
Establishing historical baselines of benthic diversity … · 1 Establishing historical baselines...
85
1 Establishing historical baselines of benthic diversity and community composition, Western Greenland Irina Chemshirova CID: 00643328 Supervisor: Dr Chris Yesson Word count: 5650 BSc Biology with a Year in Industry/ Research
Transcript of Establishing historical baselines of benthic diversity … · 1 Establishing historical baselines...

1
Establishing historical baselines of benthic diversity
and community composition, Western Greenland
Irina Chemshirova
CID: 00643328
Supervisor: Dr Chris Yesson
Word count: 5650
BSc Biology with a Year in Industry/ Research

2
Abstract
Historical data is vital to our understanding of how human-induced changes
affect ecosystems. One of these changes is overharvesting of fish stocks, sometimes
through the means of bottom trawling. This is a major disturbance to the benthos due
to the prolonged contact that fishing gear has with the seabed. It is difficult to find
comparable unimpacted sites in the present day in order to quantify the effect of
trawling. Therefore this study examines historical photographs taken by Per
Kanneworrff (Greenland Institute of Natural Resources) between 1977 and 1984. The
analyses revealed that trawling intensity had negligible effect on the communities
formed. Substrata was found to be the main factor influencing community composition.
We hypothesise that the short window of time (4 years for some stations) we looked at
was not sufficient for any changes due to trawling activity to occur.
Introduction
The human population is growing rapidly and is set to reach 8.9 billion by 2050
(Cohen, 2003). This growth is putting a strain on many ecosystems throughout the
world, with very few still remaining in a truly pristine state. It is very difficult to
quantify the effect we are having on a system after it has been exploited. Therefore we
must turn to historical data. This is an opportunity to re-construct ecosystems to an
unimpacted state, before wide-ranging exploitation began, thus giving us a reference
point for comparison with data collected more recently (Vellend et al., 2013). Swetnam,
Allen & Betancourt (1999) highlight the growing need for pursuing historical data to
determine the processes behind the current patterns and give us some predictive
power for the future. Extensive examination of historical data can also allow us to
avoid shifting the baseline of the ecosystems being studied. A shifting baseline is a

3
phenomenon which occurs when comparisons are drawn between a present day
impacted system with one which has already been significantly degraded (Lotze &
Worm, 2009, Roberts, 2003). When only looking at recent data we also tend to
underestimate the effects that even seemingly primitive methods of exploitation have
had on ecosystems (Pauly, 1995).
Whilst historical data is immensely useful it also presents many challenges. The
datasets are often incomplete. The records can be ambiguous and outdated, using old
taxonomic classifications (Mladenoff et al., 2002). Nonetheless the contribution they
make to our understanding of how the changes imposed upon ecosystems affect them
far outweighs these difficulties.
Historical data is invaluable when attempting to quantify various anthropogenic
effects. For example, it has been used to determine changes in carbon sequestration of
both forests and oceans over time (Rhemtulla, Mladenoff & Clayton, 2009, Cubillos et
al., 2007). Museum collections can often shed light on what an ecosystem looked like in
the past (Hoeksema et al., 2011). Changes in the distribution and population density of
many groups have been studied using various historical sources (Terry, 2010, Skelly et
al., 2003).
We alter much of the environment through the exploitation of populations which
are of economic importance. The Atlantic cod (Gadus morhua) is a classic example.
Rosenberg et al. (2005) have shown, using archaeological records that with increased
fishing pressure the species have decreased in size. Cod is obtained largely through the
means of bottom trawling.

4
Bottom trawling is a fishing practice which has wide ranging impacts on the
ecosystem being exploited. It usually targets organisms that live on the seabed e.g.
crustaceans and demersal fish. This practice is often thought to be highly destructive to
the benthos as the fishing gear is dragged along the seabed. The type of trawl gear used
is of importance when it comes to measuring the fisheries impact. In Western
Greenland, the otter trawl is widely used for the capture of the Northern shrimp
(Pandalus borealis). It typically consists of two trawl doors, (which can weigh up to 5
tonnes each) their function being to keep the net open. Rockhopper gear is an addition
to the otter trawl, developed around 1985. It is made up of bobbins and rollers attached
to the bottom end of the net. It evens out the seabed and dislodges any rocks which may
damage it (Valdemarsen, 2004). Collie et al. (2000) state that dredging has a much
greater impact on the seabed than trawling. Nonetheless, trawling impacts both the
target organism (in this case P. borealis) and the organisms which are caught in the net
as bycatch (Rumohr & Kujawski, 2000). Furthermore there is also the secondary
mortality imposed upon non-target organisms as they are irreparably damaged through
direct physical contact, but not necessarily removing them from their habitat (Jenkins,
Beukers-Stewart & Brand, 2001). Trawling has also been shown to alter nutrient
delivery in the benthic system. Epifaunal organisms receive nutrients in large bursts as
opposed to the usual steady stream (Pilskaln, Churchill & Mayer, 1998).
Jennings et al. (2001) have reported reduced biomass of infaunal and epifaunal
organisms in heavily trawled areas, however they did not find any changes in trophic
structure. Trawling can alter the functional diversity of a community. Tillin et al. (2006)
have shown that with extensive trawling a community shift occurs. From attached filter
feeders towards mobile organisms and infaunal scavengers.

5
The simplification of the habitat which occurs with trawling should not be
underestimated. Much of the complex structures created by sessile organisms such as
soft corals are often levelled out by the trawling gear. Krieger & Wing (2002)
highlighted the importance of these organisms in a benthic ecosystem. They form many
associations with various tropic levels, from predators to animals which seek shelter.
Andrews et al. (2002) also link thriving fish stocks to the presence of healthy Gorgonian
coral gardens.
Considerable changes to the benthic community and its habitat occur after the
first time an area has been trawled (Auster & Langton, 1999). Many studies attempting
to quantify the effects of trawling in the present day are dealing with a system which
has already undergone considerable change (Garcia, Ragnarsson & Eiríksson, 2006).
Therefore this study focuses on historical image data from 1977 until 1984.
According to Hamilton, Brown & Rasmussen (2003) the west Greenlandic
fishery primarily exploited G. morhua until the early 1970s through the means of
bottom trawling until the stock eventually crashed. It has been recorded that a
movement form G. morhua to P. borealis occurred around 1972 when catches were
estimated at 10 tonnes a year (Figure 1).
Figure 1. Showing the biomass of the cod and shrimp catch of the West Greenland fishery 1950 - 2000, Adapted from Hamilton, Brown & Rasmussen (2003).
6
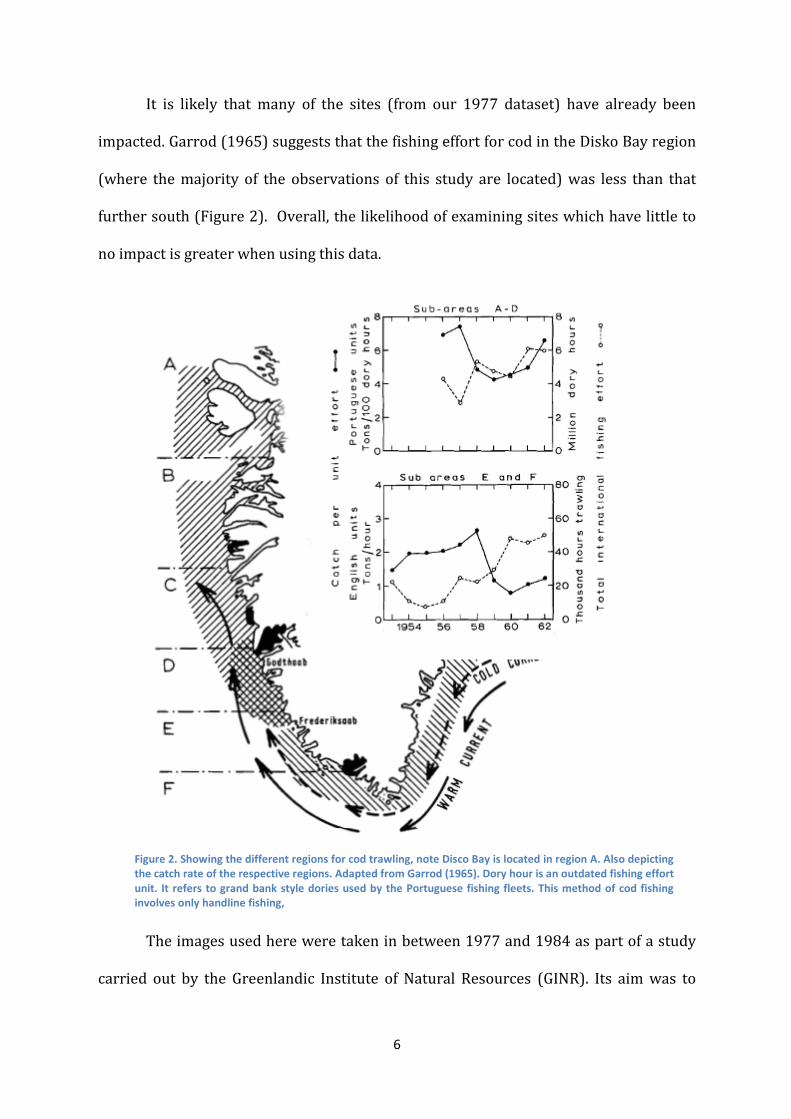
It is likely that many of the sites (from our 1977 dataset) have already been
impacted. Garrod (1965) suggests that the fishing effort for cod in the Disko Bay region
(where the majority of the observations of this study are located) was less than that
further south (Figure 2). Overall, the likelihood of examining sites which have little to
no impact is greater when using this data.
The images used here were taken in between 1977 and 1984 as part of a study
carried out by the Greenlandic Institute of Natural Resources (GINR). Its aim was to
Figure 2. Showing the different regions for cod trawling, note Disco Bay is located in region A. Also depicting the catch rate of the respective regions. Adapted from Garrod (1965). Dory hour is an outdated fishing effort unit. It refers to grand bank style dories used by the Portuguese fishing fleets. This method of cod fishing involves only handline fishing,

7
estimate the density of the shrimp population without the need for survey trawls
(Kanneworff, 1978) . This is an opportunity to observe what the seabed looked like
before large-scale trawling has occurred.
This project aims to compare data obtained from these images in the 1970s and
the 1980s. The 70s dataset will serve as a pre-intensive fishing baseline to compare
with the 80s dataset.
We also aim to collect data from a single year of the historical dataset (1984)
and use the historical trawling effort data available (1975 to 1984), in order to see if a
relationship can be inferred.
Materials and Methods
Kanneworff (1979) used a 35 mm robot camera with a flash and power unit in
waterproof housing to take the images processed here. The equipment was lowered to
the seabed using a winch wire, an image was taken once every minute after the bottom
sensor detected the seabed.. The equipment was allowed to drift whilst the ship
followed it using its echolocation system. The total area of an image is 3.39 m2
Much biodiversity and community analysis is dependent on correct taxonomic
identification. Therefore two datasets will be generated. One which contains organisms
identified to various taxonomic levels (e.g. some at order others at family level
Appendix 1.1) and another in which all are at the Class level (Appendix 1.2). This aims
to check how a variety of taxonomic levels may influence the statistical analysis.

