How the Retina Works · The retina is a filmy piece of tissue, barely half a millimeter thick, that...
8
T he retina is a filmy piece of tissue, barely half a millimeter thick, that lines the inside of the eyeball. The tis- sue develops from a pouch of the em- bryonic forebrain, and the retina is therefore considered part of the brain. This most important part of the eye has a basic structure similar to that of a three-layer cake, with the bodies of nerve cells arrayed in three rows sepa- rated by two layers packed with synaptic connections. The retina in- cludes both the sensory neurons that respond to light and intricate neural circuits that perform the first stages of image processing; ultimately, an elec- trical message travels down the optic nerve into the brain for further pro- cessing and visual perception. Intuitively, one might expect that the surface of the retina (the layer ex- posed to the liquid in the eyeball’s vit- reous chamber) would contain the sensory cells, the photoreceptors, but actually these cells lie at the very back of the retina; light rays must pass through the entire retina before reach- ing pigment molecules to excite. This is because the pigment-bearing mem- branes of the photoreceptors have to be in contact with the eye’s pigment epithelial layer, which provides a steady stream of the vital molecule, retinal or vitamin A. Retinal becomes fixed in the photoreceptors’ opsin pro- teins, where this small molecule changes its conformation in response to photons, or packets of light. Once retinal molecules are exposed to light and undergo their conformational change, they are recycled back into the pigment epithelium. This tissue be- hind the retina is usually very dark because its cells are full of melanin granules. The pigment granules ab- sorb stray photons, preventing their reflection back into the photorecep- tors, which would cause images to blur. They also protect the cells from too much exposure to light radiation. Retina Design According to Lifestyle All vertebrate retinas contain at least two types of photoreceptors—the fa- miliar rods and cones. Rods are gener- ally used for low-light vision and cones for daylight, bright-colored vision. The variations among animal eyes reveal adaptations to the different environ- ments in which they live. Most fish, frog, turtle and bird retinas have three to five types of cones and consequently very good color vision. Keep in mind that reptiles and fish are “cold blood- ed” and need to be active in the warm daytime. Most mammals have retinas in which rods predominate. When the number of mammals started to ex- plode as the dinosaurs died out, the Earth was likely a dark place covered in ash and clouds; the tiny, fur-covered early mammals were able to generate their own body heat and developed visual systems sensitive to dim light. Modern rodents such as rats and mice, which are nocturnal animals, still have retinas overwhelmingly dominated by rods; their cones are small in size and only make up 3 to 5 percent of their photoreceptors. Most other mammalian retinas also have a preponderance of rods, and the cones are often concentrated in special- ized regions. In species such as cats and dogs, images focus to a central specialized area, aptly called the area centralis, where cones predominate. The retinas of mammals such as rab- bits and squirrels, as well as those of nonmammals like turtles, have a long, horizontal strip of specialized cells called a visual streak, which can detect the fast movement of predators. Pri- mates as well as some birds have front- projecting eyes allowing binocular vi- sion and thus depth perception; their eyes are specialized for good daylight vision and are able to discriminate col- or and fine details. Primates and rap- tors, like eagles and hawks, have a fovea, a tremendously cone-rich spot devoid of rods where images focus. Primates, in fact, have what is called a duplex retina, allowing good visual discrimination in all lighting condi- tions. The fovea contains most of the cones, packed together as tightly as physically possible, and allows good daylight vision. More peripheral parts of the retina can detect the slightest glimmer of photons at night. Most mammals have two types of cones, green-sensitive and blue-sensitive, but primates have three types—red-sensi- tive as well as the other two. With our cone vision, we can see from gray dawn to the dazzling conditions of high noon with the sun burning down on white sand. Initially the cone photo- receptors themselves can adapt to the surrounding brightness, and circuitry through the retina can further modu- late the eye’s response. Similarly, the rod photoreceptors and the neural cir- cuitry to which they connect can adapt to lower and lower intensity of light. 28 American Scientist, Volume 91 How the Retina Works Much of the construction of an image takes place in the retina itself through the use of specialized neural circuits Helga Kolb © 2003 Sigma Xi, The Scientific Research Society. Reproduction with permission only. Contact [email protected]. Helga Kolb is professor emeritus of ophthalmology and visual sciences at the University of Utah. She studied at the University of Bristol, completing her Ph.D. in 1971. She conducted eye research at the Institute of Ophthalmology in London, where she became involved in studies of electrophysiology and anatomy, and later moved to positions at Johns Hopkins and the National Institutes of Health. She joined the Utah faculty in 1979. Address: Univer- sity of Utah Health Sciences Center, 75 North Medical Drive, Salt Lake City, Utah 84132. Inter- net: [email protected]
-
Upload
nguyentram -
Category
Documents
-
view
214 -
download
0
Transcript of How the Retina Works · The retina is a filmy piece of tissue, barely half a millimeter thick, that...

The retina is a filmy piece of tissue,barely half a millimeter thick, that
lines the inside of the eyeball. The tis-sue develops from a pouch of the em-bryonic forebrain, and the retina istherefore considered part of the brain.This most important part of the eyehas a basic structure similar to that of athree-layer cake, with the bodies ofnerve cells arrayed in three rows sepa-rated by two layers packed withsynaptic connections. The retina in-cludes both the sensory neurons thatrespond to light and intricate neuralcircuits that perform the first stages ofimage processing; ultimately, an elec-trical message travels down the opticnerve into the brain for further pro-cessing and visual perception.
