
FBH PHD Afhandling 2014
159
Copenhagen 2014 1 FACULTY OF SCIENCE UNIVERSITY OF COPENHAGEN PhD thesis Frederik Boëtius Hertz ESBL-Producing Escherichia coli: Antibiotic Selection, Risk Factors and Population Structure. This thesis has been submitted to the PhD School of The Faculty of Science, University of Copenhagen’ Academic advisor: Anders Løbner-Olesen Academic advisor: Niels Frimodt-Møller Submitted: 29-09-2014
Transcript of FBH PHD Afhandling 2014

Copenhagen 2014
1
F A C U L T Y O F S C I E N C E UNIVERSITY OF COPENHAGEN
PhD thesis Frederik Boëtius Hertz
ESBL-Producing Escherichia coli: Antibiotic Selection, Risk
Factors and Population Structure.
This thesis has been submitted to the PhD School of The Faculty of Science, University of
Copenhagen’
Academic advisor: Anders Løbner-Olesen
Academic advisor: Niels Frimodt-Møller
Submitted: 29-09-2014

PhD Dissertation
Institutnavn: Biologisk Institut
Name of department: Functional genomics
Author: Frederik Boëtius Hertz
Title: ESBL-Producing Escherichia coli: Antibiotic Selection,
Risk Factors and Population Structure.
Academic advisor: Anders Løbner-Olesen
Academic advisor: Niels Frimodt-Møller
Submitted: 29th of September 2014

Copenhagen 2014
3
Table of Contents
Preface 5
Abbreviations 6
List of manuscripts 7
Abstract 8
Dansk resume 9
1. Introduction 10
1.1. Research objectives 11
1.2. Outline of dissertation 12
2. Background 13
2.1. Escherichia coli 13
2.1.1. Genomic Diversity of Escherichia coli 13
2.1.2. Extraintestinal Intestinal Pathogenic E.coli 14
2.2. Antibiotic Resistance in E.coli 14
2.2.1. Extended Spectrum Beta-Lactamases 15
2.2.2. Prevalence of ESBL-producing E.coli 16
2.3. Epidemiology of Resistance in E.coli 18
2.3.1. Clinical Consequence of Resistance 19
2.4. The Gastrointestinal Microflora 19
2.4.1. E.coli and Colonization 20
2.5. Reservoirs of ESBL-producing ExPEC 20
2.5.1. The Human Gut as Reservoir 21
2.5.2. Animals, Water and Food 21
2.6. Animal Experimental Models for Investigation of Colonization 22
2.7. Composition of ExPEC Susceptibility Populations 23
2.8. Risk factors for infection with ESBL-producing E.coli 25
2.8.1. Considerations on Study Design 25
2.8.2. Case-Control Studies 26
2.8.3. Double-case control study of epidemiological factors 27
2.8.4. Case-control study in low prevalence countries 28
3. Bacteriological Typing of E.coli 29
3.1. General Considerations on Typing Methods 29
3.2. Phylogrouping by PCR 29
3.3. Multi-Locus Sequence Typing 30
3.4. Multi-Locus Variable Number of Tandem Repeats Analysis 31
3.5. Serotyping 31
4. Material & Methods 33
4.1. General Protocols 33

PhD Dissertation
4.1.1. Research Ethical Approvals 33
4.1.2. CPR Registry 34
4.1.3. Statistics Denmark 34
4.2. Mouse Intestinal Colonization Model (Manuscript I) 35
4.2.1. Selection of E.coli Isolates and Preliminary Mice Experiments 35
4.3. Study Populations (Manuscript II and III) 35
4.3.1. Sample Size Determination 35
4.3.2. Strain Collection 36
4.4. Epidemiology of Collected Isolates 36
5. Results 37
5.1. Mouse intestinal colonization model (Manuscript I) 37
5.1.1. Effect of Antibiotic Selective Pressure on the Microflora 38
5.1.2. Effect of Antibiotic Selective Pressure on Prolonged Presence of 65-Ec-09 38
5.1.3. Molecular Tests of Bacteria Isolated from Mice Faeces 39
5.2. Case-control study (Manuscript II) 43
5.2.1. Urine Samples to DCM 43
5.2.2. Description of E.coli Populations 45
5.2.3. Statistical Analysis of Risk Factors and Population Data 47
5.3. Characterization of E.coli populations (Manuscript III) 49
5.3.1. Distribution of Serogroups, MLVA codes and Sequence Types 49
6. Discussion 56
6.1. Selection of CTX-M-producing E.coli in vivo 56
6.2. Investigation of Epidemiological Factors 57
6.3. Characterization of E.coli Populations 58
7. Conclusion and Perspectives 59
8. Reference 64
9. Appendix 86

Copenhagen 2014
5
Preface
The studies included in this PhD dissertation were carried out at Department of Microbiology and
Infection Control at Statens Serum Institut (SSI) and at Department of Clinical Microbiology, Hvidovre
University Hospital (DCM). The research was carried out as part of PAR, an EU FP7-Health-2009-Single
Stage Project, with support from The Danish Centre for Antibiotic Research and Development
(DanCARD).
Work has been interrupted by two periods of paternity leave, each of three and half months.
At SSI and DCM I have worked with several skilled and generous people. I am very thankful to the many
people who have kindly supported me and contributed greatly to this PhD thesis.
First I would like to extend my gratitude to my supervisors Anders Løbner-Olesen and Niels Frimodt-
Møller for their exceptional guidance and patience as well as the good-humoured atmosphere at all times.
Your support and confidence have made this work possible.
I would like to thank the medical staff and laboratory technicians at SSI and DCM for the technical
assistance and tolerance throughout the three years of research. I must also thank Lars Villiam Pallesen
and Christian Østergaard Andersen for use of department facilities and support.
Pia Jeannette Littauer, Jenny Dahl Knudsen and especially Kristian Schønning have all engaged in
excellent scientific discussions and provided appreciated guidance and aid.
Finally, I would like to thank many of my closest current and former colleagues Anne Sandberg-Schaal,
Lotte Jakobsen, Alexandra Bugay Medina, Mette Pinholt, Steen Christian Rasmussen, Karen Leth
Nielsen and Rikke Fleron Leihof for their friendship and help during this PhD. I would especially like
thank Jesper Boye Nielsen for, not only his friendship, but also his immense efforts and contribution to
my work in the laboratory and scientific deliberations.
My love and warmest gratitude go to my wife Christin for her support and encouragement.
Frederik Boëtius Hertz, August 2014

PhD Dissertation
Abbreviations
a-MLVA Abbreviated MLVA
COI Community-onset infections
CPD-R Cefpodoxime resistance
ESBL Extended-Spectrum-Beta-lactamase
ESCMID European Society for Microbiology and Infectious Diseases
ExPEC Extraintestinal pathogenic Escherichia coli
GP General Practitioner
HCAI Healthcare-associated infections
HGT Horizontal gene transfer
MALDI-TOF Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry
MDR Multidrug resistant
MLST Multi locus sequence typing
MLVA Multi locus variable number of tandem repeat analysis
MNEC Meningitis-associated E.coli
PCR Polymerase chain reaction
PFGE Pulsed field gel-electrophoresis
SSI Statens Serum Institut
SLV Single locus variant
ST Sequence type
UTI Urinary Tract infection
UPEC Uropathogenic E.coli
VF Virulence factor
VNTR Variable number of tandem repeats

Copenhagen 2014
7
List of Manuscripts
Paper I: Hertz F.B., Løbner-Olesen A. and Frimodt-Møller N.
“Antibiotic selection of E.coli ST131 in a mouse intestinal colonization
model”. Antimicrobial Agents and Chemotherapy., 2014. Vol. 58 (10)
Manuscript II: Hertz F.B., Littauer P., Schønning K., Knudsen J.D., Løbner-Olesen A.
and Frimodt-Møller N.
“Epidemiological factors associated with ESBL- or non ESBL-
producing E.coli causing UTI in general practices.”
Manuscript III: Hertz F.B., Nielsen J.B., Littauer P., Schønning K., Knudsen J.D.,
Løbner-Olesen A., Frimodt-Møller N.
“Population structure of Drug-Susceptible, -Resistant and ESBL-
producing Escherichia coli Populations from Community-Acquired
Urinary Tract Infections as Characterized by Abbreviated MLVA,
MLST and Serogrouping in Denmark”

PhD Dissertation
Abstract
Urinary tract infection (UTI) is one the most common bacterial infections and is regularly treated in
primary health care. The most common cause of UTI is extraintestinal pathogenic Escherichia coli
(ExPEC) already present in the intestinal microflora, often as the dominating strain. Resistance in E.coli
is increasing and especially isolates producing Extended-Spectrum Beta-Lactamases (ESBL) have been
reported worldwide. Treatment of UTI is usually initiated by the general practitioners and a significant
proportion of clinical isolates are now resistant to first line antibiotics. The global dissemination of
resistant E.coli has in particular been driven by the spread of a few specific E.coli-lineages and it seems
that there is a difference between the sequence types found among resistant E.coli, ESBL-producing
E.coli and antibiotic susceptible E.coli. The overall objectives of this thesis were to investigate (i)
antibiotics involved in selection of ESBL-producing E.coli, in an experimental mouse model in vivo, (ii)
risk factors for UTI with ESBL-producing E.coli and (iii) to describe the phylogenetic composition of
E.coli populations with different resistance patterns.
We found that different antibiotics can select for the ESBL-producing E.coli, even anti-Gram-
positive antibiotics as dicloxacillin and clindamycin showed selective abilities. While dicloxacillin
has no effect on anaerobic Gram-negatives, this is the case for clindamycin. The selective abilities
of other beta-lactam antibiotics varied, with selection identified by cefotaxime, cefuroxime and
penicillin. The triple-case control study showed that exposure to antibiotics is not a good predictor
for risk of UTI with ESBL-producing E.coli. There were few differences between the case groups
when compared to the uninfected group. However, when case groups where compared to each
other, healthcare association and hospital admission proved to be independent risk factors for UTI
with ESBL-producing E.coli. When typing uropathogenic E.coli, we less frequently saw ST131
among non-ESBL than among ESBL-producing E.coli. We found that ESBL-producing E.coli,
resistant E.coli and susceptible E.coli in turn were dominated by different MLVA codes and
sequence types. Overall the susceptible E.coli was a much more diverse group, whereas resistant
and ESBL-producing E.coli were found in larger clusters, indicating that the success of resistant
lineages like O25b-ST131 is mainly due to positive selection of previously specialized UPEC with
newly gained resistance.

Copenhagen 2014
9
Abstract – Danish
Urinvejsinfektion (UVI) er en af de hyppigste bakterielle infektioner og de behandles typisk i almen
praksis. Den mest almindelige årsag til UVI er ekstra intestinale patogene Escherichia coli (ExPEC), der
forud for infektion ofte er til stede i den intestinale microflora. Resistens blandt E.coli isolater er stigende
og især isolater der producer beta-lactamaser med udvidet spektrum (ESBL) er beskrevet over hele
verden. Behandling af UVI håndteres og påbegyndes ofte af den praktiserende læge og en betydelig del af
de kliniske isolater nu er resistente over for almindeligt benyttede antibiotika. Denne globale spredning af
resistente E.coli har i høj grad været drevet af spredning af få E.coli-kloner og der er tilsyneladende en
forskel på de sekvens typer der ses blandt resistente E.coli, ESBL-producerende E.coli og fuldt følsomme
E.coli. Formålet med denne ph.d. afhandling var, at undersøge (i) hvilke almindeligt benyttede antibiotika
der er involveret i selektion af ESBL-producerende E.coli in vivo, (ii) risiko faktorer for UVI med ESBL-
producerende E.coli og endelig (iii) at beskrive den fylogenetiske sammensætning af E.coli populationer
med forskellige resistens mønstre.
Vi fandt, at forskellige antibiotika kan selektere for ESBL-producerende E.coli, selv anti-Gram-
positive antibiotika som dicloxacillin og clindamycin selekterede. Clindamycin har stor effekt på
anaerobe Gram-negative, mens dicloxacillin ingen effekt har. De selektive egenskaber af andre
beta-lactam antibiotika varierede, med positiv selektion observeret fra cefotaxim, cefuroxim og
penicillin. Case kontrol studiet viste, at antibiotikaforbrug ikke er en god variabel til at
risikostratificere patienter for infektion med ESBL-producerende E.coli. Der var kun få forskelle
mellem case grupperne når disse blev sammenlignet med den ikke inficerede gruppe. Når case
grupper blev sammenlignet med hinanden så vi dog, at kontakt med sundhedsvæsenet og hospitals
indlæggelser var risiko faktorer for UVI med ESBL-producerende E.coli. Ved den bakterielle
typning af de uropatogene E.coli, så vi at ST131 var langt mindre hyppige blandt ikke-ESBL end
blandt de ESBL-producerende E.coli. Vi fandt at ESBL-producerende E.coli, resistente E.coli og
følsomme E.coli var domineret af forskellige MLVA koder og sekvens typer. Alt i alt var de
følsomme E.coli en langt mere diverse gruppe, mens de resistente E.coli og især de ESBL-
producerende E.coli blev identificeret i større grupper, hvilket indikerer at den globale succes for
resistente linjer, som O25b-ST131, i høj grad skyldes det positive selektions tryk, der selekterer
uropatogene E.coli med erhvervet resistens.

PhD Dissertation
1. Introduction
Urinary tract infection (UTI) is one the most common bacterial infections and it is regularly treated in
primary health care by general practitioners (GPs) (1–3). The highest incidence of UTI is found in young,
healthy women, but bacterial infection of the urinary tract is seen in all ages and in both genders (3). The
most common cause of UTI are extraintestinal pathogenic Escherichia coli (ExPEC), causing more than
70% of community-acquired UTI (1,4,5). Resistance in E.coli is increasing and especially isolates
producing Extended-Spectrum Beta-Lactamases (ESBL) have, in the last decades, been reported
worldwide(3,6). Treatment of UTI is usually initiated by the GP and a significant proportion of clinical
isolates are now resistant to first line antibiotics (3,6). This poses a great challenge for the empirical
treatment of infections, caused by E.coli (3,6).
The global dissemination of resistant E.coli have in particular been driven by the spread of a few specific
E.coli-lineages, best characterized by Multi-Locus Sequence Typing (MLST) (7–10). It seems that there
is a difference between the sequence types (STs) found among resistant E.coli, ESBL-producing E.coli
and antibiotic susceptible E.coli, illustrated by recurrent findings of explicit STs in each population (7–9).
UTI is most often caused by an E.coli strain already present in the intestinal microflora, often as the
dominating strain, and as such the human intestinal tract pose a reservoir for ExPEC (11,12).
Additionally, up to 80% of recurrent UTI are caused by the index strain and 25-40% of women
experience a new case of UTI within 6 months (13,14).
Overall risk factors for UTI have been shown to be female gender, previous UTI, sexual activity and
recent exposure to antibiotics, with exposure to antibiotics also posing a major risk for UTI with resistant
bacteria (3,5,11). With the great increase in ESBL-producing E.coli, found to be the reason for higher
morbidity, prolonged hospitalization and increased health care costs, there has been an interest in the
epidemiological factors associated with infections caused by ESBL E.coli. Thus, risk factors have been
widely investigated and reports have, among other, identified age > 65 years, male sex, previous UTI, use
of antibiotics and hospitalization as common risk factors (15–18). Moreover diet, lifestyle and traveling
have been proposed as sources for colonization and infection (17–19).

Copenhagen 2014
11
1.1. Research Objectives
The overall objectives of the present dissertation were to investigate antibiotics involved in
selection of ESBL-producing E.coli, risk factors for UTI with ESBL-producing E.coli through focus
on usage of antibiotics and finally to describe E.coli populations to identify differences in the
phylogenetic compositions among resistant and susceptible E.coli.
The main objectives were therefore:
Objective I:
To evaluate the ability of nine common antibiotics, to select for a CTX-M-15-producing E.coli
isolate belonging to sequence type (ST) 131 in vivo. This was done in a mouse intestinal
colonisation model developed and tested as part of this research.
Objective II:
To investigate epidemiological factors associated with acquiring UTI with ESBL-producing E.coli,
in general practices, investigated as a case-control study involving three case groups and a single
control group.
Objective III:
To investigate if the population structure of uropathogenic E.coli populations is predominantly
composed of related isolates, regardless of resistance profile. This was done by typing of three
susceptibility groups by use of a-MLVA, MLST, serogrouping and ESBL-genotyping.

PhD Dissertation
1.2. Outline of dissertation
This thesis consists of six chapters. First, a literature review provides an overview of ExPEC,
antibiotic resistance in E.coli, possible reservoirs, previously used experimental mouse models,
known population structure of E.coli populations and risk factors for UTI with ESBL-producing
E.coli. This is followed by a brief review of typing methods used in this thesis. Secondly, a brief
method section describes methods and study designs not described in manuscript I-III, followed by
a presentation of main findings. Finally, the thesis concludes with a short discussion of main results
and an overall conclusion with future perspectives. References are presented before appendix´s are
shown and the thesis concludes with an inclusion of Manuscript I-III.

Copenhagen 2014
13
2. Background
2.1. Escherichia coli
Escherichia coli (E.coli) is a member of the family Enterobacteriaceae and is a Gram-negative,
facultative anaerobic bacterium (20). It is a highly versatile bacterial species comprised of both
harmless commensal strains and different pathogenic variants with the ability to cause either
intestinal or extraintestinal diseases (21,22). Consequently, E. coli strains are broadly classified into
three major groups of commensal E.coli, intestinal pathogenic E.coli (IPEC) and extraintestinal
pathogenic E.coli (ExPEC) (21–23). As a non-pathogenic inhabitant of the intestine of many
mammals, including humans, E.coli exists as part of the indigenous flora, often contributing to the
vital tasks performed by the intestinal microflora (20,24). Traditionally, commensal E.coli have
been described as colonizers that rarely cause infection and categorized as belonging to phylogroup
A and B1, while ExPEC isolates are mostly derived from phylogroup B2 and D (20,22,25). All four
phylogroups can, however, cause intestinal and extraintestinal infections and phylogroup B2 and D
have been found as regular colonizing strains in healthy individuals (11,20,26).
The pathogenic E.coli, IPEC and ExPEC, can each be further subcategorized into specific
pathotypes. This classification is based on clinical manifestations of disease and the pathogenic
traits such, as presence of virulence factors (VFs) (20). The most prevalent ExPEC pathotypes are
the uropathogenic E.coli (UPEC) and meningitis-associated E.coli (MNEC) (27). Often intestinal
non-pathogenic E.coli and IPEC can be distinguished by genome content and phenotypic traits, but
the discrimination between commensal E.coli and extraintestinal pathogens is not easy (21). ExPEC
strain are habitually found as part of the commensal flora of healthy individuals without causing
enteric disease (20,23). While IPEC cause diseases of the intestinal tract, ExPEC can cause a range
of diseases in almost any anatomical niche such as UTI, bacteraemia, meningitis and intra-
abdominal infections (27–29).
2.1.1. Genomic Diversity of Escherichia coli
In E.coli an extensive horizontal gene transfer (HGT) by plasmids occurs, contributing to a
noticeable genome plasticity in the species (20,21). As a result and in combination with genomic
recombination, E.coli has an amazing ability to colonize and survive in many different locations
(10,20). The genome is composed of the conserved core of genes, constituting the genetic
information for most essential cellular processes, and a flexible gene pool often consisting of

PhD Dissertation
genetic information providing the abilities to adapt to new environmental conditions (10). The
acquisition of new genomic material therefore contributes to the rapid evolution of E. coli lineages
(21). Thus, E.coli is a versatile species with a remarkable adaptive potential, alternating between
habitats and adapting rapidly to new and complex ecosystems (30,31).
2.1.2. Extra Intestinal Pathogenic Escherichia coli
Extraintestinal infections in humans have a high incidence and ExPEC is the most common Gram-
negative extraintestinal pathogen. The most frequent infection is UTI, but E.coli is also the leading
cause of bloodstream infections (9,25,32,33).
The term ExPEC was introduced by Johnson et al. in 2000 based on reports of UPEC and MNEC
isolates causing a range of extraintestinal infections (23). Several presumed virulence
genes were linked to the pathogenicity of ExPEC, enabling them to invade almost any
extraintestinal tissue (22,23). Many of these VFs are present on the chromosome, but VFs are seen
extensively on mobile elements, creating great diversity within the categories of ExPEC pathotypes
(20,24,30). Among ExPEC there is an increasing recognition of isolates resistant to antibiotics, the
most important being isolates producing plasmid-mediated β-lactamases (22).
2.2. Antibiotic resistance in E.coli
Generally, there are five main methods by which Gram-negative organism develop
resistance(34,35):
First, bacteria can carry genes coding for enzymes, such as beta-lactamases, hydrolysing and
inactivating beta-lactam antibiotics. Second, mutations can occur in the genes for binding sites for
antibiotics changing the specific target or its function. Third, alterations of the membrane porins
result in reduced permeability. Fourth, bacteria can express efflux pumps to actively transport
antibiotics out of the cell and finally, fifth, alternate metabolic pathways can by-pass paths inhibited
by antibiotics (34,35). Resistance in Gram-negative bacteria can be intrinsic, arise or be acquired
and is often composed of a combination of resistance mechanism like beta-lactamases, porin
deletions and efflux pumps (36–40). The predominant mechanism of resistance is, however, the
hydrolysis of the antibiotic by beta-lactamases (41). The ability to produce β-lactamases, including
ESBL, is frequently acquired through large plasmids holding many different genes coding
resistance against other antibiotic classes, contributing to MDR (41,42). This also means that many
different antibiotics may facilitate selection of, and colonization by, these pathogens and that the

Copenhagen 2014
15
same types of ESBLs may be recognized in clonally unrelated isolates or isolates of the same clonal
origin may encode different types of ESBLs (42). E.coli is one of the organisms most frequently
found harbouring ESBL-genes and MDR in ESBL-producing Enterobacteriaceae is rapidly
becoming a threat to the medical community (9,25,32,33,42–47).
2.2.1. Extended Spectrum Beta-Lactamases
Beta-lactams are the most widely used class of antibiotics – all characterized by having a β-lactam ring as
the chemical base and include penicillins, cephalosporins, carbapenems, monobactams and
cephamycins(48). Hydrolysis by enzymes is the most common mechanism for resistance towards this
class of antibiotics (22,49,50). Beta-lactamases belong to a very large, diverse family of enzymes capable
of hydrolysing β-lactam antibiotics by cutting open the β-lactam ring and the hydrolysing ability of β-
lactamases can be of narrow or broad spectres. Several hundred β-lactamases have been described
(http://www.lahey.org/Studies/) each with a defined spectrum (examples: penicillinases and
cephalosporinases). These enzymes are divided into molecular classes based on their amino acid
similarities (Ambler scheme A to D = Classes) or into groups by functional similarities (Bush-Jacoby-
Medeiros scheme = Groups 1-4). Ambler class A, C and D are serine- β-lactamases and class B are
metallo- β-lactamases (51). Finally, several families of β-lactamase exist, with the most common plasmid-
mediated β-lactamases being TEM and SHV, accounting for >60% of E.coli resistance to ampicillin
(41,51).
A major source for resistance in E.coli are plasmid-borne Extended-Spectrum β-Lactamases
(ESBL), which are broadly classified as enzymes capable of hydrolysing most β-lactams such as
penicillins, extended-spectrum cephalosporins and monobactams, but not cephamycins and
carbapenems. In addition, ESBLs are inhibited by β-lactamase inhibitors such as clavulanic acid,
sulbactam, and tazobactam, and are carried on large plasmids often containing various other genes,
causing resistance to other classes of antibiotics (47,50,52,53). Several ESBL families have been
described, but the majority of ESBLs belong to the four large families of SHV, TEM, CTX-M and
OXA (53,54).
The first ESBL was a SHV type detected in K.pneumoniae in 1983 in Germany and around the
same time the first TEM ESBLs were found in France and England (51). For unknown reasons,
K.pneumoniae was the bacterial species most likely to express ESBLs in the 1980s and 1990s.
These ESBLs were most frequently derivatives of common penicillinases like TEM-1 and SHV-1,
generated by point mutations creating an extended-spectrum of hydrolysis (6,42). In 1989 an ESBL

PhD Dissertation
CTX-M enzyme was described in Germany (named for cefotaxime resistance and place of
discovery, Munich) (51). CTX-M enzymes are believed to have originated from Kluyvera spp. and
are divided into five groups (CTX-M group 1, 2, 8, 9 and 25) again with several subgroups. With
the emergence of different CTX-M types the prevalence of ESBLs changed dramatically during the
early 2000s. Rapidly CTX-M types surpassed SHV and TEM as the dominating ESBL, most often
present in E.coli (22). Now, CTX-M ESBLs display a global distribution causing infections in every
part of the world (10,42,45,49).
2.2.2. Prevalence of ESBL-producing E.coli
With the emergence of CTX-M ESBLs, community-onset ESBL infections have become an important
public health issue, primarily caused by E. coli producing CTX-M type ESBLs (10,22,42). The rapid
worldwide dissemination of this particular ESBL type has been known as the “CTX-M pandemic” and the
dominance of CTX-M types ESBL are, largely, caused by dissemination of E.coli lineages, often
expressing co-resistance to other classes of antibiotics (10,41,45). It has become evident, that once a
CTX-M type enters an area, it becomes prevalent, replacing TEM and SHV as the dominating ESBL (49).
In 2007, a study based on TEST global surveillance database reported that the incidence of ESBL-
producing E.coli was highest among isolates collected in Latin America (13.5%) and Asia (12%)
followed by Europe (7.5%) and North America (2.2%) (42,55). As seen, there is a noticeable differences
in ESBL prevalence, a variation also seen in dominating CTX-M subtypes between European countries
and different parts of the world, as depicted by Hawkey and Jones in 2009 (Figure 1) (42,55). Examples
include CTX-M-1 in Italy and the Netherlands, CTX-M-2 in Argentina and Israel, CTX-M-3 enzymes in
Poland, CTX-M-9 in Spain, CTX-M-14 in China and CTX-M-15 in UK and Denmark. Nevertheless,
CTX-M-14 and -15 producing E.coli are distributed around the world and CTX-M-15 is the most
prevalent type (10,22,42,45,49,56,57).
Looking at the ESBL-prevalence in Denmark there has been a slow, but steady, increase in number of
infections caused by ESBL-producing E.coli. In clinical isolates from 1997 there were no ESBL-
producing E.coli found (58). In 2003, 0.8% of E.coli isolates were ESBL-producing and cefuroxime
resistance was found in <5% of E.coli isolates in the years 2003-2006 (59). However, a study on E.coli
isolates from 2007, reported the UPEC ESBL-prevalence to be 1.5% from general practices and 2.3% in
hospital urine, with 60% of ESBL-producing E.coli producing CTX-M-15 (60). Resistance to extended-
spectrum cephalosporins in UPEC from primary health-care, used as a marker for ESBL-production, was
found to be 4% in 2012 (61).

Copenhagen 2014
17
Figure 1. Worldwide distribution of different classes of CTX-M β-lactamases (first identified in 1989). Davies J , and Davies
D Microbiol. Mol. Biol. Rev. 2010;74:417-433

PhD Dissertation
2.3. Epidemiology of Resistance in E.coli
An important feature complicating treatment of infections caused by E.coli is the increase in
resistance to first-line antibiotics (22,61,62). Until the late 1990s ExPEC were relatively susceptible
to first-line drugs (22). Currently, resistance in Gram-negative bacteria constitutes one of the
biggest challenges to public health and the changes in antimicrobial susceptibility have the potential
to impact efficacy of antibiotics (6,45,46,63). When resistant bacteria spread to the community,
resistance creates comprehensive infection control issues, increasing morbidity for non-hospitalized
patients of all ages (63).
The estimated number of cases of uncomplicated cystitis per year, caused by E. coli alone, is 130–
175 million globally and 2-300.000 in Denmark alone (N. Frimodt-Møller, personal
communication) (64). Consequently, infections caused by E.coli, susceptible and resistant,
collectively result in considerable morbidity as well as direct and indirect financial costs seen as
increased health-care expenses, antibiotic treatment and loss of productivity (14,22,25,64).
Furthermore, UTI patients experience morbidity and impaired quality of life with an estimated 20-
40% of women having at least one UTI during their lifetime (22,64,65). It is difficult to determine
the precise incidence of UTI, but by using self-reported medical history the annual incidence in
USA was 13% among women and 3% among men (3).
Resistance in E.coli, besides β-lactam resistance, includes sulphonamides, trimethoprim and
ciprofloxacin (22). In 2008, UPEC isolates from five countries, were commonly resistant to
ampicillin (28%), sulfonamides (25%), trimethoprim (17%) and nalidixic acid (10%), with an
significant increase in resistance to nalidixic acid and trimethoprim from 2000 to 2009 (4). A total
of 60%, only, of the UPEC isolates were found to be fully susceptible (4). The antibiotic resistance
continued to increase throughout Europe, with 41% being fully susceptible in 2012, only. Especially
the current increase in resistance to extended-spectrum cephalosporins (mean = 12%) and
aminoglycosides (mean = 10%) in combination with increased resistance to at least three antibiotic
classes, are worrisome (41). The increased resistance is likewise worrying in Denmark. In 2012, the
resistance in E.coli isolated from urine (primary health care) were 40% for ampicillin with 33% for
sulphonamide and 10% were resistant to ciprofloxacin and 6% to mecillinam (61).
The continual increase in resistant E.coli has added to the enormous economic and human costs of
infections with 400.000 infections caused by MDR bacteria in Europe in 2007 (66). The economic
costs associated with these infections, counted as extra hospital costs and productivity losses
exceeds €1.5 billion in Europe and $20 billion per year in the United States (66).

Copenhagen 2014
19
2.3.1. Clinical Consequences of Resistance
There is an on-going discussion on the methods by which outcome of infections should be
investigated and uncertainty of true influence by resistant pathogens do exists (67). However,
predictors of mortality in patients with infections due to MDR Gram-negative bacteria have
previously been defined as infection severity, underlying diseases, inappropriate empiric treatment,
age and multidrug resistance (67,68). As MDR strains, including ESBL-producing strains, often are
resistant to most first line antibiotics, patients infected with these are more likely to receive
inappropriate empirical therapy why morbidity and mortality rate is generally higher (67,69–72). As
such patients with ESBL infections are more likely to suffer prolonged hospital stay and infections
are associated with higher use of broad-spectrum antibiotics (18,72–79). The increased mortality is,
however, significantly reduced if correct definitive therapy is given according to susceptibility
patterns and precise nonmedical interventions are performed (68). This, of course, makes
identification of patients at risk and carriers of resistant strains of great importance.
2.4. The Gastrointestinal Microflora
The human intestine is populated by thousands of bacterial species with the vast majority (99%)
being strict anaerobes and less than 1% being facultative aerobic and aero-tolerant (20,80). The
anaerobic Gram-negative Bacteriodetes and Gram-positive Firmicutes constitute 80–90% of the gut
bacteria (81).
The gastrointestinal microflora in the human intestine performs well recognized functions like
digestion of otherwise indigestible carbohydrates and the prevention of colonization of pathogenic
strains referred to as colonization resistance (20). The indigenous intestinal flora, therefore, acts as a
barrier against incoming pathogens and overgrowth of opportunistic microorganisms already
present in the gut. However, alterations in the microbiota can allow for colonization, with possible
subsequent infection, and antibiotic treatment is known to disturb the ecological balance of the
indigenous microflora (11,43,82). In addition, antibiotic treatment can select for resistant bacteria,
which could then disseminate in the intestine, increasing the possibility of infections (11). The level
of disturbances, caused by antibiotics, depends on route of administration, route of elimination and
spectrum of the agent (83). Resistance genes can be introduced into the gut via a travelling, non-
colonizing strain. The presence of an antibiotic, or recent exposure, will allow the resistant
microorganisms to establish themselves and proliferate (80). This overgrowth of resistant bacteria
encourages conjugation of plasmids between same and different species, increasing the resistance

PhD Dissertation
gene pool (37,43,83–88). Overgrowth and establishment of resistant strains is not seen, in nearly the
same degree, in individuals not under the influence of antibiotic (84,86,88,89). There is therefore a
likely association between last use of antibiotics and presence of resistance, but resistant E.coli have
been found in healthy individuals with no recent antibiotic exposure (84,86,88–90). Once present,
resistant pathogens may spread to other parts of the body or other individuals and the number of
colonized individuals exceeds the number of patients with identified infection (80). In clinical
settings, the gastrointestinal tracts can, as such, act as a source of infection or transmission and
previous exposure to antibiotic is a known risk factor for UTI with resistant bacteria
(37,38,43,82,83,86,87,91–93). In addition faecal carriage of ESBL-producing E.coli have been
detected in patients with ESBL infections, with risk of UTI increasing with larger quantity of the
ESBL-producing E.coli and abundance of such E.coli increasing with antibiotic exposure (42,93).
2.4.1. E.coli and Colonization
Virulence factors conventionally determining ExPEC are also found in commensal E.coli indicating
that VFs causing extraintestinal disease are also important for the intestinal colonization (10,22).
Usually the faecal flora of healthy humans is inhabited by one to five E.coli clones, with E.coli as
the dominating facultative anaerobic species and one clone habitually being dominant (11,20,89). It
has been found that E.coli belonging to classical ExPEC phylogroups B2 and D are often among the
dominating strains. Thus, there is a link between the presence of certain virulence genes,
colonization and pathogenicity with commensal E.coli often resembling ExPEC (11,20). Infections
due to ExPEC isolates, including UTI, are most often caused by E.coli already present in the
patient’s own intestinal flora (11,26,62). The human gut is now considered to be the primary
reservoir for uropathogenic E.coli (11,22,94).
2.5. Reservoirs of ESBL-producing ExPEC
Larger reservoirs have only been detected at local levels, like high rates of carriage in nursing-home
residents, with limited knowledge of true external reservoirs (52,62). Community outbreaks caused by
pathogenic E.coli have been described for ExPEC and a variety of contaminated foods have previously
been linked to outbreaks of IPEC (22,95,96). During a recent outbreak in Germany, of Shiga toxin
producing IPEC, also expressing ESBL, sprouts were identified as the likely primary source with
secondary person-to-person transmission and long-term shedding after illness (97). Transmission of E.coli

Copenhagen 2014
21
through sexual activity have been reported and infecting UPEC have been found in the faecal flora of
family members (14,22,98).
2.5.1. The Human Gut as Reservoir
Among 86 Danish army recruits, 3.5% had ESBL-producing E.coli in their faecal flora in 2011 (99). In
Sweden 2.4% of healthy individuals were colonized with ESBL-producing Enterobacteriaceae and
Jørgensen et al. found the overall ESBL carrier rate among patients in Norway, submitting faecal samples
to a hospital laboratory, to be 15.3% (100). Similar low to modest carrier rates have been seen in Japan
and Switzerland (6%), Tunisia (7%), UK (11.3%) and Saudi Arabia (13.1%), but with extremely high
carrier rates in Cairo (63.3%) and rural areas of Thailand (65.7%) and with varying findings in China (7-
50%) (56,101–104). In 2011, in the Paris area, a 10-fold increase in ESBL-carriage in healthy individuals
was detected over a 5-year period (from 0.6% to 6%) (104).
History of international travelling has been found to propose a risk for colonization of ESBL-producing
E.coli (52). Studies in the Scandinavian countries have identified considerably higher rates of carriage of
ESBL-producing Enterobacteriaceae among patients with a history of recent travelling to Africa north of
the equator, Asia and in particular the Indian subcontinent (100,101,105,106). This applied to both
hospitalized and non-hospitalized patients (100,101,105,106). These Scandinavian studies corresponded
well with a report from Canada showing that travellers were five times more likely to be carriers of ESBL
than non-travellers (107). Östholm-Balkhed et al. found that travellers >65 years had a higher risk of
ESBL acquisition than travellers aged 18-35 years and Tham et al. identified 24% of travellers as long
time-carriers (3-8 months) (101,108).
Comprehensive intestinal colonization of resistant E.coli has been seen in some nursing homes suggesting
long-term care facilities as another reservoirs for resistant bacteria (52). Rooney et al. found that 40% of
nursing home residents in Northern Ireland were colonized with ESBL-producing E.coli (109). Here
carriers of ESBL had higher levels of exposure to antibiotics, high rates of hospital admission and
frequent use of urinary catheter (109).
2.5.2. Animals, Water and Food
Finally, food, water and animal reservoirs have been suggested, with a genuine zoonotic origin of UPEC
strains previously suspected (65,110). ESBL-producing E.coli have been detected in production animals
in several studies, with the presence of known human ExPEC lineages (52,98). Pathogenic E.coli from
animals (house-hold pets and production animals) and humans do, to some extent, share a common

PhD Dissertation
genetic background based on MLST, yet with some species specific VFs (62,94,111,112). E.coli isolates
from meat have, however, been shown able to cause disease in a mouse model of ascending UTI (113).
In two Danish studies, ESBL-producing E.coli were found with increased frequency from pigs-farms with
a high consumption of extended-spectrum cephalosporin (98). Similarities between E.coli isolates and
ESBL types have additionally indicated transfer among animals, meats and humans and a voluntary ban
on use of cephalosporin in pig production, furthermore led to a decrease in ESBL-producing E.coli among
slaughter pigs in Denmark (62,98). The Netherlands have a higher level of antibiotic use in the poultry
industry, than any other European country, and the prevalence of ESBL-producing E.coli in the
gastrointestinal tract of healthy food-producing poultry were 15% in 2008 (114). Of retail chicken meat
94% of samples contained ESBL-producing E.coli isolates, of which 39% were ESBL genotypes also
found among humans (114). In Barcelona, pathogenic E.coli lineages, often associated with resistance,
were detected in sewage and river water, with high levels of clonal similarities to human isolates (110).
Lastly, ESBL-producing E. coli were detected in healthy pigs, cattle, and poultry as well as in wildlife in
China. The latter has been suspected to be due to spreading of manure containing ESBL-producing E. coli
(115).
2.6. Experimental Animal Models for Investigation of Colonization
The compositions of the intestinal microflora of laboratory mice have been examined by Krych et al. to
evaluate the similarities to the flora of the human gut (116). There were some quantitative differences, but
mouse and human faeces, to a large extent, had similar representatives of phyla and a substantial segment
of common genera. Hence, lab mice and human share the same basic bacterial species in the gut (116).
Animal models have been used extensively to study the pharmacokinetic and -dynamic parameters of
antibiotics as well as acute and chronic infections caused by bacterial pathogens with different resistance
patterns (117,118). Furthermore, since the intestinal flora of mice and men are comparable, mice are often
the mammal used to investigate the relationship between intestinal microbiome, health and disease (116).
Colonization studies of the mouse gut by bacteria have been used in a range of mouse models, with many
similarities between methods (119,120). In general, both at SSI and with foreign research groups,
colonization has been described in mice given antibiotics prior to inoculation with bacteria of interest and
conjugation between two colonizing strains have likewise been tested (37,119,121,122).
The effect of subcutaneous treatment on the indigenous intestinal microflora as well as the effect on
colonisation by a KPC-producing Klebsiella strain (KPC-Kp strain) was tested in a mouse study in 2011
by Perez et al (123). Their findings suggested that antibiotics with effect on the anaerobic intestinal flora

Copenhagen 2014
23
promoted the establishment of the KPC-Kp strain – provided the antibiotic had no effect on the strain of
interest. Hence, antibiotics with limited effect on the anaerobic flora might be less likely to promote the
colonisation of MDR Gram-negative bacteria (123). Perez et al also found that antimicrobial impact on
the total anaerobic population is proportional to the measured impact on the Bacteroides population (123).
Colonization was successfully investigated by sampling and measurement of stool, since E.coli is present
in both mucus content and intestinal content (123,124). The hypothesis gained from previous mice studies
has, however, never been confirmed or tested on more common resistant Gram-negative strains. It has
previously been shown, that a human-simulated dosing of antibiotics gives the most constant levels of
drug in mice – who have a higher elimination rate (125). Yet, a single daily subcutaneous dose can
produce similar levels of drugs in mice faeces, to those seen in humans (123). This dosing frequency of
antibiotic does, nonetheless, not precisely mimic the exposures seen in patients (123).
Mouse intestinal colonization of Gram-negative bacteria has therefore successfully been determined in
several studies, many using elimination of resident facultative bacteria prior to inoculation
(119,121,123,124,126).
2.7. Composition of ExPEC populations
With the introduction of MLST, many E.coli isolates, causing outbreaks and previously
characterized by serotypes, were re-characterized by sequence type (ST) (7). Consequently, reports
of the global spread of ExPEC belonging to a few STs followed, with reports of worldwide
occurrence of ST10, ST12, ST73, ST95, ST127, ST131 and ST393 (7). The spread of E.coli-
lineages associated with certain resistant patterns and multidrug-resistant STs were also,
retrospectively, determined.
Interestingly, the initial descriptions of ESBL-producing E.coli gave indications of a non-clonal
distribution of isolates with great diversity of MLST types and ESBL types (52,127,128). However,
reports of rapid worldwide dissemination of the CTX-M β-lactamases, and especially CTX-M-15,
were followed by descriptions of closely related groups of E.coli spreading throughout the world
and this global spread was not confined to ESBL-producing E.coli. ST69 has since 1999 appeared
worldwide as a resistant E.coli related primarily to trimethoprim-sulphamethoxazole resistance, but
not regularly as ESBL-producing strains (7,129). Conversely, ST393 emerged as a MDR ExPEC in
1986-87 in London and has since then spread to many parts of world, gaining resistance and has
now been associated with CTX-M-type ESBLs, without losing virulence (7,130). Likewise, ST131
was identified in the mid-2000s with an unknown origin and has been associated closely with

PhD Dissertation
ESBL-production and quinolone resistance (7,131,132). Today a substantial proportion of MDR
ExPEC, worldwide, belong to ST131 producing CTX-M-type ESBLs and is described as a
pandemic ST-lineage with very low incidence amongst susceptible isolates (7,52,131,132). Other
very common ExPEC lineages like ST73 and ST95 were discovered among isolates from the 1917
and 1941 and has continued as common causes of UTI, but rarely as carriers of ESBLs and rarely
multidrug resistant (52,133–135). Consequently, certain ExPEC STs seem to be associated with
resistance, ESBL and virulence genes, enabling them to cause major outbreaks at different locations
simultaneously (54,134,136). Thus, it seems that there is a distinction between resistant isolates and
fully susceptible E.coli (135,137–139). The prevalence of the different ST-lineages varies
depending on country and continent, but ST131 in particular has been reported as widely
disseminated in North America, most parts of Europe, in Asia, the Middle East and Australia
(52,140). In Norway 20% of CTX-M-producing E.coli belonged to ST131 and in Denmark, ST131
has been shown to dominate among the ESBL-producing E.coli constituting 35-45% (52,57,132). In
Canada ST131 comprised <3 % of susceptible isolates but presence of ST131 amongst non-ESBL
has never been studied in Denmark (52,57,132). As a common MDR E.coli-lineage ST131 has been
presented both as a lineages with a high virulence score and as a lineage which harboured less
virulence factors than other uropathogenic E.coli (115,135,141). Nonetheless, ST131 has proven to
have a higher metabolic potential and carry similar virulence genes regardless of location
(7,8,115,136,141–144). It might be the explicit combination of virulence and resistance features
which gives MDR ST-lineages a beneficial competitive advantage and be the possible reason for the
clonal-like global expansion (141,142,145). Thus, studies on collection of E.coli isolates has shown
that a limited set of lineages account for a large proportion of multidrug-resistant strains (7).
Sequence types can often be sub-dived by serotyping, PFGE, fimH sequencing and identification of
virotypes (133,136,146). Thus, most ST131 isolates belong to serotype O25, but a minor part of the
ST131 populations have recently been identified as O16 (132). The subdivision of STs has proven
to be a weakness of the MLST method, since most of the MLST-based lineages consist of highly
heterogeneous groups of strains (133). The spread of resistant E.coli sequence types has been
extensively investigated (7). However, only a few studies have examined non-ESBL and susceptible
E.coli (9,138,147).

