
Frontiers in Neuroendocrinology - Rutgers...
15
Review Neuroendocrinology of coping styles: Towards understanding the biology of individual variation J.M. Koolhaas * , S.F. de Boer, C.M. Coppens, B. Buwalda Dept. of Behavioural Physiology, University Groningen, P.O. Box 14, 9750 AA Haren, The Netherlands article info Article history: Available online 9 April 2010 Keywords: Coping style Animal personality Individual difference HPA-axis Serotonin Vasopressin Catecholamines Gonadal steroids abstract Individual variation in behavior and physiology is a widespread and ecologically functional phenomenon in nature in virtually all vertebrate species. Due to domestication of laboratory animals, studies may suf- fer from a strong selection bias. This paper summarizes behavioral, neuroendocrine and neurobiological studies using the natural individual variation in rats and mice. Individual behavioral characteristics appear to be consistent over time and across situations. The individual variation has at least two dimen- sions in which the quality of the response to a challenging condition (coping style) is independent from the quantity of that response (stress reactivity). The neurobiology reveals important differences in the homeostatic control of the serotonergic neuron and the neuropeptides vasopressin and oxytocin in rela- tion to coping style. It is argued that a careful exploitation of the broad natural and biologically functional individual variation in behavior and physiology may help in developing better animal models for under- standing individual disease vulnerability. Ó 2010 Elsevier Inc. All rights reserved. 1. Introduction Biomedical research has struggled with individual variation ever since its existence. Clearly, it is not the average population that gets a certain disease, but usually specific individuals under specific environmental conditions. For example, it is well recog- nized in humans that only a rather small proportion of individuals develop depression/anxiety disorders after stressful life events, be- come addicted after exposure to drugs of abuse, or express esca- lated and violent aggression after repeated social conflicts. Likewise, the therapeutic efficacy and/or side-effect profile of phar- macological treatments of these psychopathologies is highly vari- able between individuals. Hence, the major challenge of contemporary biomedical research is to understand not only this individual disease vulnerability, but also the individual differenti- ation in response to medication. Biomedical research using animal models starts to recognize this problem as well. During the last decades, preclinical animal research has obtained a wealth of evi- dence on the way in which the adult phenotype is shaped by the physical and social environment at critical periods during develop- ment [151]. Moreover, it becomes increasingly clear that these developmental processes interact with genetic predispositions in a rather complex way [90,119], and the adult phenotype is consid- ered to be the net result of this gene-environment interaction. This complex, rather subtle and poorly understood interaction is one of the main reasons why certain experimental results are often diffi- cult to reproduce in different laboratories [102,149]. The answer to this problem is generally a further reduction of genetic variation by inbreeding and standardizing rearing, housing and experimental conditions. However, even within inbred populations of mice or rats exposed to adverse (stressful) situations, susceptible and unsusceptible subpopulations can be distinguished. The result of more than a century of laboratory animal experimentation is that science can now select their experimental animals out of a list of over 500 officially registered inbred strains of mice and about 250 registered inbred rat strains. In addition, there are numerous outbred strains, unregistered strains and an almost exponentially increasing number of transgenic strains. This has resulted in the peculiar situation where it is virtually impossible to select a given strain of experimental animals on a rational scientific basis, with the exception of transgenic animals. Science is trying to correct this by starting extensive phenotypical characterization of large series of rat and mouse strains [126]. Although it is very useful to start experiments on the basis of an extensive behavioral and neuroen- docrine characterization of the strain of experimental animals, the choice may still be rather arbitrary from a biological point of view. It will be argued below that individual variation in nature is not infinite, but seems to be clustered in a limited number of pheno- types or behavioral syndromes. In laboratory animals, it is often unknown how a given strain i.e. genotype and phenotype relates to the biologically functional variation of its wild ancestors living under natural selection pressures. From this point of view, many laboratory animal strains might be considered as laboratory 0091-3022/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.yfrne.2010.04.001 * Corresponding author. Fax: +31 50 363 2331. E-mail address: [email protected] (J.M. Koolhaas). Frontiers in Neuroendocrinology 31 (2010) 307–321 Contents lists available at ScienceDirect Frontiers in Neuroendocrinology journal homepage: www.elsevier.com/locate/yfrne
-
Upload
vuongquynh -
Category
Documents
-
view
216 -
download
0
Transcript of Frontiers in Neuroendocrinology - Rutgers...

Review
Neuroendocrinology of coping styles: Towards understanding the biologyof individual variation
J.M. Koolhaas *, S.F. de Boer, C.M. Coppens, B. BuwaldaDept. of Behavioural Physiology, University Groningen, P.O. Box 14, 9750 AA Haren, The Netherlands
a r t i c l e i n f o
Article history:Available online 9 April 2010
Keywords:Coping styleAnimal personalityIndividual differenceHPA-axisSerotoninVasopressinCatecholaminesGonadal steroids
a b s t r a c t
Individual variation in behavior and physiology is a widespread and ecologically functional phenomenonin nature in virtually all vertebrate species. Due to domestication of laboratory animals, studies may suf-fer from a strong selection bias. This paper summarizes behavioral, neuroendocrine and neurobiologicalstudies using the natural individual variation in rats and mice. Individual behavioral characteristicsappear to be consistent over time and across situations. The individual variation has at least two dimen-sions in which the quality of the response to a challenging condition (coping style) is independent fromthe quantity of that response (stress reactivity). The neurobiology reveals important differences in thehomeostatic control of the serotonergic neuron and the neuropeptides vasopressin and oxytocin in rela-tion to coping style. It is argued that a careful exploitation of the broad natural and biologically functionalindividual variation in behavior and physiology may help in developing better animal models for under-standing individual disease vulnerability.
! 2010 Elsevier Inc. All rights reserved.
1. Introduction
Biomedical research has struggled with individual variationever since its existence. Clearly, it is not the average populationthat gets a certain disease, but usually specific individuals underspecific environmental conditions. For example, it is well recog-nized in humans that only a rather small proportion of individualsdevelop depression/anxiety disorders after stressful life events, be-come addicted after exposure to drugs of abuse, or express esca-lated and violent aggression after repeated social conflicts.Likewise, the therapeutic efficacy and/or side-effect profile of phar-macological treatments of these psychopathologies is highly vari-able between individuals. Hence, the major challenge ofcontemporary biomedical research is to understand not only thisindividual disease vulnerability, but also the individual differenti-ation in response to medication. Biomedical research using animalmodels starts to recognize this problem as well. During the lastdecades, preclinical animal research has obtained a wealth of evi-dence on the way in which the adult phenotype is shaped by thephysical and social environment at critical periods during develop-ment [151]. Moreover, it becomes increasingly clear that thesedevelopmental processes interact with genetic predispositions ina rather complex way [90,119], and the adult phenotype is consid-ered to be the net result of this gene-environment interaction. Thiscomplex, rather subtle and poorly understood interaction is one of
the main reasons why certain experimental results are often diffi-cult to reproduce in different laboratories [102,149]. The answer tothis problem is generally a further reduction of genetic variation byinbreeding and standardizing rearing, housing and experimentalconditions. However, even within inbred populations of mice orrats exposed to adverse (stressful) situations, susceptible andunsusceptible subpopulations can be distinguished. The result ofmore than a century of laboratory animal experimentation is thatscience can now select their experimental animals out of a list ofover 500 officially registered inbred strains of mice and about250 registered inbred rat strains. In addition, there are numerousoutbred strains, unregistered strains and an almost exponentiallyincreasing number of transgenic strains. This has resulted in thepeculiar situation where it is virtually impossible to select a givenstrain of experimental animals on a rational scientific basis, withthe exception of transgenic animals. Science is trying to correct thisby starting extensive phenotypical characterization of large seriesof rat and mouse strains [126]. Although it is very useful to startexperiments on the basis of an extensive behavioral and neuroen-docrine characterization of the strain of experimental animals, thechoice may still be rather arbitrary from a biological point of view.It will be argued below that individual variation in nature is notinfinite, but seems to be clustered in a limited number of pheno-types or behavioral syndromes. In laboratory animals, it is oftenunknown how a given strain i.e. genotype and phenotype relatesto the biologically functional variation of its wild ancestors livingunder natural selection pressures. From this point of view, manylaboratory animal strains might be considered as laboratory
0091-3022/$ - see front matter ! 2010 Elsevier Inc. All rights reserved.doi:10.1016/j.yfrne.2010.04.001
* Corresponding author. Fax: +31 50 363 2331.E-mail address: [email protected] (J.M. Koolhaas).
Frontiers in Neuroendocrinology 31 (2010) 307–321
Contents lists available at ScienceDirect
Frontiers in Neuroendocrinology
journal homepage: www.elsevier .com/locate /yfrne

artifacts having a potential low fitness in nature. This paper willtake a rather unconventional approach by considering the biologi-cal basis of individual variation as it occurs in nature as an alterna-tive to the use of laboratory animal strains. There is a rapidlygrowing interest in the biological significance of individual differ-entiation in behavior and physiology in the science of ecologyand evolutionary biology. Individual variation in coping with chal-lenges encountered in the natural habitat determines evolutionaryfitness and is considered to be the prerequesite of speciation.Moreover, it may be an important factor in the regulation of pop-ulations as well. It seems that evolution has shaped general adap-tive response patterns in reaction to everyday challenges in thenatural habitat [7,111]. The aim of the present paper is to explorethe ultimate biological basis of individual variation in stress reac-tivity in terms of individual coping styles in animals as they mightbe observed in nature and to discuss the proximate consequenceswith respect to neuroendocrine reactivity.
2. Individual variation
Variation between individuals is a well known phenomenon inboth human and animal experimentation, often regarded as a nui-sance by many researchers rather than a worthy target for investi-gation. For decades, the dogma in animal experimentation was toreduce individual variation as much as possible in order to obtainstatistically significant results with the least number of animals.Due to the 3R doctrine [105], reduction of the number of animalsused in an experiment often became a goal in itself. Reducing indi-vidual variation by using outbred or inbred strains of laboratoryanimals and through excessive and rigorous standardization maycompromise the general validity and robustness of the experimen-
tal results. It may lead to a strong selection bias in the experimen-tal results [148]. Fig. 1 shows an example of this. The frequencydistribution of total time spent on offensive aggressive behaviorin a standard outbred wistar strain of laboratory rats is comparedwith the distribution in an outbred strain of wild-derived rats, bredin the laboratory, but originating from feral breeding pairs. Despitelaboratory breeding of the wild-type rat, this wide distribution isconsidered to be representative of the individual variation as itmight be functional in nature. Clearly, the highly aggressive pheno-type is absent in the laboratory strain, most likely due to some(perhaps unintended) artificial-selection for tameness during thecentury-long domestication process of this wild animal. This isunderstandable from the point of view of an animal caretaker,but from an experimental point of view this is a scientificallyunjustified selection. One may argue that selection bias is inevita-ble and inherent to the use of animal models, starting with theselection of the animal species followed by the selection of thestrain. However, we want to show that the individual variationin behavior and physiology, as it might have a function in nature,can be used experimentally and might be an important standardin the interpretation of experimental results. After all, reducingvariation may imply a reduction of the general validity of theexperimental results at the same time. Fig. 1 also illustrates theimportance of an absolute scale. Obviously, a manipulation thatleads to a 4-fold increase in aggressive behavior requires a differ-ent biological interpretation when it changes from 2% to 8% of totaltime than a change from 20% to 80% on the absolute scale. Again,the biology of the species, including the degree of individual vari-ation can be an important reference standard in the interpretationof experimental results.
