EXPRESSION OF CFTR mRNA IN NASAL EPITHELIUM AND VAS DEFERENS · University of Toronto The gene...
96
EXPRESSION OF CFTR mRNA IN NASAL EPITHELIUM AND VAS DEFERENS Victor Mak, M.D. A thesis submitted in conformity with the requirements for the degree of Master of Science Graduate Department of Institute of Medical Science University of Toronto O Copyright by Victor Mak 1997
Transcript of EXPRESSION OF CFTR mRNA IN NASAL EPITHELIUM AND VAS DEFERENS · University of Toronto The gene...

EXPRESSION OF CFTR mRNA
IN NASAL EPITHELIUM
AND VAS DEFERENS
Victor Mak, M.D.
A thesis submitted in conformity with
the requirements for the degree of
Master of Science
Graduate Department of Institute of Medical Science
University of Toronto
O Copyright by Victor Mak 1997

Bibiiographic Services services bibliographiques 395 Wellington Street 395, rue Wellington Ottawa ON K I A ON4 Ottawa ON K I A ON4 Canada Canada
Your rï& Votre rdterence
Our file Nolre rdOrence
The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distribute or sel1 reproduire, prêter, distribuer ou copies of this thesis in microforni, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/film, de
reproduction sur papier ou sur fomat électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts fiom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

University of Toronto
The gene responsible for cystic fibrosis (CF), called the cystic fibrosis
transmembrane conductance regulator (CFTR), encodes the CAMP-regulated chloride
channel found in the apical membrane of secretory epithelial cells. It has been well
established that almost al1 males with CF are azoospermie due to atrophy or absence of
structures derived fiom the Wolffian duct. Interestingly, a higher than expected
fiequency of mutations in the CFTR gene has been identified in men with congenital
absence of vas deferens and men with epididymal obstruction. In particular, these
individuals have been found to have a significantly higher incidence of the 5-thymidine
(5T) variant of the CFTR intron 8 polypyrimidine tract (IVS8-T tract) compared to
normal or CF populations. The 5T variant results in less efficient splicing of CFTR exon
9 compared to the more common 7T and 9T variants and therefore produces less normal,
full-length CFTR mRNA. The protein produced by the CFTR transcript lacking exon 9
fails to fùnction as a CAMP-dependent chloride channel. The fact that these infertile
males have no other clinical signs of classical CF suggests that the epithelia of the male
reproductive tract may have the highest requirement for CFTR fûnction or, altematively,
splicing of CFTR mRNA in the reproductive tract is less efficient than the other CF-
associated organs. Nasal epithelia and segments of vas deferens were obtained fiom 24
healthy, previously vasectomized men who presented for vasectomy reversal.
Quantitative RT-PCR was performed on these specimens, with the region of CFTR
cDNA spanning exon 9 amplified. For both nasal and vasal tissues, a strong positive
correlation was found between the length of the TVS8-T tract and the proportion of
&A with exon 9 intact. In addition, within the same subject, a significantly higher
level of transcripts lacking exon 9 was found in vas deferens than nasal epithelia,
regardless of the NS8-T genotype. These findings suggest that the splicing of CFTR
precursor mRNA is less efficient in vasal epithelia compared to respiratory epitheiia.
Thus, differential splicing efficiency between the various tissues which express CFTR
provides one possible explanation for the reproductive tract abnormalities observed in
infertile men with CFTR gene alterations but without other manifestations of CF.

The author wishes to express sincere appreciation
to Prof. Lap-Chee Tsui, Dr. Keith Jarvi, and
Dr. Johanna Rornmens for their assistance in the
preparation of this manuscript.

.......................................................................................................................... List of Tables iv
.......................................................................................................................... List of Figures v
Abbreviations .......................................................................................................................... vi
.............................................................................................. Attribution of Labor and Data 1
1 . Introduction ...................................................................................................................... 2
............................................................................................................ I.A. Background 2
....................................................................................... 1 .A. 1 . Human Male Znfirtility 2
I.A. 1 .a. Overview of Male Testicular and
.............................................................. Reproductive Tract Embryology 2
.................................................................... I.A. 1 .ai. Development of testes 2
............................................... I.A. 1 .aii . Development of male genital ducts 3
........................................ I.A. 1 .aii i. Development of male external genitalia 4
I.A.1 . b. OveMew of Male Reproductive Physiology .............................................. 4
.......................................... I.A. 1 . b.i. Hypothdamic-pituitary-testicular axis 5
........................................................................................ 1.A.l .b.i i. The testis 5 .. ......................................................................... LA.1 .b.ii. a. Leydig cells 5
.......................................................... I.A. 1 .b.ii. b. Seminiferous tubules 6 . . I.A. 1 .b.ii. c. Spermatogenesis ................................................................ -7
......................................... I.A. 1 .b.ii i. Hormonal control of spermatogenesis 7
.................................... I.A. 1 .b.i v. Sperm maturation, storage, and transport 8
LA . 1 .b. v. Fertilization ................................................................................... -9 . . .................................*.......,................... LA 1 c. Clinical Aspects of Male Infertility 9
................................................... I.A. 1 .c .i. Evaluation of the infertile male 10
................................................................. I.A. l .c.i. a. Semen analysis 1 0
................................... l.A.l .c.i i. Etiologic classification of male infertility 11
l.A.l . c i i. Treatment of male infertility .................................................... 11

Conductance Regulator ................................................................................. 1 2
I.A.3. Cystic Fibrosis Trunsmembrane Conductance Regulator and
........................................................................ Mesonephric Duct Anomalies 13
I.A.4. The Intron 8 Polypyrimidine Tract of t he CFTR Gene and
Alternative Splicing of CFTR Exon 9 ....................................................... 1 8
I.A.5 . The 5T Variant in Iniron 8 of the CFTR Gene and
Congenital Absence of Yas Deferens ............................................................. 19
I.B. Hypothesis ........................................................................................................... 21
1.C . Purpose ................................................................................................................ 21
.................................................................................................. II . Materials and Methods 22
Patients and samples ........................................................................................... 22
Allele specific oligonucleotide analysis of NS8-T tract ................................... 22
................................. PCR amplification of CFTR exon 9+ and exon 9- cDNA 24
Analysis of CFTR mRNA by reverse transcriptase-
polymerase chain ration .............................................................................. 25
Quantification of CFTR mRNA transcripts ....................................................... 27
Analysis of data ................................................................................................... 27
............................................................................................................................. III . Resuits 29
m.A. WS8-T tract genotype ......................................................................................... 29
iII.B. Conversion of experimental to actual proportions of
................................................................................... exon 9+ CFTR mRNA 29
ïiI.C. Evaluation of exon 9+ and exon 9- CFTR mRNA transcripts .......................... 31 ........................................................... m.D. Quantification of exon 9+ CFTR mRNA 32
m.E. Relationship between level of exon 9+ C F ' mRNA
....................................................................................... and the IVSS-T tract 32
iIi.F. Cornparison of the proportion of exon Pt CFTR mRNA
in nasal epitheliurn to vas deferens ................................................................ 32

- . - - - - - - - - - - - - -
1V.A. Splicing efficiency of CFTR exon 9 is related to
the iVS8-T tract length ...... .. . ......... ............ ..... . .. .. ... ...... , .. . . .. .. ............. . . . ...... .. .33
N . B . Higher proportion of normal CFTR mRNA in
nasal epithelium than vas deferens ................................................................ 33
W.C. The CFTR IVS8-T tract modifies phenotype by variable
splicing of exon 9 in different tissues ............................................................ 34
1V.D. Proportion of normal CFTR mRNA produced by
the WS8-T aileles ........ .... ................... . ... ..... ..... ....... .... ...........,..... . ................. 3 5
W.E. Molecular mechanisrn of differential splicing efficiency
of CFTR exon 9 .............................................,................................................ 37
N . F . Summary ............................................................................................... . ............. 38
V. Future Directions .... . .......... .... ........ .., . .. . ,..... .. ........... ....., .. ...... . ..... . . . . .... ...... ....... . . . . ...... .. -40
VI. References .. . . ... . . . . ... . . . . . . . . . . .. . . . . . . . .. . . . . .. . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . ... . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . ... -42

Table 1. World Health Organization criteria for
normal semen analysis .................................................................................. 54
Table 2. CFïR gene mutations or variations and CBAVD ............................................ 55
Table 3. Frequencies of the WS8-T alleles in subjects from
present study and in general population ..................................................... 56
Table 3. Proportion of Cl?T'R exon 9+ mRNA in
nasal epithelia and vas deferens ................................................................... 57
Table 4. Estimated proportion of CFTR exon 9+ transcripts
produced by the IVS8-T tract alieles ........................................................... 58
Table 5. Estimated proportion of CFTR exon 9+ transcripts
produced by the WS8-T tract genotypes .................................................... 59

Figure 1. The intron 8 polypyrimidine tract of the CFTR gene
and alternative splicing of CFTR exon 9 .................................................... 60
Figure 2. Detection of IVSS-T genotype by
oligonucIeotide hybridization ........................ .. ..... .. ..... ... ..... .................. . ...... -62
Figure 3. Determination of proportion of exon 91 CFTR mRNA .................................. 65
Figure 4. Evaluation of RNA extraction and cDNA synthesis ........................................ 72
Figure 5. Evaluation of CFTR mRNA ......................................... ............ ... ....................... 74
Figure 6. Quantification of nasal and vas deferens CFTR mRNA ................................ 76
Figure 7. Relationship between CFTR IVSS-T genotype
and proportion of exon 9+ transcripts in
nasal epithelia or vas deferens. ....... ....... ............... ..................................... ... 78
Figure 8. Cornparison of the proportion of CFTR exon 9+
transcripts within nasal epitheiia and
vas deferens from the same subject ...... .. .. .. .. . . .... .. . . ... . . . .... . . ......... ...... ..... .. ... 80
Figure 9. Influence of CFïR genotype on phenotype by
the proportion of normal CFTR mRNA .................................................. 82

ABC
ATP
ARTS
bp
CAMP
CAVD
CBAVD
cDNA
CF
CF-PI
CF-PS
CFTR
CUAVD
DHT
DNA
ciNTP
EDTA
FSH
GAPDH
GnRH
rvF
adenosine triphosphate binding cassette
adenosine triphosphate
assisted reproductive technologies
base pair(s)
cyclic adenosine monophosphate
congenitai absence of vas deferens
congenital bilateral absence of vas deferens
complementary deoxyribonucleic acid
cystic fibrosis
pancreatic-insufficient cystic fibrosis
pancreatic-sufficient cystic fibrosis
cystic fibrosis trammembrane conductance regulator
congenital unilateral absence of vas deferens
dihydrotestosterone
deoxyribonucleic acid
deoxynucleoside triphosphate
ethylenediamine tetraacetic acid
follicle-stirnulating hormone
glyceraldehyde-3-phosphate dehydrogenase
gonadotropin-releasing hormone
in vifro fertilization

rvs
LH
M
MESA
P8
MIE:
min
cl1
ml
PM
mM
mRNA
*g
OD
PCR
RNA
RNase
RT
RT-PCR
S
SD
SDS
intervening sequence (intron)
luteinizing hormone
mole(s)/liter
microsurgical epididymal sperm aspiration
micrograrn(s)
Mullerian inhibitory factor
minute(s)
microliter(s)
milliliter(s)
micromole(s)/liter
millimole(s)/liter
messenger ribonucleic acid
nanogram(s)
optical density
polymerase chah reaction
ribonucleic acid
ri bonuclease
reverse transcriptase
reverse transcriptase-polymerase chah reaction
second(s)
standard deviation
sodium dodecyl sulfate
vii

SSC
TAE
TDF
TdT
TSH
VS.
x2
sodium citrate/sodiurn chloride
Tris-acetate, EDTA buffer
testis determining factor
terminal deoxy transferase
thyroid-stimulating hormone
versus
chi-square
viii

Mrs. Theresa Longley served as study nurse for the project and assisted in specimen
collection, patient counseling, and obtaining patient consent.
Dr. Keith Jarvi fiom the Division of Urology, Department of Surgery, Mount Sinai
Hospital, Toronto provided tissue specimens.
My contribution to this work includes the following:
collection of specirnens
obtaining patient consent
extraction of DNA fiom pet-ipheral leukocytes
genotyping of CFTR IVS8-T tract
extraction of RNA
quantitative RT-PCR experiments
determination of proportion of exon 9+ CFTR transcnpts
design of experimental protocols
9. analysis and interpretation of data

I.A. BACKGROUND
I.A.1. HUMAN MALE INFERTILITY
Approximately fifteen percent of couples are infertile [Mosher, 19851. An
abnormality is found in only the man in 30 percent of the cases, while abnomalities
detected in both partners occur in another 20 percent. Therefore, male factor infertility
accounts for one half of couples with fertility problems [Mosher, 1985; Sigman and
Howards, 19921. Infection, immunologie factors, congenital anomalies, genetic defects,
toxic insult, and sexual dystunction contribute to this important medicai problem. In this
section, the embryology of the male gonad and reproductive tract, the physiology of male
reproduction, and clinical aspects of male infertility will be reviewed.
1.A.l.a. OKERCiIEW OF MALE TESTICULAR AND REPRODUCTM TRACT EMBR YOLOGY
I.A. 1 .a.i. DEVELOPMENT OF TESTES
The gonads are derived from three sources: coelomic epithelium, mesenchyme, and
primordial germ cells Noore, 1989bl. They are f h t recognized during the f i f i week of
gestation when a thickened area of coelomic epithelium develops on the media1 aspect of
the mesonephros. Proliferation of these epithelial cells produces a bulge on the mediai side
of each mesonephros called the gonadal ridge. Primary sex cords grow fiom the gonadal
ridges into the underlying mesenchyme. The indifferent gonad now has an outer cortex and
an inner medulla. The cortex will normally differentiate into an ovary while the medulla

----- - - - -- - - - - , -- -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - i - -
sex chromosome constitution, the medulla norrnally differentiates into a testis and the
cortex regresses. Primordial germ cells migrate along the dorsal mesentery of the hindgut to
the gonadal ridges and become incorporated in the primary sex cords [Witschi, 1948;
Peters, 1 9701.
The short a m of the Y chromosome (Yp) carries the sex determining gene, SRY,
which encodes the testis determinhg factor, TDF. TDF is essential for differentiation of the
gonad into a testis and its absence renilts in formation of the gonad into an ovary [Disteche
et al., 1986; Affara et al., 1987; Page et al., 1987; Palmer et al., 1989; Sinclair et al., 1990;
Berkovitz et al., 19911. TDF pennits the prirnary sex çords to differentiate into
seminiferous tubules. The walls of the seminiferous tubules are composed of Sertoli cells,
derived fiom the surface epithelium, and spermatogonia, derived fiom the primordial germ
cells. The Sertoli cells produce Mullerian inhibitory factor (MF) [Josso, 19861. The
seminiferous tubules become separafe- by mesenchyme that gives rise to the interstitial
cells (of Leydig). These cells produce testosterone [Siiten and Wilson, 1 9741.
I.A. l .aii. DEVELOPMENT OF MALE GENITAL DUCTS
Testosterone stimulates development of the Wolffian duct into the male
reproductive tract while MIF suppresses devetopment of the pararnesonephric (Mullerian)
ducts into female genital ducts Moore, l989bI. When the mesonephros degenerates, some
mesonephric tubules near the testis persist and become efferent ductules which open into
the mesonephric duct, forrning the head of the epididymis. The mesonephric duct proper
separates into its ureteral and reproductive divisions at approximately the seventh week of

- - -- -- - - - -. - - - - . - - - - - - - - - - - - - - - - - Y d Y
the metanephric mesodem. The reproductive portion of the mesonephric (Wolfian) duct
forms the body and tail of the epididyrnis. vas deferens, seminal vesicle, and ejaculatory
duct.
I.A. 1 .a.iii. DEVELOPMENT OF MALE EXTERNAL GENITALLA
Masculinization of the indifferent extemal genitalia results fkom androgens
produced by the fetal testes. As the phallus elongates to form a penis, the urogenital folds
fuse with each other dong the ventral surface of the penis to form the penile urethra.
Consequently, the extemal urethral meatus rnoves to the glans penis. The scrotum is
formed by the fusion of the labioscrotal swellings. Feminization of the indifferent external
genitalia occurs in the absence of androgenic stimulation. The final form of the external
genitaiia is established by the twelflh week of gestation Moore, 1989bJ.
I.A.1.b. OVERCIEW OF MALE REPRODUCTIVE PWYSZOLOGY
The testis has two important iùnctions: spermatogenesis in the seminiferous tubules
and secretion of steroid hormones (androgens) by the interstitial Leydig cells. These are
intimately related, as testosterone synthesis is necessary for both spenn production and
development of secondary sexual characteristics. The two functions are oonûolled by the
anterior pituitary through the secretion of gonadotropins. The anterior pituitary itself is
regulated by gonadotropin-releasing hormone (GnRH) fkom the hypothalamus. The
hypothalamic-pituitary-testicular axis consists of a closed-loop feedback control mechanism
for maintaining normal reproductive fiction.

