
Use of various genetic markers in differentiation of Mycobacterium ...

ORIGINAL ARTICLE
Genetic differentiation of wild and cultivated Lens basedon molecular markers
K. Tewari & H. K. Dikshit & Neelu Jain & Jyoti Kumari &D. Singh
Received: 23 May 2011 /Accepted: 6 October 2011 /Published online: 30 December 2011# Society for Plant Biochemistry and Biotechnology 2011
Abstract Genetic diversity among 83 lentil genotypesincluding 23 wild types, 19 indigenous varieties, 5exotic lines and 36 advanced breeding lines was studiedusing molecular markers. A total of 112 amplicons wereproduced using 15 RAPD and 8 SSR markers. Dendro-gram based on Jaccard similarity coefficient andUPGMA analysis revealed two major clusters and oneminor cluster. Cluster I comprised 21 wild accessions ofL. orientalis and 1 L. ervoides subspecies. NineteenIndian varieties grouped together in subcluster IIAindicating their narrow genetic base. Subcluster IIBconsisted of 41 genotypes including 5 exotic and 36advanced breeding lines mainly derived from exoticgenotypes. The narrow genetic base of released cultivarsand germplasm lines emphasized the need for broadeningof genetic base of breeding material using exoticcollections and wild species to ascertain genetic improve-ment upon existing cultivars.
Keywords Genetic differentiation . Lens .Wild spp .
RAPD . SSR .Molecular markers
AbbreviationsSSR Simple sequence repeatRAPD Random amplified polymorphic DNAISSR Inter simple sequence repeats
PCR Polymerase chain reactionUPGMA Unweighted pair-group method arithmetic averageNTSYS Numerical taxonomy multivariate analysis system
Introduction
Lentil (Lens culinaris subsp. culinaris) is a self pollinatedcrop (2n=2x=14) belonging to the Vicieae tribe in Legumi-nosae family. This crop originated in Near East and was firstdomesticated in Fertile Cresent around 7000 B.C. (Zohary1972a, b). The genus Lens consists of four species and seventaxa- L.culinaris Medikus {subsp. culinaris, subsp. orientalis(Boiss.) Ponert, subsp. tomentosus (Ladizinsky) Ferguson,Maxted, Slageren & Robertson and subsp. odemensis(Ladizinsky) Ferguson, Maxted, Slageren & Robertson}, L.ervoides (Brign.) Grande, L. nigricans (M. Bieb.) Godron andL. lamottei Czefr (Ferguson 2000). Among these subsp.orientalis is considered to be progenitor of the cultivatedlentil. The cultivated lentil is classified as macrosperma ormicrosperma (Barulina 1930) which have been considered assubspecies, races or varieties by different researchers andbreeders. Lentil is grown mainly in Indian Subcontinent,Middle East, North Africa, Southern Europe, North andSouth America, Australia and West Asia. The major lentilproducing regions are Asia (58% of the area) and the WestAsia-North Africa region (37% of the acreage of developingcountries). Lentil is rich source of protein and micronutrientsand is mainly consumed as dal. Lentil is grown in winterseason as rainfed crop. Lentil occupied 1.38 million hectarein India with a production of 0.96 million tons during 2008–09. The major bottlenecks limiting high yield of this crop aresusceptibility to biotic and abiotic stresses, poor harvest
K. Tewari :H. K. Dikshit (*) :N. Jain :D. SinghDivision of Genetics, IARI,New Delhi 110012, Indiae-mail: [email protected]
J. KumariGermplasm Evaluation Division, NBPGR,New Delhi 110012, India
J. Plant Biochem. Biotechnol. (July–Dec 2012) 21(2):198–204DOI 10.1007/s13562-011-0087-9

index, lack of genetic variability and non availability ofsuitable ideotypes for different cropping systems.
