
Effects of Ammonium, L-Glutamate, L-Glutamine Nitrogen ...jb.asm.org/content/120/3/1116.full.pdf ·...
8
JOURNAL OF BACTERIOLOGY, Dec. 1974, p. 1116-1123 Copyright ( 1974 American Society for Microbiology Vol. 120, No. 3 Printed in U.S.A. Effects of Ammonium, L-Glutamate, and L-Glutamine on Nitrogen Catabolism in Aspergillus nidulans M. J. HYNES Department of Genetics, La Trobe University, Bundoora, Victoria 3083, Australia Received for publication 23 September 1974 During growth of Aspergillus nidulans in medium containing ammonium the specific activities of most enzymes involved in catabolism of nitrogen sources are low (ammonium repression). The gdhAIO lesion, which results in loss of' nicotinamide adenine dinucleotide phosphate-linked glutamate dehydrogenase activity, has been shown to lead to partial relief of ammonium repression of three amidase enzymes as well as histidase. The areA102 lesion led to altered levels of' these enzymes but did not greatly affect ammonium repression. The double mutant areA1O2,gdhA1O was almost completely insensitive to ammonium repression of' two of the amidase enzymes and histidase. This suggests that an interaction between the areA and gdhA genes in determining responses to ammonium occurs. Growth of mycelium in medium containing L-glutamate has been found to result in lowered levels of' all four enzymes, and this occurs in strains insensitive to ammonium repression. Very strong repression in all strains occurred during growth in medium containing L-glutamine. Relief' of' these repressive eff'ects of glutamate and glutamine was blocked by cycloheximide. Glutamate and glutamine had similar effects on the production of extracellular protease activity, and growth on glutamine led to low levels of urate oxidase. In contrast to the above enzymes, nitrate reductase was insensitive to the effects of' glutamine and glutamate, even though this enzyme is very sensitive to ammonium repression. Although other possibilities exist, it is suggested that there may be mechanisms of general control of nitrogen-catabolic enzymes other than ammonium repression. Enzymes involved in the catabolism of nitro- gen sources have been commonly found to be present in low amounts during growth of micro- organisms on ammonium salts (1, 6, 10, 12, 17, 20, 21). This phenomenon has been called by many names but will be referred to as ammo- nium repression in this paper, without making any specific inferences as to mechanism. Muta- tions leading to loss of' nicotinamide adenine dinucleotide phosphate (NADP)-dependent glutamate dehydrogenase (GDH) (EC 1.4.1.4) have been found to result in loss of ammonium repression in yeast (10, 11, 24) and Aspergillus nidulans (3, 16). It has been proposed that the NADP-GDH enzyme molecule has a regulatory function in controlling enzyme synthesis (10, 20). However, van de Poll has reported that, in a strain of' Saccharomyces carlsbergensis lacking NADP-GDH, allantoinase is repressed by gluta- mine and glutamate plus ammonium but not by ammonium alone (24). This has led to the suggestion that the NADP-GDH catalytic ac- tivity is necessary for ammonium repression. So far no mutants affecting the proposed regula- tory f'unction but not the catalytic activity of NADP-GDH have been described. In A. nidulans, mutations in a gene, areA (called amdT in previous publications; see ref. 2), lead to pleiotropic effects on nitrogen source utilization (2, 14). A variety of' phenotypic effects are observed in various areA mutants, including inability to grow on many nitrogen sources, the utilization of' which are subject to ammonium repression. Others show altered sen- sitivities to ammonium repression of' some of' the enzymes of' nitrogen catabolism. The areA 102 lesion results in increased growth on many nitrogen sources and leads to increased levels of some enzymes of' nitrogen catabolism without relieving ammonium repression (2, 14, 15; unpublished data). Arst and Cove (2) have concluded that the product of' the areA gene is required for the synthesis of' nitrogen-catabolic enzymes and that ammonium repression in- volves an interf'erence with the action of this positive regulatory factor. In addition, they suggest that carbon catabolite repression is mediated via a separate control system (2). 1116 on March 5, 2019 by guest http://jb.asm.org/ Downloaded from
Transcript of Effects of Ammonium, L-Glutamate, L-Glutamine Nitrogen ...jb.asm.org/content/120/3/1116.full.pdf ·...

JOURNAL OF BACTERIOLOGY, Dec. 1974, p. 1116-1123Copyright ( 1974 American Society for Microbiology
Vol. 120, No. 3Printed in U.S.A.
Effects of Ammonium, L-Glutamate, and L-Glutamine onNitrogen Catabolism in Aspergillus nidulans
M. J. HYNES
Department of Genetics, La Trobe University, Bundoora, Victoria 3083, Australia
Received for publication 23 September 1974
During growth of Aspergillus nidulans in medium containing ammonium thespecific activities of most enzymes involved in catabolism of nitrogen sources arelow (ammonium repression). The gdhAIO lesion, which results in loss of'nicotinamide adenine dinucleotide phosphate-linked glutamate dehydrogenaseactivity, has been shown to lead to partial relief of ammonium repression of threeamidase enzymes as well as histidase. The areA102 lesion led to altered levels of'these enzymes but did not greatly affect ammonium repression. The doublemutant areA1O2,gdhA1O was almost completely insensitive to ammoniumrepression of' two of the amidase enzymes and histidase. This suggests that aninteraction between the areA and gdhA genes in determining responses toammonium occurs. Growth of mycelium in medium containing L-glutamate hasbeen found to result in lowered levels of' all four enzymes, and this occurs instrains insensitive to ammonium repression. Very strong repression in all strainsoccurred during growth in medium containing L-glutamine. Relief' of' theserepressive eff'ects of glutamate and glutamine was blocked by cycloheximide.Glutamate and glutamine had similar effects on the production of extracellularprotease activity, and growth on glutamine led to low levels of urate oxidase. Incontrast to the above enzymes, nitrate reductase was insensitive to the effects of'glutamine and glutamate, even though this enzyme is very sensitive toammonium repression. Although other possibilities exist, it is suggested thatthere may be mechanisms of general control of nitrogen-catabolic enzymes otherthan ammonium repression.