8
The camera film was provided by GINR. It consisted of 56 reels of Kodak Safety
Film 5036 (ISO 400), spanning from 1975 until 1986. Location data was not available
for the stations from 1975 and 1976 and therefore could not be used. Using the location
data provided, the rest of the stations were placed on a map using QGIS (Figure 3).
Figure 3. Map of the stations used in this study, showing the cumulative amount of trawling occurred in the area between 1975 and 1984. This area overlaps with regions A and B for cod fishing (Figure 2)

9
The Kanneworff team aimed to visit the same stations each year of their survey,
however due to poor weather conditions this was not always possible. For two stations
to be regarded as a pair they can be a maximum of 5 km apart.
We combined the images from 1977, 1978, 1979 and 1980 to make the 70s pre-
intensive fishing dataset as this allowed for the largest number of paired stations (19 in
total) thi allowed for a minimum 4 year gap for most stations. The relevant stations
form the 1984 reels were used for the 1980s data. All 36 stations from 1984 were
digitized.
Ten images were digitized from each of the chosen stations using a Reflecta i-
scan 3600 at a resolution of 3600 dpi to ensure high image quality. An air duster
containing 1, 1, 1, 2 – Tetrafluroethane was frequently used to limit the dust particles
which can reduce image quality. The batch- processing function in Photoshop Elements
8 was used to improve image sharpness and colour. A total of 567 images were
digitized, only 295 (five per station) were processed, with the best quality images being
chosen for each station.
Image Processing
The processing of an image usually involves identifying an organism and
counting how many of it were present in the current image. Most of the identifications
were made based on prior knowledge of the study system. An ID guide was compiled in
2013 in order to facilitate processing of the 2011-2013 image data collected for the
same region (Appendix 1.3), this was referred to in case of doubt. As the image quality
was sometimes not as high as desired some taxa were counted as “unkown”. This
classification was used for organisms which were deemed alive but the image was not

10
high quality enough to allow for the distinction of any defining feature. Sensitive taxa
were also noted as organisms which are upright and thus more vulnerable to damage
from the trawl gear (Appendix 1.1 highlights which taxa were classed as sensitive)(de
Juan, Demestre & Thrush, 2009). After all of the organisms visible in the image were
counted and identified, a substrata type was assigned to each image (hard or soft).
Examples of the substrata classification can be seen in Figure 4. The presence/ absence
of bioturbation was also noted. Bioturbation is often a sign of the presence of infaunal
organisms which burrow into soft substrata.
Analysis
Trawling impact data was available for the period between 1975 and 1986. Since
we do not have any observational data from 1984 onwards the trawling data from the
two remaining years was excluded. A grid of 20 x 20 km was used to represent annual
trawling impact (Yesson, 2014, pers. com). Trawling data for the 70s was the sum of
trawl durations for three years before the observation. If the observation was made in
1979 the trawling impact number for that station would be the sum of the trawling
occurred from 1977 until 1979. In order to maintain consistency the same was carried
out for the 80s data, thus the trawling impact included was for the years 1982-1984.
Figure 4. Images of hard and soft substrata respectively

11
The trawling impact data for both time periods was treated as a continuous explanatory
variable. Shapiro – Wilks test was carried out on both datasets to assess if they were
normally distributed. Both failed the normality test (W=0.48, p<0.001 for 1970s and
W=0.63, p<0.001 for the 1980s). Therefore both were log transformed to the base of e.
After the transformation was carried out the data was again tested for normality, and it
failed (W=0.83, p<0.01 and W=0.85, p<0.01 for the 70s and 80s respectively). However
as this improved the distribution of the data, the log transformed version was used for
the analysis.
Historical comparisons
Species accumulation curves for each time period were composed in order to
asses if the sampling effort was sufficient (Ugland, Gray & Ellingsen, 2003).
The following diversity measures were calculated for each station: taxon
richness (α diversity), number of individuals, Pileou’s evenness and the Shannon-
Weiner Index. It is a composite measure attempting to combine both richness and
evenness of the community, it was calculated using Eq1.
∑
Pileou’s evenness was calculated using the H’ derived from the Shannon-Weiner Index,
Eq 2. It attempts to quantify how equal the community is and thus acts as a correction
factor for the Shannon – Weiner Index.
(Eq. 1)

12
All diversity measures were treated as continuous response variables. Scatter
plots of the different diversity indices from both periods against one another were
produced in order to visualise what changes may have occurred.
Wilcoxon signed-rank paired tests were performed in order to determine if
there is any significant difference in the diversity measures between the two time
periods. The changes in taxon richness were visualised using QGIS, where a map
showing the increase or decrease in richness was produced with the relative fishing
intensity. .
Non - metric Multi-dimensional Scaling (nMDS) is an ordination representing the
variation in community composition in a reduced number of dimensions. It has been
used to visualise the community composition of both time periods. Environmental
vectors were overlaid onto this analysis to determine if they can provide an explanation
of patters observed. The environmental vectors included in the analysis were: Latitude,
Longitude, Depth, Fishing (Trawling) Impact, Substrata and Bioturbation.
A hierarchical cluster analysis was also performed in order to compare and
contrast the patterns highlighted by the nMDS analysis. This classifies the different
stations based on the number of dissimilarities between them using Euclidian distance
as a measure. A dendogram using the complete clustering method was generated for
the data from both historical periods.
(Eq. 2)

13
A linear regression of taxon richness and trawling impact was carried out to
determine if there is a relationship. Both time periods were plotted on the same scatter
plot.
β diversity has been calculated using the Bray – Curtis dissimilarity method to
quantify the diversity change over time. This takes into account that, diversity indices
may not vary between the two time periods but the taxon identities might i.e. a shift
from soft corals to decapods. A linear regression was carried out to determine if
trawling impact affects the amount of change observed. A high number of β diversity
indicates a large change in the taxon identities.
Historical comparisons – Class level
All of the analyses described were then repeated for the dataset in which all
organisms are identified at the Class level.
Single year historical dataset (1984)
Further analysis was also pursued with the 1984 dataset. The same diversity
indices were calculated. A linear regression was performed to determine if there is a
relationship between trawling impact and diversity. nMDS and cluster analyses were
also performed. The ward method was used for the clustering of this dataset. A species
accumulation curve was also estimated.
With these additional analyses, many of the results did not differ from the initial
historical comparison. Therefore they have been placed in the Appendix.
All analysis was performed using the statistics software R and the following
packages: vegan and plyr (R Core Team, 2013, Oksanen et al., 2013, Wickham, 2011).

14
Results
Historical comparisons
For the comparison between the 70s and the 80s 19 stations were processed,
giving a total of 38. Overall 38 taxa were identified across both time periods, including
an additional classification for “unknown” organisms. Encrusting bryozoa was the most
abundant taxon in both time periods. Overall out of the five most numerous taxa, three
belonged to the sensitive taxa group (Figure 5). Sabellidae and ascidians were found to
decrease in abundance over time. Brittle stars, soft bryozoans, stylasterina, serpulidae
and soft coral on the other hand have increased over time (Appendix 1.4).
PterasteridaePolynoidae
IsopodaSea SpidersScaphopods
ScorpaeniformesAstropectinidae
Sea UrchinsEunicidae
RajiformesCrinoidsBivalves
AsteriidaeStarfish Other
GastropodsSea Cucumbers
GoniasteridaeZoanthids
EchinasteridaeTerebratulida
PleuronectiformesHydroids
PerciformesSoft CoralsSabellidaeAnemones
UnknownSerpulidae
Arborescent SpongesStylasterinaBrittlestars
Encrusting SpongesSoft Bryozoa
Massive SpongesDecapoda
Erect BryozoaAscidians
Encrusting Bryozoa
Total
0 200 400 600 800
Figure 5: Shows the taxa identified and their relative abundance for the 70s and 80s datasets. Sensitive taxa have been highlighted

