GYRFALCON FALCO RUSTICOLUS POST-GLACIAL ......if the birds have no former familiarity with the...
12
81 GYRFALCON FALCO RUSTICOLUS POST-GLACIAL COLONIZATION AND EXTREME LONG-TERM USE OF NEST SITES IN GREENLAND KURT K. BURNHAM, 1,2,†, WILLIAM A. BURNHAM, 2,^, AND IAN NEWTON 3 1 Edward Grey Institute, Department of Zoology, University of Oxford, South Parks Road, Oxford, OX1 3PS, UK. E-mail: [email protected] † Present address: High Arctic Institute, 603 10th Avenue, Orion, IL 61273, USA 2 The Peregrine Fund, 5668 West Flying Hawk Lane, Boise, ID 83709, USA 3 Centre for Ecology & Hydrology, Maclean Building, Benson Lane, Crowmarsh, Gifford, Wallingford, Oxon OX10 8BB, UK ^ Deceased ABSTRACT.—Gyrfalcons (Falco rusticolus) use the same nest-sites over long periods of time, and in the cold dry climate of Greenland, guano and other nest debris decay slowly. Nineteen guano samples and three feathers were collected from 13 Gyrfalcon nests with stratified faecal accumu- lation in central-west and northwest Greenland. Samples were 14 C dated, with the oldest guano sample dating to c. 2740–2360 calendar years (cal yr) before present (BP) and three others were likely > 1000 cal yr BP. Feather samples ranged from 670 to 60 cal yr BP. Although the estimated age of material was correlated with sample depth, both sample depth and guano thickness gave a much less reliable prediction of sample age than use of radiocarbon dating on which the margin of error was less. Older samples were obtained from sites farther from the current Greenland Ice Sheet and at higher elevations, while younger samples were closer to the current ice sheet and at lower elevations. Values for δ 13 C showed that Gyrfalcons nesting further from the Greenland Ice Sheet had a more marine diet while those nesting closer to the ice sheet (= further inland) fed on a more terrestrial diet. The duration of nest site use by Gyrfalcons is a probable indicator of both the time at which colonization occurred and of the palaeoenvironmental conditions and patterns of glacial retreat. Nowhere before has such extreme long-term to present use of raptor nest sites been documented. Reproduced with permission of the authors from Ibis (2009) 151:514–522. BURNHAM, K. K., W. A. BURNHAM, AND I. NEWTON. 2011. Gyrfalcon Falco rusticolus post-glacial colonization and extreme long-term use of nest sites in Greenland. Reproduced, pages 81–92 in R. T. Watson, T. J. Cade, M. Fuller, G. Hunt, and E. Potapov (Eds.). Gyrfalcons and Ptarmigan in a Changing World, Volume II. The Peregrine Fund, Boise, Idaho, USA. http://dx.doi.org/ 10.4080/gpcw.2011.0123 Key words: Carbon dating, guano, palaeoenvironmental conditions, δ 13 C. 81
Transcript of GYRFALCON FALCO RUSTICOLUS POST-GLACIAL ......if the birds have no former familiarity with the...

81
GYRFALCON FALCO RUSTICOLUS POST-GLACIAL COLONIZATION AND EXTREME LONG-TERM USE OF NEST SITES IN GREENLAND
KURT K. BURNHAM,1,2,†, WILLIAM A. BURNHAM,2,^, AND IAN NEWTON3
1Edward Grey Institute, Department of Zoology, University of Oxford, South Parks Road, Oxford, OX1 3PS, UK.
E-mail: [email protected]
†Present address: High Arctic Institute, 603 10th Avenue, Orion, IL 61273, USA
2The Peregrine Fund, 5668 West Flying Hawk Lane, Boise, ID 83709, USA
3Centre for Ecology & Hydrology, Maclean Building, Benson Lane, Crowmarsh, Gifford,Wallingford, Oxon OX10 8BB, UK
^Deceased
ABSTRACT.—Gyrfalcons (Falco rusticolus) use the same nest-sites over long periods of time, andin the cold dry climate of Greenland, guano and other nest debris decay slowly. Nineteen guanosamples and three feathers were collected from 13 Gyrfalcon nests with stratified faecal accumu-lation in central-west and northwest Greenland. Samples were 14C dated, with the oldest guanosample dating to c. 2740–2360 calendar years (cal yr) before present (BP) and three others werelikely > 1000 cal yr BP. Feather samples ranged from 670 to 60 cal yr BP. Although the estimatedage of material was correlated with sample depth, both sample depth and guano thickness gave amuch less reliable prediction of sample age than use of radiocarbon dating on which the marginof error was less. Older samples were obtained from sites farther from the current Greenland IceSheet and at higher elevations, while younger samples were closer to the current ice sheet and atlower elevations. Values for δ13C showed that Gyrfalcons nesting further from the Greenland IceSheet had a more marine diet while those nesting closer to the ice sheet (= further inland) fed ona more terrestrial diet. The duration of nest site use by Gyrfalcons is a probable indicator of boththe time at which colonization occurred and of the palaeoenvironmental conditions and patternsof glacial retreat. Nowhere before has such extreme long-term to present use of raptor nest sitesbeen documented. Reproduced with permission of the authors from Ibis (2009) 151:514–522.