2.1. Biological significance
In behavioral ecology, individual variation is considered to be aprerequisite for evolution. The general idea is that certain individ-uals under particular environmental conditions have a higher fit-ness than others leading to a higher reproduction, better survival,dispersal, etc. Differences in fitness are thought to be the basis ofspeciation. However, recent studies in feral populations demon-strate that a considerable degree of genetic variation, partly under-lying the phenotypic variation, is maintained in the population andmight be evolutionary stable [106,110]. Not only has this individ-ual variation been demonstrated to be heritable in many species,but recent computer simulations show that individual differencesin responsiveness to the environment could most certainly evolve[146,147]. Apparently, different phenotypes may have a differentfunction in the population ecology of the species. This notion goesback to the original hypothesis by Chitty [27] that genetic variationin aggressive behavior might play an important role in the popula-tion dynamics of house mice. Genetic variation for aggression wassuggested to be maintained in the population because the ex-tremes in the population have a differential fitness in differentpopulation densities or under different environmental conditions.In nature, mouse populations are known to go through phases ofgrowth and decline. Such population cycles may have a periodfrom four to seven years and can be so extreme that the populationsuddenly collapses at the end of a cycle and becomes extinct [16].There is still no satisfactory understanding and explanation for thecyclic nature of mammalian populations [83]. Factors such as pre-dation risk, and food availability seem to play a role and the cyclicnature may disappear over time [66,95]. An old hypothesis by Chit-ty [27] suggests that the cyclic nature of rodent populations mightbe due to disruptive selection for aggressive behavior in the courseof the population cycle. This hypothesis is supported by the earlystudies of Van Oortmerssen and Busser [57] on feral populationsof house mice. Phenotypic characterization of the laboratory bred
0
5
10
15
20
25
30
35
40Wistar rats (n=77)
Perc
enta
ge o
f ani
mal
s
Aggression score class (% offense)
Low aggressive animals Medium aggressive animals
A
Low aggressive animals Medium aggressive animals high aggressive animals
0 20 40 60 80 100
0 20 40 60 80 1000
5
10
15
20
25Wild-type rats (n=315)
Perc
enta
ge o
f ani
mal
s
Aggression score class (% offense)
B
Fig. 1. Frequency distribution of offensive aggressive behavior in a sample (N = 77)of adult wistar rats (A) and a sample (N = 315) of adult wild-type rats (B) asmeasured during ten minutes in the standard resident-intruder paradigm usingmale wistar intruders of the same age.
308 J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321

male offspring (F1) of breeding pairs caught from feral colonies, re-vealed a bimodal distribution of attack latencies as measured in astandardized resident-intruder paradigm. Subsequent selectivebreeding for high and low attack latencies resulted within five gen-erations in stable short attack latency (SAL) selection line. After anumber of failures due to infertility of the offspring, we managedto obtain a long attack latency (LAL) selection line as well. Embryotransfer, cross-fostering and back-cross experiments show that thephenotypic differentiation in aggressive behavior as observed inthe wild has a strong genetic basis that is only marginally influ-enced by the maternal environment [117]. Several additional datasupport the general idea that stable genetic variation for aggressivebehavior may be a factor in the population dynamics of the wildhouse mouse. Indeed, analysis of mortality in feral populations re-veals a strong increase in dead females, juveniles and pre-weanlingjuveniles just before the crash of the population [57]. It is temptingto consider the possibility that this is due to high levels of inter-male aggression in that phase of the population cycle. During thewhole population cycle, animals are migrating from the popula-tion. It is suggested that the non-aggressive phenotype is moresuccessful in establishing a new colony than the highly aggressivephenotype. More recently, the ecological significance of geneticvariation for aggression was also demonstrated in an extensivefield and laboratory study of a passerine bird, the great tit (Parusmajor) [47]. It was shown that individual variation in explorationof novel trees was stable over time and correlated with a varietyof other behavioral characteristics including aggressive behavior[137]. Subsequent selective breeding experiments showed the ge-netic basis of this differentiation. The fitness of the two phenotypeswas studied in individually characterized birds living in a wildpopulation. A differential fitness of the two phenotypes was ob-served under conditions of high and low food availability [44].
In an extensive review, Sih et al. [110] give several other exam-ples in mammals, birds, fish and insects indicating that phenotypicvariation within feral populations can be stable over time andacross situations. These behavioral syndromes appear to have afunction in the ecology of the species and are somehow maintainedwithin a single natural population. Several studies suggest that thedegree in which behavioral syndromes are expressed in naturemay depend on the magnitude of selection pressure in feral popu-lations [6,29]. Recent experimental evidence confirms that the de-gree in which different behavioral characteristics are correlateddepend on environmental selection pressure. In an elegant exper-iment in sticklebacks, it was shown that boldness in terms of thetendency of individual fish to approach a predator became corre-lated with intra-specific aggression in experimental fish popula-tions only after increasing selection pressure by adding apredator to the population [7,45]. The development of behavioralsyndromes was not only due to selection of specific phenotypesby the predator, but also due to phenotypic plasticity. This andother studies support the general view that individual variationin correlated behaviors may have an adaptive function in nature,buffering the species against fluctuations in environmental condi-tions [100].
2.2. Coping styles
Since the times of the Greek philosophers, scientists have triedto categorize individual variability in behavior and underlyingphysiology into distinct personalities or temperaments. Currently,the general view is that trait characteristics should be stable over aconsiderable period of time and should be consistent across situa-tions. Although stable trait characteristics are found in many ani-mal species, there is a lack of consistency and agreement in theliterature regarding the terminology used to describe and catego-rize trait characteristics. Various terms are used to categorize the
extremes of the individual variation such as shy versus bold, activeversus passive, proactive versus reactive, and hawk versus dove[74,78]. Behavioral ecology has adopted the term behavioral syn-drome indicating that trait characteristics should involve suits ofcorrelated behaviors [100,111]. This more neutral term may applyfor any set of correlated behaviors that shows consistency overtime and across situations. In the biomedical research field theterm coping style is commonly used for these individual traitdifferences.
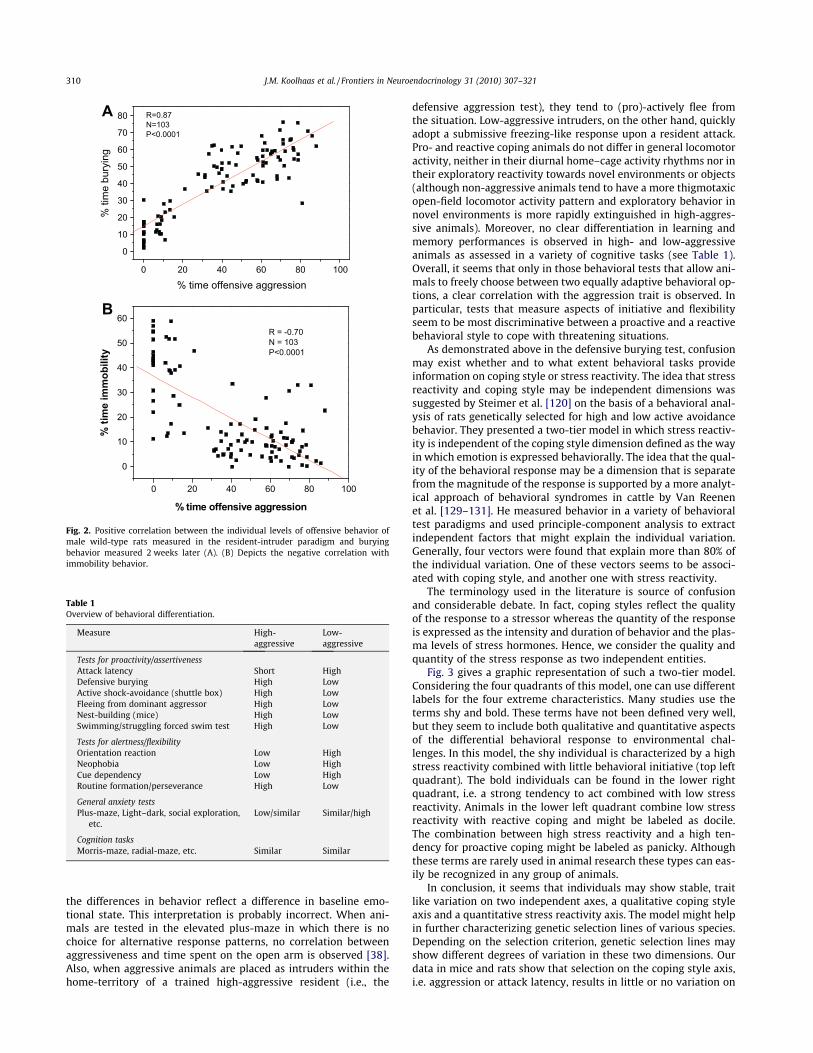
In a series of studies using rats and mice, we have presentedexperimental evidence that the concept of coping style might ex-plain a considerable proportion of the individual trait like variationin behavior and underlying physiology. Coping styles may be de-fined as alternative response patterns in reaction to a stressor.The concept is based on the observation that the individual varia-tion in offensive aggressive behavior is stable over time and relatedto the response pattern in a variety of other challenging conditions.Indeed, the amount of time spent on offensive behavior by malewild-type rats in the resident-intruder paradigm is very stable overtime in different individuals [75]. Henry and Stephens [63] werethe first to suggest that the individual variation in aggressionmight be an expression of actively coping with environmentalchallenges in general. The consistency of aggression as an expres-sion of coping style across situations can be best demonstratedin tests that allow the animal a choice of different response pat-terns. For example, in the defensive burying paradigm [124] ani-mals are confronted with an electrified probe inserted into theirhome cage. In response to a brief contact with the shock probe,the animal can either actively bury the probe with the beddingof their cage or show immobility and more passively avoid theelectrified probe. Both response patterns can be considered as suc-cessful coping, because in neither of the two response patterns theanimal will ever touch the probe again. Fig. 2a shows the positivecorrelation between offensive behavior measured in the resident-intruder paradigm and burying behavior in the defensive buryingtest. Fig. 2b shows the negative correlation between aggressivebehavior and immobility. Similar results were obtained in theSAL and LAL selection lines of mice that were mentioned above[116], and in laboratory rats that were genetically selected foreither high or low active shock-avoidance [13]. This demonstratesthe consistency of individual variation in behavior across situa-tions, i.e. the aggressive response to an intruder in the home cagepredicts an active behavioral response in this non-social buryingparadigm. Other examples of this consistency in response patternto challenging situations can be found in nest building behavior[115], active avoidance behavior [9] and in the forced swim test[135]. The behavioral differentiation observed in wild-type ratsand the selection lines of wild house mice are summarized in Ta-ble 1. This table supports the view that high levels of offensiveaggressive behavior can be considered as an expression of a moregeneral tendency to adopt a proactive coping style.
Low levels of offensive aggressive behavior can be considered asa reactive coping style [74]. It is important to notice that the copingstyle concept implies that animals may react with alternative re-sponse patterns. Interestingly, Treit [124] originally described thedefensive burying test as an anxiety test. High levels of buryingbehavior would indicate high levels of anxiety. However, it can justas well be considered as a test for the way in which emotion is ex-pressed behaviorally [38]. The same argument seems to hold forthe forced swim test in which floating behavior is frequently inter-preted as a behavioral expression of depression. Here too, aggres-sive males react with active swimming and climbing whereasnon-aggressive males show predominantly floating behavior[135]. Hence, in the interpretation of these tests one has to con-sider not only the magnitude of the behavioral response to a stres-sor but also the type of behavioral response. One may argue that
J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321 309
the differences in behavior reflect a difference in baseline emo-tional state. This interpretation is probably incorrect. When ani-mals are tested in the elevated plus-maze in which there is nochoice for alternative response patterns, no correlation betweenaggressiveness and time spent on the open arm is observed [38].Also, when aggressive animals are placed as intruders within thehome-territory of a trained high-aggressive resident (i.e., the
defensive aggression test), they tend to (pro)-actively flee fromthe situation. Low-aggressive intruders, on the other hand, quicklyadopt a submissive freezing-like response upon a resident attack.Pro- and reactive coping animals do not differ in general locomotoractivity, neither in their diurnal home–cage activity rhythms nor intheir exploratory reactivity towards novel environments or objects(although non-aggressive animals tend to have a more thigmotaxicopen-field locomotor activity pattern and exploratory behavior innovel environments is more rapidly extinguished in high-aggres-sive animals). Moreover, no clear differentiation in learning andmemory performances is observed in high- and low-aggressiveanimals as assessed in a variety of cognitive tasks (see Table 1).Overall, it seems that only in those behavioral tests that allow ani-mals to freely choose between two equally adaptive behavioral op-tions, a clear correlation with the aggression trait is observed. Inparticular, tests that measure aspects of initiative and flexibilityseem to be most discriminative between a proactive and a reactivebehavioral style to cope with threatening situations.
As demonstrated above in the defensive burying test, confusionmay exist whether and to what extent behavioral tasks provideinformation on coping style or stress reactivity. The idea that stressreactivity and coping style may be independent dimensions wassuggested by Steimer et al. [120] on the basis of a behavioral anal-ysis of rats genetically selected for high and low active avoidancebehavior. They presented a two-tier model in which stress reactiv-ity is independent of the coping style dimension defined as the wayin which emotion is expressed behaviorally. The idea that the qual-ity of the behavioral response may be a dimension that is separatefrom the magnitude of the response is supported by a more analyt-ical approach of behavioral syndromes in cattle by Van Reenenet al. [129–131]. He measured behavior in a variety of behavioraltest paradigms and used principle-component analysis to extractindependent factors that might explain the individual variation.Generally, four vectors were found that explain more than 80% ofthe individual variation. One of these vectors seems to be associ-ated with coping style, and another one with stress reactivity.