Enzymes involved in the catabolism of nitro-gen sources have been commonly found to bepresent in low amounts during growth of micro-organisms on ammonium salts (1, 6, 10, 12, 17,20, 21). This phenomenon has been called bymany names but will be referred to as ammo-nium repression in this paper, without makingany specific inferences as to mechanism. Muta-tions leading to loss of' nicotinamide adeninedinucleotide phosphate (NADP)-dependentglutamate dehydrogenase (GDH) (EC 1.4.1.4)have been found to result in loss of ammoniumrepression in yeast (10, 11, 24) and Aspergillusnidulans (3, 16). It has been proposed that theNADP-GDH enzyme molecule has a regulatoryfunction in controlling enzyme synthesis (10,20). However, van de Poll has reported that, in astrain of' Saccharomyces carlsbergensis lackingNADP-GDH, allantoinase is repressed by gluta-mine and glutamate plus ammonium but not byammonium alone (24). This has led to thesuggestion that the NADP-GDH catalytic ac-tivity is necessary for ammonium repression. Sofar no mutants affecting the proposed regula-
tory f'unction but not the catalytic activity ofNADP-GDH have been described.
In A. nidulans, mutations in a gene, areA(called amdT in previous publications; see ref.2), lead to pleiotropic effects on nitrogen sourceutilization (2, 14). A variety of' phenotypiceffects are observed in various areA mutants,including inability to grow on many nitrogensources, the utilization of' which are subject toammonium repression. Others show altered sen-sitivities to ammonium repression of' some of'the enzymes of' nitrogen catabolism. TheareA 102 lesion results in increased growth onmany nitrogen sources and leads to increasedlevels of some enzymes of' nitrogen catabolismwithout relieving ammonium repression (2, 14,15; unpublished data). Arst and Cove (2) haveconcluded that the product of' the areA gene isrequired for the synthesis of' nitrogen-catabolicenzymes and that ammonium repression in-volves an interf'erence with the action of thispositive regulatory factor. In addition, theysuggest that carbon catabolite repression ismediated via a separate control system (2).
1116
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

NITROGEN CATABOLISM IN A. NIDULANS
Results from this laboratory strongly supporttheir conclusions.
This paper reports studies on ammoniumrepression of a number of nitrogen-catabolicenzymes in strains of A. nidulans with lesions inthe gdhA gene (the proposed structural gene forNADP-GDH) and the areA gene. The effects ofgrowth on L-glutamate and L-glutamine onenzyme levels have also been investigated.These studies raise the possibility that theremay be control mechanisms, in addition toammonium repression, affecting the levels ofenzymes of nitrogen catabolism.
MATERIALS AND METHODSStrains. Details of the strains used are described in
Table 1. It should be noted that more limited experi-ments with other strains, of similar gdhA and areAgenotypes, have given similar results to those reportedhere.Media and growth conditions. The standard min-
imal medium used was that of Cove (8), where 1%(wt/vol) glucose is the sole carbon source and nonitrogen source is present. Nitrogen sources to thedesired concentration were added to this medium.Glutamate was added as sodium L-glutamate andammonium was added as ammonium tartrate. Myce-lium was grown at 30 C in a Gallenkamp orbitalincubator as described previously (13, 14). Harvestedmycelium was stored frozen at -15 C and assayedwithin 5 days.
Preparation of crude extracts. All crude extractswere made in 100 mM orthophosphate buffer, pH 7.2,by methods described previously (13). Protein concen-trations of crude extracts were determined by themethod of Lowry et al. (18).
Plate tests for extracellular protease. These werecarried out by the method of Cohen (6) using 1% driedmilk powder.Amidase enzyme assays. Formamidase and ace-
tamidase (EC 3.5.1.4) activities were determined bymethods described before (12, 13). Both of theseenzymes are subject to strong repression by ammo-
nium and derepress on transfer to medium lacking anitrogen source (12). The acetamidase is inducible byamides (12, 13). The formamidase is not apparentlyinducible (as originally thought; 12, 13). Formami-dase levels are no higher in medium containingformamide than in medium lacking a nitrogen source(unpublished data). A third amidase enzyme hasrecently been partially characterized (unpublisheddata). This enzyme hydrolyzes benzamide as well asother amides, but its precise substrate specificity hasnot been determined and so no Enzyme Commissionnumber is available. Genetic and thermal inactivationexperiments show that it is distinct from the otheramidase enzymes (unpublished data). For the pur-poses of this paper this enzyme will be called thebenzamidase. As shown here, this enzyme is subjectto ammonium repression and derepresses on transferto medium lacking a nitrogen source. The areA102lesion leads to much higher benzamidase activitiesthan those present in wild-type (areA ) strains (un-published data). Benzamidase assays were performedusing the method of assay for the other amidaseenzymes, with 10 mM benzamide (recrystallized oncefrom hot water) in 100 mM orthophosphate buffer(pH 7.2) as the substrate solution. All amidase-specific activities are expressed as milliunits permilligram of protein, where one unit is the amount ofenzyme that catalyzes the release of 1 Mmol ofammonium per min.