The hypothalamus receives messages from the central nervous system and the testis
to regulate ihe synthesis and secretion of GnRH, which stimulates the synthesis and release
of both luteinizing hormone (LH) and follicle-stimulating hormone (FSH) [Schally et al.,
19711. These two hormones are synhesized in the anterior pituitary and are secreted
episodically in response to the pulsatile release of GnRH. LH and FSH bind to specific
receptors on the membrane of Leydig cells and Sertoli cells, respectively, to stimulate
cellular metabolism.
The hypothalamic-pitulw-gonadal axis consists of a closed-loop feedback control
mechanism. Testosterone, secreted by the Leydig cells in testes, inhibits LH secretion in
males [Walsh et al., 19731. It is metabolized in peripherai tissue to dihydrotestosterone
(DHT) or estradiol. Secretion of FSH is inhibited by inhibin, a nonsteroidal compound
produced by the Sertoli cells p a n Thiel et al., 19721.
I.A. 1 .b.ii. TKE TESTIS
I.A. 1 .b.ii.a. Leydig Cells
Testosterone is secreted episodically fiom the Leydig cells in response to LH pulses
and has a diurnal pattern, with the peak level in the early morning and the nadir in the
evening [Lipsett, 19741. The biotogic effects of androgens are exerted on target organs that
contain a specific androgen receptor protein in the ce11 cytosol. Testosterone enters target
cells where it may be converted to the more potent androgen, DHT, by Sa-reducbse, and
either testosterone or DHT binds to a receptor protein. This bound receptor complex is
ûanslocated into the nucleus, where it binds to nuclear chromatin and resdts in the

mRNA then exerts its androgen action in the target ce11 [McClure, 19871.
The major functions of androgen in target tissue include regulation of gonadotropin
secretion by the hypothalarnic-pituitary axis, initiation and maintenance of spermatogenesis,
differentiation of the interna1 and extemal male genital system d d n g fetal development,
and promotion of sexual maturation at puberty.
I.A. 1 .b.ii.b. Seminiferous Tubules
Seminiferous tubules contai. germ cells and Sertoli cells. The latter are nondividing
support cells found on the basement membrane of the seminiferous tubules and extend
filamentous cytoplasmic ramifications toward the tubular lumen. Sertoli cells are linked by
tight junctions that divide the seminiferous tubule into a basal and adluminal cornpartment.
These tight junctions dong with the myoid cells of the peritubular contractile ce11 layer,
serve to form the blood-testis barrier wussell, 19801. Sertoli cells are believed to provide
structural and metabolic support for the developing spermatogenic cells. They are also
involved in the phagocytosis of excess cytoplasm cast off by spermatids [Wheater et al.,
19871.
The germ cells are arranged in an orderly manner fiom the basement membrane to
the lumen. Spermatogonia lie d'mtly on the basement membrane, and found progressing
centrally are prirnary spermatocytes, secondary spermatocytes, and spermatids [Clermont,
1963; Heller and Clermont, 19641.

Spematogenesis is the process in which primitive stem cells either divide to renew
themselves or produce daughter cells that will become spermatocytes. The most primitive
undifferentiated spermatogonia are the stem cells. For stem ce11 renewal, the dark type A
spematogonia foltowing mitotic division produce a fiesh stock of type A cells as well as
pale type A spermatogonia. The latter undergo mitotic divisions into preleptotene primary
spermatocytes via type B spermatogonia [Clermont, 19721. The primary spermatocytes
undergo the first maturation division by meiosis, reducing the number of chromosomes
fiom 46 to 23 [Kerr and deKretser, 198 Il. Each prirnary spermatocyte gives nse to two
secondary spermatocytes, and each of these divide into two spermatids. The spermatids
then transfom into spexmatozoa, a process called spermiogenesis. This process includes
nuclear condensation, acrosome formation, loss of cytoplasm, development of a tail, and
arrangement of mitochonciria into the middle piece of the sperm. The entire spermatogenic
process requires approximately 64 days to complete [Clemonf 19721.
I.A. 1 .b.iii. HORMONAL CONTROL OF SPERMATOGENESIS
LH indirectly affects spermatogenesis in that it stimulates endogenous testosterone
production p f i r g a and Sherins, 198 11. Sertoli cells, possessing specific hi&-aninty
FSH receptors, are the target for FSH Neans et al., 1980; Ritzen et al., 19811. Androgen-
binding proteins produced by Sertoli cells carry androgens intracellularly [Ritzen et al.,
1971; Hansson and Djoseland, 19721. The close proxirnity of the Leydig cells to the
seminiferous tubule and elaboration by the Sertoli cells of androgen-binding protein cause a

sperrnatozoa.
I.A. 1 .b.iv. SPERM MATURATION, STORAGE, AND TRANSPORT
While the testis is responsible for sperm production, the epididymis is involved in
the maturation, storage, and transport of spennatozoa. Spermatozoa develop an increased
capacity for motility pedford et al., 1973; Moore et al., 19831 and fertilization [Orgebin-
Cnst, 1969; Bedford et ai., 1973; Hinrischsen and Blaquier, 1980; Moore et al., 1983;
Bedford, 19881 as they migrate through the epididymis.
The epididymis consists of a single convoluted tubule, 3 to 4 metres long, and is
divided into the caput (head), corpus (body), and cauda (tail) epididymis [Jenkins et al.,
19783. Sperrn are transported by a hydrostatic pressure difference, ciliary propulsion, and
peristaltic contraction of myoid cells dong the epididymis [Johnson and Howards, 19761.
The epididymis aiso functions as a storage reservoir for sperm [Johnson and Varner, 19881.
Mature sperm fkom the cauda epididiyrnis enter the vas deferens, which transports
its contents by peristaltic motion into the ejaculatory duct [Bruschini et al., 19771. Sperm
are then propelled to the outside by emission and ejaculation. Secretions fiom the seminal
vesicles and prostate are deposited into the postenor urethra during emission. Peristaisis of
the vas deferens and contraction of the bladder neck are under sympathetic control of the
autonomie nervous system [Bell and McLean, 1 967; Owman and Sjo berg, 1 972; Lipshultz
et al., 19911. During ejaculation, the extemal sphincter relaxes and the semen is propelled
through the urethra by rhythrnic contractions of the perineal and bulbourethral muscles, both
under somatic control.

(20%). The seminai vesicles produce fructose, prostaglandins, and coagulating factors
[Mann and Mann, 19811. Seminal plasma acts as a buffer on the acidic vaginal secretions.
The coagulum formed by the ejacuiated semen liquefies within 20 minutes from the action
of prostatic proteolytic enzymes. More fluid are contributed by the bulboureùiral glands
(Cowper's glands) and urethrai glands (Littre's glands) through the penile urethra. The first
portion of the ejaculate contains most of the spermatozoa and secretions fiom the
bulbourethral glands and the prostate; the remaining portion consists mostly of seminal
vesicle secretions and contains only a few spermatozoa milja et al., 19871.
I.A. 1 . b.v. FERTILIZATION
Fertilization normally occurs in the fallopian tubes after ovulation. During mid-
cycle of the menstnial period in the fernale, cervical mucus becomes more abundant,
thinner, and watery, thus allowing entry of spem into the uterus and also protecting spem
fiom the acidic vaginal environment parnes and Toot, 19861. Spermatozoa must undergo
capacitation within the female reproductive tract in order for fertilization to take place
[Chang, 195 1; Austin, 195 11. Capacitation is followed by the acrosome reaction. The
latter is a progressive fusion between the b e r and outer acrosome membrane, resuiting in
the release of enzymes important for oocyte penetration by the spemi Noore, 1989aJ.
LA.1 .c. CL13VICAL ASPECTS OF W E llVFERTILITY
Pregnancy occurs in the majority of couples within one year of natural unprotected
intercourse. The term infertility applies to those couples who have not been able to
conceive d e r this one year period. Both partners should be evaluated for factors that may

entirely to a male factor, and an additional 20% involve both males and female factors; thus,
about one half of infertile unions is due to a male factor [Mosher, 1985; Sigman and
Howards, 19921.
I.A. 1 .ci . EVALUATION OF THE INFERTILE MALE
The work-up of a man presenting with infertility should consist of a detailed history
and physical examination. The latter must include inspection and palpation of the scrotum
to ensure that the testicles have properly descended into the scrotum and are normal in size
and consistency, and that the epididyrnides and vasa deferentia are present. Abnormal or
suspicious findings fiom the history and physical examination direct subsequent
investigations, such as serum hormonal tests (e-g., TSH, LH, FSH, testosterone, prolactin)
and/or radiological imaging studies (e.g., scrotal ultrasound, transrectal ultrasound).
However, al1 men presenting with infertility should have a semen analysis.
I.A. 1 .c.i.a. Semen analysis
The semen analysis is the cornerstone of the infertility investigation [S ipan and
Howards, 19921. Normal parameters are show in Table 1 [World Health ûrganization,
1 9871. Major abnonnali ties in semen analysis include the following :
asthenospermia (-30% progressively motile sperm)
oligospemiia (QO x 1 o6 sperm 1 mL)
pyospermia (>1 x 106 white blood ce11 1 rnL)
azoospennia (no sperm)

teratospermia (abnormal spem morphology)
I.A. 1 . c i . ETIOLOGIC CLASSIFICATION OF MALE INFERTILITY
The causes of mde infertility may be classified according to results of the semen
analysis. Approximately 10% of men presenting with infertility will have azoospermia,
whereas the remaining 90% will have oligospemia, asthenospermia or teratospermia
[Lipshultz, 19801. Of the patients with azoospermia, standardard laboratory and irnaging
studies can determine whether this abnormality is due to a pre-testicular (hypothalamic-
pituitary pathology, -10% of azoospermie men), testicular (spermatogenic deficiency
including testicular failure, -40%), or pst-testicular (functional or structural obstruction to
spem outflow, -50%). Specific causes of pst-testicular avoospermia include congenital
absence of vas deferens (CAVD), epididymal obstruction, and vasectomy.
I.A. 1 .c.iii. TREATMENT OF MALE INFERTILITY
Obviously, treatment of an individual male patient with infertility will depend on the
underlying cause. Those with a pre-testicda. cause can ofien be treated successfûlly with
exogenous GnRH analogues or gonadotropins mowards, 19951. Advances in rnicrosurgery
and assisted reproductive technologies (ARTS) now make it possible for rnany infertile men
with a testicular or pst-testicular cause to father their own children. Men with previous
vasectomies wishing to reverse their sterile status can undergo microsurgical
vasovasostomy (re-connecting the vas deferens) or, if necessary, vasoepididymostomy
(connecting the vas deferens to the epididyrnis). The latter procedure is also indicated in
cases of acquired (usually due to infection) or idiopathic epididymal obstruction. Men with

CAVU can be ettectively treated with microsurgicai epididymal sperm aspiration
(MESA) and routine in vitro fertilization (NF) or in vifro fertilization-intracytoplasmic
sperm injection (IVF-ICSI) [Temple-Smith et al., 1985; Silber et al., 1988; Palenno et al.,
1992; Silber et al., 1995; Schlegel et al., 1995; Gil-Salom et al., 19951. Patients with a
testicular cause for their infertility such as testicular failure may also be treated with IVF-
ICSI via testis-retrieved haploid sperm [Van Steirteghem et al., 1993; Vanderzwalmen et
al., 1995; Fishel et al., f 995; Fishel and Thornton, 1995; Harari et al., 1995; Silber et al.,
1995; Sherins et al., 1995; Tesarik et al., 19951. However, it must be emphasized that
ARTS should not be initiated until the patient and his partner have received rigorous
genetic counseling on the possibility that the genetic defect responsible for his
reproductive failure may be transrnitted to the offspring [de Kretser, 1995; Ln't Veld et al.,
1995; Mak and Jarvi, 1996; Morris and Gleicher, 19961.
I.A.2. CYSTIC FIBROSIS AND THE CYSTIC FIBROSIS TRANSMEMBRANE CONDUCTANCE RIEGULATOR
Cystic fibrosis (CF) is the most common fatal autosomal recessive disorder in the
Caucasian population, with an incidence of approxirnately 1 in 2,500 live births and a
carrier fiequency of 1 in 25 persons in populations of Northem European descent. The
classical form of CF is characterized by abnormalities of electrolyte, fluid and
macromolecule secretion of exocrine glands. Clinical hallmarks of CF include chronic
pulmonary obstruction and infections, exocrine pancreatic insufficiency, neonatal
meconium ileus, elevated sweat electrolytes, and male infertility [Welsh et al., 19951.
The gene responsible for CF, called cystic fibrosis transmembrane conductance
regulator (CFTR), was identified and cloned in 1989 [Rommens et al., 1989; Riordan et

- - -
-230 kilobases (kb) of genomic DNA, and produces a mRNA transcnpt of 6.2 kb. The
encoded protein contains 1,480 arnino acids and is a member of the ATP-binding cassette
(ABC) family of membrane proteins. Various studies support that CFTR is a CAMP-
regulated chloide channel found in the apical membrane of secretory epithelial cells [Welsh
et al., 19951.
Hundreds of mutations in the CFTR gene have been reported [Cystic Fibrosis
Genetic Analysis Consortium, 19941. These mutations may be classified as follows: class
1 mutations confer defective protein production; class II mutations involve defective
protein processing; class III mutations involve defective channel regdation; class IV
mutations involve defective channel conduction; and class V mutations lead to decreased
protein synthesis [Zielenski and Tsui, 19951. In general, class 1, II or HI mutations are
expected to have more serious phenotypic consequences than class IV or V mutations.
I.A.3. CYSTIC FIBROSIS TRANSMEMBRANE CONDUCTANCE REGULATOR AND MESONEPHRIC DUCT ANOMALIES
Studies have shown that more than 95% of CF men have abnorrnalities in the
structures derived fiom the Wolfian duct [Kaplan et al., 1968; Valman and France, 1969;
Landiing et al., 1969; Holsclaw et al., 1971 ; Taussig et ai., 19721. The body and tail of
epididymis, vas deferens, seminal vesicles and ejaculatory ducts are atrophic, fibrotic, or
completely absent. As a result, semen analysis typically reveals decreased volume,
complete absence of spermatozoa (azoospermia), increased acidity, decreased fructose
concentration, elevated levels of citric acid and acid phosphatase Kaplan 19681.
However, spermatogenesis is present on testicular histology [Denning, 19681.