Copenhagen 2014
25
2.8. Risk factors for infection with ESBL-producing E.coli
The epidemiology concerning infections due to ESBL-producing Enterobacteriaceae has for some time
been of great concern and as a result has been extensively investigated. The study designs used to
investigate these epidemiological characteristics of infections by resistant Gram-negative bacteria have
often been retrospective case-control studies (148–150).
In the early 2000s infections due to ESBL-producing E.coli were mainly investigated as infections
associated to hospital settings (15,72). As CTX-M became the dominating ESBL genotype, with a
preference for E.coli, these pathogens appeared increasingly from both healthcare settings and the
community, posing a threat to management of empirical treatment. Consequently, research focus
shifted towards E.coli and to some extent the community (15,72,74,76,149,151–155). Doi et al.
found that 36.8% of patients colonized or infected with ESBL-producing E.coli were community-
associated, of which 91.3% produced a CTX-M-type ESBL and 54.2% were caused by ST131.
Cases of UTI were composed of 67.4% community-onset infections caused by ESBL-producing
E.coli and UTI was identified as the most frequent source of bloodstream infections caused by
ESBL-producing E.coli (15,142).
2.8.1. Considerations on study design
The case–control study design is one of the most frequently used methods for identifying risk
factors related to antibiotic resistant pathogens. Yet, the case-case–control study design can more
accurately identify these risk factors than the classic single case–control study design (156,157).
When patients with resistant bacteria are compared to patients with susceptible bacteria, the design
makes it impossible to identify which risk factors are specific for resistant infections and which are
risk factors for infection in general. Control-patients with susceptible strains are not likely to be true
representative of the population for resistant cases and constitutes a risk for selection bias,
especially on the effect of antibiotics (156–161). Patients with the susceptible bacteria could,
however, represent a part of the true source population but are then not included in the control
group. Furthermore, using two different control-groups makes the results less comparable, even if
control-patients are more representative. The control-group should be representative of the source
population, among which all cases are found, and act as controls for all cases. As the source
population is the population from which all case-patients were selected, members of the control-
group with the target organism would have been classified in a case-group (156,157). When the
same control-patients are used in all analyses, comparison of the resulting risk models is easy and

PhD Dissertation
the comparison can identify risk factors specifically associated with one or the other case-group
(156–161).
Overall the case-case-control study design is best suited for investigations relating to infection
control and public health perspectives, when the focus is to detect patients who are at risk for
harbouring particular resistant organisms, and to further implement interventions to limit the spread
of these organisms (156). If the emphasis, on the other hand, is on estimating the likelihood that a
resistant pathogen is involved and hereby identify the most effective empiric antimicrobial
treatment regimen, the control-group must include patients who had clinical cultures performed
(156). Now the control-group does not represent the true source population, since all involved
patients have sought medical attention, but makes the collection of control patients easier (156,158).
The basis of the case-case-control design is the uses of separate case–control analyses within a
single study, performed as parallel t-tests representing the risk factor identification followed by
parallel multivariate logistic analysis (156,161,162).
2.8.2. Case-Control Studies
Many case-control studies of risk factors, included either admitted patients or patients from hospital
settings, and infections were retrospectively classified as either community-onset infections (COI)
or healthcare-associated infections (HCAI), by use of criteria similar to those proposed by Friedman
et al. (15,17,18,73,74,151,155,163–165). As one of the main foci of investigation, exposure to
antibiotics were looked at preceding infections in various periods as within the previous year
(18,73,155,166), 2-3 months (70,149,167–172) and 1 month prior to infection (15,173), with
hospitalization generally studied in similar intervals (17,70,73–75,77,149–
151,153,155,165,166,172,174,175).
Regularly identified risk factors were male sex, age >65 years, prior ESBL-producing E.coli
colonization, previous UTI, urinary catheterization, previous receipt of antibiotics and in particular
previous exposure to extended-spectrum cephalosporins and fluoroquinolones, nursing home
residence and hospitalization within the preceding 12 months (15,17–
19,69,70,155,165,172,173,176,177). Impact of comorbidities and underlying diagnoses´ as risk
factors varied between studies (33,69,73–75,148–153,167,169,170,178). Thus, important
epidemiological factors habitually involved antibiotic selective pressure, rather than individual
comorbidities, in the emergence of ESBL-producing isolates (153). Some investigations also
recognized patients with COI ESBL-producing E.coli who had no preceding health-care exposure

Copenhagen 2014
27
and ESBL-producing E.coli emerged as pathogens in children (<15 years) (15,18,19,155,172,176).
Finally, urinary catheterization and exposure to β-lactamases-inhibitors, cephalosporins, co-
trimoxazole and aminoglycosides were factors associated with an ESBL-infection in patients
previously colonized with an ESBL-producing strain (179).
The dominating β-lactamases identified were of the CTX-M type, primarily from CTX-M group 1,
while TEM and SHV types became less frequent (73,150,151,154,167). Major limitations related to
many case-control studies were the lack of integrated databases for use of medications, the inability
to control the use of non-prescribed antibiotics and the fact that many studies were single case-
studies, where ESBL infections were compared to non-ESBL (74–76,148,149,151).
2.8.3. Double-Case-Control Studies of Epidemiological Factors
A few studies have investigated epidemiological features associated with COI, by use of a double-
case or -control design (19,73,170). All were located at hospital settings including hospitalized
patients and only two studies included an uninfected control group (19,73,170).
In Thailand in 2007, Apisarnthanarak et al. performed a double-case control study including all
infections caused by ESBL-producing E.coli (73). They found previous ESBL colonization and
recent exposure to antibiotics, especially to 3rd generation cephalosporins and fluoroquinolones to
be independent risk factors for any COI with ESBL-producing E coli (73). Risk factors for non-
ESBL infections were stroke and diarrhoea while diabetes was found as a common risk factor for
both groups (73). Interestingly, they demonstrated an association between infections with ESBL-
producing E.coli and increased mortality, longer hospitalization and higher costs (73).
Nicolas-Chanoine et al. studied if patient’s origin and lifestyle was associated with infections with
CTX-M-producing E.coli, as a case-control-control study in Paris in 2012 (19). They compared
ESBL patients to patients with non-ESBL E.coli and uninfected individuals and found that birth
outside of Europe, chronic infections and antibiotic treatment after hospitalization, but before
inclusion, were risk factors for ESBL infections. Furthermore, functionally dependent and collective
housing before hospitalisation was associated with CTX-M-producing E.coli.
2.8.4. Case-Control Study in Low Prevalence Countries
Two research groups have studied the epidemiological factors in low prevalence countries (Søraas
et al., Norway 2013 and Rogers et al., Australia/New Zealand 2014) as single-case control studies

PhD Dissertation
including hospitalized and non-hospitalized patients (17,18). Søraas et al. looked at healthcare
contact and prescriptions of antibiotics and further included travel, diet and recreational swimming
in the analysis (18). Here they found patients with ESBL infections to be of younger age and
identified travel to the Middle East or Africa, recent use of fluoroquinolones and β-lactams (except
mecillinam), diabetes mellitus and recreational freshwater swimming as risk factors with decreased
risk associated with increasing number of fish meals per week (18). Rogers et al. looked at risk
factors for expanded-spectrum-cephalosporin-resistant E.coli and found birth on the Indian
subcontinent, previous urinary tract infection, travel, prior exposure to trimethoprim±
sulfamethoxazole and ±expanded-spectrum cephalosporins and health care exposure in the previous
6 months to be associated with ESBL infections in a low prevalent country.
The European Society for Microbiology and Infectious Diseases (ESCMID) treatment
guidelines for UTI, have changed in recent years, to meet the challenges of empirical treatment of
E.coli. As we have seen a constant development and rapid spread of antibiotic resistance,
continuous local surveillance for resistance combined with identification of patients at risk are
essential to avoid inadequate empirical therapy and adverse outcomes. In order to create
interventions to reduce the prevalence of ESBLs, we need to understand and identify the
dissemination of ESBL-producing E.coli and understand the impact of antibiotic selection pressure
(115).

Copenhagen 2014
29
3. Bacteriological Characterization of E.coli
The typing methods used in these studies are briefly introduced below.
3.1. General Considerations on Typing Methods
Bacterial typing is used to distinguish genetic variation among isolates of the same species. Typing
of bacteria has become a valuable tool for detecting and controlling outbreaks as well as identifying
spread of resistant strains (180). As for now, a variety of different typing methods exists, each with
advantages and shortcomings when it comes to discriminatory power and price. Furthermore, there
is a constant development of new methods in addition to improvements of known techniques and
such advances have to balance between creating new and maintaining existing nomenclature (181).
Maintaining elements of an existing nomenclature, that may have been well understood and
accepted, is essential for general acceptance and use (181).
E.coli can be typed with several different methods, such as phylogrouping, sequence-typing, multi
locus variable number of tandem repeat analysis (MLVA), pulsed-field gel electrophoresis (PFGE),
serotyping, random amplified polymorphic DNA (RAPD) etc., not including subdivision based on
virulence and pathogenic capacities. Such methods have proved crucial in investigation of the
epidemiology of pathogenic E.coli and are of great importance in disease control (181,182).
Additionally, each technique can often subdivide across each other’s boundaries. Therefore,
choosing a typing method requires consideration on economic costs, skills needed by the institution
and analytical properties in regards to what questions needs to be answered.
3.2. Phylogrouping by PCR
The subdivision of E.coli into the phylogroups A, B1, B2 and D by polymerase chain reaction
(PCR), has been a fast and inexpensive manner to divide E.coli into the four major phylogroups
(28,29,183). The first described PCR protocol was a triplex PCR developed by Clermont and
colleagues (28). A method which was adjusted by Gordon and colleagues and further revised by
Clermont and colleagues (29,183). The Clermont method was originally designed to assign E. coli
strains to four phylogroups, why the method clearly not will assign strains not belonging to A, B1,
B2 or D. To which extend strains are miss-assigned by this method will rest on the composition of
the collections evaluated, but an estimated 79% of isolates will be correctly assigned (29). In this
study we used the triplex PCR adjusted by Gordon et al. It is a rapid procedure based on primary

PhD Dissertation
PCR detection of three marker genes (i) chuA (ii) yjaA and (iii) TSPE4.C2, followed by potential
subsequent detection of a fourth gene ibeA. The presence or absence of genes (i)-(iii) generates a
dichotomous decision tree used to determine the phylogenetic group of an E.coli isolate (28,183).
The specific findings of chuA+, yjaA-, TSPE4.C2+ results in the need for secondary screening for
the gene ibeA, to correctly discriminate between phylogroup B2 and D. The presence of ibeA meant
assigning the isolate to group B2. Strains failing to yield any PCR products of the three primary
genes remain unassigned as non-type able (183).
E.coli has traditionally been divided into five main phylogroups (A, B1, B2, D and E). Recently,
other minor phylogroups have been described and now E.coli is divided into a total of seven
different E.coli sensu stricto phylogroups (A, B1, B2, C, D, E and F) with an eighth phylogroup
being the Escherichia cryptic clade Ι. The world of E.coli phylogroups has therefore become
increasingly large, but just 13 % of human faecal E.coli isolates belong to the newly identified
phylogroups C, E, F and clade I, why we here will focus on the four biggest phylogenetic lineages
(A, B1, B2 and D).
3.3. Multi-Locus Sequence Typing
Multi-locus sequence typing (MLST) is a typing method which have proved portable and easily
comparable between laboratories (180,181,184,185). It was introduced in 1998 and a range of
MLST schemes for different organisms has been established and made readily accessible in online
databases. In this study the Achtman scheme for E.coli characterisation was used, and this method
is briefly described. MLST of E.coli is based on the identification of allelic profiles of seven
different housekeeping genes coding for fundamental metabolic functions: adk, fumC, gyrB, icd,
mdh, purA and recA (181,186). It meets the requirement for multiple loci characterisation of
alleles that are distinguishable and easily analysed (180,181,184,185). MLST is a less strenuous
technique than whole genome sequencing, but requires PCR amplification and subsequent
sequencing of the seven genes. It holds the clear advantages of an international on-line
nomenclature (180,181). Each allele is sequenced and by accessing the online MLST database the
determined nucleotide sequence is assigned a unique number
(http://mlst.warwick.ac.uk/mlst/dbs/Ecoli). This produces high levels of discrimination between
isolates. As each allele is given a number, the combination of numbers is used in an MLST scheme
to give each isolates a unique sequence type (ST) (180,181,186). As for whole genome sequencing,
the success of MLST lies on decreased cost for nucleotide sequence determination. MLST is today

Copenhagen 2014
31
the gold standard of characterisation and can be used to describe evolutionary and phylogenetic
relationships (181).
3.4. Multi-Locus Variable Number of Tandem Repeats Analysis
The multi-locus variable number of tandem repeat analysis (MLVA) is a characterisation method
originally based on PCR amplification and size determination of eight variable number of tandem
repeats loci (VNTR), meaning that MLVA exploits naturally occurring differences in DNA regions
with tandem repeats. These regions are found throughout most bacteria as well as mammals, but
only identical cells will have the same number of repeats in each of the targeted regions.
Consequently, identical cells will generate PCR products of matching size (187,188). Each of the
VNTR can be assigned a number depending on size and one isolate will consequently be assigned
an eight-digit code when using the original MLVA method (187,188). The method relies, however,
on accurate size determination of each PCR product.
In this study we used a modified MLVA method described as an abbreviated MLVA (a-MLVA).
Here, only six of the eight original VNTR loci were used and reactions done as singleplex PCR
amplification with unlabelled primers. We used a-MLVA since this method uses loci with repeat
sizes between 12bp and 95bp, meaning that the size of fragments could easily be determined by the
use of an automated capillary electrophoresis system (QIAxcel, Qiagen) and a high-resolution
cartridge (57).
3.5. Serotyping
The serological classification of E.coli isolates, as proposed by Kauffman in 1944, is based on the O
(somatic) and the H (flagellar) surface antigen profiles (95,189,190). Since O and H antigens are
stable this method for strain characterization is considered to be very reliable (189). Serotyping has
therefore been an important technique for differentiating important pathogenic E.coli and classically
used to describe IPEC pathotypes (190). The characterization is based on bacterial agglutination
tests with antigen-antiserum. An E. coli strain which is inagglutinable by O antiserum, but
agglutinable when heated is said to have a K antigen (95). The specific combination of O and H
antigen are characterized as a serotype, while the determination of O antigens only are termed
serogroups (190). IPEC seems to belong to a limited number of O:H serotypes, which to some
extend applies to ExPEC, were virulent E.coli belong to a limited number of O serogroups (190).
More than 175 different O antigens, each defining a serogroup, and more than 50 H antigens are

PhD Dissertation
recognized (95). It is not the serological characterization which confers virulence, but rather the
separate presence of virulence genes (95). For this thesis O serogrouping was performed only, and
executed by GlycoVaxyn as part of an unpublished commercial project.

Copenhagen 2014
33
4. Material and Methods
Methods are described in each individual manuscript. This following paragraph is a short
description of materials and methods not specifically described in these (Figure 2).
Figure 2. Overview of used materials and methods
4.1. General Protocols
Work performed in the laboratory was done according to current guidelines from SSI, DCM as well
as stated by manufacturers of material and products. Furthermore, all media and products were
stored according to manufacturer’s recommendations. Guidelines were followed in agreement with
previously published papers and current literature.
4.1.1. Research Ethical Approvals
When necessary, needed approvals were obtained from Government institutions. Thus all animal
experiments were initially approved by the Danish Centre for Animal Welfare (Current permit:
2013-15-2934-00777 C3) and carried out at licensed animal facilities at SSI. Additionally,
collection of patient samples and patient data was approved by the Danish Health and Medicines
Authority as well as Statistics Denmark and The Regional Committee of Danish Data Protection
Objective 1
Strain: CTX-M-15 producing
E.coli ST131
Selective Agar
Disk disffussion, Etest, Vitek 2,
MALDI-TOF
Mouse Intestinal Colonization
Model
Detection and quantification of
bacteria
MLST
Objective 2
Three case-groups (98vs.174vs.
177).One control group(200)
Epidemiological data (Statistics
Denmark and hospital records)
Multivariate logistic regression
analysis
Disk disffussion, Etest, Vitek 2,
MALDI-TOF
Phylogrouping
ESBL Genotyping
Objective 3
Three case-groups (98 vs. 94 vs.
94).
Serogrouping
(94 vs. 49 vs. 51)
Abbreviated MLVA
MLST
Phylogrouping
ESBL

PhD Dissertation
Agency. The Regional Committee of Health Research Ethics Committee declared notification of
patients unnecessary.
4.1.2. CPR Registry
In 1968 the Danish Civil Registration System (CRS) was established. Now every person living in
Denmark is registered with a unique personal identification number called the CPR-number. The 10
digit CPR-number includes individual information on name, gender, date and place of birth, identity
of parents as well as continuously updated information on address, marriage and vital status. The
CPR is used for identification when in contact with governmental institutions, hospitals, general
practitioner and for prescribing medicine. The CRS is connected to all Danish national registers as
well as bio-banks and this information is available for research purposes (191,192).
4.1.3. Statistics Denmark
Statistics Denmark (DST) is a central authority that collects and publishes statistics on the Danish
society (http://www.dst.dk/en/). They are linked to other Danish national registers such as the
Danish National Prescription Registry (DNPR) and National Patient Registry (LPR)
(http://www.ssi.dk/Sundhedsdataogit/Registre). Both DNPR and LPR are linked to CRS. DNPR is
owned by the Danish Medicines Agency, currently maintained by SSI, and monitors all information
on dispensed prescriptions and drug user. Thus, DNPR collects CPR-data and details on prescribed
drug (ATC-code, dose, dispensing date etc.). However, DNPR does not register over-the-counter
drugs on an individual level or drugs prescribed in hospital settings. The LPR is the National Patient
Registry and monitors all hospital contacts (admissions, outpatient contacts and visits to emergency
room), the departments involved, diagnoses given and surgical procedures. Data on medicine
administered during hospital admission is not available, but data on prescriptions dispensed to
residents at long-term care institutions is. Additionally, diagnoses from general practices are not
available. Hence, DST governs all prescribed medicine, hospital contacts and diagnoses for each
individual with a CPR-number. This extensive data is available for research purposes. The
availability is granted as an encrypted, extern and time-limited access to databases and only for
publications on population- and not individual-level. The CRS linkage with nationwide individual-
level data sources renders DST as a very powerful epidemiological tool (192,193).

Copenhagen 2014
35
4.2. Mouse Intestinal Colonisation Model (Manuscript I)
The mouse intestinal colonisation model and the results gained from this have been thoroughly
described in manuscript I.
We based our experimental mouse model on similar animal models previously used at SSI
(paragraph 2.6.1.) and modified the model through pilot studies. Here follows a short description of
the preliminary studies performed to test and verify its use.
4.2.1. Selection of E.coli Isolates and Preliminary Mice Experiments
Among a collection of Danish E.coli blood isolates at SSI, collected as part of another study (194),
four ESBL-producing E.coli isolates were chosen as possible colonising pathogens. These isolates
were chosen based on their ESBL-genotype, their sequence type 131 and the fact that they were
susceptible to gentamicin (later used in selective plates). The four isolates ((A)64-Ec-09: B2,
ST131, CTX-M-15, AmpC hyper production (B) 65-Ec-09: B2, ST131, CTX-M-15 (C) 69-Ec-09:
B2, ST131, CTX-M-15 and (D) 71-Ec-09: B2, ST131, CTX-M-15) were subjected to an initial
mouse experiment, in the attempt to choose an E.coli isolate appropriate for a reproducible
experimental study of colonisation. The basics of each experiment were similar and described in
detail for the mouse intestinal colonisation model. Based on the mouse experiment, we chose our
colonising pathogen to be 65-Ec-09. Similarly, we tested if resistant E.coli would colonise the gut
without antibiotic treatment and likewise examined the effect of cefotaxime on the indigenous
E.coli population. Furthermore, for the selected strain, we repeated the initial mouse study with
cefotaxime and performed a study with cefuroxime, clindamycin and ciprofloxacin. In these studies,
the colonising ability was established as well as the appropriate interval of faeces collection.
Finally, we did one last pilot study with 65-Ec-09, implementing the faeces collection arrangement
and selection of resistant E.coli and Gram-negative anaerobes. This final pilot was performed with
the antibiotics cefotaxime, cefuroxime, ciprofloxacin, dicloxacillin and clindamycin. Selective
plates for Gram-positive bacteria were not introduced until the second run of the experimental
study.

PhD Dissertation
4.3. Study Populations (Manuscript II and III)
4.3.1. Sample Size Determination
Prior to the initiation of the case-control study we evaluated sample sizes of previous case-control
studies. We used these sample sizes to estimate the needed number of cases in our investigation in
concordance with expected monthly prevalence of community-onset ESBL UTI and an online
calculation of groups sizes, when p=0.05% and 80% power.
A statistician at Hvidovre Hospital calculated the number of isolates needed in the characterization
of susceptible populations.
4.3.2. Strain Collections
The triple-case-control study was based on the prospective collection and examination of urine
samples submitted from general practices to DCM from 1st. of October 2011 to 1st. of July 2012,
combined with the retrospective exploration of patient data. This was done as a non-intervention
study with independent patient-treatment being handled by health-care professionals. All received
samples were handled by the laboratory technicians according to general guidelines and patients
were treated accordingly. An initial susceptibility test was always performed on E.coli from urine
samples and included screening for ESBL-production by the use of cefpodoxime. E.coli with low
susceptibility for cefpodoxime was subjected to a MAST-test and phenotype confirmed.
Composition of case groups are described in manuscript II.
GlycoVaxyn AG determined serogroups for 94 of ESBL isolates, 49 resistant and 51 susceptible
isolates (Manuscript III).
For typing of the E.coli populations (Manuscript III) all 98 ESBL-producing E.coli isolates, 94
isolates from the resistant group and 94 isolates from the susceptible group were included. Due to
cost restraints, we were not able to include all isolates from each of the case groups. A statistician at
Hvidovre Hospital helped us calculate the number of isolates needed to identify the majority of STs
expected to be present in the two non-ESBL populations. Primarily, all isolates with a known
serogroup were selected. Secondly, isolates from patients with no immediate health-care association
(this was before data from DST were obtained) were chosen randomly.

Copenhagen 2014
37
4.4. Epidemiology of collected isolates from cases and controls
Patient data were collected from medical records, when necessary, or obtained from Statistics
Denmark (DST). Data on admissions, diagnoses and prescribed medications from general practices
were acquired from one year prior to, till 90 days after collection of urine sample. Since data from
Statistics Denmark were analysed by external online access only, it was not possible for us to
subsequently add data to the DST database.

PhD Dissertation
5. Results
This chapter includes several tables of results and a brief stating of the main results obtained during
this PhD. It is divided in three sections: The first section includes results found in the selection
study of the mouse intestinal colonization model. The second and third sections are results obtained
during the case-control study and the following characterization of selected E.coli isolates.
5.1. Mouse Intestinal Colonization Model (Manuscript I)
This first section shows results found during the mice experiments presented as major findings in
Table 1 and 2 and in Figure 3, 4 and 5.
Objective I
To evaluate the ability of nine common antibiotics, to select for a CTX-M-15-producing E.coli
isolate belonging to sequence type (ST) 131 in vivo.
By use of the described mouse intestinal colonisation model (manuscript I) we evaluated the
selective abilities of cefotaxime, cefuroxime, dicloxacillin, clindamycin, penicillin, ampicillin,
meropenem, ciprofloxacin and mecillinam on a fluoroquinolone resistant, CTX-M-15-producing
E.coli isolate (65-Ec-09) belonging to the lineage B2-O25b-ST131 (resistance pattern found in
Table 1).
Table 1. Antibiotic susceptibility profile for selected antibiotics for E.coli isolate 65-Ec-09.
Antibiotic MIC mg/l Interpretation
Cefotaxime >64 R
Cefuroxime >64 R
Ampicillin >32 R
Ciprofloxacin >4 R
Gentamicin <1 S
Meropenem < 0.25 S
Mecillinam 8 S
Dicloxacillin >256 R
Clindamycin >256 R

Copenhagen 2014
39
We used 8 mice per antibiotic and CFU of E.coli ST131, Bacteroides and Gram-positive aerobic
bacteria in faecal samples were studied on day 1, 2, 4 and 8. We used the Bacteroides population as
an indicator for antibiotic impact on the Gram-negative anaerobic population. Prolonged
colonization was studied for three antibiotics with added faecal CFU´s studied on day 10 and 14
(cefotaxime, dicloxacillin and clindamycin).
We collected 0.5 grams of faeces from each cage and calculated results of faecal bacteriology as
LOG CFU/0.5 grams of faeces as the average CFU count on two agar plates. The mean CFU from a
total of four cages were then used for statistical calculations (two cages for the Gram positive
aerobic flora). For the graphic depiction we used means and SDs.
5.1.1. Effect of antibiotic selective pressure on the microflora
As seen in Figure 3 we found that cefotaxime, dicloxacillin and clindamycin promoted colonization
and overgrowth of 65-Ec-09 from day 2 through 8 (P < 0.01 for dicloxacillin and clindamycin, P <
0.05 for cefotaxime). Interestingly, both benzylpenicillin and cefuroxime induced overgrowth on
day 2 and 4 (P < 0.05) with dicloxacillin and clindamycin showing the overall highest selective
abilities (P < 0.01 for dicloxacillin on day 2 and 4 and P < 0.01 for clindamycin on all days). For all
antibiotics, there was a decline in the colonization of cefuroxime resistant E.coli after ended
treatment on day 3.
As seen in Figure 4 the remaining antibiotics (ampicillin, mecillinam, meropenem and
ciprofloxacin) showed limited selective abilities and promoted overgrowth of the cefuroxime
resistant E.coli on day 2 only (P > 0.05). Antibiotics impact on the microbiota represented by
Bacteroides and Gram-positive growth are found in shown in Figure 3 and 4. It seems that none of
the antibiotics made an impact on the Gram-positive flora, inhibiting or promoting (P > 0.05).
Essentially we found clindamycin, only, suppressed the Gram-negative anaerobic flora, completely
eliminating the Bacteroides during treatment (P<0.05). As treatment stopped, CFU counts for
Bacteroides increased and returned to counts equal to before treatment.
5.1.2. Effect of antibiotic selective pressure prolonged presence of 65-Ec-09
Data for CFU of 65-Ec-09 on day 1-14 are found in Figure 5. Here we saw that the initial high CFU
count was followed by a rapid decrease for all three antibiotics. For two of them (dicloxacillin and
cefotaxime) the CFU counts were below the detection limit on day 10. For clindamycin, however,
the CFU count was assessable until day 14, which was the last day of the study. Nonetheless, we

PhD Dissertation
saw no significant differences in faecal CFU of cefuroxime resistant E.coli among the three
antibiotics in the study of prolonged colonization (P>0.05).
5.1.3. Molecular tests of bacteria isolated from mice faeces
Results of the molecular tests performed on isolates from mice faeces are found in Table 2.
To correctly identify species found in mice faeces we performed MALDI-TOF on selected isolates.
Furthermore, to properly ensure that cefuroxime resistant E.coli cultured were identical to 65-Ec-09
we tested 17 resistant E.coli isolates found in faeces during treatment and four E.coli isolates from
day 1 (prior to inoculation) for the presence of a blaCTX-M group 1. Finally, one E.coli isolate from
day 1 and five cefotaxime resistant E.coli isolates were characterized by MLST. For these tests we
included isolates from both study runs.
All 21 isolates suspected to be E.coli were confirmed as such by MALDI-TOF and all resistant
E.coli isolates contained a blaCTX-M group 1. None of the E.coli isolates collected prior to
inoculation contained such a gene. All tested resistant E.coli were characterized as ST131 while the
non-CTX-M-producing E.coli belonged to ST602. Table 2 shows E.coli isolates characterized by
MLST as well as a representative for Gram-negative and –positive isolates, which were all
identified as belonging to identical species.
Table 2. Overview of molecular testing of bacterial isolates from mice faeces.
Antibiotic Day of experiment MALDI-TOF blaCTX-M group-1
present
MLST
Dicloxacillin 4 E.coli Yes ST131
Clindamycin 4 E.coli Yes ST131
Cefotaxime 2 E.coli Yes ST131
Cefuroxime 2 E.coli Yes ST131
Clindamycin 10 E.coli Yes ST131
Dicloxacillin 1 E.coli No ST602
Dicloxacillin 2 Parabacteroides spp. - -
Clindamycin 2 Enterococcus faecalis - -

Copenhagen 2014
41
Figure 3. LOG/CFU per 0.5 grams of faeces shown for antibiotics with selective effect on 65-Ec-09. The three
cultured populations are shown. Where SD is not shown SD=0.
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
80
5
10
15
Cefotaxime resistant E.coli
Clindamycin
CefotaximeCefuroximeDicloxaillin
Penicillin
Days After Inoculation
LOG
CFU
/0.5
g
Anaerobic Gram-negative
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
80
5
10
15
Days After Inoculation
LOG
CFU
/0.5
g
Gram-pos. aerobic
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
80
5
10
15
Days After Inoculation
LOG
CFU
/0.5
g

PhD Dissertation
Figure 4. LOG/CFU per 0.5 grams of faeces shown for antibiotics with limited selective effect on 65-Ec-09. The
three cultured populations are shown. Where SD is not shown SD=0.
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
0
5
10
15AmpicillinMeropenemCiprofloxacinMecillinamControl+ 65-Ec-09
Cefotaxime resistant E.coli
Days After Inoculation
LOG
CFU
/0.5
g
Anaerobic Gram-negative
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
80
5
10
15
Days After Inoculation
LOG
CFU
/0.5
g
Gram-pos. aerobic
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
8
Day
1
Day
2
Day
4
Day
80
5
10
15
Days After Inoculation
LOG
CFU
/0.5
g
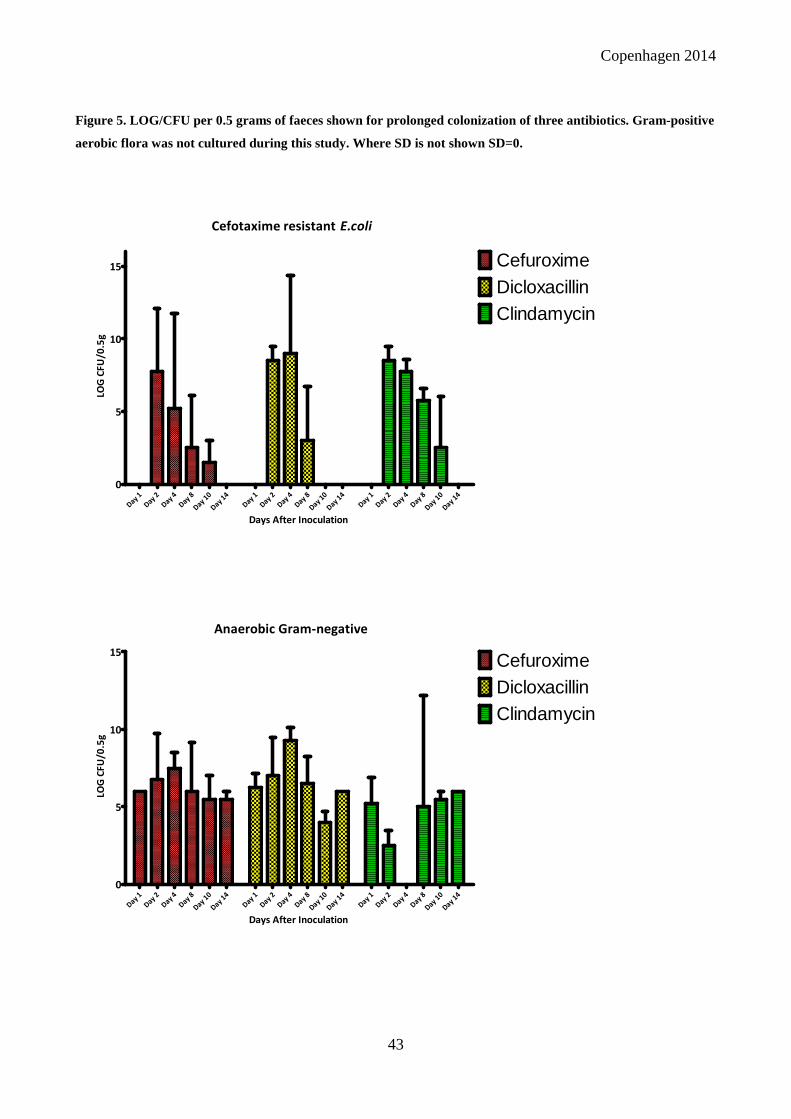
Copenhagen 2014
43
Figure 5. LOG/CFU per 0.5 grams of faeces shown for prolonged colonization of three antibiotics. Gram-positive
aerobic flora was not cultured during this study. Where SD is not shown SD=0.
Cefotaxime resistant E.coli
Day 1
Day 2
Day 4
Day 8
Day 10
Day 14
Day 1
Day 2
Day 4
Day 8
Day 10
Day 14
Day 1
Day 2
Day 4
Day 8
Day 10
Day 14
0
5
10
15 CefuroximeDicloxacillinClindamycin
Days After Inoculation
LOG
CF
U/0
.5g
Anaerobic Gram-negative
Day 1
Day 2
Day 4
Day 8
Day 10
Day 14
Day 1
Day 2
Day 4
Day 8
Day 10
Day 14
Day 1
Day 2
Day 4
Day 8
Day 10
Day 14
0
5
10
15CefuroximeDicloxacillinClindamycin
Days After Inoculation
LOG
CF
U/0
.5g

PhD Dissertation
5.2. Case Control Study (Manuscript II)
Objective II
To investigate epidemiological factors associated with acquiring UTI with ESBL-producing E.coli,
in general practices.
In the study period we collected 98 ESBL positive urine samples, 174 resistant E.coli, 177
susceptible E.coli and 200 uninfected urine samples, respectively. All associated E.coli isolates
were analysed molecularly. A few patients were, however, excluded from the analysis of
epidemiological data from DST, due to a missing date of birth. Hence, we excluded two patients
from the ESBL group, three from the resistant group and two from the susceptible group. No
patients were excluded from the uninfected group
5.2.1. Urine samples to DCM
An overview of urines submitted to DCM is shown in Table 3. We have calculated p-values by use
of Fishers exact test. These data are listed in written text when appropriate.
In the years 2011 and 2012 more than 450 general practitioners submitted 110.471 urine samples to
DCM. Of these 48.541 (44%) were positive for bacterial growth and 23.001 (47%) were positive for
growth of E.coli with 4% of E.coli isolates being resistant to cefpodoxime (CPD-R). Determination
of CPD-R is used as an indicator of possible ESBL-production.
For all 649 patient primarily included in the study, a total of 1773 urine samples were submitted to
the DCM for investigation within one year prior to sampling. More than 90% of urine samples from
the three case groups were positive for bacterial growth, with just 33% of the samples for the
uninfected control group.
Patients in the ESBL and resistant groups had submitted significantly more urine samples to DCM
compared to the uninfected group, during 365 days, as well as within 90 days, prior to sampling
(P<0.05 for all). Compared to the uninfected control group, we found that all three case groups had
significantly more urine samples positive for bacterial growth and significantly more were positive
for E.coli within 90 days before and after sampling (P<0.0001). We observed significantly more
urine samples positive for CDP-R E.coli in the ESBL group than all other groups (P<0.0001) before
and after sampling. However, the three case groups submitted significantly fewer positive urine

Copenhagen 2014
45
samples in the 90 days after the index UTI compared to the prior 90 days. Furthermore, the ESBL
group submitted significantly less CPD-R E.coli (P<0.0001). There was no change in the number of
CPD-R E.coli in the other case groups.
Table 3. An overview of urine samples submitted to DCM from 365 days prior to and within 90 days after
sampling. ×The percentage shown at positive samples are the percentages of submitted urine samples being
positive. . *P<0.05 compared to the uninfected group. ▪P<0.05 compared to the resistant group. °P>0.05
compared to the susceptible group.
Positive samples E.coli n (%) CPD Resistant E.coli n (%)
ESBL (n=98) 341*° (91.2×) 192* (56.3) 138*° (71.9)
Negative (n=200) 129 (33×) 31 (24) 3 (9.7)
One year before
index sample
Resistant (n=174) 529*° (90.6×) 339* (64.1) 12*° (3.5)
Susceptible (n=177) 388* (91.5×) 264* (68) 0
All (n=649) 1387 (78.2×) 826 (59.6) 153 (18.5)
90-0 days before
index sample
ESBL (n=98) 183* (93.4×) 126* (68.9) 105*▪° (57.4)
Negative (n=200) 77 (26.1×) 22 (28.6) 3 (3.9)
Resistant (n=174) 328* (96.2×) 229* (69.8) 8° (2.4)
Susceptible (n=177) 281* (94.9×) 215* (76.5) 0
All (n=649) 869 (77×) 592 (68.1) 116 (13.3)
0-90 days after
index sample
ESBL (n=98) 117*° (83×) 60*° (51.3) 48*▪° (80)
Negative (n=200) 39 (48.1×) 10 (25.6) 0
Resistant (n=174) 159*° (88.8×) 77*° (48.5) 4* (5.2)
Susceptible (n=177) 101* (81.5×) 44* (43.6) 1 (2.3)
All (n=649) 416 (79.2×) 191 (45.9) 53 27.7)

PhD Dissertation
5.2.2. Description of E.coli Populations
Susceptibility patterns and phylogroups
As indicated in the group criteria all isolates from the susceptible group were fully susceptible to all
tested antibiotics. Susceptibility patterns and distribution of phylogroups are shown in Figure 6 and
Table 4, respectively. We found phylogroup B2 and D to be the dominating phylogroups in all three
case groups, representing 41-58% and 22-28% respectively.
Percentage of Isolates Resistant to antibiotics
0 20 40 60 80 100
Ampicillin
Cefuroxime
Gentamycin
Aztreonam
Ciprofloxacin
*Sulfamethoxazole
Trimethroprim
*Tetracycline
Ceftazidime
Ampicillin/Clavulanic acid
Tobramycin
Nitrofurantoin
*Piperacillin/Tazobactam
Mecillinam
*Fosfomycin
Meropenem
ESBLResistant
Percentage of Isolates
Ant
ibio
tics
Figure 6. Antibiotic susceptibility pattern for E.coli isolates in the ESBL-producing and the resistant (non-ESBL)
groups.