The terminology used in the literature is source of confusionand considerable debate. In fact, coping styles reflect the qualityof the response to a stressor whereas the quantity of the responseis expressed as the intensity and duration of behavior and the plas-ma levels of stress hormones. Hence, we consider the quality andquantity of the stress response as two independent entities.
Fig. 3 gives a graphic representation of such a two-tier model.Considering the four quadrants of this model, one can use differentlabels for the four extreme characteristics. Many studies use theterms shy and bold. These terms have not been defined very well,but they seem to include both qualitative and quantitative aspectsof the differential behavioral response to environmental chal-lenges. In this model, the shy individual is characterized by a highstress reactivity combined with little behavioral initiative (top leftquadrant). The bold individuals can be found in the lower rightquadrant, i.e. a strong tendency to act combined with low stressreactivity. Animals in the lower left quadrant combine low stressreactivity with reactive coping and might be labeled as docile.The combination between high stress reactivity and a high ten-dency for proactive coping might be labeled as panicky. Althoughthese terms are rarely used in animal research these types can eas-ily be recognized in any group of animals.
In conclusion, it seems that individuals may show stable, traitlike variation on two independent axes, a qualitative coping styleaxis and a quantitative stress reactivity axis. The model might helpin further characterizing genetic selection lines of various species.Depending on the selection criterion, genetic selection lines mayshow different degrees of variation in these two dimensions. Ourdata in mice and rats show that selection on the coping style axis,i.e. aggression or attack latency, results in little or no variation on
A
0
10
20
30
40
50
60
70
80 R=0.87N=103P<0.0001
% ti
me
bury
ing
% time offensive aggression0 20 40 60 80 100
0 20 40 60 80 100
0
10
20
30
40
50
60R = -0.70N = 103P<0.0001
% ti
me
imm
obili
ty
% time offensive aggression
B
Fig. 2. Positive correlation between the individual levels of offensive behavior ofmale wild-type rats measured in the resident-intruder paradigm and buryingbehavior measured 2 weeks later (A). (B) Depicts the negative correlation withimmobility behavior.
Table 1Overview of behavioral differentiation.
Measure High-aggressive
Low-aggressive
Tests for proactivity/assertivenessAttack latency Short HighDefensive burying High LowActive shock-avoidance (shuttle box) High LowFleeing from dominant aggressor High LowNest-building (mice) High LowSwimming/struggling forced swim test High Low
Tests for alertness/flexibilityOrientation reaction Low HighNeophobia Low HighCue dependency Low HighRoutine formation/perseverance High Low
General anxiety testsPlus-maze, Light–dark, social exploration,
etc.Low/similar Similar/high
Cognition tasksMorris-maze, radial-maze, etc. Similar Similar
310 J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321

the emotionality axis [38]. It is conceivable that genetic selectionlines may show a different degree of variation on the two dimen-sions, depending on the selection criterion used. For example, ratsgenetically selected for high or low levels of anxiety like behavior(HAB and LAB respectively) vary between 0% and 63% at the emo-tionality dimension i.e. in anxiety, but differ between 0% and 30%in aggressive behavior as a component of the coping style dimen-sion [132].
Considering the five main trait dimensions in humans (big five),trait characteristics in animals are likely to be multidimensional aswell. The research in this area is still relatively young and underde-veloped. In an extensive review of the available animal literature,Reale et al. [100] suggests five trait characteristics, some of whichseem to be in line with the two dimensions suggested here. Clearly,the issue needs further experimental support using multidimen-sional scaling methods of animals tested in a wide range of situa-tions [4]. Moreover, one may argue that the dimensions and theterminology should somehow reflect the individual variation inthe proximate causal mechanisms.
2.3. Proximate behavioral considerations
So far, the concept of coping style has been discussed indescriptive terms of behavioral syndromes or sets of correlatedbehaviors. One may wonder why these behaviors are correlatedfrom a more proximate mechanistic point of view. Characteristicof the proactive coping style is that actions are principally basedon predictions. This is in contrast to the reactive coping style inwhich there is a more direct stimulus–response relationship.This fundamental behavioral difference can be demonstrated inmaze experiments showing that proactive and reactive copingstyles differ in the degree in which behavior is guided by envi-ronmental cues [74]. Animals can be trained to run through amaze for a food reward. Rats and mice can learn such a task eas-ily. When they reached a stable task performance, the reactionto a small change in the maze was studied. In one experiment,a small piece of tape was put on the floor in one of the alleysof the maze, while in another experiment the maze was turned
ninety degree with respect to the extra-maze cues. In bothexperiments, the proactive coping males paid little or no atten-tion to the change; i.e. there was no increase in time to com-plete the task and no increase in the number of errors madein the maze. Reactive coping males on the other hand startedexploring the maze again and hence took much more time toget to the goal box and made more errors in the task. This sug-gests that the reactive coping style may be much more guidedby environmental stimuli, while the proactive coping style seemsto develop routines. This fundamental difference in cue depen-dency between the two coping styles was confirmed in pigsusing reversal learning in a T-maze. Proactive coping pigs hadfar more difficulties in switching from a once learned task to anew one than reactive coping pigs [15]. This difference in behav-ioral flexibility can be demonstrated in several other situationswhere the animal has to switch suddenly from a standard situ-ation to a new one. For example, the two coping styles differstrongly in the response to a 12-h shift in light/dark cycle. Pro-active coping male mice stay in their original day–night rhythmfor a few days after which their rhythm gradually shifts to thenew cycle. Reactive coping males on the other hand start to shifttheir rhythm immediately; they are twice as fast in adapting tothe new light–dark cycle as the proactive coping males [8]. Re-cent experiments in rats and hamsters using operant condition-ing paradigms show that this individual differentiation inbehavioral flexibility may be related to a differentiation in pre-frontal cortex functioning [26,37].
These experiments show that the two coping styles differ moregenerally in behavioral flexibility. The proactive animal acts pri-marily on the basis of previous experience, i.e. feed-forward con-trol. The reactive coping animal tends to rely more on thedetailed accounts of the environment, i.e. it reacts. This fundamen-tal difference in behavioral control also relates to the adaptivecharacter of the two coping styles. A proactive coping animalmay be adapted to stable environmental conditions. The reactivecoping style may do better under variable and unpredictable envi-ronmental conditions. Indeed, field studies on feral mouse popula-tions indicate that aggressive males are more successful under
Fig. 3. Two tier model with coping style and stress reactivity as two independent dimensions of stable trait characteristics. The four quadrants indicate the type of animalwhen varying on the two dimensions simultaneously.
J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321 311

stable colony conditions, whereas non-aggressive males may dobetter during migration.
3. Neuroendocrinology of adult coping styles
Individual differentiation in behavior has frequently been asso-ciated with distinct patterns of both autonomic nervous and endo-crine (re)activity. While the majority of studies focus on hormonesof the hypothalamic–pituitary–adrenal (HPA) axis, some studiesalso included the individual difference in and the hypothalamic-pituitary-gonadal axis (HPG) and the balance between the sympa-thetic and parasympathetic branch of the autonomic nervoussystem.
3.1. Autonomic nervous system
Social defeat induces a strong release of noradrenalin andadrenalin (Fig. 4). This is consistent with earlier observations thatsocial defeat can be considered as one of the most severe stressorsin terms of the magnitude of the physiological stress response [72].The correlation diagrams in Fig. 4 clearly show that aggressivemales respond with the highest increase in both adrenaline andnoradrenaline. In the shock-prod burying test the response of plas-ma catecholamines is on average a fifth of the response to defeat,but noradrenaline is still positively correlated with offensiveaggression (Fig. 5). These data support the general view that theaggressive, proactive male is characterized by a high reactivity ofthe sympathetic nervous system. Baseline levels of adrenalineand noradrenaline are generally similar.
3.2. HPA-axis
Figs. 4 and 5 show that both in the social defeat test and duringshock-prod defensive burying plasma corticosterone is not corre-lated with the individual level of offensive aggressive behavior ofwild-type rats measured in the resident-intruder paradigm. Thisresult is in contrast to a number of other studies that suggest thatthe reactive coping style is associated with both a high baselineactivity and a higher stress reactivity of the HPA-axis. In the forcedswimming test, LAL males show a higher ACTH and corticosteroneresponse than SAL males [133]. Korte et al. [76] showed that circa-dian peak values of plasma corticosterone were more than twice ashigh in non-aggressive (reactive) LAL male house mice as com-pared to highly aggressive (proactive) SAL males. However, laterstudies in our lab failed to confirm this finding. Under basal condi-tions circadian trough values of corticosterone were shown to behigher in SAL males [133,134]. Baseline levels of plasma ACTHwere higher in SAL males indicating decreased adrenal cortex sen-sitivity for ACTH.
The general picture of the relationship between aspects of cop-ing style and HPA-axis activity is rather complicated. In great titsand geese [23,80], the non-aggressive, reactive coping animalsshow the highest response. However, in the wild-type rats, cyno-molgus monkeys [64] and stickle back [7], this association is ab-sent or variable and in trout there is only a weak associationbetween HPA-axis activity and coping style [107].
3.3. HPG-axis
Some early studies in rats and mice demonstrated a positiverelationship between intermale aggression and plasma T levels
0
2000
4000
6000
8000
0
1000
2000
3000
4000
0
10
20
30
40
50
Nor
adre
nalin
e (p
g/m
l) DefeatN=19
Adre
nalin
e (p
g/m
l)C
ortic
oste
rone
(µg/
dl)
Time (min)-15-10 -5 0 5 10 15 20 25 30 35 40 45 50 55 60 65 0 20 40 60 80
200400600800
10001200140016001800
0
20
40
60
80
100
0102030405060
CS
resp
onse
(AU
C)
Aggressiveness (% offense)
R=0.26n=19p=0.29
NA
resp
onse
(AU
C)
R=0.79n=19p<0.001
A re
spon
se (A
UC
)
R=0.78n=19p<0.001
0 20 40 60 80
0 20 40 60 80
Fig. 4. Individual response curves of noradrenalin, adrenalin and corticosterone to social defeat. The gray bar depicts the period of physical interaction. The right hand graphsshow the relationship between the total neuroendocrine response (area under the curve) and the individual level of offensive aggressive behavior measured in the resident-intruder paradigm. Blood samples were collected using permanently implanted jugular vein canula.
312 J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321

[96,109]. However, later experiments showed the inconsistency ofthis relationship [32,51]. Although there are clear indications thattestosterone secretion relates to the control of aggression in manyspecies, this mainly holds for aggressive behavior in a reproductivecontext [58,143,144] and not for trait differences in aggression orcoping. It seems that the relatively weak T-aggression relationshipexplains some of the aggressive displays in particular situationslike aggression during the mating season but certainly not in manyother situations.
In the brain, T is aromatized to estradiol acting on estrogenreceptors and via 5-a reductase reduced to di-hydro-testosteroneacting on androgen receptors. These enzymes and receptors areknown to be causally involved in aggression [112,123]. However,a recent study in song sparrows shows that the involvement ofthe androgen receptor in aggression is restricted to the breedingseason [118]. There is some evidence in mice that individual vari-ation in aromatase [33] and in the estrogen a receptors in distinctbrain areas might explain individual variation in aggression [122].
05
10152025303540
0200400600800
10001200
0
100
200
300
400
Cor
ticos
tero
ne ( µ
g/dl
)
Time (min)
Shock-prodN
orad
rena
line
(pg/
ml)
Adre
nalin
e (p
g/m
l)
-4000-2000
02000400060008000
-2000
0
2000
4000
6000
8000
-15-10 -5 0 5 10 15 20 25 30 35 40 45 50 55 60 65 0 10 20 30 40 50 60 70 80
-2000
200400600800
1000120014001600
R=0.71N = 16P= 0.0023
NA
resp
onse
(AU
C)
R=0.45N=16P=0.079
A re
spon
se (A
UC
)
R=-0.09N=16P=0.74
CS
resp
onse
(AU
C)
% time offensive aggression
Fig. 5. Individual response curves of noradrenalin, adrenalin and corticosterone during defensive burying. The right hand graphs show the relationship between the totalneuroendocrine response (area under the curve) and the individual level of offensive aggressive behavior measured in the resident-intruder paradigm. Blood samples werecollected using permanently implanted jugular vein canula.
