
Effective antimicrobial activity of Cbf-14, derived from a ... · Cbf-14, a cationic peptide...
14
Effective antimicrobial activity of Cbf-14, derived from a cathelin-like domain, against penicillin-resistant bacteria Lingman Ma, Yanrong Wang, Mengxiao Wang, Yuwei Tian, Wei Kang, Hanhan Liu, Hui Wang, Jie Dou * , Changlin Zhou * State Key Laboratory of Natural Medicines, School of Life Science and Technology, China Pharmaceutical University, Nanjing, Jiangsu 210009, PR China article info Article history: Received 2 February 2016 Accepted 4 February 2016 Available online 9 February 2016 Keywords: Cbf-14 Antimicrobial activity Penicillin-resistant bacteria NDM-1-carrying bacteria Membrane permeation DNA binding abstract Cbf-14, a cationic peptide derived from a cathelin-like domain, was designed by inserting the highly a- helical sequence RLLR into an antibacterial sequence and deleting the inactive amino acids in Cbf-K 16 . Clinical penicillin-resistant isolates as well as NDM-1-carrying Escherichia coli and a correspondingly infected mice model were employed to evaluate Cbf-14 antibacterial activity. The results showed that Cbf-14 possessed potent antimicrobial effects with an MIC of 8e64 mg/ml, and killed almost all bacteria within 240 min. Cbf-14-treated mice achieved an 80% survival rate and approximate 2.5 log unit reduction in CFU in tissues; additionally, this peptide significantly suppressed the production of pro- inflammatory cytokines by the disaggregation of lipopolysaccharide (LPS), suggesting its anti- inflammatory effects. Furthermore, Cbf-14, concentration higher than 2 MIC value, increased mem- brane uptake to NPN and PI dye by 96.2% and 63.7%, respectively, neutralised the negative zeta potential of LPS and bacteria surface, and induced 100% leakage of liposome-entrapped calcein and cytoplasmic membrane disruption of E. coli, indicating obvious membrane permeation. Finally, it bound to DNA and respectively evoked 85.0% and 63.3% inhibition of gene replication and protein expression of NDM-1 at sub-MIC concentration in E. coli BL21 (DE3)-NDM-1. These data indicated that Cbf-14 possessed effective antimicrobial activity against penicillin-resistant bacteria in vitro/vivo through membrane disruption, DNA binding, down-regulating NDM-1 expression by plasmid replication inhibition, and anti- inflammatory activity by LPS disaggregation, suggesting a potential anti-infective clinical agent. © 2016 Elsevier Ltd. All rights reserved. 1. Introduction Microbial resistance, the cause of invalidation of major antimi- crobial drugs in clinical settings, has become a worldwide health problem in recent years [1]. The “superbugs”, such as methicillin- resistant Staphylococcus aureus, vancomycin-resistant enterococci and extended-spectrum b-lactamase-producing bacteria, exhibit multidrug resistance to antibiotics as a result of carrying several antibiotic genes, especially New Delhi metallo-b-lactamase-1 (NDM-1), which was found in patients in South Asia and first reported in the Lancet in 2010 [2]. NDM-1, mainly expressed in Escherichia coli (E. coli) and Klebsiella pneumonia, is a novel broad spectrum carbapenemase with a high ability to hydrolyse and inactivate almost all carbapenems, which are used as a last resort for the treatment of multidrug-resistant bacterial infections [3]. Therefore, there is an urgent need for novel, broad-spectrum compounds with rapid antimicrobial effects, new mechanisms of action and the ability to limit the induction of antimicrobial resis- tance in “superbugs”. Cathelicidins form a family of antimicrobial peptides (AMPs) that harbour a highly conserved cathelin-like domain. They are effective against a broad range of microorganisms, including gram- positive and gram-negative bacteria, fungi, and viruses, some of which are clinical drug-resistant strains [4,5]. Cathelicidin-BF (BF- Abbreviations: NDM-1, New Delhi metallo-beta-lactamase-1; BF-30, Cath- elicidin-BF; CFU, Colony forming unit; Glu, Glutamic acid; Lys, lysine; RLLR, Arg- Leu-Leu-Arg; IPTG, Isopropyl b-D-1-thiogalactopyranoside; TEM, Transmission electron microscopy; SDS-PAGE, Sodium Dodecyl Sulphate-Polyacrylamide gel electrophoresis; NPN, N-Phenyl-1-naphthylamine; PI, Propidium iodide; SYTO 9, SYTO ® 9 nucleic acid stain; FITC, Fluorescein isothiocyanate; CCK-8, Cell counting Kit-8; DAPI, 4 0 ,6-diamidino-2-phenylindole; DPPG, 1,2-Dipalmitoyl-sn-glycero-3- phospho-(1 0 -rac-glycerol); DPPE, 1, 2-Dipalmitoyl-sn-glycero-3- phosphoethanolamine. * Corresponding authors. School of Life Science and Technology, China Pharma- ceutical University, 24 Tong Jia Xiang, Nanjing, Jiangsu 210009, PR China. E-mail addresses: [email protected] (J. Dou), [email protected] (C. Zhou). Contents lists available at ScienceDirect Biomaterials journal homepage: www.elsevier.com/locate/biomaterials http://dx.doi.org/10.1016/j.biomaterials.2016.02.011 0142-9612/© 2016 Elsevier Ltd. All rights reserved. Biomaterials 87 (2016) 32e45
Transcript of Effective antimicrobial activity of Cbf-14, derived from a ... · Cbf-14, a cationic peptide...