Germplasm evaluation and characterization data helpsin identification of diverse genotypes which can beutilized by breeders for creating the desired variation.The availability of low number of morphologicalmarkers, their poorly known genetic control andenvironmental influence on phenotypic expression atdifferent stages of growth are impediments in usingthese as stable genetic markers in diversity analysis. Theevaluation of genetic diversity and construction oflinkage maps promotes the efficient use of geneticvariations in the breeding program (Paterson et al.1991). DNA markers provide an opportunity for precisecharacterization of genotypes and measurement of geneticrelationships than other markers (Soller and Beckmann1983). In genus Lens genetic diversity have been assessedby using RFLPs (Havey and Muehlbauer 1989), RAPDand ISSR (Duran and Perez de la Vega 2004; Fikiru et al.2007), AFLP (Abo-el wafa et al. 1995; Sharma et al.1996) and SSRs (Hamwieh et al. 2009). Microsatelliteshave become the marker of choice in many crops forgenetic diversity studies because of their polymorphism,multiallelic nature, co-dominant inheritance, reproducibil-ity, relative abundance and genome coverage. In thepresent study, genetic diversity was surveyed in lentilgermplasm, released varieties and related species usingRAPD and SSRs.
Materials and methods
Eighty three genotypes including 23 wild, 5 exotic and 55indigenous collections of Lens species (Table 1) were usedin molecular diversity analysis using RAPD and micro-satellite markers.
Genomic DNA extraction
Total genomic DNA from 5 g of fresh young leaf tissue,collected from five random plants per accession, wasextracted following the cetyl trimethylammonium bromide(CTAB) method as described by Murray and Thompson1980. RNase treatment was given as suggested by Murrayand Thompson 1980. The purified DNA was quantified on0.8% agarose gel along with uncut lambda DNA (30 and60 ng).The total genomic DNA was diluted to 20 ng/μl forPCR analysis.
Primer selection and polymerase chain reaction
Twenty nine SSR primers reported by Hamwieh et al.2005 and 52 RAPD primers procured from Operon
Technologies Inc., USA (kits A, B, C, O, U, V) wereanalyzed. Eight SSRs and 15 RAPD primers among themproduced repeatable and clear bands and were used forfurther analysis (Table 2). The PCR profile for RAPDprimers consisted of : initial denaturation at 94°C for2 min, and subsequent 45 cycles, each with denaturationat 94°C for 1 min, primer annealing at 40°C for 1 minand primer extension at 72°C for 1 min. The finalextension step was performed at 72°C for 7 min. ThePCR cycling conditions for SSR primer were as follows:initial denaturation at 94°C for 3 min followed by 30cycles with 30 s at 94°C, 30 s at 52–58°C (dependingupon the primer), 1 min at 72°C with final extension at72°C for 5 min. Amplifications were carried out in 20 μlreaction mix containing 30 ng of template DNA, 10picomole of each primer, 0.2 mM dNTPs,1x PCR bufferand 1 unit Taq DNA polymerase (Bangalore Genei) forboth the markers. The amplified products with RAPDmarkers were resolved on 2% agarose gel and withmicrosatellite markers on 3% metaphor gel. A 100 bpladder was used for approximate sizing of the products.DNA samples were electrophoresed for 3 h at a constantvoltage of 100 V in 1X TBE buffer and photographedwith a CCD camera attached to a gel documentationsystem (Syngene).
Data analysis
Fragments amplified by primer sets were scored manuallyin terms of positions of the bands relative to the laddersequentially from the smallest to the largest-sized bands.Diffused bands or bands revealing ambiguity in scoring willbe considered as missing data and designated as ‘9’ incomparison with ‘1’ for the presence of a band and ‘0’ forthe absence of a band in the data matrix. A binary matrixwas then transformed to genetic similarity (GS) matrixusing Jaccard’s coefficient (Jaccard 1908). A dendrogrambased on similarity coefficients was prepared by usingUnweighted Pair Group Method with Arithmetic Mean(UPGMA) using statistical software NTSYS-PC 2.02(Rohlf 2000). Robustness of the clusters was evaluated bybootstrap analysis using WIN BOOT software (Yap andNelson 1996). Polymorphism information content (PIC)values for each band was calculated based on allelefrequencies according to Smith et al. (1997), as
PIC ¼ 1�PPi2, where Pi is the band frequency of the
ith allele. Gene diversity parameters and partitioning ofvariation between genotypes (GST) (equivalent to FST forbiallelic loci) was calculated with Popgene ver 1.32 (Yeh etal. 2000) assuming Hardy-Weinberg equilibrium. Analysisof molecular variance (AMOVA) (Excoffier et al. 2005)was used to estimate variance components among geno-types using Arlequin ver 3.01.