200
400
600
800
1000
1200
1400
1600
1800R=0.73N=14P<0.001
Cor
toco
ster
one
resp
onse
(AU
C; µ
g .m
in/d
l)
Aggressiveness (% offense)0 20 40 60 80-15-10 -5 0 5 10 15 20 25 30 35 40 45 50 55 60 65
05
1015
2025
3035
4045
5055
Cor
tioco
ster
one
( µg/
dl)
Time (min)
Conflict WinnersN=14
Fig. 6. Individual response curves of corticosterone during winning of an aggressive interaction. The gray bar depicts the period of direct physical interaction. The right handgraph shows the positive correlation of the total corticosterone response (area under the curve) with the individual level of aggressive behavior. Blood samples were collectedusing permanently implanted jugular vein canula.
J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321 313

To what extent the involvement of HPG system in aggressivebehavior may be generalized to other behavioral expressions ofcoping style is unknown.
3.4. Discussion
Little is known about the exact causal relationship between thedifferential behavioral and neuroendocrine characteristics. How-ever, based on the outcome of several studies performed in ourlab and with the exception of the HPG-axis, we consider it likelythat the differential neuroendocrine characteristics are mainly aconsequence rather that the cause of the behavioral differentiation.This is supported by the magnitude of the corticosterone responseobserved in male wild-type rats during winning of a social interac-tion (Fig. 6). In this situation there is a significant positive correla-tion between the individual level of aggressive behavior as anindependent trait characteristic and the corticosterone response.Apparently, HPA-axis reactivity is not a trait characteristic. Corre-lations with coping style that are situational dependent shouldbe considered as the consequence of the differential appraisal ofthe environmental challenge. Walker et al. [138] came to a similarconclusion in a study of the corticosterone response during socialdefeat. A similar type of reasoning may hold for the correlation be-tween coping style and sympathetic activation. Sympathetic ner-vous activity in general and plasma noradrenalin in particular isconsidered to be a direct reflection of the metabolic and cardiovas-cular demands required for physical activity. Because of the factthat proactive coping is characterized by an active response toenvironmental challenges, it is no surprise to find a positive corre-lation between proactive coping and sympathetic reactivity.
4. Neuroendocrine factors in the development of coping styles
The adult individual difference in behavioral coping styles andthe accompanying differences in neuroendocrine and physiological(re)activity are the result of a complex interaction between geneticbackground and environmental modulation. There are two periodsin the life history during which hormones have a clear organiza-tional action in neural structures that might be involved in the reg-ulation of coping behavior: the perinatal period and puberty. Theorganizational role of pre- and postnatal variation in neuroendo-crine (re)activity on the brain and its relationship to adult copingbehavior has been extensively studied. More recently, interest isincreasing in the question to what extent environmental and neu-roendocrine factors during adolescence and puberty may alterbrain circuitry and in this way contribute to differences in adultcoping style.
4.1. HPG-axis during perinatal development
The organizational role of neuroendocrinology in the ontogenyof coping styles has been addressed in several studies. Hormonesof the hypothalamic–pituitary–gonadal (HPG) axis and in particu-lar testosterone (T) received a substantial amount of attention inthis respect. Studies in SAL and LAL mice in our laboratory indi-cated that perinatally circulating T levels [33], T secreting capacityof the testis [42] and brain aromatase activity [33] were strikinglydifferent in aggressive and non-aggressive mice. The results indi-cated that the circulating T peaks earlier in LAL than in SAL mice.In LAL males T surges prenatally at embryonic day 17–18. At thisstage plasma testosterone is still low in SAL males. Neonatally, atpostnatal day 1, T has decreased to low levels in LAL males butpeaks in SAL males together with brain aromatase activity. Thepeak activity of this enzyme in brain areas of LAL mice occurs alsoon postnatal day 1 but is much lower compared to SAL males. Neo-
natal T surges are known to be involved in the permanent organi-zation of brain circuitry resulting in higher responsiveness to Tfacilitating aggression in adulthood in both male and female ani-mals [17]. This modulatory role of T on adult behavior is very ele-gantly demonstrated in avian studies on the role of maternalandrogens and estrogens that are transferred via egg yolk to theembryos (see [22,59]. Experimentally increasing T concentrationin the yolk of eggs of Japanese quails can induce a shift towardsa more proactive coping style of hatchlings irrespective of sex[35]. The described effects of maternal steroids on behavior andphysiology of the chick are mainly short-lasting. A recent studyprovides indications, however, that they can also permanentlyshape the phenotype of the offspring [50]. In this study aggressiveand sexual displays were enhanced one year after a single andro-gen injection in Black-headed gull eggs. Although androgensapparently facilitate adult aggressive behavior, modulation of neo-natal T fails to alter coping styles in SAL and LAL males. Cross-fos-tering experiments and embryo transfer with SAL and LAL malehouse mice further support the suggestion that maternal environ-mental influences are minimal in changing coping styles [117].
The ontogenetic differences in gonadal steroid related mecha-nisms within the perinatal time frame may be related to a differen-tial organization of the testosterone dependent vasopressin (AVP)system. The density of AVP neurons in the bed nucleus of the striaterminalis and its projections to the lateral septum is much higherin adult LAL mice [32]. A study in wild-type Groningen rats con-firmed this finding, showing a much denser AVP innervation ofthe lateral septum in non-aggressive animals [51]. The role of a dif-ferential central nervous AVP regulation regarding coping style willbe discussed in detail in the next chapter.
4.2. HPA-axis during perinatal development
There are numerous studies performed on effects of perinatalstress, maternal care and in specific of corticosterone on adultphysiology and behavior in various animal models and even in hu-mans [56,88,90]. All these studies focus mainly on changes on theemotionality axis in the two-tier model described previously. Vir-tually no studies exist that aimed at the specific effects of perinatalstress or corticosterone on behaviors on the coping style axis. Con-cerning the postnatal period, many studies have clearly demon-strated the powerful effects of early life stressors and/ormaternal care on the expression of adult endocrine, autonomicand behavioral responses to stressors. In general, increased behav-ioral stress responsiveness (anxiety and depressive-like behavioralreactivity) and neuroendocrine stress reactivity (increased HPAreactivity) are observed in adult animals that have experienced ad-verse early life experiences like maternal deprivation or neglect[90,98]. For example, studies by Menard and Hakvoort [92] haveshown that adult rats that received high levels of maternal lickingand grooming in infancy display dramatic reductions in shock-prodburying and aggression relative to offspring of low licking andgrooming mothers. The concurrent decrease in immobility indi-cates that differences in maternal care have little effect on copingstyle of the offspring but rather alters behaviors on the emotional-ity axis [46]. Although circulating glucocorticoid levels in lactatingmothers have been assigned an important role in the induction ofthese programming effects [24,25,101] a recent study of Menardand Hakvoort [92] failed to confirm this hormonal role. A studyof Meerlo et al. [91] showed that offspring of mothers receivingcorticosterone in drinking water during the weaning period wereless aggressive in the resident-intruder test and also respondedwith increased immobility and decreased prod-directed activityin the shock-prod defensive burying test as compared to controltreated rats. This study indicates that neonatal increases in cortico-sterone levels may in fact facilitate the development of adult reac-
314 J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321

tive coping styles. It is obvious that more research is needed to elu-cidate whether and how differences in neonatal corticosterone lev-els or other neuroendocrine factors can affect the central nervoussystem circuits involved in the selection and generation of proac-tive and reactive coping responses. However, ontogenetic studiesemploying embryo transfer, cross-fostering and cross-breedingexperiments do clearly suggest that individual differences in cop-ing styles are already detectable early in life and may have a stronggenetic basis [82,99,117,142].
4.3. Neuroendocrine factors during puberty
Puberty is marked by profound behavioral and neuroendocrinechanges. The activation of the HPG-axis leads to maturation ofreproductive behaviors [108]. Also the HPA-axis sensitivity [103]and the sensitivity of brain areas to corticosterone [86] is appar-ently higher during puberty than in adulthood. The hormonalchanges during puberty are clearly involved in organizing the ado-lescent brain and play a role in individual differences in adultbehavior [113]. The organization effects of hormones during pub-erty are not restricted to sexual behavior, but extend to a varietyof social behaviors like agonistic and anxiety-related behaviors[108,113]. No studies up till now focus on the role of neuroendo-crine maturation during puberty on adult coping style.
5. Neurobiology
It is commonly accepted that the behavioral, neuroendocrineand autonomic physiological (re)activity is the ultimate output ofa complex organization of the brain at the level of the structuraland functional activity in various neuronal networks, neurotrans-mitter systems and the molecular processes of signal transductionin neurons. Hence, animals with different coping styles can also beexpected to show consistent differences in the putative neurobio-logical characteristics that underlie the respective behavioral andphysiological coping responses. Indeed, the past 30 years have wit-nessed an explosion of research in the discipline of affective neuro-science. This has provided evidence that stress coping behaviorsand its associated physiological output are subserved by a basiccircuitry of emotional limbic brain areas including prefrontal cor-tex, nucleus accumbens, amygdala, BNST, septum, hippocampusand hypothalamus and their common output projection nodes. Aschematic presentation of this circuitry is presented in Fig. 7A.The entire functioning of this network is tightly controlled by thebrainstem ascending monoaminergic inputs [36]. A much harderto resolve but intriguingly fundamental question is whether differ-ent brain regions or distinct neurons within these brain regionsunderlie the two different coping styles, or are both coping stylesa function of the same basic brain circuitry? In other words, onepossibility is that two intermingled but distinct subpopulationsof neurons in the same region mediate proactive and reactive cop-ing patterns (i.e., qualitative distinction). Alternatively, the samepopulation of neurons mediates both proactive and reactive copingstyles through different levels or degree of neuromodulation (i.e.,quantitative difference).
Based upon the behavioral activation and inhibition effects ofseveral global regional brain lesion and stimulation experiments,Henry and Stephens [63] provocatively linked distinct brain re-gions to the two divergent coping styles. In their original scheme,the amygdala was associated with the active fight/flight reactionpattern and the hippocampus–septum with the reactive non-aggressive coping response [63]. Obviously, this picture is too sim-ple and with the subsequent advances in neuro-anatomical tracing,lesion, stimulation and imaging techniques, a more complex andrefined neuronal coping circuitry emerged. In particular the recent
studies using immediate-early gene proteins (e.g., Fos, Jun, Zenk,etc.) expression mapping, have revealed the individual neuronsin the various emotional brain regions that become activated dur-ing the expression of different coping-style reactions[2,3,60,71,93,132,136]. For example, in the resident-intruder para-digm proactive fighting residents in comparison to reactive non-fighting residents show an increased number of Fos-positive neu-rons in central amygdala, anterior bed nucleus of the stria termi-nalis, ventrolateral hypothalamus, nucleus accumbens shell,orbital frontal cortex, lateral/ventrolateral periaquaductal grayand dorsal raphe, while in the lateral septum and dorsolateral peri-aquaductal gray a reversed pattern was seen [60,128,132]. Theavailable neuro-anatomical data so far clearly support the viewthat a quantitatively different number of activated neurons withinseveral brain regions that are part of a qualitatively similar neuralcircuit underlie the initiation and execution of proactive/aggressiveand reactive coping style. However, the extent to which similar ordifferent sets of neurons within these brain regions are involved inthe distinct coping styles remains a challenging issue for futurefunctional neuro-anatomical studies.
The functional activity of this neural network, and thereby thetendency to cope either proactively or reactively with environmen-tal challenges, is determined by a wide variety of molecular sub-strates (i.e., neurotransmitters, hormones, cytokines and theirrespective metabolic enzymes, receptors and intraneuronal signal-ing molecules). Indeed, several studies in our wild-type rats andartificially selected SAL and LAL mice show a widespread centralnervous differentiation between proactive and reactive copingstyles, for example at the level of the peptidergic innervation andmodulation of the central nucleus of the amygdala [104], the vaso-pressinergic neurons in the bed nucleus of the stria terminalis andits innervation of the lateral septum [30,51] and the suprachias-matic nucleus [20], the auto-inhibitory control of serotonin neuro-transmission [21,77,127], the hippocampal mossy fiber system[114], and striatal dopaminergic mechanisms [10]. They may beconsidered as a suite of correlated neurobiological trait character-istics, which in concert may determine the tendency to cope eitherproactively or reactively with environmental challenges. With theexception of mainly serotonin and vasopressin (see Fig. 7), the cau-sal involvement of these neurobiological substrates in the individ-ual differentiation in coping style is still far from clear.
5.1. Serotonin
Variability in central serotonin (5-hydroxytryptamine; 5-HT)function is associated with individual differences in affect, temper-ament and personality. For more than 40 years now, this phyloge-netically ancient and anatomically well conserved 5-HT system hasbeen postulated to be essential in the control of aggressive andimpulsive behavioral traits in many animal species, ranging frominvertebrates like fruit flies, crickets and lobsters [81] to verte-brates like lizards [121], fish [97], birds [70], rodents [94] and pri-mates including humans [64,87]. In most animal species, highlevels and/or inappropriate forms of aggressive behavior are gener-ally associated with low levels of brain 5-HT and its metabolite 5-HIAA. This is confirmed in some recent studies in feral mice andrats from our laboratory see [41] for review.
