
Functional and immunological characterization of the stimulation of human lymphocytes with a mitogen...
9
CELLULAR IMMUNOLOGY 68, 146-154 (1982) Functional and Immunological Characterization of the Stimulation of Human Lymphocytes with a Mitogen from Group A Streptococci (SM) FRANCOQUAGLIATA,* SUSAN BECKERDITE-QUAGLIATA,* RICHARD A. ALSOBROOK,* CARLO Russo,? FRANCESCO INDIVERI,~ MICHELE A. PELLEGRINO,~AND SOLDANOFERRONE? *Department of Medicine, University of TennesseeCenter for the Health Sciences, Memphis. Tennessee38163, and TDepartments of Pathology and Surgery, College of Physicians and Surgeons, Columbia University, New York, New York 10032 Received November 15, 1981: accepted January 29, 1982 Streptococcal mitogen (SM), an extracellular product of group A streptococci, is non spe- cifically mitogenic for both B and T lymphocytes. The mitogenic activity of SM is resistant to digestion with trypsin, to heating at 100°C for 5 min, and to treatment with dithiothreitol. The proliferative response of lymphocytes from patients with a history of rheumatic fever is similar to that of lymphocytes from healthy donors when stimulated with optimal concentra- tions of SM. but is significantly reduced when low doses of SM are used. T lymphocytes stimulated with SM acquire Ia antigens and the ability to stimulate allogeneic and autologous lymphocytes in mixed lymphocyte reactions. An involvement of Ia antigens in these reactions is indicated by the specific block by monoclonal antibodies to human Ia antigens. INTRODUCTION The role of the streptococcus in the pathogenesis of rheumatic fever (RF) and rheumatic carditis has stimulated interest in the characterization of streptococcal components and of their interaction with the immune system. Our work has focused on the study of a product of group A streptococci which is mitogenic for human lymphocytes. This product, which is referred to as streptococcal mitogen (SM), was initially separated from streptolysin S (SS) preparations (1, 2) with which it had been identified (3). The proliferative responseto SM has been recently reported to be controlled by a gene closely linked to the major histocompatibility complex (MHC) (4, 5) and to be blocked by sera containing antibodies to Ia antigens (6). These antigens which share functional and structural properties with murine I-E subregion antigens have a restricted tissue distribution (7): among lymphocyte subpopulations Ia antigens are expressed on B cells, but are not detectable on most resting T cells (8-21). T cells may acquire Ia antigens following stimulation with mitogens or antigens (8, 19). The appearance of Ia antigens on activated T lym- phocytes is associatedwith the ability of the activated T cells to stimulate autologous and allogeneic T lymphocytes in mixed lymphocyte reactions (MLRs) ( 16, 17, 20). 146 000%8749/82/050146-09$02.00/O Copyright 8 1982 by Academic Press. Inc. All rights of reproduction in any form rcsctved.
-
Upload
franco-quagliata -
Category
Documents
-
view
214 -
download
2
Transcript of Functional and immunological characterization of the stimulation of human lymphocytes with a mitogen...

CELLULAR IMMUNOLOGY 68, 146-154 (1982)
Functional and Immunological Characterization of the Stimulation of Human Lymphocytes with a Mitogen from Group A
Streptococci (SM)
FRANCO QUAGLIATA,* SUSAN BECKERDITE-QUAGLIATA,* RICHARD A. ALSOBROOK,* CARLO Russo,? FRANCESCO INDIVERI,~
MICHELE A. PELLEGRINO,~AND SOLDANO FERRONE?
