocolitica. J. Bacteriol. 190:5031–5043.
2. Alamuri, P., and H. L. Mobley. 2008. A novel autotransporter of uropathogenic Proteus mirabilis is
both a cytotoxin and an agglutinin. Mol. Microbiol. 68:997–1017.
3. Amend, D. F. 1981. Potency testing of fish vaccines. In: International Symposium on Fish Biologics:
Serodiagnostics and Vaccines. Developments in Biological Standardisation. 49:447–454.
4. Buddenborg, C., D. Daudel, S. Liebrecht, L. Greune, V. Humberg, and M. A. Schmidt. 2008.
Development of a tripartite vector system for live oral immunization using a gram-negative probiotic
carrier. Int. J. Med. Microbiol. 298:105–114.
5. Chen, Z., B. Peng, S. Wang, and X. Peng. 2004. Rapid screening of highly efficient vaccine
candidates by immunoproteomics. Proteomics. 4:3203–3213.
6. Conners, R., D. J. Hill, E. Borodina, C. Agnew, S. J. Daniell, N. M. Burton, R. B. Sessions, A. R.
Clarke, L. E. Catto, D. Lammie, T. Wess, R. L. Brady, and M. Virji. 2008. The Moraxella adhesin
UspA1 binds to its human CEACAM1 receptor by a deformable trimeric coiled-coil. EMBO J.
27:1779–1789.
7. Dautin, N., and H. D. Bernstein. 2007. Protein secretion in gram-negative bacteria via the
autotransporter pathway. Annu Rev Microbiol. 61:89–112.
8. Desvaux, M., N. J. Parham, and I. R. Henderson. 2004. Type V protein secretion: simplicity gone
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

23
awry? Curr. Issues Mol. Biol. 6:111–124.
9. Economou, A., P. J. Christie, R. C. Fernandez, T. Palmer, G. V. Plano, and A. P. Pugsley. 2006.
Secretion by numbers: protein traffic in prokaryotes. Mol. Microbiol. 62:308–319.
10. Felek, S., M. B. Lawrenz, and E. S. Krukonis. 2008. The Yersinia pestis autotransporter YapC
mediates host cell binding, autoaggregation and biofilm formation. Microbiology. 154:1802–812.
11. Fink, D. L., L. D. Cope, E. J. Hansen, and J. W. Geme III. 2001. The Hemophilus influenzae Hap
autotransporter is a chymotrypsin clan serine protease and undergoes autoproteolysis via an
intermolecular mechanism. J. Biol. Chem. 276:39492–39500.
12. Henderson, I R., F. Navarro-Garcia, M. Desvaux, R. C. Fernandez RC, and D. Ala’Aldeen. 2004.
Type V protein secretion pathway: the autotransporter story. Microbiol. Mol. Biol. Rev. 68:692–744.
13. Henderson, I. R., and J. P. Nataro. 2001. Virulence functions of autotransporter proteins. Infect.
Immun. 69:1231–1243.
14. Hou, J., W. Zhang, and L. Sun. 2008. Immunoprotective analysis of two Edwardsiella tarda antigens.
J. Gen. Appl. Microbiol. 55:57–61.
15. Jose, J. 2006. Autodisplay: efficient bacterial surface display of recombinant proteins. Appl. Microbiol.
Biotechnol. 69:607–614.
16. Jose, J., F. Jahnig, and T. F. Meyer. 1995. Common structural features of IgA1 protease-like outer
membrane protein autotransporters. Mol. Microbiol. 18:378–380.
17. Jose, J., and T. F. Meyer. 2007. The autodisplay story, from discovery to biotechnical and biomedical
applications. Microbiol. Mol. Biol. Rev. 71:600–619.
18. Kajava, A. V., and A. C. Steven. 2006. The turn of the screw: variations of the abundant β-solenoid
motif in passenger domains of type V secretory proteins. J. Struct. Biol. 155:306–315.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