15
The species accumulation curve produced showed that increasing the sampling
effort would not lead to new undiscovered taxa being added (Figure 6). This was also
separetly performed for the two time periods and showed the same pattern (Appendix
1.5). The chao method estimated that there should be 41 taxa (SE=3), whilst we found
38.
0 10 20 30
01
02
03
04
0
Sites
Ta
xa
Figure 6. Shows the curve beginning to asymptote suggesting that the sampling is sufficient.
16
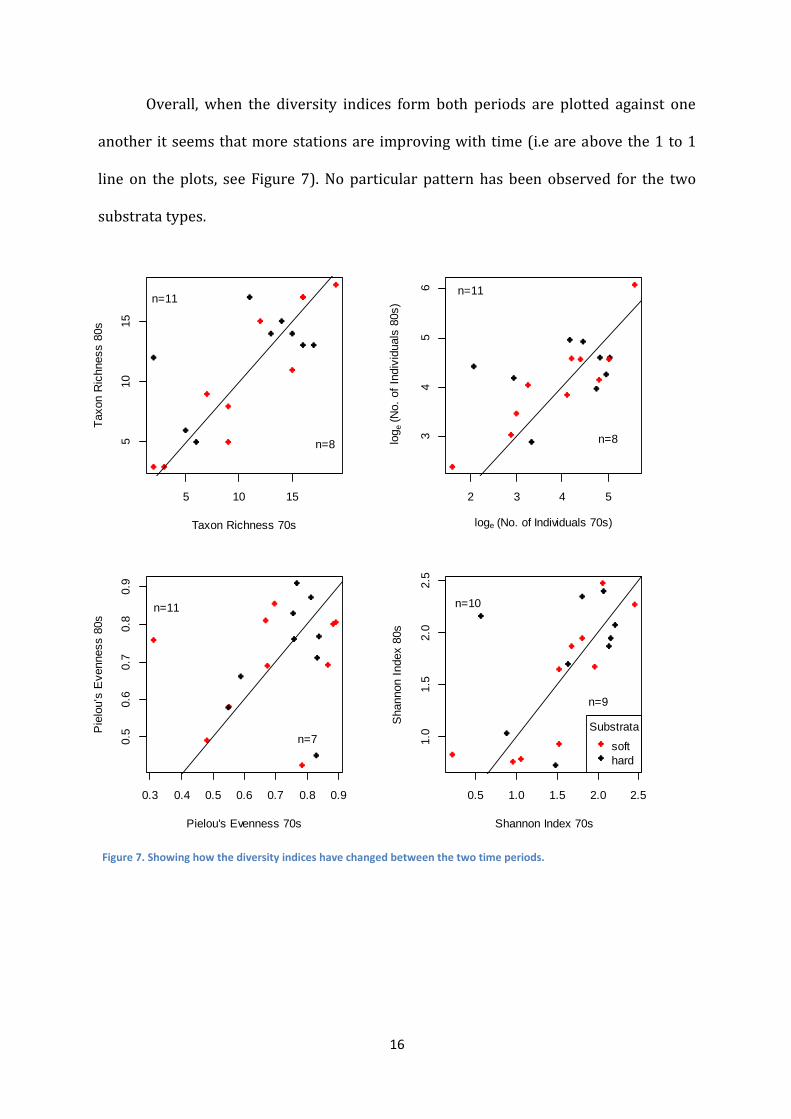
Overall, when the diversity indices form both periods are plotted against one
another it seems that more stations are improving with time (i.e are above the 1 to 1
line on the plots, see Figure 7). No particular pattern has been observed for the two
substrata types.
Figure 7. Showing how the diversity indices have changed between the two time periods.
5 10 15
510
15
Taxon Richness 70s
Taxon R
ichness 8
0s
n=11
n=8
2 3 4 5
34
56
loge (No. of Individuals 70s)
log
e(N
o.
of
Indiv
iduals
80s)
n=11
n=8
0.3 0.4 0.5 0.6 0.7 0.8 0.9
0.5
0.6
0.7
0.8
0.9
Pielou's Evenness 70s
Pie
lou's
Evenness 8
0s
n=11
n=7
0.5 1.0 1.5 2.0 2.5
1.0
1.5
2.0
2.5
Shannon Index 70s
Shannon I
ndex 8
0s
Substrata
soft
hard
n=10
n=9
17
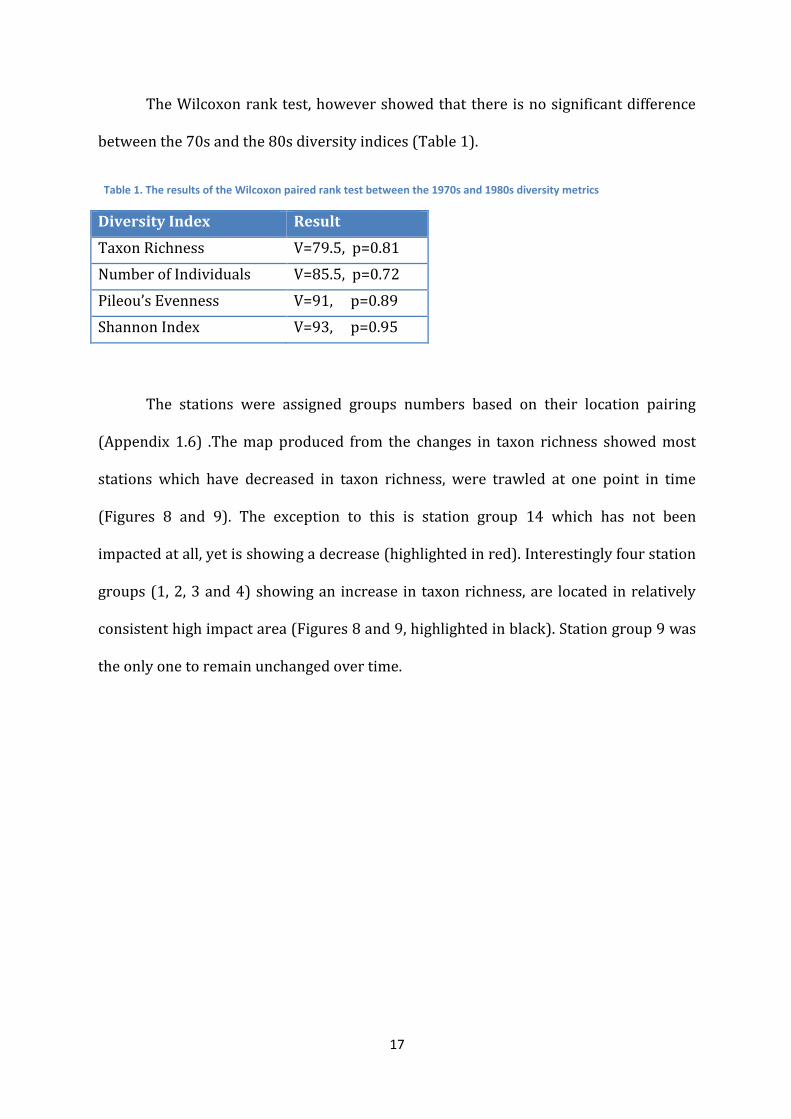
The Wilcoxon rank test, however showed that there is no significant difference
between the 70s and the 80s diversity indices (Table 1).
Diversity Index Result
Taxon Richness V=79.5, p=0.81
Number of Individuals V=85.5, p=0.72
Pileou’s Evenness V=91, p=0.89
Shannon Index V=93, p=0.95
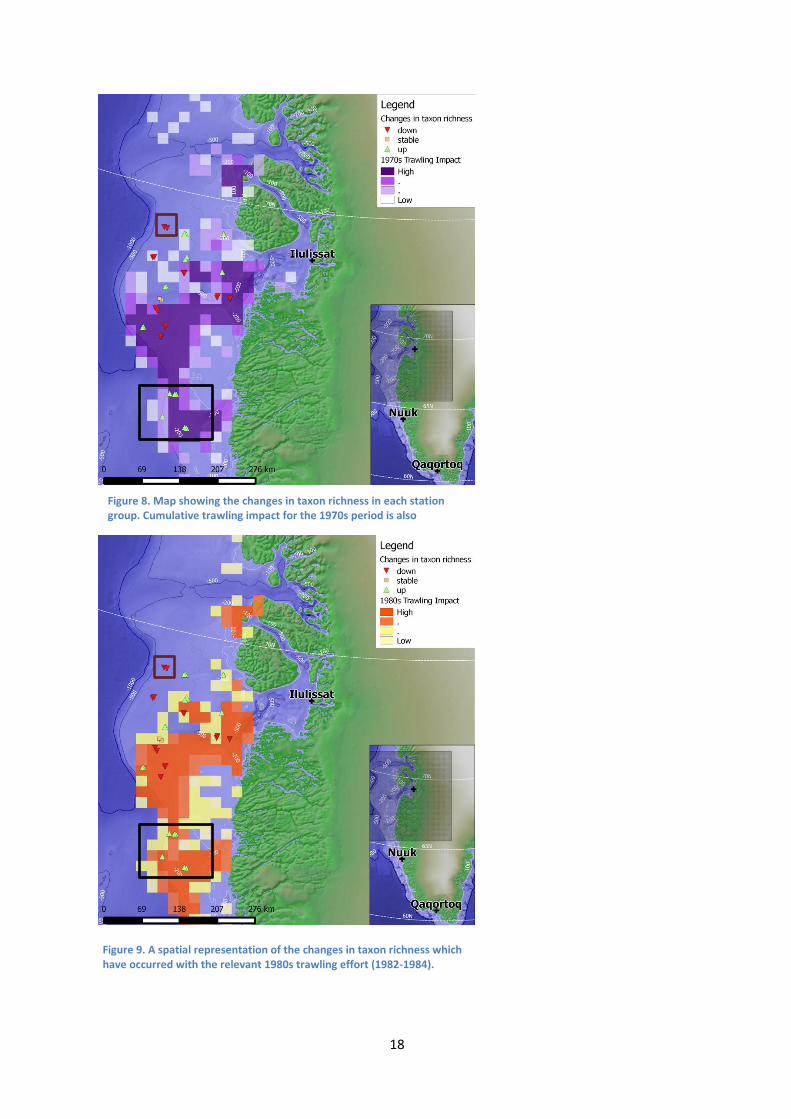
The stations were assigned groups numbers based on their location pairing
(Appendix 1.6) .The map produced from the changes in taxon richness showed most
stations which have decreased in taxon richness, were trawled at one point in time
(Figures 8 and 9). The exception to this is station group 14 which has not been
impacted at all, yet is showing a decrease (highlighted in red). Interestingly four station
groups (1, 2, 3 and 4) showing an increase in taxon richness, are located in relatively
consistent high impact area (Figures 8 and 9, highlighted in black). Station group 9 was
the only one to remain unchanged over time.
Table 1. The results of the Wilcoxon paired rank test between the 1970s and 1980s diversity metrics
18
Figure 8. Map showing the changes in taxon richness in each station group. Cumulative trawling impact for the 1970s period is also represented.
Figure 9. A spatial representation of the changes in taxon richness which have occurred with the relevant 1980s trawling effort (1982-1984).

19
Diversity indices such as these however reveal little in the way of the various
communities in the area. Therefore we carried out nMDS plots which reveal the
relevant communities for both time periods.
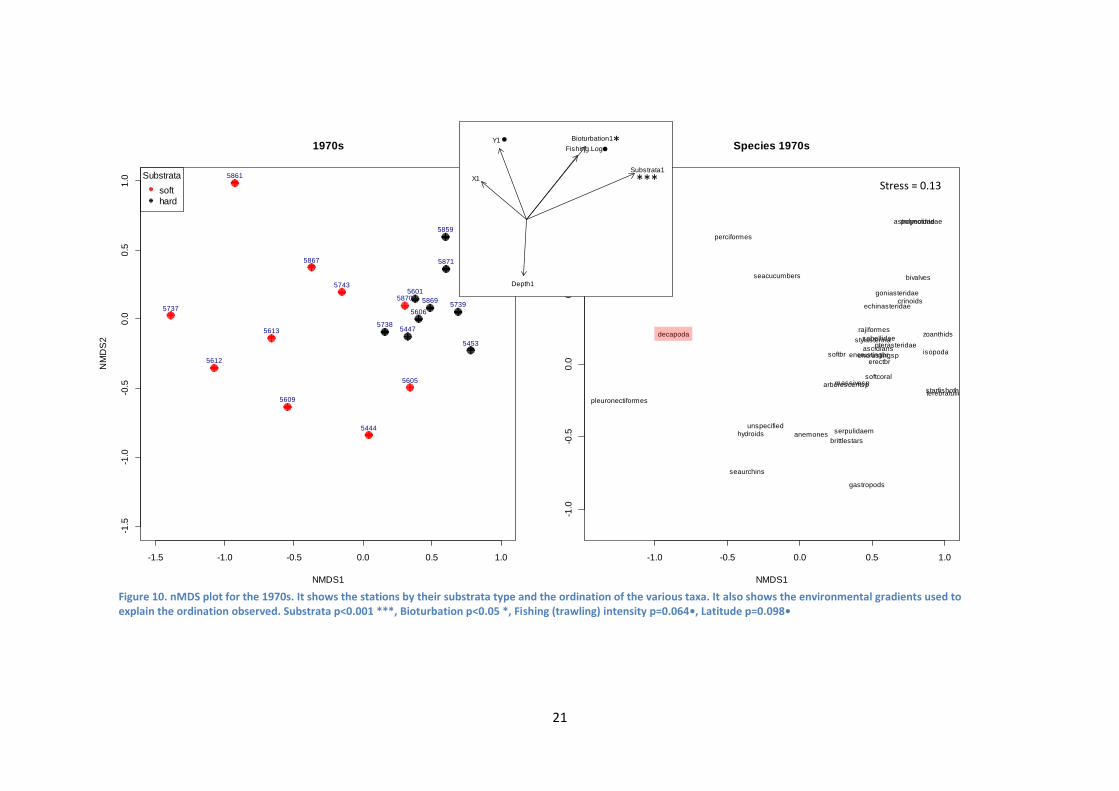
In the 1970s substrata was shown to be an important factor in differentiating
between the two communities observed in the nMDS plot (p=0.001, ***, Figure 10).
The other environmental vectors overlaid on the nMDS plots highlighted that
substrata type has a very strong influence on the community composition of the 1970s.
Interestingly, for the 1970s period, trawling effort appears to have a greater impact on
hard substrata. Considering the species plot, decapods (which primarily consist of P.
borealis, the target species of the fishery) appear to not be affected by the trawling
(Figure 10, highlighted in red). Trawling impact has shown to have very little influence
on the community composition (p=0.064, •) along with Latitude (p=0.098, •).
Bioturbation also had some effect, in a different direction to substrata type (p<0.05,*).
Depth and Longitude were not shown to explain any of the observed variation.
For the 1980s only substrata explained the patterns observed in the nMDS plot
(p=0.001, ***, Figure 11). In this species plot, decapods appeared where expected with
respect to trawling (Figure 11, highlighted in red).
An alternative approach to viewing the relationships between stations in terms
of communities is a cluster analysis. This method uses a dendogram to show how the
stations cluster together based on the taxa they contain. It showed some stations which
were designated as hard substrata in the soft substrata cluster (Figure 12). In the hard
substrata cluster (highlighted in black, Figure 12), the dominant taxa were encrusting
sponges and bryozoans along with ascidians and soft corals. In the soft substrata cluster