*Department of Medicine, University of Tennessee Center for the Health Sciences, Memphis. Tennessee 38163, and TDepartments of Pathology and Surgery, College of Physicians and
Surgeons, Columbia University, New York, New York 10032
Received November 15, 1981: accepted January 29, 1982
Streptococcal mitogen (SM), an extracellular product of group A streptococci, is non spe- cifically mitogenic for both B and T lymphocytes. The mitogenic activity of SM is resistant to digestion with trypsin, to heating at 100°C for 5 min, and to treatment with dithiothreitol. The proliferative response of lymphocytes from patients with a history of rheumatic fever is similar to that of lymphocytes from healthy donors when stimulated with optimal concentra- tions of SM. but is significantly reduced when low doses of SM are used. T lymphocytes stimulated with SM acquire Ia antigens and the ability to stimulate allogeneic and autologous lymphocytes in mixed lymphocyte reactions. An involvement of Ia antigens in these reactions is indicated by the specific block by monoclonal antibodies to human Ia antigens.
INTRODUCTION
The role of the streptococcus in the pathogenesis of rheumatic fever (RF) and rheumatic carditis has stimulated interest in the characterization of streptococcal components and of their interaction with the immune system. Our work has focused on the study of a product of group A streptococci which is mitogenic for human lymphocytes. This product, which is referred to as streptococcal mitogen (SM), was initially separated from streptolysin S (SS) preparations (1, 2) with which it had been identified (3). The proliferative response to SM has been recently reported to be controlled by a gene closely linked to the major histocompatibility complex (MHC) (4, 5) and to be blocked by sera containing antibodies to Ia antigens (6). These antigens which share functional and structural properties with murine I-E subregion antigens have a restricted tissue distribution (7): among lymphocyte subpopulations Ia antigens are expressed on B cells, but are not detectable on most resting T cells (8-21). T cells may acquire Ia antigens following stimulation with mitogens or antigens (8, 19). The appearance of Ia antigens on activated T lym- phocytes is associated with the ability of the activated T cells to stimulate autologous and allogeneic T lymphocytes in mixed lymphocyte reactions (MLRs) ( 16, 17, 20).
146
000%8749/82/050146-09$02.00/O Copyright 8 1982 by Academic Press. Inc. All rights of reproduction in any form rcsctved.

STREPTOCOCCAL MITOGEN AND HUMAN LYMPHOCYTES 147
TABLE 1
Kinetics of Stimulation” of Blood Lymphocytes from Healthy Tuberculin-Positive Subjects Cultured with SM, PHA, Con A, PWM, and PPD
Donor Incubation time (hr) Medium
[ “C]Thymidine incorporation
SM PHA Con A PWM PPD
A.I. 42 243’ 9,731 13,026 10,640 8,561 483 120 428 29,412 31,765 28,182 19,859 9822
P.L. 42 316 11,072 12,154 9,931 10,198 369 120 497 37,641 34,119 24,156 18,434 7689
L? Stimulation of lymphocytes was determined by their uptake of [‘%]thymidine. b Figures indicate average cpm values of triplicate cultures.
In this study we have characterized the mitogenic activity of SM for lymphocyte subpopulations and for lymphocytes from patients with a history of RF. Further- more, since Ia antigens are believed to mediate and restrict the cellular interactions required to generate immune responses (2 1 ), we have tested T lymphocytes stim- ulated with SM for expression of Ia antigens and analyzed some of their immu- nological properties.
MATERIALS AND METHODS
Mitogens and enzymes. The SM preparation used was an ammonium sulfate precipitate of the supernatant of a culture of strain C203-S group A streptococci grown as described (22). When the material was subjected to ion-exchange chro- matography on CM cellulose equilibrated with 10 mM acetate (pH 5), the mito- genie activity eluted at 0.1 M NaCl concentration. The isoelectric point of this component is 6.9-7.0. Purified PHA (Wellcome, Bechkenham, England) was used in 0.1 -ml aliquots of 2 mg/ml original product at lOO- or lOOO-fold dilutions. Concanavalin A (Con A) (Sigma Chemical Co., St. Louis, MO.) was used in 20 and 40 pm/ml amounts. Pokeweed mitogen (PWM) (Difco, Detroit, Mich.) was used in 40 pg/ml amounts. PPD (Parke Davis, Detroit, Mich.) was used in 10 pg/ml amounts. Trypsin and trypsin inhibitor (Worthington Biochemical Corpo- ration, Freehold, N.J.) chymotrypsin (Sigma), and dithiothreithol (Sigma) were used in excess (100: 1) to treat the SM.