Wolffian duct abnormalities but no other manifestations of CF. These conditions may
represent either a reproductive tract specific or a very mild form of CF. At present,
congenital bilateral absence of the vas deferens (CBAVD), congenital unilateral absence
of the vas deferens (CUAVD), and idiopathic epididymal obstruction have al1 been
associated with CFTR gene mutations [Durnur et al., 1990; Anguiano et al., 1992; Mickle
et al., 1993; Osborne et al., 1993; Patrizio et al., 1993; Culard et al., 1994; Oates and
Amos, 1994; Casals et al., 1995; Chillon et al., 1995; Costes et al., 1995; Durieu et al.,
1995; Jarvi et al., 1995; Mercier et al., 1995; Zielenski et al., 1995b; Schlegel et al., 1996;
Donat et al., 19971.
CBAVD is found in 1-2% of men presenting with infertility [Dubin and Amelar,
197 1 ; Greenberg et al., 1978; Jequier et al., 19851. Pnor to the identification of the CFTR
gene, CBAVD was considered to be a distinct clinical and genetic entity with an
autosomal recessive inheritance pattern [McKusick, 19921. Genetic studies found that
5042% of CBAVD men have at least one detectable CFTR gene mutation and that about
15% of these men have two detectable CFTR mutations [Durnw et al., 1990; Anguiano et
al., 1992; Osborne et al., 1993; Paûizio et al., 1993; Culard et al., 1994; Oates and Amos,
1994; Casals et al., 1995; Chillon et al., 1995; Costes et al., 1995; Durieu et al., 1995;
Jarvi et al., 1995; Mercier et al., 1995; Zielenski et al., 1995b; Schlegel et al., 1996;
Donat et al., 19971. CFTR gene mutations have also been reported in men with
congenitai unilateral absence of vas deferens (CUAVD) [Mickle et al., 1993; Schlegel et
al., 1 9961. Of 14 infertile patients with CUAVD tested for 20 CFTR mutations, 6 (43%)
had one mutation, and, in fact, one of these patients had two brothers with CBAVD

CFTR gene mutations was further extended by the observation that 47% of otherwise
healthy men with idiopathic epididymal obstruction have a CFTR gene mutation [Jarvi et
al., 19951. These findings suggest that a broad spectrurn of Wolffian duct abnormalities
(absence of the vas deferens, obstruction of the epididymis, ejaculatory duct cysts,
seminal vesicle cysts, etc.) may be associated with CFTR gene mutations.
Alterations in the CFTR gene identified in men with Wolffian duct abnomalities
can be divided into four groups. The first group consists of mutations that have been
identified in classical CF patients. These "severe" CFTR mutations associate with
pancreatic insufficiency (CF-PI) and are generally class 1-III mutations. Examples of such
mutations reported include AF508, G542X, R553X, R1 l62X, W l282X, N1303K, 1717-
1 G+A, and 2 1 84delA. The second group consists of mutations found in more benign
presentations of CF. These "mild" CFTR mutations are associated with pancreatic
sufficiency (CF-PS) and tend to be class N-V mutations. Such mutations include R117H
and R347P. The third group consists of novel mutations identified exclusively in some
CAVD men; however, as these sequence alterations are extremely rare, it is only
speculated that they contribute to the CAVD phenotype [Anguiano et al., 1992; Culard et
al., 1994; Mercier et al., 19951. The fourth group consists of CFTR gene alterations that
were previously considered as benign sequence variations [Anguiano et al., 1992; Mercier
et al., 1995; Zielenski et al., 1995aI. Such examples include R75Q, G576A, R668C, and
5T. in the majority of patients (-45-50%), only one mutation or sequence alteration has
been identified despite exhaustive screening of al1 exons and immediate flanking intron
sequences in the CFTR gene. For those with mutations identified in both CFTR alleles

It is reasonable to assume that othewise healthy men with CAVD with one or no
detectable CFTR mutation may harbour as yet unidentified CFTR gene mutations or
variants. However, not al1 cases of congenital Wolffian duct anomalies are due to
mutations in the CFTR gene. There is evidence that CUAVD or CBAVD men with
concomitant renal aplasia or ectopia (not typical features of CF) do not result fiom CFTR
gene mutations. Augarten et al. reported that of 47 CBAVD patients, 10 had rend
malformations on ultrasonography [Augarten et al., 19941. Of the 37 patients with
normal urinary tract, 18 (49%) tested positive for CFTR mutations. Interestingly, none of
the 10 individuals with renal malformations had mutant CFTR aileles or abnonnal sweat
tests. in a separate report, 12 of 70 men with CUAVD or CBAVD had renal anomalies
on ultrasound or excretory urography, and al1 12 tested negative for CFTR mutations. Of
58 patients without rend abnormalities, 46 underwent CFTR mutation analysis and 27
(59%) tested positive for mutations [Schlegel et ai., 19961. Therefore, cases with vasal
agenesis and rend malformations probably do not represent a genital form of CF but
rather a distinct clinical entity.
The aforementioned information provides insight into the timing for the effect of
the CFTR mutation on the reproductive tract. The mesonephric duct separates into its
ureteral and reproductive divisions at approximately week seven of gestation. The
ureteral bud induces rend formation as it grows into the metanephric mesoderm, while
the reproductive portion (Wolfian duct) forms the corpus and cauda epididymis, vas
deferens, serninal vesicle, and ejaculatory duct [Moore, 1989bl. It has been proposed that
in CUAVD or CBAVD not caused by a defect in the CFTR gene, anomalous changes to

-
and genital systems are afFected [Augarten et ai., 19941. In CUAVD or CBAVD patients
with CFTR mutations. changes to the Wolffian duct are believed to occur after the urinary
system has separated fiom the reproductive system; therefore. no renal malformation
resuits. n e pathogenesis probably involves intrauterine obstruction of the Wolffian duct
with inspissated secretions which results from CFTR chloride channel dysfunction, and
gives rise to a dehydrated intraluminal fluid content with secondary degeneration or
obliteration of the Wolffian duct [Taussig et al., 19721. However, a primary
morphogenetic defect in development of the Wolfian duct cannot be excluded [Olson
and Weaver, 19691.
Electrophysiological experiments on cultured hurnan epididymal cells using the
patch-clamp technique demonstrated the presence of a low-conductance chloride channel
that was activated by addition of CAMP agonists to these cells pollard et al., 19911. This
channel most likely represents CFTR. Furthemore, expression studies in human fetal
and adult reproductive tract by in situ hybndization showed that CFTR is expressed in the
ductal epithelium by week 18 of gestation and th is expression is maintained throughout
pst-natal life [Trezise et al., 1993; Tizzano et al., 19931. CFTR expression is high in the
head of the epididymis, low in the body and tail of the epididymis, low in the vas
deferens, and undetectable in the seminal vesicles [Tizzano et al., 19941. The decreased
expression of CFTR in Wolffian denvatives suggests that these structures may be more
sensitive to CFTR dysfunction than other organs.

SPLICING OF CFTR EXON 9
One of the first forms of alternative splicing discovered in CFTR was the deletion
of exon 9 (exon 9-) [Strong et al., 1991 ; Chu et al., 19911. Exon 9 encodes the initial
21% of the first nucleotide binding fold (NBFI) of CFTR, including motif A of the
Walker ATP binding consensus sequence [Walker et al., 19823. Based on quantitative
RT-PCR experirnents performed on respiratory epithelia, the amount of exon 9-
transcripts was found to Vary between different individuals [Bremer et al., 1992; Chu et
al., 1991; Chu et al., 19921. The variability in exon 9 splicing efficiency can be
explained, for the most part, by a variation in the polypyrimidine tract of the splice
acceptor site in intron 8 of CFTR (IVS8-T tract) [Chu et al., 19931. Sequencing of the
splice acceptor site preceding exon 9 demonstrated that three variations of uninterrupted
pyrimidines occur in the population: 5,7, and 9 thymidines (5T, 7T, and 9T) [Chu et al.,
19911. It was also observed that a positive correlation existed between the length of
IVS8-T tract and the proportion of branchial epithelial CFTR transcripts with exon 9
intact (exon 9+) (Fig. 1) [Chu et al., 19931.
The abundance of exon 9- CFTR eanscripts in respiratory epithelial cells lead two
groups of investigators to determine whether it encoded a functional isoform of CFTR.
Transduction of CFPAC (pancreatic adenocarcinorna ce11 line from a CF patient) or HeLa
cells with the CFTR cDNA lacking exon 9 failed to confer CAMP-regulated chloride
conductance [Delaney et al., 1993; Strong et al., 19931. Protein analysis of transduced
cells detected mutant CFTR that was immature and incompletely glycosylated in both
studies, suggesting that exon 9- CFTR mRNA transcripts are û-anslated but the protein

- - - -
protein probably undergoes degradation within the endoplasmic reticulum. Expression of
the CFTR missing exon 9 encoded amino acids in Xenopus oocytes incubated at room
temperature also did not produce a CAMP-regulated chloride conductance. This finding
is important as other CFTR mutants that are improperly processed at 37OC appear to fold
properly and regain function at lower temperatures [Drumm et al., 1991; Denning et al.,
19921. Although some have argued that the protein made by the exon 9- transcript may
have physiological function [Bremer et al., 19921, the lack of conservation of exon 9-
transcripts in mice suggests otherwise [Delaney et al., 19931. Thus, CFTR missing the
region of NBFl encoded by exon 9 does not function as a chloride channel and likely
does not have any intracellular function.
I.A.5. THE 5T VARIANT IN INTRON 8 OF THE CFTR GENE AND CONGENITAL ABSENCE OF VAS DEFERENS
Studies have shown that the fiequency of the 5T variant was significantly higher
in men with CBAVD (46%), CUAVD (25%), or epididymal obstruction (29%) than the
general population (5%) [Chillon et al., 1995; Costes et al., 1995; Jarvi et al., 1995;
Zielenski et al., 1995bl. Indeed, the 5T variant is the single rnost fkequent CFTR gene
sequence alteration found in CBAVD men (Table 2). The presumed role of the 5T allele
in the pathogenesis of CBAVD is supported by two separate studies showing that the
frequency of the 5T allele is lower in fathers (2.1 % and 1.9%) than in mothers (5.2% and
4.7%) of CF patients [Chillon et al., 1995; Zielenski et al., 1995bl. In fact, Chillon et al.
reported four fathers who had one CFTR gene with the 5T ailele and the other with a
severe CFTR mutation (G542X, Nl3O3KY 18 12-1 G+A, or 936delTA) [Chillon et al.,

and are phenotypically normal. Three hypotheses could explain the strong but incomplete
association between the 5T allele and CBAVD. First, there could be a non-random
association between 5T and CBAVD, with the allele segregating with but not being
causal for the CBAVD phenotype; however, this hypothesis is probably untrue as the
analysis of several intragenic DNA markers (Le., within the CFTR gene) in the four
fathers and in patients with CBAVD showed that several haplotypes are associated with
the 5T allele [Chillon et al., 19951. Second, the 5T variant may have a partially causal
role, together with other CFTR gene mutations. The presence of another mutation in the
same CFTR gene as the 5T allele is unlikely since al1 coding regions and their
immediately flanking intron sequences have been screened [Chillon et ai., 1995; Costes et
al., 1995; Zielenski et al., 1995b3. However, this does not d e out mutations within the
promoter region or introns of the CFTR gene. Third, the ST variant could have a causal
role in CBAVD, with other factors (genetic andor environmental) accounting for these
exceptional cases without CBAVD (Le., the four fathers with the CFTR rnutatiod5T
genotype). The association of the 5T allele with lower levels of normal, full-length
CFTR mRNA supports the hypothesis that this variant generally causes CBAVD,
especially when a CFTR mutation is present in the other copy of the gene. Nevertheless,
the association between the 5T allele and CBAVD is not absolute. Taking into account
the predicted frequency (1 in 222 men) and the reported iiequency (-1 in 1,000 men) of
CBAVD, it has been estirnated that the penetrance of the 5T allele is -0.6 wielenski et
al., 1995bl. While confirmation of this estirnate awaits M e r studies involving larger

penetrance.
I.B. HYPOTHESIS
An intriguing question pertains to the mechanism by which dterations in CFTR
expression results in reproductive tract abnormalities in the absence of pathologie
changes in other CF-associated organs (e.g., lung, nasal sinus, pancreas, liver). It is
possible that the reproductive tract is more sensitive to CFTR dysfunction than the other
tissues. Alternatively, the level of functional CFTR may vary between different tissues,
with the lowest expression in the Wolfian structures. Since the 5T variant of the NS8-T
tract of the CFTR gene is associated with inefficient splicing of exon 9 and has been
identified in a signifiant proportion of infertile men with isolated Wolffian duct
anomalies, it is proposed that the splicing efficiency of CFTR exon 9 is lower in the
reproductive tract than the other CF-associated organs.
I.C. PURPOSE
The purpose of my work was to compare the proportion of exon 9+ CFTR mRNA
transcripts within nasal epithelia and vas deferens fiom the sarne individual, and to
examine the relationship between the IVS8-T tract and the level of exon 9+ CFTR rnRNA
in these tissues.

1I.A. PATIENTS AND SAMPLES
Twenty-four healthy men with a previous vasectomy presenting to a male
infertility clinic for sterilization reversai were recniited for the study (mean
agefSD4Of3 years). This study was performed with the approval of the Human
Subjects Review Cornmittee of the University of Toronto, and informed consent was
obtained fiom each subject. None of the patients had a personai or family history of
pulmonary or gastrointestinai manifestations suggestive of CF. Each patient underwent
scrotal surgery (i.e., either vasovasostomy (re-comecting the vas deferens) or
vasoepididymostomy (connecting the vas deferens to the epididymis)). Intraoperatively,
while under general anesthesia, in each case the following were obtained: peripherai
venous blood by venipunchire for analysis of the NS8-T tract genotype, nasal epithelia
by a sterile swab and a small piece of vas deferens (-5- 10 mm, which would ordinarily be
discarded) by excision for quantitative RT-PCR analysis. Nasal samples were placed into
a 2 ml microfuge tube containîng 500 pl of phosphate buffered saline and vasal sarnples
were placed into a 2 ml microfuge tube, snap-fiozen in liquid nitrogen, and stored at -
70°C if not analyzed immediately.
1I.B. ALLELE SPECIFIC OLIGONUCLEOTIDE ANALYSIS OF IVSS-T TRACT
Genomic DNA was isolated fiom peripheral blood lymphocytes according to
standard protocols [Miller et al., 19881. Exon 9 including the WS8-T tract was amplified
by the polyrnerase chah reaction (PCR) mullis et al., 19861 with primers 9i-5 (5 ' -
TAATGGATC ATGGGCC ATGT -3 ' ) and 9i-3 (5 ' -AC AGTG'TTGAATGTGGTGC A-3 ' ),

follows: 94OC for 2 min; denaturation at 94OC for 20 S. annealing starting at 60°C then
auto-down OS°C/s over 20 s, and extension at 72OC for 30 s, for 20 cycles; followed by
denaturation at 94OC for 20 s, annealing at 50°C for 20 s, and extension at 72OC for 30 s,
for 15 cycles; followed by one final cycle of denaturation at 94OC for 20 s, annealing at
50°C for 20 s, and extension at 72OC for 7 min. The reaction mixture contained 5 pl of
PCR buffer (10 mM Tris-HC1 (pH 8.3), 50 rnM KCl, 1.75 mM MgC12, 0.001% gelatin), 5
pl of dNTP's (2 m M each of deoxyadenine triphosphate, deoxycytidine triphosphate,
deoxyguanosine triphosphate, and deoxyrhymidine triphosphate; Pharmacia); 50 ng of
each primer; and 0.5 unit of Taq DNA polymerase (Perkin Elmer/Cetus) in a final volume
of 50 pl, containing 50-100 ng of genomic DNA. Al1 PCR amplifications were
performed with the GeneAmp PCR System 9600 (Perkin ElmerlCetus). Ten pl of PCR
arnplified DNA was denatured and vacuum-blotted to nylon membrane (Hybond-N+;
Amersharn). The following allele specific oligonucleotides were used: ST (5 ' -
GTGTGmAACAGG-3') , 7T (5'-GTGTGTTTTTTTAACAG-3'), and 9T (5'-
TOTGmAAC AG-3'). Each oligonucleotide was labeled with 3 2 ~ by terminal
transferase (Pharmacia), and hybridization was performed ovemight at 42OC and the
washings were done once with 3xSSC/O.l%SDS at room temperature for 20 min, and
twice with 0.2xSSC at 36OC for 20 min. Amplified DNA fiom three individuals
previously s h o w to have the 5T/7T, 7T/7T, or 9T/9T genotype were used to control for
oligonucleotide specificity [Jarvi et al., 19951.

