
FIRST BIOCHEMISTRY EXAM Tuesday 25/10/2016 10-11 40 …
45
FIRST BIOCHEMISTRY EXAM Tuesday 25/10/2016 10-11 40 MCQs. Location : 102, 105, 106, 301, 302
Transcript of FIRST BIOCHEMISTRY EXAM Tuesday 25/10/2016 10-11 40 …

FIRST BIOCHEMISTRY EXAM
Tuesday 25/10/2016
10-11
40 MCQs.
Location : 102, 105, 106, 301, 302

The Behavior of Proteins: Enzymes,
Mechanisms, and Control

General theory of enzyme action, by
Leonor Michaelis and Maud Menten in 1913. Leonor Michaelis, German 1875–1949
Maud Menten, Canadian 1879–1960

Learning Objectives
1. What Is the Michaelis–Menten Approach to Enzyme Kinetics? 2. How Do Enzymatic Reactions Respond to Inhibitors? 3. Does the Michaelis–Menten Model Describe the Behavior of Allosteric Enzymes ? 4. What Are the Models for the Behavior of Allosteric Enzymes? 5. What is Lineweaver –Burk plot ? 5 . How Does Phosphorylation of Specific Residues Regulate Enzyme Activity? 6. What Are Zymogens, and How Do They Control Enzyme Activity? 7. What Are Coenzymes and Cofactors ? 8. What Are Isoenzymes & what is their clinical application.

They postulated that the enzyme first combines reversibly with its substrate to form an enzyme-substrate complex in a relatively fast reversible step:
The ES complex then breaks down in a slower second step to yield the free enzyme and the reaction product P:


Michaelis-Menten Model
2
V max [S] V =
K M + [S]
Michaelis-Menten equation

Michaelis-Menten Model –When [S]= KM, the equation reduces to
Vmax [S]V =
KM + [S]=
Vmax [S]
[S] + [S]=
Vmax
2
2

Michaelis-Menten Model
– It is difficult to determine Vmax experimentally
– The equation for a hyperbola (E saturation curve)
– Can be transformed into the equation for a straight line by taking the reciprocal of each side
V1 =
KM + [S]
Vmax [S]=
KM [S]
Vmax [S] Vmax [S]+
V1 =
KM
Vmax [S] Vmax
+ 1
Vmax [S]V =
KM + [S](an equation for a hyperbola)

The physiological consequence of KM is illustrated by the sensitivity of some individuals to ethanol. Such persons exhibit facial flushing and rapid heart rate (tachycardia) after ingesting even small amounts of alcohol.
In the liver, alcohol dehydrogenase converts ethanol into acetaldehyde

Normally, the acetaldehyde, which is the cause of the symptoms when present at high concentrations, is processed to acetate by acetaldehyde dehydrogenase.

Most people have two forms of the acetaldehyde dehydrogenase, a low KM mitochondrial form and a high KM cytosolic form. In susceptible persons, the mitochondrial enzyme is less active due to the substitution of a single amino acid, and acetaldehyde is processed only by the cytosolic enzyme. Because this enzyme has a high KM, less acetaldehyde is converted into acetate; excess acetaldehyde escapes into the blood and accounts for the physiological effects.

Lineweaver-Burk Plot
– which has the form y = mx + b, and is the formula for a straight line (linearization)
– a plot of 1/V versus 1/[S] will give a straight line
with slope of KM/Vmax and y intercept of 1/Vmax
– such a plot is known as a Lineweaver-Burk double reciprocal plot
V
1 =Vmax
+ 1
Vmax [S]
1
y m x + b
V
1 =KM
•
= •

Lineweaver-Burk Plot
V1
[S]1
x intercept =
y intercept =1
Vmax
-1
KM
slope =KM
Vmax
– KM is the dissociation constant for ES; the greater the value of KM, the less tightly S is bound (less affinity) to E
The units KM are units of [S] that is M or mM

Turnover number
• Turnover number (kcat): is the moles of S converted to P per mole of E per second.
–Unit of kcat is mol S*mol E-1*s-1
–The higher Kcat value the higher the reactivity of the E