lable at ScienceDirect
Biomaterials 87 (2016) 32e45
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomateria ls
Effective antimicrobial activity of Cbf-14, derived from a cathelin-likedomain, against penicillin-resistant bacteria
Lingman Ma, Yanrong Wang, Mengxiao Wang, Yuwei Tian, Wei Kang, Hanhan Liu,Hui Wang, Jie Dou*, Changlin Zhou*
State Key Laboratory of Natural Medicines, School of Life Science and Technology, China Pharmaceutical University, Nanjing, Jiangsu 210009, PR China
a r t i c l e i n f o
Article history:Received 2 February 2016Accepted 4 February 2016Available online 9 February 2016
Keywords:Cbf-14Antimicrobial activityPenicillin-resistant bacteriaNDM-1-carrying bacteriaMembrane permeationDNA binding
Abbreviations: NDM-1, New Delhi metallo-betaelicidin-BF; CFU, Colony forming unit; Glu, GlutamicLeu-Leu-Arg; IPTG, Isopropyl b-D-1-thiogalactopyraelectron microscopy; SDS-PAGE, Sodium Dodecylelectrophoresis; NPN, N-Phenyl-1-naphthylamine; PSYTO® 9 nucleic acid stain; FITC, Fluorescein isothiocKit-8; DAPI, 40 ,6-diamidino-2-phenylindole; DPPG, 1phospho-(10-rac-glycerol); DPPE, 1,phosphoethanolamine.* Corresponding authors. School of Life Science an
ceutical University, 24 Tong Jia Xiang, Nanjing, JiangsE-mail addresses: [email protected] (J. Dou), cl_z
http://dx.doi.org/10.1016/j.biomaterials.2016.02.0110142-9612/© 2016 Elsevier Ltd. All rights reserved.
a b s t r a c t
Cbf-14, a cationic peptide derived from a cathelin-like domain, was designed by inserting the highly a-helical sequence RLLR into an antibacterial sequence and deleting the inactive amino acids in Cbf-K16.Clinical penicillin-resistant isolates as well as NDM-1-carrying Escherichia coli and a correspondinglyinfected mice model were employed to evaluate Cbf-14 antibacterial activity. The results showed thatCbf-14 possessed potent antimicrobial effects with an MIC of 8e64 mg/ml, and killed almost all bacteriawithin 240 min. Cbf-14-treated mice achieved an 80% survival rate and approximate 2.5 log unitreduction in CFU in tissues; additionally, this peptide significantly suppressed the production of pro-inflammatory cytokines by the disaggregation of lipopolysaccharide (LPS), suggesting its anti-inflammatory effects. Furthermore, Cbf-14, concentration higher than 2 � MIC value, increased mem-brane uptake to NPN and PI dye by 96.2% and 63.7%, respectively, neutralised the negative zeta potentialof LPS and bacteria surface, and induced 100% leakage of liposome-entrapped calcein and cytoplasmicmembrane disruption of E. coli, indicating obvious membrane permeation. Finally, it bound to DNA andrespectively evoked 85.0% and 63.3% inhibition of gene replication and protein expression of NDM-1 atsub-MIC concentration in E. coli BL21 (DE3)-NDM-1. These data indicated that Cbf-14 possessed effectiveantimicrobial activity against penicillin-resistant bacteria in vitro/vivo through membrane disruption,DNA binding, down-regulating NDM-1 expression by plasmid replication inhibition, and anti-inflammatory activity by LPS disaggregation, suggesting a potential anti-infective clinical agent.