Lymphocytes. Lymphocytes were isolated by differential centrifugations on Fi- toll-Hypaque mixtures from blood drawn in heparinized tubes from healthy donors, various groups of patients, and at time of delivery. The mononuclear cells recovered at the plasma and Ficoll-Hypaque interface were washed twice with PBS, and resuspended to a concentration of lo8 cells/ml.
Monoclonal antibodies and polyclonal xenoantisera. The monoclonal antibody (MoAb) Q1/28 to HLA-A,B antigens and the MoAb Q5/13 to Ia antigens were prepared and characterized as described (23, 24). Antisera to Ia antigens were developed in rabbits with Ia antigens bound to insolubilized xenoantibodies to Ia antigens (25). An anti-human ,&-microglobulin (p+) xenoantiserum was from a rabbit immunized with purified urinary pZ-+
Rabbit antisera to human and mouse Ig were prepared by standard procedures

148 QUAGLIATA ET AL.
and antibodies to human and mouse Ig were purified by affinity chromatography on immunoadsorbent columns (26).
Rosette assays to detect sheep red blood cell (SRBC) receptors, Mblg, and la antigens. Receptors for SRBC were detected by a rosette assay with SRBC treated with neuraminidase (N) (Sigma) or with 2-aminoethylisothiouronium ( AET) (27). The rosette microassay with SRBC chemically coated with purified anti-Ig xe- noantibodies was used to detect MbIg and Ia antigens (26).
Isolation of B and T lymphocytes. B- and T-lymphocyte subpopulations were isolated by filtration through a nylon-wool column or by rosetting with AET- or N-treated SRBC (27).
Lymphocyte cultures. Lymphocytes (1 X 105) were cultured in 250-~1 aliquots of medium RPM1 1640 containing 20% FCS and desired amount of mitogen. Following 3 days of incubation at 37°C in a 5% CO* atmosphere, [‘“Cl- or [3H]thymidine (0.03 and 1.0 Ci, respectively) was added and incubation was con- tinued for an additional 16 hr. Cells were then harvested on glass fiber filter paper with a Model 24 cell harvester (Brandel, Gaithersburg, Md.). The washed and dried filters were mixed with a liquid scintillation cocktail and then counted in a liquid @-scintillation counter. Each culture was carried out in triplicate. Results are expressed as the difference in counts per minute between the means of triplicate mitogen-stimulated and control cultures. MLRs were performed as previously de- scribed (28). Briefly, responding cells ( 1 X 10’) were cultured with irradiated (2000 rad) stimulating cells in 250 ~1 of medium RPM1 1640 with 20% fetal calf serum. Following 5 days of culture at 37°C in a 5% CO* atmosphere, [3H Jthymidine (1 &i) was added and incubation continued for an additional 16 hr. Cells were then harvested on glass fiber and processed as described above.
RESULTS
Mitogenic activity of SM for lymphocytes from healthy donors. The kinetics of the blastogenic response to SM were determined with lymphocytes from two tuberculin-positive, otherwise normal, donors and compared with those of PHA, Con A, PWM, and PPD (Table 1). Following 42 hr of culture, there is significant response to SM and to the other mitogens, but not to PPD. However, significant response to PPD occurs after 5 days of culture. These results suggest that the response to SM is of the mitogen type and this conclusion is supported by the ability of SM to stimulate cord blood lymphocytes to an extent similar to that of lymphocytes obtained from the respective parents (Table 2). The mitogenic activity
TABLE 2
Stimulation” of Cord Blood Lymphocytes and Parental Blood Lymphocytes Cultured with SM
Lymphocytes Individuals
tested
[ %]Thymidine incorporation (cpm)
Mean Range
Cord blood 16 42,763 25,189-56,350 Maternal 16 39,836 29.664-5 1,892 Paternal 5 47,235 3 1,678-62,429
’ Stimulation of lymphocytes was determined by their uptake of [14C]thymidine.