Copenhagen 2014
47
Table 4. Distribution of phylogroups as found by use of the early Clermont et al. methodology amongst all
collected E.coli isolates. # = Number of isolates.
ESBL ISOLATES n = 98 RESISTANT ISOLATES n = 174 SUSCEPTIBLE ISOLATES n = 177
Phylogr. # Percent # Percent # Percent
A 9 9% 20 11% 19 11%
B1 6 6% 5 3% 2 1%
B2 57 58% 71 41% 91 51%
D 22 22% 49 28% 42 24%
Non-Type 4 4% 29 17% 22 12%
ESBL Genotypes
The distribution of ESBL genotypes are shown in figure 7.
Figure 7. Distribution of major ESBL Genotypes amongst the ESBL-producing E.coli isolates.
For the ESBL population 73% carried a blaCTX-M group 1 and 24% a blaCTX-M group 9. For the
remaining two isolates we were not able to detect a blaCTX-M and found blaTEM only. A total of 54%
of the isolates belonged to CTX-M-15 with CTX-M-14 and CTX-M-27 as two other large groups of
12% and 12%, respectively. Fifteen isolates also had an AmpC phenotype, but no plasmid-borne
AmpC gene was found in any of the strains.
TEM
CT-X-M Gr. 9
CTX-M Gr. 1

PhD Dissertation
5.2.3. Statistical Analysis of Risk Factors and Population Data
In the following paragraphs results of statistical analysis for risk factors are briefly listed.
Results from the initial univariate t-tests are not shown. Patient demographics and exposure to
antibiotics, defined as prescriptions one year prior to sampling, are found in Appendix 1.
Appropriate p-values from chi-square and Fisher´s exact test are found Appendix 2 and 3 in
combination with the results of the multivariate logistic regression analyses.
5.2.3.1. Multivariate Logistic Regression Models
The most important findings will be stated below.
Patient characteristics
Classifications as COI, HCAI and complicated UTI have been done according to previously stated
criteria defined in Manuscript II. Exposures to antibiotics are depicted in Appendix 1 as number of
individuals who received prescriptions and total number of prescriptions prescribed.
Patients included in the three case groups proved to be significantly older than the uninfected
controls (P<0.0001). We found, however, no significant difference between ages in the case groups
(P>0.05). Interestingly there were more men among the uninfected group than in the resistant and
the susceptible groups, but no significant difference in the number of men between the case groups.
Nonetheless, in the multivariate analysis, we found a significant difference between gender as well
as age for all case groups (P<0.005 and P<0.0001).
Epidemiological features for case groups compared to the uninfected group
The multivariate logistic regression analyses are shown in Appendix 2. We performed the
multivariate analyses for the time periods 365 days, 90 days and 30 days before and 30 days and 90
days after sampling. We have, nevertheless, decided to describe findings for 90 days and 30 days
prior to sampling and briefly included selected interesting results from the other time periods.
In the 90 days prior to sampling we found limited antibiotics and diagnoses to be independent risk
factors. For antibiotics, only use of penicillin in the backward elimination analysis was significant
for the ESBL group (P=0.015). Conversely, exposure to five and two specific antibiotics proved to
be independent risk factors for the resistant and susceptible groups, respectively (all P<0.05).

Copenhagen 2014
49
Looking at 30 days prior to sampling, exposure to antibiotics in general proved statistically
significant for all three case groups, when analysed by chi-square (P<0.0001). Yet, just mecillinam
(P<0.0001) was a risk factor for the two resistant groups with mecillinam and sulfamethoxazole
(P=0.003 and P=0.03) being independent risk factors in the susceptible group. For the ESBL group,
diagnoses “kidney infections” and “urosepsis” were risk factors (P=0.01 and P=0.03). Of note, only
for the susceptible group use of antibiotics was an independent risk factor (data not shown).
When looking, in particular, at the backward stepwise elimination procedure, only including
variables with P<0.1, use of antibiotics increased in significance for all three groups. The significant
antibiotics have been added to Appendix 2.
Among the E.coli case groups we found no significant difference in number of community- and
healthcare-associated infections, as defined by us (P>0.05, results not shown).
In the period of 90 days after sampling, use of sulfamethizole was significant in all case groups
(P=0.03, P=0.007 and P=0.001) and mecillinam in the ESBL group (P=0.006) with use of
nitrofurantoin showing significance in the two other case groups (P=0.008 and P=0.04).
Epidemiological features for the ESBL case group compared to the other case groups
These results are found in Appendix 3. When calculated by chi-square there were significantly more
HCAI patients in the ESBL group compared to the susceptible group (P=0.006).
The number of patients receiving antibiotics and total number of prescriptions made did not clearly
show that the ESBL group had a higher use of antibiotics before the index sample (Appendix 1).
The multivariate analyses in the two runs, identified the total use of antibiotics as a risk factor for
the ESBL group 90 days prior to index sample (P=0.0049 and P=0.0029). Hospital admission 30
days prior was likewise an independent risk factor for ESBL (P=0.0073 and P=0.0216).
Finally, we saw that 90 days after infection, the use of penicillin/beta-lactamase inhibitors were
significantly higher in the ESBL group compared to the resistant group (P=0.02) and the use of
penicillins with effect on Gram-negatives and specifically the use of mecillinam were higher in the
ESBL than in the susceptible group (P=0.04 and P=0.002).

PhD Dissertation
5.3. Characterization of E.coli populations (Manuscript III)
Objective III
To investigate if the population structure of uropathogenic E.coli populations is predominantly
composed of related isolates, regardless of resistance profile.
The phylogroup and ESBL genotype distributions as well as the susceptibility patterns of the E.coli
populations included here were very similar to what was found during the case control study (Table
5 and Manuscript III). Briefly, antibiotic resistance for the resistant group was found with 84%
being resistant to ampicillin and 46-68% being resistant to trimethoprim, tetracycline and
sulphonamides.
Table 5. Distribution of phylogroups amongst E.coli isolates included in the characterization study. # =
number of isolates.
ESBL n = 98 Resistant n = 94 Susceptible n = 94
Phylogroups # Percent # Percent # Percent
A 9 9% 7 7% 9 10%
B1 6 6% 3 3% 1 1%
B2 57 58% 42 45% 52 55%
D 22 22% 30 32% 22 23%
Non-Type 4 4% 12 13% 10 11%
5.3.1. Distribution of Serogroups, MLVA codes and Sequence Types
Distribution of serogroups, a-MLVA codes and Sequence Types is found in Figure 8-11 and in
detail in Appendix 4-6.
Of the ESBL-producing isolates 94 were sent for serogrouping accompanied by 49 resistant (non-
ESBL) isolates and 51 susceptible E.coli. We found a variety of serogroups amongst the selected
isolates, with 53 different serogroups. For three isolates (two in the ESBL and one in the susceptible
group) it was not possible to distinguish between two serogroups.
There were 17% of the ESBL isolates, 12% of the resistant isolates and 14% of the susceptible
isolates which were classified as either “Negative” or “Multiple” after serogrouping.

Copenhagen 2014
51
PCR amplification of the six MLVA alleles was done as previously described and this abbreviated
MLVA was used to type the three strain collections. All isolates (100%) were typable by a-MLVA
and all isolates except for one, selected for MLST, had alleles successfully sequenced. One isolate
with a unique a-MLVA code was not sent for sequence typing.
In these three populations we found a total of 83 a-MLVA-codes and 72 sequence types. Of these
13 a-MLVA´s and 13 ST´s were found in two out of three populations and six a-MLVA´s and seven
ST´s were found in all three populations. Overall 10 a-MLVA codes were classified as unique
“unknown” sequence types, here classified as “New STs”. Generally, ST131 constituted 23% of all
isolates, ST69 constituted 10% and ST73 a total of 9%. Of the found a-MLVA codes 74 identified
just one sequence type. However, there were quite a few situations where one specific ST was
subdivided by more than one a-MLVA code, but also more complex situations where several
different ST´s were assigned the same a-MLVA code.
As result:
- ST 10 were subdivided by three different a-MLVA codes
- ST38 by three codes
- ST69 by three codes
- ST73 by ten codes
- ST95 by three codes
- ST141 by three codes
- ST357 by two codes
- ST405 by two codes
- ST648 by two codes and lastly
- ST131 were subdivided by four different codes.
Likewise, we found that the a-MLVA method could not distinguish certain sequence types and in
particular eight a-MLVA-codes did not classify unique sequence types. As such
- ST58, ST101 and ST448 shared one a-MLVA code. ST58 and ST448 are double-locus
variants.
- ST998 and one of the a-MLVA codes for ST141 were identical, were ST141 and ST998 are
single-locus variants.
- It was not possible to discriminate between on isolate of ST14 and ST1193 which are also
single-locus variants.

PhD Dissertation
- ST38, ST117 and ST1177 were assigned a single a-MLVA code. Here ST38 and ST1177
are single locus variant with ST117 being unrelated.
- One a-MLVA code for ST10 also identified ST746, ST1598 and one New ST. Here ST10
and ST746 are single-locus variants while the new ST is a double-locus variant of ST10.
- A different ST10 a-MLVA code was identical to codes for ST93, ST540, ST617, ST2279
and a New ST where only ST617 belonged to ST Complex 10 and was a double-locus
variant of ST10.
- Also ST354 and a New ST had the same code.
- Finally ST88, ST410 and one New ST had a common code with ST88 and ST410 being
double locus variants and belonging to ST Complex 23.
The ESBL-producing E.coli Population
In this ESBL population we found a total of 26 serogroups, 20 a-MLVA codes and 20 different
sequence types. We found no New ST´s in this population. In the serogroups we found one large
cluster with 38% belonging to group O25 and some minor clusters with 6% being serogroup O153,
5% serogroup O16 and 4% serogroup O15 (Appendix 4).
As described above, we saw different a-MLVA codes recognized as identical ST´s and found
different ST´s with indistinguishable a-MLVA codes. Among these isolates there were seven
clusters of a-MLVA codes and five clusters of ST´s. The population was clearly dominated by a
few ST´s as 50 isolates belonged to ST131. These ST131 isolates were identified by three a-MLVA
codes, where the largest of these (153562) contained 44 isolates. Of the 44 isolates, 35 belonged to
the serogroup O25 with 2 isolates belonging to serogroup O77 and O97. The remaining isolates had
no specific serogroup identified. The two other ST131 a-MLVA codes were comprised of 5 and 1
isolates, respectively, all of which belonged to serogroup O16.
The Resistant E.coli Population
Here 19 serogroups, 36 a-MLVA codes and 30 ST´s were identified. Among the isolates there were
three larger clusters with serogroup O25, O75 and O73 totalled 16%, 14% and 10%, respectively. In
addition, there were some minor clusters with serogroup O6, O11 and O15 representing 6-8%
(Appendix 5).

Copenhagen 2014
53
The a-MLVA codes and ST´s were composed of eight MLVA clusters and eight clusters of ST´s
and the population was, like the ESBL-population, dominated by few clusters of ST´s. Hence,
ST69, ST73 and ST131 comprised the largest ST cluster with 17, 10 and 14 isolates, respectively.
The Susceptible E.coli Population
In the population of susceptible isolates we found two larger clusters of serogroups and numerous
very small collections or single-isolate serogroups. The two clusters were serogroup O2 and O6
with 18% and 10% (Appendix 6).
Among the susceptible isolates 29 serogroups, 54 a-MLVA codes and 44 ST´s were found. We saw
seven a-MLVA clusters and eight clear ST clusters. Here ST73, as identified by multiple codes and
ST95 dominated, but also a-MLVA codes for ST10/93/540 and ST141/ST998 were frequent. This
population was comprised of a more diverse and heterogeneous group of E.coli.
Percentage of identifed a-MLVA codes,STs and O-groups
MLVAMLST
Serogro
ups0
50
100
150 Susceptible
Resistant
ESBL
Typing methods
Figure 8. Shown are the percentages of a-MLVA codes, STs and O-serogroup found in each of the three
populations.

PhD Dissertation
Figure 9. Distribution of major serogroups among the three E.coli populations. We show groups with 4% or more of isolates. Groups with <4% are found among singe isolates serogroup.
O25
Single Isolate Serotypes
Multiple
O153
Negative
O16O15ESBL
O25
O75
Single Isolate Serotypes
O73
O6
O15
Multiple
O11
O2NegativeResistens
Single Isolate Serotypes
O2
O6
Multiple
O1
O4
O18
O88Negative
Susceptible
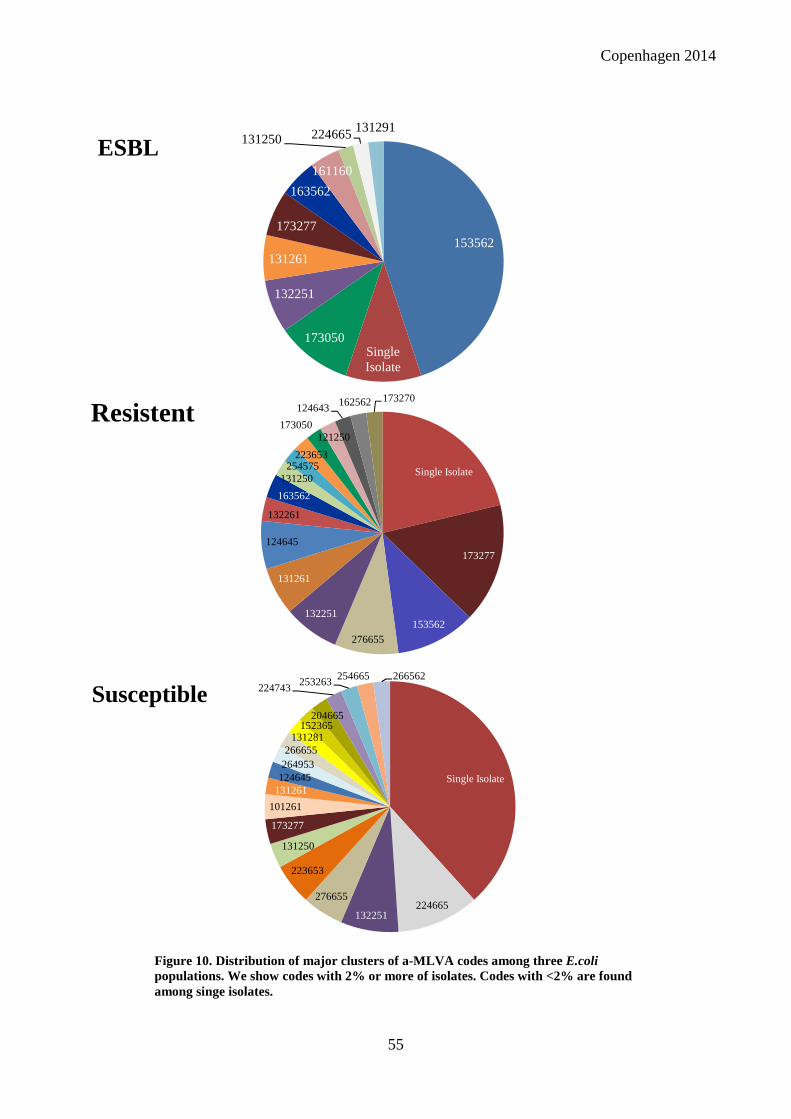
Copenhagen 2014
55
153562
Single Isolate
173050
132251
131261
173277
163562
161160
131250 224665131291
ESBL
Single Isolate
224665132251
276655
223653
131250
173277
101261
131261124645264953266655
131281152365
204665
224743253263 254665 266562
Susceptible
Single Isolate
173277
153562
276655
132251
131261
124645
132261
163562
131250254575
223653
173050121250
124643 162562 173270
Resistent
Figure 10. Distribution of major clusters of a-MLVA codes among three E.coli populations. We show codes with 2% or more of isolates. Codes with <2% are found among singe isolates.

PhD Dissertation
ST131
Single Isolate ST
ST38
ST69
ST10
ST101
ST648
ST998 ST2852
ESBL
Single Isolate ST
ST69
ST131ST73
ST10
ST1193
ST58
ST405
ST95
ST88 ST80 ST1597
Resistent
ST73
ST998
ST95
ST141ST10
ST69
ST127
ST93
ST223
ST357
ST14
ST38
ST101
ST12ST48
ST59ST540
ST4235ST131Susceptible
Figure 11. Distribution of major clusters of sequence types among three E.coli populations. We show STs with 2% or more of isolates, except for ST131 which only constituted 1% of isolates.

Copenhagen 2014
57
6. Discussion
6.1. Selection of CTX-M-producing E.coli in vivo
One of the advantages of using animal models is the relatively rapid study of large populations. In
addition, animal experiments can be repeated and are reproducible (195). Testing on animals, however,
can give an estimation of effects only and there are the obvious limitations of extrapolating from animals
to humans.
We developed a mouse model for the investigation of intestinal colonization without the need for
prior suppression of the indigenous flora by antibiotics. It was hereby possible to illustrate the
selective ability of common antibiotics with different spectra, when an ESBL-producing, virulent
E.coli lineage was introduced into the intestines. We confirmed that an antibiotic, clindamycin,
suppressing the Bacteroides population, with no activity against Enterobacteriaceae, allows for
overgrowth of an ESBL-producing E.coli in the mouse gut. Conversely, antibiotics active against
the colonizing strain will not promote overgrowth. Beta-lactam antibiotics with no inhibiting effects
on the ESBL-producing E.coli showed diverse levels of selection. Interestingly, dicloxacillin
promoted proliferation of the ESBL-producing E.coli, even with no apparent impact on the Gram-
negative anaerobic flora or other Gram-negative bacteria. On the other hand, and very surprisingly,
ciprofloxacin, with known effect on Enterobacteriaceae, had no selective abilities. Yet, treatment
with fluoroquinolones can in some instances reach faecal concentrations above MIC of even
resistant strains, a feature not measured by us (93). There are several other limitations concerning
our mouse model, besides possible problems regarding extrapolation from mice to humans. Most
importantly however, we did not investigate antibiotic impact on the total microflora of the
intestines. We could therefore not study shifts in dominating phyla or changes in the bacterial
species. Finally, it was not possible to observe alterations in the Enterobacteriaceae population
which could potentially describe features influencing colonization.
Nonetheless, we found that the indigenous microflora provides an important protection by
inhibiting colonization. Our findings strongly indicate that other parts of the flora than Gram-
negative anaerobes play an important part upholding colonization resistance. Antibiotics may
therefore disrupt the indigenous microflora, inhibiting colonization resistance and promoting
proliferation. This disturbance may be caused by a variety of antibiotics, and level of selection is

PhD Dissertation
unforeseen by spectra. Antibiotic with limited impact on Gram-negative bacteria can select for
resistant E.coli, as seen for penicillin and dicloxacillin (196).
Disruption of the anaerobic flora, or at least the Bacteroides population, seems to play a particularly
important part in preventing prolonged colonization by MDR pathogenic E.coli.
6.2. Investigations of epidemiological factors
The case-control study performed here is, to our knowledge, the first triple-case-control study of
epidemiological characteristics for UTI caused by E.coli. It has, however, been designed with
several limitations. First, the study exclusively includes non-hospitalized patients of all ages with no
knowledge on antibiotic treatment given in hospitals. Secondly, as we included patients with
previous UTI we might be collecting and investigating patients in the middle of a course of
infection, posing an apparent cause for antibiotic prescriptions. It would have been ideal, if we had
selected only cases where the E. coli infection was the first UTI with at least one year lapse to
previous UTI. The lack of data on nursing home residence as well as outpatient contacts will lead us
to underestimate the significance of healthcare exposures and make it impossible to truly identify
the impact of these locations as reservoirs for ESBL-producing E.coli.
Nevertheless, in this unique triple-case control study we found exposure to antibiotics to be
associated with UTI in general. Healthcare contact and previous UTI infection by ESBL-producing
E.coli were associated with community-onset UTI with ESBL-producing E. coli. This is not
surprising based on previous studies of epidemiological features (15,17,18,70,73,197,198).
However, the new aspects of the epidemiology identified here were the very few differences found
between the UTI case groups. There was no clear difference in exposure to antibiotics and no
significant difference in age or gender, nor was there a significant difference (P>0.05) between the
percentages of HCAI infections in the age group 18-64 and >65 (Appendix 1). There was,
conversely, an association between hospital admissions and ESBL infection when comparing the
case groups. This confirms hospital contacts as an independent risk factor for ESBL-producing
E.coli infections (15,17,18,70,73,197). The ESBL group had significantly more UTIs caused by
CPD-R E.coli, in all investigated time periods. Antibiotics found to select in the mouse intestinal
colonisation model were not identified as risk factors in this present case-control study.
Nevertheless, in Denmark cephalosporins are prescribed in hospital setting only, data we did not

Copenhagen 2014
59
investigate. Furthermore, very few patients were exposed to dicloxacillin and clindamycin which
might be the reason we did not find them as risk factors.
In conclusion, we found exposure to antibiotics to be an unreliable predictor of UTI with resistant
E.coli. We could not link specific diagnoses to the ESBL group, except kidney infections and
urosepsis. The combination of our finding with previous studies suggests that, in low prevalent
countries at least, exposure to antibiotics, comorbidity, age and gender are not necessarily decent
predictors of risk, but healthcare association, travelling and previous carriage of, and infection by,
ESBL-producing E.coli are features which needs to be explored by GPs (17–19,73,93,179).
6.3. Characterization of E.coli populations
We characterized three E.coli populations with different susceptibility patterns by use of an easily
performed and fast typing method. The abbreviated MLVA could assign 71% of isolates to
corresponding sequence types, simply comparing this novel method to a gold standard method with
known nomenclature. Data can furthermore be reproduced and compared to other laboratories. In
this study we chose to apply different typing methods on parts of our collection of isolates. This
increases the discriminatory power enabling us to identify the diversity of different UPEC
populations (190). With the availability of whole genome sequencing, at lower costs and with high
discriminatory power, new methods are at risk of being typing methods with no real impact, unless
price and speed overcomes lack of power (181). There is, however, still need for cheap analytic
tools for surveillance of resistant lineages and detection of outbreaks, a need persistently nourished
by the dissemination of MDR E.coli (11).
MLST and MLVA schemes analyse a small portion of the genome only, and these genes might be
subject to lateral transfer between strains, leading to somewhat incongruent phylogenetic trees, not
correlating with the genome content of a bacterium (20). Changes in VNTR regions will not
necessarily mean a change in ST, and vice versa. There is a higher rate of sustainable genetic
alterations within the VNTR regions, than in the housekeeping genes used for MLST. This might be
the reason for incomplete separation of all STs, but it provides a high discriminatory power and the
ability to separate closely related ST-lineages (187,188). The limitation of the abbreviated MLVA
was the inability to distinguish specific STs, which has been previously described for many of the
MLVA codes. Often the indistinguishable isolates were closely related STs, perhaps indicating a
kinship not recognized by MLST. By use of serogrouping and a-MLVA we described the STs as

PhD Dissertation
extremely heterogeneous lineages. We found, as has previously been seen, that ESBL-producing
E.coli are clustered in homogenous populations dominated by few lineages (199). We identified a
high level of diversity among susceptible E.coli isolates, a level of diversity which, to our
knowledge, has not previously been described (9,131,136,142). The successful lineage of ST131
was similar to other studies dominating this Danish ESBL-producing E.coli population.
Interestingly, a-MLVA seemed to sub-divide ST131 isolates based on O-serogroup, showing the
abilities to survey minor differences in E.coli lineages. The study is of course limited by the
relatively small number of isolates characterized and by the fact that the collection of isolates within
a very limited time period does not allow us to identify the fluctuation in dominant lineages of non-
ESBL populations.
We do, nonetheless, conclude based on serogroups, a-MLVA and MLST, that in this collection of
uropathogenic E.coli from general practices the populations were dominated by different
serogroups, MLVA-codes and ST´s and that the susceptible E.coli population was a much more
diverse group of isolates. Within a dominating ST-lineage, like ST73, we could sub-divide isolates
by a-MLVA. Conversely, the resistant E.coli populations and in particular the ESBL-producing
E.coli are disseminated as related lineages, indicating that the success of resistant lineages is mainly
due to positive selection of UPEC with gained resistance and limited loss of fitness.

Copenhagen 2014
61
7. Conclusion and Perspectives
Our knowledge on the mechanisms behind antibiotic disruption of the microbiota is limited. We
know that antibiotic therapy alters the composition of the microflora and healthy volunteers
undergoing a 5-day course of ciprofloxacin had changes in the flora not resettling for months (3,12).
Antibiotic treatment changes the microbiota and selects for antibiotic resistant pathogens increasing
the risk of dissemination of resistant clones, subsequently increasing risk of infection (11,200). Our
first objective was to evaluate the ability of common antibiotics, to select for a CTX-M-15-
producing E.coli isolate belonging to ST131 in vivo. This was successfully done in a mouse
intestinal colonisation model developed and tested as part of this research. We found that antibiotics
with variation in spectra of activity can select for MDR, virulent E.coli lineages, making new
studies on antibiotic impact on the microflora a necessity to understand selection.
The current increase in faecal carriage of ESBL-producing E.coli, seen in healthy individuals and
returning traveller’s, strongly suggests a rapid dissemination of ESBL-producing isolates in the
community (104,107,108,201). Reports of carriers with no recent hospitalization or antibiotic
treatment indicate long-term carriage proposing a further risk for person-to-person transmission
(108). To what extent ESBL-producing E. coli will spread in the community is unknown, but the
potential for dissemination of resistant lineages like ST131 has proven exceptionally high
(104,199,202). Especially the combination of virulent E.coli lineages and the CTX-M type ESBL,
often showing resistance to ciprofloxacin, shows that the dissemination could be related to the
selective pressure exerted by cephalosporins or fluoroquinolones (104). However, since different
CTX-M types differ in resistance, it might reflect the various antibiotics pressure and reservoirs
involved (104).
Our second objective was to investigate epidemiological factors associated with acquiring UTI with
ESBL-producing E.coli, in general practices. We conducted a case-control study and identified new
aspects of which risk factors are associated to COI UTI in Denmark. The results of our triple-case
control study, in concordance with previous findings, raises the question if COI ESBL infections are
in fact healthcare-associated, at least in countries of low ESBL prevalence (42). Previous antibiotic
treatment was a risk factor for UTI as has previously been seen for UTI in general and for UTI
resistant bacteria (11). We found that patients in the ESBL group were more likely to belong to this
particular group, prior to our sampling but also after sampling, than the non-ESBL case groups.
These individuals might be periodic low-level carriers or treatment can correctly eradicate ESBL-

PhD Dissertation
producing E. coli, why we found significantly less ESBL-producing E.coli during the 90 days after
sampling (93). Yet, strictly COI with ESBL-producing E.coli are being reported and true clusters of
cases in the community, and among family members, are described (42). Overestimating the risk of
ESBL-producing E.coli can lead to the use of broad-spectrum antibiotics leading to a higher
selection pressure hereby preserving existing isolates and allowing for the emergence of novel
resistant strains (179).
As our third objective we wished to investigate if the population structure of uropathogenic E.coli
populations is predominantly composed of related isolates, regardless of resistance profile. Such a
characterization is of great interest to help determine if ST-lineages producing and spreading ESBL
are successful UPEC adapting to environments with antibiotics or if plasmids, carrying ESBL, are
taken up by less prevalent UPEC gaining a new advantage.
This characterization of E.coli populations was successfully done by typing of the three
susceptibility groups by use of a-MLVA, MLST and serogrouping. The a-MLVA method was here
efficient in the characterization of resistant E.coli populations and could possibly help sub-divide
MLST-lineages (57,187,203). In non-clonal populations, there is no lineage structure why such
populations are a bigger challenge when it comes to bacterial typing, than typing of highly clonal
populations (181). Information from a single locus or few loci can be unreliable in identifying
genetic relatedness, why the use of multiple loci is essential to achieve the resolution required
(181,184). Generally it can be more efficient to examine genes displaying more diversity especially
in population investigations (181). This makes the combination of MLVA and MLST a potential
robust method for surveillance. When studying heterogenic E.coli populations by MLST only,
community ExPEC are classified as clonal groups of E. coli, regardless of drug resistance (8). This
would advocate that antibiotic resistance is not a requirement for related dissemination and suggests
that susceptible E. coli strains do disseminate clonally (8). Yet, when such E.coli populations are
described by more than one typing method, the susceptible isolates are found to be a heterogonous
group, as previously determined by fimH-typing and as determined by us in this thesis. We
conclude that the susceptible E.coli populations are not disseminated as related lineages in nearly
the same degree as ESBL-producing E.coli. Reports from other research groups, suggests that some
E.coli lineages acquire different types of mobile resistance genes, as seen with ST69 and especially
ST131, while other lineages remain fairly susceptible and rarely take up plasmids (8,204). It is
therefore very likely that antibiotic selection pressure creates this clonal-like population structure
while antibiotic free environments allows for competition and subsequent non-related population

Copenhagen 2014
63
structure (138,181). As an example, ST393 has become more frequent as its resistance profile
increases, with no change in virulence and we still see common non-ESBL and non-MDR E.coli
lineages on a global scale as we also detected a clear difference in dominating lineages (7,130). The
lineage ST69 has been found as the dominant faecal E. coli in healthy individuals. Thus, ExPEC are
well adapted to the gut of humans (11,104). As the abundance of faecal ESBL-producing E.coli is a
predictor of infection and antibiotic exposure increases ESBL-producing E.coli faecal count,
constraining use of antibiotics might maintain ESBL count at low levels, hereby minimizing the
probability of infection (93). However, ST131 must be a specialized UPEC, present in all
populations, capable of surviving in the community once present. Thus, ST131 could be one of the
UPEC lineages which are able to effectively colonize the human gut, causing UTI simply since they
dominate the microflora after antibiotic exposure (11). We speculate if resistance in E.coli is
defined by intrinsic differences in different E. coli lineages, allowing for a limited number of UPEC
ST-lineages to obtain and carry blaCTX-M.
The identified distribution of MDR E.coli indicates the existence of reservoirs for ESBL-producing
E.coli. It does, likewise, raise the question if plasmids carrying ESBL genes or not that easily transferred
between E.coli strains, why plasmids require certain E.coli lineages combined with selection pressure to
effectively spread. The likely existence of reservoirs of ESBL-producing E.coli creates a need for reliable,
cheap and fast characterization of pathogenic E.coli, to identify these reservoirs and lessen subsequent
distribution (203). Nonetheless, since not all ESBL-producing E.coli belong to known and identical
lineages, and as plasmids can be acquired by horizontal gene exchange, restraining ST131 and lineages
alike will not completely stop the spread of ESBL-producing E. coli (115). Virulence factors do not
necessarily influence abundance of faecal ESBL-producing E.coli and faecal dominance has previously
been found to be more important than virulence in the pathogenesis of UTI (93). It is therefore likely that,
even with the VFs conferring transmissibility and colonization of global lineages, other less resistant but
“classic” ExPEC as well as commensal E.coli could be adequate colonizing competitors, without
antibiotic selection pressure (115,138).
In future studies, we must identify external reservoirs of ESBL-producing E.coli, which could be nursing
homes, long-term care facilities and hospitals as previously stated, but impossible for us to determine
(205). However, a Swedish study did not find increased level of resistance in nursing homes from 2003-
12 and only very few cases of ESBL-producing E.coli with recent antibiotic treatment and hospitalisation

PhD Dissertation
during the last six months predicted higher resistance rates (206). Therefore, we must understand how
patients acquire MDR E.coli, how antibiotics affect the intestinal flora and creates a possible reservoir,
how long they carry MDR E.coli and finally discover if ESBL-producing E.coli are more likely to cause
disease than susceptible E.coli. ESBL-producing E.coli might simply be the pathogens found since they
dominate the flora. Such studies would require survey of numerous individuals of different ages and with
no history of ESBL carriages. However, carrier studies could be performed as mice colonisation
experiments with treatment given for more than three days and more than once a day. Such studies could
be prolonged to see impact on time of carriage or mice could be re-treated as CFU counts drop to detect
non-countable presence of resistant E.coli. The mouse intestinal colonisation model could furthermore be
used with different resistant UPEC or the level of conjugation between STs in the mouse gut could be
investigated. Finally, it would be of great interest to see the impact of antibiotic on the microflora,
performed as studies of the microbiome of mice, before and after antibiotic treatment. Here shifts in
domination phyla, anaerobic and aerobic and Gram-positive and –negative would be interesting to
investigate. The combination of such mice studies would help to clarify features involved in colonisation
and the dissemination of ESBL-producing E.coli.

Copenhagen 2014
65
8. References
1. Ferry S a., Holm SE, Stenlund H, Lundholm R, Monsen TJ. The natural course of uncomplicated lower urinary tract infection in women illustrated by a randomized placebo controlled study. Scand J Infect Dis. 2004 Jan;36(4):296–301.
2. Lipsky B a. Prostatitis and urinary tract infection in men: what’s new; what's true? Am J Med. 1999 Mar;106(3):327–34.
3. Foxman B. The epidemiology of urinary tract infection. Nat Rev Urol. Nature Publishing Group; 2010 Dec;7(12):653–60.
4. Kahlmeter G, Poulsen HO. Antimicrobial susceptibility of Escherichia coli from community-acquired urinary tract infections in Europe: the ECO·SENS study revisited. Int J Antimicrob Agents. Elsevier B.V.; 2012 Jan;39(1):45–51.
5. Foxman B. Urinary tract infection syndromes: occurrence, recurrence, bacteriology, risk factors, and disease burden. Infect Dis Clin North Am. Elsevier Inc; 2014 Mar;28(1):1–13.
6. Rice LB. The clinical consequences of antimicrobial resistance. Curr Opin Microbiol. 2009 Oct [cited 2011 Jul 13];12(5):476–81.
7. Riley LW. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin Microbiol Infect. 2014 Apr 26;
8. Adams-Sapper S, Diep BA, Perdreau-Remington F, Riley LW. Clonal composition and community clustering of drug-susceptible and -resistant Escherichia coli isolates from bloodstream infections. Antimicrob Agents Chemother. 2013 Jan;57(1):490–7.
9. Banerjee R, Johnston B, Lohse C, Chattopadhyay S, Tchesnokova V, Sokurenko E V, et al. The clonal distribution and diversity of extraintestinal Escherichia coli varies according to patient characteristics. Antimicrob Agents Chemother. 2013 Sep 16;(September):1–29.
10. Pitout JDD. Extraintestinal Pathogenic Escherichia coli: A Combination of Virulence with Antibiotic Resistance. Front Microbiol. 2012 Jan;3(January):9.
11. Nielsen KL, Dynesen P, Larsen P, Frimodt-Møller N. Faecal Escherichia coli from patients with E. coli urinary tract infection and healthy controls who have never had a urinary tract infection. J Med Microbiol. 2014 Apr;63(Pt 4):582–9.
12. Foxman B, Goldberg D. Why the human microbiome project should motivate epidemiologists to learn ecology. Epidemiology. 2010;21(6):757–9.
13. Ejernæs K, Stegger M, Reisner A, Ferry S, Monsen T, Holm SE, et al. Characteristics of Escherichia coli causing persistence or relapse of urinary tract infections. 2011;2(6):528–37.
14. Drekonja DM, Johnson JR. Urinary tract infections. Prim Care. 2008 Jun;35(2):345–67, vii.

PhD Dissertation
15. Doi Y, Park YS, Rivera JI, Adams-Haduch JM, Hingwe A, Sordillo EM, et al. Community-associated extended-spectrum β-lactamase-producing Escherichia coli infection in the United States. Clin Infect Dis. 2013 Mar;56(5):641–8.
16. Banerjee R, Johnston B, Lohse C, Stephen B, Clabots C, Johnson JR, et al. Escherichia coli Sequence Type 131 Is a Dominant, Antimicrobial- Resistant Clonal Group Associated with Healthcare. 2013;34(4):361–9.
17. Rogers B a, Ingram PR, Runnegar N, Pitman MC, Freeman JT, Athan E, et al. Community-Onset Escherichia coli Infection Resistant to Expanded-Spectrum Cephalosporins in Low-Prevalence Countries. Antimicrob Agents Chemother. 2014 Apr;58(4):2126–34.
18. Søraaas A, Sundsfjord AS, Sandven I, Brunborg C, Jenum PP a, Søraas A. Risk Factors for Community-Acquired Urinary Tract Infections Caused by ESBL-Producing Enterobacteriaceae – A Case – Control Study in a Low Prevalence Country. PLoS One. PLoS One; 2013 Jan;8(7):1–7.
19. Nicolas-Chanoine M-H, Jarlier V, Robert J, Arlet G, Drieux L, Leflon-Guibout V, et al. Patient’s origin and lifestyle associated with CTX-M-producing Escherichia coli: a case-control-control study. PLoS One. 2012 Jan;7(1):e30498.
20. Dobrindt U, Hacker JH. Current Topics in Microbiology and Immunology Between Pathogenicity and Commensalism.
21. Dobrindt U. (Patho-)Genomics of Escherichia coli. Int J Med Microbiol. 2005 Oct;295(6-7):357–71.
22. Pitout JDD. Extraintestinal pathogenic Escherichia coli: an update on antimicrobial resistance, laboratory diagnosis and treatment. Expert Rev Anti Infect Ther. 2012 Oct;10(10):1165–76.
23. Russo T a, Johnson JR. Proposal for a new inclusive designation for extraintestinal pathogenic isolates of Escherichia coli: ExPEC. J Infect Dis. 2000 May;181(5):1753–4.
24. Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004 Feb ;2(2):123–40.
25. Johnson JR, Russo T a. Extraintestinal pathogenic Escherichia coli: “The other bad E coli.”J Lab Clin Med. 2002 Mar;139(3):155–62.
26. Bailey JK, Pinyon JL, Anantham S, Hall RM. Distribution of human commensal Escherichia coli phylogenetic groups. J Clin Microbiol. 2010 Sep;48(9):3455–6.
27. Johnson JR, Russo T a. Uropathogenic Escherichia coli as agents of diverse non-urinary tract extraintestinal infections. J Infect Dis. 2002 Sep 15;186(6):859–64.
28. Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl Environ Microbiol. 2000 Oct;66(10):4555–8.

Copenhagen 2014
67
29. Clermont O, Christenson JK, Denamur E, Gordon DM. The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ Microbiol Rep. 2013 Feb;5(1):58–65.
30. Bleibtreu A, Gros P-A, Laouénan C, Clermont O, Le Nagard H, Picard B, et al. Fitness, stress resistance, and extraintestinal virulence in Escherichia coli. Infect Immun. 2013 Aug ;81(8):2733–42.
31. Barroso-Batista J, Sousa A, Lourenço M, Bergman M-L, Sobral D, Demengeot J, et al. The first steps of adaptation of Escherichia coli to the gut are dominated by soft sweeps. PLoS Genet. 2014 Mar;10(3):e1004182.
32. Croxall G, Hale J, Weston V, Manning G, Cheetham P, Achtman M, et al. Molecular epidemiology of extraintestinal pathogenic Escherichia coli isolates from a regional cohort of elderly patients highlights the prevalence of ST131 strains with increased antimicrobial resistance in both community and hospital care settings. 2011;(August):2501–8.
33. Skippen I, Shemko M, Turton J, Kaufmann ME. Epidemiology of infections caused by Escherichia coli and Klebsiella spp .: a nested caseecontrol study from a tertiary hospital in London. Infection. 2006;
34. Ho J, Tambyah P a, Paterson DL. Multiresistant Gram-negative infections: a global perspective. Curr Opin Infect Dis. 2010 Dec;23(6):546–53.
35. Mcdermott PF, Walker RD, White DG. Antimicrobials�: Modes of Action and Mechanisms of Resistance. 2003;135–43.
36. Sommer MO a, Dantas G, Church GM. Functional characterization of the antibiotic resistance reservoir in the human microflora. Science. 2009 Aug 28;325(5944):1128–31.
37. Schjørring S, Struve C, Krogfelt K a. Transfer of antimicrobial resistance plasmids from Klebsiella pneumoniae to Escherichia coli in the mouse intestine. J Antimicrob Chemother. 2008 Nov;62(5):1086–93.
38. Alanis AJ. Resistance to antibiotics: are we in the post-antibiotic era? Arch Med Res. 2005 ;36(6):697–705.
39. Wright GD. Antibiotic resistance in the environment: a link to the clinic? Curr Opin Microbiol. Elsevier Ltd; 2010 Oct;13(5):589–94.
40. Llarrull LI, Testero S a, Fisher JF, Mobashery S. The future of the β-lactams. Curr Opin Microbiol. Elsevier Ltd; 2010 Oct;13(5):551–7.
41. ECDC. Antimicrobial resistance surveillance in Europe 2012. 2012.
42. Falagas ME, Karageorgopoulos DE. Extended-spectrum beta-lactamase-producing organisms. J Hosp Infect. Elsevier Ltd; 2009 Dec;73(4):345–54.

PhD Dissertation
43. Donskey CJ. Antibiotic regimens and intestinal colonization with antibiotic-resistant gram-negative bacilli. Clin Infect Dis . 2006 Sep 1;43 Suppl 2(Suppl 2):S62–9.
44. Oteo J, Navarro C, Cercenado E, Delgado-Iribarren A, Wilhelmi I, Orden B, et al. Spread of Escherichia coli strains with high-level cefotaxime and ceftazidime resistance between the community, long-term care facilities, and hospital institutions. J Clin Microbiol . 2006 Jul ;44(7):2359–66.
45. Bush K. Alarming β-lactamase-mediated resistance in multidrug-resistant Enterobacteriaceae. Curr Opin Microbiol . Elsevier Ltd; 2010 Oct;13(5):558–64.
46. Novais A, Comas I, Baquero F, Cantón R, Coque TM, Moya A, et al. Evolutionary trajectories of beta-lactamase CTX-M-1 cluster enzymes: predicting antibiotic resistance. PLoS Pathog . 2010 Jan;6(1):e1000735.
47. Peirano G, Pitout JDD. Molecular epidemiology of Escherichia coli producing CTX-M beta-lactamases: the worldwide emergence of clone ST131 O25:H4. Int J Antimicrob Agents . 2010 Apr;35(4):316–21.
48. Yao JD, Moellering R. Manual of Clinical Microbiology. 8th. ed. Murray PR, Baron EJ, Jorgensen J, Pfaller MA, Yolken RH, editors. 2003.
49. Cantón R, González-Alba JM, Galán JC. CTX-M Enzymes: Origin and Diffusion. Front Microbiol . 2012 Jan;3(April):110.
50. Bush K, Jacoby G a. Updated functional classification of beta-lactamases. Antimicrob Agents Chemother . 2010 Mar;54(3):969–76.
51. Paterson DL, Bonomo RA. Extended-Spectrum Beta -Lactamases�: a Clinical Update. Society. 2005;18(4):657–86.
52. Rogers B a, Sidjabat HE, Paterson DL. Escherichia coli O25b-ST131: a pandemic, multiresistant, community-associated strain. J Antimicrob Chemother. 2011 Jan;66(1):1–14.
53. Pitout JDD, Laupland KB. Enterobacteriaceae�: an emerging public-health concern. 2008;8(March):159–66.
54. Naseer U, Sundsfjord A. The CTX-M conundrum: dissemination of plasmids and Escherichia coli clones. Microb Drug Resist . 2011 Mar;17(1):83–97.
55. Hawkey PM, Jones AM. The changing epidemiology of resistance. J Antimicrob Chemother . 2009 Sep;64 Suppl 1:i3–10.
56. Tian SF, Chen BY, Chu YZ, Wang S. Prevalence of rectal carriage of extended-spectrum b -lactamase-producing Escherichia coli among elderly people in community settings in China. 2008;785:781–5.