Intuitively, one might expect thatthe surface of the retina (the layer ex-posed to the liquid in the eyeball’s vit-reous chamber) would contain thesensory cells, the photoreceptors, butactually these cells lie at the very backof the retina; light rays must passthrough the entire retina before reach-ing pigment molecules to excite. Thisis because the pigment-bearing mem-branes of the photoreceptors have tobe in contact with the eye’s pigmentepithelial layer, which provides asteady stream of the vital molecule,retinal or vitamin A. Retinal becomes
fixed in the photoreceptors’ opsin pro-teins, where this small moleculechanges its conformation in responseto photons, or packets of light. Onceretinal molecules are exposed to lightand undergo their conformationalchange, they are recycled back into thepigment epithelium. This tissue be-hind the retina is usually very darkbecause its cells are full of melaningranules. The pigment granules ab-sorb stray photons, preventing theirreflection back into the photorecep-tors, which would cause images toblur. They also protect the cells fromtoo much exposure to light radiation.
Retina Design According to Lifestyle All vertebrate retinas contain at leasttwo types of photoreceptors—the fa-miliar rods and cones. Rods are gener-ally used for low-light vision and conesfor daylight, bright-colored vision. Thevariations among animal eyes revealadaptations to the different environ-ments in which they live. Most fish,frog, turtle and bird retinas have threeto five types of cones and consequentlyvery good color vision. Keep in mindthat reptiles and fish are “cold blood-ed” and need to be active in the warmdaytime. Most mammals have retinasin which rods predominate. When thenumber of mammals started to ex-plode as the dinosaurs died out, theEarth was likely a dark place coveredin ash and clouds; the tiny, fur-coveredearly mammals were able to generatetheir own body heat and developedvisual systems sensitive to dim light.Modern rodents such as rats and mice,which are nocturnal animals, still haveretinas overwhelmingly dominated byrods; their cones are small in size andonly make up 3 to 5 percent of theirphotoreceptors.
Most other mammalian retinas alsohave a preponderance of rods, and thecones are often concentrated in special-ized regions. In species such as catsand dogs, images focus to a centralspecialized area, aptly called the areacentralis, where cones predominate.The retinas of mammals such as rab-bits and squirrels, as well as those ofnonmammals like turtles, have a long,horizontal strip of specialized cellscalled a visual streak, which can detectthe fast movement of predators. Pri-mates as well as some birds have front-projecting eyes allowing binocular vi-sion and thus depth perception; theireyes are specialized for good daylightvision and are able to discriminate col-or and fine details. Primates and rap-tors, like eagles and hawks, have afovea, a tremendously cone-rich spotdevoid of rods where images focus.
Primates, in fact, have what is calleda duplex retina, allowing good visualdiscrimination in all lighting condi-tions. The fovea contains most of thecones, packed together as tightly asphysically possible, and allows gooddaylight vision. More peripheral partsof the retina can detect the slightestglimmer of photons at night. Mostmammals have two types of cones,green-sensitive and blue-sensitive, butprimates have three types—red-sensi-tive as well as the other two. With ourcone vision, we can see from graydawn to the dazzling conditions ofhigh noon with the sun burning downon white sand. Initially the cone photo-receptors themselves can adapt to thesurrounding brightness, and circuitrythrough the retina can further modu-late the eye’s response. Similarly, therod photoreceptors and the neural cir-cuitry to which they connect can adaptto lower and lower intensity of light.
28 American Scientist, Volume 91
How the Retina Works
Much of the construction of an image takes place in the retina itself through the use of specialized neural circuits
Helga Kolb
© 2003 Sigma Xi, The Scientific Research Society.Reproduction with permission only. Contact [email protected].
Helga Kolb is professor emeritus of ophthalmologyand visual sciences at the University of Utah. Shestudied at the University of Bristol, completing herPh.D. in 1971. She conducted eye research at theInstitute of Ophthalmology in London, where shebecame involved in studies of electrophysiology andanatomy, and later moved to positions at JohnsHopkins and the National Institutes of Health. Shejoined the Utah faculty in 1979. Address: Univer-sity of Utah Health Sciences Center, 75 NorthMedical Drive, Salt Lake City, Utah 84132. Inter-net: [email protected]
Anatomy and Physiology Understanding the anatomy of the pri-mate retina is essential to understand-ing its function. Again, the photore-ceptors lie in a layer against the backof the eyeball. In the second of threecell layers, called the inner nuclear lay-er, lie one to four types of horizontalcells, 11 types of bipolar cells and 22 to30 types of amacrine cells. The numbersvary depending on species. The sur-face layer of the retina contains about20 types of ganglion cells. Impulsesfrom the ganglion cells travel to thebrain via more than a million opticnerve fibers. The spaces separatingthese three layers are also anatomicallydistinct. The region containing synaps-es linking the photoreceptors withbipolar and horizontal cell dendrites isknown as the outer plexiform layer; thearea where the bipolar and amacrinecells connect to the ganglion cells is theinner plexiform layer.
Decades of anatomical studies haveshed light on how the retina works.
Imaging techniques ranging from old-style Golgi silver staining, first usedover a century ago by Ramón y Cajal,to electron microscopy and modern-day antibody staining have revealedthe shapes and sizes of the retina’s celltypes and how the different cells con-nect to form synapses. Staining tech-niques have revealed electrical junc-tions between cells and the identityand location of neurotransmitter recep-tors and transporters. We now knowthat the neurotransmitter (chemicalsignal) passed through the verticalpathways of the retina—from photore-ceptors to bipolar cells to ganglioncells—is glutamate. The horizontal andamacrine cells send signals using vari-ous excitatory and inhibitory aminoacids, catecholamines, peptides and ni-tric oxide.