In view of the two-tier model presented above, one may wonderhow serotonin relates to the two dimensions of the model. Numer-ous studies over the past two decades have convincingly shownthat pharmacological compounds that activate 5-HT1A or 5-HT1B
receptor subtypes, potently suppress the display of aggressivebehavior in various animal species ranging from invertebrates, fish,rodents, guinea pigs to primates, including man [39]. Despite thisoverwhelming evidence, it is still a matter of debate which brainarea is the most important site of action of these compounds. A
J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321 315

schematic presentation of a serotonergic neuron its connectionsand receptors is given in Fig. 7B. The 5-HT1A and the 5-HT1B recep-tor are not only found post-synaptically, they have also an impor-tant function in the negative feedback control of the 5-HT neuronitself. The 5-HT1B receptor is present pre-synaptically at the 5-HTaxon terminals, where it inhibits 5-HT release. The 5-HT1A receptorlocated on the soma and dendrites of the serotonergic neuron at
the level of the raphé nuclei acts as an inhibitory auto-receptorreducing the firing of the 5-HT neuron. Hence, at the presynapticlevel these agonists reduce 5-HT signaling, whereas at the levelof the postsynaptic receptors, they mimic the effects of an en-hanced 5-HT signaling. A proper interpretation of the role of sero-tonin in aggression therefore depends on the predominant site ofaction of these 5-HT receptor agonists. Using the compound S-
Fig. 7. (A) Overview of the main brain structures and their connections involved in aggressive behavior. (B) Schematic presentation of a serotonergic neuron and its mostrelevant projections involved in the control of aggression and coping. Abbreviations: AMY = amygdala, AVP = arginine vasopressin, cp = caudate putamen, DRN = dorsal raphenucleus, Glu = glutamate, HAA = hypothalamic attack area, hpc = hippocampus, LS = lateral septum, MRN = medial raphe nucleus, NAcc = nucleus accumbens, ob = olfactorybulb, PAG = periaqueductal gray, PFC = prefrontal cortex, pit = pituitary, Thal = thalamus, VP = ventral pallidum, VTA = ventral tegmental area, 5-HT = serotonin, 5-HTT =serotonin transporter.
316 J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321

15535, which is a selective, full agonist of the somatodendritic 5-HT1A auto-receptor and a partial (ant)agonist of the postsynaptic5-HT1A receptor sites, De Boer [39] showed a potent and behavioralselective reduction in aggressive behavior as well. This means thatthe aggression reducing effect of 5-HT1A agonists is most likely dueto their action on the auto-receptors. Analysis of the dose–re-sponse relationship of S-15535 reveals a 15-fold difference be-tween high- and low-aggressive male rats, i.e. the high-aggressive males have a far more sensitive 5-HT1A auto-receptormediated inhibition. Similarly, using the specific 5-HT1B receptoragonist CP-94253 an almost 20-fold difference in its anti-aggres-sive potency was observed between high- and low-aggressive rats.This strongly enhanced tonic inhibitory control of the serotonergicneuron in the aggressive males may explain the negative correla-tion between baseline levels of 5-HT and 5-HIAA and aggressionfound in many species [39].
In view of the intricate and causal involvement of serotonin inaggressive behavior and the strong individual differentiation inauto-receptor feedback control in relation to aggression as a traitcharacteristic, one may wonder whether serotonin is more gener-ally involved in coping style. Indeed, 5-HT1A and 5-HT1B receptoragonists also reduce swimming behavior in the Porsolt forcedswim test [34] and burying behavior in the defensive burying test[5,38]. So far, these and similar data have been interpreted in termsof emotionality, anxiety or depression. However, one cannot ex-clude the possibility that serotonin is related to the coping styleaxis rather than the emotionality axis. Considering the individualvariation in the behavioral effects of 5-HT1A receptor agonists inthe forced swim test, there is a clear qualitative difference in theway the extremes of the variation react to these compounds. Thereactive coping males show a reduction of immobility and an in-crease in escape attempts, whereas the proactive coping animalshows a decrease in escape and an increase in immobility behavior.In other words, activation of the 5-HT1A auto-receptor leads to areduction of the preferred behavioral response in this test, and toan increase in the alternative behavioral response rather than toa general reduction in anxiety [135]. It is tempting to interpret thisin terms of the two-tier model as presented in Fig. 3. If the seroto-nergic system is related to the coping style axis rather than thestress reactivity axis, one might indeed expect opposite dose re-sponse relations in the extremes of the variation. If serotonin wereassociated with the stress reactivity axis, one would have expecteda decrease in immobility and swimming/escape at the two ends ofthe distribution respectively, without an increase in the alternativeresponse. Preliminary data from our laboratory using the resident-intruder test in rats indeed show opposite dose response relationsin the extremes of the population.
Recently, Valentino and coworkers have proposed that theselection of active or passive coping strategies can also be medi-ated via bimodal effects of the stress-related neuropeptide CRF ininhibiting or enhancing the functional outflow of the dorsal raphe5-HT system targeting limbic and associated structures [125]. Theysupport this novel concept by their own and other findings thatthese qualitatively different functional consequences of CRF aremediated via a distinct expression level or cellular distribution ofCRF1 and CRF2 receptors [139]. CRF1-mediated inhibition andCRF2-mediated excitation of the dorsal raphe 5-HT system facili-tates active and passive coping strategies, respectively [61,89].
5.2. Vasopressin and oxytocin
Besides their important peripheral physiological functions asneurohypophysial released hormones, the neuropeptides argininevasopressin (AVP) and oxytocin (OXT) are also implicated in inter-neuronal communication within various areas of the brain to mod-ulate emotional and social behavioral and physiological
responding [65]. While AVP is generally known to increase anxietylike behaviors, stress and aggressiveness, OXT on the other handhas the opposite effects and facilitates social attachment, careand affiliation [62,68]. Existing data from the pioneering work ofthe late Bohus and de Wied [14,79] already convincingly demon-strated opposite roles for AVP and OXT in fear learning processes.More recent studies in wild-type rats, Roman high- and low-avoid-ance rats and artificially selected SAL and LAL house mice havedemonstrated that non-aggressive males are characterized by afar more extensive vasopressinergic network and a higher vaso-pressin content in several brain regions than high-aggressive males[1,19,31,51]. Interestingly, these differences in density of lateralseptum AVP fibers within males turn out to be as large as the dif-ferences between the sexes. Males are characterized by a consider-ably higher density of lateral septum AVP fibers than females. Sincelow storage levels of neuropeptide (and hence low AVP-immunore-activity and peptide content) may well be linked to enhanced orstimulated release patterns [84,85], this suggests that high-aggres-sive animals exhibit higher levels of release when compared totheir non-aggressive counterparts. Although far less intensely scru-tinized, the closely related ‘‘affiliative” neuropeptide oxytocin doesnot appear to be differently expressed between aggressive andnon-aggressive animals [40].
There is abundant experimental evidence for the suggestion of acausal function of vasopressin in proactive aggressive behavior.Micro-infusion of AVP into the cerebral ventricles [145] or withinthe anterior ventrolateral hypothalamus, BNST or lateral septum[43,52,54,69,73] facilitates offensive aggression in hamsters andrats. In addition, there is a positive correlation of CSF concentra-tions of vasopressin with life history of general aggression as wellas aggression towards individuals [28]. Furthermore, mutant micewith the vasopressin receptor V1B gene, deleted showed virtuallyno offensive aggressive behavior anymore [140,150], althoughtheir defensive aggressiveness remained intact [141]. In addition,offensive aggression is not changed in vasopressin receptor V1a
gene knock-out mice [141]. Consistent with this, systemic as wellas intra-hypothalamic administration of AVP V1A/B receptor antag-onists effectively block offensive aggressive behavior in male ham-sters [12,43,55,53]. We recently replicated this potent anti-aggressive effect of the non-peptidergic AVP V1B selective receptorantagonist SSR149415 in our wild-type rats (Fig. 8).
These findings strongly suggest that increased vasopressinergicactivity in discrete brain regions that are strongly linked to stress-coping regulation is associated with increased levels of aggressive-ness and, perhaps in general, proactive coping styles. On the otherhand, increased oxytocinergic activity in these very same neuralstructures may be implicated more in reactive/passive coping.Therefore, it can be hypothesized that an endogenous balance be-tween vasopressin and oxytocin signaling within (components of)this neural circuit may gate the expression of either proactive orreactive/passive responses to stressors.
One key brain area that importantly controls behavioral andautonomic stress-coping responses, and where vasopressin andoxytocin modulate neuronal activity in opposite ways, is the cen-tral nucleus of the amygdala (CeA) [67] Stressful stimuli increasethe levels of both peptides in the extracellular space of the CeA,possibly due to the release from local synaptic terminals [84]. Fur-thermore, a recent study in rats showed that stress-induced activa-tion of oxytocinergic system in the CeA is implicated in generatingsome of the passive/reactive stress-coping style characteristics[49]. Individual variations in concentrations of these peptidesand/or expression of their receptors within the CeA may thus pro-vide a neuro-physiological mechanism for the selection andexpression of a distinct coping style.
Clearly, the two hypotheses that the two distinct modes of cop-ing are causally associated with distinct neurobiological substrates
J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321 317

(qualitative difference) or different levels or degree of neuromodu-lation (i.e., quantitative difference) needs careful further explora-tion, preferably employing the natural occurring individualvariation within a species in tests that allow a differential expres-sion of coping style.
6. Concluding remarks and future perspectives
Contemporary animal research is characterized by a strong ten-dency to reduce variation. This has attenuated the search for prin-ciples underlying genotypic variation and phenotypic plasticity fora long time. Although this has changed during the last decadeswith studies on gene-environment interactions, there is still a lackof a conceptual framework for individual variation and its function,causes and consequences. This paper has argued that such a frame-work should be based on a thorough understanding of the role ofindividual variation in the evolutionary biology and ecology ofthe species in question (see Fig. 9). It is important to notice thatthe individual variation in behavior and physiology, as observedin nature, is not infinite; nature seems to accept only a limitednumber of phenotypes. An across species comparison suggests thatthere is a certain degree of clustering of individual characteristics.Indeed, several studies show that the concept of discrete copingstyles may hold for quite a number of species [100]. Moreover, ge-netic selection for one behavioral or physiological characteristicusually leads to co-selection of all other characteristics of a givenphenotype as well. For example, genetic selection for proactivebehavior in response to a cold environment (nest building) leadsto higher levels of aggression in a social environment. Conversely,genetic selection for aggression leads to a co-selection of nestbuilding [117]. It is intriguing that genetic selection for one aspect
of proactive coping such as active avoidance behavior [48], sympa-thetic reactivity [11], anxiety [132] and neonatal ultrasonic vocal-ization [18] seems to result in a nearly complete proactivephenotype, irrespective of the exact selection criterion.
With respect to the neuroendocrine differentiation, one has toconsider the possibility that this individual variation is a conse-quence of behavioral differentiation and its related differentiationin metabolic demands and cardiovascular requirements, ratherthan causal trait characteristic. It seems plausible that a trait-likeneuroendocrine differentiation may develop with increasing selec-tion pressure (or continuing artificial genetic selection), that willconsequently become a constraint in phenotypic plasticity. In themodel presented in Fig. 3, coping style is considered as a qualita-tive dimension that is independent from the quantity of the stressresponse. Considering stress reactivity as a dimension separatefrom the type of stress response might explain some of the incon-sistencies observed in the wide variety of genetic selection lines oflaboratory animals. Although these selection lines are generallydeveloped to study the genetic basis of stress reactivity, theymay differ in the selection criterion. Some selection lines have beenselected exclusively on the coping style dimension, whereas othershave been selected on the stress reactivity dimension or a mixturebetween the two. Multidimensional models and better concepts ofindividual variation may help in a better phenotypic characteriza-tion required for a scientifically solid rationale for the selection of acertain strain from the vast amount of laboratory animal strains.
We would argue that the individual variation in behavior andphysiology as it occurs in nature should: (1) serve an importantfunction in nature that (2) can and should be wisely used experi-mentally in the laboratory and (3) might be an important standardto obtain biologically more meaningful and robust results that (4)better translate to the clinical situation. In particular for scientificquestions related to stress vulnerability, it is crucial to avoid selec-tion bias in the early stages of the research. The choice for a certainphenotype for more detailed mechanistic research should ideallybe based on a screening of the full spectrum of natural individualvariation.