STREPTOCOCCAL MITOGEN AND HUMAN LYMPHOCYTES 149
I 4x103 4X' j 1x103 1x107 1x103 1x103
Mitogen Concentration ( pg /ml )
FIG. I. Proliferative response of lymphocytes from normal donor after 96 hr stimulation with SM (right panel) and PHA (left panel). Stimulation of responder cells was determined by their uptake of [ Wlthymidine.
of SM is dose dependent, the optimal mitogenic concentration at 96 hr of culture being in the range of 100 to 0.1 pg/ml. Results of a representative experiment are shown in Fig. 1.
Treatment of SM with an excess of trypsin (100: 1) for 30 min at 37°C followed by trypsin inhibitor only reduces the mitogenic activity by 20% while treatment with chymotrypsin under the same conditions enhances the mitogenic activity by approximately 20%; heating at 100°C for 5 min and treatment with dithiothreitol for 60 min at 37°C produces a reduction in mitogenic activity of only 10%.
Proliferative response of lymphocytes to SM does not require the presence of the mitogen in the culture throughout the incubation period; maximal response is ob-
AL :. : 4x103
Mitogen Concentration ( Pg /ml)
FIG. 2. Proliferative response of lymphocytes from 12 subjects with a history of rheumatic fever (m) and 8 age-matched controls (0) stimulated with SM (right panel) and PHA (left panel). Stimulation of responder cells was determined by their uptake of [ “C]thymidine.
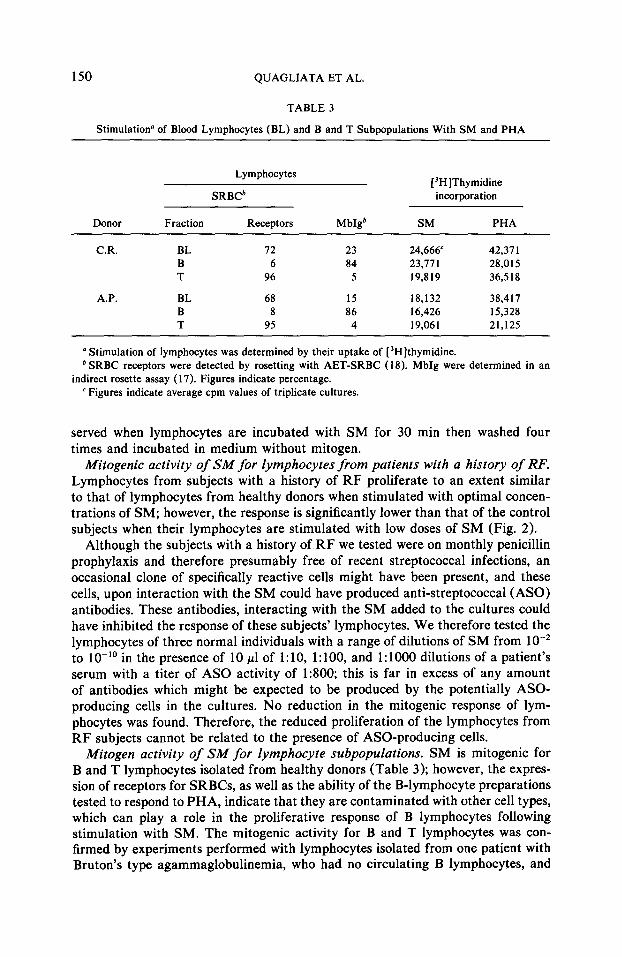
150 QUAGLIATA ET AL.
TABLE 3
Stimulation” of Blood Lymphocytes (BL) and B and T Subpopulations With SM and PHA
Lymphocytes
SRBCb [“H]Thymidine incorporation
Donor Fraction Receptors Mb@ SM PHA
CR. BL 72 23 24,666 42,371 B 6 84 23,77 1 28,015 T 96 5 19,819 36,518
A.P. BL 68 15 18,132 38,417 B 8 86 16,426 15,328 T 95 4 19,061 21,125
a Stimulation of lymphocytes was determined by their uptake of [‘Hlthymidine. b SRBC receptors were detected by rosetting with AET-SRBC (18). MbIg were determined in an
indirect rosette assay ( 17). Figures indicate percentage. ’ Figures indicate average cpm values of triplicate cultures.
served when lymphocytes are incubated with SM for 30 min then washed four times and incubated in medium without mitogen.