Electrophysiological investigationsof the retina started 60 years ago. Stud-ies of the optic nerve fibers showed thatthey could be stimulated to give tradi-tional depolarizing action potentials,
like those observed in other neurons.However, the first recordings of im-pulses within the retina by GunnarSvaetichin in the 1950s showed veryodd responses to light. Neurons in theouter retina—it was not immediatelyclear which cells he was recordingfrom—responded to stimulation notwith depolarizing spikes but with slowhyperpolarization. These “S potentials”are now known to originate with thephotoreceptors and to be transmitted tohorizontal cells and bipolar cells. Themembrane hyperpolarization starts onexposure to light, follows the timecourse of a light flash and then returnsto the baseline value when the light isoff. This reflects the counterintuitivefact that both rods and cones releaseneurotransmitters during the dark,when the membrane is depolarizedand sodium ions flow freely across thephotoreceptors’ cell membranes. Whenexposed to light, ion channels in thecell membranes close. The cells go intoa hyperpolarized state for as long as
2003 January–February 29
© 2003 Sigma Xi, The Scientific Research Society. Reproductionwith permission only. Contact [email protected].
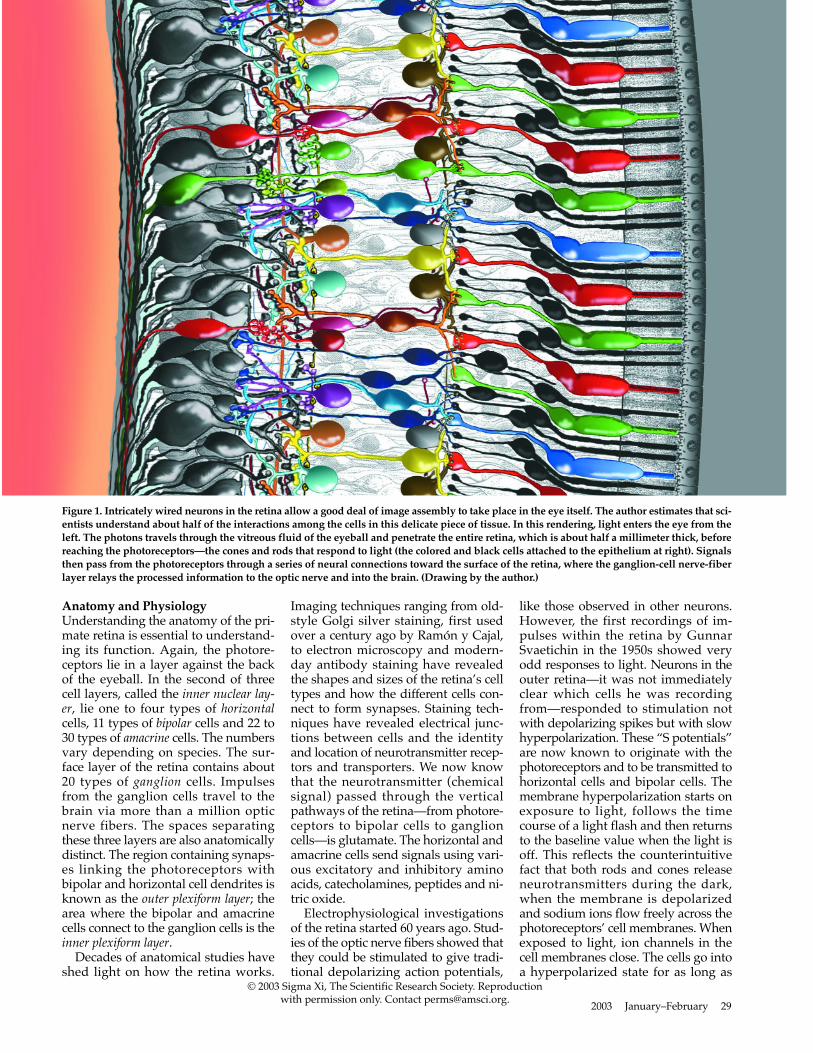
Figure 1. Intricately wired neurons in the retina allow a good deal of image assembly to take place in the eye itself. The author estimates that sci-entists understand about half of the interactions among the cells in this delicate piece of tissue. In this rendering, light enters the eye from theleft. The photons travels through the vitreous fluid of the eyeball and penetrate the entire retina, which is about half a millimeter thick, beforereaching the photoreceptors—the cones and rods that respond to light (the colored and black cells attached to the epithelium at right). Signalsthen pass from the photoreceptors through a series of neural connections toward the surface of the retina, where the ganglion-cell nerve-fiberlayer relays the processed information to the optic nerve and into the brain. (Drawing by the author.)
the light continues to shine on themand do not release a neurotransmitter.
Although both rods and cones re-spond to light with a slow hyperpolar-izing response, they report quite differ-ent image properties. Rods, detectingdim light, usually respond to relativelyslow changes. Cones, dealing withbright signals, can detect rapid lightfluctuations. In both cases, photorecep-tors begin the process of decomposing
images into separate parts. Both rodsand cones respond to light directlyover them. Thus, their receptive fieldsare very narrow.