Copenhagen 2014
69
57. Nielsen JB, Albayati A, Jørgensen RL, Hansen KH, Lundgren B, Schønning K. An abbreviated MLVA identifies Escherichia coli ST131 as the major extended-spectrum β-lactamase-producing lineage in the Copenhagen area. Eur J Clin Microbiol Infect Dis . 2013 Nov 6;32(3):431–6.
58. Kerrn MB, Klemmensen T, Espersen F. Susceptibility of Danish Escherichia coli strains isolated from urinary tract infections and bacteraemia , and distribution of sul genes conferring sulphonamide resistance. 2002;513–6.
59. Kjerulf A, Hansen DS, Sandvang D, Hansen F, Frimodt-Møller N. The prevalence of ESBL-producing E. coli and Klebsiella strains in the Copenhagen area of Denmark. APMIS . 2008 Feb;116(2):118–24.
60. Hansen DS, Schumacher H, Hansen F, Stegger M, Hertz FB, Schønning K, et al. Extended-spectrum β-lactamase (ESBL) in Danish clinical isolates of Escherichia coli and Klebsiella pneumoniae: prevalence, β-lactamase distribution, phylogroups, and co-resistance. Scand J Infect Dis . 2012 Mar;44(3):174–81.
61. DANMAP 2012. DANMAP. 2012;
62. Jakobsen L, Kurbasic A, Skjøt-rasmussen L, Ejrnæs K, Porsbo LJ, Pedersen K, et al. Escherichia coli Isolates from Broiler Chicken Meat , Broiler Chickens , Pork , and Pigs Share Phylogroups and Antimicrobial Resistance with Community-Dwelling. 2010;7(5).
63. Tenover FC. Mechanisms of antimicrobial resistance in bacteria. Am J Infect Control . 2006 Jun;34(5 Suppl 1):S3–10; discussion S64–73.
64. Russo T a, Johnson JR. Medical and economic impact of extraintestinal infections due to Escherichia coli: focus on an increasingly important endemic problem. Microbes Infect . 2003 Apr;5(5):449–56.
65. Jakobsen L, Garneau P, Bruant G, Harel J, Olsen SS, Porsbo LJ, et al. Is Escherichia coli urinary tract infection a zoonosis? Proof of direct link with production animals and meat. Eur J Clin Microbiol Infect Dis . 2012 Jun;31(6):1121–9.
66. Bush K, Courvalin P, Dantas G, Davies J, Eisenstein B, Huovinen P, et al. Tackling antibiotic resistance. Nat Rev Microbiol . Nature Publishing Group; 2011 Dec;9(12):894–6.
67. Vardakas KZ, Rafailidis PI, Konstantelias A a, Falagas ME. Predictors of mortality in patients with infections due to multi-drug resistant Gram negative bacteria: the study, the patient, the bug or the drug? J Infect . Elsevier Ltd; 2013 May;66(5):401–14.
68. Kang C-I, Wi YM, Ko KS, Chung DR, Peck KR, Lee NY, et al. Outcomes and risk factors for mortality in community-onset bacteremia caused by extended-spectrum beta-lactamase-producing Escherichia coli, with a special emphasis on antimicrobial therapy. Scand J Infect Dis . 2013 Mar 19;(November 2012):1–7.

PhD Dissertation
69. Rodríguez-Baño J, Picón E, Gijón P, Hernández JR, Cisneros JM, Peña C, et al. Risk factors and prognosis of nosocomial bloodstream infections caused by extended-spectrum-beta-lactamase-producing Escherichia coli. J Clin Microbiol . 2010 May;48(5):1726–31.
70. Hayakawa K, Gattu S, Marchaim D, Bhargava A, Palla M, Alshabani K, et al. Epidemiology and Risk Factors for Isolation of Escherichia coli Producing CTX-M-Type Extended-Spectrum β-Lactamase in a Large U.S. Medical Center. Antimicrob Agents Chemother . 2013 Aug;57(8):4010–8.
71. Vardakas KZ, Tansarli GS, Rafailidis PI, Falagas ME. Carbapenems versus alternative antibiotics for the treatment of bacteraemia due to Enterobacteriaceae producing extended-spectrum β-lactamases: a systematic review and meta-analysis. J Antimicrob Chemother . 2012 Aug 21;(August):2793–803.
72. Marchaim D, Gottesman T, Schwartz O, Korem M, Maor Y, Rahav G, et al. National multicenter study of predictors and outcomes of bacteremia upon hospital admission caused by Enterobacteriaceae producing extended-spectrum beta-lactamases. Antimicrob Agents Chemother . 2010 Dec;54(12):5099–104.
73. Apisarnthanarak A, Kiratisin P, Saifon P, Kitphati R, Dejsirilert S, Mundy LM. Clinical and molecular epidemiology of community-onset, extended-spectrum beta-lactamase-producing Escherichia coli infections in Thailand: a case-case-control study. Am J Infect Control . 2007 Nov;35(9):606–12.
74. Hsieh C, Shen Y, Hwang K. Clinical Implications , Risk Factors and Mortality Following Community-onset Bacteremia Caused by Extended-spectrum b-lactamase ( ESBL ) and non-ESBL Producing Escherichia coli. J Microbiol Immunol Infect . Taiwan Society of Microbiology; 2010;43(3):240–8.
75. Kanafani ZA, Mehio-sibai A, Araj GF, Kanaan M, Kanj SS. Epidemiology and risk factors for producing organisms�: A case control study at a tertiary care center in. Control. 2005;21–3.
76. Kang C, Song J-H, Chung DR, Peck KR, Ko KS, Yeom J, et al. Risk factors and treatment outcomes of community-onset bacteraemia caused by extended-spectrum beta -lactamase-producing Escherichia coli. Int J Antimicrob Agents . 2010 Sep;36(3):284–7.
77. Lautenbach E, Patel JB, Bilker WB, Edelstein PH, Fishman NO. Extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae: risk factors for infection and impact of resistance on outcomes. Clin Infect Dis . 2001 Apr 15;32(8):1162–71.
78. Melzer M, Petersen I. Mortality following bacteraemic infection caused by extended spectrum beta-lactamase (ESBL) producing E. coli compared to non-ESBL producing E. coli. J Infect . 2007 Sep [cited 2012 Feb 8];55(3):254–9.
79. Tumbarello M, Spanu T, Di Bidino R, Marchetti M, Ruggeri M, Trecarichi EM, et al. Costs of bloodstream infections caused by Escherichia coli and influence of extended-spectrum-

Copenhagen 2014
71
beta-lactamase production and inadequate initial antibiotic therapy. Antimicrob Agents Chemother . 2010 Oct;54(10):4085–91.
80. Tosh PK, McDonald LC. Infection control in the multidrug-resistant era: tending the human microbiome. Clin Infect Dis . 2012 Mar 1;54(5):707–13.
81. Dantas G, Sommer MO a, Degnan PH, Goodman AL. Experimental approaches for defining functional roles of microbes in the human gut. Annu Rev Microbiol . 2013 Sep 8;67:459–75.
82. Leatham MP, Banerjee S, Autieri SM, Mercado-Lubo R, Conway T, Cohen PS. Precolonized human commensal Escherichia coli strains serve as a barrier to E. coli O157:H7 growth in the streptomycin-treated mouse intestine. Infect Immun . 2009 Jul;77(7):2876–86.
83. Edlund C, Nord CE. Effect on the human normal microflora of oral antibiotics for treatment of urinary tract infections. J Antimicrob Chemother . 2000 Sep;46 Suppl 1:41–8; discussion 63–5.
84. Raum E, Lietzau S, von Baum H, Marre R, Brenner H. Changes in Escherichia coli resistance patterns during and after antibiotic therapy: a longitudinal study among outpatients in Germany. Clin Microbiol Infect . 2008 Jan;14(1):41–8.
85. Licht TR, Struve C, Christensen BB, Poulsen RL, Molin S, Krogfelt K a. Evidence of increased spread and establishment of plasmid RP4 in the intestine under sub-inhibitory tetracycline concentrations. FEMS Microbiol Ecol . 2003 May 1;44(2):217–23.
86. Feld L, Schjørring S, Hammer K, Licht TR, Danielsen M, Krogfelt K, et al. Selective pressure affects transfer and establishment of a Lactobacillus plantarum resistance plasmid in the gastrointestinal environment. J Antimicrob Chemother . 2008 Apr;61(4):845–52.
87. Harmoinen J, Mentula S, Heikkila M, Rest M Van Der, Rajala-schultz J, Donskey CJ, et al. Orally Administered Targeted Recombinant Beta-Lactamase Prevents Ampicillin-Induced Selective Pressure on the Gut Microbiota�: a Novel Approach to Reducing Antimicrobial Resistance. Society. 2004;48(1):75–9.
88. Faure S, Perrin-Guyomard a, Delmas JM, Chatre P, Laurentie M. Transfer of plasmid-mediated CTX-M-9 from Salmonella enterica serotype Virchow to Enterobacteriaceae in human flora-associated rats treated with cefixime. Antimicrob Agents Chemother . 2010 Jan;54(1):164–9.
89. Bailey JK, Pinyon JL, Anantham S, Hall RM. Commensal Escherichia coli of healthy humans: a reservoir for antibiotic-resistance determinants. J Med Microbiol . 2010 Nov;59(Pt 11):1331–9.
90. Kristiansson C, Grape M, Gotuzzo E, Samalvides F, Chauca J, Larsson M, et al. Socioeconomic factors and antibiotic use in relation to antimicrobial resistance in the Amazonian area of Peru. Scand J Infect Dis . 2009 Jan;41(4):303–12.

PhD Dissertation
91. Gustafsson I, Sjölund M, Torell E, Johannesson M, Engstrand L, Cars O, et al. Bacteria with increased mutation frequency and antibiotic resistance are enriched in the commensal flora of patients with high antibiotic usage. J Antimicrob Chemother . 2003 Oct;52(4):645–50.
92. Olofsson SK, Marcusson LL, Strömbäck A, Hughes D, Cars O. Dose-related selection of fluoroquinolone-resistant Escherichia coli 2007.pdf. J Antimicrob Chemother . 2007 Oct ;60(4):795–801.
93. Ruppé E, Lixandru B, Cojocaru R, Büke C, Paramythiotou E, Angebault C, et al. Relative Fecal Abundance of Extended-Spectrum-β-Lactamase-Producing Escherichia coli Strains and Their Occurrence in Urinary Tract Infections in Women. Antimicrob Agents Chemother . 2013 Sep;57(9):4512–7.
94. Kluytmans J a JW, Overdevest ITM a, Willemsen I, Kluytmans-van den Bergh MFQ, van der Zwaluw K, Heck M, et al. Extended-spectrum β-lactamase-producing Escherichia coli from retail chicken meat and humans: comparison of strains, plasmids, resistance genes, and virulence factors. Clin Infect Dis . 2013 Feb;56(4):478–87.
95. Nataro JP, Kaper JB. Diarrheagenic Escherichia coli. Clin Microbiol Rev . 1998 Jan;11(1):142–201.
96. Chapman PA. Sources of Escherichia coli 0157 and experiences over the past 15 years in Sheffield , UK. 2000;
97. Vonberg RP, Höhle M, Aepfelbacher M, Bange FC, Belmar Campos C, Claussen K, et al. Duration of Fecal Shedding of Shiga Toxin-Producing Escherichia coli O104:H4 in Patients Infected During the 2011 Outbreak in Germany: A Multicenter Study. Clin Infect Dis . 2013 Apr;56(8):1132–40.
98. Hammerum AM, Larsen J, Andersen VD, Lester CH, Skovgaard Skytte TS, Hansen F, et al. Characterization of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli obtained from Danish pigs, pig farmers and their families from farms with high or no consumption of third- or fourth-generation cephalosporins. J Antimicrob Chemother . 2014 Jun 7;1–8.
99. Hammerum a M, Lester CH, Jakobsen L, Porsbo LJ. Faecal carriage of extended-spectrum β-lactamase-producing and AmpC β-lactamase-producing bacteria among Danish army recruits. . Clinical microbiology and infection�: the official publication of the European Society of Clinical Microbiology and Infectious Diseases. 2011. p. 566–8.
100. Jørgensen SB, Samuelsen O, Sundsfjord A, Bhatti SA, Jørgensen I, Sivapathasundaram T, et al. High prevalence of faecal carriage of ESBL-producing Enterobacteriaceae in Norwegian patients with gastroenteritis. Scand J Infect Dis . 2014 Apr 4;(1478):1–4.
101. Ostholm-Balkhed A, Tärnberg M, Nilsson M, Nilsson LE, Hanberger H, Hällgren A. Travel-associated faecal colonization with ESBL-producing Enterobacteriaceae: incidence and risk factors. J Antimicrob Chemother . 2013 Sep [cited 2014 Jul 7];68(9):2144–53.

Copenhagen 2014
73
102. Wickramasinghe NH, Xu L, Eustace A, Shabir S, Saluja T, Hawkey PM. High community faecal carriage rates of CTX-M ESBL-producing Escherichia coli in a specific population group in Birmingham, UK. J Antimicrob Chemother . 2012 Mar 8;1–6.
103. Luvsansharav U-O, Hirai I, Nakata A, Imura K, Yamauchi K, Niki M, et al. Prevalence of and risk factors associated with faecal carriage of CTX-M β-lactamase-producing Enterobacteriaceae in rural Thai communities. . The Journal of antimicrobial chemotherapy. 2012. p. 1769–74.
104. Nicolas-Chanoine M-H, Gruson C, Bialek-Davenet S, Bertrand X, Thomas-Jean F, Bert F, et al. 10-Fold increase (2006-11) in the rate of healthy subjects with extended-spectrum β-lactamase-producing Escherichia coli faecal carriage in a Parisian check-up centre. J Antimicrob Chemother . 2013 Mar;68(3):562–8.
105. Lausch KR, Fuursted K, Larsen CS, Storgaard M. Colonisation with multi-resistant Enterobacteriaceae in hospitalised Danish patients with a history of recent travel: a cross-sectional study. Travel Med Infect Dis . Elsevier Ltd; 2013;11(5):320–3.
106. Tham J, Odenholt I, Walder M, Brolund A, Ahl J, Melander E. Extended-spectrum beta-lactamase-producing Escherichia coli in patients with travellers’ diarrhoea. Scand J Infect Dis . 2010 Apr;42(4):275–80.
107. Peirano G, Laupland KB, Gregson DB, Pitout JDD. Colonization of Returning Travelers With CTX-M-Producing Escherichia coli. J Travel Med . 2011 Sep;18(5):299–303.
108. Tham J, Walder M, Melander EVA, Odenholt I. Duration of colonization with extended-spectrum beta-lactamase-producing Escherichia coli in patients with travellers’ diarrhoea. Scand J Infect Dis . 2012 Jan 31;46(December 2011):1–5.
109. Rooney PJ, O’Leary MC, Loughrey AC, McCalmont M, Smyth B, Donaghy P, et al. Nursing homes as a reservoir of extended-spectrum beta-lactamase (ESBL)-producing ciprofloxacin-resistant Escherichia coli. J Antimicrob Chemother . 2009 Sep;64(3):635–41.
110. Colomer-Lluch M, Mora A, López C, Mamani R, Dahbi G, Marzoa J, et al. Detection of quinolone-resistant Escherichia coli isolates belonging to clonal groups O25b:H4-B2-ST131 and O25b:H4-D-ST69 in raw sewage and river water in Barcelona, Spain. J Antimicrob Chemother . 2013 Apr;68(4):758–65.
111. Clermont O, Olier M, Hoede C, Diancourt L, Brisse S, Keroudean M, et al. Animal and human pathogenic Escherichia coli strains share common genetic backgrounds. Infect Genet Evol . 2011 Apr;11(3):654–62.
112. Johnson JR, Delavari P, Stell a L, Whittam TS, Carlino U, Russo T a. Molecular comparison of extraintestinal Escherichia coli isolates of the same electrophoretic lineages from humans and domestic animals. J Infect Dis . 2001 Jan 1;183(1):154–9.
113. Jakobsen L, Hammerum AM, Frimodt-Møller N. Detection of clonal group A Escherichia coli isolates from broiler chickens, broiler chicken meat, community-dwelling humans, and

PhD Dissertation
urinary tract infection (UTI) patients and their virulence in a mouse UTI model. Appl Environ Microbiol . 2010 Dec;76(24):8281–4.
114. Leverstein-van Hall M a, Dierikx CM, Cohen Stuart J, Voets GM, van den Munckhof MP, van Essen-Zandbergen a, et al. Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin Microbiol Infect . 2011 Jun;17(6):873–80.
115. Foxman B. Extended-spectrum β-lactamase-producing escherichia coli in the united states: time to rethink empirical treatment for suspected E. coli infections? Clin Infect Dis . 2013 Mar;56(5):649–51.
116. Krych L, Hansen CHF, Hansen AK, van den Berg FWJ, Nielsen DS. Quantitatively Different, yet Qualitatively Alike: A Meta-Analysis of the Mouse Core Gut Microbiome with a View towards the Human Gut Microbiome. PLoS One . 2013 Jan;8(5):e62578.
117. Knudsen JD, Frimodt-møller N. Animal Models in Bacteriology. 2001;9:1–14.
118. Jakobsen L, Cattoir V, Jensen KS, Hammerum AM, Nordmann P, Frimodt-Møller N. Impact of low-level fluoroquinolone resistance genes qnrA1, qnrB19 and qnrS1 on ciprofloxacin treatment of isogenic Escherichia coli strains in a murine urinary tract infection model. J Antimicrob Chemother . 2012 Oct;67(10):2438–44.
119. Leatham MP, Stevenson SJ, Gauger EJ, Krogfelt KA, Lins JJ, Haddock TL, et al. Mouse Intestine Selects Nonmotile flhDC Mutants of Escherichia coli MG1655 with Increased Colonizing Ability and Better Utilization of Carbon Sources. 2005;73(12):8039–49.
120. Donskey CJ, Helfand MS, Pultz NJ, Rice LB. Effect of Parenteral Fluoroquinolone Administration on Persistence of Vancomycin-Resistant Enterococcus faecium in the Mouse Gastrointestinal Tract. Society. 2004;48(1):326–8.
121. Hoyen CK, Pultz NJ, Paterson DL, Aron DC, Donskey CJ. Effect of Parenteral Antibiotic Administration on Establishment of Intestinal Colonization in Mice by Klebsiella pneumoniae Strains Producing Extended-Spectrum Beta-Lactamases. Society. 2003;47(11):3610–2.
122. Pultz MJ, Donskey CJ. Effects of imipenem-cilastatin, ertapenem, piperacillin-tazobactam, and ceftriaxone treatments on persistence of intestinal colonization by extended-spectrum-beta-lactamase-producing Klebsiella pneumoniae strains in mice. Antimicrob Agents Chemother . 2007 Aug;51(8):3044–5.
123. Perez F, Pultz MJ, Endimiani A, Bonomo R a, Donskey CJ. Effect of antibiotic treatment on establishment and elimination of intestinal colonization by KPC-producing Klebsiella pneumoniae in mice. Antimicrob Agents Chemother . 2011 Jun;55(6):2585–9.
124. Møller AK, Leatham MP, Conway T, Nuijten PJM, Haan LAM De, Krogfelt KA, et al. An Escherichia coli MG1655 Lipopolysaccharide Deep-Rough Core Mutant Grows and Survives in Mouse Cecal Mucus but Fails To Colonize the Mouse Large Intestine. 2003;71(4):2142–52.

Copenhagen 2014
75
125. Flückiger U, Segessenmann C, Gerber a U. Integration of pharmacokinetics and pharmacodynamics of imipenem in a human-adapted mouse model. Antimicrob Agents Chemother . 1991 Sep;35(9):1905–10.
126. Stiefel U, Pultz NJ, Donskey CJ. Effect of carbapenem administration on establishment of intestinal colonization by vancomycin-resistant enterococci and Klebsiella pneumoniae in mice. Antimicrob Agents Chemother . 2007 Jan;51(1):372–5.
127. Oteo J, Diestra K, Juan C, Bautista V, Novais A, Pérez-Vázquez M, et al. Extended-spectrum beta-lactamase-producing Escherichia coli in Spain belong to a large variety of multilocus sequence typing types, including ST10 complex/A, ST23 complex/A and ST131/B2. Int J Antimicrob Agents . 2009 Aug;34(2):173–6 .
128. Hernández J, Martínez-Martínez L, Cantón R, Coque T, Pascual A. Nationswide Study of Escherichia Coli and Klebsiella pneumoniae Producing Extended-Spectrum beta -Lactamases in Spain. 2005;49(5):2122–5.
129. Colpan A, Johnston B, Porter S, Clabots C, Anway R, Thao L, et al. Escherichia coli Sequence Type 131 (ST131) Sub-clone H30 as an Emergent Multidrug-Resistant Pathogen Among US Veterans. Clin Infect Dis . 2013 Aug 29;131(9):1–10.
130. Olesen B, Scheutz F, Menard M, Skov MN, Kolmos HJ, Kuskowski M a, et al. Three-decade epidemiological analysis of Escherichia coli O15:K52:H1. J Clin Microbiol . 2009 Jun;47(6):1857–62.
131. Johnson JR, Tchesnokova V, Johnston B, Clabots C, Roberts PL, Billig M, et al. Abrupt emergence of a single dominant multidrug-resistant strain of Escherichia coli. J Infect Dis . 2013 Mar 15;207(6):919–28.
132. Olesen B, Hansen DS, Nilsson F, Frimodt-Møller J, Leihof RF, Struve C, et al. Prevalence and characteristics of the epidemic multiresistant Escherichia coli ST131 clonal group among extended-spectrum beta-lactamase-producing E. coli isolates in Copenhagen, Denmark. J Clin Microbiol . 2013 Jun;51(6):1779–85.
133. Johnson JR, Tchesnokova V, Johnston B, Clabots C, Roberts PL, Billig M, et al. Abrupt emergence of a single dominant multidrug-resistant strain of Escherichia coli. J Infect Dis . 2013 Mar 15;207(6):919–28.
134. Mora A, Blanco M, López C, Mamani R, Blanco JE, Alonso MP, et al. Emergence of clonal groups O1:HNM-D-ST59, O15:H1-D-ST393, O20:H34/HNM-D-ST354, O25b:H4-B2-ST131 and ONT:H21,42-B1-ST101 among CTX-M-14-producing Escherichia coli clinical isolates in Galicia, northwest Spain. Int J Antimicrob Agents . 2011 Jan;37(1):16–21.
135. Gibreel TM, Dodgson AR, Cheesbrough J, Fox AJ, Bolton FJ, Upton M. Population structure, virulence potential and antibiotic susceptibility of uropathogenic Escherichia coli from Northwest England. J Antimicrob Chemother . 2012 Feb;67(2):346–56.

PhD Dissertation
136. Blanco J, Mora A, Mamani R, López C, Blanco M, Dahbi G, et al. National survey of Escherichia coli causing extraintestinal infections reveals the spread of drug-resistant clonal groups O25b:H4-B2-ST131, O15:H1-D-ST393 and CGA-D-ST69 with high virulence gene content in Spain. J Antimicrob Chemother . 2011 Sep;66(9):2011–21.
137. Naseer U, Haldorsen B, Tofteland S, Hegstad K, Scheutz F, Simonsen GS, et al. Molecular characterization of CTX-M-15-producing clinical isolates of Escherichia coli reveals the spread of multidrug-resistant ST131 (O25:H4) and ST964 (O102:H6) strains in Norway. APMIS . 2009 Jul;117(7):526–36.
138. Bengtsson S, Naseer U, Sundsfjord A, Kahlmeter G, Sundqvist M. Sequence types and plasmid carriage of uropathogenic Escherichia coli devoid of phenotypically detectable resistance. J Antimicrob Chemother . 2012 Jan;67(1):69–73.
139. Brisse S, Diancourt L, Laouénan C, Vigan M, Caro V, Arlet G, et al. Phylogenetic distribution of CTX-M- and non-extended-spectrum-β-lactamase-producing Escherichia coli isolates: group B2 isolates, except clone ST131, rarely produce CTX-M enzymes. J Clin Microbiol . 2012 Sep;50(9):2974–81
140. Aibinu I, Odugbemi T, Koenig W, Ghebremedhin B. Sequence type ST131 and ST10 complex (ST617) predominant among CTX-M-15-producing Escherichia coli isolates from Nigeria. Clin Microbiol Infect . 2012 Mar;18(3):E49–51.
141. Gibreel TM, Dodgson AR, Cheesbrough J, Bolton FJ, Fox AJ, Upton M. High metabolic potential may contribute to the success of ST131 uropathogenic Escherichia coli. J Clin Microbiol . 2012 Oct;50(10):3202–7.
142. Peirano G, van der Bij AK, Gregson DB, Pitout JDD. Molecular epidemiology over an 11-year period (2000 to 2010) of extended-spectrum β-lactamase-producing Escherichia coli causing bacteremia in a centralized Canadian region. Journal of clinical microbiology. 2012. p. 294–9.
143. Novais A, Pires J, Ferreira H, Costa L, Montenegro C, Vuotto C, et al. Characterization of globally spread Escherichia coli ST131 isolates (1991 to 2010). Antimicrob Agents Chemother . 2012 Jul;56(7):3973–6.
144. Kudinha T, Johnson JR, Andrew SD, Kong F, Anderson P, Gilbert GL. Distribution of phylogenetic groups, sequence type ST131, and virulence-associated traits among Escherichia coli isolates from men with pyelonephritis or cystitis and healthy controls. Clin Microbiol Infect . 2013 Apr;19(4):E173–80.
145. Johnson JR, Johnston B, Clabots C, Kuskowski M a, Castanheira M. Escherichia coli sequence type ST131 as the major cause of serious multidrug-resistant E. coli infections in the United States. Clin Infect Dis . 2010 Aug 1;51(3):286–94.
146. Blanco J, Mora A, Mamani R, López C, Blanco M, Dahbi G, et al. Four Main Virotypes among Extended-Spectrum β-Lactamase-Producing Isolates of Escherichia coli O25b:H4-

Copenhagen 2014
77
B2-ST131: Bacterial, Epidemiological, and Clinical Characteristics. J Clin Microbiol . 2013 Aug 7;(August):1–35.
147. Adams-Sapper S, Sergeevna-Selezneva J, Tartof S, Raphael E, Diep BA, Perdreau-Remington F, et al. Globally dispersed mobile drug-resistance genes in gram-negative bacterial isolates from patients with bloodstream infections in a US urban general hospital. J Med Microbiol . 2012 Jul;61(Pt 7):968–74.
148. Graffunder EM, Preston KE, Evans AM, Venezia R a. Risk factors associated with extended-spectrum b-lactamase-producing organisms at a tertiary care hospital. J Antimicrob Chemother . 2005 Jul;56(1):139–45.
149. Ben-Ami R, Rodríguez-Baño J, Arslan H, Pitout JDD, Quentin C, Calbo ES, et al. A multinational survey of risk factors for infection with extended-spectrum beta-lactamase-producing enterobacteriaceae in nonhospitalized patients. Clin Infect Dis . 2009 Sep 1;49(5):682–90.
150. Goyal A, Prasad KN, Prasad A, Gupta S, Ghoshal U, Ayyagari A. Extended spectrum β -lactamases in Escherichia coli & Klebsiella pneumoniae & associated risk factors. Indian J Med Res. 2009;(June):695–700.
151. Cassier P, Lallechère S, Aho S, Astruc K, Neuwirth C, Piroth L, et al. Cephalosporin and fluoroquinolone combinations are highly associated with CTX-M β-lactamase-producing Escherichia coli: a case-control study in a French teaching hospital. Clin Microbiol Infect . 2011 Nov;17(11):1746–51.
152. Rodríguez-Baño J, Navarro MD, Ngugro MD. Extended-spectrum beta-lactamases in ambulatory care: a clinical perspective. Clin Microbiol Infect . 2008 Jan;14 Suppl 1:104–10.
153. Wu U-I, Yang C-S, Chen W-C, Chen Y-C, Chang S-C. Risk factors for bloodstream infections due to extended-spectrum beta-lactamase-producing Escherichia coli. J Microbiol Immunol Infect . Taiwan Society of Microbiology; 2010 Aug;43(4):310–6.
154. Rodríguez-Baño J, Alcalá J, Cisneros JM, Grill F, Oliver A, Horcajada JP, et al. Escherichia coli producing SHV-type extended-spectrum beta-lactamase is a significant cause of community-acquired infection. J Antimicrob Chemother . 2009 Apr;63(4):781–4.
155. Banerjee R, Strahilevitz J, Johnson JR, Nagwekar PP, Schora DM, Shevrin I, et al. Predictors and Molecular Epidemiology of Community-Onset Extended-Spectrum β-Lactamase-Producing Escherichia coli Infection in a Midwestern Community. Infect Control Hosp Epidemiol . 2013 Sep;34(9):947–53.
156. Kaye KS, Harris AD, Samore M, Carmeli Y. The Case-Case-Control Study Design: Addressing The Limitations of Risk Factor Studies for antimicrobial resistance. 2005;26(4):346–51.

PhD Dissertation
157. Harris AD, Karchmer TB, Carmeli Y, Samore MH. Methodological principles of case-control studies that analyzed risk factors for antibiotic resistance: a systematic review. Clin Infect Dis . 2001 Apr 1;32(7):1055–61.
158. Wacholder S, Silverman DT, Mclaughlin JK, Mandel JS. Selection of Controls in Case-Control Studies. 1992;135(9):1029–41.
159. Harris AD, Samore MH, Carmeli Y. Control Group Selection Is an Important but Neglected Issue in Studies of Antibiotic Resistance. Ann Intern Med . 2000 Jul 18;133(2):158.
160. Wacholder S, Mclaughlin JK, Silverman DT, Mandel JS. Selection of controls in injury case-control studies. Ann Epidemiol . 2000 Oct 1;10(7):455.
161. Wener KM, Schechner V, Gold HS, Wright SB, Carmeli Y. Treatment with fluoroquinolones or with beta-lactam-beta-lactamase inhibitor combinations is a risk factor for isolation of extended-spectrum-beta-lactamase-producing Klebsiella species in hospitalized patients. Antimicrob Agents Chemother . 2010 May;54(5):2010–6.
162. Søraas A, Sundsfjord AS, Sandven I, Brunborg C, Jenum P. Risk Factors for Community-Acquired Urinary Tract Infections Caused by ESBL-Producing Enterobacteriaceae -A Case-Control Study in a Low Prevalence Country. PLoS One; 2013.
163. Friedman ND, Kaye KS, Stout JE, Mcgarry SA, Trivette SL, Briggs JP, et al. Health Care – Associated Bloodstream Infections in Adults�: A Reason To Change the Accepted Definition of Community-Acquired Infections. 2002;791–8.
164. Kang C-I, Wi YM, Lee MY, Ko KS, Chung DR, Peck KR, et al. Epidemiology and risk factors of community onset infections caused by extended-spectrum β-lactamase-producing Escherichia coli strains. J Clin Microbiol . 2012 Feb;50(2):312–7.
165. Briongos-Figuero LS, Gómez-Traveso T, Bachiller-Luque P, Domínguez-Gil González M, Gómez-Nieto a, Palacios-Martín T, et al. Epidemiology, risk factors and comorbidity for urinary tract infections caused by extended-spectrum beta-lactamase (ESBL)-producing enterobacteria. Int J Clin Pract . 2012 Sep;66(9):891–6.
166. Meier S, Weber R, Zbinden R, Ruef C, Hasse B. Extended-spectrum β-lactamase-producing Gram-negative pathogens in community-acquired urinary tract infections: an increasing challenge for antimicrobial therapy. Infection . 2011 Aug;39(4):333–40.
167. Azap OK, Arslan H, Serefhanoğlu K, Colakoğlu S, Erdoğan H, Timurkaynak F, et al. Risk factors for extended-spectrum beta-lactamase positivity in uropathogenic Escherichia coli isolated from community-acquired urinary tract infections. Clin Microbiol Infect . 2010 Feb;16(2):147–51.
168. Rodríguez-Baño J, Navarro MD, Romero L, Muniain M a, de Cueto M, Ríos MJ, et al. Bacteremia due to extended-spectrum beta -lactamase-producing Escherichia coli in the CTX-M era: a new clinical challenge. Clin Infect Dis . 2006 Dec 1;43(11):1407–14.

Copenhagen 2014
79
169. Quirante OF, Cerrato SG, Pardos SL. Risk factors for bloodstream infections caused by extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. . The Brazilian journal of infectious diseases�: an official publication of the Brazilian Society of Infectious Diseases. p. 370–6.
170. Park SH, Choi S-M, Lee D-G, Kim J, Choi J-H, Kim S-H, et al. Emergence of extended-spectrum β-lactamase-producing escherichia coli as a cause of community-onset bacteremia in South Korea: risk factors and clinical outcomes. Microb Drug Resist . 2011 Dec;17(4):537–44.
171. Rodríguez-Baño J, Picón E, Gijón P, Hernández JR, Ruíz M, Peña C, et al. Community-onset bacteremia due to extended-spectrum beta-lactamase-producing Escherichia coli: risk factors and prognosis. Clin Infect Dis . 2010 Jan 1;50(1):40–8.
172. Park K-H, Oh WS, Kim ES, Park SW, Hur J-A, Kim YK, et al. Factors associated with ciprofloxacin- and cefotaxime-resistant Escherichia coli in women with acute pyelonephritis in the emergency department. Int J Infect Dis . International Society for Infectious Diseases; 2014 Mar 18;
173. Qureshi Z a, Paterson DL, Pakstis DL, Adams-Haduch JM, Sandkovsky G, Sordillo E, et al. Risk factors and outcome of extended-spectrum β-lactamase-producing Enterobacter cloacae bloodstream infections. Int J Antimicrob Agents . Elsevier B.V.; 2011 Jan;37(1):26–32.
174. Bhavnani SM, Ambrose PG, Craig W a, Dudley MN, Jones RN. Outcomes evaluation of patients with ESBL- and non-ESBL-producing Escherichia coli and Klebsiella species as defined by CLSI reference methods: report from the SENTRY Antimicrobial Surveillance Program. Diagn Microbiol Infect Dis . 2006 Mar;54(3):231–6.
175. Han JH, Maslow J, Han X, Xie SX, Tolomeo P, Santana E, et al. Risk Factors for the Development of Gastrointestinal Colonization With Fluoroquinolone-Resistant Escherichia coli in Residents of Long-Term Care Facilities. J Infect Dis . 2013 Sep 17;209:420–5.
176. Fan N-C, Chen H-H, Chen C-L, Ou L-S, Lin T-Y, Tsai M-H, et al. Rise of community-onset urinary tract infection caused by extended-spectrum β-lactamase-producing Escherichia coli in children. J Microbiol Immunol Infect . Elsevier Taiwan LLC; 2013 Jul 5;1–7.
177. Park K-H, Oh WS, Kim ES, Park SW, Hur J-A, Kim YK, et al. Factors associated with ciprofloxacin- and cefotaxime-resistant Escherichia coli in women with acute pyelonephritis in the emergency department. Int J Infect Dis . International Society for Infectious Diseases; 2014 Mar 19;23:8–13.
178. Rodríguez-Baño J, Alcala JC, Cisneros JM, Grill F, Oliver A, Horcajada JP, et al.
Community Infections Caused by Extended-Spectrum -Lactamase–Producing. Arch Intern Med. 2008;168(17):1897–902.
179. Goulenok T, Ferroni a, Bille E, Lécuyer H, Join-Lambert O, Descamps P, et al. Risk factors for developing ESBL E. coli: can clinicians predict infection in patients with prior colonization? J Hosp Infect . 2013 Aug;84(4):294–9.

PhD Dissertation
180. Urwin R, Maiden MCJ. Multi-locus sequence typing: a tool for global epidemiology. Trends Microbiol . 2003 Oct;11(10):479–87.
181. Maiden MCJ. Multilocus sequence typing of bacteria. Annu Rev Microbiol . 2006 Jan;60:561–88.
182. Nielsen KL, Godfrey P a, Stegger M, Andersen PS, Feldgarden M, Frimodt-Møller N. Selection of unique Escherichia coli clones by random amplified polymorphic DNA (RAPD): Evaluation by whole genome sequencing. J Microbiol Methods . Elsevier B.V.; 2014 Jun 6;103C:101–3.
183. Gordon DM, Clermont O, Tolley H, Denamur E. Assigning Escherichia coli strains to phylogenetic groups: multi-locus sequence typing versus the PCR triplex method. Environ Microbiol . 2008 Oct;10(10):2484–96.
184. Maiden MCJ, Bygraves JA, Feil E, Morelli G, Russel JE, Urwin R, et al. Multilocus sequence typing�: A portable approach to the identification of clones within populations of. 1998;95(March):3140–5.
185. Enright MC, Spratt BG. Multilocus sequence typing. Trends Microbiol . 1999 Dec;7(12):482–7.
186. Wirth T, Falush D, Lan R, Colles F, Mensa P, Wieler LH, et al. Sex and virulence in Escherichia coli: an evolutionary perspective. Mol Microbiol . 2006 Jun;60(5):1136–51.
187. Manges AR, Tellis P a, Vincent C, Lifeso K, Geneau G, Reid-Smith RJ, et al. Multi-locus variable number tandem repeat analysis for Escherichia coli causing extraintestinal infections. J Microbiol Methods . Elsevier B.V.; 2009 Nov;79(2):211–3.
188. Keys C, Kemper S, Keim P. Highly diverse variable number tandem repeat loci in the E. coli O157:H7 and O55:H7 genomes for high-resolution molecular typing. J Appl Microbiol . 2005 Jan;98(4):928–40.
189. Pop C, Brenner F, Fields P, Wells J, Strockbine N. Manual of Clinical Microbiology. 8th, volum. Murray PR, Baron EJ, Jorgensen JH, Pfaller MA, Yolken RH, editors. ASM Press; 2003.
190. Köhler C-D, Dobrindt U. What defines extraintestinal pathogenic Escherichia coli? Int J Med Microbiol . 2011 Dec;301(8):642–7.
191. Pedersen CB, Gøtzsche H, Møller JO, Mortensen PB. The Danish Civil Registration System. A cohort of eight million persons. Dan Med Bull . 2006 Nov;53(4):441–9.
192. Pedersen CB. The Danish Civil Registration System. Scand J Public Health . 2011 Jul;39(7 Suppl):22–5.
193. Kildemoes HW, Sørensen HT, Hallas J. The Danish National Prescription Registry. Scand J Public Health . 2011 Jul;39(7 Suppl):38–41.

Copenhagen 2014
81
194. Hansen F, Olsen SS, Heltberg O, Justesen US, Fuglsang-Damgaard D, Knudsen JD, et al. Characterization of Third-Generation Cephalosporin-Resistant Escherichia coli from Bloodstream Infections in Denmark. Microb Drug Resist . 2014 Feb 11;00(00):1–9.
195. Zak O, Sande MA, O’Reilly T. Handbook of Animal Models of Infection. Zak O, Sande MA, editors. Academic Press;
196. Frimodt-moller N, Ostri P, Pedersen IBK, Poulsen SR. Antibiotic Prophylaxis in Pulmonary Surgery - A Double-Blind Study of Penicillin Versus Placebo. 1980;195(4):444–50.
197. Ben-ami R, Arslan H, Pitout JDD, Quentin C, Calbo ES, Azap K, et al. A Multinational Survey of Risk Factors for Infection with Extended-Spectrum b -Lactamase – Producing Enterobacteriaceae in Nonhospitalized Patients. 2009;49.
198. Lautenbach E, Bilker WB, Tolomeo P, Maslow JN. Impact of diversity of colonizing strains on strategies for sampling Escherichia coli from fecal specimens. J Clin Microbiol . 2008 Sep ;46(9):3094–6.
199. Riley LW. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin Microbiol Infect . 2014 May;20(5):380–90.
200. Sullivan a, Edlund C, Nord CE. Effect of antimicrobial agents on the ecological balance of human microflora. Lancet. Elsevier Ltd; 2001 Dec 2;1:101–14.
201. Van der Bij AK, Pitout JDD. The role of international travel in the worldwide spread of multiresistant Enterobacteriaceae. J Antimicrob Chemother . 2012 Sep;67(9):2090–100.
202. Nicolas-Chanoine M-H, Blanco J, Leflon-Guibout V, Demarty R, Alonso MP, Caniça MM, et al. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J Antimicrob Chemother . 2008 Feb;61(2):273–81.
203. Naseer U, Olsson-Liljequist BE, Woodford N, Dhanji H, Cantón R, Sundsfjord A, et al. Multi-locus variable number of tandem repeat analysis for rapid and accurate typing of virulent multidrug resistant Escherichia coli clones. PLoS One . 2012 Jan;7(7):e41232.
204. Bengtsson S, Naseer U, Sundsfjord A, Kahlmeter G, Sundqvist M. Sequence types and plasmid carriage of uropathogenic Escherichia coli devoid of phenotypically detectable resistance. J Antimicrob Chemother . 2012 Jan;67(1):69–73.
205. Rooney PJ, O’Leary MC, Loughrey AC, McCalmont M, Smyth B, Donaghy P, et al. Nursing homes as a reservoir of extended-spectrum beta-lactamase (ESBL)-producing ciprofloxacin-resistant Escherichia coli. J Antimicrob Chemother . 2009 Sep;64(3):635–41.
206. Sundvall P-D, Elm M, Gunnarsson R, Mölstad S, Rodhe N, Jonsson L, et al. Antimicrobial resistance in urinary pathogens among Swedish nursing home residents remains low: a cross-sectional study comparing antimicrobial resistance from 2003 to 2012. BMC Geriatr . 2014 Jan;14:30.