Mitogenic activity of SM for lymphocytes from patients with a history of RF. Lymphocytes from subjects with a history of RF proliferate to an extent similar to that of lymphocytes from healthy donors when stimulated with optimal concen- trations of SM; however, the response is significantly lower than that of the control subjects when their lymphocytes are stimulated with low doses of SM (Fig. 2).
Although the subjects with a history of RF we tested were on monthly penicillin prophylaxis and therefore presumably free of recent streptococcal infections, an occasional clone of specifically reactive cells might have been present, and these cells, upon interaction with the SM could have produced anti-streptococcal (ASO) antibodies. These antibodies, interacting with the SM added to the cultures could have inhibited the response of these subjects’ lymphocytes. We therefore tested the lymphocytes of three normal individuals with a range of dilutions of SM from lo-* to 10-l’ in the presence of 10 ~1 of I : 10, 1: 100, and 1: 1000 dilutions of a patient’s serum with a titer of AS0 activity of 1:800; this is far in excess of any amount of antibodies which might be expected to be produced by the potentially ASO- producing cells in the cultures. No reduction in the mitogenic response of lym- phocytes was found. Therefore, the reduced proliferation of the lymphocytes from RF subjects cannot be related to the presence of ASO-producing cells.
Mitogen activity of SM for lymphocyte subpopulations. SM is mitogenic for B and T lymphocytes isolated from healthy donors (Table 3); however, the expres- sion of receptors for SRBCs, as well as the ability of the B-lymphocyte preparations tested to respond to PHA, indicate that they are contaminated with other cell types, which can play a role in the proliferative response of B lymphocytes following stimulation with SM. The mitogenic activity for B and T lymphocytes was con- firmed by experiments performed with lymphocytes isolated from one patient with Bruton’s type agammaglobulinemia, who had no circulating B lymphocytes, and

STREPTOCOCCAL MITOGEN AND HUMAN LYMPHOCYTES 151
TABLE 4
Effect of B/T Cell Ratio on Stimulation” of Lymphocytes Cultured with SM, PHA, and PWM
Donor Disease
type
Lymphocytes [ ?Z]Thymidine incorporation
Bb Tb SM PHA PWM
J.A. Bruton’s 0 82 46,823’ 39,462 29.98 1 H.G. CLL 86 11 43,568 25,638 32,736 E.R. CLL 95 0.5 39,431 19,251 35,165
u Stimulation of lymphocytes was determined by their uptake of [ “C]thymidine. * SRBC receptors were detected by rosetting with N-SRBC (18). MbIg were determined in an indirect
rosette assay ( 17). Figures indicate percentage. ’ Figures indicate average cpm values of triplicate cultures.
from two patients with chronic lymphocytic leukemia who had 86 and 95% cir- culating B lymphocytes, respectively. After 4 days in culture, irrespective of their B/T ratios, the lymphocytes from these three patients were stimulated by SM to levels higher than those obtained with PHA and PWM (Table 4).
Appearance of la antigens on T lymphocytes stimulated with SM. Less than 105 of nylon-wool column-purified T lymphocytes react with the MoAb QS/ 13 to human Ia antigens in an indirect rosette microassay. Following 3 days of culture with SM, up to 86% of these T cells become reactive with the MoAb Q5/ 13 (Table 5). The reactivity of SM-stimulated T lymphocytes with the MoAb Q5/13 is spe- cific since it is blocked by coating T lymphocytes with rabbit antisera to human Ia antigens, but not with rabbit anti-human &-microglobulin antisera (results not shown). No significant change was found in the percentage of cells binding either SRBC, anti-human Ig xenoantisera, anti-HLA-A,B MoAb Q1/28 (Table 5). Cul- turing nylon-wool column-purified T lymphocytes without SM for 3 days did not significantly change the percentage of cells reacting with the MoAb Q5/ 13, with the MoAb Q1/28, with anti-human Ig xenoantisera, or rosetting with AET-SRBC.