An image continues to be brokeninto component elements at the firstsynapses of the visual pathway, thosebetween photoreceptors and bipolarcells. Different bipolar cells have differ-ent types of receptors for the neuro-transmitter glutamate, allowing the
cells to respond to photoreceptor inputdifferently (Figure 7). Some bipolar cellsare tuned to faster and some to slowerfluctuations in the visual signal; someglutamate receptors resensitize rapidlyand others more gradually. The cellsthus fire either quickly in succession orrelatively slowly in response to thesame amount of stimulation. These re-ceptors respond to glutamate by acti-vating what’s known as an OFF path-way in the visual process, detectingdark images against a lighter back-ground. (Recall that photoreceptorsconstantly release glutamate unless ex-posed to light.) Other bipolar cellshave inhibitory glutamate receptors;in other words, they prevent the bipo-lar cell from firing when the cell is ex-posed to the neurotransmitter. Thesereceptors activate the ON pathway, de-tecting light images against a darkerbackground.
Parallel ProcessingThe parallel sets of visual channels forON (detecting light areas on darkbackgrounds) and OFF (detecting darkareas on light backgrounds) qualitiesof an image are fundamental to ourseeing. Vertebrate vision depends onperceiving the contrast between im-ages and their backgrounds. For ex-ample, we read black letters against awhite background using the OFF chan-nels that start in the retina. Parallelbipolar channels transmit inputs toganglion cells. Early in developmentthe architecture of the inner plexiformlayer, full of synapses between bipolarand ganglion cells, shows that synapticconnections become segregated in dis-tinct, parallel pathways. Connectionsoccur between ON bipolar cells andON ganglion cells and also betweenOFF bipolar cells and OFF ganglioncells in demarcated portions of the in-ner plexiform layer.
If the retina were simply to transmitopposite-contrast images directly fromthe photoreceptors to the brain, the re-sulting vision would probably becoarse-grained and blurry. Further pro-cessing in the retina defines preciseedges to images and allows us to focuson fine details. The honing of the im-age starts at the first synaptic level inthe retina, where horizontal cells re-ceive input from cones. Each horizontalcell actually receives input from manycones, so its collection area or receptivefield is large. Horizontal cells’ receptivefields become even broader because
30 American Scientist, Volume 91
© 2003 Sigma Xi, The Scientific Research Society. Reproductionwith permission only. Contact [email protected].
sclera choroid
retinacornea
pupil
lens
iris
ciliarybody
opticnerve
ganglion cells
photoreceptors
outer plexiform
inner nuclear
inner plexiform
pigment epithelium
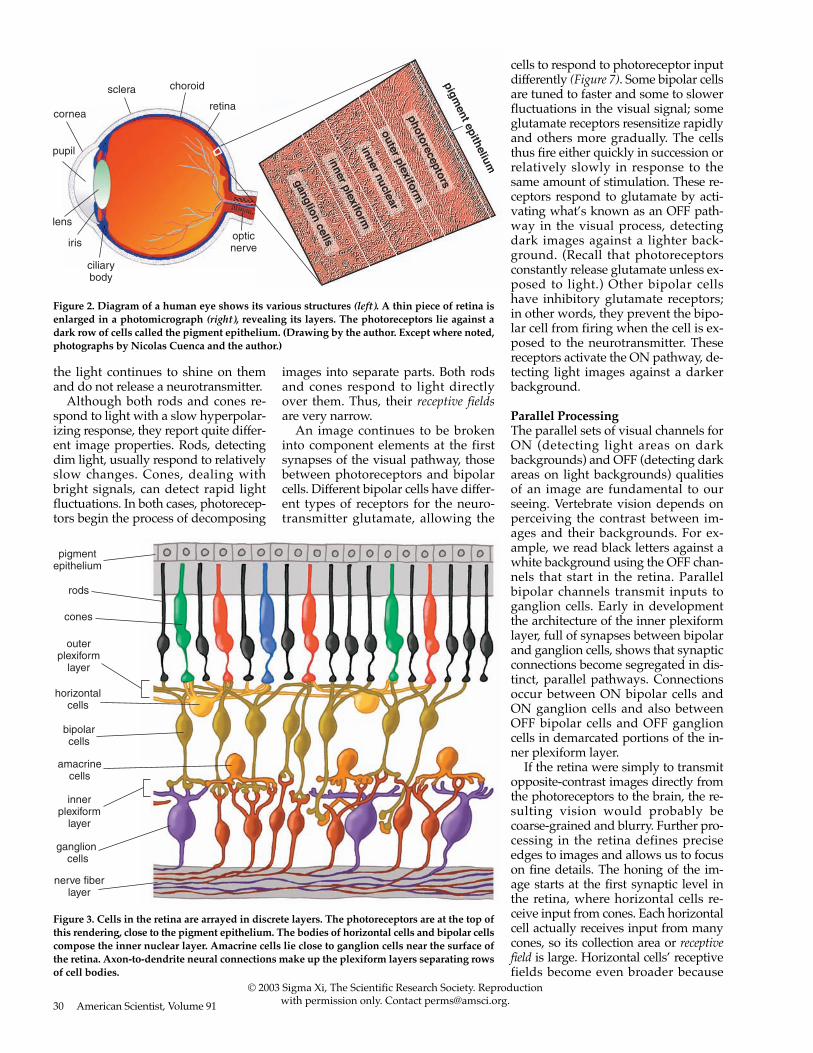
Figure 2. Diagram of a human eye shows its various structures (left). A thin piece of retina isenlarged in a photomicrograph (right), revealing its layers. The photoreceptors lie against adark row of cells called the pigment epithelium. (Drawing by the author. Except where noted,photographs by Nicolas Cuenca and the author.)
pigmentepithelium
rods
cones
horizontalcells
bipolarcells
amacrinecells
ganglion cells
nerve fiberlayer
outerplexiform
layer
innerplexiform
layer
Figure 3. Cells in the retina are arrayed in discrete layers. The photoreceptors are at the top ofthis rendering, close to the pigment epithelium. The bodies of horizontal cells and bipolar cellscompose the inner nuclear layer. Amacrine cells lie close to ganglion cells near the surface ofthe retina. Axon-to-dendrite neural connections make up the plexiform layers separating rowsof cell bodies.