24
19. Kawai, E., A. Idei, H. Kumura, K. Shimazaki, H. Akatsuka, and K. Omori. 1999. The
ABC-exporter genes involved in the lipase secretion are clustered with the genes for lipase, alkaline
protease, and serine protease homologues in Pseudomonas fluorescens no. 33. Biochim. Biophys. Acta
1446:377–382.
20. Kim, Y. R., S. E. Lee, C. M. Kim, S. Y. Kim, E. K. Shin, D. H. Shin, S. S. Chung, H. E. Choy, A.
Progulske-Fox, J. D. Hillman, M. Handfield, and J. H. Rhee JH. 2003. Characterization and
pathogenic significance of Vibrio vulnificus antigens preferentially expressed in septicemic patients.
Infect. Immun. 71:5461–5471.
21. Kjaergaard, K., H. Hasman, M. A. Schembri, and P. Klemm. 2002. Antigen 43-mediated
autotransporter display, a versatile bacterial cell surface presentation system. J. Bacteriol.
184:4197–4204.
22. Kolodziejek, A. M., D. J. Sinclair, K. S. Seo, D. R. Schnider, C. F. Deobald, H. N. Rohde, A. K.
Viall, S. S. Minnich, C. J. Hovde, S. A. Minnich, and G. A. Bohach. 2007. Phenotypic
characterization of OmpX, an Ail homologue of Yersinia pestis KIM. Microbiology. 153:2941–2951.
23. Konieczny, M. P., M. Suhr, A. Noll, I. B. Autenrieth, and M. A. Schmidt. 2000. Cell surface
presentation of recombinant (poly-) peptides including functional T-cell epitopes by the AIDA
autotransporter system. FEMS Immunol. Med. Microbiol. 27:321–332.
24. Kumar, R., S. C. Mukherjee, R. Ranjan, and S. K. Nayak. 2008. Enhanced innate immune
parameters in Labeo rohita (Ham.) following oral administration of Bacillus subtilis. Fish Shellfish
Immunol. 24:168–72.
25. Lawrenz, M. B., J. D. Lenz, and V. L. Miller. 2008. A novel autotransporter adhesin is required for
efficient colonization during bubonic plague. Infect. Immun. 77:317–326.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

25
26. Leduc, I., B. Olsen, and C. Elkins. 2008. Localization of the domains of the Haemophilus ducreyi
trimeric autotransporter DsrA involved in serum resistance and binding to the extracellular matrix
proteins fibronectin and vitronectin. Infect. Immun. 77:657–666.
27. Loveless, B. J., and M. H. Saier, Jr. 1997. A novel family of channel forming, autotransporting,
bacterial virulence factors. Mol. Membr. Biol. 14:113–123.
28. Ma, Q., Y. Zhai, J. C. Schneider, T. M. Ramseier, and M. H. Saier. 2003. Protein secretion systems
of Pseudomonas aeruginosa and P. fluorescens. Biochimica et Biophysica Acta 1611: 223–233.
29. Maurer, J., J. Jose, and T. F. Meyer. 1997. Autodisplay: one-component system for efficient surface
display and release of soluble recombinant proteins from Escherichia coli. J. Bacteriol. 179:794–804.
30. Meng, G., N. K. Surana, J. W. St. Geme, and G. Waksman. 2006. Structure of the outer membrane
translocator domain of the Haemophilus influenzae Hia trimeric autotransporter. EMBO J.
25:2297–2304.
31. Nordstrom, T., A. M. Blom, A. Forsgren, and K. Riesbeck. 2004. The emerging pathogen Moraxella
catarrhalis interacts with complement inhibitor C4b binding protein through ubiquitous surface
proteins A1 and A2. J. Immunol. 173:4598–4606.
32. Oomen, C. J., P. van Ulsen, P. van Gelder, M. Feijen,J. Tommassen, and P. Gros. 2004. Structure
of the translocator domain of a bacterial autotransporter. EMBO J. 23:1257–1266.
33. Pallen, M. J., R. R. Chaudhuri, and I. R. Henderson. 2003. Genomic analysis of secretion systems.
curr. Opin. Microbiol. 6:519-527.
34. Patel, S. K., J. Dotson, K. P. Allen, and J. M. Fleckenstein. 2004. Identification and molecular
characterization of EatA, an autotransporter protein of enterotoxigenic Escherichia coli. Infect. Immun.
72:1786–1794.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