Histidase enzyme assays. A method (M. Polk-inghorne and M. J. Hynes, unpublished data) basedon measuring the formation of urocanate from histi-dine by histidase (EC 4.3.1.3) was used. The reactionwas carried out at 30 C in a Unicam SP 1700spectrophotometer using 1-cm2 quartz cuvetts. Thereaction mixture contained 2.8 ml of 100 mM ortho-phosphate buffer (pH 8.0) and 0.1 ml of crude extract.The reaction was started by the addition of 0.1 ml ofL-histidine solution (3 mg per ml of water), and theincrease in absorbance at 277 nm followed. Specificactivities are expressed as milliunits per milligram ofprotein, where one unit is the amount of enzyme thatcatalyzes the conversion of 1 umol of histidine tourocanate per min. It has been shown that histidase issubject to ammonium repression and does not require
TABLE 1. Details of strains used in this study
Strain Genotypea Origin References
Wild type biAl Glasgow wild typegdhAlO biA; gdhA10 H. Arst 3areA102 biAI; areA102; niiA4 Formerly strain amdT102 2, 13-15areA102,gdhA10 yAl; areA102,gdhA10; riboB2 Isolate from cross in this
laboratorymauA2 biAl; mauA2 D. Cove 1,4mauA2; gdhAlO biAI; gdhA10; mauA2 Isolate from cross in this
laboratorynirA C yAI,adE20; nirA c, riboB2 J. Pateman 19a biAl, Biotin auxotrophy; niiA4, lack of nitrite reductase; yAl, yellow conidia; riboB2, riboflavin
auxotrophy; mauA2, lack of mono-amine oxidase; adE20, adenine auxotrophy. These genetic markers have nosignificance for the experiments reported here. The significance of the gdhA10, areA102 and nirA,c lesions isdescribed in the text.
VOL. 120, 1974 1117
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

1J. BACTERIOL.
external induction by histidine (Polkinghorne andHynes, unpublished data; this paper).
Urate oxidase enzyme assays. The assay methodfor urate oxidase (EC 1.7.3.3) was based on that ofScazzocchio and Darlington (23). The reaction mix-ture contained 2.8 ml of' 100 mM orthophosphatebuffer (pH 7.2), 0.1 ml of crude extract, and 0.1 ml ofuric acid solution (0.336 mg per ml of' water). Thedecline in absorbance at 292 nm at 30 C was f'ollowedusing a Unicam SP 1700 spectrophotometer with1-cm2 quartz cuvettes. This enzyme has been shownto require induction by uric acid and to be subject toammonium repression (23). Specific activities areexpressed as milliunits per milligram of protein,where one unit is the amount of' enzyme that catalyzesthe oxidation of 1 Mmol of uric acid per min.
Nitrate reductase enzyme assays. The method ofCove (8), with the modif'ied control of Lewis andFincham (17), was used to assay nitrate reductase(EC 1.6.6.3). This enzyme has an absolute require-ment for induction by nitrate and is repressed byammonium (8). Specif'ic activities are expressed asmilliunits per milligram of protein, where one unit isthe amount of enzyme that catalyzes the reduction of1 umol of' nitrate to nitrite per min.
Extracellular protease assays. Cohen (6, 7) hasshown that A. nidulans produces protease enzymeactivity in the external medium when mycelium istransf'erred to medium lacking a nitrogen source andthat this activity does not appear when mycelium isgrowing in medium containing ammonium. In thisstudy protease activity has been measured by f'ollow-ing release of trichloroacetic acid-soluble coloredcompounds from azocasein (5). Mycelium was har-vested by the normal method, and the externalmedium was collected and used for extracellularprotease assays. The mycelium was dried and the dryweight was determined. One milliliter of azocaseinsolution (4 to 5 mg per ml of 100 mM orthophosphatebuffer, pH 7.2) was added to plastic centrit'uge tubes,and the reaction was initiated by adding 1 ml of' themedium to be assayed. Incubation was carried out at37 C in a shaking water bath. Af'ter a suitable time thereaction was stopped by addition of' 5 ml of' 59%(wt/vol) trichloroacetic acid. The insoluble materialwas spun down in a bench centrifuge, and theabsorbance of the supernatant was read at 440 nm.The assay was linear within the times of incubationused. A control where the trichloroacetic acid wasadded before addition of' the medium for assay wasused in each case. Results were calculated as absorb-ance units per minute per milliliter of' medium permilligram (dry weight) of' mycelium and are expressedas relative arbitrary units in Table 6.