© 2016 Elsevier Ltd. All rights reserved.
1. Introduction
Microbial resistance, the cause of invalidation of major antimi-crobial drugs in clinical settings, has become a worldwide healthproblem in recent years [1]. The “superbugs”, such as methicillin-resistant Staphylococcus aureus, vancomycin-resistant enterococci
-lactamase-1; BF-30, Cath-acid; Lys, lysine; RLLR, Arg-noside; TEM, TransmissionSulphate-Polyacrylamide gelI, Propidium iodide; SYTO 9,yanate; CCK-8, Cell counting,2-Dipalmitoyl-sn-glycero-3-2-Dipalmitoyl-sn-glycero-3-
d Technology, China Pharma-u 210009, PR [email protected] (C. Zhou).
and extended-spectrum b-lactamase-producing bacteria, exhibitmultidrug resistance to antibiotics as a result of carrying severalantibiotic genes, especially New Delhi metallo-b-lactamase-1(NDM-1), which was found in patients in South Asia and firstreported in the Lancet in 2010 [2]. NDM-1, mainly expressed inEscherichia coli (E. coli) and Klebsiella pneumonia, is a novel broadspectrum carbapenemase with a high ability to hydrolyse andinactivate almost all carbapenems, which are used as a last resortfor the treatment of multidrug-resistant bacterial infections [3].Therefore, there is an urgent need for novel, broad-spectrumcompounds with rapid antimicrobial effects, new mechanisms ofaction and the ability to limit the induction of antimicrobial resis-tance in “superbugs”.
Cathelicidins form a family of antimicrobial peptides (AMPs)that harbour a highly conserved cathelin-like domain. They areeffective against a broad range of microorganisms, including gram-positive and gram-negative bacteria, fungi, and viruses, some ofwhich are clinical drug-resistant strains [4,5]. Cathelicidin-BF (BF-

L. Ma et al. / Biomaterials 87 (2016) 32e45 33
30) is a 30 amino acids cathelicidin-like polypeptide from thenon-mammalian vertebrate Bungarus fasciatus. Its precursor has acathelin-like domain at the N-terminus and the maturecathelicidin-BF at the C-terminus [6]. Our previous study indicatedthat BF-30 could destabilize the lipid head groups and producepores in the cell membrane, resulting in the leakage of cytoplasmiccontent [7]. Despite their incomparable advantages compared withconventional antibiotics, source and biological stability limit thewidespread clinical application of natural AMPs. In a previousreport, we introduced a single amino acid mutation, Cbf-K16, byreplacing Glu16 with Lys16 in BF-30 to increase the cationic charge[8]. Consequently, the cathelicidin-BF Lys16 mutant, Cbf-K16, has ahigher net positive charge (þ13) at neutral pH than BF-30, and thismutant can permeate bacterial membrane and interfere with thefunction of specific macromolecular, such as DNA and protein, toinduce bacterial cell death [8].
However, it remained unclear whether the mutated Cbf-K16 wascompetent in terms of bactericidal activity and whether theimprovement of its permeation ability enhanced its antibacterialactivity against penicillin-resistant gram-negative strains. Thus, wedesigned a modified small peptide, Cbf-14, by inserting a highly a-helical sequence RLLR into the N-terminal region of Cbf-K16.Consequently, the cost of Cbf-14 was significantly reduced becausewe retained the antibacterial sequence (KFFRKLKKSV) but deletedthe inactive amino acids of Cbf-K16 (Fig. 1A and B). Cbf-14, with thesequence RLLRKFFRKLKKSV, has a higher fat-soluble index thanCbf-K16 (104.29 vs 71.33) because of the high proportion of leucineresidues. Furthermore, its a-helical content in a bio-mimeticmembrane, 0.2 DPPG/DPPE þ DPPG molar ratio system, wasincreased from 7.1% to 57.0% when compared to the 50% TFE/watersystem (Fig. 1C).