J. Plant Biochem. Biotechnol. (July–Dec 2012) 21(2):198–204 199

Table 1 Sources of accession of different Lens species used in molecular analysis
S.No. Genotypes Species Seed source
Wild Species
1 ILWL 4 Lens orientalis ICARDA, Syria
2 ILWL 72 Lens orientalis ICARDA, Syria
3 ILWL 74 Lens orientalis ICARDA, Syria
4 ILWL 76 Lens orientalis ICARDA, Syria
5 ILWL 77 Lens orientalis ICARDA, Syria
6 ILWL 80 Lens orientalis ICARDA, Syria
7 ILWL 94 Lens orientalis ICARDA, Syria
8 ILWL 115 Lens orientalis ICARDA, Syria
9 ILWL 117 Lens orientalis ICARDA, Syria
10 ILWL 144 Lens orientalis ICARDA, Syria
11 ILWL145 Lens orientalis ICARDA, Syria
12 ILWL147 Lens orientalis ICARDA, Syria
13 ILWL148 Lens orientalis ICARDA, Syria
14 ILWL150 Lens orientalis ICARDA, Syria
15 ILWL154 Lens orientalis ICARDA, Syria
16 ILWL171 Lens orientalis ICARDA, Syria
17 ILWL177 Lens orientalis ICARDA, Syria
18 ILL-6821 Lens orientalis ICARDA, Syria
19 ILWL485 Lens ervoides ICARDA, Syria
20 ILL1005 Lens orientalis ICARDA, Syria
21 ILWL356 Lens orientalis ICARDA, Syria
22 ILWL75 Lens orientalis ICARDA, Syria
23 ILWL200 Lens orientalis ICARDA, Syria
Indigenous lines
24 L 4076 Lens culinaris subsp. culinaris IARI, New Delhi
25 L 4147 Lens culinaris subsp. culinaris IARI, New Delhi
26 L 4594 Lens culinaris subsp. culinaris IARI, New Delhi
27 L 4596 Lens culinaris subsp. culinaris IARI, New Delhi
28 PL 01 Lens culinaris subsp. culinaris G.B.P.U.A&T. Pantnagar
29 PL 04 Lens culinaris subsp. culinaris G.B.P.U.A&T. Pantnagar
30 PL 05 Lens culinaris subsp. culinaris G.B.P.U.A&T. Pantnagar
31 PL 406 Lens culinaris subsp. culinaris G.B.P.U.A&T. Pantnagar
32 DPL 58 Lens culinaris subsp. culinaris IIPR Kanpur
33 DPL 62 Lens culinaris subsp. culinaris IIPR Kanpur
34 L 4378 Lens culinaris subsp. Culinaris IARI, New Delhi
35 L 4582 Lens culinaris subsp. culinaris IARI, New Delhi
36 PKVL 01 Lens culinaris subsp. culinaris PKV, Akola
37 Sehore 74-3 Lens culinaris subsp. culinaris JNKVV, Jabalpur
38 JL 03 Lens culinaris subsp. culinaris JNKVV, Jabalpur
39 L 5120 Lens culinaris subsp. culinaris IARI, New Delhi
40 L 4691 Lens culinaris subsp. culinaris IARI, New Delhi
41 PL 02 Lens culinaris subsp. culinaris GBPUA&T Pantnagar
42 L 7764 Lens culinaris subsp. culinaris IARI, New Delhi
Exotic lines and derivatives
43 EC-382703 Lens culinaris subsp. culinaris ICARDA, Syria
44 P 27240 Lens culinaris subsp. culinaris ICARDA, Syria
45 Precoz Lens culinaris subsp. Culinaris ICARDA, Syria
46 Bari masoor 4 Lens culinaris subsp. culinaris ICARDA, Syria
200 J. Plant Biochem. Biotechnol. (July–Dec 2012) 21(2):198–204

Results and discussion
Diversity analysis based on RAPD and SSR markers
Eighty three lentil genotypes produced a total of 92amplicons by using 15 RAPD primers, the percentagepolymorphism being 57% to 100%. Nine primers OPA01,OPA03, OPA04, OPA10, OPB01, OPC02, OPC18, OPC05and OPV17 exhibited 100% polymorphism. The PIC valueranged from 0.20 (OPA04) to 0.85 (OPA16) (Table 2). Total