For reconstruction studies, a segment of exon 9+ cDNA was obtained fiom a
plasmid containing full-length CFTR cDNA [Rommens et al., 19911 following two
rounds of nested PCR. First round PCR was performed with plasmid wild-type CFTR
cDNA, 5' primer X5-5 (5'-GCTGTCAAGCCGTGTTCTAG-3'' in exon 5) and 3' primer
13i-3sA (5'-TGGTCGAAAGAATCACATCC-3', in exon 13) for 35 cycles (94OC, 20 s;
60°C, 20 s; 72OC, 30 s) (Fig. 3A). The reaction mixture contained 5 pl of PCR buffer (10
rnM Tris-HCl (pH 8.3), 50 mM KCl, 1.75 m M MgCI2, 0.001% gelatin), 5 pl of dNTP's
(2 rnM of each of the four nucleotides triphosphate; Pharmacia); 200 ng of each primer;
and 0.5 unit of Taq DNA polymerase (Perkin ElmerKetus) in a final volume of 50 pl,
containing 400 ng of plasmid wild-type CFTR cDNA. Nested PCR was performed on 115
of the reaction product fiom the first round under identical PCR conditions, except that 5'
primer 7i-5s (5'-TTCAATAGCTCAGCCTTC-3', in exon 7) and 3' primer X12-3 (5'-
GTTAAAACATCTAGGTATCC-3', in exon 12) were used. The PCR products were
size fiactionated on 1% agarose gel in 1 X TAE (40 mM Tris-acetate; 1 mM EDTA, pH
8.0). The exon 9+ cDNA fhgments were extracted fiom the agarose gel and purified
according to the manufacturer's protocol (QIAquick Gel Extraction KitlQLAGEN). Exon
9- cDNA fragments were isolated fkom commercially-obtained hurnan lung total RNA
(Clontech) after conversion to cDNA and amplification by two rounds of nested PCR as
described above, except the exon 9- hgments were extracted fiom the agarose gel and
purified (Fig. 3B).

POLYMERASE CHAIN REACTION
Nasal epithelial cells were collected by centrifugation and lysed in 800 pl of
TRIzot Reagent (Gibco/BRL). Vas deferens sarnples were homogenized with a rotor-
stator homogenizer (PRO 200 Handheld Homogenizer, PRO Scientific Inc.) in 800 pl of
TNzol Reagent. Total RNA was extracted fiom the lysates according to the
manufacturer's instructions (Gibco/BRL). First strand cDNA was synthesized fiom total
RNA using Superscript II RNase H- reverse transcriptase (RT) and random
hexanucleotide pnmers. The reaction (21 pl) contained up to 1 1 pl of total RNA, 2 pl
lOxPCR buffer (100 mM Tns-HCl (pH 8.4), 500 rnM K I ) , 2 pl MgCl2 (25mM), 1 pl 10
m M dNTP mix, 2 pl DTT (0.1 M), 1 pl random hexamers (5Ong/pL), 1 pl Superscnpt II
RT (200 unitslpl), and 1 pl E. coli RNase H (2 unitdpl), with al1 components obtained
comrnercially fiom Gibco/BRL.
To ensure that total RNA was successfidly extracted and synthesized into cDNA,
rnRNA of the human housekeeping gene glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) [Tokunaga et al., 19871 fiom each sample was amplified using the PCR
conditions described below. Primers used in the reaction were S'primer GAPDHSB (5'-
GGTCGGAGTCAACGGATTTGGTCG-3') and 3' primer GAPDH3B (5' -
CCTCCG ACGCCTGCTTCACCAC-3 '). The ampli fied products (78 8 bp) were size
fractionated by agarose gel (1.5%) electrophoresis, transferred to Hybond N+ nylon
membrane (Amersham) by the rnethod of Southem [Southem, 19751 and evaluated with a
32~-labeled human GAPDH cDNA probe (GAPDH 5A; 5'-
CCAATATGATTCCACCCATG-3') interna1 to the amplified region. The resultant

3 5 SoftwareProtein Databases Inc).
AAer being adjusted with GAPDH mRNA transcnpt level for each individual
sample, transcripts of the CFTR gene were amplified by PCR. The oligonucleotide
primers included 5 ' primer X8-5 (5 '-ACGACTACAGAAGTAGTGATGGAG-3 ' , in
exon 8) and 3' primer C 16D (5'-GTT'GGCATGCTITGATGACGCTTC-3', in exon 10)
(Fig. SA), and amplification was performed as follows: 94OC for 2 min; denahiration at
94OC for 20 s, annealing at 5S°C for 20 s, and extension at 72OC for 30 s, for 30 cycles;
followed by one final cycle of denaturation at 94OC for 20 s, mealing at 50°C for 20 s,
and extension at 72OC for 7 min. The PCR products were blotted to nylon membrane and
hybridized with a CFTR cDNA oligonucleotide probe derived fiom exon 10, which
would anneal to both exon 9+ and exon 9- fragments (C16B; 5'-
GT'ITTCCTGGATTATGCCTGGCAC-3') (Fig. 5B). The expected size of the exon 9+
and exon 9- products were 4 16 bp and 233 bp, respectively. To ensure that the 4 1 6 bp
and 233 bp fragments respectively represented exon 9+ and exon 9-, two oligonucleotide
probes were designed: 9i-5s (5'-ACAGGGATITGGGGAATTATTTG-3'), a sequence
within exon 9, and ex8/10 (5'-TGGGAGGAGACTTCACTT-3'), a sequence which spans
the 3' end of exon 8 and the 5' end of exon 10 (Fig. SB).
The absence of contaminants in RT-PCR assays was regularly assessed by
controls that did not contain any cDNA template, starting RNA, or RT enzyme.

Nasal epithelial and vas deferens total RNA were extracted, converted to cDNA,
amplified by PCR with CFTR primers, and subjected to Southem analysis using a '*P-
labeled exon 10 probe (C 16B) as described in Section H.D. The resulting autoradiograph
was analyzed by scanning densitometry, and the proportion of exon 9+ transcripts, as a
percentage of total CFTR transcripts, was denved by using the total densitometric units of
both transcripts as 100%.
As standards for quantification of exon 9+ and exon 9- mRNA species, the
isolated exon 9+ and exon 9- cDNA fragments (see Section II.C.) were serially mixed in
varying, known quantities and subjected to the same PCR conditions as for the nasal and
vas deferens samples as outlined in Section U.D. Quantification of exon 9+ and exon 9-
was then determined as described above. The results were plotted and graphed. Al1
experimental proportions of exon 9+ and exon 9- CFTR transcripts were accordingly
adjusted, based on this graph.
1I.G. ANALYSIS OF DATA
Differences in IVS8-T allele frequencies between study subjects and the general
population were compared by the X2 statistic or Fisher's exact test. The mean proportions
of CFTR exon 9+ mRNA transcripts associated with the different WS8-T genotypes were
compared uing analysis of variance (ANOVA). Post-hoc pairwise comparisons between
the different means were evaluated by the Bonferroni test. The correlation between the
proportion of exon 9+ transcripts and length of the IVS8-T tract was analyzed by the
Spearman rank correlation. Intra-individual nasal and vas deferens levels of exon 9+

cornparisons and those less than 0.05 were considered to indicate statistical significance.
Al1 statisticai analyses were performed using SPSS for Windows 1994.

1II.A. IVSS-T TRACT GENOTYPE
Leukocyte genomic DNA was isolated and amplified by PCR using CFTR
primers which encompass the IVS8-T tract and exon 9 region (Fig. 2A). Subsequent
ethidium bromide staining and gel electrophoresis confirmed successfùl amplification
(Fig. 2B). Slot blot analysis using allele specific oligonucleotides for the 5T, 7T, and 9T
variants revealed that, of the 24 patients, 8 were 7Tl9T heterozygotes, 14 were 7Tl7T
homozygotes, and the remaining 2 were 5Tl7T heterozygotes (Fig. 2C). As shown in
Table 3, the fiequencies of the 9T, 7T, and 5T alleles were not significantly different
between individuals in the present study and the general population [Kiesewetter et al.,
1993; Cuppens et al., 1994; Dork et al., 1994; Chillon et al., 19951 (9T: 16.7% vs.
1 1.5%, p=0.28; 7T: 79.2% vs. 83.3%, e . 4 6 ; 5T: 4.2% vs. 5.2%, p=0.75).
1II.B. CONVERSION OF EXPERIMENTAL TO ACTUAL PROPORTIONS OF EXON 9+ CFTR mRNA
To establish accurate measurement of the proportion of exon 9+ CFTR
transcnpts, it was fmt necessary to establish a standard c u v e using known concentrations
of isolated fiagments of exon 9+ and exon 9- transcripts. A plasmid containing Ml-
length CFTR cDNA served as the source of exon 9+ transcnpts pomrnens et al., 19911.
M e r two rounds of nested PCR using primers as shown in Fig. 3A, the exon 9+
fragments were extracted fiom agarose gel and purified. Exon 9- CFTR trmscnpts were
similarly obtained, except iung total RNA served as the source (Fig. 3B). Analysis by
electrophoresis and ethidium bromide staining revealed that a greater concentration of

. -
was then serially diluted and mixed with a constant quantity of the latter, and analyzed by
Southern transfer and scanning densitometry. This demonstrated that the concentration of
the isolated exon 9+ species was higher than that of exon 9- (Fig. 3D). Therefore, the
former was diluted to obtain equivalent concentrations of exon 9+ and exon 9-
transcripts. We then verified our calibration and the purity of the isolated exon 9+ and
exon 9- fragments by subjecting them to Southern analysis and hybridization with both
specific and cornmon radiolabeled oligonucleotide probes (Fig. 3E). Hybridization with
the probe comrnon to both transcripts (C16B) and subsequent densitomeûic anaiysis
confmed that we had accwately obtained equivaient concentrations of the exon 9+ and
exon 9- fiagments (Fig. 3F). Furthemore, hybridization with probes unique for exon 9+
or exon 9- rnRNA, validated the purity of the exon 9+ and exon 9- isolates, as the exon
9+ specific probe (9i-5s) and the exon 9- specific probe (ex8/10) annealed only to the
exon 9+ and exon 9- cDNA hgrnents, respectively (Fig. 3F).
Next, the exon 9+ and exon 9- fiagments were serially mixed in known
proportions and subjected to the identical PCR conditions and quantification protocol as
for the nasal and vas deferens specirnens (Fig. 3G-1). The experimental proportions of
exon 9+ and exon 9- transcripts were derived from densitometric scanning, taking the
surn of densitometric units for the exon 9+ and exon 9- bands as 100%. The
corresponding actual and experirnental proportions of exon 9+ transcripts were then
recorded graphically (Fig. 35). Consequently, al1 experhental proportions of exon 9+
transcripts from our tissue samples were adjusted according to this graph.

Total RNA was extracted fiom nasal and vas deferens specimens, converted to
cDNA, arnplified initially with primers for the transcript of the ubiquitously expressed
GAPDH gene, and analyzed by Southem hybridization. Quantification of the
autoradiographic bands by scanning densitometry showed differences in the yield of RNA
extraction and cDNA synthesis (Fig. 4). This information was then employed to estimate
the volume of cDNA to be used for PCR amplification with CFTR primers. For
example, Fig. 4 illustrates that cDNA obtained fiom the vas of patient 5 was -3.4 times
less than that of patient 1, hence 3.4 times the amount of cDNA fiom patient 5 was used
for the CFTR transcript analysis. These steps minimized inter-sample variability in RNA
extraction and cDNA synthesis, thus allowing cornparison of the proportion of exon 9+
CFTR transcripts within and between subjects.
Evaluation of CFTR mRNA transcripts fiom nasal epithelial and vas deferens
cells in the region encompassing exons 8 to 10 d e r conversion to cDNA and PCR
amplification revealed two different kgments. The difference in size between these two
bands (416 bp and 233 bp) corresponded to the size of exon 9 (183 bp) (Fig. 5A).
Southem analysis with an exon 10 specific probe (C16B) showed that both fragments
contained exon 10 sequences (Fig. 33; Fig. SC, lane 1). However, Southern analysis with
the exon 9+ specific probe (9i-5s) annealeci to the larger fiagrnent only (Fig. 5B; Fig. SC,
lane 2), while the exon 9- specific probe (ex8/10) detected only the smaller transcript
(Fig. 5B; Fig. SC, lane 3). These results confirm that the larger transcript represented the
normal, full-length transcript with exon 9 intact, and that the smaller transcript
represented the in-fi-ame exon 9 deleted transcript.

1ll.U. VUAN'I'Ik'1LA171VN OF EXVfl Y+ CFI'K mKRA
Al1 proportions of exon 9+ transcripts reported hereafter refer to the corrected,
actual proportions, based on Fig. 35. The proportion of exon 9+ nasal transcripts were
93&2%, 82t3%, and 74&1% (memSD) for the 7T/9T, 7T/7T, and 5T/7T genotypes,
respectively, while in the vas deferens sarnples, the proportion of exon 9+ transcripts
were 88&3%, 76k3%, and 64k4% for the 7T/9T, 7T/7T, and 5W7T groups, respectively
(Fig. 6 & 7; Table 4).
1XI.E. RELATIONSHIP BETWEEN LEVEL OF EXON 9+ CFTR mRNA AND THE IVSS-T TRACT
For both the nasal and vas deferens samples, significant differences were noted in
the mean exon 9+ levels for the different IVS8-T genotypes (p<0.0001, ANOVA; p<O.O5,
Bonferroni). In addition, a strong positive correlation was found between the length of
the IVS8-T tract and the proportion of exon 9+ &anscripts in nasal epithelia ( ~ 0 . 8 8 ,
p<0.001, Spearman) and in vas deferens ( ~ 0 . 8 7 , p<0.001, Spearman) (Fig. 7).
1II.F. COMPARISON OF THE PROPORTION OF EXON 9+ CFTR mRNA IN NASAL EPITHELIUM TO VAS DEFERENS
The mean levels of nasal and vasal exon 9+ transcripts were 85I7% and 79I8%
(meankSD), respectively. When evaluating the proportions of exon 9+ transcripts for
these two different CFTR expressing tissues fiom the same subject as paired samples, a
significantly higher proportion of exon 9+ transct-ipts was found within the nasd
epitheliurn compared to the vas deferens (p<0.001, paired t-test) (Fig. 8).