20
(highlighted in red, Figure 12) decapods were found to be most numerous. Stations
5606 and 6716 (group 3) have been shown to be very different in terms of their
composition compared to the other stations (highlighted in green, Figure 12). Many of
the stations occur very far from their paired station in the dendogram, e.g. 5605 and
6720 (group 4, highlighted in blue, Figure 12). It is also worth noting that many stations
pairs which have been placed in the same substrate cluster are still described as quite
different from one another e.g. 5609 and 6713, group 1, highlighted in purple, Figure
12.
21
-1.5 -1.0 -0.5 0.0 0.5 1.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1970s
NMDS1
NM
DS
2
5609
5453
5606
5605
5601
5739
5447
5867
5612
5613
5743
5738
5870 5869
5871
5859
5444
5861
5737
Substrata
soft
hard
-1.0 -0.5 0.0 0.5 1.0-1
.0-0
.50
.00
.51
.0
Species 1970s
NMDS1
unspecified
softcoral
anemones
zoanthids
hydroids
stylasterinapterasteridae
echinasteridae
goniasteridae
astropectinidae
starfishother
brittlestars
seaurchins
seacucumbers
crinoids
encrustingsp
massivesparborescentsp
sabellidae
serpulidaem
polynoidae
decapoda
isopoda
gastropods
bivalves
terebratulida
erectbrencrustingbrsoftbr
ascidians
rajiformes
perciformes
pleuronectiformes
NMDS1
NM
DS
2
X1
Y1
Depth1
Fishing.Log
Substrata1
Bioturbation1
*
***
• •
Figure 10. nMDS plot for the 1970s. It shows the stations by their substrata type and the ordination of the various taxa. It also shows the environmental gradients used to explain the ordination observed. Substrata p<0.001 ***, Bioturbation p<0.05 *, Fishing (trawling) intensity p=0.064•, Latitude p=0.098•
Stress = 0.13

22
-1.0 -0.5 0.0 0.5 1.0 1.5
-1.0
-0.5
0.0
0.5
1.0
1980s
NMDS1
NM
DS
2
6713
6715
6716 6717
6720
6721
6722
6724
6727
6728
6729
67306731
6732
6733
6736
6737
6738
6749
Substrata
soft
hard
-0.5 0.0 0.5 1.0 1.5
-0.5
0.0
0.5
1.0
1.5
Species 1980s
NMDS1
unspecified
softcoral
anemoneszoanthids hydroids
stylasterina
asteriidae
echinasteridae
goniasteridae
astropectinidae
starfishother
brittlestars
seacucumbers
encrustingsp
massivesp
arborescentspsabellidae
eunicidae
serpulidaemdecapoda
seaspiders
gastropods
bivalves
scaphopods
terebratulida
erectbr
encrustingbr
softbr
ascidians
rajiformes
scorpaeniformes
perciformes
pleuronectiformes
NMDS1
NM
DS
2
X2
Y2
Depth2
Fishing.LogSubstrata2
Bioturbation2
***
Figure 11. nMDS plot for the 1980s. It shows the stations by substrata type and the ordination of the various taxa. It also shows the environmental gradients used to explain the ordination observed - Substrata p<0.001 ***
Stress = 0.14

23
67
37
67
33
56
05
67
13
58
67
67
24
67
28
67
49
56
13
57
37
67
21
67
29
54
44
56
09
57
43
56
12
58
61
67
27
57
38
58
70
67
38
58
59
67
30
67
31
58
71
54
47
56
01
58
69
67
15
67
32
54
53
67
22
57
39
67
20
67
17
67
36
56
06
67
16
05
01
00
15
02
00
Cluster Dendrogram
hclust (*, "complete")
all.dist
He
igh
t
Figure 12. Dendogram of the hierarchical clustering. Highlighted in red are the stations with a soft substrata classification, stations highlighted in black classed as hard substrata. Stations with an * beside them are classed as hard substrata but have been grouped with the soft by the analysis. The numbers below denote the station groups based on their location.
17 15 4 1 8 8 10 19 10 19 6 11 17 1 11 9 18 9 12 13 18 16 12 13 15 7 5 14 2 14 2 7 6 5 4 16 3 3
* * *

24
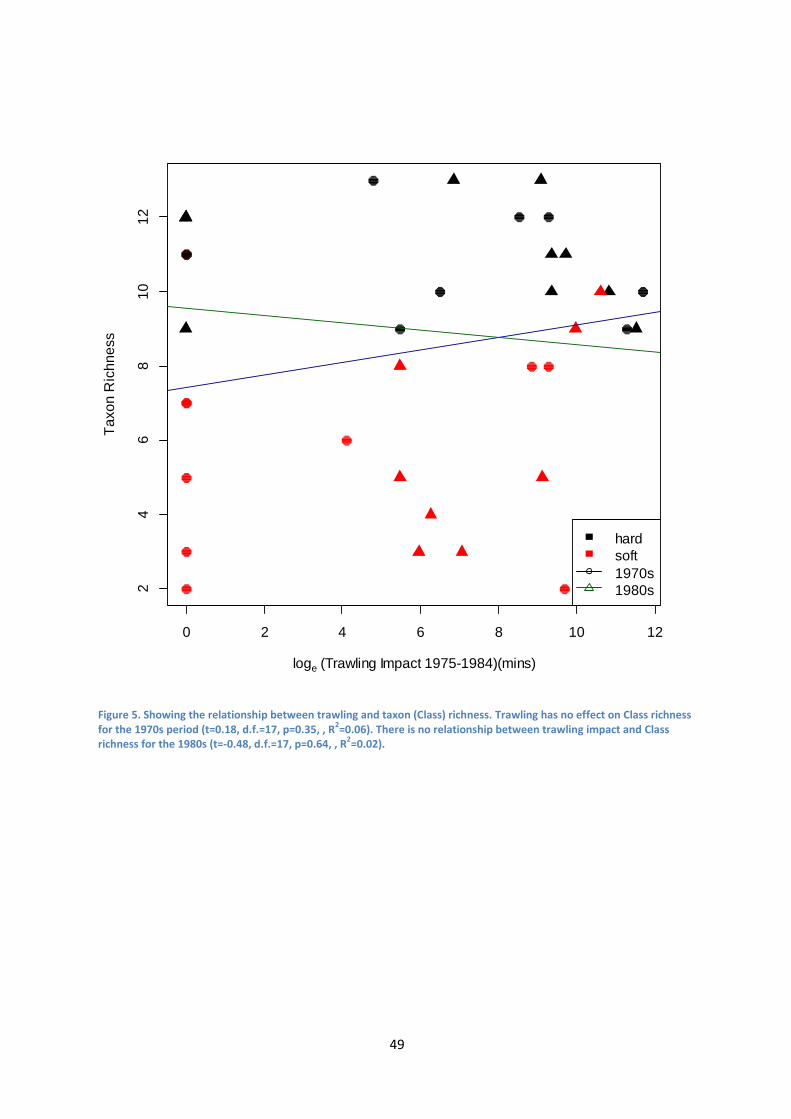
No relationship could be inferred between increasing trawling intensity and
taxon richness. Linear regressions for both the 1970s and 1980s showed no particular
patterns in terms of positive or negative effects of increasing trawling intensity on
taxon richness (Figure13).
There is no relationship between β diversity and trawling pressure as shown by
the linear regression in Figure 14.
0 2 4 6 8 10 12
51
01
5
loge (Trawling Impact 1975-1984)(mins)
Ta
xo
n R
ich
ne
ss
hard
soft
1970s
1980s
Figure 13. Showing the relationship between trawling intensity and taxon richness. Trawling has no effect on taxon richness for the 1970s period (t=1.4, d.f.=17, p=0.17). There is no relationship between trawling impact and taxon richness for the 1980s (t=0.3, d.f.=17, p=0.54).