their plasma membranes fuse withthose of neighboring horizontal cells atgap junctions. The membrane potentialsof a whole sheet of cells become thesame; consequently, horizontal cells re-spond to light over a very large area.Meanwhile, a single bipolar cell receivesinput from a handful of cones and thushas a medium-size receptive field.
Whereas a single bipolar cell with itsOFF or ON light response would carrya fairly blurry response to its ganglioncell, horizontal cells add an opponentsignal that is spatially constrictive, giv-ing the bipolar cell what is known as acenter surround organization (Figure 9).The bipolar center signals either ON orOFF, and the horizontal cells add anOFF or ON surround signal, by one oftwo means. The horizontal cells can ei-ther signal the bipolar cell or feed infor-mation back to the cone photoreceptorsthemselves, which then feed forwardinformation to the bipolar cells thecones contact. Feedback to the cones isnow proposed to occur by means of anunusual electrical synapse consisting ofhalf a gap junction; these hemi gap junc-tions are thought to change the ionic en-vironment across the membrane of the
cone photoreceptor. This complicated cir-cuit from horizontal cell to cone to bipo-lar cells is still a subject of hot debate inthe community of retina scientists.
Horizontal-cell function has occupiedmany vision scientists for decades, andmuch is now known about the role of
these cells in the organization of visualmessages. Horizontal cells respond tomore than the photoreceptors that linkto them. Feedback signals from the innerplexiform layer influence horizontal-cellactivity as well. These feedback signalsare transmitted via substances such as
2003 January–February 31
rhodopsin disc
rhodopsin
surface
N
C
retinal 11-cis retinal 11-trans
light
O
O
Figure 4. Cone photoreceptors from a monkey are stained with a fluorescent green dye (left). When the outer segments of cones or rods are mag-nified further, stacked membrane disks are visible inside (middle). The disks are studded with thousands of rhodopsin complexes. Eachrhodopsin consists of a membrane-traversing protein with a retinal molecule embedded in its core (right). When exposed to light, one of thebonds in the retinal molecule rotates, changing the shape of the protein (lower right). (Middle photograph courtesy of Carlos Rozas.)
fovea opticnerve
visual streak
optic nerve
human retina turtle retina
Figure 5. An ophthalmologist’s view of the human retina (left) shows the optic nerve head,from which blood vessels radiate to nourish every part of the tissue, as well as the fovea,which is where images focus most directly and is specialized for sharp daylight vision. Thefovea is densely packed with more than half of the human cone photoreceptors. The turtle reti-na (right) has no blood vessels radiating from the optic nerve and no fovea. Instead, it has a vis-ual streak, a region of specialized cells running horizontally above the optic nerve, which candetect fleeting movements and orient those movements relative to the horizon.
© 2003 Sigma Xi, The Scientific Research Society. Reproductionwith permission only. Contact [email protected].

dopamine, nitric oxide and retinoicacid. The result is that horizontal cellsmodulate the photoreceptor signal un-der different lighting conditions—al-lowing signaling to become less sensi-tive in bright light and more sensitivein dim light—as well as shaping the re-ceptive field of the bipolar cells, as wehave seen. The horizontal cells can
even make the bipolar cells’ responsecolor-coded, all apparently throughfeedback circuits to the cones.
The ganglion cells have a receptivefield organized as concentric circles.The amacrine-cell circuitry in the innerplexiform layer conveys additional in-formation to the ganglion cell, possiblysharpening the boundary between cen-
ter and surround even further than thehorizontal-cell input does. In humanretinas, two basic types of ganglioncells—ON center and OFF center—form the major output of the retina tothe visual centers in the brain (Figure10, left). ON-center ganglion cells are ac-tivated when a spot of light falls in thecenter of their receptive field and areinactivated when light falls on thefield’s periphery. OFF-center ganglioncells react in the opposite way: Their ac-tivity increases when the periphery oftheir receptive field is lit and decreaseswhen light falls on the center of thefield. (The receptive fields of ganglioncells are modeled as the difference be-tween Gaussian distributions, givingthem a so-called Mexican-hat shape.)
In contrast to the rest of the retina, thehuman fovea contains midget ganglioncells, which have minute dendritic treesconnected in a one-to-one ratio withmidget bipolar cells (Figure 10, right). Thechannel from midget bipolar to midgetganglion cell carries information from asingle cone, thus relaying a point-to-point image from the fovea to the brain.Each red or green cone in the centralfovea connects to two midget ganglioncells, so at all times each cone can eithertransmit a dark-on-light (OFF) signal or alight-on-dark (ON) message. The mes-sage that goes to the brain carries bothspatial and spectral information of thefinest resolution.
Messages from blue cones are notprocessed in the same way as from redand green cones for some reason, possi-bly because the blue system is older inevolutionary terms. Blue cones arefound in the retinas of most species.The typical mammalian retina also hasgreen cones; primates have the addi-tional red cones. Blue cones transmit in-formation through a special blue conebipolar cell to a different type of gan-glion cell, which can carry both a blueON and a yellow OFF response.