RESULTSPreliminary observations on the eff'ects of'
glutamate on histidase levels in the areA 102,gdhAJO strain (Polkinghorne and Hynes, un-published data) led to a more detailed study of'this problem. Since strains containing the gdhAlesion grow poorly in medium lacking gluta-mate, glutamine, or a source of glutamate (3),
the initial experiments in this study involvedgrowing mycelium in medium containing gluta-mate plus ammonium and then transferring totreatment media containing various nitrogensources. It was found that, compared withwild-type and areA 102 strains, the gdhA 10 andareAl02,gdhA1O strains were at least partiallyinsensitive to ammonium repression of' form-amidase, acetamidase, benzamidase, and his-tidase. In addition. it was f'ound that mvceliumtransf'erred to medium containing glutamate (inboth the presence and absence of' ammonium)led to reduction in the levels of' these enzymescompared with the levels in mycelium trans-ferred to medium lacking a nitrogen source.Also, when mycelium was transferred to me-dium containing glutamine as the sole nitrogensource, the enzyme levels were very low in thewild-type and areA 102 strains and were atlevels consistent with repression of' enzymesynthesis and dilution during growth in thegdhAlO and areAl(2,gdhAIO strains. Figure1 shows the results of' a more detailed experi-ment on the areA 102 and areAl02,gdhAIOstrains. It is seen that growth of' areA102 onglutamate plus ammonium led to very lowlevels of' acetamidase and histidase, showingthat this strain is sensitive to ammonium re-pression. These levels were maintained inmycelium transferred to medium containingglutamine. Transfer to medium containingglutamate alone led to enzyme levels approxi-mately 50.17 of' those in mycelium transf'erredto medium lacking a nitrogen source. In con-trast to these results, the areA 102,gdhA 10strain grown on glutamate plus ammoniumproduced substantial levels of' acetamidase andhistidase, and this level was maintained ontransfer to medium containing glutamate alone.Approximately 50"'/ derepression occurred inmedium lacking a nitrogen source. Transfer tomedium with glutamine as the sole nitrogensource caused a decline in enzyme activityconsistent with repression and dilution of' en-zyme.These experiments led to the conclusion that
the gdhAIO lesion causes relief' of' ammoniumrepression but not relief' of' repression arisingfrom growth on glutamate or glutamine. Forconvenience the effects of glutamate and gluta-mine will be referred to as glutamate repressionand glutamine repression, respectively, in therest of this communication (see Discussion).
It was then possible to carry out experimentsin which mycelium was grown on medium withglutamine as the sole nitrogen source and thentransferred to various treatment media. Thisavoided the complication of' transferring myce-
1118 HYNES
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
NITROGEN CATABOLISM IN A. NIDULANS
P200- ~~~~~40-
1'00 2
2 4 6
300 are A102 0dhA10 L A 102 gdh A10ACETAMIDASE _ HISTIDASE
20 40
10
0 2 4 60
TIME (HOURS)
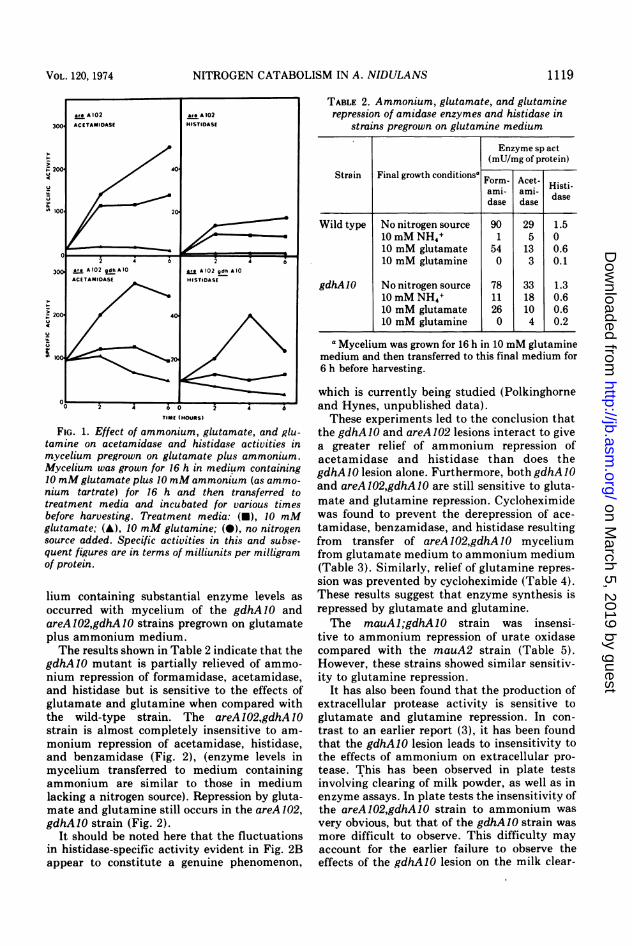
FIG. 1. Effect of ammonium, glutamate, and glu-tamine on acetamidase and histidase activities inmycelium pregrown on glutamate plus ammonium.Mycelium was grown for 16 h in medium containing10 mMglutamate plus 10 mM ammonium (as ammo-nium tartrate) for 16 h and then transferred totreatment media and incubated for various timesbefore harvesting. Treatment media: (U), 10 mMglutamate; (A), 10 mM glutamine; (0), no nitrogensource added. Specific activities in this and subse-quent figures are in terms of milliunits per milligramof protein.
lium containing substantial enzyme levels as
occurred with mycelium of the gdhA1O andareA102,gdhA10 strains pregrown on glutamateplus ammonium medium.The results shown in Table 2 indicate that the
gdhAlO mutant is partially relieved of ammo-nium repression of formamidase, acetamidase,and histidase but is sensitive to the effects ofglutamate and glutamine when compared withthe wild-type strain. The areA 102,gdhA 10
strain is almost completely insensitive to am-
monium repression of acetamidase, histidase,and benzamidase (Fig. 2), (enzyme levels inmycelium transferred to medium containingammonium are similar to those in mediumlacking a nitrogen source). Repression by gluta-mate and glutamine still occurs in the areA 102,gdhA10 strain (Fig. 2).