25
Historical comparisons – Class
The class level analysis was performed because community and diversity indices
can often be influenced by the level of taxonomic resolution used for the identification
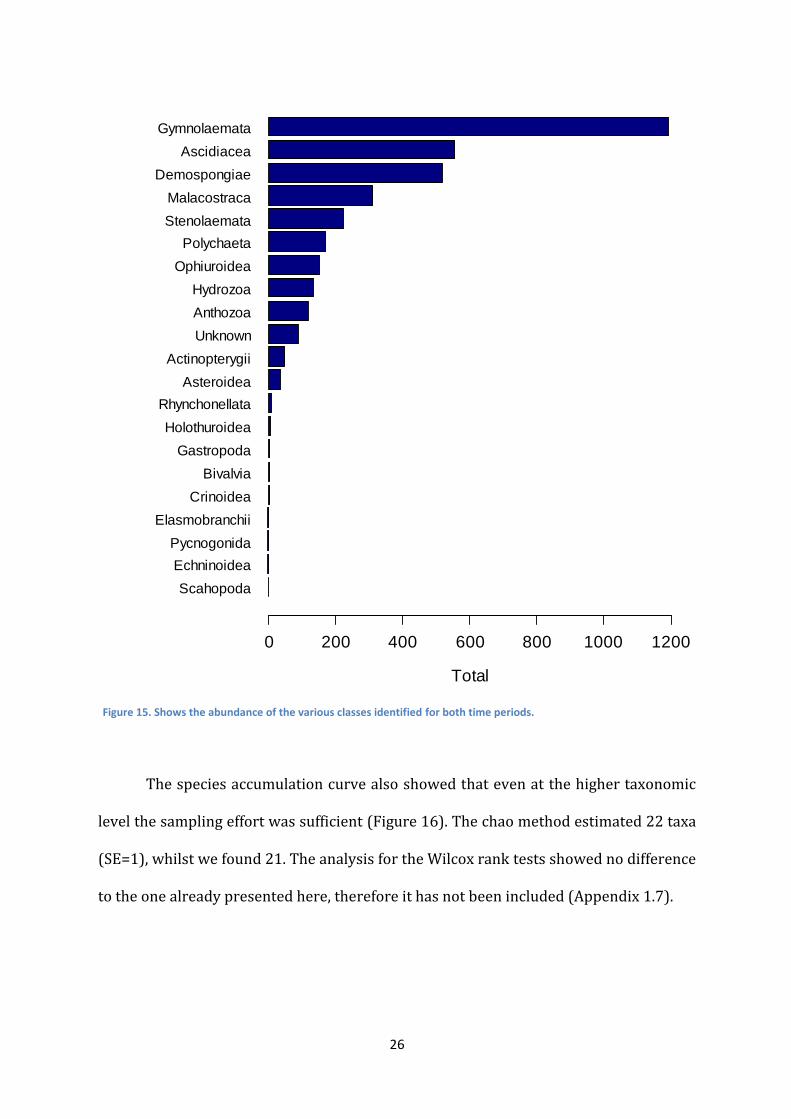
of organisms. Bryozoa were still the most abundant of taxa (Gymnolaemata) at class
level, followed by Ascidiacea and Demospongiae (Figure 15).
0 2 4 6 8 10
0.2
0.3
0.4
0.5
0.6
0.7
loge (Trawling Impact 1982-1984)(mins)
Be
ta D
ive
rsity
Substrata
hard
soft
Figure 14. Showing the relationship between β diversity and trawling impact. Increasing trawling impact had no effect on the β diversity observed (t=0.77, d.f.=17, p=0.45)
26
The species accumulation curve also showed that even at the higher taxonomic
level the sampling effort was sufficient (Figure 16). The chao method estimated 22 taxa
(SE=1), whilst we found 21. The analysis for the Wilcox rank tests showed no difference
to the one already presented here, therefore it has not been included (Appendix 1.7).
Scahopoda
Echninoidea
Pycnogonida
Elasmobranchii
Crinoidea
Bivalvia
Gastropoda
Holothuroidea
Rhynchonellata
Asteroidea
Actinopterygii
Unknown
Anthozoa
Hydrozoa
Ophiuroidea
Polychaeta
Stenolaemata
Malacostraca
Demospongiae
Ascidiacea
Gymnolaemata
Total
0 200 400 600 800 1000 1200
Figure 15. Shows the abundance of the various classes identified for both time periods.

27
The nMDS plots for both periods showed that substrata type is the most
important environmental factor governing the community composition (Appendix 1.8).
The cluster analysis mostly maintained the main patterns observed in the
previous dataset (Appendix 1.9). The linear regressions carried out for the taxon
richness and β diversity again were unaffected by increasing trawling intensity
(Appendix 2.0 and 2.1 respectively).
0 10 20 30
51
01
52
0
Sites
Ta
xa
Figure 16. The number of new taxa being identified is shown to asymptote with increasing sampling effort.

28
Single year historical dataset (1984)
The species accumulation curve again showed that increased sampling effort
would unlikely uncover additional taxa (Appendix 2.2). Chao estimate stated that there
could be 40 taxa (SE=3), we found 38. The nMDS plots revealed substrate to be the most
important environmental factor in the community composition (Appendix 2.3).
Cluster analysis revealed three clusters. Cluster A contained true hard substrata
stations (Figure 17). Cluster B contained mainly soft substrata stations. Cluster C was
designated as mixed as it contained an equal number of hard and soft substrata
stations. Two stations were notably different from all the rest – 6716 and 6743
(highlighted in orange, Figure 17).
Linear regression analysis found that trawling intensity has no effect on taxon
richness in the 1984 dataset (Appendix 2.4).

29
67
42
67
44
67
48
67
20
67
17
67
36
67
14
67
25
67
23
67
21
67
26
67
29
67
19
67
41
67
27
67
34
67
28
67
49
67
38
67
24
67
40
67
30
67
31
67
46
67
18
67
37
67
15
67
35
67
22
67
32
67
45
67
47
67
13
67
33
67
16
67
43
02
00
40
06
00
80
0Cluster Dendrogram
hclust (*, "ward")
dist84
He
igh
t
* * * * *
Figure 17. A dendogram showing the 3 distinct clusters, referred to A, B and C respectively left to right.

30
Discussion
The number of taxa identified did not differ significantly when compared to
other studies. Jones, Bett & Tyler (2007) studied the megabenthos off the coast of East
Greenland and found 45 taxa at varying taxonomic levels through the means of bottom
photography. Whilst Cusson, Archambault & Aitken (2007) found 68 orders and 29
classes, when examining historical records from 1955 – 1977. The area they examined
was much larger, from Beaufort Sea in the west to the Southern Davis Strait in the west.
In terms of abundance, we found that the numbers of soft corals increased with time,
whilst Strain et al. (2012) found a decline. The Strain study was at a relatively shallow
depth (30m). They also had a temporal gap of 24 years compared to a minimum 4-year
gap in our study. Both of those factor may have contributed to the discrepancies with
our results. The increase of soft coral with time we have observed is a curious
phenomenon as they are vulnerable to fishing gear as they cannot escape it as easily as
mobile taxa. Looking back at the specific images where they were found the 70s dataset
contains more large soft corals than the 80s one. There was also an increase in brittle
stars, which have been classed as sensitive to trawling by Olsgard et al. (2008). The
reason for this dissimilarity could be that they have a different functional role in this
community e.g. as scavengers as opposed to filter feeders.
We did observe a slight decline in ascidian abundance, which is in line with
Strain’s (2012) findings. Sabellidae worms decreased in abundance greatly. Simpson &
Watling (2006) found a similar abundance pattern in the gulf of Maine when comparing
trawled and untrawled areas.
No significant change was observed in the diversity indices between the
two time periods. This would suggest that the communities have remained relatively

31
stable over the 4-year period. Kenchington et al. (2007) found little variation in species
richness when comparing two time periods with a 30-year gap. However their study
was based in the Bay of Fundy, Canada which is quite sheltered. This could be the
reason they did not see difference.
When the changes in richness are placed on a map, several station groups show
interesting patterns. Station group 14 (highlighted in red, Figures 8 and 9) is the most
curious one by far. It shows a decrease in both richness and number of individuals, yet
it has not experienced any trawling activity. When looking at this group closely it, the
80s station has only one taxon less than the 70s one. This is due to rare taxa, more
specifically Pterasteridae as it only appears once in the whole dataset, in that station. It
is therefore possible that the analyses are skewed due to rare taxa. Some studies have
excluded taxa with abundance lower than 5%, this would be something to consider
testing out for this study in the future (Frid et al., 2000). Four stations increased in
richness over time even though the area was heavily trawled in the four-year period.
Hill et al. (1999) suggest that this could be due to trawling activity sometimes bringing
up more stones to the surface creating more opportunities for colonisation by sessile
organisms. Diversity indices show if there has been a significant change in terms of
number of taxa but do not highlight how the community composition has changed.
Therefore we carried out an nMDS analysis.
The nMDS plot for the 1970s period reveal two main communities, one
associated with soft substrata and another with hard substrata. The environmental
vectors also highlighted the same pattern as substrata type being a significant driver in
determining the composition of the stations. Kenchington et al. (2007) show a much
weaker substrata effect in a similar analysis. The communities seemed to be largely

32
differentiated by the time period they belonged to as opposed to substrata.
Kenchington et al. (2007) attributed this to dredging and trawling activity occurring
between 1967 and 1997 changing the community composition. The area they
investigated was at depths between 100-200m so it is very likely that a different
community is concerned to the one this study has sampled. When both time periods
were plotted for this study, no pattern was observed (Appendix 2.5).
Bioturbation also affected community structure, this is likely to be due to the fact
that soft substrata is inhabited by infaunal and burrowing organisms which modify the
sediment regularly. However, this does not point in the same direction as substrata
which is unexpected, it seems to overlap with the fishing effort variable instead.
The trawling effort is giving a weak signal in terms of community structure. It is
possible that the strong substrata signal is masking the effect fishing is having on the
communities. Kaiser et al. (1998) analysed communities separately after they
concluded that there was a significant difference between the two due to substrata
type. This is something to consider if developing this project further. Another
possibility for the apparent lack of fishing influence on communities is the quality of the
fishing data itself. This dataset only includes one location for the trawls, i.e. it could be
start or end. Therefore a coarse grid of 20 x 20 km needed to be used in order to
account that an average trawl could be of length of 20 km. This means that the data is of
a relatively low resolution. Stations within the 20km square will show the same amount
of impact even if the trawl has not passed through there. Bradshaw, Veale & Brand
(2002) had a relatively poor quality fishing dataset when comparing benthic
community data from 1938-1950 with that from 1990. They attempted to solve this
issue by creating several proxies for fishing impact. The two that showed changes

33
between the communities were “number of years since first fished” and a “Fishermen’s
Index”. The number of years is a count of how long an area has been exploited. The
Fishermen’s Index was developed by conferring with local fishermen on how intensive
the fishing has been in the area by giving a score from 1 (being low) to 7 (being high).
Both of those methods for quantifying fishing effort would be interesting and possibly
more realistic alternatives to the current dataset.
It is possible that the cod fishery, which was active in the area almost 60 years
before the images were taken, has had a greater impact on the benthos than
anticipated. Therefore, the change which we are trying to quantify may have already
happened. We may be sampling a system in an already degrading state, thus shifting the
baseline.
When looking at the position of certain taxa in the nMDS plot it is strange to see
that the decapods were ordered with taxa at stations which are less affected by
trawling, This however, could still be the substrate effect, as the soft substrata were
trawled less intensively on average.
The 1980s MDS ordination showed similar patterns to the 1970s with one
notable difference. Substrata was the only environmental variable to explain the
communities observed. The fact that fishing did not have an effect was expected
because the initial impact should have been obvious in the first time period (70s) as the
initial impact on a community is often the greatest. Even though the effect of fishing
was negligible, decapods were placed where expected with respect to fishing.
The cluster analysis reinforced the main message of the nMDS analysis, that
substrata is the main factor determining the communities in this study. Kaiser et al.

34
(2002) described the importance of the sediment type when considering the effect
trawling may have on the benthos. Three stations which have been classed as hard have
however clustered with the soft substrata stations (6733, 6721 and 5738). The reason
for this is likely to be that these stations were relatively impoverished and contained a
few individuals from taxa generally associated with soft substrata e.g. decapods and
serpulids. Two stations (5606 and 6716) were very distinct from any of the other
stations and form one another. When the images from the stations were examined, it
was obvious that they were quite different substrata, being mostly large boulders with
few organisms. Considering the different station groupings it was surprising to see that
stations which have similar locations (within 5 km) would appear so far away from one
another in the dendogram e.g. group 4. When their substrata class was taken into
account the difference in clustering was likely because one station was soft substrata
and the other hard substrata. However groups which have been given the same
substrate classification e.g. group 1 have also occurred on distant branches on the
dendogram. This difference could be attributed to trawling pressure, as effort was
shown to increase for that station. It is worth noting that fishing effort is often patchy
thus it can be difficult to pin-point if the increase in effort occurred evenly in the 20 x 20
km square or if only a stretch of a few kilometres was affected (Cryer, Hartill & O'shea,
2002).
Linear regressions of trawling against taxon richness showed no relationship.
Separating out the different time periods had the same result. Although a non-
significant positive slope can be seen for the 1970s. This however is a substrate effect
again (Kaiser et al., 1998). It can be inferred from this that on average hard substrata
are more taxon rich than soft substrata.