average of polymorphic bands per primer was 5.6 whichwas higher than described by Abo-el wafa et al. (1995).Polymorphism generated by RAPD markers was lower incultivated varieties compared to wild ones as describedpreviously by other workers in lentil (Duran and Perez de laVega 2004; Abo-el wafa et al. 1995). Six polymorphicSSRs amplified 18 alleles and polymorphic informationcontent ranged from 0.10 to 0.78 (Table 2). Microsatelliteprimer SSR11 revealed 2 unique alleles (present in singlegenotype) (ILWL75 and L4147) and one rare allele (present
Table 1 (continued)
S.No. Genotypes Species Seed source
47 FLIP 96-51 Lens culinaris subsp. culinaris IARI, New Delhi
48 L 404 Lens culinaris subsp. culinaris IARI, New Delhi
49 L 830 Lens culinaris subsp. culinaris IARI, New Delhi
50 L 4583 Lens culinaris subsp. culinaris IARI, New Delhi
51 L 4602 Lens culinaris subsp. culinaris IARI, New Delhi
52 L 4605 Lens culinaris subsp. culinaris IARI, New Delhi
53 L 4618 Lens culinaris subsp. culinaris IARI, New Delhi
54 L 4649 Lens culinaris subsp. culinaris IARI, New Delhi
55 L 4661 Lens culinaris subsp. culinaris IARI, New Delhi
56 L 5125 Lens culinaris subsp. culinaris IARI, New Delhi
57 L 7434 Lens culinaris subsp. culinaris IARI, New Delhi
58 L 7774 Lens culinaris subsp. culinaris IARI, New Delhi
59 L 7820 Lens culinaris subsp. culinaris IARI, New Delhi
60 L 7903 Lens culinaris subsp. culinaris IARI, New Delhi
61 L 4908 Lens culinaris subsp. culinaris IARI, New Delhi
62 L 7917 Lens culinaris subsp. culinaris IARI, New Delhi
63 L 7920 Lens culinaris subsp. culinaris IARI, New Delhi
64 L 7931 Lens culinaris subsp. culinaris IARI, New Delhi
65 L 7933 Lens culinaris subsp. culinaris IARI, New Delhi
66 L 7939A Lens culinaris subsp. culinaris JNKVV, Jabalpur
67 JLS 01 Lens culinaris subsp. culinaris IARI, New Delhi
68 MC 06 Lens culinaris subsp. Culinaris PAU, Ludhiana
69 LL 884 Lens culinaris subsp. culinaris IARI, New Delhi
70 10-3-Y-26 Lens culinaris subsp. culinaris IARI, New Delhi
71 SKL 259 Lens culinaris subsp. culinaris IARI, New Delhi
72 MC 01 Lens culinaris subsp. culinaris IARI, New Delhi
73 LC 74-1-5-1 Lens culinaris subsp. culinaris IARI, New Delhi
74 L 4603 Lens culinaris subsp. culinaris IARI, New Delhi
75 Globe mutant Lens culinaris subsp. culinaris IARI, New Delhi
76 Fasciated mutant Lens culinaris subsp. culinaris IARI, New Delhi
77 L 3685 Lens culinaris subsp. culinaris IARI, New Delhi
78 L 358 Lens culinaris subsp. culinaris IARI, New Delhi
79 L 7828 Lens culinaris subsp. culinaris IARI, New Delhi
80 L 7830 Lens culinaris subsp. culinaris IARI, New Delhi
81 L 7916 Lens culinaris subsp. culinaris IARI, New Delhi
82 10-2-B-2 Lens culinaris subsp. culinaris IARI, New Delhi
83 RL 01 Lens culinaris subsp. Culinaris IARI, New Delhi
J. Plant Biochem. Biotechnol. (July–Dec 2012) 21(2):198–204 201

in <5% frequency). Wild lentil genotypes amplified locicomparable to those of cultivated species indicating theiramenability to PCR and microsatellite sets.