BURNHAM, K. K., W. A. BURNHAM, AND I. NEWTON. 2011. Gyrfalcon Falco rusticolus post-glacialcolonization and extreme long-term use of nest sites in Greenland. Reproduced, pages 81–92 inR. T. Watson, T. J. Cade, M. Fuller, G. Hunt, and E. Potapov (Eds.). Gyrfalcons and Ptarmigan ina Changing World, Volume II. The Peregrine Fund, Boise, Idaho, USA. http://dx.doi.org/10.4080/gpcw.2011.0123
Key words: Carbon dating, guano, palaeoenvironmental conditions, δ13C.
81

IT IS WELL KNOWN that raptors may re-use nestsites for generations and some possibly forcenturies (Newton 1979). Hickey (1942)referred to these nest sites as “ecological mag-nets.” These locations are evidently so desir-able that they are re-used again and again, evenif the birds have no former familiarity with thelocation. For example, Peregrine Falcons(Falco peregrinus) disappeared from largeareas of North America and Europe during the1960s as a result of organochlorine pesticideuse. Many years later, following the restrictionin the use of these chemicals, released or recol-onizing Peregrines usually reoccupied formerterritories first, frequently re-using the samenest ledges as their predecessors (Newton1979, Ratcliffe 1993, Oakleaf 2003).
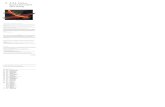
Gyrfalcons (Falco rusticolus) and PeregrineFalcons both breed in the Arctic. Falcons donot build nests, but lay eggs in bowl-shapeddepressions they scrape into existing sub-strates, including old nests made by otherbirds. Arctic Peregrine Falcons typically useopen ledges on cliffs for nesting with little pro-tection from the weather (Cade 1960). Gyrfal-cons usually nest on cliff ledges overhung byrock, in potholes, or in sheltered stick-nestsbuilt by Northern Ravens Corvus corax (Cade1960, Burnham and Mattox 1984). Thesenesting situations provide protection fromfalling rock and the extreme weather condi-tions often found during the early part of theGyrfalcon breeding season. While stick nestsare frequently damaged beyond re-use in a sin-gle season, some ledges and potholes are usedlong-term by Gyrfalcons (Burnham and Mat-tox 1984). Gyrfalcons and Northern Ravensdo not seem to alternate the use of the samenest site from year to year in Greenland. At re-used sites, faecal accumulation frequentlyoccurs where Gyrfalcons roost and nest.Deposited over periods of years, the stratifiedaccumulation of guano can become greaterthan 1.5 m thick in locations protected fromerosion and where dry and cold environmentalconditions enhance preservation (Figure 1).
There are many such nest sites throughout ice-free areas of Greenland.
The Greenland Ice Sheet covers 82% of theland mass of Greenland (Ohmura et al. 1999),and ice-free land occurs only along the periph-ery of the island. Nest-site availability for fal-cons may be affected by climate-inducedglacial retreat and advance covering andexposing cliffs. Long-term use of nest-sites byGyrfalcons is a potential indicator forpalaeoenvironments and of stable glacial con-ditions. As the ice sheet retreated, areas athigher elevations, having a thinner layer of iceand snow, were exposed first. Land at lowerelevations, particularly valley bottoms, had thethickest covering of ice and were exposed last(Fristrup 1966). We therefore hypothesizedthat guano in Gyrfalcon nests at lower eleva-tions and closer to the current ice sheet wouldhave accumulated over shorter periods thanthat at nests at higher elevations and furtherfrom the current Greenland Ice Sheet. Toinvestigate this question we sampled andradiocarbon dated guano from Gyrfalcon nest-sites in two areas, in Kangerlussuaq, central-west (66.50–67.00° N), and Thule, northwest(76.25–77.17° N), Greenland. These studyareas were separated by about 9° latitude and1100 km.
82
– BURNHAM ET AL. –
Figure 1. Gyrfalcon nest site 123 being sampledfor radiocarbon dating in Kangerlussuaq,central-west Greenland, with a 14C age of 1160 to920 cal yr BP.

STUDYAREAS
The Kangerlussuaq study area is located at thehead of a 175-km-long fjord and about 25 kmfrom the current ice sheet margin. The LowArctic tundra landscape in this area wassculpted by glaciation, with rolling hills andvalleys, moraines and lakes, dissected by sev-eral meltwater rivers, and dominated mainlyby shrubs up to 2 m in height. Primary preyspecies for Gyrfalcons in Kangerlussuaqinclude both Rock Ptarmigan (Lagopus mutus)and Arctic Hare (Lepus arcticus), with lesserquantities of passerines and waterfowl con-sumed (Booms and Fuller 2003). It is one ofthe largest deglaciated land areas in Greenlandand, because of extensive past research, pro-vides one of the most complete records ofGreenland’s glacial history (Ten Brink andWeidick 1974, Eisner et al. 1995). From pro-jected rates of deglaciation (see below), we canestimate that some Gyrfalcon nest-sites mayhave been uncovered c. 6500–6000 yearsbefore present (yr BP).
The Thule study area is centered aroundPituffik/Thule Air Base and the current icesheet margin lies up to 26 km inland from thesea, but it reaches the sea at several locations.The environment is High Arctic with anappearance of recent deglaciation, and sparselyvegetated prostrate growing herbs and shrubs.In this area, Gyrfalcons fed primarily on LittleAuks (Alle alle), Rock Ptarmigan and ArcticHares, with seabirds, waterfowl, and passer-ines taken in smaller numbers (Burnham2008). Information on glaciation in Thule ismore limited than for Kangerlussuaq, and gla-cial history is predicted more from inference ofpast climates than from moraine locations andmeasurement (Davies et al. 1963, Fredskild1985, Kelly et al. 1999). Deglaciation suffi-cient to allow for consistent use of existingGyrfalcon nest sites may not have occurreduntil 1350 yr BP or more recently (see below).