References
[1] J.M. Aubry, V. Bartanusz, P. Driscoll, P. Schulz, T. Steimer, J.Z. Kiss,Corticotropin-releasing factor and vasopressin mRNA levels in romanhigh- and low-avoidance rats: response to open-field exposure,Neuroendocrinology 61 (2) (1995) 89–97.
[2] R. Bandler, K.A. Keay, N. Floyd, J. Price, Central circuits mediating patternedautonomic activity during active vs. passive emotional coping, Brain Res. Bull.53 (1) (2000) 95–104.
[3] R. Bandler, J.L. Price, K.A. Keay, Brain mediation of active and passiveemotional coping, Prog. Brain Res. 122 (2000) 333–349.
[4] M. Bardi, A.E. Bode, S.M. Ramirez, L.Y. Brent, Maternal care anddevelopment of stress responses in baboons, Am. J. Primatol. 66 (3)(2005) 263–278.
[5] T. Barf, S.M. Korte, G. Korte-Bouws, C. Sonesson, G. Damsma, B. Bohus, H.Wikstrom, Potential anxiolytic properties of R-(+)-8-OSO2CF3-PAT, a 5-HT 1Areceptor agonist, Eur. J. Pharmacol. 297 (3) (1996) 205–211.
[6] A.M. Bell, Behavioural differences between individuals and two populationsof stickleback (Gasterosteus aculeatus), J. Evol. Biol. 18 (2) (2005) 464–473.
[7] A.M. Bell, A. Sih, Exposure to predation generates personality in threespinedsticklebacks (Gasterosteus aculeatus), Ecol. Lett. 10 (9) (2007) 828–834.
[8] R.F. Benus, J.M. Koolhaas, G.A. Van Oortmerssen, Aggression and adaptation tothe light–dark cycle: role of intrinsic and extrinsic control, Physiol. Behav. 43(1988) 131–137.
[9] R.F. Benus, B. Bohus, J.M. Koolhaas, G.A. Van Oortmerssen, Behaviouralstrategies of aggressive and non-aggressive male mice in active shockavoidance, Behav. Proc. 20 (1989) 1–12.
[10] R.F. Benus, B. Bohus, J.M. Koolhaas, G.A. Van Oortmerssen, Behaviouraldifferences between artificially selected aggressive and non-aggressive mice:response to apomorphine, Behav. Brain Res. 43 (1991) 203–208.
[11] D.F. Berger, J.J. Starzec, Contrasting lever-press avoidance behaviors ofspontaneously hypertensive and normotensive rats (Rattus norvegicus), J.Comp. Psychol. 102 (3) (1988) 279–286.
[12] R.J. Blanchard, G. Griebel, C. Farrokhi, C. Markham, M. Yang, D.C. Blanchard,AVP V1b selective antagonist SSR149415 blocks aggressive behaviors inhamsters, Pharmacol. Biochem. Behav. 80 (1) (2005) 189–194.
0102030405060708090
SSR149415 Vehicle (n=10) 1.0 mg/kg (n=11) 5.0 mg/kg (n=10) 25 mg/kg (n=11)
**
*
*
GroomingInactivityNon-social explore
Socialinteraction
Socialexplore
Offense
*
*
Dur
atio
n (%
tim
e)
Fig. 8. Aggression reducing effect of the AVP V1B selective receptor antagonistSSR149415 (see de Boer et al. (1998), for methodological details). (!p < 0.05).
correlated behaviors
correlated physiologicaland endocrine traits
epigenesisexperience
Genes
survival reproduction dispersalresilience
fitness
Fig. 9. General framework of scientific approach and interpretation.
318 J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321

[13] G.J. Boersma, A.J. Scheurink, P.Y. Wielinga, T.J. Steimer, L. Benthem, Thepassive coping Roman Low Avoidance rat, a non-obese rat model for insulinresistance, Physiol. Behav. 97 (3–4) (2009) 353–358.
[14] B. Bohus, D. de Wied, The vasopressin deficient Brattleboro rats: a naturalknockout model used in the search for CNS effects of vasopressin, Prog. BrainRes. 119 (1998) 555–573.
[15] J.E. Bolhuis, W.G. Schouten, J.A. de Leeuw, J.W. Schrama, V.M. Wiegant,Individual coping characteristics, rearing conditions and behaviouralflexibility in pigs, Behav. Brain Res. 152 (2) (2004) 351–360.
[16] R. Boonstra, C.J. Krebs, N.C. Stenseth, Population cycles in small mammals:the problem of explaining the low phase, Ecology 79 (5) (1998) 1479–1488.
[17] F.H. Bronson, C. Desjardins, Aggression in adult mice: modification byneonatal injections of gonadal hormones, Science 161 (842) (1968) 705–706.
[18] S.A. Brunelli, M.A. Hofer, Selective breeding for infant rat separation-inducedultrasonic vocalizations: developmental precursors of passive and activecoping styles, Behav. Brain Res. 182 (2) (2007) 193–207.
[19] A. Bult, E.A. Van der Zee, J.C. Compaan, C.B. Lynch, Differences in the numberof arginine–vasopressin–immunoreactive neurons exist in thesuprachiasmatic nuclei of house mice selected for differences in nest-building behavior, Brain Res. 578 (1–2) (1992) 335–338.
[20] A. Bult, L. Hiestand, E.A. Van der Zee, C.B. Lynch, Circadian rhythms differbetween selected mouse lines: a model to study the role of vasopressinneurons in the suprachiasmatic nuclei, Brain Res. Bull. 32 (1993) 623–627.
[21] D. Caramaschi, S.F. De Boer, J.M. Koolhaas, Differential role of the 5-HT1Areceptor in aggressive and non-aggressive mice: an across-strain comparison,Physiol. Behav. 90 (4) (2007) 590–601.
[22] C. Carere, J. Balthazart, Sexual versus individual differentiation: thecontroversial role of avian maternal hormones, Trends Endocrinol. Metab.18 (2) (2007) 73–80.
[23] C. Carere, T.G. Groothuis, E. Mostl, S. Daan, J.M. Koolhaas, Fecal corticosteroidsin a territorial bird selected for different personalities: daily rhythm and theresponse to social stress, Horm. Behav. 43 (5) (2003) 540–548.
[24] A. Catalani, P. Casolini, S. Scaccianoce, F.R. Patacchioli, P. Spinozzi, L.Angelucci, Maternal corticosterone during lactation permanently affectsbrain corticosteroid receptors, stress response and behaviour in ratprogeny, Neuroscience 100 (2) (2000) 319–325.
[25] A. Catalani, P. Casolini, G. Cigliana, S. Scaccianoce, C. Consoli, C. Cinque, A.R.Zuena, L. Angelucci, Maternal corticosterone influences behavior, stressresponse and corticosteroid receptors in the female rat, Pharmacol.Biochem. Behav. 73 (1) (2002) 105–114.
[26] M.C. Cervantes, Y. Delville, Individual differences in offensive aggression ingolden hamsters: a model of reactive and impulsive aggression?,Neuroscience 150 (3) (2007) 511–521
[27] D. Chitty, The natural selection of self-regulatory behaviour in animalpopulations, Proc. Ecol. Soc. Aust. 2 (1967) 51–78.
[28] E.F. Coccaro, R.J. Kavoussi, R.L. Hauger, T.B. Cooper, C.F. Ferris, Cerebrospinalfluid vasopressin levels: correlates with aggression and serotonin function inpersonality-disordered subjects, Arch. Gen. Psychiatry 55 (8) (1998) 708–714.
[29] K. Coleman, D. Wilson, Shyness and boldness in pumpkinseed sunfish:individual differences are context-specific, Anim. Behav. 56 (4) (1998) 927–936.
[30] J.C. Compaan, R.M. Buijs, C.W. Pool, A.J.H. De Ruiter, J.M. Koolhaas, Differentiallateral septal vasopressin innervation in aggressive and nonaggressive malemice, Brain Res. Bull. 30 (1992) 1–6.
[31] J.C. Compaan, J.M. Koolhaas, R.M. Buijs, C.W. Pool, A.J. de Ruiter, G.A. VanOortmerssen, Vasopressin and the individual differentiation in aggression inmale house mice, Ann. N.Y. Acad. Sci. 652 (1992) 458–459.
[32] J.C. Compaan, R.M. Buijs, C.W. Pool, A.J. de Ruiter, J.M. Koolhaas, Differentiallateral septal vasopressin innervation in aggressive and nonaggressive malemice, Brain Res. Bull. 30 (1–2) (1993) 1–6.
[33] J.C. Compaan, J.B. Hutchison, A. Wozniak, A.J. de Ruiter, J.M. Koolhaas, Brainaromatase activity and plasma testosterone levels are elevated in aggressivemale mice during early ontogeny, Brain Res. Dev. Brain Res. 82 (1–2) (1994)185–192.
[34] J.F. Cryan, A.M. Redmond, J.P. Kelly, B.E. Leonard, The effects of the 5-HT1Aagonist flesinoxan, in three paradigms for assessing antidepressant potentialin the rat, Eur. Neuropsychopharmacol. 7 (2) (1997) 109–114.
[35] N.J. Daisley, V. Bromundt, E. Mostl, K. Kotrschal, Enhanced yolk testosteroneinfluences behavioral phenotype independent of sex in Japanese quail chicksCoturnix japonica, Horm. Behav. 47 (2) (2005) 185–194.
[36] T. Dalgleish, The emotional brain, Nat. Rev. Neurosci. 5 (7) (2004) 583–589.[37] J.W. Dalley, R.N. Cardinal, T.W. Robbins, Prefrontal executive and cognitive
functions in rodents: neural and neurochemical substrates, Neurosci.Biobehav. Rev. 28 (7) (2004) 771–784.
[38] S.F. De Boer, J.M. Koolhaas, Defensive burying in rodents: ethology,neurobiology and psychopharmacology, Eur. J. Pharmacol. 463 (1-3) (2003)145–161.
[39] S.F. De Boer, J.M. Koolhaas, 5-HT1A and 5-HT1B receptor agonists andaggression: a pharmacological challenge of the serotonin deficiencyhypothesis, Eur. J. Pharmacol. 526 (1–3) (2005) 125–139.
[40] S.F. De Boer, B.J. Van Der Vegt, J.M. Koolhaas, Individual variation inaggression of feral rodent strains: a standard for the genetics of aggressionand violence?, Behav Genet. 33 (5) (2003) 485–501.
[41] S.F. De Boer, D. Caramaschi, D. Natarajan, J.M. Koolhaas, The vicious cycletowards violence: focus on the negative feedback mechanisms of brainserotonin neurotransmission, Front. Behav. Neurosci. 3 (2009) 52.
[42] A.J. de Ruiter, L.E. Feitsma, J.N. Keijser, J.M. Koolhaas, G.A. Van Oortmerssen, B.Bohus, Differential perinatal testosterone secretory capacity of wild housemice testes is related to aggressiveness in adulthood, Horm. Behav. 27 (2)(1993) 231–239.
[43] Y. Delville, K.M. Mansour, C.F. Ferris, Serotonin blocks vasopressin-facilitatedoffensive aggression: interactions within the ventrolateral hypothalamus ofgolden hamsters, Physiol. Behav. 59 (4–5) (1996) 813–816.
[44] N.J. Dingemanse, C. Both, P.J. Drent, J.M. Tinbergen, Fitness consequences ofavian personalities in a fluctuating environment, Proc. Biol. Sci. 271 (1541)(2004) 847–852.
[45] N.J. Dingemanse, F. Van der Plas, J. Wright, D. Reale, M. Schrama, D.A. Roff, E.Van der Zee, I. Barber, Individual experience and evolutionary history ofpredation affect expression of heritable variation in fish personality andmorphology, Proc. Biol. Sci. 276 (1660) (2009) 1285–1293.
[46] J. Diorio, M.J. Meaney, Maternal programming of defensive responses throughsustained effects on gene expression, J. Psychiatry Neurosci. 32 (4) (2007)275–284.
[47] P.J. Drent, O.K. van, A.J. van Noordwijk, Realized heritability of personalities inthe great tit (Parus major), Proc. Biol. Sci. 270 (1510) (2003) 45–51.
[48] P. Driscoll, J. Demek, M. D’Angio, Y. Claustre, B. Scatton, A genetically-based model for divergent stress responses: behavioral, neurochemicaland hormonal aspects, in: V. Pliska, G. Stranzinger (Eds.), FarmAnimals in Biomedical Research, Verlag Paul Parey, Hamburg, 1990,pp. 97–107.