TABLE 5
Expression of Cell Surface Markers on Human Purified T Lymphocytes Cultured with SM for 72 hr
Percentage of cells positive for”
Donor
SF. L.D. N.G.
SRBC receptors
95 98 90
94.3 f 9.1b (90.2 f 7.5)
MbIg
5 6 3
4.6 rtr 2.1 (2.7 f 1.2)
Ia-like antigens
86 75 81
80.6 f 8.9 (3.4 k 1.7)
HLA-A, B antigens
98 91 99
91.6 + 9.8 (98.1 f 3.3)
’ SRCB receptors were detected by rosetting with AET-SRBC (18). MbIg, Ia-like antigens, and HLA- A, B antigens were determined in an indirect rosette assay (17) with the appropriate indicator cells.
b Mean + SD. Figures in parentheses indicate mean + SD values for T lymphocytes cultured for 72 hr in absence of SM.

152 QUAGLIATA ET AL.
FIG. 3. Stimulatory activity in allogeneic and autologous mixed lymphocyte reactions of T lymphocytes cultured with SM. Nylon-wool-purified T lymphocytes from donor B.C. (upper panels) and donor C.R. (lower panels), cultured for 72 hr in absence (left panels) and presence (right panels) of SM, were irradiated with 2000 rad and then used as stimulators in unidirectional MLRs with freshly purified T lymphocytes from donor CR. MLRs were performed in RPM1 1640 medium containing either fetal calf serum (O), HLA-A, B, C, MoAb, Q1/28 (a), MoAb Q5/13 to Ia antigens (k@, or the xenoantiserum No. 8802 to Ia antigens (m). Stimulation of responder cells was determined by their uptake of [ ‘Hlthymidine.
Ability of la antigen-bearing T lymphocytes to stimulate allogenic and autol- ogous lymphocytes in MLRs. T lymphocytes cultured for 3 days in the presence of SM were washed five times and tested for their ability to act as stimulators in MLRs. Figure 3 shows that SM-stimulated T lymphocytes could induce prolifer- ation of both autologous and allogeneic lymphocytes in MLRs. An involvement of Ia antigens in these reactions is indicated by the significant reduction in the extent of stimulation when the MoAb Q5/ 13 or the xenoantiserum No. 8802 to Ia antigens was added to the cultures. This blocking is specific since addition of the anti-HLA- A, B MoAb Q1/28 to the cultures did not have any significant effect (Fig. 3).
DISCUSSION
Our study shows that SM is a nonspecific mitogen for both B and T human lymphocytes, that the proliferative response of lymphocytes from subjects with a history of RF is decreased, and that SM induces the expression of immunologically functional Ia antigens on T lymphocytes.
Several extracellular streptococcal products have been reported to be mitogenic for lymphocytes (29-32). SM bears some similarities with the blastogen from group A streptococci, however, the latter is sensitive to heat and proteolytic enzyme treatment (30). SM is not pyrogenic (33). Hirshhorn et al. (3) reported that a decreased proliferative response of lymphocytes to an optimal dose of SS was pathognomonic for the first attack of RF, but this finding was not confirmed by Cuppari et al. (34). The present study supports the possibility of abnormalities of

STREPTOCOCCAL MITOGEN AND HUMAN LYMPHOCYTES 153
the mitogenic activity of SM for lymphocytes from patients with a history of RF since they display a reduced proliferative response to suboptimal doses of SM.