Electrical recordings show that sev-eral types of ganglion cells do not haveconcentric organization, especially inanimals whose eyes lack a fovea. Thisincludes most nonmammalian speciesand mammalian species that have reti-nas with visual streaks. Comparedwith species with foveas, the specieswith visual streaks do even more im-age processing in the retina itself be-fore sending a message to the brain;their retinas can immediately synthe-size information about image motionand direction of motion.
32 American Scientist, Volume 91
© 2003 Sigma Xi, The Scientific Research Society. Reproductionwith permission only. Contact [email protected].
Figure 6. A single green-sensitive cone photoreceptor responds to the presence of green lightby becoming hyperpolarized; that is, the membrane’s electrical potential becomes more nega-tive. The hyperpolarization lasts as long as the light flash (top right). The cone only responds tolight immediately directed to it, so its receptive field is very narrow (bottom right).
pola
rizat
ion
time
hype
rpol
ariz
atio
n
receptive field
green flash
rod
ONbipolar
cell
ONbipolar
cell
inhibitoryglutamatereceptor
inhibitoryglutamatereceptor
excitatoryglutamatereceptor
glutamate glutamate
cones
OFFbipolar
cell
a b c
Figure 7. Photoreceptors transmit information to bipolar cells using the molecule glutamate,but different bipolar cells respond differently to the presence of the molecule; some fire in re-sponse, whereas others cease firing, depending on the kind of glutamate receptor on their sur-face. ON bipolar cells have a depolarizing receptive field (a, b); OFF cells have a hyperpolar-izing receptive field (c). Contrary to what one might expect, photoreceptors stop releasingglutamate when stimulated by light, in turn causing ON bipolar cells to release glutamate.

Building Images with Amacrine CellsThere is more to understand about themessages ganglion cells receive beforethey transmit a signal to the brain. Forthat, it is important to appreciate theorganization of the inner plexiform lay-er, where 22 or more different types ofamacrine cells make synaptic connec-tions with about 20 different types ofganglion cells.
It was already clear from Cajal’s de-scription in the 19th century thatamacrine-, ganglion- and bipolar-celldendrites and axons were organizedinto distinct layers; Cajal himself divid-ed the inner plexiform layer into fivestrata. But what sorts of synapses wereformed among the tangle of intermesh-ing processes and what this organiza-tion meant were not immediately ap-parent. Electron microscopy helped tounravel this neurocircuitry. Now theinterconnections of nine types of bipo-lar cells, 14 types of amacrine cells andeight types of ganglion cells are under-stood quite well. We can say we arehalf way to the goal of understandingthe neural interplay between all thenerve cells in the retina.
Much is now known about whattypes of neurotransmitters differentamacrine cells contain and about theorganization of receptors at the differ-ent synapses. Amacrine cells are aboutequally divided between those that useglycine and those that use GABA(gamma-aminobutyric acid) neuro-transmitters.
Glycinergic amacrine cells are usu-ally “small field.” Their processes canspread vertically across several stratawithin the inner plexiform layer, butthey extend relatively short distanceshorizontally. Glycinergic amacrine cellsreceive information from bipolar cellsand transmit information to ganglioncells and to other bipolar and amacrinecells. Some glycinergic amacrine cellsprovide interconnections between ONand OFF systems of bipolar and gan-glion cells. The most famous of theseis called the AII cell; the AII and aGABA-releasing amacrine cell calledA17 are pivotal in the circuitry of rod-based, dim-light vision in the mam-malian retina. These cells aren’t foundin mammalian species that are activesolely in daylight and have very fewrods—for example, squirrels.
In the earlier discussion of ON andOFF channels emanating from cones, Ineglected to talk about the channelsfrom rod cells. Whereas cones connect
in a direct pipeline to bipolar cells toganglion cells, the bipolar cells that re-ceive input from rods do not synapsewith ganglion cells directly. The bipolarcells connected to rods are all of onetype, solely transmitting an ON signal,and use the AII and A17 amacrine cellsas intermediaries to get signals to gan-glion cells. The small-field AII cell col-lects from about 30 rod-connectedbipolar cells and transmits a depolariz-ing message both to ON (light-detect-ing) cone bipolar cells and to their ONganglion cells and to OFF cone bipolarcells and OFF ganglion cells (Figure 11).It is as if the AII cells developed in therod-dominated parts of the retina as anafterthought to the cone-to-ganglioncell architecture and now takes advan-tage of the preexisting cone pathwaycircuitry.
At the same time, the A17 amacrinecell collects rod messages from thou-sands of rod-connected bipolar cells. Itsomehow amplifies and modulates theinformation from the rod bipolar cellsto transmit to the AII cells, but how itdoes this is not completely understood.In any case, the rod pathway with itsseries of convergent and then diver-gent intermediary neurons is clearlywell designed to collect and amplifyscattered vestiges of light for twilightand night vision.
Wide-field amacrine cells sometimesstretch horizontally across the innerplexiform layer for hundreds of mi-crons and interact with hundreds ofbipolar cells and many ganglion cells.Such amacrines are usually confined toone of the five different strata of the in-
ner plexiform layer and create elegantmeshworks of dendrites. Usually, theyemit GABA as a neurotransmitter.Sometimes they connect to neighbor-ing amacrine cells by gap junctions, in-creasing their sphere of influence andthe speed at which signals transmitacross large areas of retina.