1V.A. SPLICING EFFICIENCY OF CFTR EXON 9 IS RELATED TO THE IVSS- T TRACT LENGTH
In the present study, we found a relationship between the WS8-T genotype and
the proportion of normal, full-length CFTR mRNA ûmscripts (exon 9+) in nasal
epithelia and vas deferens. It appears that the longer the WS8-T tract, the greater the
proportion of normal CFTR transcripts in nasal epithelia and vas deferens cells (Fig. 7;
Table 4). This fmding is consistent with that reported by Chu et al., who also
demonstrated a positive correlation between the proportion of exon 9+ transcripts in
branchial epitheliai cells and the length of the NS8-T tract [Chu et al., 19931. It is likely
that this relationship holds true for other CFTR-expressing tissues as well.
N.B. HlGHER PROPORTION OF NORMAL, CFTR mRNA IN NASAL EPITHELIUM THAN VAS DEFERlENS
Our study aisu showed that there is a greater proportion of the normal, Ml-length
CFTR message in nasal epithelia than in vas deferens fiom the same subject, regardless of
the NS8-T genotype @<0.001, paired t-test) (Fig. 8). In other words, the precise excision
of intron 8 with in-£&ne joining of exon 8 and exon 9 of CFTR *A may be less
efficient in epithelia of the reproductive tract compared to those of the respiratory tract.
Two recent reports corroborate our findings. Teng et al. found that, for the same NS8-T
tract genotype, the proportion of exon 9+ transcripts was lower in a series of vas deferens
samples obtained fiom vasectornized men than in a series of nasal biopsies obtained fiom
different men and women with chronic nasal obstruction or sinusitis [Teng et al., 19971.
Rave-Harel et al. documented that three men with CBAVD had an increased level of exon

19971. Taken together, these and our observations support the hypothesis that splicing
efficiency varies between the different tissues affected in CF.
W.C. THE CFTR WSS-T TRACT MODIFIES PHENOTYPE BY VARIABLE SPLICING OF EXON 9 IN DIFFERENT TISSUES
The discovery of differential splicing effrciency between the various tissues which
express CFTR provides important insights into the relationship between tevels of normal
CFTR and phenotypic variation. For instance, the R117H mutation is associated with
pancreatic-sufficient CF (CF-PS) mst id i s et al., 19921. CF-PS patients with this
mutation have pulmonary dysfunction but do not have pancreatic exocrine insufficiency
and their sweat chloride measurements are only modestly elevated [Harnosh et al., 19941.
Not surprisingly, the rnild R117H mutation has been identified in otherwise healthy males
with CBAVD. However, M e r genetic analysis uncovered that individuals
heterozygous for the R117H mutation on a 5T background (i.e., R117H and 5T on the
same chromosome) and a "severe" CFTR mutation (e.g., AF508, G55 1 D) developed lung
disease characteristic of CF, whereas the RI 17H mutation found in CBAVD men is
associated exclusively with the more efficient splice acceptor 7T (Le., R117H and 7T on
the same chromosome) [Kiesewetter et al., 19931. It is also important to note that the
R117H mutation gives rise to a partially functional CFTR protein [Sheppard et al., 19931.
Therefore, the R117W5T allele results in a low enough level of partially functioning
CFTR in the lung and an even lower level in the reproductive tract such that both organs
are affected. In contrast, the R117W7T allele, although producing a sufficient level of
partially functional CFTR in the lung to prevent disease, the lower level in the genital

results strongly suggest that the specific IVS8-T tract background on which a CFTR
mutation resides can modulate disease severity in a tissue-specific manner.
1V.D. PROPORTION OF NORMAL CFTR mRNA PRODUCED BY THE IVSS-T ALLELES
The frequencies of the 9T, 7T, and 5T alleles of the IVS8-T tract of the CFTR
gene in our study sample are similar to those in the general population wesewetter et ai.,
1993; Cuppens et al., 1994; Dork et al., 1994; Chillon et al., 19951 (Table 3). This
finding is not unexpected as both groups consist of normal, healthy subjects. The fact
that our study lacked subjects with the 9T/9T, 5T/9T, or 5TET genotype is likely a
consequence of the relatively small sarnple size. Despite the absence of these groups,
based on our subjects with the 7T/9T, 7Tl7T and 5T/7T genotypes (Table 4) and the
assumption that each of the two CFTR alleles contributes equally to the total arnount of
CFTR transcripts, it is inferred that the 7T ailele produces -41% exon 9+ transcripts (ie.,
-41% of the total arnount of CFTR transcripts is exon Pt) in nasal epithelium and -38%
exon 9+ transcripts in vasal epithelium. It follows, then, that the 5T allele produces
-32% and -26.5%, and the 9T ailele produces -49.5% and 4 9 % , exon 9+ CFTR mRNA
in nasal epithelial and vas deferens cells, respectively (Table 5). Therefore, although the
present study does not consist of any individuals with the 9T/9T, 5T/9T, or 5T/5T
genotypes, it c m be deduced that they would have 99% and 98%, 82% and 76%, and 64%
and 53%, exon 9+ CFTR transcripts in nasal epithelia and vas deferens, respectively
(Table 6).

phenotype and the amount of normal CFTR message (Fig. 9). A phenotypically normal,
male CF carrier typically has the 7T variant on one chromosome and a severe CFTR gene
mutation (e.g., AF508) on the other. The latter will result in absent or non-functional
CFTR protein while the 7T variant may give rise to -41% normal CFTR in respiratory
tract and -38% normal CFTR in reproductive tract, enough to sustain a normal phenotype
in these tissues. On the other hand, a typical CBAVD patient may harbor the 5T variant
on one chromosome and a severe CFTR mutation on the other. He may produce -32%
normal CFTR in the lung, which is adequate to sustain normal pulmonary f ict ion, but
-26% in the reproductive tract, an insufficient level to confer a normal genital duct
phenotype. However, the occurrence of fertile males with the severe CFTR mutation6T
genotype, such as fathers of some CF patients [Chillon et al., 19951, suggests that other
genetic factors (e.g., expression of dternative chloride channels) andfor environmental
influences can ameliorate the unfavorable eflects of certain CFTR gene sequence
alterations. Alternatively, since a range of exon 9+ mRNA level does exist for the same
ïVS8-T genotype (Fig. 7; Table 4), these heaithy, fertile men with a severe CFTR
mutation and the 5T allele may produce a level of normal CFTR fkom the 5T
chromosome that exceeds a minimal essential threshold. Furthermore, CBAVD men with
the severe CFTR mutatiod5T genotype may harbor additional mutations not detectable
by our curent DNA mutation screening methods (e.g., mutations within the promoter
region or introns of the CFTR gene).
Various studies have reported different mean values for the proportion of exon 9+
transcripts produced fiom the various IVSS-T tract genotypes. For example, with respect

ro me 1 i I I i genorype, u u er ai. reporcea a mean proporcion or exon Y+ nasal epitheliai
transcripts to be 75% [Chu et al., 19931 while Teng et al. docurnented 86% [Teng et al.,
19971. Although these estimates are not significantly different fiom ours (82%), the
slight discrepancy may be partly explained by the different methods used in the
quantitative RT-PCR analysis, such as the nurnber of cycles and nested rounds of PCR
employed, which pairs of primers were used in the PCR reaction, whether oligo-
deoxythymidine or random primers were utilized in the first strand cDNA synthesis, etc.
in addition, these studies employed a differential RT-PCR which may lead to preferential
amplification of the smaller exon 9- cDNA [Walsh et ai., 19923. Recently, Rave-Harel et
al. designed nondifferential RT-PCR reactions in which both exon 9+ and exon 9- cDNA
products were of the sarne size [Rave-Harel et al., 19971. However, this necessitated the
use of different primers and oligonucleotide probes. The former may result in different
amplification efficiencies while the latter will require deprobing and rehybridization
procedures which may lead to inadequate stripping of the previous probe andor some
loss of membrane-bound PCR products. In our approach, we minimize the number of
PCR cycles and introduce a standardization curve which should more accurately reflect
the actual proportion of exon 9+ transcripts (Fig. 3). The methodology involved is simple
and may be applied broadly to other similar quantitative RT-PCR analyses.
N.E. MOLECULAR MECNANISM OF DIFFERENTIAL SPLICING EFFICIENCY OF CFTR EXON 9
Several consensus sequences are found within introns of higher organisms that are
important for the efficient splicing of nuclear pre-mRNA. These include the s'-GU splice
donor, 3'-AG splice acceptor, branch-point A at -20 to 50 bases fiom the 3' splice site,

consensus has eleven consecutive nucleotides consisting of thymidine andfor cytosine
[Krainer and Maniatis. 19881. Extensive polypyrimidine tracts can make these sites more
cornpetitive as splice acceptor sites [Helfman and Ricci, 1989; Smith and Nadal-Ginard,
19891, while deletions in the polypyrimidine tract has been s h o w to inhibit the
S'cleavage reaction [Frendeway and Keller, 1985; Reed and Maniatis, 1986; Ruskin and
Green, 19851, binding of the splicing factors U2A.F and U2snRNP [Ruskin et al, 1 9881,
and assembly of the splicing complex [Frendeway and Keller, 19851. Furthemore, the
observation that identical pre-mRNA transcripts are processed into alternatively spliced
forms in a cell-type specific manner strongly suggests that the splicing environment of
different cells varies for these transcripts [Breibart and Nadal-Ginard, 1987; Wieczorek et
al., 19881. This tissue-specific difference in the splicing environment may be due to the
presence of specific alternative splicing factors or variations in the activities or levels of
constitutive splicing factors. Thus, the short polypyrimidine tract in CFTR intron 8,
especially with ST, likely makes it underutilized as a splice acceptor site, resulting in
reduced efficiency of exon 9 splicing. This effect is presumably more pronounced in the
reproductive tract than in the other CF-associated organs.
1V.F. StJMMAFtY
In sumrnary, o u study confums the hypothesis that splicing efficiency of CFTR
pre-mRNA varies between the different organs affected in CF. Specifically, depending
on the IVS8-T tract background, the splicing efficiency of CFTR exon 9 may be poor in
the genital tract but adequate in other tissues, thus explaining some cases of the CAVD or

- A . - . - . . . . - - - - - - - - - .
However, the existence of fertile males with the 5T genotype [Chillon et al., 19951 imply
that other genetic factors (e.g., expression of alternative chloride channels) and
environmental influences contribute to the overall phenotype. Studies such as ours may
have important future implications for CF gene therapy, which will involve delivery of
the appropriate level of normal CFTR into affected cells. Moreover, the study of the
molecular pathogenesis underlying CF-associated male infertility is especially relevant
given that the barrier to conception for men with CF, CAVD, or epididymal obstruction
has been overcome by advances made in the assisted reproductive technologies, thus
raising the concem of passing deleterious genetic traits ont0 subsequent generations [Mak
and Jarvi, 19961.

Alternative splicing variants other than exon 9- have been detected in CFTR
transcripts by RT-PCR [Bremer et al., 1992; Delaney et al., 19931. including ones which
involve the 5' end, portions of TM1, portions of NBD1, or the 3' end of the CFTR gene.
To date, these variants have not been dernonstrated to result in functional CFTR isoforms.
These patterns of alternative RNA splicing have been identified for the CFTR gene in a
variety of tissues, including lung, pancreas, intestine, and testis; however, similar
experiments have not been conducted in Wolffian-derived tissues. Since the latter
structures are highly sensitive to aberrant splicing as our study suggests, it would be
reasonable to assume that altematively spliced transcripts may occur with greater
fiequency in the reproductive tract. Therefore, we hypothesize that aberrantly spliced
transcripts other than exon 9-, previously discovered or novel, occur in the vas deferens
andor epididyrnis.
The tissue-specific difference in the splicing environment between the various
organs which express CFTR may be due to the presence of specific alternative splicing
factors. The identification and characterization of alternative splicing factors can be
approached by developing in viho splicing systems able to accurately reproduce cell-
specific splicing patterns. This should be possible either by using nuclear extracts fiom
the different ce11 types, or by complementing HeLa nuclear extracts [Smith et al., 1989;
Hodges and Bernstein, 19941. Other strategies for the isolation of factors include co-
injection of nuclei or nuclear extracts into Xenopur oocytes, dong with the experimental
transcnpt [Kelley and Perry, 1986; Kedes and Steitz, 19881. This system would also

vilru. With these approaches, alternative splicing factors which act on CFTR rnay be
identified and their mode of action subsequently characterized. The elucidation of
alternative splicing mechanisms could play an important role in development of novel
therapeutic strategies for CF. For example, it may be possible to design phmacologic
agents which upregulate the level andlor activity of constitutive splicing factors, or which
downregulate that of alternative splicing factors as means of augmenting levels of the
normal CFTR gene product.
Our findings may have a more immediate application to ow study population in
tems of providing prognostic information. Although too early in the post-operative
period for us to address this issue critically, it would be interesting to see whether a
correlation exists (after controlling for variables such as patient age, interval between
vasectomy and vasectorny reversal, and CF carrier statu) between the length of the IVS8-
T tract and the presence of sperm in the ejaculate pst-vasectomy reversal. Indeed, it
appears that azoospemiic men with idiopathic epididymal obstruction (who have a
significantly higher fiequency of the 5T variant (29%) compared to the general population
(5%) [Jarvi et al., 19951) have a lower rate of surgical success (def5ned as presence of
sperm in the ejaculate pst-vasoepididymostomy) compared to men with acquired
epididymal or vasal obstruction [Jarvi, K., unpublished data]. Although the difficulty of
surgery and the actual site of anastomosis are paramount in prognosticating post-
operative success, inadequate CFTR chloride function in the former group may contribute
to re-stenosis of the excurrent genital ducts.