26
35. Pohlner, J., R. Halter, K. Beyreuther, and T. F. Meyer. 1987. Gene structure and extracellular
secretion of Neisseria gonorrhoeae IgA protease. Nature 325:458–462.
36. Rizos, K., C. T. Lattemann, D. Bumann, T. F. Meyer, and T. Aebischer. 2003. Autodisplay:
efficacious surface exposure of antigenic UreA fragments from Helicobacter pylori in Salmonella
vaccine strains. Infect. Immun. 71:6320–6328.
37. Roggenkamp, A., N. Ackermann, C. A. Jacobi, K. Truelzsch, H. Hoffmann, and J. Heesemann.
2003. Molecular analysis of transport and oligomerization of the Yersinia enterocolitica adhesin YadA.
J. Bacteriol. 185:3735–3744.
38. Qu, J., C. Mayer, S. Behrens, O. Holst, and J. H. Kleinschmidt. 2007. The trimeric periplasmic
chaperone Skp of Escherichia coli forms 1:1 complexes with outer membrane proteins via hydrophobic
and electrostatic interactions. Mol. Biol. 374:91–105.
39. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a laboratory manual, 2nd ed.
Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
40. Serruto, D., J. Adu-Bobie, M. Scarselli, D. Veggi, M. Pizza, R. Rappuoli, and B. Arico. 2003.
Neisseria meningitidis App: a new adhesin with autocatalytic serine protease activity. Mol. Microbiol.
48:323–334.
41. Sjolinder, H., J. Eriksson, L. Maudsdotter, H. Aro, and A. B. Jonsson. 2008. Meningococcal outer
membrane protein NhhA is essential for colonization and disease by preventing phagocytosis and
complement attack. Infect. Immun. 76:5412–5420.
42. Sheets, A. J., S. A. Grass, S. E. Miller, and J. W. St Geme III. 2008. Identification of a novel trimeric
autotransporter adhesin in the cryptic genospecies of Haemophilus. J. Bacteriol. 190:4313–4320.
43. Skillman, K. M., T. J. Barnard, J. H. Peterson, R. Ghirlando, and H. D. Bernstein. 2005. Efficient
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

27
secretion of a folded protein domain by a monomeric bacterial autotransporter. Mol. Microbiol.
58:945–958.
44. Stein, M., B. Kenny, M. A. Stein, and B. B. Finlay. 1996. Characterization of EspC, a 110-kilodalton
protein secreted by enteropathogenic Escherichia coli which is homologous to members of the
immunoglobulin A protease-like family of secreted proteins. J. Bacteriol. 178:6546–6554.
45. Sun, K., W. Zhang, J. Hou, and L. Sun. 2009. Immunoprotective analysis of VhhP2, a Vibrio harveyi
vaccine candidate. Vaccine. In press. DOI:10.1016/j.vaccine.2009.03.012.
46. Szabady, R. L., J. H. Peterson, K. M. Skillman, and H. D. Bernstein HD. 2005. An unusual signal
peptide facilitates late steps in the biogenesis of a bacterial autotransporter. Proc. Natl. Acad. Sci. USA
102:221–226.
47. Tong, S., H. Li, and H. Z. Miao. 1997. The establishment and partial characterization of a continuous
fish cell line FG-9307 from the gill of flounder Paralichthys olivaceus. Aquaculture 156:327-333.
48. Valle, J., A. N. Mabbett, G. C. Ulett, A. Toledo-Arana, K. Wecker, M. Totsika, M. A. Schembri, J.
M. Ghigo, and C. Beloin. 2008. UpaG, a new member of the trimeric autotransporter family of
adhesins in uropathogenic Escherichia coli. J Bacteriol. 190:4147–4161.
49. Van Gerven, N., H. De Greve, J. P. Hernalsteens. N. Van Gerven, H. De Greve, and J. P.
Hernalsteens. 2008. Inactivated Salmonella expressing the receptor-binding domain of bacterial
adhesins elicit antibodies inhibiting hemagglutination. Vet. Microbiol. 131:369–375.
50. Voulhoux, R., M. P. Bos, J. Geurtsen, M. Mols, and J. Tommassen. 2003. Role of a highly
conserved bacterial protein in outer membrane protein assembly. Science. 299:262–265.
51. Wagner, J. K., J. E. Heindl, A. N. Gray, S. Jain, and M. B. Goldberg. 2008. Contribution of the
periplasmic chaperone Skp to efficient presentation of the autotransporter IcsA on the surface of
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