Variation between different genotypes was analyzed usingdifferent estimators-effective number of alleles/loci, percent-age of polymorphic loci, expected heterozygosity andshannon’s information index. RAPD marker showed 96.74%polymorphic loci, 0–0.49 heterozygosity range over all theloci (Table 3). The overall Gst which is a measure of genedifferentiation was found to be 0.30 for all the genotypes.Wild Lens genotypes seemed to be most diverse consideringall the estimators followed by released varieties. For micro-satellite markers, average number of polymorphic lociamong the genotypes was 90%. Shannon’s informationindex ranged from 0 to 0.69 and average heterozygositywas 0.24. Further analysis of molecular variance (AMOVA)among and within accessions revealed that 36.8% of thevariation resides among the genotypes and 63.1% betweenthe genotypes. The overall Fst value for all the loci was
calculated to be 0.31 which is comparable to the Gst values.Nei’s unbiased genetic distance revealed more distancebetween wild types and exotic lines with RAPD markersand between released varieties and exotic lines with SSRmarkers. However, genetic distance between paired popula-tions by AMOVA for the combined data was congruent withRAPD analysis. Released varieties were closer to wild typeLens (0.29) compared to exotic ones (0.38), which might be
Table 2 Polymorphic informa-tion content (PIC) and numberof scorable bands produced by15 RAPD and eight SSRsmarkers in 83 Lens genotypes
Tm Annealing temperature, NSBnumber of scorable bands, NPBnumber of polymorphic bands,%PB% polymorphic Bands,PIC–Polymorphism InformationContent
S. No. Primers Sequence Tm (°C) NSB NPB %PB PIC
1. OPA18 5′AGGTGACCGT 3′ 38 4 3 75 0.01
2. OPA01 5′CAGGCCCTTC3′ 38 4 4 100 0.24
3. OPA13 5′CAGCACCCAC3′ 38 6 5 83 0.66
4. OPB11 5′GTAGACCCGT3′ 38 7 4 57 0.55
5. OPA04 5′AATCGGGCTG3′ 38 3 3 100 0.20
6. OPB01 5′GTTTCGCTCC3′ 38 7 7 100 0.72
7. OPC02 5′GTGAGGCGTC3′ 38 7 7 100 0.73
8. OPA10 5′GTGATCGCAG3′ 38 9 9 100 0.58
9. OPC18 5′TGAGTGGGTG3′ 38 5 5 100 0.74
10. OPO11 5′GACAGGAGGT3′ 38 6 5 83 0.65
11. OPA03 5′AGTCAGCCAC3′ 38 5 5 100 0.34
12. OPV17 5′ACCGGCTTGT3′ 38 7 7 100 0.69
13. OPA16 5′AGCCAGCGAA3′ 38 12 11 91 0.85
14. OPU19 5′GTCAGTGCGG3′ 38 6 5 93 0.77
15. OPC05 5′GATGACCGCC3′ 38 4 4 100 0.65
16. LC16 5′GACTCCCAACTTGTATG3′ 53 1 0 0 03′GTACATTGAACAGCATCATC5′
17. LC02 5′CAAGCATGACGCCTATGAAG3′ 56 3 3 100 0.483′CTTTCACTCACTCAACTCTC5′
18. SSR17 5′GAGATTTCCAATACTGAATA3′ 52 2 2 100 0.333′TTCTACTTCTTCTTTCTTGA5′
19. SSR11 5′CAACCTCACTTACCTTAC3′ 52 4 4 100 0.043′GCTCTTTATCATCATTCTAC5′
20. SSR19 5′TGGTAGCGTAAAAAAGTGTC3′ 58 1 0 0 03′TTTAGTGCTGGTATGATCGC5′
21. SSR10 5′CACCAATCACCAACACAC3′ 54 4 2 50 0.783′GAGCTGTGAAGTCTTATCTG5′
22. SSR07 5′CCAGAACAAACGTAAACC3′ 52 2 2 100 0.423′CTATCGCATATGAGTGAAC5′
23. SSR04 5′GGTAGTGGTGAGGAATGAC3′ 55 3 2 66 0.103′GCATCACTGCAACAGACC5′
Table 3 Gene diversity parameters for RAPD and SSR markersamong 83 lentil genotypes
Parameters RAPD SSR
Percentage Polymorphic loci 96.74 90
Mean Nei Heterozygosity 0.29 0.24
Shanonn’s information index 0.44 0.37
Partitioning of variation using Gst 0.3 0.2
202 J. Plant Biochem. Biotechnol. (July–Dec 2012) 21(2):198–204

due to less number of markers used and hence insufficientcoverage of genomic region.