Affara, N.A., Ferguson-Smith, M.A., Magenis, R.E., Tolmie, J.L., Boyd, E., Cooke, A., Jarnieson, D., Kwok, K., Mitchell, M. and Snadden, L.: Mapping testis determinants by an analysis of Y-specific sequences in males with apparent XX and XO karyotypes and females with XX karyotypes. Nucleic Acids Res., 15: 7325, 1987.
Anguiano, A., Oates, RD., Amos, LA., Dean, M., Gerrard, B., Stewart, C., Maher, T.A., White, M.B. and Milunsky, A.: Congenital bilateral absence of the vas deferens. A primarily genital f o m of cystic fibrosis. J.A.M.A., 267: 1794, 1992.
Augarten, A., Yahav, Y., Kerem, B.S., Halle, D., Laufer, J., Szeinberg, A., Dor, J., Mashiach, S., Gazit, E. and Madgar, L: Congenital bilateral absence of vas deferens in the absence of cystic fibrosis. Lancet, 344: 1473, 1994.
Austin, CR.: Observation on the penetration of sperm into the mammalian egg. Aust. J. Sci. Res., B4: 581, 195 1.
Barnes, A.D. and Toot, P.J.: Ovulation, fertilization, and implantation. In: Essentials of Obstetrics and Gynecology. Edited by N.F. Hacker and J.O. Moore. Philadelphia: W. B. Saunders Co., chapt. 3, pp.25-33, 1986.
Bedford, J.M., Calvin, H.I. and Cooper, G.W.: The maturation of spermatozoa in the human epididymis. J. Reprod. Fertil., suppl., 18: 199, 1973.
Bedford, J.M.: The bearing of epididymal fûnction in stratedes for in vitro fertilization and garnete intrafdlopian transfer. Ann. N. Y. Acad. Sci., 541: 284, 1988.
Bell, C. and McLean, J.R.: Localization of norepinephrine and acetylcholinesterase in separate neurons supplying the guinea pig vas deferens. 3. Pharmacol. Exp. Ther., 157: 69, 1967.
Berkovitz, G.D., Fechner, P.Y., Zacur, H.W., Rock, J.A., Snyder, H.M., Migeon, C.J. and Perhan, E.J.: Clinicd and pathologie spectrum of 46,XY gonadal dysgenesis: Its relevance to the understanding of sex differentiation. Medicine, 70: 375, 199 1.
Breibart, R.E. and Nadal-Ginard, B.: Developmentaily induced, muscle specific tram factors control the differential splicing of aitemative and constitutive troponin-T exons. Cell, 49: 793, 1987.
Bremer, S., Hoof, T., Wilke, M., Busche, R., Scholte, B., Riordan, J.R., Maass, G. and Turnmler, B.: Quantitative expression patterns of multidmg-resistance P-glycoprotein (MDRI) and differentially spliced cystic-fibrosis trammembrane-conductance regulator mRNA transcnpts in human epithelia. Eur. J. Biochem., 206: 137, 1992.

vas deferens. Invest. Urol., 15: 1 12. 1977.
Casals, T., Bassas, L., Ruiz-Romero, J., Chillon, M., Gimenez, J., Ramos, M., Tapia, G., Narvaez, H., Nunes, V. and Estivill, X.: Extensive analysis of 40 infertile patients with congenital absence of the vas deferens: In 50% of cases only one CFTR allele could be detected. Hum. Genet. 95: 205, 1995.
Chang, M.C.: Fertilizing capacity of spematozoa deposited in the fallopian tube. Nature, 168: 697, 195 1.
Chillon, M., Casais, T., Mercier, B., Bassas, L., Lissens, W., Silber, S., Romey, MX., Ruiz-Romero, J., Verlingue, C., Claustres, M., Nunes, V., Ferec, C. and Estivill, X.: Mutations in the cystic fibrosis gene in patients with congenital absence of the vas deferens. N. Engl. J. Med., 332: 1475, 1995.
Chu, C.-S., Trapneil, B.C., Murtagh, J.J., Moss, J., Dalemans, W., Jallat, S., Mercenier, A., Pavirani, A., Lecocq, J--P., Cutting, GR, Guggino, W.B. and Crystal, R.G.: Variable deletion of exon 9 coding sequences in cystic fibrosis transrnembrane conductance regulator gene mRNA transcripts in normal branchial epithelium. E.M.B.O.J., 10: 1 355, 1991.
Chu, CA., Trapnell, B.C., Cunistin, S.M., Cutting, G.R. and Crystal, R.G.: Extensive posttranscriptional deletion of the coding sequences for part of nucleotide-binding fold 1 in respiratory epithelial mRNA bsuiscripts of the cystic fibrosis transmembrane conductance regulator gene is not associated with the clinical manifestations of cystic fibrosis. J. Clin. Invest., 90: 785, 1992.
Chu, C.-S., Trapneli, B.C., Curristin, S.M., Cutting, G.R. and Crystal, R.G.: Genetic basis of variable exon 9 skipping in cystic fibrosis transmembrane conductance regulator rnRNA. Natwe Genet, 3: I 5 1, 1 993.
Clermont, Y.: Kinetics of spermatogenesis in mamals: Serniniferous epithelium cycle and spermatogonial renewal. Physiol. Rev., 52: 198, 1972.
Clermont, Y .: The cycle of the seminiferous epithelium in man. Amer. J. Anat., 112: 35, 1963.
Costes, B., Girodon, E., Ghanem, N., Flori, E., Jardin, A., Soufi, J.C. and Goossens, M.: Frequent occurrence of the CFTR intron 8 (TG)n 5T allele in men with congenitai bilateral absence of the vas deferens. Eur. J. Hum. Genet., 3: 285, 1995.

Demaille, J. and Claustres, M.: Analysis of the whole CFTR coding regions and splice junctions in azoospermie men with congenital bilateral aplasia of epididymis or vas deferens. Hum. Genet., 93: 467, 1994.
Cuppens, H., Teng, H., Raeymaekers, P., De Boeck, C. and Cassiman, J.J.: CFTR haplotype backgrounds on normal and mutant CFTR genes. Hum. Molec. Genet., 3: 607, 1 994.
Cystic Fiborsis Genetic Analysis Consortium: Population variations of common cystic fibrosis mutations. Hum. Mutation, 4: 167, 1994.
deKretser, D.M.: The potential of intracytoplasmic sperm injection (ICSI) to transmit genetic defects causing male infertility. Reprod. Fertil. Develop., 7: 137, 1995.
Delaney, S.J., Rich, D.P., Thomson, S.A., Hargtave, M.R., Lovelock, P.K., Welsh, M.J. and Wainwright, B.J.: Cystic fibrosis transmembrane conductance regulator splice variants are not consemed and fail to produce chlonde channels. Nat. Genet., 4: 426, 1993.
Denning, C.R., Sommers, S.C. and Quigley Jr., H.J.: Infertility in male patients with cystic fibrosis. Pediatrics, 41: 7, 1968.
Denning, G.M., Anderson, M.P., Amara, J.F., Marshall, J., Smith, AE. and Welsh, M.J.: Processing of mutant cystic fibrosis transmembrane conductance regulator is temperature- sensitive. Nature, 358: 76 1, 1992.
Disteche, C.M., Casanova, M., Saal, H., Friedman, C., Sybert, V., Graham, J., Thuline, H., Page, D.C. and Fellous, M.: Small deletions of the short arm of the Y chromosome in 46,XY fernales. Proc. Natl. Acad. Sci. USA, 83: 784 1, 1 986.
DiZerga, G.S. and Sherins, R.J.: Endocrine control of adult testicular function. In: The Testis. Edited by H. Burger and D. deKretser. New York: Raven Press, pp. 127, 198 1.
Donat, R., McNeill, A.S., Fitzpatrick, D.R. and Hargreave, T.B.: The incidence of cystic fibrosis gene mutations in patients with congenital bilateral absence of the vas deferens in Scotland. Brit. J. Urol., 79: 74, 1997.
Dork, T., Fislage, R., Neumann, T., Wulf, B. and Turnmler, B.: Exon 9 of the CFTR gene: splice site haplotypes and cystic fibrosis mutations. Hum. ene et., 93: 67, 1994.
Dnimm, M.L., Wilkinson, D.J., Smit L.S., Worrell R.T., Strong, T.V., Frizzell, R.A., Dawson, D.C. and Collins, F.S: Chloride conductance expressed by delta F508 and other mutant CFTRs in Xenopus oocytes. Science, 254: 1797, 199 1.

- infertility. Fertil. Steril., 22: 469, 197 1.
Dumur, V., Gervais, R., Rigot, J.-M., Lafitte, LJ . , Manouvrier, S., Biserte, J., Mazeman, E. and Roussel. P.: Abnormal distribution of CF AF508 allele in azoospermie men with congenital aplasia of epididymis and vas deferens. Lancet, 336: 5 12, 1990.
Dwieu, I., Bey-Omar, F., Rollet, J., Calemard, L., Boggio, D., Lejeune, H., Gilly, R., Morel, Y. and Durand, D.V.: Diagnostic criteria for cystic fibrosis in men with congenital absence of the vas deferens. Medicine, 74: 42, 1995.
Fishel, S., Green, S., Bishop, M., Thornton, S., Hunter, A., Fleming, S. and al-Hassan, S.: Pregnancy afier intracytoplasmic injection of spermatid. Lancet, 345: 164 1, 1995.
Fishel, S.B. and Thornton, S.: The use of assisted reproduction technology (ART) for achieving conception in male infertility. Brit. J. Urol., suppl. 1,75: 50, 1995.
Frendeway, D. and Keller, W.: The stepwise assembly of a pre-mRNA splicing complex requires U-snRNPs and specific intron sequences. Cell, 42: 355, 1985.
Gil-Salom, M., Minguez, Y., Rubio, C., Remohi, J. and Pellicer, A.: Intracytoplasmic testicular sperm injection: an effective treatment for otherwise intractable obstructive azoosperrnia. J. Urol., 154: 2074, 1995.
Greenberg, S.H., Lipshultz, L.I. and Wein, A.J.: Experience with 425 subfertile male patients. J. Urol., 119: 507, 1978.
Hamosh, A., Corey, M. and the Cy stic fibrosis genotypdphenotype consortium: Correlation between genotype and phenotype in patients with cystic fibrosis. N. Engl. J. Med., 329: 1308, 1994.
Hansson, V. and Djoseland, O.: Prelirninary characterization of the 5-alpha dihydrotestosterone binding protein in the epididymal cytosol fraction: In vivo studies. Acta. Endocrinol., 71: 614, 1972.
Harari, O., Borne, H., McDonald, M., Richings, N., Speirs, A.L., Johnston, W.I. and Baker, H.W.: htracytoplasmic sperm injection: a major advance in the management of severe male subfertility. Fertil. Steril., 64: 360, 1995.
Helfnian, D.M. and Ricci, W.M.: Branch point selection in alternative splicing of tropomyosin pre-mRNAs. Nucleic Acids Res., 17: 5633, 1989.
Heller, C.G. and Clermont, Y.: Kinetics of the germinal epithelium in man. Recent Prog. Hom. Res., 20: 545, 1964.

maturation in the human epfdidyrnis. J. Reprod. ~erti1.i 60: 291, 1980.
Hodges, D. and Bernstein, S.I.: Genetic and biochemical analysis of alternative RNA splicing. Adv. Genet., 31: 207, 1994.
Holsclaw, D.S., Lobel, B., Jockin, H. and Schwachman, H.: Genital abnonnalities in male patients with cystic fibrosis. J. Urol., 106: 568, 1971.
Howards, S.S.: Treatment of male infertility. N. Engl. J. Med., 332: 3 12, 1995.
In't Veld, P., Brandenburg, H., Verhoeff, A., Dhont, M. and Los, F.: Sex chrornosomal abnormalities and intracytoplasmic sperm injection. Lancet, 346: 773, 1995.
Jarvi, K., Zielenski, J., Wilschanski, M., Durie, P., Suckspan, M., Tullis, E., Markiewicz, D. and Tsui, L.-C.: Cystic fibrosis trammembrane conductance regulator and obstructive azoosperrnia. Lancet, 345: 1578, 1995.
Jenkins, A.D., Turner, T.T. and Howards, S.S.: Physiology of the male reproductive system. Urol. Clin. North Amer., 5: 437, 1978.
Jequier, A.M., Ansell, I.D. and Bullimore, N.J.: Congenital absence of the vasa deferentia presenting with infertility . J. Androl., 6: 1 5, 1 985.
Johnson, A.L. and Howards, S.S.: Intratubular hydrostatic pressure in the testis and epididyrnis before and after long-term vasectomy in the guinea pig. Biol. Reprod., 14: 371,1976.
Johnson, L. and Vamer, D.D.: Effect of daily spermatozoan production but not age on transit time of spermatozoa through the human epididyrnis. Biol. Reprod., 39: 8 12, 1988.
Josso, N.: Antirnullerian hormone: New perspectives for a sexist molecule. Endocrinol. Rev., 7: 421, 1986.
Kaplan, E., Shwachrnan, H., Perlmutter, A.D., Rule, A., Khaw, K.T. and Holsclaw, D.S.: Reproductive failure in males with cystic fibrosis. N. Engl. J. Med., 279: 65, 1968.
Kedes, D.H. and Steitz, J.A.: Correct in vivo'splicing of the moue irnmunoglobulin K
light-chah pre-niRNA is dependent on the 5' splice-site position even in the absence of transcription. Genes Dev., 2: 1448, 1988.
Kelley, D.E. and Perry, R.P.: Transcriptional and posttranscriptional control of irnmunoglobulin mRNA production during B lymphocyte development. Nucleic Acids Res., 14: 543 1,1986.

Buchwald, M. and Tsui, L.-C.: Identification of the cystic fibrosis gene: genetic analysis. Science, 245: 1073, 1989.
Kerr, J.B. and deKretser, D.M.: The cytology of the human testis. In: The Testis. Edited by H. Burger and D. deketser. New York: Raven Press, pp. 141, 1981.
Kiesewetter, S., Macek, M Jr., Davis, C., Curristin, S. M., Chu, CS., Graham, C., Shrimpton, A.E., Cashman, S.M., Tsui LX., Mickle, J., Amos, J., Highsmith, W.E., Shuber, A., Witt, D.R., Crystal, R.G. and Cutting, G.R.: A mutation in CFTR produces different phenotypes depending on chromosomal background. Nat. Genet., 5: 274, 1993.
Krainer, A.R. and Maniatis, T.: RNA splicing. In: Transcription and Splicing. Edited by B.D. Harnes and D.M. Glover. Oxford: IRL Press, pp. 13 1-206,1988.
Kristidis, P., Bozon, D., Corey, M., Markiewicz, D., Rommens, J., Tsui, L.-C. and Durie, P.: Genetic determination of exocrine pancreatic function in cystic fibrosis. Amer. J. Hum. Genet., 50: 1 178, 1992.
Landing, B.H., Wells, T.R. and Wang, C.-1.: Abnomality of the epididymis and vas deferens in cystic fibrosis. Arch. Path., 88: 569, 1969.
Lilja, H., Oldbring, J., Rannevik, G. and Laurell, C.B.: Seminal vesicle-secreted proteins and their reactions during gelation and liquefaction of human semen. J. Clin. invest., 80: 281,1987.
Lipsett, M.B.: Steroid secretion by the testis in man. In: The Endocrine Function of the Human Testis. New York: Academic Press, vol. 2, 1974.
Lipshultz, L.I., Howards, S.S. and Buch, J.P.: Male Idertility. In: Adult and Pediatric Urology. Edited by J.Y. Gilienwater, J.T. Grayhack, S.S. Howards and J.W. Duckett. St. Louis: Mosby, vol. 2, chapt. 36, pp. 1425-1477, 19%.
Lipshultz, L.I.: Current Urologic Therapy. Philadelphia: W.B. Saunders Co., 1980.
Lodish, H., Baltimore, D., Berk, A., Zipursky, S.L., Matsudaira, P. and Darnell, J.: Transcription termination, RNA processing, and posttranscriptional control. In: Molecular Ce11 Biology. Edited by H. Lodish, D.Baltimore, A. Berk, Zipurslcy, S.L., P. Matsudaira and J. Darnell. New York: W.H. Freeman and Company, pp. 485-542, 1995.
Mak, V. and Jarvi, K.A.: The genetics of male infertility. J. Urol., 156: 1245, 1996.
Mann, T. and Mann C.L.: Reproductive Function and Semen. New York: Springer- Verlag, 198 1.