METHODS
Gyrfalcon nest-sites are distributed widely andirregularly throughout the two study areas andGreenland as a whole. All sites are on cliffsand usually inaccessible without technical rockclimbing equipment. The amount of faecalmaterial build-up at sites varies greatly, as doesthe structure and size of the nest. The rocksubstrate upon which falcon guano accumu-lates is usually irregular, sloping, and seldomflat. Particularly on large ledges, the actualnest scrape is not always in the same locationeach year, nor is the associated faecal deposi-tion resulting from breeding. Also, the rates oferosion are probably not constant across thenest, as some locations in the nest are moreprotected from weather than are others. There-fore, the guano is not of constant depththroughout the site, and it is difficult to knowwhere the thickest and/or oldest deposits maylie. At nest-sites where deposits seemed ofmore-or-less uniform thickness throughout, asingle sample was collected where guano camein contact with the rock, while at other sitesmore than one sample was collected in anattempt to obtain the oldest guano. Bulk sam-ple materials were collected from several cen-timeters of stratified guano, probablyrepresenting accumulation over decades orlonger. Notes were made of sample depth(cm), cliff height (m), and nest ledge elevation(meters above sea level, m asl). All nests sam-pled had been occupied by Gyrfalcons withinthe past 25 years (Table 1).
When collecting samples, layers of faecalbuild-up were excavated carefully to preventdamage of the site for future use by Gyrfal-cons. At nest-sites with only a few centimetersof faecal build-up, we dug vertically down intothe guano, extracted samples at its base, andthen refilled the hole to minimize damage tothe nest site. At sites with substantial build-up,samples were taken from the side by using a
83
– GYRFALCON LONG-TERM NEST USE –

84
– BURNHAM ET AL. –
Table 1. Results of the 14C measurements on guano material and feathers from Gyrfalcon nest sites inKangerlussuaq, central-west, and Thule, northwest Greenland, collected from 2002 to 2004. Samplesfrom the same nest site are designated by the use of A, B, C, or D following the number.
aGyrfalcon feather samples.bpMC (percent Modern Carbon) analyzed material was post-1950 and had more 14C than did the AD 1950 referencestandard due to atomic bomb testing and subsequent fall-out, 50 years used as conventional 14C age whileCalibrated Age Range is outside calibration range.
019
053A
053B
053Ca
068A
068B
082
087A
087B
087C
087Da
123
163
170
201A
201B
500A
500B
501
502
503A
503Ba
195586
191125
191126
191295
191127
195576
195577
168839
168840
168838
168837
195578
195587
195579
195580
195581
195583
195582
195584
195585
191124
191348
20–25
30–35
15–20
15
15–20
10–15
25–30
30–35
25–30
0–3
15
20–25
100-110
15–20
35–40
10–15
5–8
3–6
2–4
15–20
20–25
20
15
15
15
15
152
152
76
46
46
46
46
46
152
24
23
23
46
46
117
23
76
76
355
304
304
304
55
55
441
365
365
365
365
258
395
103
200
200
152
152
122
152
304
304
9
11
11
11
14
14
5
54
54
54
54
62
72
28
78
78
14
14
2
9
17
17
-22.7
-22.4
-22.4
-22.5
-22.9
-20.7
-23.2
-21.9
-20.9
-24.2
-21.3
-21.1
-20.8
-21.3
-17.8
-21.3
-23.3
-21.2
-23.2
-20.2
-19.1
-19.5
1992
1991
1991
1991
2000
2000
1999
2002
2002
2002
2002
~1985
2003
2004
2000
2000
2003
2003
2004
2004
2004
2004
Nest-site
Betaanalyticsample #
Sampledepth (cm)
Cliff height
(m)
Nestelevation(m asl)
Distancefrom icemargin(km)
δ13C (‰)
(PDB)Lastused
Conventional 14C age
(yr BP ± 1σ)
Calibrated age range
(cal yr BP ± 2σ)
Kangerlussuaq, central-west Greenland
Thule, northwest Greenland
790 ± 60
1290 ± 50
1090 ± 80
320 ± 30
570 ± 50
240 ± 60
830 ± 70
2480 ± 40
350 ± 40
105.09 ± 0.66 pMCb
170 ± 40
1090 ± 60
960 ± 60
690 ± 60
1430 ± 70
820 ± 60
113.34 ± 0.71 pMCb
122.08 ± 0.73 pMCb
115.29 ± 0.66 pMCb
640 ± 50
650 ± 70
650 ± 40
790–650
1300–1080
1180–900, 850–810
470–300
650–520
450–260, 220–140,30–0
920–660
2740–2360
500–300
outside calibration rangeb
300–60, 40–0
1160–920
970–740
710–550
1480–1470, 1430–1250
910–660
outside calibration rangeb
outside calibration rangeb
outside calibration rangeb
670–540
690–530
670–550

masonry hammer and a small trowel to exca-vate horizontally until rock was reached. Theamount of bulk material collected from thestratified samples varied, but in all cases wassufficient for dating using standardized radio-carbon procedures. Within the nest sites,feathers (from probable prey and/or Gyrfal-cons) and bones (prey) were found duringexcavation. Three feathers that were easilyidentifiable as from Gyrfalcons were alsoradiocarbon dated.