PhD Dissertation
9. Appendix
Appendix 1. Patient demographics. Community-onset infection (COI) and Healthcare associated infections (HCAI) have been divided into age groups and patients have been classified a complicated UTI. These classifications have been done according to previously defined criteria as stated in Manuscript II. Prescr. = Prescriptions.
Cases
ESBL n=96 Resistant n=171 Susceptible n=175 Controls n=200
Demographics n(%) n(%) n(%) n(%)
Age (Mean) 60.5 96(100) 57.95 171(100) 53,25 175(100) 42.10 200(100)
Age>18 (Mean) 65.4 88(92) 58,5 169(99) 56,07 165(94) 46.16 180(90)
Age >65 (Mean) 76.5 48(50) 79.09 85(50) 79.72 66(38) 76.49 37(19)
Male 18 (19)
17(10) 16(9)
54(27)
Female 78(81) 154(90) 159(91) 146(73)
Community-Onset-Infection (COI) 71(74)
139(81) 153(87)
163(82)
Health-Care Associated-Infection (HCAI) 25(26) 32(19) 22(13) 37(19)
COI Age < 18 years 8(100)
1(50) 10(100)
18(90)
HCAI Age < 18 years 0 1(50) 0 2(10)
COI Age 18-65 30(75) 0.5661 77(92) 0.5750 90(91) 0.7870 119(83)
HCAI Age 18-65 10(25) 0.0553 7(8) 0.0624 9(9) 0.3759 24(17)
COI Age >65 years 33(69) 61(72) 53(80)
26(70)
HCAI Age >65 years 15(31) 24(28) 0.4049 13(20) 1 11(30)
Complicated UTI 1(1) 1(0.6) 0
Complicated UTI due to Age > 65 years 44(46) 86(50) 66(38)
Appendix 1 cont. Exposure to antibiotics within one year prior to index sample.
ESBL n=96 Resistant n=171 Susceptible n=175 Controls n=200
Exposure to Antibiotics N (%) Prescr. N (%) Prescr. N (%) Prescr. N (%) Prescr.
Antibiotics in general 85 (89) 480 155 (91) 733 153 (87) 457
131 (66)
Penicillins 25 (26) 40 40 (23) 52 41 (23) 48 42 (21) 59
Trimethoprim 12 (13) 43 22 (13) 93 8 (5) 8 6 (3) 7
Sulphonamides 27 (28) 46 67 (39) 110 50 (29) 78 42 (21) 53
Metronidazole 7 (7) 11 11 (6) 13 8 (5) 8 7 (4) 8
Quinolones 21 (22) 31 22 (23) 36 14 (8) 18 18 (9) 23
Macrolides 13 (14) 29 23 (13) 31 23 (13) 25 27 (14) 43
Mecillinam 74 (77) 149 118 (69) 233 104 (59) 193 60 (30) 97
Nitrofurantoin 19 (20) 53 35 (20) 104 19 (11) 29 11 (6) 18
Tetracycline 5 (5) 8 9 (5) 21 6 (3) 9 6 (3) 9
2nd generations cephalosporins 2 (2) 3 2 (1) 6 0 0 2 (1) 3
Penicillins w. beta-lactamase inhibitors 8 (8) 11 7 (4) 8 5 (3) 5 5 (3) 11
Penicillinase stable penicillins 10 (10) 13 9 (5) 19 6 (3) 6 12 (6) 14
Penicillins with effect on Gram-negative 69 193 187 267 125 228 71 (36) 127

Copenhagen 2014
83
Appendix 2. Results of the Multivariate Logistic Regression Analysis comparing case groups with the uninfected group. We performed the multivariate analysis based on the initial student´s t-test, with all variables with P<0.1. Subsequently we did a simple manual backward step analysis including variables with P<0.1 in the multivariate analysis. The second multivariate analysis was not possible to perform in full for the ESBL group due to lack of goodness of fit. Therefore we have included results from 30 days after sampling. *P-values found by chi-square analysis. Empty squares means estimates not computed by SAS. OD = Odds Ratio.
Multivariate Logistic Regression. First run.
ESBL, n=96 Resistant, n=171 Susceptible, n=175
Variables OR 95 CI P-Value OR 95 CI P-Value OR 95 CI P-Value
Sex (Females) 3.1 1.4 6.6 0.0044 3.7 1.8 7.7 0.0004 3.5 1.807 6.9 0.0002
Age 1.02 1.01 1.04 <.0001 1.0 1.0 1.0 <.0001 1.02 1.010 1.0 <.0001
30 Days Prior
Antibiotics . . . <.0001* . . . <.0001* . . . <.0001*
Mecillinam 2.6 0.9 7.2 0.0713 4.1 1.5 11.1 0.0053 6.09 1.846 20.1 0.0030
Trimethoprim 2.01 0.19 21.1 0.5621 4.1 0.9 19.4 0.0712 1.6 0.186 12.9 0.6850
Kidney Infection Infty Infty Infty 0.0106* 1.1 0.3 4.1 0.8366 . . . 1
Urosepsis Infty 0.99 10.7 0.0334* . . . . . . . .
90 Days Prior
Mecillinam 1.8 0.98 3.3 0.0572 2.4 1.1 5.4 0.0261 2.3 0.910 5.8 0.0781
Nitrofurantoin 1.6 0.5 5.1 0.3871 3.8 1.3 11.4 0.0182 3.2 0.975 10.4 0.0552
Penicillin 2.7 0.8 8.7 0.1040 5.0 1.7 15.1 0.0041 2.6 0.848 8.2 0.0940
Sulphonamides 1.03 0.4 2.5 0.9511 2.9 1.3 6.4 0.0091 2.4 1.078 5.2 0.0318
Trimethoprim 0.99 0.2 4.4 0.9848 8.0 1.99 32.1 0.0034 1.9 0.268 13.5 0.5201
Kidney Infection 0.95 0.4 2.5 0.9187 0.5 0.2 1.6 0.2469 0.2 0.052 0.9 0.0387
90 Days After
Mecillinam 9.88 1.9 50.4 0.0059 2.99 0.9 9.6 0.0668 0.8 0.265 2.4 0.6714
Nitrofurantoin 3.2 0.9 11.4 0.0708 4.7 1.5 14.8 0.0076 3.3 1.050 10.4 0.0410
Penicillin (+ effect on Gram-neg.)
0.5 0.1 2.9 0.4727 1.3 0.4 4.2 0.6394 4.5 1.379 14.5 0.0126
Sulphonamides 6.2 1.1 33.8 0.0336 6.4 1.7 24.5 0.0070 7.8 2.284 26.9 0.0011
Trimethoprim . . . 0.0263* 9.7 1.9 50.4 0.0071 4.1 0.726 23.2 0.1100
Appendix 2 cont. Simplified manual backward stepwise elimination procedure including variables with P<0.1
Variables OD 95 CI P-Value
OD 95 CI P-Value OD 95 CI P-Value
30 Days Prior
Mecillinam 3.01 1.8 4.9 <.0001
3.9 2.5 6.4 <.0001 3.3 2.0 5.2 <.0001
Sulphonamides 0.97 0.3 2.4 0.9533
2.6 1.4 4.99 0.0041 2.1 1.1 3.99 0.0201
Trimethoprim 3.33 0.4 27.2 0.2616
4.6 1.2 17.4 0.0249 1.0 0.1 7.2 0.9894
90 Days Prior
Mecillinam 2.30 1.6 3.4 <.0001
3.1 1.8 5.4 <.0001 2.8 1.6 4.9 0.0003
Penicillin 2.78 1.2 6.3 0.0148
3.1 1.2 8.3 0.0225 2.3 0.8 6.4 0.1048
Sulphonamides 0.92 0.4 1.95 0.8234
2.0 1.1 3.9 0.0326 1.9 0.98 3.6 0.0548
30 Days After
Antibiotics 1.52 0.8 2.98 0.1549
2.3 1.4 3.8 0.0017 . . . <.0001*

PhD Dissertation
Kidney Infection 2.62 1.2 5.8 0.0171 . . . 0.0816* 0.8 0.1 6.3 0.8726
Mecillinam 11.5 3.6 36.4 <.0001 7.1 2.6 19.4 0.0001 6.9 2.8 17.0 <.0001
Nitrofurantoin 34.8 3.9 312.6 0.0015 8.0 0.9 68.0 0.0567 9.9 1.3 77.8 0.0291
Sulphonamides 8.9 0.8 98.1 0.0729 6.2 0.7 55.0 0.1036 20.7 2.7 158.7 0.0036
90 Days After
Mecillinam 2.9 0.9 9.1 0.0574 2.5 1.5 4.1 0.0006
Nitrofurantoin 3.6 1.5 8.4 0.0030 2.8 1.4 5.4 0.0954
Penicillin (+ effect on Gram-neg)
1.4 0.5 3.8 0.5569 6.5 1.96 21.5 0.0027
Sulphonamides 5.5 1.6 18.4 0.0055 7.1 2.4 21.1 0.0022

Copenhagen 2014
85
Appendix 3. . Results of multivariate logistic regression analyses comparing the ESBL group to other case groups. These tests were performed as previously described in Table 6. *P-values found by chi-square analysis. OR = Odds Ratio.
Multivariate Logistic Regression. First run.
ESBL vs. Resistant ESBL vs. Susceptible
Variables OR 95 CI P-Value OR 95 CI P-Value
COI vs. HCAI 1.5 0.8 2.8 0.1660* 2.4 1.3 4.6 0.0058*
Sex 0.7 0.3 1.7 0.4335 0.7 0.3 1.7 0.4606
Age 1.0 0.99 1.02 0.4387 1.0 0.99 1.02 0.1837
30 Days Prior
Admissions 4.95 1.5 15.9 0.0073 3.9 1.2 12.7 0.0216
Sulphonamides 0.3 0.1 0.9 0.0381 0.3 0.08 0.9 0.0299
90 Days Prior
Antibiotics 2.0 1.2 3.3 0.0049 2.5 1.4 4.7 0.0029
Insulin 1.3 0.8 2.4 0.3218 2.5 1.0 6.1 0.0447
Nitrofurantoin 0.4 0.2 0.8 0.0089 0.6 0.3 1.3 0.1840
Sulphonamides 0.3 0.1 0.6 0.0010 0.3 0.1 0.8 0.0091
Trimethoprim 0.3 0.1 0.8 0.0096 0.6 0.2 2.2 0.4663
30 Days After
Alimentary tract and metabolism 0.8 0.5 1.2 0.2260 0.6 0.4 0.96 0.0344
Antibiotics 1.1 0.6 1.9 0.7451 1.9 1.0 3.4 0.0470
Mecillinam 1.7 0.4 6.7 0.4336 4.03 0.99 16.4 0.0515
90 Days After
Genitourinary and sex hormones 0.2 0.04 0.8 0.0225 0.2 0.03 0.8 0.0245
Mecillinam 1.6 0.7 4.5 0.3806 6.9 2.04 23.5 0.0019
Penicillin (with effect on Gram-negative) 0.9 0.3 2.8 0.9081 0.2 0.075 0.9 0.0414
Penicillin in combination w. beta-lactamase inhibitors
5.3 1.2 22.2 0.0240 1.2 0.33 4.3 0.7795
Appendix 3 cont. Simplified manual backward stepwise elimination procedure including variables with P<0.1
30 Days Prior Variables OR 95 CI P-Value OR 95 CI P-Value
Trimethoprim 0.3 0.1 0.99 0.0486 1.3 0.1 11.7 0.8100
Sulphonamides 0.4 0.2 0.99 0.0486 0.3 0.1 0.7 0.0088
Quinolones 3.4 1.08 10.6 0.0365 2.08 0.6 7.3 0.2489
Antibiotics 0.98 0.6 1.5 0.9194 1.8 1.02 3.1 0.0414
Admissions 4.1 1.35 12.1 0.0124 3.4 1.1 10.5 0.0295
90 Days Prior
Antibiotics 1.2 0.97 1.4 0.1044 1.4 1.1 1.8 0.0024
Sulphonamides 0.6 0.3 0.95 0.0316 0.6 0.4 1.05 0.0723
90 Days After
Mecillinam 1.4 0.5 3.9 0.4624 4.1 1.4 11.8 0.0096 Penicillin in combination w. beta-
lactamase inhibitors 4.4 1.1 17.7 0.0362 1.767 0.8 4.0 0.1734
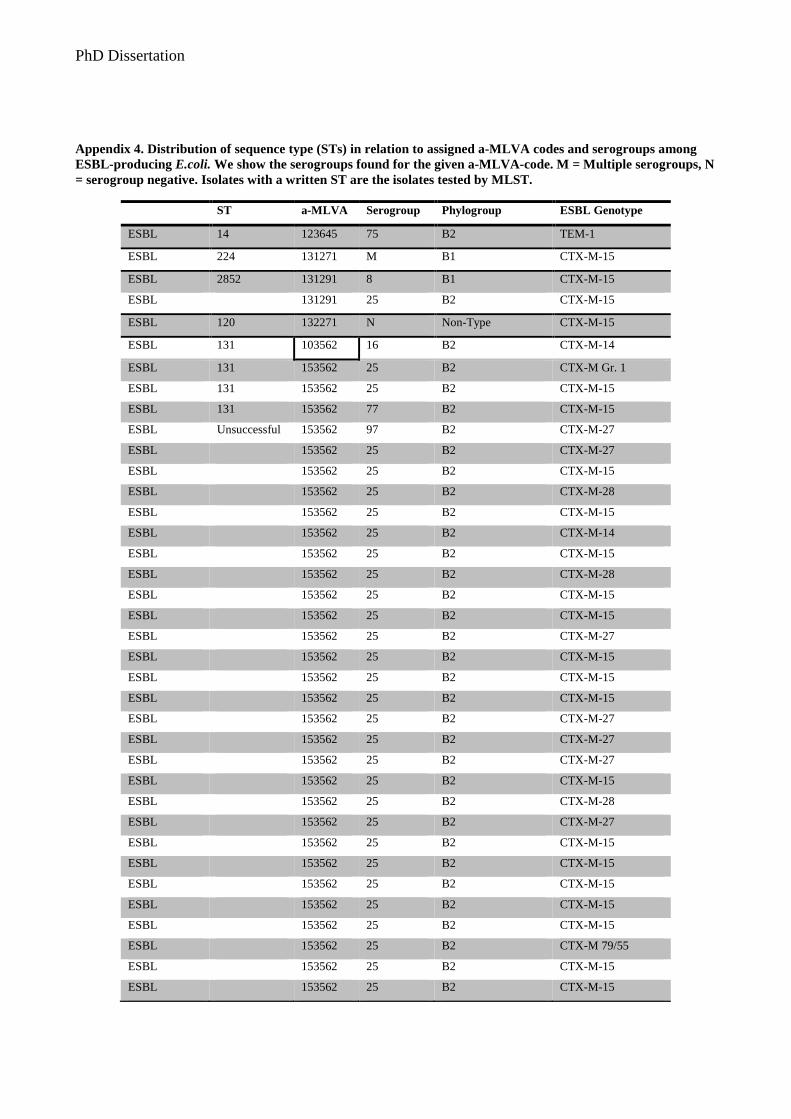
PhD Dissertation
Appendix 4. Distribution of sequence type (STs) in relation to assigned a-MLVA codes and serogroups among ESBL-producing E.coli. We show the serogroups found for the given a-MLVA-code. M = Multiple serogroups, N = serogroup negative. Isolates with a written ST are the isolates tested by MLST.
ST a-MLVA Serogroup Phylogroup ESBL Genotype
ESBL 14 123645 75 B2 TEM-1
ESBL 224 131271 M B1 CTX-M-15
ESBL 2852 131291 8 B1 CTX-M-15
ESBL 131291 25 B2 CTX-M-15
ESBL 120 132271 N Non-Type CTX-M-15
ESBL 131 103562 16 B2 CTX-M-14
ESBL 131 153562 25 B2 CTX-M Gr. 1
ESBL 131 153562 25 B2 CTX-M-15
ESBL 131 153562 77 B2 CTX-M-15
ESBL Unsuccessful 153562 97 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-28
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-14
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-28
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-28
ESBL 153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M 79/55
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15

Copenhagen 2014
87
ESBL 153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL 153562 M B2 CTX-M-27
ESBL 153562 M B2 CTX-M-15
ESBL 153562 M B2 CTX-M-15
ESBL 153562 M B2 CTX-M-15
ESBL 153562 MISSING B2 CTX-M-28
ESBL 153562 MISSING B2 CTX-M-27
ESBL 153562 N B2 CTX-M-28
ESBL 131 163562 16 B2 CTX-M-15
ESBL 131 163562 16 B2 CTX-M-15
ESBL 163562 16 B2 CTX-M-15
ESBL 163562 16 B2 CTX-M-1
ESBL 163562 153 B2 CTX-M-15
ESBL 648 161160 1 D CTX-M-15
ESBL 648 161160 102,130 D CTX-M-15
ESBL 161160 153 D CTX-M-15
ESBL 161160 MISSING D CTX-M-15
ESBL 62 161370 MISSING D CTX-M-14
ESBL 69 173277 15 D CTX-M Gr. 1
ESBL 173277 15 D CTX-M Gr. 1
ESBL 173277 15 D CTX-M-79/55
ESBL 173277 15 D CTX-M-27
ESBL 173277 44 D TEM-1
ESBL 173277 73 D CTX-M-14
ESBL 315 173552 25 B2 CTX-M-14
ESBL 428 252365 117 B2 CTX-M-1
ESBL 354 151170 153 D CTX-M-14
ESBL 998 224665 2 B2 CTX-M-15
ESBL 998 224665 M B2 CTX-M-1
ESBL 88 132261 119 A CTX-M-1
ESBL 636 293852 21 B2 CTX-M-15
ESBL 101 131261 N B1 CTX-M-1
ESBL 448 131261 111 B1 CTX-M-15
ESBL 131261 170 NT CTX-M Gr. 9
ESBL 131261 81 B1 CTX-M-79/55
ESBL 131261 100 NT CTX-M-1
ESBL 131261 M B1 CTX-M-15
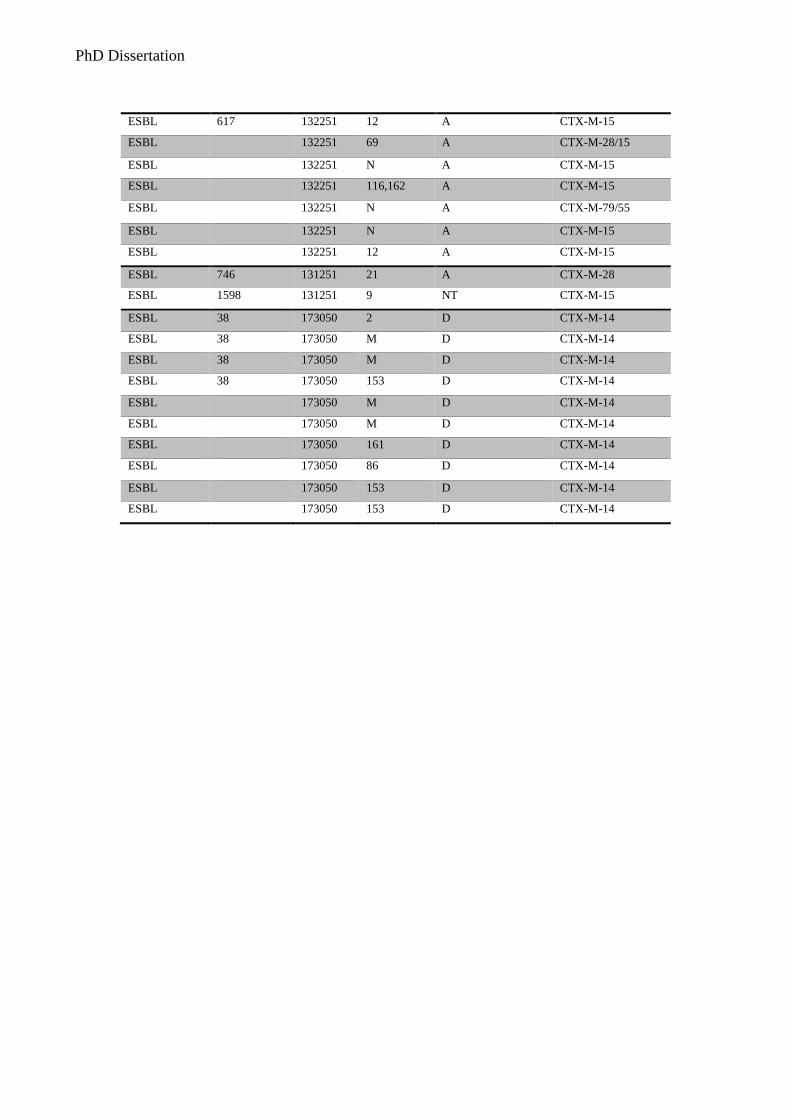
PhD Dissertation
ESBL 617 132251 12 A CTX-M-15
ESBL 132251 69 A CTX-M-28/15
ESBL 132251 N A CTX-M-15
ESBL 132251 116,162 A CTX-M-15
ESBL 132251 N A CTX-M-79/55
ESBL 132251 N A CTX-M-15
ESBL 132251 12 A CTX-M-15
ESBL 746 131251 21 A CTX-M-28
ESBL 1598 131251 9 NT CTX-M-15
ESBL 38 173050 2 D CTX-M-14
ESBL 38 173050 M D CTX-M-14
ESBL 38 173050 M D CTX-M-14
ESBL 38 173050 153 D CTX-M-14
ESBL 173050 M D CTX-M-14
ESBL 173050 M D CTX-M-14
ESBL 173050 161 D CTX-M-14
ESBL 173050 86 D CTX-M-14
ESBL 173050 153 D CTX-M-14
ESBL 173050 153 D CTX-M-14

Copenhagen 2014
89
Appendix 5. Distribution of sequence type (STs) in relation to assigned a-MLVA-codes and serogroups, for the resistant group. Not all resistant isolates were characterized by serogroup. Here we show the serogroups found for the given a-MLVA-code. M = Multiple serogroups, N = serogroup negative. Isolates with a written ST are the isolates tested by MLST.
ST a-MLVA Serogroup Phylogroup
Resistant 405 121250 M D
Resistant 121250 D
Resistant 405 131250 D
Resistant 131250 D
Resistant 14 123645 75 B2
Resistant 362 143250 D
Resistant 131 143562 B2
Resistant 131 153562 25 B2
Resistant 131 153562 B2
Resistant 153562 25 B2
Resistant 153562 25 B2
Resistant 153562 25 B2
Resistant 153562 B2
Resistant 153562 B2
Resistant 153562 B2
Resistant 153562 B2
Resistant 153562 B2
Resistant 131 163562 B2
Resistant 163562 B2
Resistant 163562 B2
Resistant 648 161150 1 D
Resistant 62 161370 D
Resistant 457 161371 11 D
Resistant 1597 162562 39 B2
Resistant 1597 162562 B2
Resistant 1193 124645 75 B2
Resistant 1193 124645 6 B2
Resistant 124645 B2
Resistant 124645 D
Resistant 124645 B2
Resistant 124645 75 B2
Resistant 393 171577 D
Resistant 69 173270 73 D
Resistant 173270 D
Resistant 69 173277 15 D
Resistant 69 173277 25 D
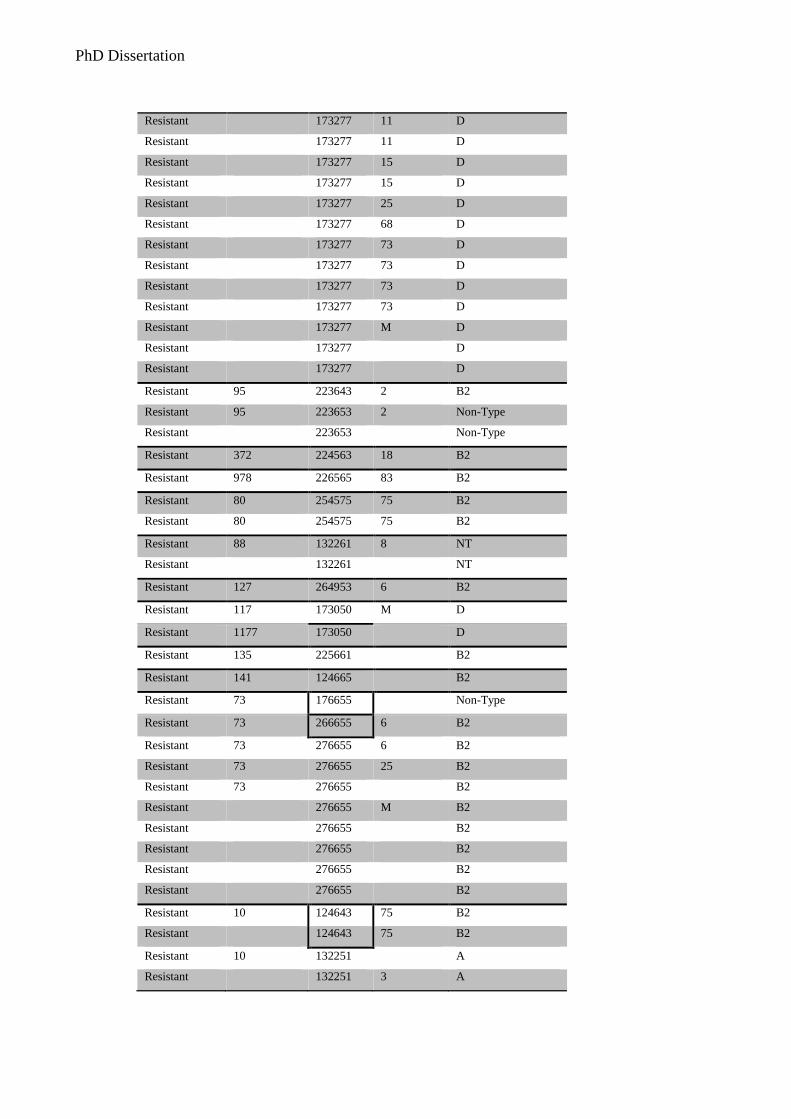
PhD Dissertation
Resistant 173277 11 D
Resistant 173277 11 D
Resistant 173277 15 D
Resistant 173277 15 D
Resistant 173277 25 D
Resistant 173277 68 D
Resistant 173277 73 D
Resistant 173277 73 D
Resistant 173277 73 D
Resistant 173277 73 D
Resistant 173277 M D
Resistant 173277 D
Resistant 173277 D
Resistant 95 223643 2 B2
Resistant 95 223653 2 Non-Type
Resistant 223653 Non-Type
Resistant 372 224563 18 B2
Resistant 978 226565 83 B2
Resistant 80 254575 75 B2
Resistant 80 254575 75 B2
Resistant 88 132261 8 NT
Resistant 132261 NT
Resistant 127 264953 6 B2
Resistant 117 173050 M D
Resistant 1177 173050 D
Resistant 135 225661 B2
Resistant 141 124665 B2
Resistant 73 176655 Non-Type
Resistant 73 266655 6 B2
Resistant 73 276655 6 B2
Resistant 73 276655 25 B2
Resistant 73 276655 B2
Resistant 276655 M B2
Resistant 276655 B2
Resistant 276655 B2
Resistant 276655 B2
Resistant 276655 B2
Resistant 10 124643 75 B2
Resistant 124643 75 B2
Resistant 10 132251 A
Resistant 132251 3 A

Copenhagen 2014
91
Resistant 132251 A
Resistant 132251 A
Resistant 132251 A
Resistant 132251 N A
Resistant 2279 132251 15 A
Resistant 38 173051 25 D
Resistant 58 131261 19 B1
Resistant 131261 B1
Resistant 131261 NT
Resistant 131261 N NT
Resistant 131261 69 B1
Resistant 131261 NT
Resistant New ST 123552 B2
Resistant New ST 131251 NT
Resistant New ST 132261 NT
Resistant New ST 162438 NT
Resistant MISSING 266561 NT

PhD Dissertation
Appendix 6. Distribution of sequence type (STs) in relation to assigned a-MLVA-codes and serogroups for the susceptible group. Not all susceptible isolates were characterized by serogroup. Here we show the serogroups found for the given a-MLVA-code. M = Multiple serogroups, N = serogroup negative. Isolates with a written ST are the isolates tested by MLST.
ST a-MLVA Serogroup Phylogroup
Susceptible 223 101261 B2
Susceptible 101261 Non-Type
Susceptible 101261 B2
Susceptible 69 103277 B1
Susceptible 69 173277 D
Susceptible 173277 D
Susceptible 173277 D
Susceptible New ST 112871 D
Susceptible 1161 124675 B2
Susceptible 405 131250 2 D
Susceptible 48 131281 Non-Type
Susceptible 131281 119 Non-Type
Susceptible 357 150365 B2
Susceptible 357 152365 73 B2
Susceptible 152365 M B2
Susceptible 538 152363 13 B2
Susceptible 131 153562 B2
Susceptible 62 161370 D
Susceptible 501 163777 D
Susceptible New ST 171250 D
Susceptible 95 203653 B2
Susceptible 95 223653 1 B2
Susceptible 223653 1 B2
Susceptible 223653 M B2
Susceptible 223653 B2
Susceptible 223653 Non-Type
Susceptible 73 176655 18 B2
Susceptible 73 203655 2 D
Susceptible 73 206653 B2
Susceptible 73 206655 B2
Susceptible 73 226655 B2
Susceptible 73 256653 D
Susceptible 73 266655 2 B2
Susceptible 266655 B2
Susceptible 73 276555 25 B2
Susceptible 73 276655 6 B2

Copenhagen 2014
93
Susceptible 73 276655 22 B2
Susceptible 73 276655 M B2
Susceptible 276655 2 B2
Susceptible 276655 B2
Susceptible 73 276665 120 B2
Susceptible 162 211251 95 A
Susceptible 681 224452 8 D
Susceptible 1858 224645 6 B2
Susceptible 4235 224743 88 B2
Susceptible 224743 88 B2
Susceptible 3672 226962 B2
Susceptible New ST 232251 105 A
Susceptible 59 241370 D
Susceptible 59 251380 D
Susceptible 80 254575 B2
Susceptible 127 264953 B2
Susceptible 264953 6 B2
Susceptible 127 274953 6 B2
Susceptible 12 266562 4 B2
Susceptible 12 266562 B2
Susceptible New ST 272275 133 B2
Susceptible 1444 273673 4 B2
Susceptible 582 274242 B2
Susceptible 1331 274655 6 D
Susceptible 589 287572 B2
Susceptible 714 473050 D
Susceptible 3846 254665 B2
Susceptible 420 254665 82 B2
Susceptible 10 131251 109 A
Susceptible 10 131251 M A
Susceptible 131251 107,117 A
Susceptible 10 132251 81 A

Manuscript I to III

Published Ahead of Print 4 August 2014. 10.1128/AAC.03021-14.
2014, 58(10):6139. DOI:Antimicrob. Agents Chemother. Frimodt-MøllerFrederik Boetius Hertz, Anders Løbner-Olesen and Niels Colonization ModelSequence Type 131 in a Mouse Intestinal Antibiotic Selection of Escherichia coli
http://aac.asm.org/content/58/10/6139Updated information and services can be found at:
These include:
REFERENCEShttp://aac.asm.org/content/58/10/6139#ref-list-1at:
This article cites 31 articles, 16 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Septem
ber 27, 2014 by guesthttp://aac.asm
.org/D
ownloaded from
on S
eptember 27, 2014 by guest
http://aac.asm.org/
Dow
nloaded from

Antibiotic Selection of Escherichia coli Sequence Type 131 in a MouseIntestinal Colonization Model
Frederik Boetius Hertz,a,b,c Anders Løbner-Olesen,b Niels Frimodt-Møllera
Department of Clinical Microbiology, Hvidovre University Hospital, Hvidovre, Denmarka; Department of Biology, University of Copenhagen, Copenhagen, Denmarkb;Statens Serum Institut, Copenhagen, Denmarkc
The ability of different antibiotics to select for extended-spectrum �-lactamase (ESBL)-producing Escherichia coli remains atopic of discussion. In a mouse intestinal colonization model, we evaluated the selective abilities of nine common antimicrobials(cefotaxime, cefuroxime, dicloxacillin, clindamycin, penicillin, ampicillin, meropenem, ciprofloxacin, and amdinocillin) againsta CTX-M-15-producing E. coli sequence type 131 (ST131) isolate with a fluoroquinolone resistance phenotype. Mice (8 pergroup) were orogastrically administered 0.25 ml saline with 108 CFU/ml E. coli ST131. On that same day, antibiotic treatmentwas initiated and given subcutaneously once a day for three consecutive days. CFU of E. coli ST131, Bacteroides, and Gram-posi-tive aerobic bacteria in fecal samples were studied, with intervals, until day 8. Bacteroides was used as an indicator organism forimpact on the Gram-negative anaerobic population. For three antibiotics, prolonged colonization was investigated with addi-tional fecal CFU counts determined on days 10 and 14 (cefotaxime, dicloxacillin, and clindamycin). Three antibiotics (cefo-taxime, dicloxacillin, and clindamycin) promoted overgrowth of E. coli ST131 (P < 0.05). Of these, only clindamycin suppressedBacteroides, while the remaining two antibiotics had no negative impact on Bacteroides or Gram-positive organisms. Only clin-damycin treatment resulted in prolonged colonization. The remaining six antibiotics, including ciprofloxacin, did not promoteovergrowth of E. coli ST131 (P > 0.95), nor did they suppress Bacteroides or Gram-positive organisms. The results showed thatantimicrobials both with and without an impact on Gram-negative anaerobes can select for ESBL-producing E. coli, indicatingthat not only Gram-negative anaerobes have a role in upholding colonization resistance. Other, so-far-unknown bacterial popu-lations must be of importance for preventing colonization by incoming E. coli.
Escherichia coli is a versatile and ubiquitous species that is regu-larly represented in the commensal flora of the gut. The species
includes nonpathogenic, intestinal pathogenic, and extraintesti-nal pathogenic E. coli (ExPEC), all of which can be variably presentin the human gut. ExPEC can cause a wide range of infections,from uncomplicated cystitis to life-threatening sepsis (1–3). Ma-jor sources for resistance in E. coli are plasmid-borne extended-spectrum �-lactamases (ESBL), which are enzymes capable of hy-drolyzing, and thus conferring resistance toward, most �-lactamantibiotics, except for the cephamycins and carbapenems. In ad-dition, ESBLs are inhibited by �-lactamase inhibitors, such as cla-vulanic acid, sulbactam, and tazobactam (1, 4). Plasmids carryingESBL genes often carry various other genes that cause resistance toother classes of antibiotics (e.g., aminoglycosides) (1, 4, 5). One ofthe most common types of ESBLs identified in the world is CTX-M-15, and the spread of its areas of endemicity seems to be asso-ciated with a few E. coli sequence types (ST), such as ST131. ST131is most frequently linked to quinolone resistance and CTX-M-15,but it also harbors specific virulence genes coding for factors suchas adhesins (fimH, papEF), toxins (sat), and capsules (kpsM II),contributing to its ability to colonize the human gut (2, 4–7). Thepossible human colonization is of great concern, since ST131 iso-lates often are resistant to several antibiotics and are known tocause urosepsis to a higher degree than non-ST131 isolates. Inclinical settings, multiresistance, including production of ESBL,delays appropriate treatment, leading to extended hospital stays aswell as increased mortality and morbidity (8–10). Finally, urinarytract infections are primarily caused by E. coli present in the pa-tient’s own microbiota, making knowledge on colonization byresistant E. coli of great importance (1, 4, 11).
Several case-control studies have identified recent antibiotic
exposure, especially to cephalosporins and fluoroquinolones, andhospitalization as significant risk factors for acquiring an infectionwith an ESBL-producing Enterobacteriaceae (within 30 days) (10,12–14). A systematic study of the selective ability of all antibioticclasses has been lacking. In this study, we wished to evaluate theability of nine common antimicrobials, including antibiotics usedfor Gram-positive infections, to select for a CTX-M-15-producingE. coli ST131 isolate in a mouse intestinal colonization model.
(This work was presented as a poster at the 52nd InterscienceConference on Antimicrobial Agents and Chemotherapy, SanFrancisco, CA, 9 to 12 September 2012.)
MATERIALS AND METHODSStrain. For the colonizing pathogen, we used a clinical blood isolate of E.coli that belongs to the lineage B2-O25b-ST131. This isolate (65-Ec-09)carries some of the virulence factors previously seen in ST131 isolatesfound throughout Denmark in 2009, and it is similarly resistant towardmany commonly used antibiotics (MICs when resistant: cefotaxime, �32�g/ml; cefuroxime, �256 �g/ml; ceftazidime, 32 �g/ml; ampicillin, �256�g/ml; aztreonam, 32 �g/ml; amoxicillin-clavulanate, �256 �g/ml; cip-rofloxacin, �32 �g/ml; cloxacillin, �256 �g/ml; clindamycin, �256 �g/ml. MICs when susceptible are as follows: amdinocillin, 2 �g/ml; trim-ethoprim-sulfamethoxazole, �0.125 �g/ml; gentamicin, �0.5 �g/ml;
Received 8 April 2014 Returned for modification 24 May 2014Accepted 31 July 2014
Published ahead of print 4 August 2014
Address correspondence to Frederik Boetius Hertz, [email protected].
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AAC.03021-14
October 2014 Volume 58 Number 10 Antimicrobial Agents and Chemotherapy p. 6139 – 6144 aac.asm.org 6139
on Septem
ber 27, 2014 by guesthttp://aac.asm
.org/D
ownloaded from

piperacillin-tazobactam, 2 �g/ml; meropenem, �0.064 �g/ml; nitrofu-rantoin, 13 millimeters in zone diameter [mm]; trimethoprim, 27 mm)(15). Pheno- and genotypic characterizations were performed by usingthe MAST-test, PCR, and DNA sequence analysis according to methodsused at the clinical laboratory of Department of Clinical Microbiology,Hvidovre Hospital, Denmark (HVH) or at Statens Serum Institut (SSI) aspreviously described (15). Sequence type verification was performed viafull multilocus sequence typing (MLST, using the Achtmann scheme[http://mlst.warwick.ac.uk/mlst/dbs/Ecoli]) by SSI, who also found thatthe isolate harbored the virulence factors kpsM II and iutA (15–17).
Media. To test bacterial growth in collected fecal samples from mice,we used selective agar plates, all from SSI Diagnostica, Hilleroed, Den-mark. For the CTX-M-15-producing E. coli population, ID Flexicult agarcontaining cefotaxime at 32 mg/liter and vancomycin at 6 mg/liter wereused. The Gram-positive aerobic population was selected on 5% bloodagar plates containing gentamicin at 5 mg/liter, and the Gram-negativeanaerobic population was selected on anaerobic plates containing genta-micin at 32 mg/liter and vancomycin at 16 mg/liter. Culturing of anaero-bic species were performed under anaerobic conditions in GasPak EZcontainers and an anaerobic atmosphere created by using AnaeroGen(Oxoid) (18).
MIC determinations. Antibiotic susceptibility testing was performedas Etests when possible and as disc diffusion tests where no Etest wasavailable (the MIC for dicloxacillin was found by using an Etest for clox-acillin, since neither Etest nor disks for dicloxacillin were available). Thediffusion test methodology has been described elsewhere (15), and a sim-ilar Etest methodology was used according to guidelines from the Depart-ment of Clinical Microbiology, Hvidovre Hospital. All results were inter-preted according to current recommendations from the EuropeanCommittee on Antimicrobial Susceptibility Testing (http://www.eucast.org/clinical_breakpoints/).
Antibiotics used for treatment. Commonly used antibiotics werechosen for this study. These included different �-lactam antibiotics, cip-rofloxacin, and clindamycin. All mouse dosages were calculated based onhuman doses (in mg per kg of body weight) from pharmacokinetic (PK)studies performed at SSI or from previously published mouse studies(Table 1) (19–26). Doses were chosen to mimic the serum antibiotic con-centrations achieved in humans on standard doses. All antibiotics wereadministered subcutaneously once each day for 3 consecutive days. Con-centrations of antibiotics in mouse feces had been measured in a previ-ously published study for a �-lactam antibiotic, an expanded-spectrumcephalosporin, a carbapenem, clindamycin, and ciprofloxacin (19). Thedoses used in this study were similar or higher.
Mouse intestinal colonization model. (i) Mice. The animal experi-ment was approved by the Danish Centre for Animal Welfare and carried
out at Statens Serum Institut in Copenhagen, Denmark. In all studies, 7-to 10-week-old female albino, outbred NMRI mice (Harlan, the Nether-lands) weighing 26 to 30 g were used. The mice used in each study were allfrom the same litter and were brought simultaneously to the stable andhoused in pairs of two per cage. At the end of the study, all mice weresacrificed to ensure that no mice were kept alive with no immediate pur-pose. Animals were housed, treated, and sacrificed according to currentguidelines.
(ii) The mouse model. The mouse intestinal colonization model wasan experimental model where all mice were kept in pairs of two per cage.Two cages constituted one group and each group received one antibiotic.Thus, each antibiotic was given to a total of four mice in two differentcages, and a total of 20 to 22 cages were included in the study. Treatmentwas given subcutaneously in the neck once a day for three consecutivedays (day 1 to day 3). Inoculation of mice with the bacterial strain wasdone through a stainless steel orogastric feeding tube on day 1 prior toinitiation of treatment. The intestinal flora was unaltered prior to thestudy, and no mice were anesthetized during the study. The experimentwas conducted from day 1 to day 8, and cages were changed daily. At theend of day 8 all mice were sacrificed.
(iii) Experimental study. We executed our full experimental study byusing the described mouse intestinal colonization model. The study wasconducted from day 1 to day 8 with feces collected prior to inoculation onday 1 and on days 2, 4, and 8. On day 1, mice were inoculated once with0.25 ml of saline containing 108 CFU/ml of 65-Ec-09. Thus, each mousewas given an inoculum of 2.5 � 107 CFU.
The mice were left for 3 h before the first doses of antibiotics wereadministered subcutaneously. Each treatment group, consisting of fourmice housed in two different cages, received cefotaxime, cefuroxime, am-picillin, dicloxacillin, amdinocillin, meropenem, clindamycin, cipro-floxacin, or benzylpenicillin (Table 1). A control group received 65-Ec-09but no antibiotic treatment. The complete selection study of E. coli ST131CTX-M-15 (65-Ec-09) in the mouse intestinal colonization model wasperformed twice, with the exception of CFU counts of the Gram-positiveaerobic flora, which were only studied in the second run. The controlgroup receiving 65-Ec-09 only was included in both runs. Furthermore, agroup of mice that received treatment with cefotaxime without receivingthe ESBL-producing strain was included in the first run only.
(iv) Prolonged presence of resistant pathogen after completed anti-biotic treatment. Additionally, the prolonged presence of the E. coliST131 after completed antibiotic treatment was studied in the first run forthree antibiotics (cefotaxime, dicloxacillin, and clindamycin). These threeantibiotics were chosen based on their selective abilities found in thestudy. Treatment stopped, for all groups, on day 3. Cages were changed
TABLE 1 Antibiotics used as treatmentsa
Antibiotic
Humanb Mouse
Dose (g, i.v.) Cmax (�g/ml)Needed dose(mg/kg) Cmax (�g/ml)
Dose given(mg/mouse/day)
Cefuroxime 1.5 65 120 50–60 4Cefotaxime 1 40 60 100 2Ampicillin 1 40 50 75 1.5Dicloxacillin 1 30–40 60 90 2Benzylpenicillin 2 mill. IEb 60 70 60 2Amdinocillin 0.4 30–40 60 30 2Meropenem 0.5 26 50 50 1.5Clindamycin 1.8 6 36 8 1.4Ciprofloxacin 0.4 4 15 2 0.5a All doses were administered subcutaneously once a day, and the needed doses were calculated based on the expected average weight of the mice (weights given by provider), asdescribed in previously published studies (18–25). All mouse dosages were calculated based on human doses from PK studies performed at SSI or from previously published mousestudies (19–26). i.v., intravenous.b Benzylpenicillin (1.2 grams � 2 millions units � 2 mill. IE) was administered intravenously.
Boetius Hertz et al.
6140 aac.asm.org Antimicrobial Agents and Chemotherapy
on Septem
ber 27, 2014 by guesthttp://aac.asm
.org/D
ownloaded from