28
Shigella flexneri. J. Bacteriol. 191:815–821.
52. Wang, H., Y. Hu, W. Zhang W, and L. Sun. 2009. Construction of an attenuated Pseudomonas
fluorescens strain and evaluation of its potential as a cross-protective vaccine. Vaccine. In press. DOI:
10.1016/j.vaccine.2009.04.023.
53. Weiser, J. N., and E. C. Gotschlich. 1991. Outer membrane protein A (OmpA) contributes to serum
resistance and pathogenicity of Escherichia coli K-1. Infect. Immun. 59:2252–2258.
54. Wells, T. J., O. Sherlock, L. Rivas, A. Mahajan, S. A. Beatson, M. Torpdahl, R. I. Webb, L. P.
Allsopp, K. S. Gobius, D. L. Gally, and M. A. Schembri. 2008. EhaA is a novel autotransporter
protein of enterohemorrhagic Escherichia coli O157:H7 that contributes to adhesion and biofilm
formation. Environ. Microbiol. 10:589–604.
55. Wells, T. J., J. J. Tree, G. C. Ulett, and M. A. Schembri. 2007. Autotransporter proteins: novel
targets at the bacterial cell surface. FEMS Microbiol. Lett. 274: 163–172.
56. Wu, T., J. Malinverni, N. Ruiz, S. Kim, T. J. Silhavy, and D. Kahne. 2005. Identification of a
multicomponent complex required for outer membrane biogenesis in Escherichia coli. Cell
121:235–245.
57. Yen, M. R., C. R. Peabody, S. M. Partovi, Y. Zhai, Y. H. Tseng, and M. H. Saier. 2002.
Protein-translocating outer membrane porins of Gram-negative bacteria. Biochim. Biophys. Acta
1562:6–31.
58. Zhang, M., K. Sun, and L. Sun. 2008. Regulation of autoinducer 2 production and luxS expression in
a pathogenic Edwardsiella tarda strain. Microbiology 154:2060–2069.
59. Zhang, W., K. Sun, S. Cheng, and L. Sun. 2008. Characterization of DegQVh, a serine protease and a
protective immunogen from a pathogenic Vibrio harveyi strain. Appl. Environ. Microbiol.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

29
74:6254–6262.
60. Zhang, W., and L. Sun. 2007. Cloning, characterization and molecular application of a beta-agarase
gene from Vibrio sp. strain V134. Appl. Environ. Microbiol. 73:2825–2831.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

30
FIGURE LEGENDS
FIG. 1. qRT-PCR analysis of pfa1 expression in relation to growth phase (A) and growth conditions (B and
C). (A) qRT-PCR was performed with total RNA extracted from TSS cultured in LB medium to different
densities. (B) and (C) qRT-PCR analyses were performed with total RNA extracted from TSS grown in LB
medium of different pH (B) or from TSS grown in LB medium at different temperatures (C). In all panels,
pfa1 mRNA level was normalized to that of the 16S rRNA. Data are means for three assays and presented
as the means ± SE. ∗∗, P < 0.01; ∗, P < 0.05.
FIG. 2. SDS-PAGE (A) and Western immunoblotting (B) analysis of Pap in the supernatants of TSS and
TSM. (A) Equal amounts of TSS and TSM supernatants were analyzed by SDS-PAGE. The TSS-unique
protein is indicated by an arrow. (B) TSS and TSM supernatants were resolved by SDS-PAGE as in (A); the
proteins were transferred to a nitrocellulose membrane and blotted with anti-Pap antibodies.
FIG. 3. Respiratory burst activities of blood neutrophils of Japanese flounder after infection by TSS and
TSM. Two groups of Japanese flounder were infected with the same dose of TSS or TSM for various times.
Blood was collected from the infected fish at the indicated time points and used for respiratory burst
activity assay. Data are presented as the means ± SE. ∗∗, P < 0.01; ∗, P < 0.05.
FIG. 4. Effects of pH (A) and temperature (B) on the activity of purified recombinant Pap. (A) The effect of
pH was determined in three different buffers: 50 mM citric acid-sodium phosphate (pH 4 to 6; ■), 50 mM
sodium phosphate (pH 6 to 9; ◇), and 50 mM glycine-NaOH (pH 9 to 11; ▲). (B) The effect of
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