In the present study, we aimed to utilize penicillin-resistantisolates and mice model induced by NDM-1-carrying E. coli toelucidate the antimicrobial activity and potential mode-of-action ofCbf-14 against penicillin-resistant bacteria in vitro and in vivo.
2. Materials & methods
2.1. Materials
Cbf-14 (�98%) and FITC-Cbf-14were synthesized using standardFmoc solid-phase synthesis protocols (GL Biochem Co., Ltd,Shanghai, China). The following reagents and kits were utilized:IPTG (Amresco, Canada, US); TNF-a, IL-6, IL-10 and IL-4 ELISA Kits(MultiSciences Biotech Co., Ltd, Hangzhou, Zhejiang, China); bac-terial genomic DNA extraction kit (Sangon Biotech Co., Ltd,Shanghai, China); tigecycline (Xianhang Medical Technology Co.,Ltd, Nanjing, Jiangsu, China); ampicillin (Sunshine BiotechnologyCo., Ltd, Nanjing, Jiangsu, China); penicillin V (Sangon Biotech Co.,Ltd, Shanghai, China); caspase 1 polyclonal antibody and NF-kB p65polyclonal antibody (ABclonal Biotechnology Co., Ltd, Wuhan,Hubei, China); LIVE/DEAD BacLight™ Bacterial Viability kit L7012(Molecular Probes Inc., Eugene, OR, US); FITC-conjugated LPS(SigmaeAldrich, St. Louis, MO, US); DPPG and DPPE (CordenPharmaInternational, Plankstadt, Germany). Other analytical grade chem-ical reagents were acquired from commercial sources. All drugswere determined using the E-TOXATE kit. Test samples used in thestudy did not show detectable levels of endotoxin within thesensitivity limit of kit (0.1e1.0 EU/ml).
2.2. Bacterial strains
The NDM-1-carrying bacteria, recombinant E. coli BL21 (DE3)-NDM-1, was constructed by inserting the Klebsiella pneumoniaeNDM-1 gene (GenBank accession no. HQ328085) into a pET-28(a)
vector and transforming it into E. coli BL21 (DE3) cells to obtain highNDM-1 activity [9]. E. coli ATCC25922, Pseudomonas aeruginosaATCC27853, S. aureus ATCC25923 were obtained from AmericanType Culture Collection; K. pneumonia, P. aeruginosa, E. coli, S. epi-dermidis, and S. aureus were penicillin-resistant strains isolatedfrom clinical specimens that had been identified by ZhongdaHospital Southeast University (Nanjing, Jiangsu, China).
2.3. Antimicrobial activity assay in vitro
The minimal inhibitory concentration (MIC) of Cbf-14 wasmeasured using a standard micro-dilution method [10]. The mini-mal bactericidal concentration (MBC) was determined as previ-ously described [11]. Bacterial cells were diluted to achieve log(CFU/ml) values in the range of 5e6 and incubated with Cbf-14 orother drugs for 0, 10, 30, 60, 120, 240, or 480 min. At each timepoint, samples (50 ml/time point) were plated onto agar plates atappropriate diluted concentration. After overnight culture, thesurviving colonies were counted, and an untreated inoculum groupwas used as a negative control.
2.4. The establishment of the E. coli BL21 (DE3)-NDM-1 infectedmodel in vivo
Mice were intravenously injected with 0.6 g/kg IPTG once ormultiple times, n ¼ 6 mice per group. Before injection and at 2 hintervals after injection, blood samples were collected to determinethe serum IPTG levels using HPLC analysis. Column: Agilent, HederaNH2 4.6 � 250 mm, 5 mm (Hanbon Sci. & Tech. Co., Ltd., Jiangsu,China).