Most GABA-releasing amacrinecells also release at least one otherneuroactive substance. The secondary
2003 January–February 33
© 2003 Sigma Xi, The Scientific Research Society. Reproductionwith permission only. Contact [email protected].
hype
rpol
ariz
atio
n
Figure 8. Photomicrograph of a cat retina shows the elongated cone photoreceptors and the hor-izontal and bipolar cells to which they connect (left). The horizontal cells are stained yellow;the bipolar and amacrine cells lie below them, stained green and red. Horizontal cells modu-late the responses of photoreceptors and bipolar cells. The receptive fields of horizontal cellsare very wide (right) because of electrical coupling among the cells.
cones
horizontalcells
gapjunctions
center surround
bipolarcell
Figure 9. Horizontal cells accumulate informa-tion from a wide field of cones and influencethe signals bipolar cells transmit by adding anopponent surround signal to their receptivefields. The horizontal cells influence bipolarcells either directly or by feeding back infor-mation to the cones—probably both.
34 American Scientist, Volume 91
© 2003 Sigma Xi, The Scientific Research Society. Reproductionwith permission only. Contact [email protected].
bipolarcells
midgetbipolarcells
ON
ON-center
ON OFF
OFF
OFF OFFOFF
ON
horizontalcells
ganglion cells
midgetganglion
cells
to brain to brainto
brain
in fovea
ON center,OFF surround
OFF center,ON surround
OFF center,ON surround
ON center,OFF surround
depo
lariz
atio
n
depo
lariz
atio
n
hyperpolarization
hyperpolarization
OFF-center
ON ON
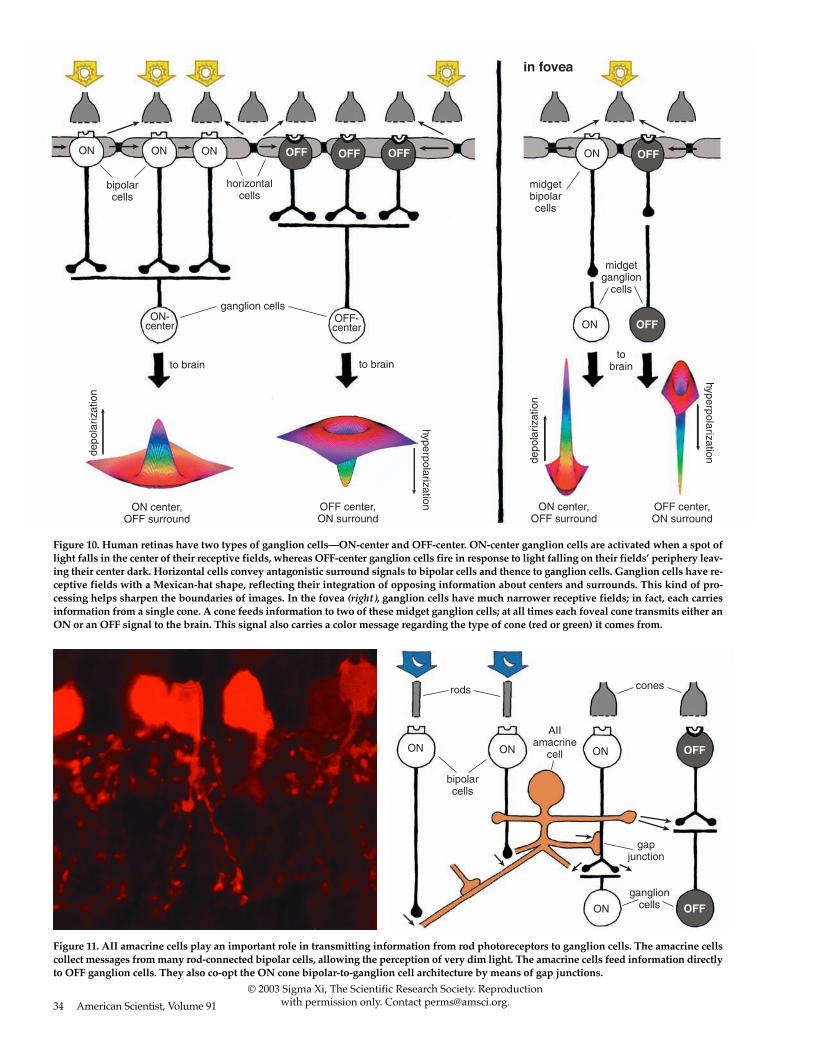
Figure 10. Human retinas have two types of ganglion cells—ON-center and OFF-center. ON-center ganglion cells are activated when a spot oflight falls in the center of their receptive fields, whereas OFF-center ganglion cells fire in response to light falling on their fields’ periphery leav-ing their center dark. Horizontal cells convey antagonistic surround signals to bipolar cells and thence to ganglion cells. Ganglion cells have re-ceptive fields with a Mexican-hat shape, reflecting their integration of opposing information about centers and surrounds. This kind of pro-cessing helps sharpen the boundaries of images. In the fovea (right), ganglion cells have much narrower receptive fields; in fact, each carriesinformation from a single cone. A cone feeds information to two of these midget ganglion cells; at all times each foveal cone transmits either anON or an OFF signal to the brain. This signal also carries a color message regarding the type of cone (red or green) it comes from.