Radiocarbon dating was carried out by BetaAnalytic Inc, Miami, Florida, USA, usingeither the conventional radiometric technique(samples > 30 g) or accelerator mass spectrom-etry (AMS) (samples < 30 g). Guano sampleswere pre-treated using an “acid wash”, whilean “acid/alkali/acid” wash was used for feathersamples. For the conventional radiometrictechnique, materials were analyzed by synthe-sizing carbon to benzene (92% C) and thenmeasuring for 14C in a scintillation spectrome-ter from which the radiocarbon age was calcu-lated (Beta Analytic). AMS results wereobtained by the reduction of sample carbon tographite (100% C) along with standards andbackgrounds (Beta Analytic). Graphite wasthen sent for 14C measurement in an accelera-tor-mass-spectrometer to a research facilitycollaborating with Beta Analytic. The meas-ured radiocarbon ages were returned to BetaAnalytic where verification, isotopic fraction-ation correction using δ13C, and calendar cali-bration took place (Beta Analytic). Calibratedresults provide both a maximum and minimumage for each sample in calendar years (cal yr)before present (BP), with a 95% confidencethat the actual age falls within this range. Cal-ibrations were made using calibration datapublished in Stuiver et al. (1998) using cubicspline fit mathematics, as described by Talmaand Vogel (1993).
From 2002 to 2004, 19 bulk guano samplesand three feathers were collected from 13 Gyr-falcon nest sites. Five guano samples and onefeather were from four nests in Thule, and 14
guano samples and two feathers were fromnine nests in Kangerlussuaq. To test for rela-tionships between age of guano samples andnest variables (using JMP IN, vs. 4, SAS Insti-tute Inc.), we analyzed the distribution of thevariables and relationships using SpearmanCorrelation Analysis, as was most appropriatebased on the non-normal distribution of thedata. Conventional 14C age was used as themaximum age for each sample.
RESULTS
The oldest guano sample was from nest site087 in Kangerlussuaq and was dated 2740 to2360 cal yr BP (Table 1). Three nests inKangerlussuaq showed evidence of occupation> 1000 cal yr BP, with the most recent nestbeing occupied from 650 to 520 cal yr BP. InThule, the oldest nest was between 690 and530 cal yr BP with two nests indicating useonly within the past 50 years (Table 1). Sitesin Kangerlussuaq have been used approxi-mately 1800 to 2000 years longer than thosefurther north in Thule. Sites with multiplesamples collected showed an increase in 14Cage with sample depth (Table 1). The threeanalyzed Gyrfalcon feathers were between 670and 60 cal yr BP. Sample 087D had two pos-sible calibrated age ranges, 300–60 and 40–0cal yr BP, and the older range was most likelyaccurate based on other samples from the samenest and sample depth. While the bulk sam-ples of guano represent a number of years ofsite use, the feathers were from a precisemoment in time.
The maximum conventional 14C age for eachindividual nest was significantly correlatedwith the current distance from the ice margin(rs = 0.56, P < 0.05, n = 13, Figure 2), nest ele-vation (rs = 0.61, P < 0.05, n = 13, Figure 3),and sample depth (rs = 0.88, P < 0.0001, n =13, Figure 4). While sample depth could beused as a predictor of 14C age, the margin oferror would be much greater than if using radiocarbon dating directly (e.g., 087A and 163,Figure 4)
85
– GYRFALCON LONG-TERM NEST USE –

In general, organisms from marine food chainsor animals that feed on them have less negativeδ13C values, while more negative δ13C valuesare typically associated with terrestrial ecosys-tems (Rounick and Winterbourn 1986, Anger-björn et al. 1994). The δ13C values from theoldest guano sample from each nest variedslightly from a median of –21.9‰ (n = 9, σ2 =2.7) for Kangerlussuaq to –21.5‰ (n = 4, σ2 =4.5) for Thule, with no statistical differencebetween the two areas. A significant correla-tion exists between δ13C for the oldest guanosample from each nest and their current dis-tance to the Greenland Ice Sheet (rs = 0.63, P< 0.05, n = 13), with less negative values beingassociated with nests further from the currentice sheet (Figure 5).
DISCUSSION
Radiocarbon dating of Gyrfalcon nest materialfrom Kangerlussuaq and Thule, Greenland,indicate much longer occupancy times thancan be determined from historical records.Sites from Kangerlussuaq were colonizedapproximately 1800–2000 years earlier thanthose further north in Thule, probably an effectof earlier deglaciation and a more stable andwarmer climate in the Kangerlussuaq area.These are some of the longest used raptor nestsites ever documented.
Similar studies of this type have been carriedout on other Arctic and Antarctic species.These have included Snow Petrels (Pago-droma nivea) (Hiller et al. 1988), Thick-billedMurres (Uria lomvia) (Gaston and Donaldson1995), and Adelie Penguins (Pygoscelisadeliae) (Emslie et al. 2007), for which radio-carbon age has been calculated using solidifiedstomach oil deposits, peat moss deposits, andbone and feather samples from molting sites,respectively, and minimum dates for the estab-lishment or colonization of bird colonies havebeen given (~34000 yr BP, 1500–3800 yr BP,and > 44000 yr BP, respectively). In addition,archaeological studies have sometimesrevealed long-term use of raptor nest-sites.
86
– BURNHAM ET AL. –
0
10
20
30
40
50
60
70
80
90
0 500 1000 1500 2000 2500 3000
conventional c-14 age (yrs BP)
dist
ance
from
ice
mar
gin
(km
)
201a163
123
087a
170
504a
053a
082019
500a
502
068a
503
Figure 2. Relationship between maximum 14Cage and nest site distance from current icemargin (rs = 0.56, P < 0.05, n = 13).