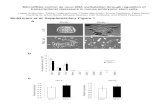
Genetic relationships among the accessions were studiedbased on cluster analysis (Fig. 1). Genetic similaritycoefficient values for both the markers ranged from 0.27(between ILWL 147 and RL 01) to 0.90 (between L 4649and L 4661) for all the genotypes. The critical evaluation ofdendrogram revealed three major clusters. Cluster I com-prised 21 wild accessions of Lens orientalis and 1 wildaccession Lens ervoides (ILWL485) inferring sufficientdiversity at genomic level among different Lens spp.Subcluster II A comprised of 19 released Indian varietieswhich pointed towards their low genetic base. This mightbe due to tendency to add desirable and improvedcharacters from few elite lines repeatedly used in breedingprogramme resulting in narrow genetic diversity. SubclusterII B consisted of 41 genotypes including 5 exoticgermplasm (EC-782703, P27240, Precoz, Bari masoor 4and FLIP 96–51) and 36 advanced breeding lines/exoticderivatives. However, Cluster III had only one genotype oforientalis (ILWL 200) exhibiting only 45% similarity withcluster I and II. The cophenetic correlation betweensimilarity matrix and cluster analysis based on combineddata represented a very high goodness of fit (r=0.9).Numbers at branch point in Fig. 1 indicated support forgenotypes clustered to the right of the number, values arepercent of bootstrap samples that exhibited the cluster. Highbootstrap value meant strength or high precision ofclustering for example ILL 177 and ILL 200 grouped with
85% precision and MC1 and globe mutant clustered with72% strength.
Although RAPD and SSR markers have been used byseveral workers to assess genetic relationship in lentil, thesuccess of our study in identifying polymorphism is due tothe use of a number of randomly selected prescreenedhighly informative markers. Geographically isolated pop-ulations accumulate genetic differences as they acclimatizethemselves to different environment. The clustering ofgenotypes within group was not similar when RAPD andSSR derived dendrograms were compared. These differ-ences may be attributed to marker sampling error and/or thelevel of polymorphism detected, importance of the numberof loci and their coverage of overall genome. Theaccessions with most distinct DNA profile are likely tocontain the greatest number of novel alleles. The advancedbreeding lines used in this study are derived from exoticlines and shared more alleles with exotic varieties comparedto past released varieties with narrow base indicating thealternate preference of breeders to select exotic genes.
In the present study the level of inter and intra specificdiversity and species relationship among different cultivatedand wild genotypes of Lens species were assessed. Differen-tiation of wild and released varieties into separate clustersindicated clear-cut distinction at genomic level and groupingof released cultivars in one cluster depicting their narrowgenetic base. ICARDA has found sources of resistance tovascular wilt and ascochyta blight, and cold tolerance in thecrossable wild progenitor L. culinaris spp. orientalis
Fig. 1 Dendrogram obtained by analysis of combined data of RAPDand SSR. G1-83 represents the same serial number of genotypes asgiven in Table 1; numbers at branch point indicate support for
genotypes clustered to the right of the number, values are percent ofbootstrap samples that exhibited the cluster
J. Plant Biochem. Biotechnol. (July–Dec 2012) 21(2):198–204 203

(Ferguson 2000). L. culinaris spp. orientalis is crossablewith the cultivated lentil. Therefore for further broadeningof the genetic base intra-specific hybridization involvingearly maturing Mediterranean germplasm lines and indig-enous agronomic bases and inter-specific hybridization arewarranted for successful lentil improvement programme.
Acknowledgements The authors are extremely thankful to Dr. K.V.Prabhu, Head, Division of Genetics, Dr. (Mrs.) Shanti Chandrashekaran,Professor, Division of Genetics and Dr. T.R. Sharma, Principal Scientist,NRC (PB) for their kind support and guidance.