35
β diversity showed no relationship, but it can be argued that overall soft
substrata have been subject to grater changes in the identities of species. One likely
reason for the lack of pattern observed in the regressions is the quality of the trawling
data.
The analysis performed at the Class level showed no major differences from the
data at various taxonomic levels. Therefore, our choice of taxon categories were similar
to choosing a consistent class level grouping. It is possible that more specific
identification e.g. genus would change our results. Nonetheless this is not realistic or
possible for many of the organisms when collecting data from images. Many taxa (e.g.
Porifera) need to be examined under a microscope in order to be assigned a more
specific taxonomic classification.
The analysis carried out on the 1984 data only, again differed little from the
original one. The only real difference was in the cluster analysis. Instead of forming two
distinct clusters, three were formed. Two were deemed hard and soft, whilst a third one
could be described as mixed. This could be due to the different clustering method used.
Two stations again showed to differ from the rest of the stations – 6716 and 6743. The
reasons for this are the same as pointed out earlier.
The main outcome of this study is that substrata is the key variable in
determining the communities in the Greenlandic benthos. It is possible that the effects
of trawling were not apparent in this study because the 4-year window is not enough
for the effects of trawling to truly manifest themselves. Processing more images per
station may give us a better idea of the community composition of this area. It would
also be interesting to compare present date image data with the historical data. Then
the community changes may become more apparent.

36
Acknowledgments
I would like to thank Kirsty Kemp and Chris Yesson at the Institute of Zoology for the
extensive help and guidance throughout this project. Many thanks to Henrik Lund at the
Greenland Institute of Natural Resources for alerting us to the existence of the historical
images and GINR for kindly providing them.
References
Andrews, A. H., Cordes, E. E., Mahoney, M. M., Munk, K., Coale, K. H., Cailliet, G. M. & Heifetz, J. (2002) Age, growth and radiometric age validation of a deep-sea, habitat-forming gorgonian (Primnoa resedaeformis) from the Gulf of Alaska. Hydrobiologia. 471 (1-3), 101-110.
Auster, P. J. & Langton, R. W. (1999) The effects of fishing on fish habitat. American Fisheries Society Symposium.
Bradshaw, C., Veale, L. & Brand, A. (2002) The role of scallop-dredge disturbance in long-term changes in Irish Sea benthic communities: a re-analysis of an historical dataset. Journal of Sea Research. 47 (2), 161-184.
Cohen, J. E. (2003) Human population: the next half century. Science (New York, N.Y.). 302 (5648), 1172-1175.
Collie, J. S., Hall, S. J., Kaiser, M. J. & Poiner, I. R. (2000) A quantitative analysis of fishing impacts on shelf‐sea benthos. Journal of Animal Ecology. 69 (5), 785-798.
Cryer, M., Hartill, B. & O'shea, S. (2002) Modification of marine benthos by trawling: Toward a generalization for the deep ocean? Ecological Applications. 12 (6), 1824-1839.
Cubillos, J., Wright, S., Nash, G., De Salas, M., Griffiths, B., Tilbrook, B., Poisson, A. & Hallegraeff, G. (2007) Calcification morphotypes of the coccolithophorid Emiliania huxleyi in the Southern Ocean: changes in 2001 to 2006 compared to historical data. Marine Ecology-Progress Series. 348 (October), 47-54.
Cusson, M., Archambault, P. & Aitken, A. (2007) Biodiversity of benthic assemblages on the Arctic continental shelf: historical data from Canada. Marine Ecology Progress Series. 331, 291-304.
de Juan, S., Demestre, M. & Thrush, S. (2009) Defining ecological indicators of trawling disturbance when everywhere that can be fished is fished: A Mediterranean case study. Marine Policy. 33 (3), 472-478.
Frid, C. J., Harwood, K., Hall, S. & Hall, J. (2000) Long-term changes in the benthic communities on North Sea fishing grounds. ICES Journal of Marine Science: Journal Du Conseil. 57 (5), 1303-1309.

37
Garcia, E. G., Ragnarsson, S. Á & Eiríksson, H. (2006) Effects of scallop dredging on macrobenthic communities in west Iceland. ICES Journal of Marine Science: Journal Du Conseil. 63 (3), 434-443.
Garrod, D. (1965) The West Greenland Cod Fishery. Lowestoft, United Kingdom, MAFF Fisheries Laboratory. Report number: 7.
Hamilton, L. C., Brown, B. C. & Rasmussen, R. O. (2003) West Greenland's cod-to-shrimp transition: local dimensions of climatic change. Arctic. 56 (3), 271-282.
Hill, A., Veale, L., Pennington, D., Whyte, S., Brand, A. & Hartnoll, R. (1999) Changes in Irish Sea benthos: possible effects of 40 years of dredging. Estuarine, Coastal and Shelf Science. 48 (6), 739-750.
Hoeksema, B. W., van der Land, J., van der Meij, Sancia ET, van Ofwegen, L. P., Reijnen, B. T., van Soest, R. W. & de Voogd, N. J. (2011) Unforeseen importance of historical collections as baselines to determine biotic change of coral reefs: the Saba Bank case. Marine Ecology. 32 (2), 135-141.
Jenkins, S., Beukers-Stewart, B. & Brand, A. (2001) Impact of scallop dredging on benthic megafauna: a comparison of damage levels in captured and non-captured organisms. Marine Ecology Progress Series. 215, 297-301.
Jennings, S., Pinnegar, J. K., Polunin, N. V. & Warr, K. J. (2001) Impacts of trawling disturbance on the trophic structure of benthic invertebrate communities. Marine Ecology Progress Series. 213, 127-142.
Jones, D. O., Bett, B. J. & Tyler, P. A. (2007) Depth-related changes in the arctic epibenthic megafaunal assemblages of Kangerdlugssuaq, East Greenland. Marine Biology Research. 3 (4), 191-204.
Kaiser, M. J., Collie, J. S., Hall, S. J., Jennings, S. & Poiner, I. R. (2002) Modification of marine habitats by trawling activities: prognosis and solutions. Fish and Fisheries. 3 (2), 114-136.
Kaiser, M., Edwards, D., Armstrong, P., Radford, K., Lough, N., Flatt, R. & Jones, H. (1998) Changes in megafaunal benthic communities in different habitats after trawling disturbance. ICES Journal of Marine Science: Journal Du Conseil. 55 (3), 353-361.
Kanneworff, P. (1979) Density of shrimp (Pandalus borealis) in Greenland waters observed by means of photography. Rapports Et Proces-Verbaux Des Réunions. Conseil International Pour L'Éxploration De La Mer. 175, 134-138.
Kanneworff, P. (1978) Estimated density of shrimp, Pandalus borealis, in Greenland waters and calculation of biomass on the offshore grounds based on bottom photography. Sel.Pap.ICNAF,(4). , 61-65.
Kenchington, E. L., Kenchington, T. J., Henry, L., Fuller, S. & Gonzalez, P. (2007) Multi-decadal changes in the megabenthos of the Bay of Fundy: the effects of fishing. Journal of Sea Research. 58 (3), 220-240.
Krieger, K. J. & Wing, B. L. (2002) Megafauna associations with deepwater corals (Primnoa spp.) in the Gulf of Alaska. Hydrobiologia. 471 (1-3), 83-90.

38
Lotze, H. K. & Worm, B. (2009) Historical baselines for large marine animals. Trends in Ecology & Evolution. 24 (5), 254-262.
Mladenoff, D. J., Dahir, S. E., Nordheim, E. V., Schulte, L. A. & Guntenspergen, G. G. (2002) Narrowing Historical Uncertainty: Probabilistic Classification of Ambiguously Identified TreeSpecies in Historical ForestSurvey Data. Ecosystems. 5 (6), 539-553.
Oksanen, J., Guillaume Blanchet, F., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., Simpson, G. l., Solymos, P., Henry H. Stevens, M. & Wagner, H. (2013) vegan: Community Ecology Package (R package version 2.0-8) .
Olsgard, F., Schaanning, M. T., Widdicombe, S., Kendall, M. A. & Austen, M. C. (2008) Effects of bottom trawling on ecosystem functioning. Journal of Experimental Marine Biology and Ecology. 366 (1), 123-133.
Pauly, D. (1995) Anecdotes and the shifting baseline syndrome of fisheries. Trends in Ecology & Evolution. 10 (10), 430.
Pilskaln, C. H., Churchill, J. H. & Mayer, L. M. (1998) Resuspension of sediment by bottom trawling in the Gulf of Maine and potential geochemical consequences. Conservation Biology. 12 (6), 1223-1229.
R Core Team. (2013) R: A Language and Environment for Statistical Computing (3.0.1) Vienna, Austria, R Foundation for Statistical Computing.
Rhemtulla, J. M., Mladenoff, D. J. & Clayton, M. K. (2009) Historical forest baselines reveal potential for continued carbon sequestration. Proceedings of the National Academy of Sciences of the United States of America. 106 (15), 608ci2-6087.
Roberts, C. M. (2003) Our shifting perspectives on the oceans. Oryx. 37 (02), 166-177.
Rosenberg, A. A., Bolster, W. J., Alexander, K. E., Leavenworth, W. B., Cooper, A. B. & McKenzie, M. G. (2005) The history of ocean resources: modeling cod biomass using historical records. Frontiers in Ecology and the Environment. 3 (2), 78-84.
Rumohr, H. & Kujawski, T. (2000) The impact of trawl fishery on the epifauna of the southern North Sea. ICES Journal of Marine Science: Journal Du Conseil. 57 (5), 1389-1394.
Simpson, A. W. & Watling, L. (2006) An investigation of the cumulative impacts of shrimp trawling on mud-bottom fishing grounds in the Gulf of Maine: effects on habitat and macrofaunal community structure. ICES Journal of Marine Science: Journal Du Conseil. 63 (9), 1616-1630.
Skelly, D. K., Yurewicz, K. L., Werner, E. E. & Relyea, R. A. (2003) Estimating decline and distributional change in amphibians. Conservation Biology. 17 (3), 744-751.
Strain, E., Allcock, A., Goodwin, C., Maggs, C., Picton, B. & Roberts, D. (2012) The long-term impacts of fisheries on epifaunal assemblage function and structure, in a Special Area of Conservation. Journal of Sea Research. 67 (1), 58-68.
Swetnam, T. W., Allen, C. D. & Betancourt, J. L. (1999) Applied historical ecology: using the past to manage for the future. Ecological Applications. 9 (4), 1189-1206.