0
50100
150
200
250300
350
400450
500
0 500 1000 1500 2000 2500 3000
conventional c-14 age (yr BP)
nest
ele
vatio
n (m
)082
163087a019
503a 053a
201a
123
502
170
500a501
068a
Figure 3. Relationship between maximum 14Cage and nest site elevation (m above sea level)(rs = 0.61, P < 0.05, n = 13).
0
20
40
60
80
100
120
0 500 1000 1500 2000 2500 3000
conventional c-14 age (yr BP)
sam
ple
dept
h (c
m) 163
087a
500a501
201a
053a123
082
019503a
068a502170
Figure 4. Relationship between maximum 14Cage and sample depth (rs = 0.88, P < 0.0001, n= 13).

For example, remains of Peregrine Falcons,probably adults and nestlings, were found dur-ing an archaeological investigation of prehis-toric human occupation of a cave on HunterIsland, Tasmania (Bowdler 1984). Bones werefound in layers 990 ± 90 to ca. 19000 yr BPsuggesting use by falcons during that time butnot more recently.
There can be little doubt about the identity ofthe Gyrfalcon sites in Greenland. Only twofalcon species breed there (Salomonsen 1950)and although Peregrines sometimes use formerGyrfalcon nest sites, these species typicallyselect different nesting situations, as explainedabove. Cliff-nesting seabirds breed in differentsituations than do falcons. Gulls nest colo-nially and sites cannot be confused with falconnests. Except for Iceland Gulls (Larus glau-coides) and Glaucous Gulls (L. hyperboreus),all other avian species breeding in the researchareas construct nests containing sticks, grass,and feathers, and/or breed on the ground.Ravens are the only other cliff-nesting speciesthat nest in similar locations to Gyrfalcons, butthey construct stick-nests. There can be nodoubt that the accumulation of guano at sites
sampled resulted from long-term use by fal-cons, and in particular Gyrfalcons.
The feather samples could be easily identifiedto Gyrfalcons. Although interesting because oftheir condition and long-term preservation,they did not contribute to knowledge on dura-tion of nest site use or palaeoenvironmentbeyond what could be inferred from guanosamples. They did, however, provide confir-mation of dates of past use.
Some nest-sites provided much older radiocar-bon dated samples than others. Newton (1979)attributed the repeated occupancy of nest sitesto “the superiority of particular places overlocal alternatives.” This superiority couldresult from a nearby abundance of prey, asuperior hunting platform, a location in appro-priate spatial relationship to other territorialpairs, or a site offering good protection frommammalian predators or inclement weather(rain, snow, sun, and/or wind). Over time, thedesirability of sites may change due to alteredenvironmental conditions, including rockstructure. This likelihood may be particularlytrue in areas of recent glacial activity and cli-mate change.
87
– GYRFALCON LONG-TERM NEST USE –
0
20
40
60
80
-24 -23 -22 -21 -20 -19 -18 -17
80
60
40
20
0
163
123
087A
201A
503
502053A
019
068A500A
501082
BlackGuillemot
Little Auk
Rock Ptarmigan
(-24.4)
SnowshoeHare
(-26.4)
dist
ance
from
ice
mar
gin
(km
)
δ13
C ‰
170
Figure 5. Relationship between δ13C and distance from current ice margin (rs = 0.63, P < 0.05, n =13). Frequent prey of Gyrfalcons shown with associated δ13C value. Snowshoe Hare (Lepusamericanus) δ13C value was substituted for Arctic Hare as no values were found in the literature forArctic Hare. δ13C values for Snowshoe Hare, Rock Ptarmigan, Little Auk, and Black Guillemot are fromRoth et al. 2007, Ricca et al. 2007, Hobson et al. 2002, and Hobson et al. 2002, respectively.

The Greenland Ice Sheet gradually retreatedabout 175 km in the Kangerlussuaq areabecause of world-wide climate warming sincethe end of the last glaciation (~15000 yr BP),but with frequent re-expansions (Ten Brinkand Weidick 1974, Funder 1989, Figure 6). Aslow retreat of the ice sheet (1 km/100 yr)occurred from c. 15000 to 10000 yr BP fol-lowed by an oscillatory but more rapid retreat(3 km/100 yr) of nearly 100 km from c. 9500to 6500 yr BP (Ten Brink and Weidick 1974).By 6000 yr BP the ice sheet had reached itspresent position, although between 5- and 10-km re-advances occurred from c. 4800 to 4000and from 2500 to 2000 yr BP (Ten Brink andWeidick 1974, Figure 6). At that time the sealevel was nearly equal to the present day. Eis-ner et al. (1995) reported that Kangerlussuaqexperienced a “climatic optimum” from c.4400 to 3400 yr BP and that a climatically sta-ble period is believed to have encompassed theperiod from 2000 to 1200 yr BP (Figure 6).Based on lichenometry, the period from c. 700yr BP to the present was characterized byoscillatory advance and retreat of the inland icewithin about a 3-km-wide zone (Ten Brink andWeidick 1974).
In Kangerlussuaq, nest-sites 019, 053, 068,082, and 170 were probably covered by iceduring re-advances of glaciers from the Green-land Ice Sheet between c. 4800 and 4000 andfrom 2500 to 2000 yr BP. All five nest-sitesare in glacial valleys or river valleys and fourof the five are on low cliffs. The fifth site(053) is near to the top edge of a deep valley.Furthermore, 068 and 170 are on small cliffsnear low elevation rivers and even a 10 m risein sea level could have affected use of thesesites by Gyrfalcons. Nest sites 087, 123, 163,and 201 are less than 50 km from the ice edgeand, based on the projected rate of ice-sheetretreat, these nest sites were ice-free by 6500–6000 yr BP. That does not preclude, however,the possibility of isolated snow banks and gla-ciers covering nest sites, particularly on smallcliffs and at lower elevations, during periods ofclimatic cooling and glacial expansion. In
addition, it was probably not until the “climaticoptimum,” from 4400 to 3400 yr BP, that thenecessary vegetation was established to sup-port the prey-base utilized by Gyrfalcons inKangerlussuaq.