McKusick, V.A.: Mendelian Inheritance in Man. Catalogs of Autosomal Dominant, Autosomal recessive, and X-Linked Phenotypes. Baltimore: John Hopkins University Press, vol. 2, p. 1746, 1992.
Means, A.R., Dedman, J.R., Tash, H.S., Tindall, D.J., Van Sickel, M. and Welsh, M.J.: Regulation of the testis Sertoli ce11 by follicle stimulating hormone. Am. Rev. Physiol., 42: 59, 1980.
Mercier, B., Verlingue, C., Lissens, W., SiIber, S.J., Novelli, G., Bondueile, M., Audrezet, M.P. and Ferec, C.: 1s congenital bilateral absence of vas deferens a primary fonn of cystic fibrosis? Analyses of the CFTR gene in 67 patients. Amer. J. Hum. Genet., 56: 272, 1995.
Mickle, J.E., Oates, R.D., Colin, A., Maher, T.A., Milunsky, A. and Amos, J.A.: Increased fiequency of cystic fibrosis (CF) mutations in males with unilateral absence of the vas deferens (UAVD). Amer. J. Hum. Genet., suppl., 53: abstract 1204, 1993.
Miller S.A., Dykes D.D. and Polesky, H.F.: A simple salting out procedure for extracting DNA fiom human nucleated cells. Nucleic Acids Res., 16: 1215, 1988.
Moore K.L.: The fmt week of human development. In: Before We Are Born. Basic Embryology and Birth Defects, 3rd ed. Philadelphia: W. B. Saunders Co., chapt. 3, pp. 25-32,1989a.
Moore K.L.: The urogenitaI system. In: Before We Are Born. Basic Embryology and Birth Defects, 3rd ed. Philadeiphia: W. B. Saunders Co., chapt. 14, pp. 180-201, 1989b.
Moore, H.D., Hartman, T.D. and Pryor, J.P.: Development of the oocyte-penetrating capacity of spermatozoa in the human epididymis. Int. J. Androl., 6: 3 10,1983.
Morris, R.S. and Gleicher, N.: Genetic abnormalities, male infertility, and ICSI. Lancet, 347: 1277,1996.
Mosher, W.E.: Reproductive Ünpainnents in the United States 1 965- 1982. Demography, 22: 415, 1985.
Mullis, K., Faloona, F., Scharf, S., Saiki, R., Hom, G. and Erlich, H.: Specific enzymatic amplication of DNA in vitro: the polymerase chah reaction. Cold Spnng Harbor Symp. Quant. Biol., 51: 263, 1986.
Oates, R.D. and Amos, J.A.: The genetic basis of congenital bilateral absence of the vas deferens and cystic fibrosis. J. Androl., 15: 1, 1994.

Olson, J.R. and Weaver, D.K.: Congenital mesonephric defects in male infants with mucoviscidosis. J. Clin. Path., 22: 725, 1969.
Orgebin-Crist, M.C.: Studies on the fünction of the epididymis. Biol. Reprod., suppl., 1: 155, 1969.
Osborne, L.R., Lynch, M., Middleton, P.G., Alton, E.W.F.W., Geddes, D.M., Pryor, J.P., Hodson, M.E. and Santis, G.K.: Nasal epithelial ion transport and genetic analysis of infertile men with congenital bilateral absence of the vas deferens. Hum. Molec. Genet., 2: 1605, 1993.
Owan, C. and Sjoberg, N.O.: The importance of short adrenergic neurons in the seminal emission mechanism of rat, guinea pig and man. J. Reprod. Fertil., 28: 379, 1972.
Page, D.C., Mosher, R., Simpson, E.M., Fisher, E.M.C., Mardon, G., Pollack, J., McGillivray, B., de la Chapelle, A. and Brown, L.G.: The sex-determining region of the hurnan Y chromosome encodes a finger protein. Cell, 51 : 109 1, 1987.
Palermo, G., Joris, H., Dewoey, P. and Van Steirteghem, AC.: Pregnancies after intracytoplasmic injection of single sperrnatozoon into an oocyte. Lancet, 340: 17, 1992.
Palmer, M.S., Sinclair, A.H., Berta, A.P., Ellis, N.A., Goodfellow, P.N., Abbas, N.E. and Fellous, M.: Genetic evidence that ZFY is not the testis-detennining factor. Nature, 342: 937, 1989.
Patrizio, P., Asch, R.H., Handelin, B. and Silber, S.J.: Aetiology of congenital absence of vas deferens: genetic study of three generations. Hum. Reprod., 8: 21 5, 1993.
Peters, H.: Migration of gonocytes into the marnmalian gonad and their differentiation. Philos. Trans. R. Soc. Lond. (Biol.), 259: 91, 1970.
Pollard, C.E., Harris, A., Coleman, L. and Argent, B.E.: Chloride channels on epithelial cells culhired fiom human fetal epididymis. J. Membr. Biol., 124: 275, 199 1.
Rave-Harel, N., Kerem, E., Nissim-Rafinia, M., Madjar, I., Goshen, R., Augarten, A., Rahat, A., Hurwitz, A., Darvasi, A. and Kerem, B.: The molecular basis of partial penetrance of splicing mutations in cystic fibrosis. Amer. J. Hum. Genet., 60: 87, 1997.
Reed, R. and Maniatis, T.: A role for exon sequences and splice-site proximity in splice- site selection. Cell, 46: 68 1, 1986.

L U V I U L U I , .#.Ab., A\UA..AAAW..", W.. 7 . 0 , ..W.- .a., Y . , . ..Y... . .., a. VY..iUI.W., a.., V.Y...V*., -.* Zielenski, J., Lok, S., Plavsic, N., Chou, J.-L., D m m , M.L., Iannuzzi, M.C., Collins, F.S. and Tsui, L.-C.: Identification of the cystic fibrosis gene: cloning and characterization of complernentary DNA. Science, 245: 1066, 1989.
Ritzen, E.M.. Hansson, V. and French, F.S.: The Sertoli cell. In: The Testis. Edited by H. Burger and D. deketser. New York: Raven Press, pp. 1 7 1, 198 1.
Ritzen, E.M., Nayfeh, S.N., French, F.S. and Dobbins, M.C.: Dernonstration of androgen binding components in rat epididymis cytosol and cornparison with binding components in prostate and other tissues. Endocrinol., 89: 142, 197 1.
Romrnens, J.M., Dho, S., Bear, C.E., Kartner, N., Kennedy, D., Riordin, J.R., Tsui, L X . and Foskett, J.K.: CAMP-inducible chloride conductance in mouse fibroblast lines stably expressing the human cystic fibrosis transmembrane conductance regulator. Proc. Natl. Acad. Sci. USA, 88: 7500,199 1.
Rommens, J.M., Iannuzzi, M.C., Kerem, B., Drumm, M.L., Melmer, G., Dean, M., Romahel, R., Cole, J.L., Kennedy, D., Hidaka, N., Zsiga, M., Buchwald, M., Riordan, J.R., Tsui, L X . and Collins, F.S.: Identification of the cystic fibrosis gene: chromosome walking and jumping. Science, 245: 1059, 1989.
Ruskin, B. and Green, M.R.: Role of the 3' splice site consensus sequence in marnmalian pre-mRNA splicing. Nature, 317: 732, 1985.
Ruskin, B., Zamore, P.D. and Green, M.R.: A factor, U2AF, is required for U2 snRNP binding and splicing complex assembly. Cell, 52: 207, 1988.
Russell, L.D.: Sertoli-genn ce11 interactions: A review. Gamete Res., 3: 179, 1980.
Schally, A.V., Mair, R.M.G., Arimura, A. and Redding, T.W.: Isolation of the luteinizing hormone and follicle-stimulating hormone-releasing hormone from porcine hypothalami. J. Biol. Chem., 246: 7230, 1971.
Schlegel, P.N., Palermo, G.D., Alikani, M., Adler, A., Reing, A.M., Cohen, J. and Rosenwaks, 2.: Micropuncture retrieval of epididymal sperm with in vitro fertilization: importance of in vitro micromanipulation techniques. Urology, 46: 238, 1995.
Schlegel, P.N., Shin, D. and Goldstein, M.: Urogenital anomalies in men with congenital absence of the vas deferens. J. Urol., 155: 1644, 1996.
Sheppard, D.N., Rich, D.P., Ostedgaard, L.S., Gregory, R.J., Smith, A.E. and Welsh, M.J.: Mutations in CFTR associated with mild-disease form Cl' channels with altered pore properties. Nature, 362: 160, 1993.

Coularn, C.B. and Schulman, J.D.: lntracytoplasm~ sperm injection facilitates fertilization even in the most severe forms of male infertility: pregnancy outcome correlates with materna1 age and number of eggs available. Fertil. Stenl., 64: 369, 1995.
Sigman, M. and Howards, S.S.: Male infertility. in: Campbell's Urology, 6th ed. Edited by P.C. Walsh, A.B. Retik, T.A. Stamey and E.D. Vaughan, Jr. Philadelphia: W.B. Saunders Co., vol. 2, chapt. 15, pp. 661-705, 1992.
Siiteri, P.K. and Wilson, J.D.: Testosterone formation and metabolisrn during male sexual differentiation in the hurnan embryo. J. Clin. Endocrinol. Metab., 38: 11 3, 1974.
Silber, S.J., Balmaceda, J., Borrero, C., Ord, T. and Asch, R.: Pregnancy with sperm aspiration fiom the proximal head of the epididymis: a new treatment for congenital absence of the vas deferens. Fertil. Steril., 50: 525, 1988.
Silber, S.J., Van Steirteghem, A.C., Liu, J., Nagy, Z., Tournaye, H. and Devroey, P.: High fertilization and pregnancy rate after intracytoplasmic sperm injection with spermatozoa obtained fiom testicle biopsy. Hum. Reprod., 10: 1 48, 1 995.
Sinclair, A.H., Berta, P., Palmer, MaS., Hawkins, J.R., Griffiths, B.L., Smith, M.J., Foster, J.W., Frischauf, A.-M., Levell-Badge, R. and Goodfellow, P.N.: A gene fiom the human sex-determining region encodes a protein with homology to a conserved DNA- binding motif. Nature, 346: 240, 1990.
Smith, C.W.J. and NadaMinard, B.: Mutually exclusive splicing of alpha-tropomyosin exons enforced by an unusual lariat branch point location: implications for constitutive splicing. Cell, 56: 749, 1989.
Smith, C.W.J., Patton, J.G. and Nadal-Ginard, B.: Alternative splicing in the control of gene expression. Annu. Rev. Genet., 23: 527, 1989.
Southem, E.M.: Detection of specific sequences among DNA hgments separated by gel electrophoresis. J. Mol. Biol., 98: 503, 1975.
Strong, T.V., Koh, J., Tsui, L.-C. and Collins, F.S.: Analysis of CFTR transcription. Pediat. Pulrnonol., suppl., 5: abstract 195, 1991.
Strong, T.V., Wilkinson, d.J., Mansoura, M.K., Devor, D.C., Genze, K., Yang, Y., Wilson, J.M., Cohn, J.A., Dawaon, D.C., Frizzell, R.A. and CoIlins, F.S.: Expression of an abundant alternatively spliced form of the cystic fibrosis transmernbrane conductance regulator (CFTR) gene is not asociated with a CAMP-activated chloride conductance. Hum. Mol. Genet., 2: 225,1993.

~er t i l i6 in males with cystic fibrosis. N T E ~ ~ ~ . J. Med., 287: 586, 1972.
Temple-Smith, P.D., Southwick, G.J., Yates, C.A., Trounson, A.O. and de Dretser, D.M.: Hurnan pregnancy by in vitro fertilization (IVF) using sperm aspirated fiom the epididymis, J. In Vitro Fertil. Embryo Transfer, 2: 1 19, 1985.
Teng, H., Jorissen, M.,Van Poppel, H., Legius, E., Cassiman, J.-J. and Cuppens, H.: Increased proportion of exon 9 altematively spliced CFTR transcnpts in vas deferens compared with nasal epithelial cells. Hum. Mol. Genet., 6: 85, 1997.
Tesarik, J., Mendoza, C. and Testart, J.: Viable embryos fiom injection of round spermatids into oocytes. N. Engl. J. Med., 333: 525, 1995.
Tizzano, E.F., Chitayat, D. and Buchwald, M.: Cell-specific localization of CFTR mRNA shows developmentally regulated expression in hurnan fetal tissues. Hum. Molec. Genet., 2: 21 9,1993.
Tizzano, E.F., Silver, M.M., Chitayat, D., Benichou, J.-C. and Buchwald, M.: Differential cellular expression of cystic fibrosis transmembrane regulator in hurnan reproductive tissues. Clues for the infertility in patients with cystic fibrosis. Amer. J. Path., 144: 906, 1994.
Tokunaga, K., Nakmura, Y., Sakata, K., Fujimori, K., Ohkubo, M., Sawada, K. and Sakiyama, S.: Enhanced expression of a glyceraldehyde-3-phosphate dehydrogenase gene in human lung cancers. Cancer Res., 47: 5616, 1987.
Trezise, A.E.O., Chambers, J.A., Wardle, C.J., Gould, S. and Harris, A.: Expression of the cystic fibrosis gene in human foetal tissues. Hum. Molec. Genet., 2: 21 3, 1993.
Valman, H.B. and France, N.E.: The vas deferens in cystic fibrosis. Lancet, 2: 566, 1969.
Van Steirteghem, A.C., Nagy, Z., Joris, H., Liu, J., Staessen, C., Smitz, J., Wisanto, A. and Devroey, P.: High fertilization and implantation rates after intracytoplasmic sperm injection. Hum. Reprod., 8: 1061, 1993.
Van Thiel, D.H., Sherins, R.J., Myers, G.H., Jr. and DeVita, V.T., Jr.: Evidence for a specific seminiferous tubular factor affecting follicle-stimulating hormone secretion in man. J. Clin. Invest., 51: 1009, 1972.
Vanderzwalrnen, P., Lejeune, B., Nijs, M., Segal-Bertin, G., Vandamme, B. and Schoysman, R.: Fertilization of an oocyte microinseminated with a spermatid in an in- vitro fertilization programme. Hum. Reprod., 10: 502, 1995.

1
the alpha and beta subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. E.M.B.O.J., 1: 945, 1982.
Walsh, P.C., Swerdloff, R.S. and Odell, W.D.: Effect of bilateral orchiectomy on s e m luteinizing hormone (LH) and follicle-stimulating hormone (FSH) concentrations in three men. Acta. Endocrinol., 74: 449, 1973. Walsh, P.S., Erlich, H.A. and Higuchi, R.: Preferential PCR amplification of alleles: mechanisms and solutions. PCR Methods Appl., 1: 241, 1992.
Welsh, M.J., Tsui, L.-C., Boat, T.F. and Beaudet, A.L.: Cystic fibrosis. In: The Metabolic and Molecular Bases of inherited Disease. Edited by C.R. Scriver, A.L. Beaudet, W.S. Sly and D. Valle. New York: McGrwHill, vol. 3, chapt. 127, pp. 3799- 3876,1995.
Wheater, P.R., Burkitt, H.G. and Daniels, V.G.: Male reproductive system. In: Functional Histology. New York: Churchill Livingstone, chapt. 18, pp.277-288, 1987.
Wieczorek, D., Smith, C.W.J. and Nadal-Ginard, B.: The rat a-tropomyosin gene generates a minimun of six different rnRNAs coding for striated, smooth and non-muscle isoforms by alternative splicing. Mol. Ce11 Biol., 8: 679, 1988.
Witschi, E.: Migration of the germ cells of human embryos fiom the yolk sac to the primitive gondal folds. Carnegie Contributions to Embryology, 32: 69, 1948.
World Health Organization: WHO Laboratory Manual for the Examination of Human Semen and Semen-Cervical Mucus Interaction. Cambridge, U.K.: Cambridge University Press, p. 30, 1987.
Zielenski, J., Markiewicz, D., Bombieri, C., Pignatti, P.F. and Tsui, L.-C.: Haplotype analysis of chromosomes carrying the R75Q CFTR variant. Pediatr. Pulmonol., suppl., 51: abstract 88, 1995a.
Zielenski, J., Patrizio, P., Corey, M., Handelin, B., Markiewicz, D., Asch, R. and Tsui, L.-C.: CFTR gene variant for patients with congenital absence of vas deferens. Amer. J. Hum. Genet., 57: 958, 1995b.
Zielenski, J., Romahel, R., Bozon, D., Kerem, B., Grzelczak, Z., Riordan, J.R., Rornmens, J. and Tsui, LX.: Genomic DNA sequence of the cystic fibrosis trammembrane regulator (CFTR) gene. Genornics, 10: 2 14,199 1.
Zielenski, J. and Tsui, L.-C.: Cystic fibrosis: genotypic and phenotypic variations. Annu. Rev. Genet., 29: 777, 1995.