39
Terry, R. C. (2010) The dead do not lie: using skeletal remains for rapid assessment of historical small-mammal community baselines. Proceedings.Biological Sciences / the Royal Society. 277 (1685), 1193-1201.
Tillin, H., Hiddink, J., Jennings, S. & Kaiser, M. (2006) Chronic bottom trawling alters the functional composition of benthic invertebrate communities on a sea-basin scale. Marine Ecology Progress Series. 318, 31-45.
Ugland, K. I., Gray, J. S. & Ellingsen, K. E. (2003) The species–accumulation curve and estimation of species richness. Journal of Animal Ecology. 72 (5), 888-897.
Valdemarsen, J. (2004) A New Ground Grear for bottom-trawls, incorporating spreading features. Norway, Havforskningsinstituttet, Institue of Marine Research. Report number: 4.
Vellend, M., Brown, C. D., Kharouba, H. M., McCune, J. L. & Myers-Smith, I. H. (2013) Historical ecology: using unconventional data sources to test for effects of global environmental change. American Journal of Botany. 100 (7), 1294-1305.
Wickham, H. (2011) The Split-Apply-Combine Strategy for Data Analysis. Journal of Statistical Software. 40 (1), 1-29.
Yesson, C.(2014) Personal Communication. How the fishing data was prepared for analysis Chemshirova, I.

40
Appendix 1.1
Table 1. Showing the classification system used in this study and the sum of individuals, sensitive taxa are highlighted.
Taxa Total
Encrusting Bryozoa 783
Ascidians 539
Erect Bryozoa 300
Decapoda 298
Massive Sponges 192
Soft Bryozoa 191
Encrusting Sponges 188
Brittle stars 153
Stylasterina 98
Arborescent Sponges 92
Serpulidae 91
Unknown 83
Anemones 71
Sabellidae 53
Soft Corals 36
Perciformes 32
Hydroids 13
Pleuronectiformes 11
Terebratulida 10
Zoanthids 9
Echinasteridae 9
Goniasteridae 8
Sea Cucumbers 6
Asteriidae 5
Starfish Other 5
Gastropods 5
Bivalves 4
Crinoids 3
Astropectinidae 2
Sea Urchins 2
Eunicidae 2
Rajiformes 2
Pterasteridae 1
Polynoidae 1
Isopoda 1
Sea Spiders 1
Scaphopods 1
Scorpaeniformes 1

41
Appendix 1.2
Table 2. Classification system all at the same taxonomic level.
Class Total
Gymnolaemata 1194
Ascidiacea 557
Demospongiae 520
Malacostraca 312
Stenolaemata 225
Polychaeta 171
Ophiuroidea 153
Hydrozoa 133
Anthozoa 119
Unknown 89
Actinopterygii 48
Asteroidea 36
Rhynchonellata 10
Holothuroidea 6
Gastropoda 5
Bivalvia 4
Crinoidea 3
Echninoidea 2
Pycnogonida 2
Elasmobranchii 2
Scahopoda 1

1
Appendix 1.3: Greenland Benthic ID Guide

2
DSC_0041
DSC_0043
DSC_0043
DSC_0042
DSC_0201
Cnidaria> Anthozoa> Octocorals> Soft Corals
DSC_0040 DSC_0041 DSC_0041
DSC_0224
DSC_0503
DSC_0628
DSC_0628
DSC_0650
DSC_0799
DSC_0802
DSC_0948
DSC_1036
DSC_1255-gersemia?

3
DSC_0280
Cnidaria> Anthozoa> Hexacorals> Actiniaria (Sea Anemones)
DSC_0501
DSC_1015
DSC_0388
DSC_0390
DSC_0043
DSC_0076
DSC_0402
DSC_0472
DSC_0712 DSC_0710
DSC_0820
DSC_0665
DSC_0873
DSC_0876 DSC_1081
DSC_1119
DSC_0826 DSC_1192

4
Cnidaria> Anthozoa> Hexacorals> Zoantharia
DSC_0200 DSC_0321
DSC_0041 DSC_0082 DSC_0082 DSC_0233 DSC_0257 DSC_0302 DSC_0804
DSC_1193
DSC_0710
DSC_0124-11

5
Cnidaria> Hydrozoa> Hydroids
DSC_0233
DSC_0234
DSC_0238
DSC_0238
DSC_0240
DSC_0253
DSC_0254
DSC_0256
DSC_0256
DSC_0258
DSC_0289
DSC_0511
DSC_0639
DSC_0639

6
Cnidaria> Hydrozoa> Hydroidolina> Anthoathecata> Stylasteridae
DSC_0223
DSC_0244 DSC_0251 DSC_0077
DSC_0077
DSC_0082
DSC_0251
DSC_0252
DSC_0255
DSC_0256

7
Echinodermata> Asteroidea
DSC_0820-Echinasteridae
DSC_0043-Solasteridae
DSC_0078-Solasteridae
DSC_0224-Goniasteridae
DSC_0243-ilac-Asteriidae? °
DSC_0255-Pterasteridae?
DSC_0300-Echinasteridae
DSC_0385-Goniasteridae
DSC_0500-Echinasteridae
DSC_0593-Echniasteridae
DSC_0606-Echniasteridae
DSC_0620-Goniasteridae
DSC_0628-Echinasteridae
DSC_0708-Solasteridae
DSC_0708-Astropectinidae?
DSC_0921-Solasteridae?
DSC_1010-Ptreasteridae
DSC_1013-Pterasteridae
DSC_1016-Pterasteridae DSC_1017-purple
DSC_1230-Goniasteridae?
DSC_1233-Goniasteridae

8
DSC_0233
DSC_0108
DSC_0236
DSC_0236
Echinodermata> Ophiuroidea
DSC_0233
DSC_0233
DSC_0237
DSC_0251
DSC_0251
DSC_0251
DSC_0252
DSC_0273
DSC_0274
DSC_0275
DSC_0277
DSC_0276 DSC_0284
DSC_0285
DSC_0285
DSC_0046

9
DSC_0256
DSC_0893 DSC_1000
Echinodermata> Echinoidea
DSC_0242 DSC_0251 DSC_0275
DSC_0284

10
Echinodermata> Holouthuroidea
DSC_0385
DSC_0385
DSC_0501
DSC_507
DSC_0507
DSC_0511 DSC_0542
DSC_0544
DSC_0550 DSC_0623
DSC_0640 DSC_0644
DSC_0684
DSC_0704 DSC_0705 DSC_0725
11
DSC_0254
Echinodermata> Crinoidea
DSC_0251
DSC_0252
DSC_0277
DSC_0280
DSC_0510 DSC_0541
DSC_0542
DSC_0544
DSC_0586 DSC_0589
DSC_0590
DSC_0620
12
Porifera> Encrusting
DSC_0042-Axinellidae
DSC_0042-Axinellidae DSC_0042-pink
DSC_0042-Pachastrellidae
DSC_0046-Acarnidae
DSC_0242-Crellidae DSC_0255– Ancorinidae
DSC_0274
DSC_0046-Micricionidae
DSC_0059-11-Hymedesmiidae

13
Porifera> Massive
DSC_0107– Polymastiidae DSC_0267-Tethyidae DSC_0258-Polymastiidae
DSC_0041-Sycettidae DSC_0252-Grantiidae
DSC_0239-Polymastiidae
DSC_0239-Suberitidae
DSC_0390
DSC_0501
DSC_00628—Esperiopsidae
DSC_0644
DSC_0256-Aninellidae
DSC_0060-11– Grantiidae
14
Porifera> Arborescent
DSC_0239-Suberitidae
DSC_0501
DSC_0639-Microcionidae
DSC_0639
DSC_0009-11-(should be in massive)

15
Annelida> Polychaetes
DSC_0077-Sabellidae
DSC_0644_Serpulidae
DSC_1123
DSC_0125-11-Polynoide
DSC_0059-11-Serpulidae

16
Arthropoda> Malacostraca> Eumalacostraca> Decapoda
DSC_0413-Majidae
DSC_0430-Majidae
DSC_0743-Paguroidea
DSC_1131
DSC_0862-Paguroidea

17
Arthropoda> Malacostraca> Eumalacostraca> Decapoda
DSC_0413
DSC_0040-shrimp?
DSC_0038-shrimp?

18
Arthropoda> Pycnogonida> Pantopoda
DSC_0221
DSC_0703
DSC_0804
DSC_0919
DSC_1071
DSC_1084
DSC_1092 DSC_1131
DSC_0125-11
19
Mollusca> Gastropoda
DSC_0301 DSC_0501
DSC_0504
DSC_0511 DSC_0626
DSC_0667
DSC_0684
DSC_0733
DSC_0733
DSC_0799 DSC_0838
DSC_1071
DSC_1191

20
Mollusca> Chitons
DSC_0772
DSC_0618
DSC_0125-11
DSC_0130-11

21
Mollusca> Bivalves
DSC_0076
DSC_0077
DSC_0107
DSC_0107
DSC_0107
DSC_0218
DSC_0227
DSC_0507
DSC_0507
DSC_0510
DSC_0511 DSC_0592
DSC_0642
DSC_0642
DSC_0643

22
Mollusca> Scaphopoda
Mollusca> Cephalopods> Sepiida
DSC_0076
DSC_0078
DSC_0584 DSC_0584
DSC_0746
DSC_0776

23
Brachiopoda> Articulata> Terebratulida
DSC_0283
DSC_0387
DSC_0390
DSC_0390 DSC_0519
DSC_0592
DSC_0628
DSC_0643-?
DSC_0995

24
Bryozoa> Erect
DSC_0255
DSC_0040
DSC_0642 DSC_0642
DSC_0860
DSC_1255
DSC_1030
DSC_0862

25
Bryozoa> Encrusting
DSC_0743
DSC_0107
DSC_0669
DSC_0937
DSC_0937

26
Bryozoa> Soft
Chordata> Ascidiacea
DSC_0043
DSC_0043
DSC_0041 DSC_0822
DSC_0822
DSC_0822
DSC_0511
DSC_1214
DSC_0043-stalked-Polyclinidae
DSC_0038-Molgulidae
DSC_0078-Ascidiidae
DSC_1238-grey-Polycitoridae
DSC_0239-Styelidae
DSC_0125-11-Didemnidae

27
Chordata> Chondrichthyes> Rajiformes

28
Chordata> Osteichthyes> Scorpaeniformes

29
Chordata> Osteichthyes> Perciformes
DSC_0205
DSC_0418
DSC_0882
DSC_0892
DSC_0964

30
Chordata> Osteichthyes> Pleuronectiformes
DSC_0273
DSC_1073
DSC_1229

31
Dr Kirsty Kemp
Poppy Simon
Irina Chemshirova

42
Appendix 1.4
Table 3. Abundance of taxa in the two time periods
Taxa 1970s 1980s
Encrusting Bryozoa 379 404
Ascidians 330 209
Decapoda 152 146
Erect Bryozoa 116 184
Massive Sponges 95 97
Encrusting Sponges 75 113
Soft Bryozoa 74 117
Brittle stars 52 101
Arborescent Sponges 50 42
Sabellidae 43 10
Serpulidae 37 54
Anemones 34 37
Stylasterina 33 65
Unknown 30 53
Perciformes 13 19
Soft Coral 12 24
Terebratulida 7 3
Echinasteridae 5 4
Hydroids 5 8
Pleuronectiformes 5 6
Crinoids 3 0
Goniasteridae 3 5
Seacucumbers 3 3
Seaurchins 2 0
Starfish Other 2 3
Zoanthids 2 7
Astropectinidae 1 1
Bivalves 1 3
Gastropods 1 4
Isopoda 1 0
Polynoidae 1 0
Pterasteridae 1 0
Rajiformes 1 1
Amphipoda 0 0
Aphroditidae 0 0
Asteriidae 0 5
Chitons 0 0
Eunicidae 0 2
Octopoda 0 0