Turnover Numbers and KM Values for some typical enzymes
Enzyme Function
Catalase C onversion of H 2 O 2 to H 2 O + O 2
4 x 10 7 25
Carbonic anhydrase
Hydration of CO 2 1 x 10 6 12
Acetylcholin- esterase
Regeneration of acetylcholine
1.4 x 10 4 9.5 x 10 -2
Chymotrypsin Proteolytic enzyme 1.9.x 10 2 6.6 x 10 -1
Lysozyme Hydrolysis of bacterial cell wall polysaccharides
0.5 6 x 10 -3
K M
(mmol•liter -1
)
Turnover numbr
[(mol S)•(mol E) -1
•s -1
]

Enzyme Inhibition
• Reversible inhibitor: a substance that binds to an enzyme to inhibit it, but can be released – competitive inhibitor: binds to the active (catalytic)
site and blocks access to it by substrate – noncompetitive inhibitor: binds to a site other than
the active site; inhibits the enzyme by changing its conformation
• Irreversible inhibitor: a substance that causes inhibition that cannot be reversed – usually involves formation or breaking of covalent
bonds to or on the enzyme – Cyanides, Penicillin, Heavy metals, Nerve gas

competitive inhibitor
A competitive inhibitor diminishes the rate of
catalysis by reducing the proportion of enzyme molecules bound to a substrate. At any given inhibitor concentration, competitive inhibition can be relieved by increasing the substrate concentration. Under these conditions, the substrate "outcompetes" the inhibitor for the active site.

Methotrexate is a structural analog of tetrahydrofolate, a coenzyme for the enzyme dihydrofolate reductase, which plays a role in the biosynthesis of purines and pyrimidines .
It binds to dihydrofolate reductase 1000-fold more tightly than the natural substrate and inhibits nucleotide base synthesis. It is used to treat cancer.

The cofactor tetrahydrofolate and its structural analog
methotrexate. Regions with structural differences are shown in red.

noncompetitive inhibitor
In noncompetitive inhibition, which also is reversible, the inhibitor and substrate can bind simultaneously to an enzyme molecule at different binding sites . A noncompetitive inhibitor acts by decreasing the turnover number rather than by diminishing the proportion of enzyme molecules that are bound to substrate. Noncompetitive inhibition, in
contrast with competitive inhibition, cannot be overcome by increasing the substrate concentration.

Competitive Vs noncompetitive Inhibition

Competitive Inhibition Illustrated on a Double-Reciprocal Plot. A double-reciprocal plot of enzyme kinetics in the presence ( I ) and absence ( I ) of a competitive inhibitor illustrates that the
inhibitor has no effect on V max but increases K M.

Noncompetitive Inhibition
Illustrated on a Double-Reciprocal Plot. A double-reciprocal plot of enzyme kinetics in the presence ( I ) and absence ( I ) of a noncompetitive inhibitor shows that K M is unaltered and V max is decreased.

Irreversible inhibitor
Penicillin acts by covalently modifying the enzyme transpeptidase, thereby preventing the synthesis of bacterial cell walls and thus killing the bacteria .
Aspirin acts by covalently modifying the
enzyme cyclooxygenase, reducing the synthesis of inflammatory signals.

Structure of Penicillin
Formation of a Penicilloyl-Enzyme Complex.

Chymotrypsin Inhibition
Treatment with organofluorophosphates such as
diisopropylphosphofluoridate (DIPF) was found to inactivate the enzyme irreversibly .
Despite the fact that the enzyme possesses 28 serine residues, only one, serine 195, was modified, resulting in a total loss of enzyme activity. This chemical modification reaction suggested that this unusually reactive serine residue plays a central role in the catalytic mechanism of chymotrypsin.

An Unusually Reactive Serine in Chymotrypsin. Chymotrypsin is inactivated by treatment with diisopropylphosphofluoridate (DIPF), which reacts only with serine 195 among 28 possible serine residues

Enzyme regulation
Enzyme activity can be regulated by several ways: 1. Covalent modification (Phosphorylation) The side chain -OH groups of Ser, Thr, and Tyr can form
phosphate esters phosphorylation by ATP can convert an inactive
precursor into an active enzyme Protein kinases phosphorylate enzymes Protein phosphatases remove phosphate
groups

Enzyme regulation
Covalent modification (Phosphorylation)

2. Noncovalent modification (Allosteric regulation)
Using allosteric effectors (activator and allosteric inhibitor )
3. Control amount of enzyme: enzyme synthesis: gene regulation enzyme degradation