It should be noted here that the fluctuationsin histidase-specific activity evident in Fig. 2Bappear to constitute a genuine phenomenon,
TABLE 2. Ammonium, glutamate, and glutaminerepression of amidase enzymes and histidase in
strains pregrown on glutamine medium
Enzyme sp act(mU/mg of protein)
Strain Final growth conditionsa Form- Acet- Histi-ami- ami - daseidase dase dase
Wild type No nitrogen source 90 29 1.510 mM NH4+ 1 5 010 mM glutamate 54 13 0.610 mM glutamine 0 3 0.1
gdhAlO No nitrogen source 78 33 1.310 mM NH4+ 11 18 0.610 mM glutamate 26 10 0.610 mM glutamine 0 4 0.2
a Mycelium was grown for 16 h in 10 mM glutaminemedium and then transferred to this final medium for6 h before harvesting.
which is currently being studied (Polkinghorneand Hynes, unpublished data).These experiments led to the conclusion that
the gdhA10 and areA 102 lesions interact to givea greater relief of ammonium repression ofacetamidase and histidase than does thegdhA 10 lesion alone. Furthermore, both gdhA 10and areA102,gdhA1O are still sensitive to gluta-mate and glutamine repression. Cycloheximidewas found to prevent the derepression of ace-tamidase, benzamidase, and histidase resultingfrom transfer of areA1O2,gdhA1O myceliumfrom glutamate medium to ammonium medium(Table 3). Similarly, relief of glutamine repres-sion was prevented by cycloheximide (Table 4).These results suggest that enzyme synthesis isrepressed by glutamate and glutamine.The mauAl;gdhA1O strain was insensi-
tive to ammonium repression of urate oxidasecompared with the mauA2 strain (Table 5).However, these strains showed similar sensitiv-ity to glutamine repression.
It has also been found that the production ofextracellular protease activity is sensitive toglutamate and glutamine repression. In con-trast to an earlier report (3), it has been foundthat the gdhAlO lesion leads to insensitivity tothe effects of ammonium on extracellular pro-tease. This has been observed in plate testsinvolving clearing of milk powder, as well as inenzyme assays. In plate tests the insensitivity ofthe areA102,gdhA1O strain to ammonium wasvery obvious, but that of the gdhAlO strain wasmore difficult to observe. This difficulty mayaccount for the earlier failure to observe theeffects of the gdhAlO lesion on the milk clear-
1119VOL. 120, 1974
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
J. BACTERIOL.
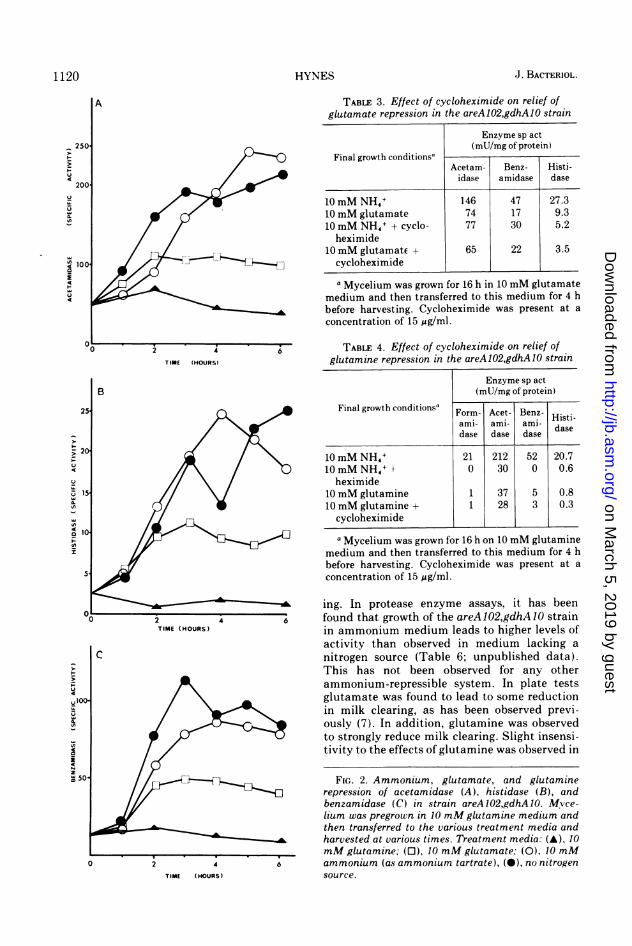
TABLE 3. Effect of cycloheximide on relief ofglutamate repression in the areA102,gdhA10 strain
Enzyme sp act(mU/mg of protein)
Final growth conditionsaAcetam- Benz- Histi-idase amidase dase
10 mM NH4+ 146 47 27.310 mM glutamate 74 17 9.310 mM NH4+ + cyclo- 77 30 5.2heximide
10 mM glutamate + 65 22 3.5cycloheximide
a Mycelium was grown for 16 h in 10 mM glutamatemedium and then transferred to this medium for 4 hbefore harvesting. Cycloheximide was present at aconcentration of 15 ;ig/ml.