daily from days 1 to 14, and feces samples were collected on days 10 and14. On day 14, the study was terminated and mice were killed.
Detection and quantification of bacteria in feces. On specified days,0.5 g of feces was collected from each cage. Feces were dissolved in 5 ml ofsaline and further diluted 10-fold in saline for a total of 6 times. Dilutionswere plated on the different selective agar plates, and the log CFU per 0.5g of stool for 65-Ec-09, the Gram-negative anaerobic flora, and the Gram-positive aerobic flora were calculated for each cage and two or three col-onies from each day frozen. Each CFU count was performed on two agarplates and calculated as the average CFU count for the two plates. Toensure no presence of cefotaxime-resistant E. coli prior to inoculation,dilutions of feces from day 1, from each cage, were spotted on selectiveplates and this showed no growth of resistant E. coli. The lower detectionlimit was 10 CFU per 0.5 g of feces.
Molecular tests. To ensure that the E. coli found in feces was identicalto the isolate given through inoculation, we tested a total of 17 cefotaxime-resistant E. coli isolates found in feces during treatment and 4 E. coliisolates from day 1 for the presence of a CTX-M group 1 gene. Samplesfrom both experimental runs were included (see below). Three to fourisolates per frozen sample were used for DNA purification. As these testswere performed on isolates from both study runs, they were not per-formed until both runs had been completed. Thus, isolates tested wereisolated from frozen samples. We were therefore likely to isolate the dom-inating E. coli strain and thereby determine if 65-Ec-09 had become thedominating E. coli of the microbiota (11, 27).
All of these 21 isolates were identified as E. coli by matrix-assisted laserdesorption–time of flight analysis as described elsewhere (28). All resis-tant E. coli isolates contained a CTX-M group 1 gene, whereas none of theisolates from day 1 contained a CTX-M group 1 gene.
Isolates from the following groups were tested: cefotaxime (day 2 fromboth study runs and day 1 from the first run), cefuroxime (day 2 fromboth study runs and day 1 from the first run), ampicillin (day 2 from bothstudy runs), dicloxacillin (days 1, 2, 4, and 8 from the first run and days 2and 8 from the second run), clindamycin (days 1, 2, 4, and 10 from the firstone and days 2 and 8 from the second run), and benzylpenicillin (day 2from second run).
Additionally, five of the cefotaxime-resistant E. coli isolates and oneisolate from day 1 were characterized by MLST (cefotaxime, cefuroxime[day 2], dicloxacillin [days 1 and 4], and clindamycin [days 4 and 10]).The five resistant E. coli were identified as ST131 and the one isolate fromday 1 belonged to ST602.
Statistical analysis. Data were analyzed with the use of SAS software,version 9.3 (SAS Institute). When just one group receiving one antibioticwas compared to the control group, this was done via a one-way analysisof variance (ANOVA). When multiple CFU counts were compared, it wasdone as a multiple variant analysis, adjusted for multiple comparisonswith the Bonferroni correction. The conservative Bonferroni correction
was used to avoid multiple comparisons. A P value of �0.05 was consid-ered significant.
RESULTS
Results of fecal bacteriology were calculated as the log CFU/0.5 gof feces, and the mean CFU of four cages (two cages for Gram-positive aerobic flora) were used for statistical calculations. Forgraphic depictions, means and standard deviations (SD) wereused. Data for the selective abilities of the different antibiotics on65-Ec-09 are shown in Table 2 and Fig. 1 and 2 for E. coli, Gram-negative anaerobic flora, and Gram-positive flora, respectively.
Effect of antibiotic treatment on establishment of resistantpathogens and on the indigenous microflora. Data for the effectson colonization of 65-Ec-09 by the different antibiotics are shownin Fig. 1. Cefotaxime, dicloxacillin, and clindamycin promotedthe colonization and overgrowth of 65-Ec-09 from day 2 throughday 8 (P � 0.01 for dicloxacillin and clindamycin; P � 0.05 forcefotaxime). Benzylpenicillin and cefuroxime showed less over-growth but selective abilities on days 2 and 4 (P � 0.05). Dicloxa-cillin and clindamycin showed the highest selective abilities (P �0.01 for dicloxacillin on days 2 and 4 and P � 0.01 for clindamycinon all days). After treatment was completed on day 3, there was adecline in the colonization of 65-Ec-09 from day 4 to 8. In com-parison, neither ampicillin, amdinocillin, meropenem, nor cipro-floxacin promoted overgrowth of 65-Ec-09 beyond day 2 (P �0.05).
Data for the impact on the original microbiota as Gram-nega-tive anaerobic flora, represented by Bacteroides, and Gram-posi-tive flora, respectively, are shown in Fig. 2. None of the antibioticsused had an inhibiting or promoting impact on the Gram-positiveflora (P � 0.05). Only clindamycin had an impact on the Gram-negative anaerobic flora, as it completely eliminated the Gram-negative anaerobic flora during treatment (P � 0.05). After treat-ment, the CFU counts for the Gram-negative anaerobic floraincreased to counts equal to those before treatment.
Prolonged presence of 65-Ec-09 after completed antibiotictreatment. Data for CFU of 65-Ec-09 on all sampling days arefound in Fig. 1. After the initial increase, the CFU decreased for allthree groups of antibiotics. For dicloxacillin and cefotaxime, theCFU counts dropped below the detection limit on day 10. Forclindamycin, the CFU was measurable until day 14, the last day ofthe study. There were no significant differences in fecal CFU of65-Ec-09 over time among the three antibiotics (P � 0.05).
TABLE 2 Results of the CFU counts for E. coli ST131
Antibiotic
Log CFU � SD (P value) ona:
Day 2 Day 4 Day 8 Day 10 Day 14
Cefotaxime 7.75 � 4.33 (�0.01) 5.25 � 6.54 (�0.01) 3.61 � 2.5 (0.011) 1.5 � 1.5 (0.98) 0.5* � 3.55 (1)Clindamycin 8.5 � 1 (�0.01) 7.75 � 0.87 (�0.01) 5.75 � 0.87 (�0.01) 2.5 � 3.55 (0.44) 1* � 1.41 (1)Dicloxacillin 8.5 � 1 (�0.01) 9 � 5.35 (�0.01) 3 � 3.74 (�0.01) 0 (1) 0 (1)Cefuroxime 7.5 � 3 (�0.01) 4.25 � 5.89 (0.015) 1.75 � 4 (0.07)Penicillin 6.25 � 3.57 (0.02) 3.75 � 6.76 (0.03) 0 (1)Amdinocillin 2.25 � 2.96 (0.3) 1.25 � 5.55 (0.5) 0 (1)Meropenem 3.5 � 2.96 (1) 0 (1) 0 (1)Ciprofloxacin 2.25 � 2.6 (0.3) 0 (1) 0 (1)Ampicillin 3.25 � 4.33 (0.8) 2.5 � 6.33 (0.14) 0.75* � 2.6 (0.4)Control 3.5 � 1.73 0 0a P values for days 2 to 8 were determined via an ANOVA, while P values from days 10 and 14 were found by a multiple variant analysis with the Bonferroni correction.*, below thelower detection limit.
Antibiotic Selection of E. coli ST131
October 2014 Volume 58 Number 10 aac.asm.org 6141
on Septem
ber 27, 2014 by guesthttp://aac.asm
.org/D
ownloaded from

DISCUSSION
To our knowledge, this is the first study to experimentally evaluatein vivo selection in the gut of a CTX-M-15-producing E. coli ST131isolate by a range of commonly used antibiotics, including antibi-otics with no activity against Gram-negative bacteria. With themouse intestinal colonization model, we were able to illustrate theselective abilities of different antibiotics on the intestinal florawhen a virulent isolate of CTX-M-15-producing E. coli ST131 wasintroduced. We found several interesting aspects of selection.First, we confirmed that antibiotics with activity against the colo-nizing strain did not promote proliferation, since neither amdi-nocillin nor meropenem resulted in colonization. Second, ourfindings confirmed that an antibiotic, clindamycin, that elimi-nates the Gram-negative anaerobic flora with no in vitro activityagainst Enterobacteriaceae promotes proliferation of ESBL-pro-ducing E. coli. It has been shown that antimicrobial impact on thetotal anaerobic population is proportional to the impact on theBacteroides population (19). Third, of the �-lactams with no in-hibiting effects on the ESBL-producing E. coli, cefotaxime showedthe highest level of selection. Cefuroxime and benzylpenicillin did,however, show higher selection propensities than ampicillin, inagreement with what has previously been seen for high doses ofpenicillin (29). Furthermore, we discovered that dicloxacillin,with no obvious influence on either the Gram-negative anaerobicflora or other Gram-negative bacteria, promoted colonizationwith the ESBL-producing E. coli, while ciprofloxacin, with a lim-ited in vitro effect on the Gram-negative anaerobic bacteria but aneffect on Enterobacteriaceae, showed no abilities to select. Theseresults could indicate that not only Gram-negative anaerobes havea key responsibility upholding colonization resistance. It appearsthat, as for penicillin (29), an antibiotic with limited impact onGram-negative bacteria can select for a resistant E. coli isolate,suggesting that other bacterial populations, not measured in ourstudy, such as certain anaerobic Gram positives, are of importancefor preventing colonization from an incoming E. coli strain.
Finally, we found that the antibiotic with the highest impact on
the anaerobic population, here represented by the Bacteroidespopulation, seems to give room for prolonged colonization, i.e.,clindamycin showed high to medium levels of 65-Ec-09 until day14, compared to dicloxacillin and cefotaxime. This points to thepossibility that even if unknown Gram-positive populations, an-aerobic or aerobic, display colonization resistance, the Bacteroidespopulation seems to play a role in preventing prolonged coloni-zation.
Our study suggests that selection of E. coli ST131 is not derivedalone by the antibiotic impact on the major population, such asanaerobes, as seen with dicloxacillin versus ampicillin and cipro-floxacin. This finding could potentially alter the perception ofwhich antibiotics drive the spread of ESBL-producing E. coli, in-cluding ST131, even if clindamycin was the antibiotic that showedthe highest level of selection and longest duration of colonization.
The model has some limitations, since it does not include clin-ical aspects of selection, such as treatment with multiple antimi-crobials or long-term treatment, nor does it take into accountreexposure to resistant pathogens or retreatment after exposure.Also, we did not investigate the antibiotics’ effects on the totalbacterial population of the intestines, including shifts in the dom-inating phyla. Such an investigation of changes in species andchanges in the total Enterobacteriaceae population could poten-tially have described, in detail, factors influencing colonization.Future studies of the microbiota should be designed to fully de-scribe antibiotic impacts on the different phyla. The lack of selec-tive ability seen from ciprofloxacin on the CTX-M-15-producingE. coli ST131 isolate was not expected and was surprising. We havenot tested the mice used in this study for the presence of cipro-floxacin-resistant strains, and such a presence could explain thelack of selection seen here. Furthermore, extrapolation of resultsfrom the model is limited by the physiological differences betweenmice and humans, even though studies have shown that subcuta-neous treatment once a day in mice gives fecal antibiotic concen-trations equivalent to those of humans (19). Finally, we studiedonly the fecal—and not the mucosal—microflora of mice. Yet, it
FIG 1 The mean and SD of the log CFU/0.5 g of feces for 65-Ec-09 from day 1 to day 14. The arrow indicates treatment from day 1 to day 3.
Boetius Hertz et al.
6142 aac.asm.org Antimicrobial Agents and Chemotherapy
on Septem
ber 27, 2014 by guesthttp://aac.asm
.org/D
ownloaded from

has been postulated that, first, the mouse fecal flora is a mixture ofmucosal and luminal flora and, second, the intestinal microfloraof laboratory mice is comparable to the intestinal flora in humans(30, 31). We have no obvious explanation for the selective abilityof dicloxacillin, since this drug had no impact on the Gram-neg-ative anaerobic or the aerobic Gram-positive flora. More detailedevaluation of the mouse and human intestinal microbiomes mayprovide the reason for the change in intestinal flora imposed bydicloxacillin. A study of antibiotics’ general impacts on the micro-flora of mice and men would further illustrate the elements in-volved in selection.
In summary, our study confirms that antibiotics with an im-pact on Gram-negative anaerobes support overgrowth and colo-nization of a CTX-M-15 producing E. coli ST131 isolate. None-theless, our study shows that selection could be driven byantibiotics with limited effect on anaerobes and no effect on com-peting Gram negatives, but not by other antimicrobials with abroader spectrum of activity. The results indicate a need for inves-tigation of selective mechanisms of different drugs, to fully de-velop rules and guidelines for stewardship of antibiotics.
ACKNOWLEDGMENTS
This study was performed with financial support from PAR, an EU FP7-Health-2009-Single-Stage project. Additionally, the work was supportedby The Danish Council for Strategic Research (DanCARD project 09-
067075/DSF), Roskilde University, the Aase & Ejnar Danielsens Founda-tion, and the SSAC Foundation.
Furthermore, the work was performed in cooperation between StatensSerum Institut, Hvidovre University Hospital, Roskilde University, andCopenhagen University. We are grateful to and thank Anette M. Ham-merum and Frank Hansen from SSI, who provided strain 65-Ec-09.
REFERENCES1. Rogers BA, Sidjabat HE, Paterson DL. 2011. Escherichia coli O25b-
ST131: a pandemic, multiresistant, community-associated strain. J. Anti-microb. Chemother. 66:1–14. http://dx.doi.org/10.1093/jac/dkq415.
2. Pitout JDD. 2012. Extraintestinal pathogenic Escherichia coli: a combi-nation of virulence with antibiotic resistance. Front. Microbiol. 3:9. http://dx.doi.org/10.3389/fmicb.2012.00009.
3. Kaper JB, Nataro JP, Mobley HL. 2004. Pathogenic Escherichia coli. Nat.Rev. Microbiol. 2:123–140. http://dx.doi.org/10.1038/nrmicro818.
4. Pitout JDD, Laupland KB. 2008. Extended-spectrum beta-lactamase-producing Enterobacteriaceae: an emerging public-health concern. Lan-cet 8:159 –166.
5. Peirano G, Pitout JDD. 2010. Molecular epidemiology of Escherichia coliproducing CTX-M beta-lactamases: the worldwide emergence of cloneST131 O25:H4. Int. J. Antimicrob. Agents 35:316 –321. http://dx.doi.org/10.1016/j.ijantimicag.2009.11.003.
6. Johnson JR, Urban C, Weissman SJ, Jorgensen JH, Lewis JS, Hansen G,Edelstein PH, Robicsek A, Cleary T, Adachi J, Paterson D, Quinn J,Hanson ND, Johnston BD, Clabots C, Kuskowski MA. 2012. Molecularepidemiological analysis of Escherichia coli sequence type ST131 (O25:H4) and blaCTX-M-15 among extended-spectrum �-lactamase-producing
FIG 2 The mean and SD of the log CFU/0.5 g of feces for Gram-negative anaerobic flora and Gram-positive aerobic flora, shown at day 1 and day 4 to comparethe impact of 3 days of treatment. The SD was 0 for bars where the SD is not shown.
Antibiotic Selection of E. coli ST131
October 2014 Volume 58 Number 10 aac.asm.org 6143
on Septem
ber 27, 2014 by guesthttp://aac.asm
.org/D
ownloaded from

E. coli from the United States, 2000 to 2009. Antimicrob. Agents Che-mother. 56:2364 –2370. http://dx.doi.org/10.1128/AAC.05824-11.
7. Dahbi G, Mora A, López C, Alonso MP, Mamani R, Marzoa J, Coira A,García-Garrote F, Pita JM, Velasco D, Herrera A, Viso S, Blanco JJE,Blanco M. 2013. Emergence of new variants of ST131 clonal group amongextraintestinal pathogenic Escherichia coli producing extended-spectrum�-lactamases. Int. J. Antimicrob. Agents 42:347–351. http://dx.doi.org/10.1016/j.ijantimicag.2013.06.017.
8. Tumbarello M, Spanu T, Di Bidino R, Marchetti M, Ruggeri M,Trecarichi EM, De Pascale G, Proli EM, Cauda R, Cicchetti A, Fadda G.2010. Costs of bloodstream infections caused by Escherichia coli and in-fluence of extended-spectrum beta-lactamase production and inadequateinitial antibiotic therapy. Antimicrob. Agents Chemother. 54:4085– 4091.http://dx.doi.org/10.1128/AAC.00143-10.
9. Pitout JDD. 2010. Infections with extended-spectrum beta-lactamase-producing Enterobacteriaceae: changing epidemiology and drug treat-ment choices. Drugs 70:313–333. http://dx.doi.org/10.2165/11533040-000000000-00000.
10. Rodríguez-Baño J, Alcala JC, Cisneros JM, Grill F, Oliver A, HorcajadaJP, Tortola T, Mirelis B, Navarro G, Cuenca M, Peña C, Llanos AC,Canton R, Pascual A. 2008. Community infections caused by extended-spectrum beta-lactamase-producing. Arch. Intern. Med. 168:1897–1902.http://dx.doi.org/10.1001/archinte.168.17.1897.
11. Nielsen KL, Dynesen P, Larsen P, Frimodt-Møller N. 2014. FaecalEscherichia coli from patients with E. coli urinary tract infection andhealthy controls who have never had a urinary tract infection. J. Med.Microbiol. 63:582–589. http://dx.doi.org/10.1099/jmm.0.068783-0.
12. Rodríguez-Baño J, Navarro MD, Romero L, Muniain MA, de Cueto M,Rios MJ, Hernandez JR, Pascual A. 2006. Bacteremia due to extended-spectrum beta-lactamases producing Escherichia coli in the CTX-M era: anew clinical challenge. Clin. Infect. Dis 43:1407–1414. http://dx.doi.org/10.1086/508877.
13. Lautenbach E, Patel JB, Bilker WB, Edelstein PH, Fishman NO. 2001.Extended-spectrum beta-lactamase-producing Escherichia coli and Kleb-siella pneumoniae: risk factors for infection and impact of resistance onoutcomes. Clin. Infect. Dis. 32:1162–1171. http://dx.doi.org/10.1086/319757.
14. Bellíssimo-Rodrigues, F, Carolina A, Gomes F, Dinis A, Passos C,Achcar JA, Castro S, Martinez R, Gomes ACF, Passas ADC, PerdonáGDSC. 2006. Clinical outcome and risk factors related to extended-spectrum beta-lactamase-producing Klebsiella spp. infection among hos-pitalized patients. Mem. Inst. Oswaldo Cruz 101:415– 421. http://dx.doi.org/10.1590/S0074-02762006000600012.
15. Hansen F, Olsen SS, Heltberg O, Justesen US, Fuglsang-Damgaard D,Knudsen JD, Hammerum AM. 2014. Characterization of third-generation cephalosporin-resistant Escherichia coli from bloodstream in-fections in Denmark. Microb. Drug Resist. 20:316 –3240. http://dx.doi.org/10.1089/mdr.2013.0157.
16. Clermont O, Dhanji H, Upton M, Gibreel T, Fox A, Boyd D, MulveyMR, Nordmann P, Ruppé E, Sarthou JL, Frank T, Vimont S, Arlet G,Branger C, Woodford N, Denamur E. 2009. Rapid detection of theO25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 64:274 –277. http://dx.doi.org/10.1093/jac/dkp194.
17. Nielsen JB, Albayati A, Jørgensen RL, Hansen KH, Lundgren B, Schøn-ning K. 2013. An abbreviated MLVA identifies Escherichia coli ST131 asthe major extended-spectrum �-lactamase-producing lineage in the Co-penhagen area. Eur. J. Clin. Microbiol. Infect. Dis. 32:431– 436. http://dx.doi.org/10.1007/s10096-012-1764-x.
18. Imhof A. 1996. Continuous monitoring of oxygen concentrations in sev-eral systems for cultivation of anaerobic bacteria. J. Clin. Microbiol. 34:1646 –1648.
19. Perez F, Pultz MJ, Endimiani A, Bonomo RA, Donskey CJ. 2011. Effectof antibiotic treatment on establishment and elimination of intestinal col-onization by KPC-producing Klebsiella pneumoniae in mice. Antimicrob.Agents Chemother. 55:2585–2589. http://dx.doi.org/10.1128/AAC.00891-10.
20. Erlendsdottir H, Knudsen JD, Odenholt I, Cars O, Espersen F, Fri-modt-møller N, Fuursted K, Kristinsson KG, Gudmundsson S. 2001.Penicillin pharmacodynamics in four experimental pneumococcal infec-tion models. Antimicrob. Agents Chemother. 45:1078 –1085. http://dx.doi.org/10.1128/AAC.45.4.1078-1085.2001.
21. Knudsen JD, Fuursted K, Frimodt-Møller N, Espersen F. 1997. Com-parison of the effect of cefepime with four cephalosporins against pneu-mococci with various susceptibilities to penicillin, in vitro and in themouse peritonitis model. J. Antimicrob. Chemother. 40:679 – 686. http://dx.doi.org/10.1093/jac/40.5.679.
22. Jakobsen L, Cattoir V, Jensen KS, Hammerum AM, Nordmann P, Fri-modt-Møller N. 2012. Impact of low-level fluoroquinolone resistance genesqnrA1, qnrB19 and qnrS1 on ciprofloxacin treatment of isogenic Escherichiacoli strains in a murine urinary tract infection model. J. Antimicrob. Che-mother. 67:2438–2444. http://dx.doi.org/10.1093/jac/dks224.
23. Sandberg A, Jensen KS, Baudoux P, Van Bambeke F, Tulkens PM,Frimodt-Møller N. 2010. Intra- and extracellular activities of dicloxacillinagainst Staphylococcus aureus in vivo and in vitro. Antimicrob. AgentsChemother. 54:2391–2400. http://dx.doi.org/10.1128/AAC.01400-09.
24. Kerrn MB, Frimodt-Møller N, Espersen F. 2003. Effects of sulfamethi-zole and amdinocillin against Escherichia coli strains (with various sus-ceptibilities) in an ascending urinary tract infection mouse model. Anti-microb. Agents Chemother. 47:1002–1009. http://dx.doi.org/10.1128/AAC.47.3.1002-1009.2003.
25. Asahi Y, Miyazaki S, Yamaguchi K. 1995. In vitro and in vivo antibac-terial activities of BO-2727, a new carbapenem. Antimicrob. Agents Che-mother. 39:1030 –1037. http://dx.doi.org/10.1128/AAC.39.5.1030.
26. Frimodt-Møller N, Thomsen V. 1987. The pneumococcus and the mouseprotection test: correlation of in vitro and in vivo activity for beta-lactamantibiotics, vancomycin, erythromycin and gentamicin. Acta Pathol. Mi-crobiol. Immunol. Scand. 95:159 –165.
27. Achá SJ, Kühn I, Mbazima G, Colque-Navarro P, Möllby R. 2005.Changes of viability and composition of the Escherichia coli flora in faecalsamples during long time storage. J. Microbiol. Methods 63:229 –238.http://dx.doi.org/10.1016/j.mimet.2005.04.024.
28. Haigh J, Degun A, Eydmann M, Millar M, Wilks M. 2011. Improvedperformance of bacterium and yeast identification by a commercial ma-trix-assisted laser desorption ionization-time of flight mass spectrometrysystem in the clinical microbiology laboratory. J. Clin. Microbiol. 49:3441.http://dx.doi.org/10.1128/JCM.00576-11.
29. Frimodt-moller N, Ostri P, Pedersen IBK, Poulsen SR. 1980. Antibioticprophylaxis in pulmonary surgery: a double-blind study of penicillin ver-sus placebo. Ann. Surg. 195:444 – 450.
30. Wilson KH, Brown RS, Andersen GL, Tsang J, Sartor B. 2006. Com-parison of fecal biota from specific pathogen free and feral mice. Anaerobe12:249 –253. http://dx.doi.org/10.1016/j.anaerobe.2006.09.002.
31. Krych L, Hansen CHF, Hansen AK, van den Berg FWJ, Nielsen DS.2013. Quantitatively different, yet qualitatively alike: a meta-analysis ofthe mouse core gut microbiome with a view towards the human gut mi-crobiome. PLoS One 8:e62578. http://dx.doi.org/10.1371/journal.pone.0062578.
Boetius Hertz et al.
6144 aac.asm.org Antimicrobial Agents and Chemotherapy
on Septem
ber 27, 2014 by guesthttp://aac.asm
.org/D
ownloaded from

“Epidemiological factors associated with ESBL- or non ESBL-producing E.coli causing UTI in
general practices.”
Keywords: UTI; Escherichia coli; Risk Factors; Community-acquired; ESBL.
Running title: Risk factors for E.coli UTI in general practices
(1,2) Frederik Boëtius Hertz, (1) Kristian Schønning, (1) Steen Christian Rasmussen (1), Pia
Littauer, (1) Jenny Dahl Knudsen, (2) Anders Løbner-Olesen and (1) Niels Frimodt-Møller
1) Dept. of Clinical Microbiology, Hvidovre University Hospital, Hvidovre, Denmark.
2) Dept. of Biology, University of Copenhagen, Denmark.
Corresponding author:
Frederik B. Hertz, MD
Hvidovre University Hospital
Dept. of Clinical Microbiology
Copenhagen
004538626971

Abstract
Background: The frequency of community-onset urinary tract infections (UTI), caused by ESBL-
producing E.coli is increasing. This demands general practitioners to rapidly identify patients at
risk. Epidemiological differences between UTIs caused by ESBL-producing, resistant (non-ESBL)
and susceptible E.coli have never been thoroughly investigated.
Methods: This was a triple-case-control study with three case groups and one group of uninfected
controls, all from general practice. We performed prospective collection of urine samples at Dept.
of Clinical Microbiology at Hvidovre Hospital, Denmark, among samples from general practices
from October 2011 to July 2012 followed by a retrospective analysis of epidemiological
characteristics. All included cases had a urine sample positive for E.coli > 1000 CFU/ml.
Results: We included 98 cases with UTI caused by ESBL-producing E.coli, 174 patients with
antibiotic-resistant (non-ESBL) E.coli and 177 patients with susceptible E.coli. These were
individually compared to a group of 200 individuals with negative urine samples. Case groups had a
significantly higher use of antibiotics than the control group within 30 days prior to infection
(P<0.0001). Prior to infection the ESBL group had significantly more hospital admissions, higher
use of antibiotics and more urinary tract infections with cefpodoxime resistant E.coli, than the other
case groups (P<0.05).
Conclusion: We found previous exposure to antibiotics to be a risk factor for UTI with E.coli in
general. Antibiotics did not prove to be indisputable predictors of ESBL infection. Healthcare
contact and prior UTI with ESBL-producing E.coli within 90 days of infection were independent
risk factors for community onset UTI by ESBL-producing E.coli.

Introduction
Urinary tract infection (UTI) is one of the most common bacterial infections in hospital settings and in the
community and Escherichia coli is the most common etiology (1,2). E.coli is additionally becoming
increasingly resistant, in part due to the presence of plasmid-borne Extended-Spectrum Beta-Lactamases
(ESBL) (2–4). Plasmids with genes coding for co-resistance to beta-lactams, fluoroquinolones,
aminoglycosides and sulphonamides are now a common observation, leaving the isolates multi-drug
resistant (3,5). Community-acquired UTI is the infection most often caused by ESBL-producing E.coli
and community-onset infections (COI) represent a major clinical challenge (5–8). Furthermore, resistant
E.coli are able to colonize the gut of humans for long periods of time, and current studies report increased
carriage of ESBL-producing E.coli among all age groups (9–12). Numerous studies have consequently
investigated epidemiological characteristics of patients infected with ESBL-producing E.coli, reporting
frequently identified risk factors such as age > 65, male sex, recent use of antibiotics (in particular
cephalosporins and fluoroquinolones), recent hospitalization and the presence of underlying disease
(3,5,6,13–15). Previous colonization with ESBL-producing E.coli, the presence of an urinary catheter and
previous UTI were likewise independent risk factors (14–16). Finally, recent publications have identified
travel to high-prevalence countries and lifestyle to be involved in colonization and infection (10,13,15).
Antibiotic resistance and subsequent inappropriate empirical therapy are associated with increased
mortality, a higher use of broad-spectrum antibiotics as well as increased healthcare costs (6,8,14). The
described risk factors apply to both healthcare and community associated infections, but ESBL -
producing E.coli among patients with no recent healthcare exposure are emerging (5). Uncomplicated
infections with a community-onset are often diagnosed and treated empirically at general practices
without preceding susceptibility tests. There is therefore an essential need to rapidly identify patients at
risk to avoid the continuous selection of resistant strains and inappropriate antibiotic therapy (5,17). The
association between use of antibiotics and infections caused by ESBL-producing E.coli from primary care

has been sparsely investigated in low-ESBL prevalence countries. A triple-case-control study focusing on
E.coli has never been performed, and very few case-control studies have included an uninfected group
(15,18,19). In the 2012 Danish population, 4% and 6% of E.coli from urine samples from general practice
and hospitals were resistant to extended-spectrum cephalosporins, respectively. In Denmark, we have
comprehensive nationwide databases on hospital contacts and prescription of medicine from general
practices. Thus, Denmark is an appropriate site for investigation of epidemiological factors associated
exclusively with non-hospitalized patients with E.coli UTI. Our purpose with this triple-case-control
study was therefore to evaluate the risk factors, antibiotics in particular, for contracting an UTI with an
ESBL-producing E.coli compared to less resistant and fully susceptible E.coli.
Material and methods
Research ethical approvals
This study was approved by the Danish Health and Medicines Authority, Statistics Denmark (DST),
The Regional Committee of Danish Data Protection Agency and patient notification was declared
not necessary by the Regional Committee of Health Research Ethics Committee (HVH-2012-017,
J.nr. 3-3013-230/1/KWH and H-4-2012-088).
Study design and population
Three parallel retrospective case-control analyses were carried out as a triple-case-control study at
Department of Clinical Microbiology at Hvidovre University Hospital in Copenhagen, Denmark
(DCM). This is an urban area with approximately 950.000 inhabitants and DCM serves more than
450 general practitioners. Three different case groups and one control group were included. All
three case groups were individually compared to the control group. All groups consisted of a unique
collection since a patient could only be included in one group and only once. The study period was
from the 1st of October 2011 to the 30th of June 2012. Patients, including controls, were eligible for

inclusion if a urine sample had been submitted in the given time period, disregarding age, gender
and previous ESBL-positive samples. Exclusion criteria were no Danish social security (CPR)
number meaning no available records on prescriptions or hospital contacts. No extended matching
of controls was performed.
Collection of subjects was conducted at the laboratory at DCM and samples were collected only
among urine specimens from general practices. Case groups were all patients with a urine sample
positive for E.coli > 1000 CFU/ml and the control group consisted of patients, randomly collected,
with a urine sample without bacterial growth (referred to as uninfected) during the study period.
The uninfected group was therefore represented by patient data only, with no existing
microbiological analysis. Case group 1 was the group with ESBL-producing E.coli (referred to as
ESBL), case group 2 consisted of patients in which the E.coli was resistant to at least one of the
tested antimicrobials (ampicillin, cefuroxime, aztreonam, ampicillin/clavulanic acid,
piperacillin/tazobactam, mecillinam, ceftazidime, meropenem, ciprofloxacin, sulfamethoxazole,
trimethoprim, tetracycline, gentamicin, tobramycin, nitrofurantoin, fosfomycin,) but without an
ESBL phenotype (referred to as resistant) and case group 3 comprised patients with UTI caus
Patient variables and definitions
The following variables were assessed from Statistics Denmark (DST) and DCM records one year
prior to until 90 days after index sampling: Age, sex, hospital admission, prescriptions of
medication, reported use of urinary catheter and number of urine samples submitted to DCM for
investigation. Urine samples were not included in the multiple statistical analyses, since data from
DST are available by extern access only, with no possible merging of DCM and DST data.
Retrospectively, UTI was defined as either complicated or uncomplicated by previously established
definitions: An uncomplicated UTI occurs in patients with no structural or functional abnormalities,

are not pregnant or who have not been submitted to instrumented procedures (as catheterization)
(2). Furthermore, UTI was categorized as either true community onset infections (COI) or
healthcare associated (HCAI), by modified Friedman criteria (23): HCAI was defined from a
sample obtained from a patient with at least one of the two criteria: (і) Patient admitted to a hospital
>2 days within 3 months prior to urine sampling or (іі) patient who initiated hospital associated
outpatient clinic contact within 30 days prior to index sample (obtained from DST for this
classification). Prescription of medicine was not available from hospital settings and data included
community-associated prescriptions only. On the contrary, diagnoses were just obtainable from
hospital settings.
Microbiological analysis
All microbiological analyses were performed at DCM.
Susceptibility patterns
Bacterial susceptibility testing was performed by either Antibiotic Disc Diffusion Test
Methodology or by The Vitek 2 automated system (Cards AST-N209 and AST-N122) (24,25). Disc
Diffusion was performed using the Disk Diffusion Test Methodology (Version 1.0, December 18,
2009) as described in the European Committee on Antimicrobial Susceptibility Testing (EUCAST))
(26). Vitek 2 testing was performed according to guidelines of DCM and in accordance with
direction from the manufacturer (bioMérieux) and as described elsewhere (24,25). For Vitek 2,
isolates found resistant to mecillinam were tested by disk diffusion or Etest to confirm resistance.
An ATCC 25922 E.coli was used for quality control for all methods and results interpreted
according to recommendations from EUCAST (www.eucast.org/clinical_breakpoints/). All strain

found as I (intermediately resistant) and R were reported as resistant and strains found as S were
reported as susceptible.
ESBL-producing strains were phenotypically identified by the MAST-test, performed as a
combined-disk method, using disk containing Cefpodoxime ±ESBL and/or AmpC inhibitors
(MAST®, Merseyside, UK). A difference in zone of inhibition >5mm between disks with and
without inhibitor combinations was considered a positive phenotypic result as previously described
(26,27).
Molecular tests
Phylogroup testing
All isolates were classified according to phylogroups A, B1, B2, D or Non-typeable, respectively,
by singleplex PCR amplification of three genes as previously described and isolates were tested for
the presence of ibeA when necessary (28,29). The method was chosen albeit a recently published
paper indicates that some isolates are possibly assigned incorrect phylogroups (30).
ESBL genotype identification
As previously described, identification of ESBL genotype, on ESBL-producing isolates, was done
by screening for the presence of blaCTX-M by PCR with subsequent identification of specific CTX-M
gene by sequencing (23,31).
Statistical analysis
The triple-case-control study was analysed using a conservative two step evaluation. We performed
an initial univariate t-test with a no-hypothesis strategy. We left out variables with no immediate
relevance for the study and included all medications prescribed, admissions to hospitals and hospital
diagnoses. We divided data into time intervals as 90 and 30 days prior to urine sampling. Variables

with P<0.1, as found by the univariate analysis, were subsequently included in the multivariate
logistic-regression models to identify individual risk factors. Variables were included in the time
period where P<0.1 and similar variables were tested for all three case groups. A second run was
performed as a simplified manual backward stepwise elimination procedure including variables
with P<0.1 in the multivariate tests only. Finally, the multivariate analyses were repeated separately
comparing the ESBL case group to the other case groups. Outcome was set as the ESBL group.
Odds ratios were estimated with the reservations needed for the study design and are therefore not
shown (33). Hosmer and Lemeshow Goodness of Fit test was evaluated for all multivariate
analyses. When appropriate, chi-square or Fisher´s exact tests were used. All analyses were
performed using SAS version 9.2 and 9.3 (SAS Institute, Cary, NC) and P<0.05 was considered
significant. Results from the initial univariate t-tests are not shown, but appropriate p-values from
chi-square and Fisher´s exact test are presented in written text and appropriate tables.
We did not include COI and HCAI in the multivariate statistical analysis. We simply assessed the
number of patients in each group rather than to compare HCAI with COI (34).
Results
Patient characteristics and multivariate analysis
In the study period we included 98 patients to the ESBL case group, 174 patients to the resistant
groups, 177 to the susceptible group and 200 patients to the uninfected group. All collected E.coli
isolates were included in the molecular analysis but a few patients were excluded during analysis of
data from DST, due to a missing date of birth. We excluded two patients from the ESBL group,
three from the resistant group and two from the susceptible group. No patients were excluded from
the uninfected group. Relevant data on patient characteristics and use of antibiotics are shown in
Table 1. Results of the multivariate analysis are found in Table 2 and 3. We found that patients in

the three case groups were significantly older than among the uninfected controls (P<0.0001).
Conversely, there was no significant difference in age between the case groups (P>0.05).
Furthermore, there were more men among the uninfected group than in the resistant and the
susceptible groups, but no significant difference among the case groups. When assessed as part of
the multivariate analysis, there was a significant difference between both gender and age for all case
groups compared to the uninfected group (P<0.005 and P<0.0001).
Risk factors for case groups compared to the uninfected group
Results of the multivariate analyses are found in Table 2. Data were analysed during two periods,
i.e. 90 and 30 days prior to sampling. During 90 days prior to sampling, few antibiotics and
diagnoses were found as independent risk factors. In the ESBL group only use of mecillinam in the
backward elimination analysis was found to be significant (P=0.0426). The use of five and two
specific antibiotics were independent risk factors for the resistant and susceptible groups,
respectively (all P<0.05). In the period of 30 days prior to index sample, prescription of antibiotics
in general was significant in all case groups, when analysed by chi-square (P<0.0001), but only
mecillinam (P<0.0001), mecillinam and trimethoprim (P<0.0001 and P=0.0249), mecillinam and
sulphamethizole (P<0.0001 and P=0.0205) were independent risk factors in the two resistant groups
and the susceptible group, respectively. The diagnoses “kidney infections” and “urosepsis” were
risk factors for the ESBL group (P=0.0106 and P=0.0334). There was no significant difference in
number of community- and healthcare-associated infections, (P>0.05, data not shown).
Risk factors for the ESBL case group compared to the other case groups
These results are illustrated in Table 3. There were significantly more HCAI patients in the ESBL
group than in the susceptible group (P=0.0058) as calculated by the chi-square test. Looking at

number of patients receiving antibiotics and prescriptions made, it was not evident that the ESBL
group had a higher use of antibiotics 90 days before index sample (Table 1). Analysis of the time
periods and the multivariate analyses performed in two runs, showed that use of antibiotics in
general was a risk factor for the ESBL group 90 days prior to index sample (P=0.0049 and
P=0.0029) and hospital admission 30 days prior was an independent risk factor for ESBL
(P=0.0073 and P=0.0216). The use of trimethoprim and sulphamethizole, in percentages, were
higher or equivalent in the resistant and the susceptible groups compared to the ESBL group (Table
1). There was a significant difference in use of trimethoprim (P<0.0298) and sulphamethizole
(P<0.0381) between the ESBL and the resistant groups and in use of sulphamethizole between the
ESBL group and the susceptible group (P=0.0299). Only with the manual backward selection
model, did use of quinolones prove significantly higher in the ESBL group 30 days prior to
sampling, compared to the resistant group (P=0.0365) and total use of antibiotics comparing the
ESBL and susceptible groups (P=0.04).
Urine Samples to Dept. of Clinical Microbiology
The sum of positive urines sample submitted from general practice to DCM as related to the
different patient case groups, as shown in Table 4, were analysed by Fishers exact test. P-values are
listed in the text where appropriate.
Overall, significantly more urine samples were submitted from patients in the ESBL and resistant
groups than from patients in the uninfected group during one year prior as well as during 90 days
before the index sample (P<0.05 for all). All case groups had significantly more positive urine
samples and significantly more samples positive for E.coli than the control group, 90 days before
and after index sampling (P<0.0001 for all). The ESBL group had significantly more urine samples
positive for CDP-R E.coli than the other three groups (P<0.0001) before and after the index sample.