[49] K. Ebner, C.T. Wotjak, R. Landgraf, M. Engelmann, Neuroendocrine andbehavioral response to social confrontation: residents versus intruders, activeversus passive coping styles, Horm. Behav. 47 (1) (2005) 14–21.
[50] C.M. Eising, W. Muller, T.G. Groothuis, Avian mothers create differentphenotypes by hormone deposition in their eggs, Biol. Lett. 2 (1) (2006)20–22.
[51] H.G. Everts, A.J. de Ruiter, J.M. Koolhaas, Differential lateral septal vasopressinin wild-type rats: correlation with aggression, Horm. Behav. 31 (2) (1997)136–144.
[52] C.F. Ferris, Vasopressin/oxytocin and aggression, Novartis Found. Symp. 268(2005) 190–198.
[53] C.F. Ferris, M. Potegal, Vasopressin receptor blockade in the anteriorhypothalamus suppresses aggression in hamsters, Physiol. Behav. 44 (2)(1988) 235–239.
[54] C.F. Ferris, H.E. Albers, S.M. Wesolowski, B.D. Goldman, S.E. Luman,Vasopressin injected into the hypothalamus triggers a stereotypic behaviorin golden hamsters, Science 224 (4648) (1984) 521–523.
[55] C.F. Ferris, S.F. Lu, T. Messenger, C.D. Guillon, N. Heindel, M. Miller, G. Koppel,B.F. Robert, N.G. Simon, Orally active vasopressin V1a receptor antagonist,SRX251, selectively blocks aggressive behavior, Pharmacol. Biochem. Behav.83 (2) (2006) 169–174.
[56] F. Fumagalli, R. Molteni, G. Racagni, M.A. Riva, Stress during development:impact on neuroplasticity and relevance to psychopathology, Prog. Neurobiol.81 (4) (2007) 197–217.
[57] G.A. Van Oortmerssen, J. Busser, Studies in wild house mice III: disruptiveselection on aggression as a possible force in evolution, in: P.F. Brain, D.Mainardi, S. Parmigiani (Eds.), House Mouse Aggression: A Model ForUnderstanding the Evolution of Social Behavior, Harwood Acad. Publ., Chur,1989, pp. 87–117.
[58] L. Gould, T.E. Ziegler, Variation in fecal testosterone levels, inter-maleaggression, dominance rank and age during mating and post-matingperiods in wild adult male ring-tailed lemurs (Lemur catta), Am. J. Primatol.(2007).
[59] T.G. Groothuis, W. Muller, N. von Engelhardt, C. Carere, C. Eising, Maternalhormones as a tool to adjust offspring phenotype in avian species, Neurosci.Biobehav. Rev. 29 (2) (2005) 329–352.
[60] J. Haller, M. Toth, J. Halasz, S.F. De Boer, Patterns of violent aggression-induced brain c-fos expression in male mice selected for aggressiveness,Physiol. Behav. 88 (1–2) (2006) 173–182.
[61] S.E. Hammack, K.J. Richey, M.J. Schmid, M.L. LoPresti, L.R. Watkins, S.F. Maier,The role of corticotropin-releasing hormone in the dorsal raphe nucleus inmediating the behavioral consequences of uncontrollable stress, J. Neurosci.22 (3) (2002) 1020–1026.
[62] M. Heinrichs, B. von Dawans, G. Domes, Oxytocin, vasopressin, and humansocial behavior, Front. Neuroendocrinol. 30 (4) (2009) 548–557.
[63] J.P. Henry, P.M. Stephens, Stress, Health and the Social Environment: ASociobiological Approach to Medicine, Springer, Berlin, 1977.
[64] J.D. Higley, P.T. Mehlman, D.M. Taub, S.B. Higley, S.J. Suomi, J.H. Vickers, M.Linnoila, Cerebrospinal fluid monoamine and adrenal correlates ofaggression in free-ranging rhesus monkeys, Arch. Gen. Psychiatry 49 (6)(1992) 436–441.
[65] A. Holmes, M. Heilig, N.M. Rupniak, T. Steckler, G. Griebel, Neuropeptidesystems as novel therapeutic targets for depression and anxiety disorders,Trends Pharmacol. Sci. 24 (11) (2003) 580–588.
[66] B. Hornfeldt, T. Hipkiss, U. Eklund, Fading out of vole and predator cycles?,Proc Biol. Sci. 272 (1576) (2005) 2045–2049.
[67] D. Huber, P. Veinante, R. Stoop, Vasopressin and oxytocin excite distinctneuronal populations in the central amygdala, Science 308 (5719) (2005)245–248.
[68] T.R. Insel, J.T. Winslow, Z. Wang, L.J. Young, Oxytocin, vasopressin, and theneuroendocrine basis of pair bond formation, Adv. Exp. Med. Biol. 449 (1998)215–224.
J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321 319

[69] R.W. Irvin, P. Szot, D.M. Dorsa, M. Potegal, C.F. Ferris, Vasopressin in the septalarea of the golden hamster controls scent marking and grooming, Physiol.Behav. 48 (5) (1990) 693–699.
[70] M. Ison, C. Fachinelli, E.L. Rodriguez Echandia, Effect of the i.c.v. injection of5,7-di-hydroxytryptamine on the aggressive behavior of dominant andsubmissive pigeons (Columba livia), Pharmacol. Biochem. Behav. 53 (4)(1996) 951–955.
[71] K.A. Keay, R. Bandler, Parallel circuits mediating distinct emotional copingreactions to different types of stress, Neurosci. Biobehav. Rev. 25 (7–8) (2001)669–678.
[72] J.M. Koolhaas, S.F. de Boer, A.J.H. De Ruiter, P. Meerlo, A. Sgoifo, Social stress inrats and mice, Acta Physiol. Scand. 161 (S640) (1997) 69–72.
[73] J.M. Koolhaas, H. Everts, A.J. de Ruiter, S.F. De Boer, B. Bohus, Coping withstress in rats and mice: differential peptidergic modulation of the amygdala–lateral septum complex, Prog. Brain Res. 119 (1998) 437–448.
[74] J.M. Koolhaas, S.M. Korte, S.F. De Boer, B.J. Van Der Vegt, C.G. Van Reenen, H.Hopster, I.C. De Jong, M.A. Ruis, H.J. Blokhuis, Coping styles in animals:current status in behavior and stress-physiology, Neurosci. Biobehav. Rev. 23(7) (1999) 925–935.
[75] J.M. Koolhaas, S.F. De Boer, B. Buwalda, C.G. Van Reenen, Individual variationin coping with stress: a multidimensional approach of ultimate andproximate mechanisms, Brain Behav. Evol. 70 (4) (2007).
[76] S.M. Korte, G.A. Bouws, J.M. Koolhaas, B. Bohus, Neuroendocrine andbehavioral responses during conditioned active and passive behavior in thedefensive burying/probe avoidance paradigm: effects of ipsapirone, Physiol.Behav. 52 (2) (1992) 355–361.
[77] S.M. Korte, O.C. Meijer, E.R. De Kloet, B. Buwalda, J. Keijser, F. Sluyter, G.A. VanOortmerssen, B. Bohus, Enhanced 5-HT1a receptor expression in forebrainregions of aggressive house mice, Brain Res. 736 (1996) 338–343.
[78] S.M. Korte, J.M. Koolhaas, J.C. Wingfield, B.S. McEwen, The Darwinian conceptof stress: benefits of allostasis and costs of allostatic load and the trade-offs inhealth and disease, Neurosci. Biobehav. Rev. 29 (1) (2005) 3–38.
[79] G.L. Kovacs, D. de Wied, Peptidergic modulation of learning and memoryprocesses, Pharmacol. Rev. 46 (3) (1994) 269–291.
[80] S. Kralj-Fiser, I.B. Scheiber, A. Blejec, E. Moestl, K. Kotrschal, Individualities ina flock of free-roaming greylag geese: behavioral and physiologicalconsistency over time and across situations, Horm. Behav. 51 (2) (2007)239–248.
[81] E.A. Kravitz, R. Huber, Aggression in invertebrates, Curr. Opin. Neurobiol. 13(6) (2003) 736–743.
[82] K.G. Lambert, K. Tu, A. Everette, G. Love, I. McNamara, M. Bardi, C.H. Kinsley,Explorations of coping strategies, learned persistence, and resilience in Long-Evans rats: innate versus acquired characteristics, Ann. N.Y. Acad. Sci. 1094(2006) 319–324.
[83] X. Lambin, V. Bretagnolle, N.G. Yoccoz, Vole population cycles in northern andsouthern Europe: is there a need for different explanations for singlepattern?, J Anim. Ecol. 75 (2) (2006) 340–349.
[84] R. Landgraf, I.D. Neumann, Vasopressin and oxytocin release within thebrain: a dynamic concept of multiple and variable modes ofneuropeptide communication, Front. Neuroendocrinol. 25 (3–4) (2004)150–176.
[85] R. Landgraf, C.T. Wotjak, I.D. Neumann, M. Engelmann, Release of vasopressinwithin the brain contributes to neuroendocrine and behavioral regulation,Prog. Brain Res. 119 (1998) 201–220.
[86] P.R. Lee, D. Brady, J.I. Koenig, Corticosterone alters N-methyl-D-aspartatereceptor subunit mRNA expression before puberty, Brain Res. Mol. Brain Res.115 (1) (2003) 55–62.
[87] K.P. Lesch, U. Merschdorf, Impulsivity, aggression, and serotonin: a molecularpsychobiological perspective, Behav. Sci. Law 18 (5) (2000) 581–604.
[88] S. Maccari, M. Darnaudery, S. Morley-Fletcher, A.R. Zuena, C. Cinque, O. vanReeth, Prenatal stress and long-term consequences: implications ofglucocorticoid hormones, Neurosci. Biobehav. Rev. 27 (1–2) (2003) 119–127.
[89] S.F. Maier, L.R. Watkins, Stressor controllability and learned helplessness: theroles of the dorsal raphe nucleus, serotonin, and corticotropin-releasingfactor, Neurosci. Biobehav. Rev. 29 (4–5) (2005) 829–841.
[90] M.J. Meaney, Maternal care, gene expression, and the transmission ofindividual differences in stress reactivity across generations, Annu. Rev.Neurosci. 24 (2001) 1161–1192.
[91] P. Meerlo, K.M. Horvath, P.G. Luiten, L. Angelucci, A. Catalani, J.M. Koolhaas,Increased maternal corticosterone levels in rats: effects on brain 5-HT1Areceptors and behavioral coping with stress in adult offspring, Behav.Neurosci. 115 (5) (2001) 1111–1117.
[92] J.L. Menard, R.M. Hakvoort, Variations of maternal care alter offspring levelsof behavioural defensiveness in adulthood: evidence for a threshold model,Behav. Brain Res. 176 (2) (2007) 302–313.
[93] J.L. Menard, D.L. Champagne, M.J. Meaney, Variations of maternal caredifferentially influence ‘fear’ reactivity and regional patterns of cFosimmunoreactivity in response to the shock-probe burying test,Neuroscience 129 (2) (2004) 297–308.
[94] K.A. Miczek, E.W. Fish, J.F. De Bold, R.M. De Almeida, Social and neuraldeterminants of aggressive behavior: pharmacotherapeutic targets atserotonin, dopamine and gamma-aminobutyric acid systems,Psychopharmacology (Berl.) 163 (3–4) (2002) 434–458.
[95] K. Norrdahl, Population cycles in northern small mammals, Biol. Rev. Camb.Philos. Soc. 70 (4) (1995) 621–637.
[96] G.A. Van Oortmerssen, R.F. Benus, F. Sluyter, Studies on wild house mice IV:on the heredity of testosterone and readiness to attack, Aggressive Behav.(1991) 18.
[97] O. Overli, C.A. Harris, S. Winberg, Short-term effects of fights for socialdominance and the establishment of dominant–subordinate relationships onbrain monoamines and cortisol in rainbow trout, Brain Behav. Evol. 54 (5)(1999) 263–275.
[98] P.M. Plotsky, M.J. Meaney, Early, postnatal experience alters hypothalamiccorticotropin-releasing factor (CRF) mRNA, median eminence CRF contentand stress-induced release in adult rats, Brain Res. Mol. Brain Res. 18 (3)(1993) 195–200.
[99] P. Prakash, Z. Merali, M. Kolajova, B.M. Tannenbaum, H. Anisman, Maternalfactors and monoamine changes in stress-resilient and susceptible mice:cross-fostering effects, Brain Res. 1111 (1) (2006) 122–133.
[100] D. Reale, S.M. Reader, D. Sol, P.T. McDougall, N.J. Dingemanse, Integratinganimal temperament within ecology and evolution, Biol. Rev. Camb. Philos.Soc. 82 (2) (2007) 291–318.