References
Abo-el wafa A, Murai K, Shimanda T (1995) Intra and Inter-specificvariation in Lens revealed by RAPD markers. Theor Appl Genet90:335–340
Barulina H (1930) Lentils of the USSR and other countries. TrudyPrikl Bot Seleck Suppl 40:1–319
Duran Y, Perez de la Vega M (2004) Assessment of genetic variationand species relationships in a collection of Lens using RAPD andISSR. Spanish J Agric 2:538–544
Excoffier I, Laval LG, Schneider S (2005) Arlequin ver 3.01: Anintegrated software package for population genetics data analysis.Evol Bioinform Online 1:47–50
Ferguson M (2000) Lens spp. conserved resources, priorities forcollection and future prospects. In: Knight R (ed) Proceedings ofThird International Food Legume Research Conference (IFLRCIII): linking research and marketing opportunities for pulses inthe 21st Century. Adelaide Australia, 22–26 September 1997.Kluwer Academic Publishers, Dordrecht, pp 613–620
Fikiru E, Tesfaye T, Bekele E (2007) Genetic diversity and populationstructure of Ethiopian lentil (Lens culinaris Medikus) landracesas revealed by SSR markers. African J Biotech 6(12):1460–1468
Hamwieh A, Udupa SM, Choumane W, Sarker A, Dreyer F, Jung C,Baum M (2005) A genetic linkage map of lens species based onmicrosatellite and AFLP markers and the localization of fusariumvascular wilt resistance. Theor Appl Genet 110:669–677
Hamwieh A, Udupa S, Sarker A, Jung C, Baum M (2009)Development of new microsatellite markers and their applicationin the analysis of genetic diversity in lentils. Breed Sci 59:77–86
Havey MJ, Muehlbauer FJ (1989) Linkages between restrictionfragment length, isozyme, and morphological markers in lentil.Theor Appl Genet 77:395–401
Jaccard P (1908) Nouvelles recherces sur la distribution florale. BullSoc Vaudoise Sci Nat 44:223–270
Murray MG, Thompson WF (1980) Rapid isolation of high molecularweight plant DNA. Nucleic Acids Res 8:4321–4325
Paterson AH, Tanksley SD, Sorrells ME (1991) DNA markers in plantimprovement. Adv Agron 46:39–90
Rohlf FJ (2000) NTSYS-pc: numerical taxonomy and multivariateanalysis system. Version 2.1 Exeter Publications, New York
Sharma SK, Knox MR, Ellis THN (1996) AFLP analysis of diversityand phylogeny of Lens and its comparison with RAPD analysis.Theor Appl Genet 93:751–758
Smith JSC, Chin ECL, Shu H (1997) An evaluation of the utility ofSSR loci as molecular markers in maize (Zea mays L.):comparisons with data from RFLP and pedigree. Theor ApplGenet 95:163–173
Soller M, Beckmann JS (1983) Genetic polymorphism in varietalidentification and genetic improvement. Theor Appl Genet67:25–33
Yap IV, Nelson R (1996) WinBoot: a program for performingbootstrap analysis of binary data to determine the confidencelimits of UPGMA-based dendrograms. IRRI Discussion PaperSeries No 14, IRRI, Los Banos, Philippines
Yeh FC, Yang R, Boyle TJ, Ye Z, Xiyan M (2000) PopGene 32,Microsoft window based freeware for population genetic analysis,version 1.32. University of Alberta, Edmonton, Canada
Zohary D (1972a) The wild progenitor and place of origin of thecultivated lentil Lens culinaris. Econ Bot 26:176–183
Zohary D (1972b) The wild progenitor and place of origin of thecultivated lentil Lens culinaris. Econ Bot 26:326–336
204 J. Plant Biochem. Biotechnol. (July–Dec 2012) 21(2):198–204








![289 ' # '6& *#2 & 7and increased 11 -HSD2 mRNA expression) markers of syncytiotrophoblast differentiation. ... (PPAR ) which is regulated by VEGF-A expression [59, 60]. In addition,](https://static.fdocuments.in/doc/165x107/60f700cb21111f656f07cf7c/289-6-2-7-and-increased-11-hsd2-mrna-expression-markers-of-syncytiotrophoblast.jpg)