43
Scaphopods 0 1
Scorpaeniformes 0 1
Seaspiders 0 1
Appendix 1.5
Figure 1. Species accumulation curves for both time periods, blue is for the 1970s, green for the 1980s.
5 10 15
05
10
15
20
25
30
35
Sites
Ta
xa

44
Appendix 1.6
Table 4. Stations from both time periods paired based on their location, including the group number based on their location pairings
Stations 1970s
Stations 1980s
Station groups
5609 6713 1
5453 6715 2
5606 6716 3
5605 6717 4
5601 6720 5
5739 6721 6
5447 6722 7
5867 6724 8
5612 6727 9
5613 6728 10
5743 6729 11
5738 6730 12
5870 6731 13
5869 6732 14
5871 6733 15
5859 6736 16
5444 6737 17
5861 6738 18
5737 6749 19

45
Appendix 1.7
Table 5. The results of the Wilcoxon paired rank test between the 1970s and 1980s diversity metrics with the Class dataset
Diversity Index Result
Taxon Richness V=52.5, p=0.5
Number of Individuals V=85.5, p=0.7
Pileou’s Evenness V=105, p=0.7
Shannon Index V=95, p=1

46
Appendix 1.8
Figure 2. nMDS plot for the 1970s with taxonomic identification to Class level. Substrata 0.001 *** Stress = 0.12
-1.5 -1.0 -0.5 0.0 0.5 1.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1970s
NMDS1
NM
DS
2
5609
5453
5606
5605
5601
5739
5447
5867
5612
5613
5743
57385870
5869
5871
5859
5444
58615737
Substrata
soft
hard
-1.5 -1.0 -0.5 0.0 0.5 1.0
-1.0
-0.5
0.0
0.5
1.0
Species 1970s
NMDS1
unspecified
anthozoahydrozoa
asteroidea
ophiuroidea
echinoidea
holothuroidea
crinoidea
demospongiaepolychaeta
malacostraca
gastropoda
bivalvia
rhynchonellata
gymnolaematastenolaemata
ascidiacea
elasmobranchii
actinopterygii
NMDS1
NM
DS
2
X1
Y1
Depth1
Fishing.Log
Substrata1
Bioturbation1

47
Figure 3. nMDS plot for the 1980s with taxonomic identification to Class level. Substrata 0.003 *** Stress = 0.15
-1.0 -0.5 0.0 0.5 1.0 1.5
-1.0
-0.5
0.0
0.5
1.0
1980s
NMDS1
NM
DS
2
6713
6715
6716
6717
6720
6721
6722
6724
6727
6728
6729
6730
6731
67326733 6736
6737
6738
6749
Substrata
soft
hard
-1.0 -0.5 0.0 0.5 1.0 1.5
-0.5
0.0
0.5
1.0
1.5
Species 1980s
NMDS1
unspecified
anthozoa
hydrozoaasteroidea
ophiuroidea
holothuroidea
demospongiae
polychaeta
malacostraca
pycnogonida
gastropoda
bivalvia
scaphopoda
rhynchonellata
gymnolaemata
stenolaemata
ascidiacea
elasmobranchii
actinopterygii
NMDS1
NM
DS
2
X2
Y2
Depth2
Fishing.LogSubstrata2
Bioturbation2
48
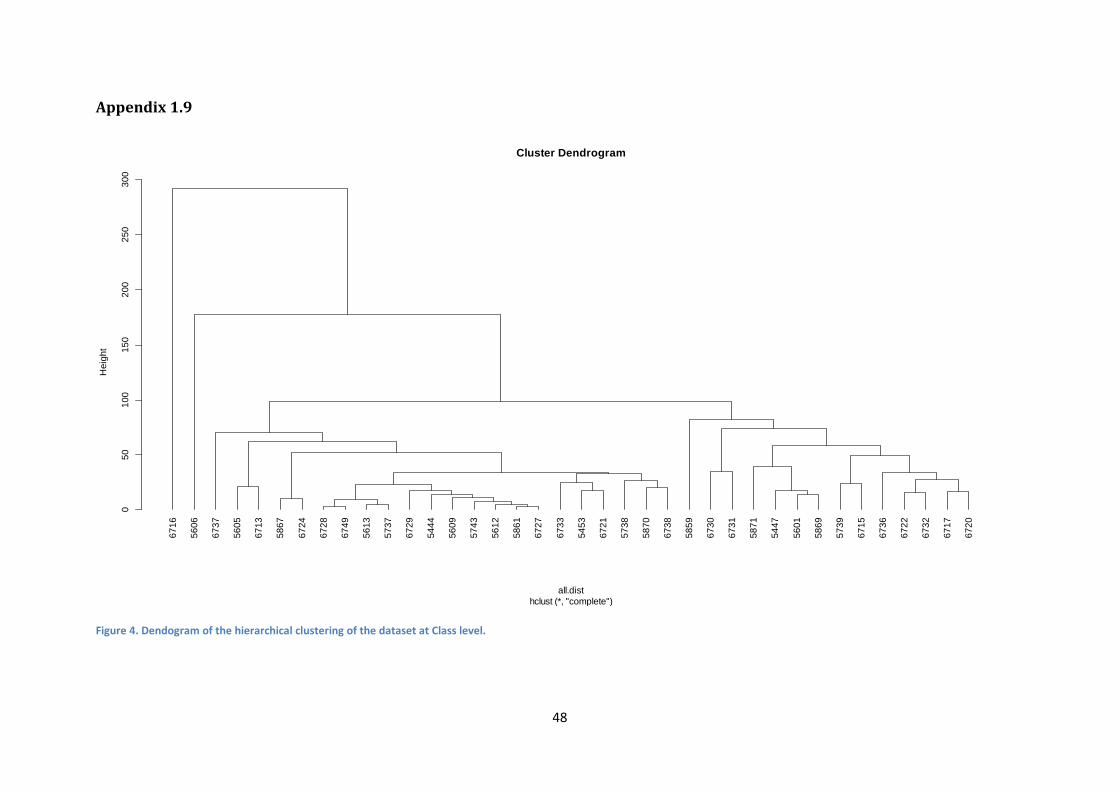
Appendix 1.9
Figure 4. Dendogram of the hierarchical clustering of the dataset at Class level.
67
16
56
06
67
37
56
05
67
13
58
67
67
24
67
28
67
49
56
13
57
37
67
29
54
44
56
09
57
43
56
12
58
61
67
27
67
33
54
53
67
21
57
38
58
70
67
38
58
59
67
30
67
31
58
71
54
47
56
01
58
69
57
39
67
15
67
36
67
22
67
32
67
17
67
20
05
01
00
15
02
00
25
03
00
Cluster Dendrogram
hclust (*, "complete")
all.dist
He
igh
t
49
Figure 5. Showing the relationship between trawling and taxon (Class) richness. Trawling has no effect on Class richness for the 1970s period (t=0.18, d.f.=17, p=0.35, , R
2=0.06). There is no relationship between trawling impact and Class
richness for the 1980s (t=-0.48, d.f.=17, p=0.64, , R2=0.02).
0 2 4 6 8 10 12
24
68
10
12
loge (Trawling Impact 1975-1984)(mins)
Ta
xo
n R
ich
ne
ss
hard
soft
1970s
1980s

50
Appendix 2.1
Figure 6. Showing the relationship between β diversity and trawling impact. Increasing trawling impact had no effect on the β diversity observed at Class level (t=0.55, d.f.=17, p=0.59, R
2=0.017).
0 2 4 6 8 10
0.1
0.2
0.3
0.4
0.5
0.6
loge (Trawling Impact 1982-1984)(mins)
Be
ta D
ive
rsity

51
Appendix 2.2
Figure 7. Species accumulation curve for the 1984 dataset, further sampling will most likely not uncover new species.
0 5 10 15 20 25 30 35
01
02
03
04
0
Sites
Sp
ecie
s

52
Appendix 2.3
Figure 8. nMDS plot of the 1984 dataset. Substrata was the only environmental vector which explained the variation seen (p<0.01 **, Stress = 0.19)
-1.5 -1.0 -0.5 0.0 0.5 1.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
Stations
NMDS1
NM
DS
2
Substrata
soft
hard
-1.0 -0.5 0.0 0.5 1.0
-1.0
-0.5
0.0
0.5
1.0
Species
NMDS1
unspecifiedsoftcoral
anemones
zoanthids
hydroids
stylasterina
asteriidae
pterasteridae
echinasteridae
solasteridae
goniasteridae
astropectinidae
starfishother
brittlestars
seaurchins
seacucumbers
crinoids
encrustingspmassivesp
arborescentspsabellidae
eunicidae
serpulidaem
decapoda
seaspidersgastropods
bivalves
scaphopods
sepioida
terebratulidaerectbr
encrustingbrsoftbr
ascidians
rajiformes
scorpaeniformes
perciformes
pleuronectiformes
NMDS1N
MD
S2
Substrata
Depth2
X
Y
Fishing.Log

53
Appendix 2.4
Figure 9. Linear regessions showing the relationship between trawling intensity and the following diversity indices: Taxon richness (t=-0.62, d.f.=34, p=0.54, R
2=0.011), Number of Individuals (t=-0.98, d.f.=34, p=0.33, R
2=0.027), Pielou’s
Evenness (t=0.84, d.f.=34, p=0.41, R2=0.020), Shannon Index (t=0.29, d.f.=34, p=0.77, R
2=0.002).
0 2 4 6 8 10 12
510
15
20
loge (Trawling Impact)(mins)
Specie
s R
ichness
0 2 4 6 8 10 12
34
56
loge (Trawling Impact)(mins)
log
e(N
o.
of
Indiv
iduals
)
0 2 4 6 8 10 12
0.4
0.5
0.6
0.7
0.8
0.9
loge (Trawling Impact)(mins)
Pie
lou's
Evenness
0 2 4 6 8 10 12
0.5
1.0
1.5
2.0
2.5
loge (Trawling Impact)(mins)
Shannon I
ndex

54
Appendix 2.5
Figure 10. nMDS plot of both time periods showing a clear separation of substrata (Stress = 0.16)
-1.0 -0.5 0.0 0.5 1.0 1.5
-1.0
-0.5
0.0
0.5
Overall
NMDS1
NM
DS
2
Substrata and Year
hard
soft
1970s
1980s


