Information from Thule is far less completeand more contentious than that from Kanger-lussuaq. In Thule, Malaurie et al. (1972) useddata from marine deposits of terraces to esti-mate that deglaciation began c. 8000 yr BP,and Kelly et al. (1999) postulate that much ofthe area has been ice-free for at least 9000years (Figure 6). From the period extending8000—3000 yr BP, the Thule area experienceda climatic optimum, with significant surfacemelting occurring (Reeh 1984, Figure 6).Archaeological evidence indicates unfavor-able, cool conditions from 2500 to 1900 yr BPfollowed by three centuries with a warmer cli-mate (McGhee 1972, Figure 6). During theperiod c. 1350—900 yr BP, a moist and likelywarm climate prevailed, with conditions cool-ing and vegetation changing to drier heath atthe end of this period (Fredskild 1973, Figure6). From c. 900 yr BP to the 20th century, con-siderable climatic variability existed, but evi-dence from ice cores and oxygen isotopicrecords from Camp Century show that gener-ally cooler conditions probably persisted in theThule area until c. 1900 AD (Johnsen et al.1970). Meteorological records from Uper-navik, Greenland (72° 50' N), show a 2°Cmean annual temperature increase through themiddle of the 20th Century (Dowdeswell1996).
In Thule, the existence of consistent long-termnest-site availability is more recent than inKangerlussuaq. While some nest sites mayhave been ice-free as early as c. 9000—6000yr BP, they may have been later re-covered byre-advances of the ice sheet. Furthermore,despite some nest-sites being ice-free forextended periods of time, suitable environmen-tal conditions for Gyrfalcons probably did notexist until between 1350 and 900 yr BP orlater. Even now, because of cool mean sum-
88
– BURNHAM ET AL. –

mer temperatures, localized large snow banksand glaciers can develop in just a few years ordecades and preclude the use of affected nestsites.
The significant correlation between δ13C val-ues and distance to the current ice sheet islikely a reflection of diet, with Gyrfalconsbreeding nearer the coast feeding on a mixeddiet of marine and terrestrial prey items, andthose breeding more inland having an almostcompletely terrestrial diet. Primary terrestrialprey items, such as Rock Ptarmigan and Arctichare, have likely δ13C values in the rangeof –24 to –27‰ (Figure 5, Ricca et al. 2007,Roth et al. 2007) while marine prey items,such as Little Auk and Black Guillemots (Cep-phus grylle), have δ13C values rangingfrom –18 to –20‰ (Figure 5, Hobson et al.2002). Other prey species taken in more lim-ited quantities, such as shorebirds (e.g., RedKnot [Calidris canutus] and Ruddy Turnstone[Arenaria interpres]) and waterfowl (e.g.,Long-tailed Duck [Clangula hyemalis]) have
δ13C values with much wider ranges (–16.6 to–24.7‰, –18.1 to –24.1‰, ~ –17 to –21‰,respectively), mainly as a result of seasonalshifts in foraging between marine and terres-trial areas (Morrison and Hobson 2004, Brauneet al. 2005).
In Kangerlussuaq, δ13C values for each of theoldest 14C dated nests ranged from –23.2 to –17.8‰. Nest 082 has the most negative δ13Cvalue and is closest to the current ice sheet (5km), and nest 201A has the least negative δ13Cvalue and is farthest from the ice sheet (78 km,Figure 5). Of particular interest is nest 087A,which is the oldest sampled nest in Kangerlus-suaq by ~1000 years, and has the median δ13Cof the nine nests sampled in Kangerlussuaq.While this nest is 54 km from the current icesheet, it is approximately 1 km from a largefjord, allowing adult Gyrfalcons potentialaccess to both a marine and terrestrial diet.The benefit of readily accessing such a largevariety of prey may have allowed for earliercolonization of the nest-site. For Thule, nests500A and 501 have very negative δ13C values,with nest 500A almost 100 km from the near-est breeding seabird colonies and prey remainsconsisting almost solely of Arctic Hare inrecent years, and nest 501 in a large wetlandarea, with prey remains primarily consisting ofRock Ptarmigan. The other two Thule nestsites, 502 and 503A, are in areas with largenumbers of Little Auks and Black Guillemots.For both study areas, multiple samples fromthe same nest site show changes of up to –3‰in δ13C over time (e.g., site 087 in Kangerlus-suaq, Table 1), which is likely the result ofchanges in prey composition over time.
On the basis of palaeoenvironmental investi-gations by others, we hypothesized that Gyr-falcon nest sites of higher elevations, andfurther from the ice margin, would showlonger usage patterns than nest sites closer tothe ice edge, and at lower elevations. Bothpredictions were confirmed using 14C dating.Carbon dating of stratified guano accumulationat Gyrfalcon nest-sites could thus be used to
89
– GYRFALCON LONG-TERM NEST USE –
0 yr BP
5,000
10,000
Kangerlussuaq Thule
1
4
3
2
9
8
7
6
14
13
12
11
Wolstenholme Fjord glaciation
Holocene climatic optimum
cooling period
warming periodstable climate
minor ice re-advance
climatic optimum
minor ice re-advance
rapid ice retreat
slow ice retreat
ice sheet at current position
?
oscillatory advance/retreat
?
first Gyrfalcon nest
first Gyrfalcon nest
Figure 6. Glacial and climate history for the past15000 yr BP for the Kangerlussuaq and Thulestudy areas. For sources on dates used in thefigure please see text.