4. Zymogen : an inactive precursor of an enzyme; cleavage of one or more covalent bonds transforms it into the active enzyme
– Involved in blood clotting and digestion
– Example chymotrypsinogen
Chymotrypsinogen
– synthesized and stored in the pancreas
– a single polypeptide chain of 245 amino acid residues cross linked by five disulfide (-S-S-) bonds
– enzyme trypsin cleaves it to give chymotrypsin

Secretion of Zymogens by an Acinar Cell of the Pancreas

Proteolytic Activation of Chymotrypsinogen. The three chains of α-chymotrypsin are linked by two interchain disulfide bonds

Zymogen Activation by Proteolytic Cleavage
Active enzymes are shown in yellow
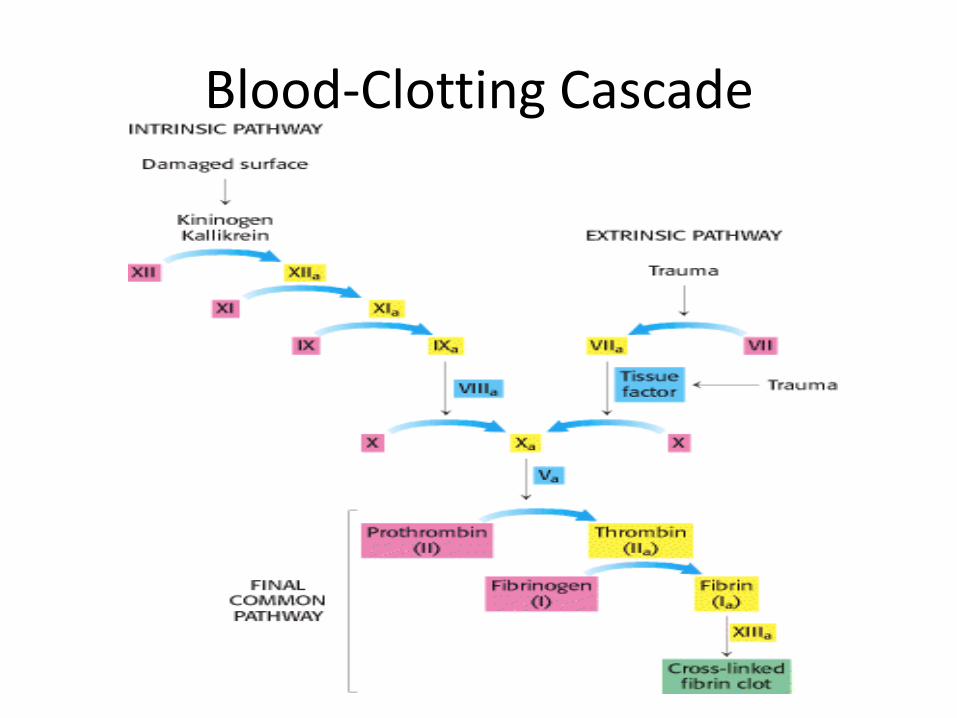
Blood-Clotting Cascade

Many Enzymes Require Cofactors for Activity

COENZYMES……Cofactors

Enzyme cofactors Metal Enzyme
Zn2+ Carbonic anhydrase
Zn2+ Carboxypeptidase
Mg2+ Hexokinase
Ni2+ Urease
Se Glutathione peroxidase
Mn2+ Superoxide dismutase
Cu2 Cytochrome oxidase

ISOENZYMES
Isozymes or isoenzymes, are enzymes that differ in amino acid sequence yet catalyze the same reaction. Usually, these enzymes display different kinetic parameters, such as K M, or different regulatory properties.
They are encoded by different genetic loci, which usually arise through gene duplication and divergence.

ISOENZYMES The existence of isozymes permits the fine-tuning of
metabolism to meet the particular needs of a given tissue or developmental stage. Consider the example of lactate dehydrogenase (LDH), an enzyme that functions in anaerobic glucose metabolism and glucose synthesis. Human beings have two isozymic polypeptide chains for this enzyme: the H isozyme highly expressed in heart and the M isozyme found in skeletal muscle. The amino acid sequences are 75% identical.

The functional enzyme is tetrameric, and many different combinations of the two subunits are possible. The H4 isozyme, found in the heart, has a higher affinity for substrates than does the M4 isozyme. The two isozymes also differ
in that high levels of pyruvate allosterically inhibit the H4 but not the M4 isozyme. The other combinations, such as H3M, have intermediate properties depending on the ratio of the two kinds of chains.

ISOENZYMES……… LDH


• END
• Chapter 7


