Figure 11. AII amacrine cells play an important role in transmitting information from rod photoreceptors to ganglion cells. The amacrine cellscollect messages from many rod-connected bipolar cells, allowing the perception of very dim light. The amacrine cells feed information directlyto OFF ganglion cells. They also co-opt the ON cone bipolar-to-ganglion cell architecture by means of gap junctions.
rods cones
AIIamacrine
cell
bipolarcells
ganglioncells
ON ON ON
ON
OFF
OFF
gapjunction

substances are usually neuromodulatorsrather than fast-acting neurotransmit-ters. The substances include peptides—“substance P,” somatostatin, vasointesti-nal peptide and cholecystokinin—aswell as the more familiar biomoleculesserotonin, dopamine, acetylcholine,adenosine and nitric oxide. A variety ofreceptors have been found on ganglionand bipolar cells—for example, recep-tors for peptides, nicotine and mus-carine (mushroom toxin) in addition todifferent forms of GABA receptors—in-dicating that amacrine cells are releasingsuch agents. Most of these neuro-modulators are not active at convention-al synapses; their release is thought toinfluence neurons even at a distance bydiffusion. Such neuromodulators appar-ently influence the retinal circuitry un-der changing light conditions or evencause retinal activity to reflect the differ-ent times of day in the circadian clock.
A specialized amacrine cell releasesdopamine when the retina is stimulat-ed with intermittent flashing light.Dopamine causes the gap junctionsamong horizontal cells to become un-coupled, reducing the size of their re-ceptive fields. Furthermore, the neuro-transmitter affects the glutamatereceptor on horizontal cells so that theamplitude of the light response de-clines. Again in the inner plexiformlayer, dopamine closes gap junctions,this time the ones that link AII ama-crine cells in large networks. The re-
sulting uncoupling of the AII cellsmakes the effective field of influence ofthe rod-system amacrine cells muchless significant in lighter conditions.Similarly in bright light conditions, an-other wide-field amacrine cell releasesnitric oxide to uncouple the AII cellfrom the cone-bipolar system. All thisremoves the interference of the large-field rod pathway from the narrow-field cone pathways.
The above broad sketch of retinalcircuitry suggests that the retina is re-markably complex. As vision researchadvances, the retina seems to take onan increasingly active role in percep-tion. Although we do not fully under-stand the neural code that the gan-glion-cell axons send as trains of spikesinto the brain, we are coming close tounderstanding how ensembles of gan-glion cells respond differently to as-pects of the visual scene and how fieldsof influence on particular ganglioncells are constructed. Much of the con-struction of the visual images doesseem to take place in the retina itself,although the final perception of sightis indisputably done in the brain.
Given how much is now known, itmight be fair to ask, are we finishedwith the retina, or are there more sur-prises on the horizon? Earlier surprisesincluded finding that much of the in-formation transfer depended on elec-trical connections among cells ratherthan standard chemical synapses. For
example, the major neural pathwayfrom the rods depends on direct elec-trical connections. Some other fast-act-ing signals pass from amacrine cellsinto ganglion cells at gap junctions.Neuromodulators change the milieu ofthe neuron circuits but act from a dis-tance by diffusion rather than at closelyapposed synapses. Again, this is a sur-prising concept compared to the previ-ous view that all neural interactionstake place via neurotransmitters at spe-cialized isolated patches of membraneapposition—that is, synapses. Themost recent surprise has been that apreviously unknown ganglion cell typeappears to function as a giant photore-ceptor itself, without needing inputfrom rods or cones. This ganglion’s cellmembrane contains light-reactive mol-ecules known as melanopsins. Givensuch unexpected findings, it appearsthat there may still be much more tolearn about how the retina works.
BibliographyDowling, J. E. 1987. The Retina: An Approach-
able Part of the Brain. Cambridge, Mass.:Belknap Press.
Hattar, S., H.-W. Liao, M. Takao, D. M. Bersonand K.-W Yau. 2002. Melanopsin-containingretinal ganglion cells: Architecture, projec-tions, and intrinsic photosensitivity. Science295:1065–1070.
Kolb, H. and E. V. Famiglietti. 1974. Rod andcone pathways in the inner plexiform layerof the cat retina. Science 186:47–49.
Kolb, H. , R. Nelson, P. Ahnelt and N. Cuenca.2001. Cellular organization of the vertebrateretina. In Concepts and Challenges in RetinalBiology: A Tribute to John E. Dowling, pp.3–26, ed. H. Kolb, H. Ripps and S. Wu. Am-sterdam: Elsevier Press.
Kolb, H., E. Fernandez and R. Nelson. 2002. Web-vision: The Organization of the Retina and VisualSystem. http://www.webvision.med.utah.edu
Nelson, R., E. V. Famiglietti and H. Kolb. 1978.Intracellular staining reveals different lev-els of stratification for on-center and off-cen-ter ganglion cells in the cat retina. Journal ofNeurophysiology 41:427–483.
Rodieck, R. W. 1998. The First Steps in Seeing.Sunderland, Mass.: Sinauer Associates.
2003 January–February 35
© 2003 Sigma Xi, The Scientific Research Society. Reproductionwith permission only. Contact [email protected].
Figure 12. Photographs highlighting two different kinds of amacrine cells show their densenetwork of dendrites and axons, which send information to various types of cells. One isstained for its neuromodulator, dopamine (left), and the other for its neurotransmitter, acetyl-choline (right). Many amacrine cells are electrically coupled by gap junctions, creating a mas-sive sheet of cells able to transmit information quickly and in unison.
Links to Internet resources for furtherexploration of “How the Retina
Works” are available on the American
Scientist Web site:
http://www.americanscientist.org/articles/03articles/kolb.html