confirm and date when local environmentswere suitable for breeding by this species andwhen colonization probably occurred. Fur-thermore, past use of sites indicates that favor-able environmental conditions existed for preyspecies as well. Values from δ13C provideinsight into likely prey species that existed atthe time of colonization and possible changesin prey species over time. While results fromthis study are specific to Kangerlussuaq andThule, it is somewhat likely that similar corre-lations exist in other regions of Greenland.These results shed further light on local, andprobably regional, palaeoenvironmental condi-tions and glaciation in the Arctic.
ACKNOWLEDGMENTS
First and foremost we thank Andrew Gosler,Jennifer Burnham, Jeff Johnson, Chris Perrins,and David Houston for comments and sugges-tions. Additionally, Konrad Steffen, Univer-sity of Colorado, USA, provided helpfulremarks on this manuscript and suggested cita-tions related to glaciation and paleoenviron-ments in Greenland. Edward Hanna,University of Plymouth, United Kingdom, pro-vided further citations related to glaciation andpaleoenvironments in Greenland. Thank youto the Greenland Home Rule Government andDanish Polar Center for providing permits forthis research. Additionally, the support ofKISS, VECO, 109th Air National Guard, andthe United States Air Force were critical insupporting field work. Brian and Ruth Mutch,Jack Cafferty, Jack Stephens, Robin Abbott,Earl Vaughn, Ed Stockard, and the residents ofboth Kangerlussuaq and Thule Air Basedeserve special thanks for their continuedfriendship and assistance. Financial supportwas provided by The Peregrine Fund, Ruth O.Mutch, The Offield Family Foundation, PeterPfendler, Comer Science and Education Foun-dation, and The G. Unger Vetlesen Foundationand is greatly appreciated.
LITERATURE CITED
ANGERBJÖRN, A., P. HERSTEINSSON, K. LIDÉN,AND E. NELSON. 1994. Dietary variation inArctic Foxes (Alopex lagopus)—An analy-sis of stable carbon isotopes. Oecologia99:226–232.
BETA ANALYTIC, INC. 2005. 4985 S. W. Court,Miami, FL, 33155, USA. www.radiocarbon.com.
BOOMS, T., AND M. FULLER. 2003. Gyrfalcondiet in central west Greenland during thenesting period. Condor 105:528–537.
BOWDLER, S. 1984. Hunter Hill, Hunter Island:archeological investigation of a prehistoricTasmanian site. Terra Australis 8. Aus-tralian National University Press, Canberra,Australia.
BRAUNE, B. M., K. A. HOBSON, AND B. J. MAL-ONE. 2005. Regional differences in collagenstable isotope and tissue trace element pro-files in populations of Long-tailed Duckbreeding in the Canadian Arctic. Science ofthe Total Environment 346:156–168.
BURNHAM, K. K. 2008. Inter- and intraspecificvariation of breeding biology, movements,and genotype in Peregrine Falcon Falcoperegrinus and Gyrfalcon F. rusticolus pop-ulations in Greenland. D. Phil. thesis, Uni-versity of Oxford, Oxford, UK.
BURNHAM, W. A., AND W. G. MATTOX. 1984.Biology of the Peregrine and Gyrfalcon inGreenland. Meddelelser om Grønland: Bio-science 14:1–25.
CADE, T. J. 1960. Ecology of the Peregrine andGyrfalcon populations in Alaska. Univer-sity of California Publications in Zoölogy63:151–290.
CADE, T. J. 1982. The Falcons of the World.William Collins Sons and Co Ltd., London,UK.
DAVIES, W. E., D. B. KRINSLEY, AND A. H.NICOL. 1963. Geology of North Star Bayarea, northwest Greenland. Meddelelser omGrønland 162:1–68.
DOWDESWELL, J. A. 1996. Glaciers in the HighArctic and recent environmental change.Pages 121–134 in P. Wadhams, J. A.
90
– BURNHAM ET AL. –

Dowdeswell, and A. N. Schofield (Eds.).The Arctic and Environmental Change.Gordon and Breach Publishers, London,UK.
EISNER, W. R., T. E. TORNQVIST, E. A. KOSTER,O. BENNIKE, AND J. F. N. VAN LEEUWEN.1995. Paleoecological studies of aHolocene lacustrine record from theKangerlussuaq (Søndre Strømfjord) regionof West Greenland. Quaternary Research43:55–66.
EMSLIE, S. D., L. COATS, AND K. LICHT. 2007.A 45,000 yr old record of Adélie Penguinsand climate change in the Ross Sea,Antarctica. Geology 35:61–64.
FREDSKILD, B. 1973. Studies in the vegeta-tional history of Greenland. Meddelelserom Grønland 198:1–245.
FREDSKILD, B. 1985. The Holocene vegeta-tional development of Tugtuligssuaq andQeqertat, northwest Greenland. Med-delelser om Grønland, Geoscience 14:1–20.
FRISTRUP, B. 1966. The Greenland Ice Cap.Rhodos, International Science Publishers,Copenhagen, Denmark.