After the injection of 1.8 g/kg IPTG, mice were inoculatedintraperitoneally with 1 � 1010 CFU/ml (0.5 ml/mice) of E. coli BL21(DE3)-NDM-1 in the presence (n ¼ 15 mice per group, 10 animalsfor survival study and other 5 for NDM-1 activity measurement) orabsence (n¼ 10mice per group) of additional IPTG and then treatedwith Cbf-14 (10 mg/kg, i.p.), ampicillin (80 mg/kg, i.v.), tigecycline(16 mg/kg, i.v.) or the same amount of saline (Fig. 2C). Survival wasmonitored over seven days, mice liver and lung samples werecollected 24 h after infection, homogenized and centrifuged. Thetissue pellets were washed, re-suspended and ultrasonicated onice; then, the sonicated lysates were centrifuged to harvest thecrude enzymes in the supernatant. NDM-1 activity in bacterial-infected liver or lung was determined as previously described[12]. The metallo-b-lactamase producing bacteria in mice liver orlung were analysed by the imipenem-EDTA disk method as previ-ously reported [13]. Briefly, imipenem disks containing 10 mg imi-penem were incubated with or without 4.5 ml of a 0.5 M EDTAsolution (pH 7.8). After drying, the imipenem disks (�/þ EDTA)were placed onto the agar surface evenly coated with 100 ml of liveror lung homogenate. The inhibition zone diameter (IZD) of eachdisk was determined and imaged after incubation for 18 h.
Manipulations of animals were performed in accordance withthe standards set by Science and Technology Department of JiangsuProvince (SYXK 2012-0035). In this experiment, a total of 137 maleICR mice were used to study the E. coli BL21 (DE3)-NDM-1 infectedmodel in vivo.
2.5. Antimicrobial and anti-inflammatory activity assay of Cbf-14 inE. coli BL21 (DE3)-NDM-1-infected mice
Mice (n¼ 10mice per group)were infected, induced and treatedas described above (Fig. 2C). After 12 h of treatment, blood sampleswere obtained from the mice to determine the serum levels ofinterleukin-10 (IL-10), interleukin-4 (IL-4), interleukin-6 (IL-6) andtumour necrosis factor (TNF-a). The lung was bluntly isolated and
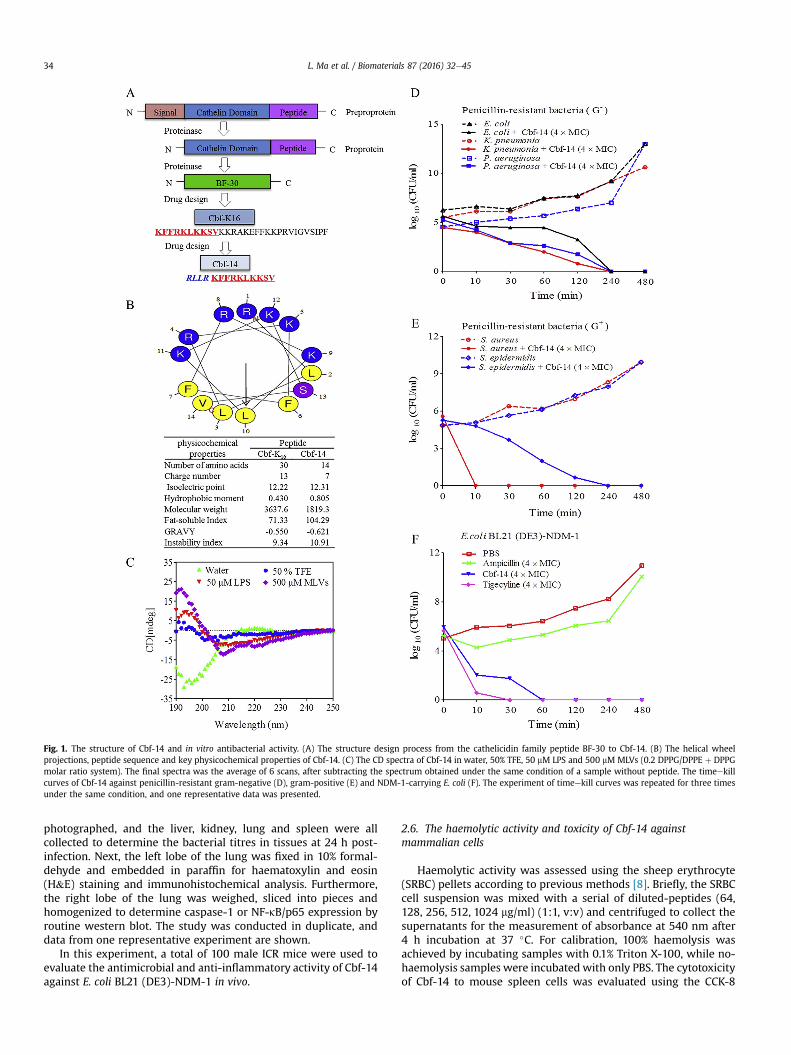
Fig. 1. The structure of Cbf-14 and in vitro antibacterial activity. (A) The structure design process from the cathelicidin family peptide BF-30 to Cbf-14. (B) The helical wheelprojections, peptide sequence and key physicochemical properties of Cbf-14. (C) The CD spectra of Cbf-14 in water, 50% TFE, 50 mM LPS and 500 mM MLVs (0.2 DPPG/DPPE þ DPPGmolar ratio system). The final spectra was the average of 6 scans, after subtracting the spectrum obtained under the same condition of a sample without peptide. The timeekillcurves of Cbf-14 against penicillin-resistant gram-negative (D), gram-positive (E) and NDM-1-carrying E. coli (F). The experiment of timeekill curves was repeated for three timesunder the same condition, and one representative data was presented.