31
temperature (■) was determined in the assay buffer. Thermostability (□) was determined by preincubating
the enzyme in the assay buffer at the indicated temperature for 1 h before initiating enzymatic reaction by
the addition of azocasein. Data are means for three independent assays and presented as the means ± SE.
.
FIG. 5. Examination of the cytotoxic effect of purified recombinant Pap on cultured FG cells by cell
viability analysis (A) and microscopy (B). (A) FG cells were treated for 0, 30, and 60 min with the same
amount of recombinant Pap or PapM. Viabilities of the cells were determined by the MTT method. Data are
the means of three independent assays and presented as the means ± SE. *, P < 0.05; **, P <0.001. (B) and
(C) FG cells were treated with Pap (B) or PapM (C) as above. The cells were stained with Giemsa and
observed under an inverted microscope. Images were taken at 120 × magnification. Bar = 50 µm.
FIG. 6. Subcellular localization of the His-tagged AT1-Et18 chimera in DH5α/pAT18. Proteins of the
outermembrane (OM), cytoplasm (CP), periplasm (PP), and supernatant (SN) were prepared from
DH5α/pAT18 and analyzed by SDS-PAGE. As a negative control, outermembrane proteins of DH5α/pAT1
(lane 6) were also subjected to SDS-PAGE. After electrophoresis, the proteins were transferred to a
nitrocellulose membrane and blotted with anti-His antibody. NC, negative control. Lane 1, proteins
markers.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

32
TABLES
TABLE 1. Plasmids used in this study
Plasmid Relevant characteristicsa Source or reference
p7TS Tcr; suicide plasmid 52
pAT1 Apr; carrying Pfa1 autotransporter regions This study
pAT2 Apr; carrying Pfa1 autotransporter regions This study
pAT3 Apr; carrying Pfa1 autotransporter regions This study
pAT1V Apr; pAT1 carrying agaV This study
pAT2V Apr; pAT2 carrying agaV This study
pAT3V Apr; pAT3 carrying agaV This study
pAT1V Apr; pAT1 carrying agaV This study
pAT18 Apr; pAT1 carrying et18 This study
pB328 pBR322 with mutated ScaI site This study
pBR322 Apr, Tcr; general cloning vector Takara
pBSAT1 Apr; carrying the 5’ portion of pfa1 This study
pBS-T Apr, cloning vector Tiangen (Beijing, China)
pEPA1 Knr; carrying the coding sequence of Pap This study
pEPA2 Knr; carrying the coding sequence of PapM This study
pET258 Knr; expression plasmid 60
pJT Tcr; broad host range vector 45
aAp
r: ampicillin resistant; Kn
r: kanamycin resistant; Tc
r: tetracycline resistant.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from

33
TABLE 2. Primers used in this study
Primers Sequences (5’→ 3’)a
18F9 GATATCGGTGGTGGTGGTGGTGGTGGTGGTTGCG
TCGCCGCCGT (EcoRV)
18R11 GATATCGTGGTGGTGGTGGTGGTGCTTCAGCAGC
GAGAACG (EcoRV)
ApF8 AGATCACTGAATTCTTGAAGACGAAAGG (EcoRI)
ApMF1 CTTGGTTGAATACTCACCAGTCACAGAA
ApMR1 TGGTGAGTATTCAACCAAGTCATTCTGAG
ApR7 TATCTCGAGCCAATGCTTAATCAGTGA
F6 TAAGTAAGTAAGCCGGATATCGAAGGCA
F11 CGCGGCGTTAACATTTAAATTGATGACGCACATATCCA (SwaI)
F12 AGTACTACGCTGACCGGTGGC (ScaI)
F15 GCATATGTCCGGCCTGGACCG (NdeI)
MF2 GGCACCGGAGCTCTGGCGTTG
MR2 CAGAGCTCCGGTGCCGTTGGCATT
R9 CCCGGGATATCAGAGTACTATTGACCTGAGCTTCC (SmaI, EcoRV, ScaI)
R12 CAGTACTATTTAAATTAGAACAGGAAGGTAAACCCG (ScaI, SwaI)
R13 CTCGAGGCCAACCGGCGTGT (XhoI)
R14 GTACTATTTAAATAGCTGTTCTGGCCTTCCA (ScaI, SwaI)
RTF33 GCAACGCAATCGCACCC
RTR10 GCTGTCCTGAACGCCGATT
UAF13 CCCGGGAGAAGACTGGCGAGAAAT (SmaI)
UAR14 CCCGGGTTGTATAAATTTCAATCGCTGGT (SmaI)
aUnderlined nucleotides are restriction sites of the enzymes indicated in the brackets at the ends.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from
34
FIGURES
FIG. 1.
on April 5, 2018 by guest
http://aem.asm
.org/D
ownloaded from


