[101] S.L. Rees, S. Panesar, M. Steiner, A.S. Fleming, The effects of adrenalectomyand corticosterone replacement on maternal behavior in the postpartum rat,Horm. Behav. 46 (4) (2004) 411–419.
[102] S.H. Richter, J.P. Garner, H. Wurbel, Environmental standardization: cure orcause of poor reproducibility in animal experiments?, Nat Methods 6 (4)(2009) 257–261.
[103] R.D. Romeo, S.J. Lee, B.S. McEwen, Differential stress reactivity in intact andovariectomized prepubertal and adult female rats, Neuroendocrinology 80(6) (2004) 387–393.
[104] B. Roozendaal, A. Wiersma, P. Driscoll, J.M. Koolhaas, B. Bohus,Vasopressinergic modulation of stress responses in the central amygdala ofthe Roman high-avoidance and low-avoidance rat, Brain Res. 596 (1992) 35–40.
[105] W.M.S. Russell, R.L. Burch, The Principles of Humane ExperimentalTechnique, Methuen, London, 1959.
[106] I. Saccheri, I. Hanski, Natural selection and population dynamics, Trends Ecol.Evol. 21 (6) (2006) 341–347.
[107] J. Schjolden, T. Backstrom, K.G. Pulman, T.G. Pottinger, S. Winberg, Divergencein behavioural responses to stress in two strains of rainbow trout(Oncorhynchus mykiss) with contrasting stress responsiveness, Horm. Behav.48 (5) (2005) 537–544.
[108] K.M. Schulz, C.L. Sisk, Pubertal hormones, the adolescent brain, and thematuration of social behaviors: lessons from the Syrian hamster, Mol. CellEndocrinol. 254–255 (2006) 120–126.
[109] T. Schuurman, Hormonal correlates of agonistic behavior in adult male rats,Prog. Brain Res. 53 (1980) 415–420.
[110] A. Sih, A. Bell, J.C. Johnson, Behavioral syndromes: an ecological andevolutionary overview, Trends Ecol. Evol. 19 (7) (2004) 372–378.
[111] A. Sih, A.M. Bell, J.C. Johnson, R.E. Ziemba, Behavioral syndromes: anintegrative overview, Q. Rev. Biol. 79 (3) (2004) 241–277.
[112] N.G. Simon, Q. Mo, S. Hu, C. Garippa, S.F. Lu, Hormonal pathways regulatingintermale and interfemale aggression, Int. Rev. Neurobiol. 73 (2006) 99–123.
[113] C.L. Sisk, J.L. Zehr, Pubertal hormones organize the adolescent brain andbehavior, Front. Neuroendocrinol. 26 (3–4) (2005) 163–174.
[114] F. Sluyter, L. Jamot, G.A. Van Oortmerssen, W.E. Crusio, Hippocampal mossyfiber distribution in mice selected for aggression, Brain Res. 646 (1994) 145–148.
[115] F. Sluyter, A. Bult, C.B. Lynch, G.A. Van Oortmerssen, J.M. Koolhaas, Acomparison between house mouse lines selected for attack latency or nest-building: evidence for a genetic basis for alternative behavioral strategies,Behav. Genet. 25 (1995) 247–252.
[116] F. Sluyter, S.M. Korte, B. Bohus, G.A. Van Oortmerssen, Behavioral stressresponse of genetically selected aggressive and nonaggressive wild housemice in the shock-probe/defensive burying test, Pharmacol. Biochem. Behav.54 (1996) 113–116.
[117] F. Sluyter, J.J. van der Vlucht, G.A. Van Oortmerssen, J.M. Koolhaas, F. van derHoeven, P. de Boer, Studies on wild house mice. VII. Prenatal maternalenvironment and aggression, Behav. Genet. 26 (5) (1996) 513–518.
[118] T.S. Sperry, D.W. Wacker, J.C. Wingfield, The role of androgen receptors inregulating territorial aggression in male song sparrows, Horm. Behav. 57 (1)(2010) 86–95.
[119] T. Steimer, P. Driscoll, Divergent stress responses and coping styles inpsychogenetically selected Roman high-(RHA) and low-(RLA) avoidance rats:behavioural, neuroendocrine and developmental aspects, Stress 6 (2) (2003)87–100.
[120] T. Steimer, F.S. La Fleur, P.E. Schulz, Neuroendocrine correlates of emotionalreactivity and coping in male rats from the Roman high (RHA/Verh)- and low(RLA/Verh)-avoidance lines, Behav. Genet. 27 (6) (1997) 503–512.
[121] C.H. Summers, W.J. Korzan, J.L. Lukkes, M.J. Watt, G.L. Forster, O. Overli, E.Hoglund, E.T. Larson, P.J. Ronan, J.M. Matter, T.R. Summers, K.J. Renner, N.Greenberg, Does serotonin influence aggression? Comparing regional activitybefore and during social interaction, Physiol. Biochem. Zool. 78 (5) (2005)679–694.
[122] B.C. Trainor, K.M. Greiwe, R.J. Nelson, Individual differences in estrogenreceptor alpha in select brain nuclei are associated with individualdifferences in aggression, Horm. Behav. 50 (2) (2006) 338–345.
[123] B.C. Trainor, H.H. Kyomen, C.A. Marler, Estrogenic encounters: howinteractions between aromatase and the environment modulate aggression,Front. Neuroendocrinol. 27 (2) (2006) 170–179.
320 J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321

[124] D. Treit, Animal models for the study of anti-anxiety agents: a review,Neurosci. Biobehav. Rev. 9 (1989) 203–222.
[125] R.J. Valentino, I. Lucki, E. van Bockstaele, Corticotropin-releasing factor in thedorsal raphe nucleus: linking stress coping and addiction, Brain Res. (2009).
[126] F.J. Van der Staay, T. Steckler, Behavioural phenotyping of mouse mutants,Behav. Brain Res. 125 (1-2) (2001) 3–12.
[127] B.J. Van Der Vegt, S.F. De Boer, B. Buwalda, A.J. de Ruiter, J.G. de Jong, J.M.Koolhaas, Enhanced sensitivity of postsynaptic serotonin-1A receptors in ratsand mice with high trait aggression, Physiol. Behav. 74 (1–2) (2001) 205–211.
[128] B.J. Van Der Vegt, N. Lieuwes, E.H. van de Wall, K. Kato, L. Moya-Albiol, S.Martinez-Sanchis, S.F. De Boer, J.M. Koolhaas, Activation of serotonergicneurotransmission during the performance of aggressive behavior in rats,Behav. Neurosci. 117 (4) (2003) 667–674.
[129] C.G. Van Reenen, J.T. Van der Werf, R.M. Bruckmaier, H. Hopster, B. Engel, J.P.Noordhuizen, H.J. Blokhuis, Individual differences in behavioral andphysiological responsiveness of primiparous dairy cows to machinemilking, J. Dairy Sci. 85 (10) (2002) 2551–2561.
[130] C.G. Van Reenen, B. Engel, L.F.M. Ruis-Heutinck, J.T.N. Van der Werf, W.G.Buist, R.B. Jones, H.J. Blokhuis, Behavioural reactivity of heifer calves inpotentially alarming test situations: a multivariate and correlational analysis,Appl. Anim. Behav. Sci. 85 (1–2) (2004) 11–30.
[131] C.G. Van Reenen, N.E. O’Connell, J.T. Van der Werf, S.M. Korte, H. Hopster, R.B.Jones, H.J. Blokhuis, Responses of calves to acute stress: individualconsistency and relations between behavioral and physiological measures,Physiol. Behav. 85 (5) (2005) 557–570.
[132] A.H. Veenema, I.D. Neumann, Neurobiological mechanisms of aggression andstress coping: a comparative study in mouse and rat selection lines, BrainBehav. Evol. 70 (4) (2007) 274–285.
[133] A.H. Veenema, O.C. Meijer, E.R. De Kloet, J.M. Koolhaas, B.G. Bohus,Differences in basal and stress-induced HPA regulation of wild housemice selected for high and low aggression, Horm. Behav. 43 (1) (2003)197–204.
[134] A.H. Veenema, J.M. Koolhaas, E.R. De Kloet, Basal and stress-induceddifferences in HPA axis, 5-HT responsiveness, and hippocampal cellproliferation in two mouse lines, Ann. N.Y. Acad. Sci. 1018 (2004) 255–265.
[135] A.H. Veenema, T.I. Cremers, M.E. Jongsma, P.J. Steenbergen, S.F. De Boer, J.M.Koolhaas, Differences in the effects of 5-HT(1A) receptor agonists on forcedswimming behavior and brain 5-HT metabolism between low and highaggressive mice, Psychopharmacology (Berl.) 178 (2–3) (2005) 151–160.
[136] J.G. Veening, L.M. Coolen, T.R. de Jong, H.W. Joosten, S.F. De Boer, J.M.Koolhaas, B. Olivier, Do similar neural systems subserve aggressive andsexual behaviour in male rats? Insights from c-Fos and pharmacologicalstudies, Eur. J. Pharmacol. 526 (1–3) (2005) 226–239.
[137] M.E.M. Verbeek, P.J. Drent, P.R. Wiepkema, Consistent individual differencesin early exploratory behavior of male great tits, Anim. Behav. 48 (1994)1113–1121.
[138] F.R. Walker, L.M. Masters, R.A. Dielenberg, T.A. Day, Coping with defeat: acuteglucocorticoid and forebrain responses to social defeat vary with defeatepisode behaviour, Neuroscience 162 (2) (2009) 244–253.
[139] M. Waselus, C. Nazzaro, R.J. Valentino, E.J. Van Bockstaele, Stress-inducedredistribution of corticotropin-releasing factor receptor subtypes in thedorsal raphe nucleus, Biol. Psychiatry 66 (1) (2009) 76–83.
[140] S.R. Wersinger, E.I. Ginns, A.M. O’Carroll, S.J. Lolait, W.S. Young III,Vasopressin V1b receptor knockout reduces aggressive behavior in malemice, Mol. Psychiatry 7 (9) (2002) 975–984.
[141] S.R. Wersinger, H.K. Caldwell, M. Christiansen, W.S. Young III, Disruption ofthe vasopressin 1b receptor gene impairs the attack component of aggressivebehavior in mice, Genes Brain Behav. 6 (7) (2007) 653–660.
[142] A. Wigger, P. Loerscher, P. Weissenbacher, F. Holsboer, R. Landgraf, Cross-fostering and cross-breeding of HAB and LAB rats: a genetic rat model ofanxiety, Behav. Genet. 31 (4) (2001) 371–382.
[143] J.C. Wingfield, R.E. Hegner, A.M. Dufty, G.F. Ball, The ‘‘Challenge Hypothesis”:theoretical implications for patterns of testosterone secretion, matingsystems, and breeding strategies, Am. Nat. 136 (1990) 829–846.
[144] J.C. Wingfield, S. Lynn, K.K. Soma, Avoiding the ‘costs’ of testosterone:ecological bases of hormone–behavior interactions, Brain Behav. Evol. 57 (5)(2001) 239–251.
[145] J.T. Winslow, T.R. Insel, Effects of central vasopressin administration to infantrats, Eur. J. Pharmacol. 233 (1) (1993) 101–107.
[146] M. Wolf, G.S. van Doorn, O. Leimar, F.J. Weissing, Life-history trade-offsfavour the evolution of animal personalities, Nature 447 (7144) (2007) 581–584.
[147] M. Wolf, G.S. van Doorn, F.J. Weissing, Evolutionary emergence of responsiveand unresponsive personalities, Proc. Natl. Acad. Sci. USA 105 (41) (2008)15825–15830.
[148] H. Wurbel, Behaviour and the standardization fallacy, Nat. Genet. 26 (3)(2000) 263.
[149] H. Wurbel, Behavioral phenotyping enhanced–beyond (environmental)standardization, Genes Brain Behav. 1 (1) (2002) 3–8.
[150] L.J. Young, R. Nilsen, K.G. Waymire, G.R. MacGregor, T.R. Insel, Increasedaffiliative response to vasopressin in mice expressing the V1a receptor from amonogamous vole, Nature 400 (6746) (1999) 766–768.
[151] T.Y. Zhang, R. Bagot, C. Parent, C. Nesbitt, T.W. Bredy, C. Caldji, E. Fish, H.Anisman, M. Szyf, M.J. Meaney, Maternal programming of defensiveresponses through sustained effects on gene expression, Biol. Psychol. 73(1) (2006) 72–89.
J.M. Koolhaas et al. / Frontiers in Neuroendocrinology 31 (2010) 307–321 321


