TABLE 4. Effect of cycloheximide on relief ofglutamine repression in the areA102,gdhA10 strain
Enzyme sp act(mU/mg of protein)
Final growth conditionsa Form- Acet- Benz-
ami- ami- ami- Histi-dase dase dase dase
10 mM NH4+ 21 212 52 20.710 mM NH4+ t 0 30 0 0.6heximide
10 mM glutamine 1 37 5 0.810 mM glutamine + 1 28 3 0.3
cycloheximidea Mycelium was grown for 16 h on 10 mM glutamine
medium and then transferred to this medium for 4 hbefore harvesting. Cycloheximide was present at aconcentration of 15 jig/ml.
ing. In protease enzyme assays, it has beenfound that growth of the areA 102,gdhA10 strainin ammonium medium leads to higher levels ofactivity than observed in medium lacking anitrogen source (Table 6; unpublished data).This has not been observed for any otherammonium-repressible system. In plate testsglutamate was found to lead to some reductionin milk clearing, as has been observed previ-ously (7). In addition, glutamine was observedto strongly reduce milk clearing. Slight insensi-tivity to the effects of glutamine was observed in
FIG. 2. Ammonium, glutamate, and glutaminerepression of acetamidase (A), histidase (B), andbenzamidase (C) in strain areA102,gdhA10. Mvce-lium was pregrown in 10 mMglutamine medium andthen transferred to the various treatment media andharvested at various times. Treatment media: (A), 10mM glutamine; (0), 10 mM glutamate; (0), 10 mMammonium (as ammonium tartrate), (-), no nitrogensource.
2Z.
,v
4
2(
0
I
!n
"I
TIME (HOURS)
TIME (HOURS)
1120 HYNES
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
NITROGEN CATABOLISM IN A. NIDULANS
the areA1O2,gdhA1O strain. The data presentedin Table 6 support these results. The gdhAlOand areA102,gdhA10 strains were found to besensitive to glutamate and glutamine repressionof extracellular protease activity, although theareA 102,gdhA 1O strain had slightly higheractivity than areA102 in glutamine medium. Itshould be noted that the strains containing theareA102 lesion had higher protease activitiesthan the wild-type strains (areA+), in agree-ment with the results of plate tests (14).An acidic amino-acid uptake system has been
TABLE 5. Glutamine repression of urate oxidase
Urate oxidaseStrain Final growth con- U/mgofStrain ~ditionSa (mu/mg of
protein)
mauA2 Uric acid 96Uric acid + 10 mM 13NH4+
Uric acid + 10 mM 24glutamine
mauA2; gdhA 10 Uric acid 172Uric acid + 10 mM 95NH4+
Uric acid + 10 mM 25glutamine
a Mycelium was grown on 10 mM glutamine me-dium for 16 h and then transferred to this medium for4 h before harvesting. Uric acid was present in themedium at a concentration of 250 gg/ml.
TABLE 6. Ammonium, glutamate, and glutaminerepression of extracellular protease activity
Strain Final growth con- RelativeStrain ~ditionsa proteaseactivityb
Wild type No nitrogen source 4510 mM NH4+ 310 mM glutamine 710 mM glutamate 16
gdhAlO No nitrogen source 2510 mM NH4+ 4110 mM glutamine 710 mM glutamate 10
areA102 No nitrogen source 10010mM NH4+ 210 mM glutamine 210 mM glutamate 75
areA102,gdhA10 No nitrogen source 12210 mM NH4+ 22210 mM glutamine 2110 mM glutamate 63
a Mycelium was grown for 16 h on 10 mM glutaminemedium and then transferred to this medium for 4 hbefore harvesting mycelium and collecting mediumfor extracellular protease assays.
b Arbitrary units (see Materials and Methods).
shown to be subject to ammonium and gluta-mine repression (22), and lesions in the gdhAgene have been shown to lead to insensitivity toammonium repression (16, 20). In an experi-ment measuring [14C]glutamate uptake by theareA 102,gdhA 10 strain, it was found thatuptake was completely insensitive to ammo-nium repression, but growth of mycelium inglutamine medium led to approximately 70%reduction in glutamate uptake capacity com-pared with uptake capacity of mycelium incu-bated in medium lacking a nitrogen source(unpublished data). Therefore, the acidic ami-no-acid uptake system appears to be sensitive toglutamine repression.
In contrast to all the systems mentionedabove, there were clear indications that nitratereductase was much less sensitive to glutaminerepression than to ammonium repression. Asfound previously (3) the presence of the gdhA 10lesion led to insensitivity of nitrate reductase toammonium repression (Table 7). The presenceof glutamine in the medium did not repressnitrate reductase greatly. The degree of repres-sion can probably be accounted for by ammo-nium released from glutamine during growth.This suggestion is supported by the lower activ-ity of the glutamine-grown culture of themauA2 strain compared with the mauA2,gdhA1O strain. It is suggested that the effects ofammonium derived from glutamine on theother sysems investigated is largely obscuredby strong repression by glutamine. Some gluta-minase activity has been observed in A.nidulans (unpublished data).The findings of Cove (9) raise a possible
objection to the conclusion that nitrate reduc-tase is not very sensitive to glutamine repres-
TABLE 7. Ammonium and glutamine repression ofnitrate reductase
NitrateFinal growth con- reductaseStrain ditionsa (mU/mg of
protein)
mauA2 10 mM NOs- 8810 mM NOs- + 1
10 mM NH4+10 mM NOs- + 47
10 mM glutaminemauA2; gdhA 10 10 mM NOs- 125
10mM NOs- + 9110 mM NH4+
10mMNO3- + 10010 mM glutamine
a Mycelium was grown for 16 h on 10 mM glutaminemedium and then transferred to this medium for 4 hbefore harvesting. NO3- was added as sodium nitrate.