An interesting aspect of our study is that T lymphocytes stimulated with SM, like those stimulated with PHA (8, 20) express Ia antigens and acquire the ability to stimulate autologous lymphocytes. One might argue that this stimulation is mediated by residual SM bound to T lymphocytes. However, we believe this mech- anism to be unlikely, since the stimulation is specifically blocked by monoclonal antibodies and conventional xenoantisera to human Ia antigens. The latter reagents are more effective than the former ones, as already observed with PHA-activated T lymphocytes (20). This difference, if not caused by contaminating antibodies present in the rabbit antiserum, may reflect a more extensive coating of Ia antigen- bearing molecules by the various antibody populations present in the rabbit anti- serum than by the monoclonal antibody. At any rate, the results of the blocking experiments suggest that the newly expressed Ia antigens on activated T cells play a role in autologous MLRs. It has been proposed that blocking of MLRs by an- tibodies occurs through an antibody-dependent cell-mediated cytotoxicity ( ADCC) mechanism (35). Therefore, it could be postulated that the MoAb Q5/13 to Ia antigens block MLRs by mediating ADCC. However, the lack of blocking of MLRs by the anti-HLA-A,B MoAb Q1/28, which belongs to the same immunoglobulin class as the MoAb Q5/13 to Ia antigens, argues against this possibility.
The nature of the cells with stimulatory activity in MLRs in the populations of T cells stimulated with SM is worth discussing in view of the recent findings of Shen et al. (36). These authors used MoAb to T-cell subsets and to monocytes to analyze the antigenic profile of lymphocytes with receptors for SRBCs: these lym- phocytes acquire the ability to stimulate in autologous and allogeneic MLRs fol- lowing activation with allogeneic lymphocytes. They concluded that the stimulatory activity is displayed by cells which express Ia antigens and do not react with MoAb to monocytes and to T-cell subsets. Whether this conclusion applies also to our data cannot be assessed at this time since we have not analyzed the SM-stimulated T lymphocytes with MoAb to T-cell subsets. Furthermore, we have isolated T lymphocytes by nylon-wool filtration (27), while Shen et al. (36) isolated them by rosetting with SRBCs.
It has been reported that T lymphocytes may acquire Ia antigens in vivo following exposure to an appropriate antigen (19). If this occurs following streptococcal infections, our in vitro findings suggest that Ia antigen-bearing T cells may play a role in the immunopathological events which can follow a streptococcal infection. The system we have described may be a useful in vitro model to analyze these immunopathological events and to develop therapeutic approaches.
ACKNOWLEDGMENTS
This work was supported by the National Institutes of Health Grants DA 01965, AI 19189, CA 24329, CA 32609, CA 32634 CA 32635, and a Research Career Development Award (M.A.P).
REFERENCES
1. Taranta, A., Cuppari, G., and Quagliata, F., J. Exp. Med. 129, 605, 1969. 2. Taranta, A., Cuppari, G., and Quagliata, F., Nafure (London) 219, 757, 1968. 3. Hirschhorn, K., Schreibman, R. R., Verbo, S., and Grushkin, R. H., Proc. Nat. Acad. Sci. C/S,4
52, 1151, 1964.

154 QUAGLIATA ET AL.
4. Greenberg, L. J., Bradley, P. W., Chopyk, R. L., and Lalouel, J. M., Immunogenetics 11, 145, 1980. 5. Greenberg, L. J., Chopyk, R. L., Bradley, P. W., and Lalouel, J. M., Immunogenetics 11, 161, 1980. 6. Greenberg, L. J., Chopyk, R. L., Noreen, H., Gray, E. D., Yunis, E. J., and Ferrara, G. V., VOX
Sang. 34, 136, 1978. 7. Ferrone, S., Allison, J. P., and Pellegrino, M. A., Contemp. Top. Mol. Immunol. 7, 239, 1978. 8. Fu, S. M., Chiorazzi, N., Wang, C. Y., Montazeri, G., Kunkel, H. G., Ko, H. S., and Gottlieb,
A. B., J. Exp. Med. 148, 1423, 1978. 9. Reinherz, E. L., Kung, P. C., Pesando, J. M., Ritz, J., Goldstein, G., and Schlossman, S. F., J.