Organization, 19871
Semen volume:
Spenn count:
Percentage motility:
Percentage nonnal morphology:
White blood ce11 count:

'L'able 2. CFI'K gene mutations or vanations ana cHAvu
CFTR genotype 1 No. of CBAVD men (%y 1 Percentage of CBAVD mena
CFTR mutation / 1 43 1 CFTR mutation (19.8) CFTR mutation / -b 34
(1 5.7) CFTR mutation / 5T 85
5T / 5T -46% involve 5T variant
5T/-
- . .
-75% involve
CFTR gene
mutations
-1 8% have no CFTR gene mutations identified
aCalculated fiom a total of 217 CBAVD men [Chillon et al., 1995; Costes et al., 1995; Zielenski et
al., 1 995 b] . "'-" indicates no CFTR mutations detected.

general population
Group IVSS-T aflele frequency
9T 7T 5T
no. with allele / no. studied (%)
Present study 8/48 3 8/48 2/48 (48 chromosomes) (1 6.71~ (79.2)' (4.2)d
General population 571498 4 15/498 26/498 (498 chromosomes)a (1 1.5) (83.3) (5.2)
Data pooled from Kiesewetter et al., 1993, Cuppens et al., 1994, Dork et al., 1994, and
Chillon et al., 1995.
b x 2, 1 . 1 , p=0.28 for cornparison with the 9T allele fiequency in the generai population.
c x 2- -0.5, p=0.46 for cornparison with the 7T allele fiequency in the general population.
dp=0.75, Fisher's exact test, for comparison with the ST allele fiequency in the general
population.

Subject IVS8-T Nasal exon Vas Meanf SD MeankSD number genotype 9+ (Yo) deferens nasal vas deferens
exon 9+ (%) exon 9+ (%) exon 9+ (%)
aValues represent corrected proportions of exon 9+ transcripts, according to Fig. 35.

T tract allelesa
IVSS-T allele Proportion of nasal exon Pt Proportion of vasal exon 9+ ("/.) (%)
Values represent corrected proportions of exon 9+ transcripts, according to Fig. 35.

T tract genotypesa
IVS8-T genotype Proportion of nasal exon 9+ Proportion of vasal exon 9+ (%) (%)
'values represent corrected proportions of exon 9+ transcripts, according to Fig. 3J.

a' .SUI b a. 1 u)i S U - a VU u y u m j pj m ~ m m m m u m m m t - 8 ur- v a s u - U A r A\ 6 Y 1 4 - aUU - 4 - b . U 4 6 . V )i
splicing of CFTR exon 9. Three alleles have been identified in the polypy-rimidine tract
within the splice acceptor site of intron 8 of the CFTR gene: one with 9 thymidines, one
with 7 thymidines, and one with 5 thymidines (9T. 7T, 5T) [Chu et al., 19911. Al1 three
alleies are associated with two transcripts, one that is normally spliced with exon 9 intact
(exon 9+) and one that is aberrantly spliced, with skipping of exon 9 but i n - M e joining
of exons 8 to 10 (exon 99) [Chu et al., 1993 3. Transfection studies demonstrated that the
protein made from the exon 9- transcript is incapable of functioning as a CAMP-regulated
chloride channel [Strong et al., 1993; Delaney et ai., 19931. In addition, quantitative RT-
PCR analysis performed on branchial epithelial cells showed that individuals with the
longer T-tract length had a higher proportion of exon 9+ transcripts [Chu et al., 19931.
Therefore, CBAVD men with the 5T variant likely produce lower levels of normal CFTR
mRNA due to a less efficient splicing machinery.

V p I I L W 9 W U W r b V I a8bW GTGTG 7 A A A C A G 7T Of CFTR intron * GTGTG77777AAACAG 5T
CFTR 5'- 8 j gene I 3' -
/ Transcription
5 r l 3 E D 118 1 9 1 10 11 Exon 9- CFTR mRNA Exon 9+ CFTR mRNA
1 Translation 4 Inactive CFTR protein Functional CFTR protein
5Tl5T 5T17T 5T19T TTlTT R19T 9T19T IVS8-T genotype
Figure 1.

Figure 2. Detection of WS8-T genotype by oligonucleotide bybridization.
A) Leukocy-te genomic DNA fiom study subjects were extracted and the region
encompassing the intron 8 polypyrimidine tract was amplified by PCR with
primers indicated in the schematic to generate a predicted product of 560 bp.
B) Amplification was evaluated by ethidium bromide staining and gel
electrophoresis. PCR product was generated fiom al1 subjects (numbered 1 to
24) (M=100 bp marker; Neg==no DNA template). C) An aliquot of amplified
DNA was denatured and vacuum-blotted to nylon membrane and hybridized with
radiolabeled oligonucleotides which specifically recognize ST, 7T, or 9T aleles.
Autoradiography revealed that subjects 1, 9, and 23 possess the 7T/9T, 7T/7T,
and 5T/7T genotypes, respectively.

poly-T tract
CFTR 5' 1 8 1 gene 3'
Figure 2A.
Figure 2B.

Subject number
1 9 23
probe TI 9 5G-GTGTGTTTllTAACAG-31 Probe sequence
TTIST 7TffT ST/7T
NS8-T genotype
Figure 2C.

type (WT) CFTR cDNA (exon 9+) were subjected to two rounds of nested PCR with
primers as shown. Amplified products were assessed by ethidium bromide staining and
agarose gel electrophoresis (Neg controkno DNA). Second round PCR fragments (834
bp) were extracted fiom the gel and purified. B) Fragments of CFTR exon 9- transcripts
(651 bp) were obtained in a similar marner from human lung total RNA. C) Sarne
volumes of the isolated exon 9+ and exon 9- fragments were loaded ont0 separate lanes
and size fractionated by agarose gel electrophoresis for cornparison. Note that a higher
yield was obtained for the exon 9+ hgments, as reflected by their greater intensity under
ultraviolet light. D) To obtain equivalent concentrations of exon 9+ and exon 9-
transcripts, the exon 9+ fragments were serially diluted and mixed with a constant volume
of the exon 9- frsigments, and evaluated by Southem analysis. Hybridization with an
exon 10 specific probe (C16B) produced bands of the two kgments, and quantification
by optical density (OD) measurements demonstrateci that the concentration of the isolated
exon 9+ fragments was -1.82 times that of exon 9- fragments. Thus, the exon 9+
fi-agments were diluted 1.82 tirnes, making them equivalent in concentration to the exon
9- fragments. E) In order to ven@ calculations and the purity of the exon 9+ and exon 9-
isolates, Southem analysis and hybridization with a common probe (C16B), exon 9+
specific probe (9i-Ss), and exon 9- specific probe (ex8/10) were used. F) Hybridization
with C 16B and OD measurements confinned that concentrations of exon 9+ (OD=2.482)
and exon 9- (OD=1.478) fragments were equivalent. Hybridization with 9i-5s and ex8/10
validated the purity of the isolates, as s h o w by their annealing to only the exon 9+ and
exon 9- fkgrnents, respectively. G-1) The exon 9+ and exon 9- fragments were serially

subjected to the same PCR conditions and quantification procedure as for the nasal and
vas deferens specimens. The experimental proportions of exon 9+ and exon 9- transcripts
were evaluated by scanning densitometry, with the sum of OD measurements for exon 9+
and exon 9- taken as 100%. J) Graph of actual versus experimental proportions of CFTR
exon 9+ transcripts. Al1 experimental proportions of nasal and vas deferens exon 9+
mRNA were adjusted according to this graph.


Figure 3C.
Vol (pl) 10.0 10.0 10.0 10.0 10.0 10.0 10.0 10.0 10.0 10.0
Figure 30.

Figure 3E.
Exon 9- fragments
c c e c c c r e r O O s d a w 8 Y 8
w l u w
7 1 8 1 1 0 ( 11 1 1 2 651bp
Exon 9+ OD 1.482
Exon 9- OD 1.478
Figure 3F.

Exon 9- fragments
Figure 36.
Exon 9+ fragments
Exon 9- fragments
Figure 3H.

Figure 31.
Figure 35.

Figure 4. Evaluation of RNA extraction and cDNA synthesis. Total RNA
was extracted fiom nasal and vas deferens specimens, converted to cDNA, and
PCR amplified for a region of the GAPDH transcript. The products were
analyzed by gel electrophoresis, Southern blotted, and hybridized with a 32P-
labeled GAPDH probe specific for the amplified region. The resuitant
autoradiographic bands were quantified by densitometric scanning. Differences
in the yield of RNA extraction and cDNA synthesis were adjusted for, prior to
amplification with CFTR primers. For example, the concentration of cDNA
obtained from the vas of subject 5 was -3.4 times l e s than that from subject 1,
hence 3.4 times the amount of cDNA from subject 5 was used for CFTR mRNA
analy sis.

Vas sample no.: I 2 3 4 5 6 7 8
OD:
Figure 4.

Figure 5. Evaluation of CFTR mRNA. A) Total RNA was extracted fiom
nasal and vas deferens samples, converted to cDNA, and PCR amplified using
primers fiom exons 8 and 10 (X8-5 and C16D, respectively), producing two
different sized frzigments which corresponded to the exon 9+ (416 bp) and exon
9- (233 bp) transcripts. B) The CFTR cDNA products were then separated by
agarose gel electrophoresis and transferred to nylon memebrane by Southem
blotting, followed by hybridization with 32P-labeled probes specific for the exon
9+ or exon 9- fragments (9i-5s and ex8110, respectively) and one common to
both (C 16B). C) C 16B, a sequence within exon 10, identified two species (lane
1). The exon 9+ specific probe (9i-5s) detected only the larger transcript,
corresponding to CFTR mRNA with exon 9 intact (Xane 2), while the exon 9-
specific probe (ex8/10) detected only the smaller transcript (lane 3) which
represented the i n - h e deleted exon 9 message.

Exon Q + ~ R N A 1 8 1 9 1 10 1 * -
X l b C16D - 416 bp
Exon s - mRNA lIia3rl - -
Xô-S Cl60
Figure SA.
Exon 9+ 1 ~ R N A 8 1 9 1 10 1416bp = 913s
Exon 9- mRNA
Figure 5B.
Figure 5C.

Figure 6. Quantification of nasal and vas deferens CFTR m W A . The
density of cDNA fragments on the autoradiographs was measured by scanning
densitometry. The proportions of nasal and vas deferens exon 9+ mWA, as a
percentage of total CFTR transcripts, were determined by using the total
densitometric units of both transcripts as 100%.

I 7T19T 1 7Tff T 1 5 ~ 1 7 ~ 1 Sublect no. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Figure 6.

Figure 7. Relationship between CFïR IVS8-T genotype and proportion of
exon 9+ transcripts in nasal epitheiia (*) or vas deferens (O). Each specimen
is represented by a separate symbol. Horizontal bars indicate mean values.

Vas Nasal Via8 Nasal V Nasal
5 T M 7TRT 7T19T
Figure 7.

Figure 8. Cornparison of the proportion of C F ï R exon 9+ trsnscnpts
within nasal epithelia and vas deferens from the same subject. Intra-
individuai values are c o ~ e c t e d by straight lines.
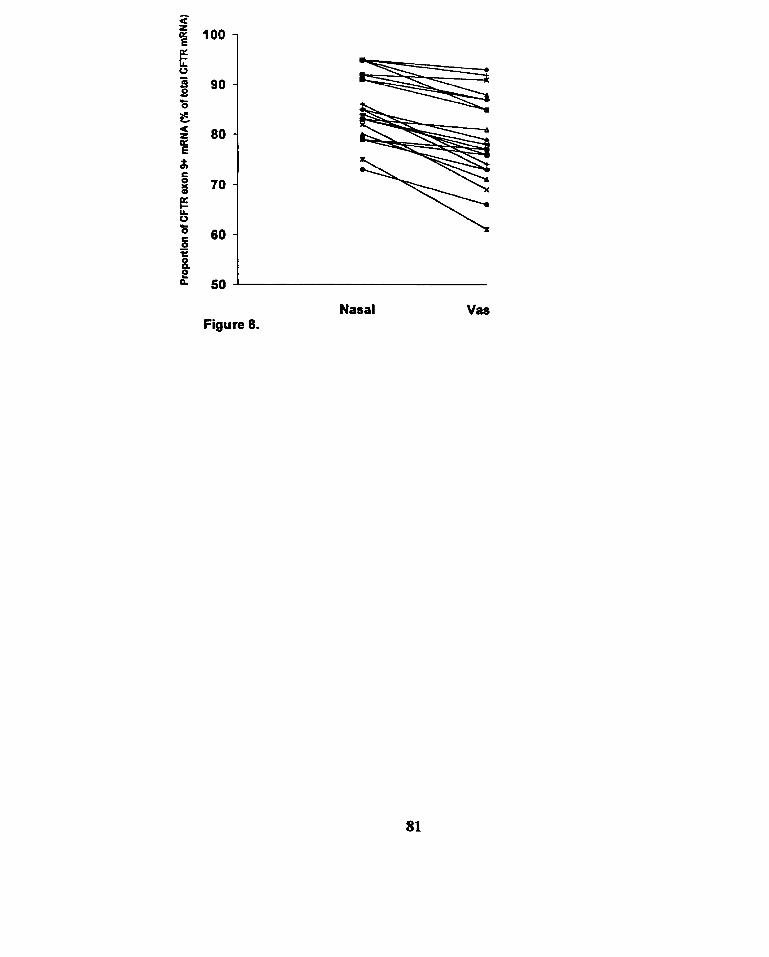
Nasal Figure 8.
Vas

Figure 9. Influence of CFTR genotype on phenotype by the proportion of
normal CM.R mRNA. A typical patient with CBAVD rnay have a severe
CFTR gene mutation (e.g., AF508) in one CFTR allele and the less efficiently
splicing 5T variant in the other. He may produce a suficient proportion of
normal (exon 9+) CFTR mRNA (and therefore protein) fiom the 5T ailele in the
lung (32% of total CFTR transcripts) to confer a normal phenotype but not so in
the reproductive tract (26%), thus giving him the isolated absence of vas deferens
phenotype without any pulmonary manifestations. In contrast, a typical CF
carrier may have a severe CFTR mutation in one copy of the CFTR gene but the
more eficiently splicing 7T variant in the other chromosome. Although he will
produce less of the normal CFTR mRNA in the reproductive tract (38%)
comparecl to the lung (41%) fiom the 7T allele, lmth proportions exceed the
threshold for normalcy, thus giving him a completely normal phenotype. The
threshold level for normal/abnomal phenotype depicted in the figure is arbitrary
and may vary between different tissues.

Congenital 811. Absence of Vas Deferens Cystic Fibrosis Camer
CFTR mut IlF cFTRmu/7T
1 :g
nomulphmnotyp. H . . . . . . . . . . . . . . . . . . . rbnomul ph- g 4kKmnrl phmnotyp.
;ii 1: &
Lung WdCRan Lung Wolffian
Figure 9.

APPLIED A IMAGE. lnc 1653 East Main Street - -. - - Rochester, NY 14609 USA -- -- - - Phone: il 61482-0300 -- -- - - Fax: 7161288-5989
Q 1993. Applied Image, Inc., All Righls Reserved