L. Ma et al. / Biomaterials 87 (2016) 32e4534
photographed, and the liver, kidney, lung and spleen were allcollected to determine the bacterial titres in tissues at 24 h post-infection. Next, the left lobe of the lung was fixed in 10% formal-dehyde and embedded in paraffin for haematoxylin and eosin(H&E) staining and immunohistochemical analysis. Furthermore,the right lobe of the lung was weighed, sliced into pieces andhomogenized to determine caspase-1 or NF-kB/p65 expression byroutine western blot. The study was conducted in duplicate, anddata from one representative experiment are shown.
In this experiment, a total of 100 male ICR mice were used toevaluate the antimicrobial and anti-inflammatory activity of Cbf-14against E. coli BL21 (DE3)-NDM-1 in vivo.
2.6. The haemolytic activity and toxicity of Cbf-14 againstmammalian cells
Haemolytic activity was assessed using the sheep erythrocyte(SRBC) pellets according to previous methods [8]. Briefly, the SRBCcell suspension was mixed with a serial of diluted-peptides (64,128, 256, 512, 1024 mg/ml) (1:1, v:v) and centrifuged to collect thesupernatants for the measurement of absorbance at 540 nm after4 h incubation at 37 �C. For calibration, 100% haemolysis wasachieved by incubating samples with 0.1% Triton X-100, while no-haemolysis samples were incubated with only PBS. The cytotoxicityof Cbf-14 to mouse spleen cells was evaluated using the CCK-8
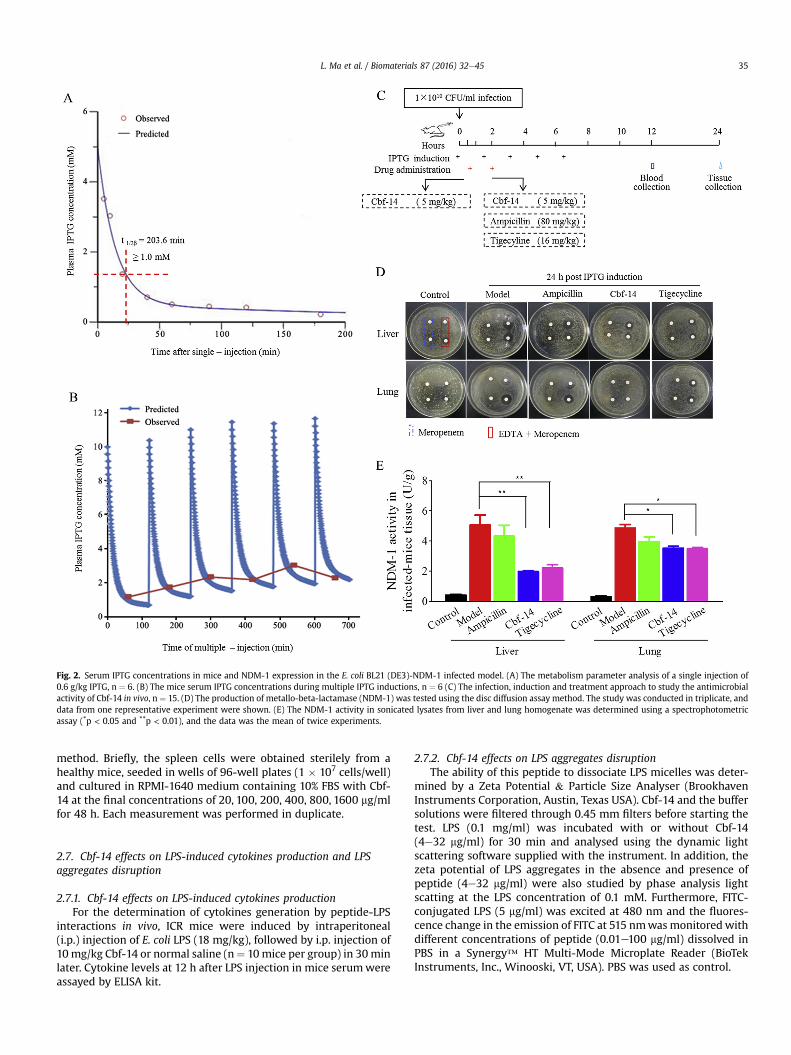
Fig. 2. Serum IPTG concentrations in mice and NDM-1 expression in the E. coli BL21 (DE3)-NDM-1 infected model. (A) The metabolism parameter analysis of a single injection of0.6 g/kg IPTG, n ¼ 6. (B) The mice serum IPTG concentrations during multiple IPTG inductions, n ¼ 6 (C) The infection, induction and treatment approach to study the antimicrobialactivity of Cbf-14 in vivo, n ¼ 15. (D) The production of metallo-beta-lactamase (NDM-1) was tested using the disc diffusion assay method. The study was conducted in triplicate, anddata from one representative experiment were shown. (E) The NDM-1 activity in sonicated lysates from liver and lung homogenate was determined using a spectrophotometricassay (*p < 0.05 and **p < 0.01), and the data was the mean of twice experiments.
L. Ma et al. / Biomaterials 87 (2016) 32e45 35