1121VOL. 120, 1974
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

HYNES
sion. He found that nitrate interferes with theutilization of many other nitrogen sources.Therefore, it is possible that the presence ofnitrate as inducer results in reduced glutamineuptake and metabolism and so relieves gluta-mine repression. Therefore, the effects of growthin glutamine on nitrate reductase activities inthe strain nirA ,, which produces nitrate reduc-tase in the absence of inducer (19), was investi-gated. This strain is very sensitive to ammo-nium repression but not to glutamine repression(Table 8). Again, the effects of ammoniumderived from glutamine might account for the50% repression (comparison with the urea-grown culture) occurring during growth on glu-tamine. These experiments also show thatgrowth on glutamate does not lead to loweractivity than growth on urea, suggesting thatnitrate reductase may not be sensitive to gluta-mate repression. This conclusion is not surpris-ing in view of the inhibition of glutamateutilization by nitrate (9). It can be concludedthat nitrate reductase is much less sensitive toglutamine repression than to ammoniumrepression.
DISCUSSIONAn important finding of this work is that the
areA102,gdhA10 strain is less sensitive than thegdhAlO strain to ammonium repression of anumber of enzymes. The areA102 lesion leads tohigher levels of some but not all enzymes ofnitrogen catabolism, but these enzymes are stillsubject to strong ammonium repression in theabsence of the gdhAlO lesion (this paper; un-published data). These results therefore providesupport for the proposed involvement of theareA gene in ammonium repression (2). Thereare at least two possible explanations for theobserved interaction between areA 102 andgdhA10. The first suggestion is that a regulatorycomplex is formed between the products ofthese two genes and that this acts to stimulateenzyme synthesis in the absence of ammonium.In the presence of ammonium the action of thiscomplex would be reduced, due to interaction
TABLE 8. Effect of growth on glutamine, glutamate,and ammonium on nitrate reductase activity in the
nirA ,c constitutive strain
Growth condition8a Nitrate reductase(mU/mg of protein)
5 mM urea 5510 mM glutamate 5510 mM glutamine 2610 mM NH4+ 1
a Mycelium was grown for 16 h in this medium andthen harvested.
between ammonium ion and the NADP-GDHpart of the complex. This proposal combines thesuggestions of Pateman et al. (20) and Arst andCove (2). The second possibility is that thegdhAlO lesion results in altered pool levels ofammonium (or a metabolite of ammonium),which then results in insensitivity to ammo-nium repression. If the areA102 gene productwas slightly altered in its interaction withammonium ion (or a metabolite of ammonium),then this might result in a greater relief ofammonium repression in the double mutant.The proposed regulatory function for NADP-GDH can only be resolved by studies on moremutants, including those with defined effects onenzyme structure (e.g., deletion and nonsensemutants), as well as by looking for alterations inthe regulatory function of NADP-GDH inde-pendent of catalytic alterations.Growth of mycelium in glutamate medium
leads to approximately 50% repression of someenzymes of nitrogen catabolism, when com-pared with mycelium incubated in mediumlacking a nitrogen source. Since this occurs instrains insensitive to ammonium repression, itis unlikely that glutamate repression occurs viaproduction of ammonium. Furthermore, there isno evidence for glutamate repression of nitratereductase (8; Table 8) which is very sensitive toammonium repression. Strong glutaminerepression of many systems of nitrogen catabo-lism has been found. This occurs in mutantsinsensitive to ammonium repression, and ni-trate reductase is much less sensitive to gluta-mine repression than to ammonium repression.These data raise the question of whether the
effects of ammonium, glutamate, and gluta-mine are all manifestations of the same regula-tory mechanism or are due to different mech-anisms. One possible way in which there couldbe a common regulatory mechanism is for thegdhAlO lesion to affect the entry of ammoniuminto a pool of ammonium (or a metabolite ofammonium) which is the effector responsible forrepression. The gdhAlO lesion would not affectthe levels of this pool of effector during growthon glutamate or glutamine medium. The effectsof the gdhAlO lesion on processing of ammo-nium could be attributed to loss of catalyticactivity of NADP-GDH (as suggested by vande Poll [24]) or to an unknown role of NADP-GDH in the distribution of ammonium in vari-ous pools. In this connection it is known thatmutants affecting ammonium pool levels can bealtered in sensitivity to ammonium repression(4, 20). A common mechanism for repression bythese compounds, however, is difficult to recon-cile with strong ammonium repression of nitratereductase (which is relieved by the gdhA 10
1122 J. BACTERIOL.
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

NITROGEN CATABOLISM IN A. NIDULANS
lesion) but insensitivity to repression by gluta-mate or glutamine.
Therefore, it is tentatively suggested thatregulation by glutamate and glutamine may beby different mechanism(s) from ammoniumrepression. Whether repression during growthon glutamate and glutamine is due to a singlemechanism (e.g., repression by glutamine or a
metabolite) cannot be determined at present.Glutamine is a strong nitrogen source for A.nidulans and therefore might be utilized inpreference to many other nitrogen sources. Inaddition, glutamine and glutamate are themajor end products of nitrogen catabolism.Therefore, mechanisms of regulation by thesecompounds are not unlikely.
Further studies on glutamine and glutamatemetabolism are necessary to clarify the issuesraised here. In addition, investigation of gluta-mine and glutamate repression in various areAmutants are necessary since the areA gene playsa fundamental role in nitrogen-catabolic regula-tion (2). Such experiments are complicated bythe effects of gene areA on the utilization ofglutamate and glutamine as nitrogen sources
(2, 14; unpublished data). Pateman et al. (20)have proposed that it is the external concentra-tion of ammonium which determines ammo-
nium repression. At present there is no data on
this point from this laboratory, except that in a
preliminary experiment growth of mycelium inglutamine did not result in high levels of ammo-nium in the external medium. Experiments willbe reported elsewhere (M. J. Hynes, Mol. Gen.Genet., in press) showing that methylammo-nium and cesium ion result in repression ofsome enzymes of nitrogen catabolism in strainsinsensitive to ammonium repression. These re-
sults further emphasize the complicated natureof regulation of nitrogen catabolism.