FUNDER, S. 1989. Quaternary geology of theice-free areas and adjacent shelves ofGreenland. Pages 741–792 in R. J. Fulton(Ed.). Quaternary Geology of Canada andGreenland, Geological Survey of Canada,Ottawa, Canada.
GASTON, A. J., AND G. DONALDSON. 1995. Peatdeposits and Thick-billed Murre colonies inHudson Strait and Northern Hudson Bay:Clue to post-glacial colonization of the areaby seabirds. Arctic 48:354–358.
HICKEY, J. J. 1942. Eastern population of theDuck Hawk. Auk 59:176–204.
HILLER, A., U. WAND, H. KÄMPF, AND W.STACKEBRANDT. 1988. Occupation of theAntarctic continent by petrels during thepast 35,000 years: Inferences from a 14Cstudy of stomach oil deposits. Polar Biol-ogy 9:69–77.
HOBSON, K. A., G. GILCHRIST, AND K. FALK.2002. Isotopic investigations of seabirds ofthe north water polynya: Contrasting
trophic relationships between the easternand western sectors. Condor 104:1–11.
JOHNSEN, S. J., W. DANSGAARD, H. B.CLAUSEN, AND C. C. LANGWAY. 1970. Cli-matic oscillations 1200–2000 AD. Nature227:482–483.
KELLY, M., S. FUNDER, M. HOUMARK-NIELSEN,K. L. KNUDSEN, C. KRONBORG, J. LANDVIK,AND L. SORBY. 1999. Quaternary glacialand marine environmental history of north-west Greenland: A review and reappraisal.Quaternary Science Reviews 18:373–392.
MALAURIE, J., Y. VASARI, H. HYVARINEN, G.DELIBRIAS, AND J. LABEYRIE. 1972. Prelim-inary remarks on Holocene paleoclimatesin the regions of Thule and Inglefield Land,above all since the beginning of our ownera. Pages 10–133 in Y. Vasari, H. Hyvari-nen, and S. Hicks (Eds.). Climatic Changesin Arctic Areas during the last Ten-thou-sand Years: A Symposium Held at Oulankaand Kevo, 4–10 October 1971. Acta Uni-versitatis Ouluensis, Series A, ScientiaeRerum Naturalium No. 3, Geologica No. 1,University of Oulu, Oulu, Finland.
MCGHEE, R. 1972. Climatic change and thedevelopment of Canadian Arctic culturaltraditions. Pages 39–57 in Y. Vasari, H.Hyvarinen, and S. Hicks (Eds.). ClimaticChanges in Arctic Areas during the lastTen-thousand Years: A Symposium Held atOulanka and Kevo, 4–10 October 1971.Acta Universitatis Ouluensis, Series A, Sci-entiae Rerum Naturalium No. 3, GeologicaNo. 1, University of Oulu, Oulu, Finland.
MORRISON, R. I. G., AND K. A. HOBSON. 2004.Use of body stores in shorebirds afterarrival on High-Arctic breeding grounds.Auk 121:333–334.
NEWTON, I. 1979. Population Ecology of Rap-tors. T. & A. D. Poyser, Berkhamsted, UK.
OAKLEAF, R. 2003. Peregrine restoration froma state biologist’s perspective. Pages 297–304 in T. J. Cade and W. Burnham (Eds.).Return of the Peregrine: a North Americansaga of tenacity and teamwork. The Pere-grine Fund, Boise, Idaho, USA.
91
– GYRFALCON LONG-TERM NEST USE –

OHMURA, A., P. CALANCA, M. WILD, AND M.ANKLIN. 1999. Precipitation, accumulationand mass balance of the Greenland IceSheet. Zeitschrift für Gletsherkunde undGlazialgeologie 35:1–20.
RATCLIFFE, D. 1993. The Peregrine Falcon.T. & A. D. Poyser, London, UK.
REEH, N. 1984. Reconstruction of the glacialice covers of Greenland and the CanadianArctic islands by three-dimensional, per-fectly plastic ice-sheet modeling. Annals ofGlaciology 5:115–121.
RICCA, M. A., A. K. MILES, R. G. ANTHONY, X.DENG, AND S. S. O. HUNG. 2007. Effect oflipid extraction on analyses of stable car-bon and stable nitrogen isotopes in coastalorganisms of the Aleutian archipelago.Canadian Journal of Zoology 85:40–48.
ROTH, J. D., J. D. MARSHALL, D. L. MURRAY,D. M. NICKERSON, AND T. D. STEURY. 2007.Geographical gradients in diet affect popu-lation dynamics of Canada Lynx. Ecology88:2736–2743.
ROUNICK, J. S., AND M. J. WINTERBOURN. 1986.Stable carbon isotopes and carbon flow inecosystems. BioScience 36:171–177.
SALOMONSEN, F. 1950. Grønlands Fugle. TheBirds of Greenland. Munksgaard, Copen-hagen, Denmark.
STUIVER, M., P. J. REIMER, E. BARD, J. W.BECK, G. S. BURR, K. A. HUGHEN, B.KROMER, G. MCCORMAC, J. VAN DER
PLICHT, AND M. SPURK. 1998. IntCal98radiocarbon age calibration, 24,000–0 calBP. Radiocarbon 40:1041–1083.
TALMA, A. S., AND J. C. VOGEL. 1993. A sim-plified approach to calibrating C14 Dates.Radiocarbon 35:317–322.
TEN BRINK, N. W., AND A. WEIDICK. 1974.Greenland Ice Sheet history since the lastglaciation. Quaternary Research 4:429–440.
92
– BURNHAM ET AL. –