Compared to the period prior to the index sample, all three case groups had significantly fewer
positive urine samples during the 90 days after the index sample and the ESBL group had
significantly less urine samples positive for cefpodoxime resistant E.coli (P<0.0001) with no
significant changes in the other case groups.
Antibiotic susceptibility, phylogroups and ESBL genotypes
The ESBL-producing E.coli isolates displayed increased resistance to most of the tested antibiotics
except towards mecillinam (2%), nitrofurantoin (7%), fosfomycin (3%) and piperacillin/tazobactam
(4%). Regarding prescriptions 90 days prior to index sample (Table 1), mecillinam was given to
55% of patients, nitrofurantoin to 14% and sulphamethizole to 19%, respectively.
E.coli from the resistant group showed lower rates of resistance, than the ESBL group, towards
most antibiotics (1%-15%) except towards trimethoprim (46%), sulphamethizole (68%),
tetracycline (47%), ampicillin (84%) and to some extent towards ciprofloxacin (22%), with low
resistance to towards mecillinam (3%), nitrofurantoin (5%), fosfomycin (3%) and
piperacillin/tazobactam (3%). In this group 58% received mecillinam and 13% nitrofurantoin,
respectively with 27% receiving sulphamethizole.
Distributions of phylogroups were very similar among the three case groups with phylogroup B2
dominating and group D being the second largest group: Phylogroup A , 7-10%; phylogroup B1, 1-
6%; phylogroup B2 , 45-58%; phylogroup D ,22-32%; and non-typeable, 4-13%, respectively.
All 98 ESBL-producing E.coli were investigated for the presence of an ESBL gene. A total of 98%
carried a blaCTX-M type ESBL and 2% did not have an ESBL gene identified. blaCTX-M group 1
represented 73% of all isolates with 54% of all isolates carrying CTX-M-15. Furthermore, 24%
belonged to blaCTX-M group 9 with two clusters of CTX-M-14 (12%) and CTX-M-27 (12%).

Discussion
To our knowledge, this is the first triple-case-control study of epidemiological characteristics for
UTI caused by E.coli. It is moreover the first study to exclusively include non-hospitalized patients
of all ages in a country with low ESBL prevalence. As reported from previous, mainly hospital
based, investigations, we found healthcare contact and previous UTI with ESBL-producing E.coli to
be associated with community-onset UTI with ESBL-producing E. coli (3,5,6,13–15,35). However,
we have also exposed some new and interesting aspects of the epidemiology of UTI caused by
E.coli. First of all, surprisingly few differences were found between the UTI case groups, when
these were compared to an uninfected control group and here no clear difference was found
regarding exposure to antibiotics as risk factors for ESBL-producing E.coli. We did find our case
groups to be of older age than the control group, but there was no significant difference in age or
gender among the case groups, even though the ESBL group did have a higher average age and a
higher number of men, respectively. In Norway, another low prevalence country, the ESBL
population was found to be younger but they also found travel to be a risk factor, indicating that
other features than healthcare contact has an influence for infections with ESBL-producing
Enterobacteriaceae in younger individuals (8,36). In our study, we found no significant difference
(P>0.05) between the percentages of HCAI ESBL infections in the age group 18-64 and >65,
respectively (Table 1). Travel history was not studied.
Regarding the prescriptions of antibiotics, the case groups had a significantly higher use before
index sampling than in the uninfected control group. Interestingly, use of mecillinam posed a risk
factor for UTI for all case groups within 90 days of infection, which is in contradiction to what has
been seen in Norway where mecillinam was found not to be associated with community-onset
ESBL infections (8). The Norwegian study did however not include an uninfected control group
and we did not find mecillinam to be associated with ESBL infection when compared to the other

case groups. Mecillinam is widely used as a first-line drug for treatment of UTI in Denmark and
considered to be an effective agent, also against most ESBL-producing E.coli, with an inhibiting
effect on E.coli in the intestines (37,38). It was previously seen that combination drugs of beta-
lactams/beta-lactamase-inhibitors were a risk factor for ESBL infection, but it remains unclear why
antibiotics with an effect on most resistant E.coli, including ESBL, seems to select for these
uropathogens (16). The healthcare aspect of hospital admissions as a risk factor became evident
when comparing the case groups. Here, we found the ESBL group to have significantly more
healthcare associated infection compared to the group of susceptible E.coli, and significantly more
admissions than both of the other case groups. This confirms hospital contact as an independent risk
factor for ESBL-producing E.coli infections as has been establish previously (3,5,6,13–15). Total
use of antibiotics was significantly higher in the ESBL group. However, exposure to antibiotics was
not unmistakeably higher among patients from the ESBL group, with varied antibiotics being used
in the groups (Table 1 and 3).
We found that the ESBL group had significantly more urine samples being positive for growth of
CPD-R E.coli. This was the case one year prior, 90 days prior and also 90 days after index
sampling, clearly showing that previous UTI plays an important role when assessing patients at risk
(14–16).
We chose to study epidemiological characteristics of ESBL-producing E.coli as a triple-case-control
study in accordance with what was previously suggested for investigations of risk factors (19,33).
When designing such a study, it is of great importance that the design includes an uninfected
control group, especially to reduce the risk of overestimating the use of antibiotics (19,33).
Furthermore, the control group should represent the background population from which all
individuals are included (19,33). As we chose a control group among individuals with a negative
urine culture, there is a clear risk of selection bias, as this group seeks medical attention and might

not represent the true background population. Our triple-case-control design is therefore not suited
for the investigation of possible infection control. However, the design can address the issue of
recommendations for empiric treatment when susceptibility patterns of the suspected pathogen are
unknown (19). Other limitations of our study are the lack of data on nursing home residence and
outpatient contacts, which will possibly lead us to underestimate the significance of healthcare
exposures and make it difficult to identify possible reservoirs for ESBL-producing E.coli. It would
also have been informative to include diagnoses from general practices as well as medications from
hospitals to give us the full overview of illnesses and treatments. Since we only included patients
who had urine samples submitted to the laboratory, we have not been able to include patients
treated empirically with no sampling or patients treated by general practitioners performing in-
house susceptibility testing. Finally, as we have had no direct contact to the included individuals,
we have not been able to investigate issues like travel and lifestyle. Furthermore, as all case groups
have more positive urine samples submitted to DCM, with no matching of patients based on UTI-
free periods, there is a risk that we are actually studying three case-groups of patients with recurrent
UTI, and this would pose an obvious reason for antibiotic prescriptions. However, our data on
hospital diagnoses argues against this bias.
Nonetheless, ESBL-producing E.coli are becoming increasingly frequent as community-acquired
infections and underestimating the risk of ESBL-producing E.coli may lead physicians to treat
patients with inappropriate empirical regimens. This may lead to selection of ESBL-positive
isolates, treatment failure and risk of severe morbidity. The selection pressure can be enforced by
several different antibiotics. Overestimating the risk of ESBL-producing E.coli can lead to the use
of broad-spectrum antibiotics leading to a higher selection pressure and the emergence of other
resistant strains (16). Therefore, to fight the spread of ESBL-producing E.coli, the physicians need
to rapidly identify patients infected with resistant E.coli and treat them accordingly. The use of

drugs with potentially good effect on ESBL should lower the risk of selection of ESBL-producing
E.coli. Nevertheless, our findings and previous studies indicate that this is not enough to avoid an
increase in ESBL prevalence, even if urine samples with CPD-R E.coli did significantly decrease
after treatment. We found mecillinam to be an independent risk factor for all case groups (16). This
could be as a result of selection of present ESBL-producing E. coli due to inefficient gut
concentrations, thereby maintaining ESBL in the faeces, or due to constant re-exposure to resistant
pathogens from an unknown reservoir (9). This reservoir could be nursing homes and long-term
care facilities, as previously stated (39) and which is not possible for us to determine. The reservoir
could also be hospital settings suggested by fact that the number of HCAI and hospital admissions
seemed to be strictly linked to ESBL infection – at least within 90 days of UTI. We did not find
admissions within one year prior to infection to be a risk factor, which corresponds to a recent
publication describing six months as the duration that healthcare exposure increases the risk of a
ESBL infection (15). We speculate if the hospital use of antibiotics with broad spectra select for
ESBL-producing E.coli in healthcare settings while use of oral antibiotics in the community select
for resistant, non-ESBL-producing E.coli.
In conclusion, we found similar risk factors for ESBL-producing E.coli infections in non-
hospitalized patients as previously proposed. However, in our study exposure to antibiotics proved
to be a problematic variable to use as guidance for general practitioners when assessing patients. All
in all, patients at risk of UTI caused by E.coli are likely to have had a UTI within 90 days and to
have received antibiotics. Nonetheless, only previous UTI by ESBL-producing E.coli and
healthcare exposure including hospital admission within 90 days are true risk factors for UTI caused
by ESBL-producing E.coli.

Acknowledgements
Work has been performed as cooperation between HVH, Roskilde University and University of
Copenhagen with laboratory facilities provided by HVH. This work was, in parts, presented at ECCMID
in Berlin 2013 and at 53rd Interscience Conference on Antimicrobial Agents and Chemotherapy, Denver,
CO, 9th-13th September 2013.
Funding
This work was supported by the EU-FP funded project PAR7, DanCARD (Danish National
Strategic Research Foundation, project 09-067075/DSF), Roskilde University, University of
Copenhagen and the SSAC Foundation (SLS-327421 and SLS-251761).
Transparency declarations
None to declare.
1. Kahlmeter G, Poulsen HO. Antimicrobial susceptibility of Escherichia coli from community-
acquired urinary tract infections in Europe: the ECO·SENS study revisited. Int J Antimicrob
Agents. Elsevier B.V.; 2012 Jan [cited 2012 Nov 27];39(1):45–51.
2. Foxman B. The epidemiology of urinary tract infection. Nat Rev Urol [Internet]. Nature
Publishing Group; 2010 Dec;7(12):653–60.
3. Doi Y, Park YS, Rivera JI et al. Community-associated extended-spectrum β-lactamase-
producing Escherichia coli infection in the United States. Clin Infect Dis. 2013 Mar.

4. Livermore DM. Fourteen years in resistance. Int J Antimicrob Agents. Elsevier B.V.; 2012
Apr [cited 2014 Mar 29];39(4):283–94.
5. Ben-ami R, Arslan H, Pitout JDD et al. A Multinational Survey of Risk Factors for Infection
with Extended-Spectrum b -Lactamase – Producing Enterobacteriaceae in Nonhospitalized
Patients. 2009;49.
6. Hayakawa K, Gattu S, Marchaim D et al. Epidemiology and Risk Factors for Isolation of
Escherichia coli Producing CTX-M-Type Extended-Spectrum β-Lactamase in a Large U.S.
Medical Center. Antimicrob Agents Chemother. 2013 Aug;57(8):4010–8.
7. Peirano G, van der Bij AK, Gregson DB et al. Molecular epidemiology over an 11-year
period (2000 to 2010) of extended-spectrum β-lactamase-producing Escherichia coli causing
bacteremia in a centralized Canadian region. Journal of clinical microbiology. 2012. p. 294–
9.
8. Søraas A, Sundsfjord AS, Sandven I et al. Risk Factors for Community-Acquired Urinary
Tract Infections Caused by ESBL-Producing Enterobacteriaceae -A Case-Control Study in a
Low Prevalence Country. PLoS One; 2013.
9. Ruppé E, Lixandru B, Cojocaru R et al. Relative Fecal Abundance of Extended-Spectrum-β-
Lactamase-Producing Escherichia coli Strains and Their Occurrence in Urinary Tract
Infections in Women. Antimicrob Agents Chemother. 2013 Sep;57(9):4512–7.
10. Jørgensen SB, Samuelsen O, Sundsfjord A et al. High prevalence of faecal carriage of ESBL-
producing Enterobacteriaceae in Norwegian patients with gastroenteritis. Scand J Infect Dis.
2014 Apr 4;(1478):462–5.

11. Fernández-Reyes M, Vicente D, Gomariz M et al. High Rate of Fecal Carriage of Extended-
Spectrum-β-Lactamase-Producing Escherichia coli in Healthy Children in Gipuzkoa,
Northern Spain. Antimicrob Agents Chemother. 2014 Mar;58(3):1822–4.
12. Dobrindt U, Hacker JH. Current Topics in Microbiology and Immunology Between
Pathogenicity and Commensalism.
13. Søraas A, Sundsfjord A, Sandven I et al. Risk factors for community-acquired urinary tract
infections caused by ESBL-producing enterobacteriaceae--a case-control study in a low
prevalence country. PLoS One. 2013 Jan;8(7):e69581.
14. Apisarnthanarak A, Kiratisin P, Saifon P et al. Clinical and molecular epidemiology of
community-onset, extended-spectrum beta-lactamase-producing Escherichia coli infections
in Thailand: a case-case-control study. Am J Infect Control. 2007 Nov;35(9):606–12.
15. Rogers BA, Ingram PR, Runnegar N et al. Community-Onset Escherichia coli Infection
Resistant to Expanded-Spectrum Cephalosporins in Low-Prevalence Countries. Antimicrob
Agents Chemother. 2014 Apr;58(4):2126–34.
16. Goulenok T, Ferroni A, Bille E et al. Risk factors for developing ESBL E. coli: can clinicians
predict infection in patients with prior colonization? J Hosp Infect. 2013 Aug;84(4):294–9.
17. Foxman B. Extended-spectrum β-lactamase-producing escherichia coli in the united states:
time to rethink empirical treatment for suspected E. coli infections? Clin Infect Dis. 2013
Mar;56(5):649–51.

18. Søraaas A, Sundsfjord A, Sandven I et al. Risk Factors for Community-Acquired Urinary
Tract Infections Caused by ESBL-Producing Enterobacteriaceae – A Case – Control Study in
a Low Prevalence Country. 2013;8(7):1–7.
19. Kaye KS, Harris AD, Samore M et al. The Case-Case-Control Study Design: Addressing The
Limitations of Risk Factor Studies for antimicrobial resistance. 2005;26(4):346–51.
20. Lausch KR, Fuursted K, Larsen CS et al. Colonisation with multi-resistant
Enterobacteriaceae in hospitalised Danish patients with a history of recent travel: A cross-
sectional study. Travel Med Infect Dis. Elsevier Ltd; 2013 Jun 27;4–7.
21. Hansen DS, Schumacher H, Hansen F et al., Stegger M, Hertz FB, Schønning K, et al.
Extended-spectrum β-lactamase (ESBL) in Danish clinical isolates of Escherichia coli and
Klebsiella pneumoniae: prevalence, β-lactamase distribution, phylogroups, and co-resistance.
Scand J Infect Dis. 2012 Mar;44(3):174–81.
22. DANMAP 2012. DANMAP. 2012;
23. Nielsen JB, Albayati A, Jørgensen RL et al. An abbreviated MLVA identifies Escherichia
coli ST131 as the major extended-spectrum β-lactamase-producing lineage in the
Copenhagen area. Eur J Clin Microbiol Infect Dis. 2013 Nov 6;32(3):431–6.
24. Livermore DM, Struelens M, Amorim J et al. et al. Multicentre evaluation of the VITEK 2
Advanced Expert System for interpretive reading of antimicrobial resistance tests. J
Antimicrob Chemother. 2002 Feb;49(2):289–300.
25. Nyberg SD, Meurman O, Jalava J et al. Evaluation of detection of extended-spectrum beta-
lactamases among Escherichia coli and Klebsiella spp. isolates by VITEK 2 AST-N029

compared to the agar dilution and disk diffusion methods. Scand J Infect Dis. 2008
Jan;40(5):355–62.
26. Hansen F, Olsen SS, Heltberg O et al. Characterization of Third-Generation Cephalosporin-
Resistant Escherichia coli from Bloodstream Infections in Denmark. Microb Drug Resist.
2014 Feb 11;00(00):1–9.
27. Hsieh C, Shen Y, Hwang K. Clinical Implications , Risk Factors and Mortality Following
Community-onset Bacteremia Caused by Extended-spectrum b-lactamase ( ESBL ) and non-
ESBL Producing Escherichia coli. J Microbiol Immunol Infect. Taiwan Society of
Microbiology; 2010;43(3):240–8.
28. Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli
phylogenetic group. Appl Environ Microbiol. 2000 Oct;66(10):4555–8.
29. Gordon DM, Clermont O, Tolley H et al. Assigning Escherichia coli strains to phylogenetic
groups: multi-locus sequence typing versus the PCR triplex method. Environ Microbiol.
2008 Oct;10(10):2484–96.
30. Clermont O, Christenson JK, Denamur E et al. The Clermont Escherichia coli phylo-typing
method revisited: improvement of specificity and detection of new phylo-groups. Environ
Microbiol Rep. 2013 Feb;5(1):58–65.
31. Jørgensen RL, Nielsen JB, Friis-Møller A et al. Prevalence and molecular characterization of
clinical isolates of Escherichia coli expressing an AmpC phenotype. J Antimicrob
Chemother. 2010 Mar;65(3):460–4.

32. Pérez-pérez FJ, Hanson ND. Detection of Plasmid-Mediated AmpC Beta -Lactamase Genes
in Clinical Isolates by Using Multiplex PCR. 2002;40(6):2153–62.
33. Harris AD, Karchmer TB, Carmeli Y et al. Methodological principles of case-control studies
that analyzed risk factors for antibiotic resistance: a systematic review. Clin Infect Dis. 2001
Apr 1;32(7):1055–61.
34. Marchaim D, Gottesman T, Schwartz O et al. National multicenter study of predictors and
outcomes of bacteremia upon hospital admission caused by Enterobacteriaceae producing
extended-spectrum beta-lactamases. Antimicrob Agents Chemother. 2010 Dec;54(12):5099–
104.
35. Lautenbach E, Bilker WB, Tolomeo P et al. Impact of diversity of colonizing strains on
strategies for sampling Escherichia coli from fecal specimens. J Clin Microbiol. 2008
Sep;46(9):3094–6.
36. Rogers B a, Sidjabat HE, Paterson DL. Escherichia coli O25b-ST131: a pandemic,
multiresistant, community-associated strain. J Antimicrob Chemother. 2011 Jan;66(1):1–14.
37. Jansåker F, Frimodt-Møller N, Sjögren I et al. Clinical and bacteriological effects of
pivmecillinam for ESBL-producing Escherichia coli or Klebsiella pneumoniae in urinary
tract infections. J Antimicrob Chemother. 2013 Oct 9;(October 2013):769–72.
38. Sullivan a, Edlund C, Svenungsson B et al. Effect of perorally administered pivmecillinam
on the normal oropharyngeal, intestinal and skin microflora. J Chemother. 2001
Jun;13(3):299–308.

39. Rooney PJ, O’Leary MC, Loughrey AC et al. Nursing homes as a reservoir of extended-
spectrum beta-lactamase (ESBL)-producing ciprofloxacin-resistant Escherichia coli. J
Antimicrob Chemother. 2009 Sep;64(3):635–41.

Table 1. Patient characteristics. Demographics of individuals included in the study and a list
of exposure to antibiotics shown for 90 days prior to index sample. The list is given as number
of patients receiving antibiotics and the total number of prescriptions. N* = Number of
patients. ▪Prescr. = Number of prescriptions.
Case groups
ESBL n=96 Resistant n=171 Susceptible n=175 Controls n=200
Demographics N* (%) N* (%) N* (%) N* (%)
Age (Mean), years 60.45 96(100) 57.95 171(100) 53,25 175(100) 42.10 200(100)
Age>18 (Mean), years 65.37 88(92) 58,5 169(99) 56,07 165(94) 46.16 180(90)
Age >65 (Mean), years 76.49 48(50) 79.09 85(50) 79.72 66(38) 76.49 37(19)
Male 18 (19) 17(10) 16(9) 54(27)
Female 78(81) 154(90) 159(91) 146(73)
Community-Onset-Infection (COI) 71(74) 139(81) 153(87) 163(82)
Health-Care-Associated-Infection (HCAI) 25(26) 32(19) 22(13) 37(19)
COI < 18 8(100) 1(50) 10(100) 18(90)
HCAI < 18 0 1(50) 0 2(10)
COI Age 18-64 30(75) 77(92) 90(91) 119(83)
HCAI Age 18-64 10(25) 7(8) 9(9) 24(17)
COI Age >65 33(69) 61(72) 53(80) 26(70)
HCAI Age >65 15(31) 24(28) 13(20) 11(30)
Complicated urinary tract infection 1(1) 1(0.6) 0
Prescriptions of antibiotics ESBL n=96 Resistant n=171 Susceptible n=175 Controls n=200
90 Days Prior N* (%) ▪Prescr. N*
(%)
▪Prescr. N* (%) ▪Prescr. N* (%) ▪Prescr.
Total antibiotics 78 (81) 199 148
(87)
329 137 (78) 231 92 (46) 164
Nitrofurantoin 13 (14) 21 22
(13)
38 16 (9) 18 8 (4) 8
Fluoroquinolones 11 (11) 13 7 (4) 9 6 (3) 7 12 (6) 13
Trimethoprim 4 (4) 8 20
(12)
31 4 (2) 4 4 (2) 4
Sulphonamides 18 (19) 22 47
(27)
58 39 (22) 47 31 (26) 31
Macrolides 6 (6) 10 5 (3) 6 5 (3) 6 6 (3) 8
Penicillins 13 (14) 15 19
(11)
21 13 (7) 14 9 (5) 10
Mecillinam 53 (55) 77 99
(58)
137 90 (51) 118 49 (25) 62
Penicillinase stable penicillins 3 (3) 5 2 (1) 3 1 (0.5) 1 4 (2) 5
Penicillins with effect on Gram-negative
(minus mecillinam)
56 (58) 86 105
(61)
153 98 (56) 130 53 (27) 75
Penicillins/beta-lactamase inhibitors 4 (4) 5 3 (2) 3 1 (0.5) 1 3 (2) 4

Table 2. Results of the Multivariate Logistic Regression analysis comparing case groups with the
uninfected group. First we performed the multivariate analysis based on results from the initial
student’s t-test followed by the manual backward stepwise elimination analysis. Finally, results of
statistical analyses on diagnoses are shown. All variables with a P<0.1 were included in both methods.
Empty squares mean estimates not computed by the statistic tool SAS. Only variables with P<0.05 are
shown. *P-values found by chi-square analyses. **P-values found by manual backward stepwise
elimination analysis.
ESBL, n=96
Resistant, n=171 Susceptible, n=175
Variables P-Value P-Value P-Value
30 Days Prior
Sex(Female) 0.0044 0.0004 0.0002
Age <.0001 <.0001 <.0001
Total antibiotics <.0001* <.0001* <.0001*
Mecillinam** <.0001** <.0001** <.0001**
Sulphamethizole 0.7379 0.0654 0.0205
Trimethoprim** 0.2616 0.0249** 0.9894
Kidney infection 0.0106* 0.9874 0.9939
Urosepsis 0.0334* . .
90 Days Prior
Mecillinam** <.0426** <.0001** 0.0001**
Nitrofurantoin 0.3871 0.0112 0.07
Penicillin** 0.1258 0.0216** 0.1048
Sulphonamides 0.9511 0.0091 0.0077
Trimethoprim 0.9848 0.0034 0.9873
Underlying diseases
90 Days Prior
Endocrine and Metabolic Diseases
0.0001 0.0005 0.5901
Diseases of the Circulatory System
<0.0001 <0.0001 0.1253
Diseases of the Respiratory System
<0.0001 0.0710 0.1919
Diseases of the Circulatory System
<0.0001 <0.0001 0.1253
Diseases of the Urinary/Reproductive
Organs <0.0001 0.0020 0.0034

Table 3. Results of the Multivariate Logistic Regression Analysis comparing the ESBL case
group with the two other case groups. These tests were performed as previously described for
case groups compared to the uninfected control group. Here we, in turn, compared the ESBL
group to each of the two other case groups. *P-values found by chi-square analyses. **P-
values found by manual backward stepwise elimination analysis.
ESBL vs. Resistant
ESBL vs. Susceptible
Variables P-Value P-Value
HCAI vs. COI 0.1660* 0.0058*
Sex (Female) 0.4335 0.4606
Age 0.4387 0.1837
30 Days Prior to index sample
Admissions 0.0073 0.0216
Trimethoprim 0.0298 0.9998
Sulphamethizole 0.0381 0.0299
Fluoroquinolones 0.0365** 0.25**
Total antibiotics 0.9194** 0.04**
90 Days Prior to index sample
Total antibiotics 0.0049 0.0029
Insulin 0.3218 0.0447
Nitrofurantoin 0.0089 0.1840
Sulphamethizole 0.0010 0.0091
Trimethoprim 0.0096 0.4663

Table 4. The ratio between patients and numbers of urine samples submitted to Dept. of
Clinical Microbiology from general practices from 365 days prior to and within 90 days after
sampling. The number of urine samples submitted within one year prior to sampling until 90
days after has been evaluated and counted for each individual patient. All isolates found in
urine are submitted to susceptibility testing towards cefpodoxime (CPD). Resistance to CPD is
used as an indication of ESBL production and leads to MAST-testing, results of which we do
not have. We use CPD-R as a possibility of ESBL-production, with the limitations it includes.
We show the ratio between total number of urine samples, number of positive samples, sum of
samples positive for E.coli and finally number of E.coli resistant to CPD divided to patients
included in each group. *P<0.05 compared to the uninfected group. ▪P<0.05 compared to the
resistant group. °P>0.05 compared to the susceptible group.
Case groups No. of urine samples
No. of positive samples (%)
No. of samples with E.coli (%)
No. of CPD Resistant E.coli (%)
ESBL (n=98) 3.8*° 3.5*° (91.2) 2* (56.3) 1.4*▪° (71.9)
Negative (n=200)
2 0.6 (33) 0.155 (24) 0.015 (9.7)
One year before index sample
Resistant (n=174)
3.4*° 3*° (90.6) 1.9* (64.1) 0.07*° (3.5)
Susceptible (n=177)
2.4 2.2* (91.5) 1.5* (68) 0
90-0 days before index sample
ESBL (n=98) 2* 1.9* (93.4) 1.3* (68.9) 1.1*▪° (57.4)
Negative (n=200)
1.5 0.4 (26.1) 0.11 (28.6) 0.02 (3.9)
Resistant (n=174)
2* 1.9* (96.2) 1.3* (69.8) 0.05° (2.4)
Susceptible (n=177)
1.7 1.6* (94.9) 1.2* (76.5) 0
0-90 days after index sample
ESBL (n=98) 1.4*° 1.2*° (83) 0.6*° (51.3) 0.5*▪° (80)
Negative (n=200)
0.4 0.2 (48.1) 0.05 (25.6) 0
Resistant (n=174)
1*° 0.9*° (88.8) 0.4*° (48.5) 0.02* (5.2)
Susceptible (n=177)
0.7* 0.6* (81.5) 0.3* (43.6) 0.01 (2.3)

1
“Population structure of Drug-Susceptible, -Resistant and ESBL-producing Escherichia coli
Populations from Community-Acquired Urinary Tract I nfections as Characterized by
Abbreviated MLVA, MLST and Serogrouping in Denmark”
Keywords: Escherichia coli; lineages; urinary tract infection; community-acquired; ESBL.
Running title: Characterization of E. coli according to susceptibility
(1,2) Frederik Boëtius Hertz, (1) Jesper Boye Nielsen, (1) Kristian Schønning (1) Pia Littauer, (1) Jenny Dahl
Knudsen, (2) Anders Løbner-Olesen and (3) Niels Frimodt-Møller
1) Dept. of Clinical Microbiology, Hvidovre Hospital, Denmark
2) Dept. of Biology, University of Copenhagen, Denmark
3) Dept. of Clinical Microbiology, Rigshospitalet, Denmark
Corresponding author:
Frederik B. Hertz, MD
Hvidovre University Hospital
Dept. of Clinical Microbiology
Copenhagen
004538626971

2
Abstract
Background: Escherichia coli is the most common cause of urinary tract infection in all age
groups. The pathogenic isolates are becoming increasingly resistant to antibiotics; with a worldwide
dissemination of resistant sequence types (ST). We characterized three different uropathogenic
E.coli (UPEC) populations, from non-hospitalized patients, by use of three typing methods, to
describe the genetic kinship between resistant and susceptible isolates.
Methods: We used MLST, a-MLVA, O-serogrouping and ESBL-genotyping. Urine samples
submitted for testing, by general practitioners, were identified at Dept. of Clinical Microbiology at
Hvidovre Hospital, Denmark, from Oct. 2011 to July 2012.
Results: We included 94 fully susceptible, 94 resistant (non-ESBL) and 98 ESBL-producing E.coli
isolates. The ESBL population was dominated by serogroup O25 constituting 39% of isolates. In
the ESBL population 51% belonged to ST131, 9% to ST38 and 6% to ST69. In the resistant group
ST69, ST73 and ST131 were the largest clusters with 18%, 11% and 15%, respectively. The
susceptible population had significantly more STs and a-MLVA codes than the other groups and
ST73 and ST95 were found as the only clusters with 16% and 6%, respectively. Ninety-eight per
cent of the ESBL-producing E.coli isolates were CTX-M-producers, 54% produced CTX-M-15.
Conclusion: Sequence type 131 dominated the population of community-associated uropathogenic
ESBL-producing E.coli, but was less frequent among non-ESBL-producing E.coli. The fully
susceptible E.coli population was a much more diverse group than the resistant and ESBL-
producing E.coli populations. Over findings suggest that dominant ESBL-producing lineages are
derived from UPEC lineages already established in the general population.

3
Introduction
Escherichia coli is the most common Gram-negative extraintestinal pathogen and the primary cause
of urinary tract infection (UTI) in all age groups (1,2). It is a highly heterogonous species with
certain lineages becoming increasingly resistant to antibiotics (2–5). Thus, resistant E.coli isolates
are emerging worldwide, including E.coli producing extended-spectrum beta-lactamase (ESBL),
now primarily producing the CTX-M type ESBLs (2,6). This dissemination of ESBL-producing
E.coli in hospital settings as well as in the community has been reported as spread of extraintestinal
pathogenic E.coli (ExPEC) belonging to a limited number of lineages or sequence types (STs) (2,6).
However, ExPEC lineages, like ST73 and ST95 continue to be common causes of primarily urinary
tract infection (UTI), but are rarely multidrug resistant and seldom associated with ESBL
production (7–10). There is a need to describe the prevalent lineages among non-ESBL-producing
E.coli to recognize if ESBL-producing E.coli lineages are identified in all populations or are
relatively newly emerged ExPEC (3,11). Different typing methods have previously sub-typed many
of the frequent E.coli ST-lineages and an abbreviated-MLVA (a-MLVA) method has been
evaluated and proved efficient in the characterization of ESBL-producing E.coli and the method is
congruent with multi-locus sequence typing (MLST) (12). However, only a few studies have
specifically studied the heterogeneity among susceptible, resistant and ESBL-producing E.coli
using several typing methods (2–4,6).
In the present study we characterize three different E.coli populations, all from non-hospitalized
patients, by use of O-serogrouping, a-MLVA and subsequent MLST of a-MLVA codes found.
MLST was performed to link a-MLVA codes to an international known nomenclature and we chose
urine samples from general practices to describe prevalence in an environment with reduced
selection pressure, compared to hospital settings. Hereby, we describe differences and kinship
among E.coli populations to assess if the spread of resistant E.coli are caused by successful

4
uropathogenic E.coli (UPEC) or other less successful UPEC with newly gained resistance (13).
Differences in dominating E.coli lineages among the populations would indicate that resistance and
ESBL-production generates selective advantages in certain bacterial populations and environments.
Materials and Methods
Strain collections
The Department of Clinical Microbiology at Hvidovre Hospital, Denmark (DCM) provides services
to more than 450 general practitioners in an area of approximately 950.000 inhabitants. From the 1st
of October 2011 until the 30th of June 2012 we collected 449 E.coli isolates from urine samples,
submitted from general practices, as part of an on-going case-control study. Hence, from non-
hospitalized patients, one unique strain was collected per patient and three susceptibility groups
were included; (i) ESBL-producing E.coli, (ii) E.coli resistant to at least one tested antibiotic (non-
ESBL) and (iii) fully susceptible E.coli. O-serogrouping was performed by GlycoVaxyn on the 94
ESBL-positive isolates as well as on 49 resistant and 51 susceptible isolates available at time of
serogrouping. From the complete case-control collections of isolates, we randomly chose isolates
for characterization by MLVA and MLST. Not all isolates or patients had been thoroughly
identified at the time of typing: First we included all isolates with a serogroup from GlycoVaxyn
and we included the remaining four ESBL-producing E.coli isolates. Secondly, we identified
resistant and susceptible isolates from patients with feasible community-onset infections (COI) and
randomly chose 45 and 43 isolates, respectively. Thus, in this study we included all 98 ESBL-
producing E.coli isolates, 94 isolates resistant (non-ESBL) isolates and 94 fully susceptible E.coli.

5
Strain identification and susceptibility testing
The UPEC isolates were all identified at species level by MALDI-TOF MS (Bruker, Germany) and
antimicrobial susceptibility testing was performed by Disk Diffusion Test Methodology as
described in the European Committee on Antimicrobial Susceptibility Testing (EUCAST, Version
1.0, December 18, 2009) or by The Vitek 2 automated system by use of cards AST-N209 and AST-
N122 (bioMérieux, France). Methods were performed according to guidelines of DCM in
agreement with direction from manufacturers and as described elsewhere (14–17). The ATCC
25922 E.coli was used for quality control and susceptibility interpreted as recommended by
EUCAST (www.eucast.org/clinical_breakpoints/). Isolates denoted as I (intermediately resistant)
were reported as resistant. ESBL phenotypes were identified by a double-disk diffusion method,
here performed as combined-disk diffusion using a AmpC + ESBL detection set (MAST®,
Merseyside, UK) as previously described (16).
Molecular characterization
Phylogrouping
The major phylogroups (group A, B1, B2 and D) were determined using singleplex PCR
amplification of three genes (chuA, yjaA and TSPE4.C2) (18,19). Strains with no detected PCR
products were classified as non-typeable (NT) (19,20).
ESBL-genotyping
For all ESBL-producing E.coli, phenotypically recognized by the double-disk diffusion MAST®
test, the ESBL genotypes were identified by PCR and subsequent sequencing. Isolates were
screened for the presence of blaCTX-M genes by multiplex PCR assay, detecting alleles encoding

6
the five CTX-M groups 1, 2, 8, 9 and 25 (12,21). Positive samples were re-amplified and products
sequenced to identify the exact genotypes (12,21).
MLVA and MLST
All included isolates were typed using a-MLVA, with PCR amplification of six variable number of
tandem repeats (VNTR) loci, as described by Nielsen et al. (12). In short, PCR were done as
singleplex PCR with unlabelled primers. Subsequent size determination were done using an
automated capillary electrophoresis system (QIAxcel, Qiagen) and a high-resolution cartridge (12).
Each of the VNTR loci was manually binned depending on size and assigned a number. As result
each isolate were given a six-digit a-MLVA code. For isolates without a measured band size, for a
VNTR locus, PCR amplification was performed twice for the given locus. The identified codes
were primarily translated to STs by an in-house library, but to further evaluate the a-MLVA and
confirm our findings, at least one isolate from each identified a-MLVA code were verified by
MLST, with the exception of one a-MLVA code. In groups with several isolates having identical a-
MLVA codes, we included isolates with extreme band sizes within bins, to effectively cohere
clusters. In the resistant and susceptible groups we furthermore included some isolates with
different O-serogroups. In all, MLST was performed on 31 isolates from the ESBL population, on
45 isolates from the resistant population and on 52 isolates from the susceptible population. MLST
was done using the Achtmann MLST scheme with PCR and sequencing of seven housekeeping
genes followed by assignment of an allelic number from the MLST-database
(http://mlst.warwick.ac.uk/mlst/dbs/Ecoli) (12,22).

7
Statistics
For statistical analysis categorical variables were compared by a Chi-square or Fisher exact test
when appropriate and p<0.05 was considered significant.
Results
For each of the three typing methods used in this investigation, we defined a cluster as three or
more isolates identified as the same O-serogroup, ST or a-MLVA code.
Characterization of the E.coli populations
The distributions of serogroups, a-MLVA codes and STs in the three populations are found in
Figure 1.
O-Serogroup distribution
We identified several serogroups among the selected 194 isolates; 24 different serogroups among
the 94 ESBL-producing E.coli, 17 among the 49 resistant E.coli and 27 among the 51 susceptible
E.coli isolates. Interestingly, we found dissimilar patterns of dominating serogroups in the three
populations. The five largest clusters in the ESBL population constituted 57% of all isolates.
Furthermore, 39% belonged to one large cluster of serogroup O25, constituting the largest cluster
found in any of the populations. The five major clusters in the resistant population amounted to
25% of isolates and in the susceptible population the two largest clusters constituted 7% of isolates
only. In addition, several serogroups with one or two isolates were identified in each group.
Characterization by MLVA and MLST
Results for the a-MLVA are presented in Table 1, 2 and 3.

8
PCR amplification of the six MLVA alleles was successfully performed on all isolates. A total of 83
unique a-MLVA codes were identified among the 286 isolates. Seventy four codes were associated
to one unique sequence type. There were, however, several occasions where isolates belonging to
one ST were subdivided into more than one a-MLVA codes. As examples, ST69 was subdivided by
three different a-MLVA codes, ST73 subdivided by ten different codes and ST131 was subdivided
by four different codes (Table 1-3).
Results from the MLST are likewise presented in Table 1-3. We found 72 different sequence types
in the 141 selected isolates from the three populations. We discovered ten STs, not previously
identified, here labelled “New ST”. Complex situations existed where the a-MLVA method did not
distinguish between two or more STs. Thus for example, ST38, ST117 and ST1177 were assigned
same a-MLVA code. ST38 and ST1177 are single locus variant with ST117 being unrelated.
A single a-MLVA code was likewise assigned to ST10, ST746, ST1598 and one New ST (Table 1-
3). Here ST10 and ST746 are single-locus variants with the new ST being a double-locus variant of
ST10. For a total of eight a-MLVA codes it was not possible to discriminate between isolates of
different STs. Phenotypic relations were tested by use of eBURST, on online tool displaying the
relationships between closely-related isolates (http://eburst.mlst.net/v3), which showed that the
majority of STs in each population were unrelated, with only a few isolates being single locus
variants (data not shown).
In the ESBL population we found 20 a-MLVA codes and 20 different STs. We found no unknown
ST´s in this population, but had one isolate unsuccessfully typed. This isolate belonged to a a-
MLVA cluster identified by MLST performed on other isolates. The population was dominated by a
few lineages with seven a-MLVA clusters and five clear ST-clusters. Fifty isolates belonged to
ST131 with code “153562” containing 44 isolates. Of the 44 isolates 35 belonged to the serogroup
O25 and phylogroup B2. Of the isolates identified as ST131 by the other a-MLVA codes, in this

9
population, five belonged to serogroup O16 and one to O153. Overall, the ESBL population had
significantly more isolates belonging to ST131 than the two other populations (p<0.0001 and p
=0.0001). ST73 and ST95 were not found among the ESBL-producing isolates.
In the resistant E.coli population, 36 a-MLVA codes and 30 ST´s were identified with a single a-
MLVA code for which MLST was not performed. These codes and STs were grouped in eight a-
MLVA- and eight ST-clusters. Here ST69, ST73 and ST131 amounted to the largest ST clusters
with 18%, 11% and 15%, respectively.
In the susceptible E.coli population 54 a-MLVA codes and 44 ST´s were found with seven a-
MLVA- and ten ST clusters. The susceptible population had significantly more STs and a-MLVA
codes compared to the ESBL population (p =0.0097 and p =0.0005). ST73 and ST95 dominated
with 16% and 6%, respectively, and ST73´s were sub-divided by ten a-MLVA codes.
Overall, ST131 constituted 23% of all isolates, ST69 constituted 10% and ST73 a total of 9%.
During the time period of sampling four per cent of E.coli isolates, from urine samples from general
practices at Hvidovre Hospital, were resistant to cefpodoxime, used as an indicator for ESBL-
production.
Antibiotic resistance patterns
Results of antibiotic susceptibility testing are found in Figure 2 and 3. Generally the ESBL
population showed high levels of resistance with 100% of the isolates showing resistance towards
three or more of the tested antibiotics. In the resistant population 66% of the isolates were resistant
to >3 of the antibiotics. All isolates were susceptible to meropenem and most to mecillinam (97-
99%), fosfomycin (96-97%), nitrofurantoin (93-96), and piperacillin/tazobactam (92-95%). Table 2
shows that resistant, non-ESBL ST131 have a broader spectrum of resistance than resistant (non-
ESBL) ST73 and ST69.