ACKNOWLEDGMENTSThe help of Iole Barbieri and Jan Dickinson and communi-
cations by H. Arst are appreciated.This work was supported by the Australian Research
Grants Committee.
ADDENDUM IN PROOFRecent work has shown that areA lesions can alter
ammonium repression of the acetamidase without af-fecting glutamate or glutamine repression of this en-
zyme. This supports the conclusion that repressionby these compounds may be distinct from ammoniumrepression.
LITERATURE CITED1. Arst, H. N., and D. J. Cove. 1969. Methylammonium
resistance in Aspergillus nidulans. J. Bacteriol.98:1284-1293.
2. Arst, H. N., and D. J. Cove. 1973. Nitrogen metaboliterepression in Aspergillus nidulans. Mol. Gen. Genet.126:111-141.
3. Arst, H. N., and D. W. MacDonald. 1973. A mutant ofAspergillus nidulans lacking NADP-linked glutamatedehydrogenase. Mol. Gen. Genet. 122:261-265.
4. Arst, H. N., and M. M. Page. 1973. Mutants of Aspergil-lus nidulans altered in the transport of methylammo-nium and ammonium. Mol. Gen. Genet. 121:239-245.
5. Charney, J., and R. M. Tomarelli, 1947. A colorimetricmethod for the determination of the proteolytic activityof duodenal juice. J. Biol. Chem. 171:501-505.
6. Cohen, B. L. 1972. Ammonium repression of extracellularprotease in Aspergillus nidulans. J. Gen. Microbiol.71:293-299.
7. Cohen, B. L. 1973. Regulation of intracellular and extra-cellular neutral and alkaline proteases in Aspergillusnidulans. J. Gen. Microbiol. 79:311-320.
8. Cove, D. J. 1966. The induction and repression of nitratereductase in the fungus Aspergillus nidulans. Biochim.Biophys. Acta 113:51-56.
9. Cove, D. J. 1972. The control of nitrogen metabolism inAspergillus nidulans. Heredity 29:119.
10. Dubois, E., M. Grenson, and J. M. Wiame. 1973. Releaseof the "ammonia effect" on three catabolic enzymes byNADP-specific glutamate dehydrogenase-less muta-tions in Saccharomyces cerevisiae. Biochem. Biophys.Res. Commun. 50:967-972.
11. Grenson, M., E. Dubois, M. Piotrowska, R. Drillien, andM. Aigle. 1974. Ammonia assimilation in Sac-charomvces cerevisiae as mediated by the two gluta-mate dehydrogenases. Mol. Gen. Genet. 128:73-85.
12. Hynes, M. J. 1970. Induction and repression of amidaseenzymes in Aspergillus nidulans. J. Bacteriol.103:482-487.
13. Hynes, M. J. 1972. Mutants with altered glucose repres-sion of amidase enzymes in Aspergillus nidulans. J.Bacteriol. 111:717-722.
14. Hynes, M. J. 1973. Pleiotropic mutants affecting thecontrol of nitrogen metabolism in Aspergillus nidulans.Mol. Gen. Genet. 125:99-107.
15. Hynes, M. J., and J. A. Pateman. 1970. The geneticanalysis of regulation of amidase synthesis in Aspergil-lus nidulans. 1. Mutants able to utilize acrylamide.Mol. Gen. Genet. 108:97-106.
16. Kinghorn, J. R., and J. A. Pateman. 1973. NAD andNADP L-glutamate dehydrogenase activity and am-monium regulation in Aspergillus nidulans. J. Gen.Microbiol. 78:39-46.
17. Lewis, C. M., and J. R. S. Fincham. 1970. Regulation ofnitrate reductase in the basidiomycete Ustilagomaydis. J. Bacteriol. 103:55-61.
18. Lowry, 0. H., M. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
19. Pateman, J. A., and D. J. Cove. 1967. Regulation ofnitrate reduction in Aspergillus nidulans. Nature (Lon-don) 215:1234-1237.
20. Pateman, J. A., J. R. Kinghorn, E. Dunn, and E. Forbes.1973. Ammonium regulation in Aspergillus nidulans. J.Bacteriol. 114:943-950.
21. Prival, M. J., and B. Magasanik. 1971. Resistance tocatabolite repression of histidase and proline oxidaseduring nitrogen-limited growth of Klebsiella aerogenes.J. Biol. Chem. 246:6288-6296.
22. Robinson, J. H., C. Anthony, and W. T. Drabble. 1973.Regulation of the acidic amino-acid permease of Asper-gillus nidulans. J. Gen. Microbiol. 79:65-80.
23. Scazzocchio, C., and A. J. Darlington. 1968. The induc-tion and repression of the enzymes of purine breakdownin Aspergillus nidulans. Biochim. Biophys. Acta166:557-568.
24. van de Poll, K. W. 1973. Ammonium repression in amutant of Saccharomyces carlsbergensis lackingNADP dependent glutamate dehydrogenase activity.FEBS Lett. 32:265-266.
1123VOL. 120, 1974
on March 5, 2019 by guest
http://jb.asm.org/
Dow
nloaded from


