Exp. Med. 150, 1472, 1979. IO. DeWolf, W. C., Schlossman, S. F., and Yunis, E. J., J. Immunol. 122, 1780, 1979. 11. Greaves, M. F., Verbi, W., Festenstein, H., Papasteriadis, C., Jaraquemada, D., and Hayward, A.,
Immunology 9, 356, 1979. 12. Evans, K. L., Faldetta, T. J., Humphreys, R. E., Pratt, D. M. Yunis, E. J., and Schlossman, S. F.,
J. Exp. Med. 148, 1440, 1978. 13. Suciu-Foca, N., Susirmo, E., McKinnan, P., Rohowsky, C., Weiner, J., and Rubinstein, P., Trans-
plant. Proc. 10, 845, 1978. 14. Metzgar, R. S., Beitoglio, J., Anderson, J. K., Bonnard, G. D., and Ruscetti, F. W., J. Immunol.
122, 949, 1979. 15. Ko. H. S., Fu, S. M., Winchester, R. J., Yu, D. T. Y., and Kunkel, H. G., J. Exp. Med. 150, 246,
1979. 16. Engelman, E. G., Benike, C. J., and Charron, D. J., J. Exp. Med. 152, 114S, 1980. 17. Indiveri, F., Wilson, B. S., Russo, C., Quaranta, V., Pellegrino, M. A., and Ferrone, S., J. Immunol.
125, 2673, 1980. 18. Yu, D. T. Y., McCune, J. M., Fu, S. M., Winchester, R. J., and Kunkel, H. G., J. Exp. Med. 152,
89S, 1980. 19. Yu, D. T. Y., Winchester, R. J., Fu, S. M., Gibofsky, A., Ko, H. S., and Kunkel, H. G., J. Exp.
Med. 151, 91, 1980. 20. Russo, C., Indiveri, F., Quaranta, V., Molinaro, G. A., Pellegrino, M. A., and Ferrone, S., Im-
munogenetics 12, 267, 1981. 21. Moller, G. (Ed.), Transplant. Rev. 30, 1, 1976. 22. Quagliata, F., and Taranta, A., Ann. Rheum. Dis. 31, 500, 1972. 23. Quaranta, V., Walker, L. E., Ruberto, G., Pellegrino, M. A., and Ferrone, S., Immunogenetics 13,
285, 1981. 24. Quaranta, V., Walker, L. E., Pellegrino, M. A., and Ferrone, S., J. Immunol. 125, 1421, 1980. 25. Wilson, B. S., Indiveri, F., Pellegrino, M. A., and Ferrone, S., J. Immunol. 122, 1967, 1979. 26. Indiveri, F., Wilson, B. S., Pellegrino, M. A., and Ferrone, S., J. Immunol. Methods 29, 101, 1979. 27. Indiveri, F., Huddlestone, J., Pellegrino, M. A. and Ferrone, S., J. Immunol. Methods 34, 107,
1980. 28. Hartzman, R. J., Segall, M., Bach, M. L. and Bach, F. H., Transplantation 11, 268, 1971. 29. Plate, G. M., and Amos, B., Cell. Immunol. 1, 476, 1971. 30. Gray, E. D., J. Exp. Med. 149, 1438, 1979. 31. Hrynieurca, W., Gray, E. D., Tagg, J. R., Wannamaker, L. W., Kanclerski, K., and Laible, N.,
Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg Abt. I Orig. Reihe A. A242, 327, 1978. 32. Hrynieurca, W., and Pryjma, J., Infect. Immun. 16, 730, 1977. 33. Barsumian, E. L., Schlievert, P. M., and Watson, D. W., Infect. Immun. 22, 681, 1978. 34. Cuppari, G., Quagliata, F., Ieri, A., and Taranta, A., J. Lab. Clin. Med. 80, 165, 1972. 35. Geier, S. S., and Cresswell, P., Cell Immunol. 28, 341, 1977. 36. Shen, H. H., Irigoyen, O., Friedman, S. M., and Chess, L., J. Immunof. 128, 501, 1982.


