10
Phylogrouping of E.coli isolates
The distribution of phylogroup A, B1, B2, D and non-typeables (NT) were similar among the three
populations of uropathogenic E.coli. Phylogroup A was found in 7-9%, phylogroup B1 in 1-6%,
phylogroup B2 in 45-58%, phylogroup D in 22-32% and NT´s were found in 4-11% of isolates in
the three populations, respectively.
ESBL-genotyping
We found that 98% of the ESBL-producing E.coli isolates were positive for the presence of a
blaCTX-M. blaCTX-M group 1 dominated with 73% of all isolates carrying this type. Furthermore, 54%
of all isolates produced CTX-M-15. A total of 24% of isolates belonged to CTX-M group 9 with
two large clusters of CTX-M-14 (12%) and CTX-M-27 (12%).
Discussion
This study was carried out to describe the diversity among susceptible, resistant (non-ESBL) and
ESBL-producing E.coli using three typing methods (13). The objective was to clarify if E.coli
isolates from these dissimilar populations are genetically related. This would determine if ESBL are
carried by current successful UPEC gaining resistance or spread by less efficient UPEC effective in
the presence of antibiotic. Overall we found ST131, ST73 and ST69 to be the dominating lineages
among all isolates, in accordance with previous studies (23). Of these, non-ESBL-producing ST131
seemed to have a broader spectrum of resistance than ST73 and non-ESBL-producing ST69, as
described by Horner et al. (Figure 3) (23). However, based on serogroups, a-MLVA and MLST, in
this collection of uropathogenic E.coli from general practices, we found that ESBL-producing
E.coli, resistant E.coli and susceptible E.coli in turn were dominated by different serogroups, a-
MLVA codes and STs. In general, the susceptible E.coli population was a much more diverse group

11
of isolates with more STs and fewer and smaller clusters. Here the biggest ST-lineage, ST73, was
extensively sub-divided by a-MLVA indicating high diversity among these isolates.
The resistant E.coli and ESBL-producing E.coli were found in larger clusters. Especially the ESBL-
population was a more homogenous population with fewest a-MLVA codes and STs. The STs
found differed to some extent from those of the susceptible population with an overlap of some a-
MLVA codes and STs among the three populations. The prevalence of STs were, however,
different. We found that ST131 completely dominated this ESBL-producing E.coli population with
the combination of serogroup O25, phylogroup B2 and ST131 being by far the largest cluster, as
previously seen in the Copenhagen area (12,24). A single ST131 isolate was found among the
susceptible isolates and 14 detected in the resistant population and still ST131 was able to
effectively dominate the ESBL-producing UPEC. The presence of ESBL might not cause higher
levels of virulence or fitness in environments with limited use of antibiotics, to which the ESBL-
producing E.coli are resistant. Thus, ESBL-producing STs does not necessarily hold an advantage
in the community in Denmark, where use of extended spectrum cephalosporins, ciprofloxacin and
aminoglycosides are limited, but can spread in settings were these groups of antibiotics are
intensely used. Nonetheless, ST131 seems to be a specialized UPEC adapted as an efficient resistant
and ESBL-producing lineage, sustainable in the community once present, at least for some time. Of
the resistant ST73 just four out of ten were resistant >3 antibiotics, an absence of multidrug
resistance also seen in previous studies (2,6–10). Furthermore, our result indicates that resistance to
one or few antibiotics creates some positive selection in the community or among patients from
general practices. ST131 could be one of the UPEC lineages, among these resistant and ESBL-
producing isolates, with the highest ability to colonize the human gut, creating a high prevalence in
these populations. Urinary tract infections are often caused by the dominating ExPEC already
present in the gut of the patient and ST131, and other less successful ESBL-producing UPEC-

12
lineages, might be found to cause UTI simply since they dominate the microflora during and after
antibiotic exposure (25). Less prevalent ESBL-producing ST-lineages could be less effective
UPEC, allowing for the few specialized UPEC actually carrying ESBL, like ST131, to fiercely
dominate the population. We speculate if resistance in different E.coli populations are somewhat
defined by intrinsic differences in distinct E. coli lineages, making a limited number of UPEC ST-
lineages capable of obtaining and spreading blaCTX-M. Some lineages successfully acquire and
maintain different types of mobile resistance genes like ST69 and especially ST131, while other
E.coli lineages remains fairly susceptible and rarely take up plasmids, as seen with ST73 and ST95
(3,4). However, productions of ESBLs are found among specific UPEC present, to some, extent in
all populations. Antibiotic selection therefore creates a less varied population structure of related
isolates while antibiotic free environments allows for competition and diverse non-related
population structure (26). This is seen as difference in diversity of O-serogroups, a-MLVA codes
and STs in the three populations. The UPEC we found in our three populations have been found in
other population-studies of E.coli isolates causing bacteremia, strongly indicating that E.coli
isolates capable one invasive disease can cause severe invasive infections, independent of
susceptibility (23).
The a-MLVA and MLST had very high typeability and showed somewhat similar discriminatory
power dividing the populations into 83 a-MLVA codes and 72 STs, respectively. Though, MLVA
seemed slightly more discriminatory; subdividing several STs. There is a higher rate of sustainable
genetic variations within the VNTR regions used in MLVA, than in the housekeeping genes used
for MLST. However, a change in a VNTR region does not necessarily lead to a change in ST, and
vice versa. This poses a challenge in identifying isolates with new VNTR-sequences, here seen as
limitations in incomplete separation of all STs, but it also provides high discriminatory power in the
ability to separate closely related ST-lineages and thereby describe E.coli populations in detail

13
(27,28). Some of the undistinguishable STs was found to share a-MLVA codes by Nielsen et al,
such as ST101 and ST448 (12). The missing identification of variation in STs are, however,
unlikely to cause bias in our overall conclusion as the prevalence and identification of the major
lineages in the three populations are unique. The a-MLVA typing method was successful in the
characterization and sub-division of some large cluster of ST-lineages (Table 1-3). Thus, ST69 was
the largest cluster in the resistant population and found in all populations with three different a-
MLVA codes. ST73 and ST95 amounted for larger clusters in both the resistant and susceptible
populations and here the a-MLVA method could describe the STs as heterogeneous lineages, by
classifying each lineage by different a-MLVA codes. Finally, the lineage of ST131 was sub-divided
by a-MLVA. All isolates identified as ST131 belonged to phylogroup B2, but we detected a large
ST131 cluster with one a-MLVA code primarily belonging to O25, all of which were resistant to
ciprofloxacin. A minor ST131 cluster, with another a-MLVA code belonging to O16, with no
isolates showing resistance to ciprofloxacin, was likewise detected. This corresponds well with
reports that most ST131 isolates belong to serogroup O25 and a minor part being identified as O16
(24,29). Furthermore, sub-division of other ST-lineages have successfully been done by fimH
sequencing and more discriminatory typing method like PFGE and our results are in line with the
reports identifying susceptible ST-lineages as more heterogeneous populations (2,6,29,30). This is
in clear contradiction to what has previously been stated, in a study based on sequence typing only,
where all E.coli isolates were found to be of clonal distribution, disregarding resistance (3).
The study is limited by the relatively small number of isolates characterized and the limited time
period of collection, which does not allow us to identify any fluctuation in dominant lineages of
non-ESBL populations. Furthermore, the ESBL-population is a more selected group, allowing us to
detect less successful STs with limited impact as UPEC. As all isolates are from general practices, it

14
would have been beneficial to precisely know to what extent UTIs were devoid of healthcare
association.
Nonetheless, our results strongly suggest that the observed dissemination of ESBL-producing
E.coli are due to the spread of certain UPEC lineages already present in the general UPEC
population (3). Less prevalent ESBL-producing E.coli might be non-specialized UPEC, why they
are not found in the non-ESBL populations. In contrast, lineages like ST69 and ST131 are UPEC
present in all populations and there able to dominate in the ESBL-population. It seems that the level
of virulence could be of limited epidemiological importance for the spread of resistant E.coli, since
this most likely occurs due to selection of previously specialized UPEC with limited fitness loss due
to ESBL-production (31).
Acknowledgements
Work has been performed as cooperation between HVH, Roskilde University and University of Copenhagen with
laboratory facilities provided by HVH. This work was, in parts, presented at ECCMID in Barcelona 2014.
Funding
This work was supported by the EU-FP funded project PAR7, DanCARD (Danish National Strategic Research
Foundation, project 09-067075/DSF), Roskilde University, University of Copenhagen and the SSAC
Foundation (SLS-327421 and SLS-251761).
Transparency declarations
None to declare.

15
1. Croxall G, Hale J, Weston V, Manning G, Cheetham P, Achtman M, et al. Molecular
epidemiology of extraintestinal pathogenic Escherichia coli isolates from a regional cohort of
elderly patients highlights the prevalence of ST131 strains with increased antimicrobial
resistance in both community and hospital care settings. 2011;(August):2501–8.
2. Banerjee R, Johnston B, Lohse C, Chattopadhyay S, Tchesnokova V, Sokurenko E V, et al.
The clonal distribution and diversity of extraintestinal Escherichia coli varies according to
patient characteristics. Antimicrob Agents Chemother. 2013 Sep 16;(September):1–29.
3. Adams-Sapper S, Diep BA, Perdreau-Remington F, Riley LW. Clonal composition and
community clustering of drug-susceptible and -resistant Escherichia coli isolates from
bloodstream infections. Antimicrob Agents Chemother. 2013 Jan;57(1):490–7.
4. Bengtsson S, Naseer U, Sundsfjord A, Kahlmeter G, Sundqvist M. Sequence types and
plasmid carriage of uropathogenic Escherichia coli devoid of phenotypically detectable
resistance. J Antimicrob Chemother. 2012 Jan;67(1):69–73.
5. Nicolas-Chanoine M-H, Blanco J, Leflon-Guibout V, Demarty R, Alonso MP, Caniça MM,
et al. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-
M-15. J Antimicrob Chemother. 2008 Feb;61(2):273–81.
6. Blanco J, Mora A, Mamani R, López C, Blanco M, Dahbi G, et al. National survey of
Escherichia coli causing extraintestinal infections reveals the spread of drug-resistant clonal
groups O25b:H4-B2-ST131, O15:H1-D-ST393 and CGA-D-ST69 with high virulence gene
content in Spain. J Antimicrob Chemother. 2011 Sep;66(9):2011–21.
7. Johnson JR, Tchesnokova V, Johnston B, Clabots C, Roberts PL, Billig M, et al. Abrupt
emergence of a single dominant multidrug-resistant strain of Escherichia coli. J Infect Dis.
2013 Mar 15;207(6):919–28.

16
8. Mora A, Blanco M, López C, Mamani R, Blanco JE, Alonso MP, et al. Emergence of clonal
groups O1:HNM-D-ST59, O15:H1-D-ST393, O20:H34/HNM-D-ST354, O25b:H4-B2-
ST131 and ONT:H21,42-B1-ST101 among CTX-M-14-producing Escherichia coli clinical
isolates in Galicia, northwest Spain. Int J Antimicrob Agents. 2011 Jan;37(1):16–21.
9. Rogers B a, Sidjabat HE, Paterson DL. Escherichia coli O25b-ST131: a pandemic,
multiresistant, community-associated strain. J Antimicrob Chemother. 2011 Jan;66(1):1–14.
10. Gibreel TM, Dodgson AR, Cheesbrough J, Fox AJ, Bolton FJ, Upton M. Population
structure, virulence potential and antibiotic susceptibility of uropathogenic Escherichia coli
from Northwest England. J Antimicrob Chemother. 2012 Feb;67(2):346–56.
11. Foxman B. Extended-spectrum β-lactamase-producing escherichia coli in the united states:
time to rethink empirical treatment for suspected E. coli infections? Clin Infect Dis. 2013
Mar;56(5):649–51.
12. Nielsen JB, Albayati A, Jørgensen RL, Hansen KH, Lundgren B, Schønning K. An
abbreviated MLVA identifies Escherichia coli ST131 as the major extended-spectrum β-
lactamase-producing lineage in the Copenhagen area. Eur J Clin Microbiol Infect Dis. 2013
Nov 6;32(3):431–6.
13. Köhler C-D, Dobrindt U. What defines extraintestinal pathogenic Escherichia coli? Int J Med
Microbiol. 2011 Dec;301(8):642–7.
14. Livermore DM, Struelens M, Amorim J, Baquero F, Bille J, Canton R, et al. Multicentre
evaluation of the VITEK 2 Advanced Expert System for interpretive reading of antimicrobial
resistance tests. J Antimicrob Chemother. 2002 Feb;49(2):289–300.
15. Nyberg SD, Meurman O, Jalava J, Rantakokko-Jalava K. Evaluation of detection of
extended-spectrum beta-lactamases among Escherichia coli and Klebsiella spp. isolates by

17
VITEK 2 AST-N029 compared to the agar dilution and disk diffusion methods. Scand J
Infect Dis. 2008 Jan;40(5):355–62.
16. Hansen F, Olsen SS, Heltberg O, Justesen US, Fuglsang-Damgaard D, Knudsen JD, et al.
Characterization of Third-Generation Cephalosporin-Resistant Escherichia coli from
Bloodstream Infections in Denmark. Microb Drug Resist. 2014 Feb 11;00(00):1–9.
17. Haigh J, Degun A, Eydmann M, Millar M, Wilks M. Improved performance of bacterium
and yeast identification by a commercial matrix-assisted laser desorption ionization-time of
flight mass spectrometry system in the clinical microbiology laboratory. J Clin Microbiol.
2011 Sep;49(9):3441.
18. Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli
phylogenetic group. Appl Environ Microbiol. 2000 Oct;66(10):4555–8.
19. Gordon DM, Clermont O, Tolley H, Denamur E. Assigning Escherichia coli strains to
phylogenetic groups: multi-locus sequence typing versus the PCR triplex method. Environ
Microbiol. 2008 Oct;10(10):2484–96.
20. Clermont O, Christenson JK, Denamur E, Gordon DM. The Clermont Escherichia coli
phylo-typing method revisited: improvement of specificity and detection of new phylo-
groups. Environ Microbiol Rep. 2013 Feb;5(1):58–65.
21. Jørgensen RL, Nielsen JB, Friis-Møller A, Fjeldsøe-Nielsen H, Schønning K. Prevalence and
molecular characterization of clinical isolates of Escherichia coli expressing an AmpC
phenotype. J Antimicrob Chemother. 2010 Mar;65(3):460–4.
22. Wirth T, Falush D, Lan R, Colles F, Mensa P, Wieler LH, et al. Sex and virulence in
Escherichia coli: an evolutionary perspective. Mol Microbiol. 2006 Jun;60(5):1136–51.

18
23. Horner C, Fawley W, Morris K, Parnell P, Denton M, Wilcox M. Escherichia coli
bacteraemia: 2 years of prospective regional surveillance (2010-12). J Antimicrob
Chemother. 2014 Jan;69(1):91–100.
24. Olesen B, Hansen DS, Nilsson F, Frimodt-Møller J, Leihof RF, Struve C, et al. Prevalence
and Characteristics of the Epidemic Multiresistant Escherichia coli ST131 Clonal Group
among Extended-Spectrum Beta-Lactamase-Producing E. coli Isolates in Copenhagen,
Denmark. J Clin Microbiol.2013 Jun;51(6):1779–85.
25. Nielsen KL, Dynesen P, Larsen P, Frimodt-Møller N. Faecal Escherichia coli from patients
with E. coli urinary tract infection and healthy controls who have never had a urinary tract
infection. J Med Microbiol. 2014 Apr;63(Pt 4):582–9.
26. Maiden MCJ. Multilocus sequence typing of bacteria. Annu Rev Microbiol. 2006
Jan;60:561–88.
27. Keys C, Kemper S, Keim P. Highly diverse variable number tandem repeat loci in the E. coli
O157:H7 and O55:H7 genomes for high-resolution molecular typing. J Appl Microbiol. 2005
Jan;98(4):928–40.
28. Manges AR, Tellis P a, Vincent C, Lifeso K, Geneau G, Reid-Smith RJ, et al. Multi-locus
variable number tandem repeat analysis for Escherichia coli causing extraintestinal
infections. J Microbiol Methods. Elsevier B.V.; 2009 Nov;79(2):211–3.
29. Peirano G, van der Bij AK, Gregson DB, Pitout JDD. Molecular epidemiology over an 11-
year period (2000 to 2010) of extended-spectrum β-lactamase-producing Escherichia coli
causing bacteremia in a centralized Canadian region. Journal of clinical microbiology. 2012.
p. 294–9.

19
30. Johnson JR, Tchesnokova V, Johnston B, Clabots C, Roberts PL, Billig M, et al. Abrupt
emergence of a single dominant multidrug-resistant strain of Escherichia coli. J Infect Dis.
2013 Mar 15;207(6):919–28.
31. Bengtsson S, Naseer U, Sundsfjord A, Kahlmeter G, Sundqvist M. Sequence types and
plasmid carriage of uropathogenic Escherichia coli devoid of phenotypically detectable
resistance. J Antimicrob Chemother. 2012 Jan;67(1):69–73.

20
Figure 1. Here we show the percentages of found a-MLVA codes, STs and O-serogroups found in each of the three populations.
Percentage of identifed a-MLVA codes,STs and O-groups
MLVA
MLST
Serogro
ups0
20
40
60
80
100
120
140
160 Susceptible Resistant ESBL
Typing methods
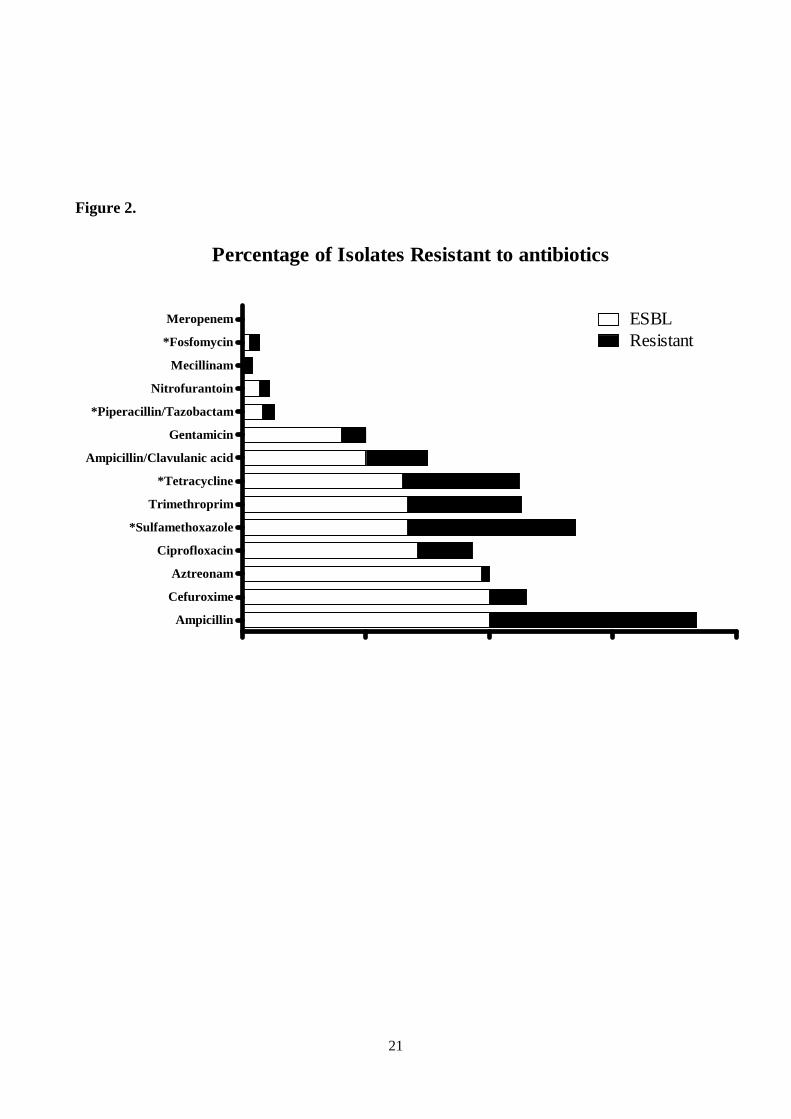
21
Figure 2.
Percentage of Isolates Resistant to antibiotics
Ampicillin
Cefuroxime
Aztreonam
Ciprofloxacin
*Sulfamethoxazole
Trimethroprim
*Tetracycline
Ampicillin/Clavulanic acid
Gentamicin
*Piperacillin/Tazobactam
Nitrofurantoin
Mecillinam
*Fosfomycin
Meropenem ESBLResistant

22
Figure 3. Percentages of isolates resistant to chosen antibiotics used for oral administration. We show resistance patterns for three common UPEC ST lineages, here all isolates are non-ESBL-producers but present in either the susceptible or resistant population. N: ST131 = 17, ST73 = 25, ST69 = 20.
Resistance patterns for non-ESBL UPEC lineages
0 20 40 60 80 100 120 140 160 180 200
Ampicillin
Ciprofloxacin
*Sulfamethoxazole
Trimethroprim
Nitrofurantoin
Mecillinam
*Fosfomycin
Resistance to >3 antibiotics
Fully susceptible
ST131ST73ST69
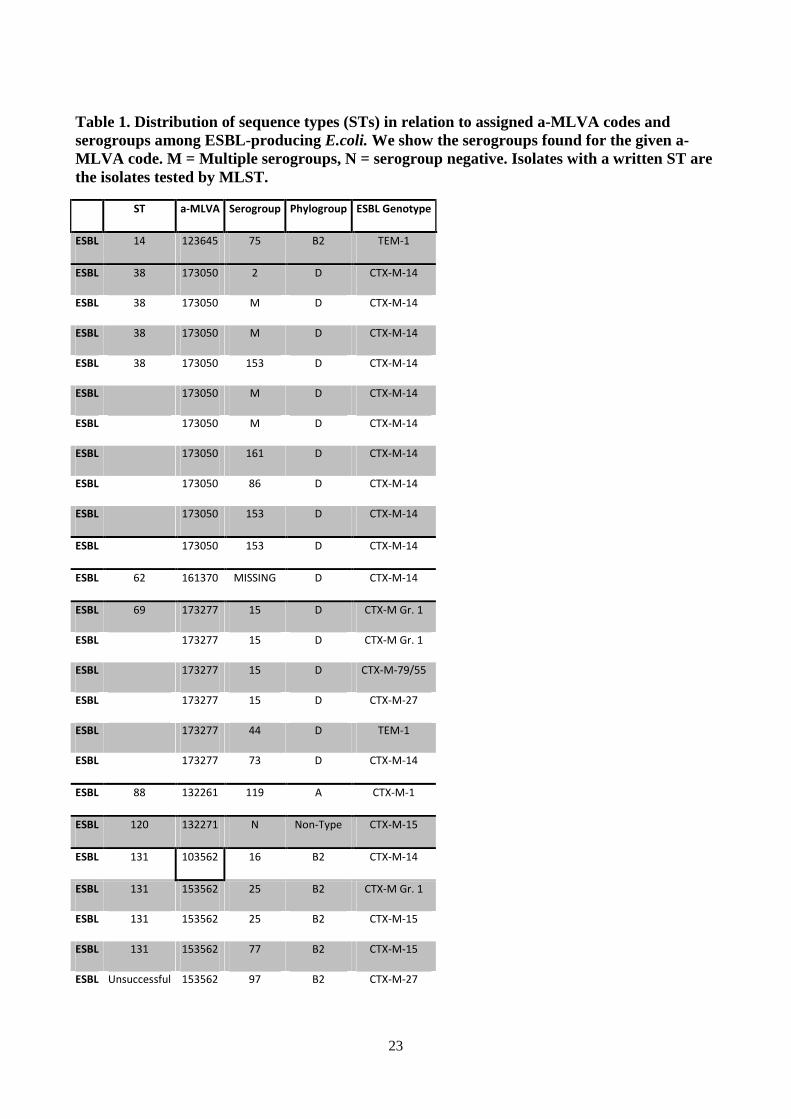
23
Table 1. Distribution of sequence types (STs) in relation to assigned a-MLVA codes and serogroups among ESBL-producing E.coli. We show the serogroups found for the given a-MLVA code. M = Multiple serogroups, N = serogroup negative. Isolates with a written ST are the isolates tested by MLST.
ST a-MLVA Serogroup Phylogroup ESBL Genotype
ESBL 14 123645 75 B2 TEM-1
ESBL 38 173050 2 D CTX-M-14
ESBL 38 173050 M D CTX-M-14
ESBL 38 173050 M D CTX-M-14
ESBL 38 173050 153 D CTX-M-14
ESBL 173050 M D CTX-M-14
ESBL
173050 M D CTX-M-14
ESBL 173050 161 D CTX-M-14
ESBL
173050 86 D CTX-M-14
ESBL 173050 153 D CTX-M-14
ESBL 173050 153 D CTX-M-14
ESBL 62 161370 MISSING D CTX-M-14
ESBL 69 173277 15 D CTX-M Gr. 1
ESBL
173277 15 D CTX-M Gr. 1
ESBL 173277 15 D CTX-M-79/55
ESBL
173277 15 D CTX-M-27
ESBL 173277 44 D TEM-1
ESBL 173277 73 D CTX-M-14
ESBL 88 132261 119 A CTX-M-1
ESBL 120 132271 N Non-Type CTX-M-15
ESBL 131 103562 16 B2 CTX-M-14
ESBL 131 153562 25 B2 CTX-M Gr. 1
ESBL 131 153562 25 B2 CTX-M-15
ESBL 131 153562 77 B2 CTX-M-15
ESBL Unsuccessful 153562 97 B2 CTX-M-27
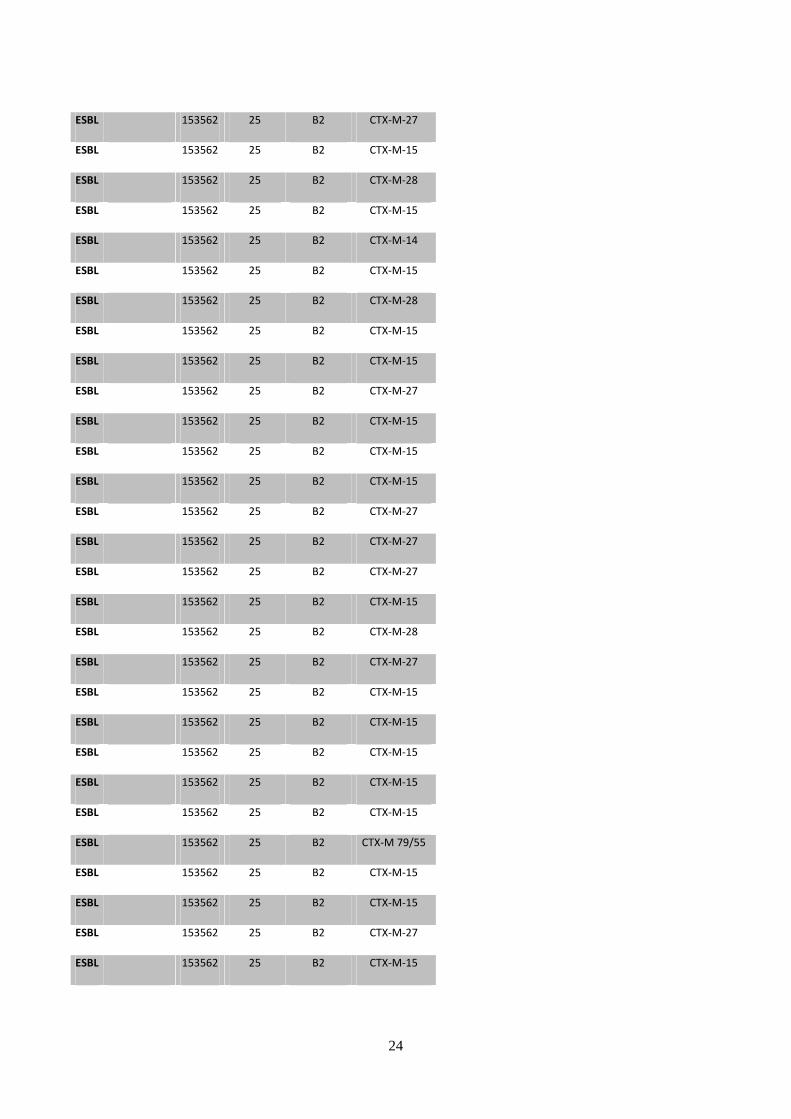
24
ESBL 153562 25 B2 CTX-M-27
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-28
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-14
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-28
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-27
ESBL
153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-28
ESBL 153562 25 B2 CTX-M-27
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M 79/55
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-27
ESBL 153562 25 B2 CTX-M-15

25
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 25 B2 CTX-M-15
ESBL 153562 25 B2 CTX-M-15
ESBL
153562 M B2 CTX-M-27
ESBL 153562 M B2 CTX-M-15
ESBL
153562 M B2 CTX-M-15
ESBL 153562 M B2 CTX-M-15
ESBL
153562 MISSING B2 CTX-M-28
ESBL 153562 MISSING B2 CTX-M-27
ESBL 153562 N B2 CTX-M-28
ESBL 131 163562 16 B2 CTX-M-15
ESBL 131 163562 16 B2 CTX-M-15
ESBL 163562 16 B2 CTX-M-15
ESBL
163562 16 B2 CTX-M-1
ESBL 163562 153 B2 CTX-M-15
ESBL 224 131271 M B1 CTX-M-15
ESBL 315 173552 25 B2 CTX-M-14
ESBL 354 151170 153 D CTX-M-14
ESBL 428 252365 117 B2 CTX-M-1
ESBL 617 132251 12 A CTX-M-15
ESBL 132251 69 A CTX-M-28/15
ESBL
132251 N A CTX-M-15
ESBL 132251 116,162 A CTX-M-15
ESBL
132251 N A CTX-M-79/55
ESBL 132251 N A CTX-M-15
ESBL 132251 12 A CTX-M-15
ESBL 636 293852 21 B2 CTX-M-15
ESBL 648 161160 1 D CTX-M-15

26
ESBL 648 161160 102,13 D CTX-M-15
ESBL
161160 153 D CTX-M-15
ESBL 161160 MISSING D CTX-M-15
ESBL 998 224665 2 B2 CTX-M-15
ESBL 998 224665 M B2 CTX-M-1
ESBL 2852 131291 8 B1 CTX-M-15
ESBL 131291 25 B2 CTX-M-15
ESBL 101 131261 N B1 CTX-M-1
ESBL 448 131261 111 B1 CTX-M-15
ESBL
131261 170 NT CTX-M Gr. 9
ESBL 131261 81 B1 CTX-M-79/55
ESBL
131261 100 NT CTX-M-1
ESBL 131261 M B1 CTX-M-15
ESBL 746 131251 21 A CTX-M-28
ESBL 1598 131251 9 NT CTX-M-15
Table 2. Distribution of sequence types (STs) in relation to assigned a-MLVA codes and serogroups, for the resistant group. Not all resistant isolates were characterized by serogroup. Here we show the serogroups found for the given a-MLVA code. M = Multiple serogroups, N = serogroup negative. Isolates with a written ST are the isolates tested by MLST.
ST a-MLVA Serogroup Phylogroup
Resistant 14 123645 75 B2
Resistant 38 173051 25 D
Resistant 58 131261 19 B1
Resistant
131261
B1
Resistant
131261
NT
Resistant
131261 N NT
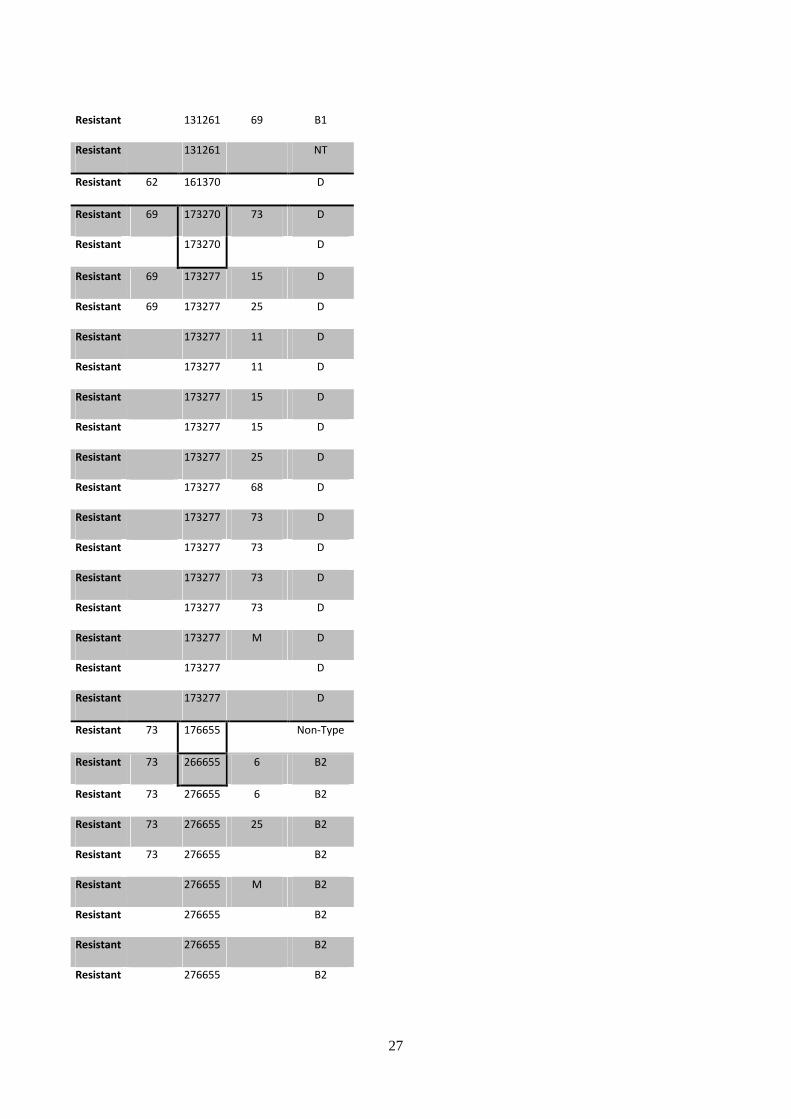
27
Resistant
131261 69 B1
Resistant
131261
NT
Resistant 62 161370
D
Resistant 69 173270 73 D
Resistant
173270
D
Resistant 69 173277 15 D
Resistant 69 173277 25 D
Resistant
173277 11 D
Resistant
173277 11 D
Resistant
173277 15 D
Resistant
173277 15 D
Resistant
173277 25 D
Resistant
173277 68 D
Resistant
173277 73 D
Resistant
173277 73 D
Resistant
173277 73 D
Resistant
173277 73 D
Resistant
173277 M D
Resistant
173277
D
Resistant
173277
D
Resistant 73 176655
Non-Type
Resistant 73 266655 6 B2
Resistant 73 276655 6 B2
Resistant 73 276655 25 B2
Resistant 73 276655
B2
Resistant
276655 M B2
Resistant
276655
B2
Resistant
276655
B2
Resistant
276655
B2

28
Resistant
276655
B2
Resistant 80 254575 75 B2
Resistant 80 254575 75 B2
Resistant 88 132261 8 NT
Resistant 132261 NT
Resistant New ST 132261
NT
Resistant 95 223643 2 B2
Resistant 95 223653 2 Non-Type
Resistant
223653
Non-Type
Resistant 127 264953 6 B2
Resistant 131 143562
B2
Resistant 131 153562 25 B2
Resistant 131 153562
B2
Resistant
153562 25 B2
Resistant
153562 25 B2
Resistant
153562 25 B2
Resistant
153562
B2
Resistant
153562
B2
Resistant
153562
B2
Resistant
153562
B2
Resistant
153562
B2
Resistant 131 163562
B2
Resistant
163562
B2
Resistant
163562
B2
Resistant 135 225661
B2
Resistant 141 124665
B2
Resistant 362 143250
D
Resistant 372 224563 18 B2

29
Resistant 393 171577
D
Resistant 405 121250 M D
Resistant
121250
D
Resistant 405 131250
D
Resistant
131250
D
Resistant 457 161371 11 D
Resistant 648 161150 1 D
Resistant 978 226565 83 B2
Resistant 1193 124645 75 B2
Resistant 1193 124645 6 B2
Resistant 124645 B2
Resistant 124645 D
Resistant
124645
B2
Resistant
124645 75 B2
Resistant 1597 162562 39 B2
Resistant 1597 162562
B2
Resistant New ST 123552
B2
Resistant New ST 131251
NT
Resistant New ST 162438
NT
Resistant MISSING 266561
NT
Resistant 10 124643 75 B2
Resistant 124643 75 B2
Resistant 10 132251 A
Resistant 132251 3 A
Resistant 132251 A
Resistant
132251
A
Resistant
132251
A
Resistant
132251 N A
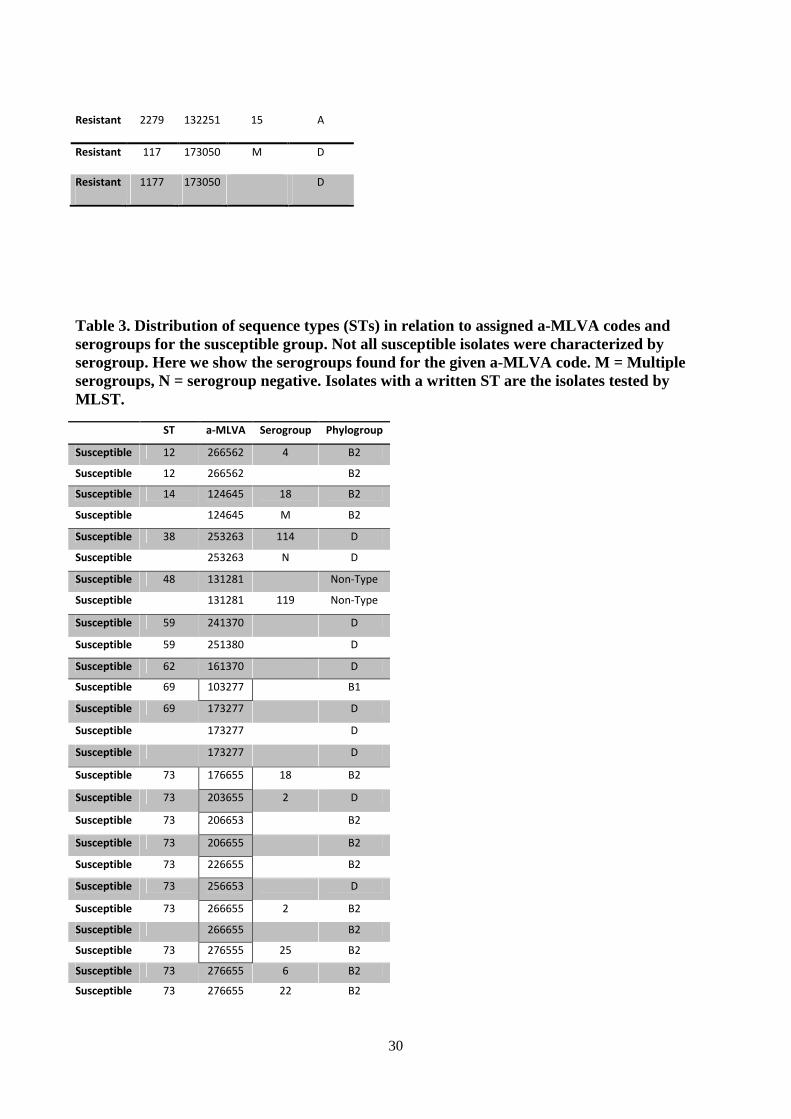
30
Resistant 2279 132251 15 A
Resistant 117 173050 M D
Resistant 1177 173050 D
Table 3. Distribution of sequence types (STs) in relation to assigned a-MLVA codes and serogroups for the susceptible group. Not all susceptible isolates were characterized by serogroup. Here we show the serogroups found for the given a-MLVA code. M = Multiple serogroups, N = serogroup negative. Isolates with a written ST are the isolates tested by MLST.
ST a-MLVA Serogroup Phylogroup
Susceptible 12 266562 4 B2
Susceptible 12 266562 B2
Susceptible 14 124645 18 B2
Susceptible 124645 M B2
Susceptible 38 253263 114 D
Susceptible 253263 N D
Susceptible 48 131281 Non-Type
Susceptible 131281 119 Non-Type
Susceptible 59 241370 D
Susceptible 59 251380 D
Susceptible 62 161370 D
Susceptible 69 103277 B1
Susceptible 69 173277 D
Susceptible 173277 D
Susceptible 173277 D
Susceptible 73 176655 18 B2
Susceptible 73 203655 2 D
Susceptible 73 206653 B2
Susceptible 73 206655 B2
Susceptible 73 226655 B2
Susceptible 73 256653 D
Susceptible 73 266655 2 B2
Susceptible 266655 B2
Susceptible 73 276555 25 B2
Susceptible 73 276655 6 B2
Susceptible 73 276655 22 B2

31
Susceptible 73 276655 M B2
Susceptible 276655 2 B2
Susceptible 276655 B2
Susceptible 73 276665 120 B2
Susceptible 80* 254575 B2
Susceptible 95 203653 B2
Susceptible 95 223653 1 B2
Susceptible 223653 1 B2
Susceptible 223653 M B2
Susceptible 223653 B2
Susceptible 223653 Non-Type
Susceptible 101 131261 174 Non-Type
Susceptible 131261 Non-Type
Susceptible 127 264953 B2
Susceptible 264953 6 B2
Susceptible 127 274953 6 B2
Susceptible 131 153562 B2
Susceptible 141 204665 B2
Susceptible 204665 D
Susceptible 162 211251 95 A
Susceptible 223 101261 B2
Susceptible 101261 Non-Type
Susceptible 101261 B2
Susceptible 357 150365 B2
Susceptible 357 152365 73 B2
Susceptible 152365 M B2
Susceptible 405 131250 2 D
Susceptible 410 132261 A
Susceptible 501 163777 D
Susceptible 538 152363 13 B2
Susceptible 582 274242 B2
Susceptible 589 287572 B2
Susceptible 681 224452 8 D
Susceptible 714 473050 D
Susceptible 1161 124675 B2
Susceptible 1331 274655 6 D
Susceptible 1444 273673 4 B2
Susceptible 1858 224645 6 B2
Susceptible 3672 226962 B2
Susceptible 4235 224743 88 B2
Susceptible 224743 88 B2

32
Susceptible New ST 1 151170 D
Susceptible New ST 5 112871 D
Susceptible New ST 6 272275 133 B2
Susceptible New ST 8 232251 105 A
Susceptible New ST 9 171250 D
Susceptible 10 131251 109 A
Susceptible 10 131251 M A
Susceptible 131251 107,117 A
Susceptible 10 132251 81 A
Susceptible 93 132251 7 Non-Type
Susceptible 132251 162 A
Susceptible 132251 N A
Susceptible 540 132251 15 Non-Type
Susceptible 540 132251 Non-Type
Susceptible New ST 3 132251 11 Non-Type
Susceptible 141 224665 2 D
Susceptible 141 224665 2 B2
Susceptible 141 224665 B2
Susceptible 998 224665 2 B2
Susceptible 998 224665 2 D
Susceptible 224665 B2
Susceptible 224665 2 B2
Susceptible 224665 138 D
Susceptible 224665 161 B2
Susceptible 224665 B2
Susceptible 420 254665 82 B2
Susceptible 3846 254665 B2


