method. Briefly, the spleen cells were obtained sterilely from ahealthy mice, seeded in wells of 96-well plates (1 � 107 cells/well)and cultured in RPMI-1640 medium containing 10% FBS with Cbf-14 at the final concentrations of 20, 100, 200, 400, 800, 1600 mg/mlfor 48 h. Each measurement was performed in duplicate.
2.7. Cbf-14 effects on LPS-induced cytokines production and LPSaggregates disruption
2.7.1. Cbf-14 effects on LPS-induced cytokines productionFor the determination of cytokines generation by peptide-LPS
interactions in vivo, ICR mice were induced by intraperitoneal(i.p.) injection of E. coli LPS (18 mg/kg), followed by i.p. injection of10mg/kg Cbf-14 or normal saline (n¼ 10mice per group) in 30minlater. Cytokine levels at 12 h after LPS injection in mice serumwereassayed by ELISA kit.
2.7.2. Cbf-14 effects on LPS aggregates disruptionThe ability of this peptide to dissociate LPS micelles was deter-
mined by a Zeta Potential & Particle Size Analyser (BrookhavenInstruments Corporation, Austin, Texas USA). Cbf-14 and the buffersolutions were filtered through 0.45 mm filters before starting thetest. LPS (0.1 mg/ml) was incubated with or without Cbf-14(4e32 mg/ml) for 30 min and analysed using the dynamic lightscattering software supplied with the instrument. In addition, thezeta potential of LPS aggregates in the absence and presence ofpeptide (4e32 mg/ml) were also studied by phase analysis lightscatting at the LPS concentration of 0.1 mM. Furthermore, FITC-conjugated LPS (5 mg/ml) was excited at 480 nm and the fluores-cence change in the emission of FITC at 515 nmwasmonitoredwithdifferent concentrations of peptide (0.01e100 mg/ml) dissolved inPBS in a Synergy™ HT Multi-Mode Microplate Reader (BioTekInstruments, Inc., Winooski, VT, USA). PBS was used as control.

L. Ma et al. / Biomaterials 87 (2016) 32e4536
2.8. Cbf-14 induced membrane rupture
2.8.1. Zeta-potential measurementsThe zeta potential of the bacteria, in the absence and presence of
different Cbf-14 concentrations (0.25, 1, 4 � MIC), was determinedat 25 �C from the mean of 15 measurements and obtained by phaseanalysis light scattering in a Zeta Potential & Particle Size Analyseras described before [14]. Bacterial concentrations were fixed at3 � 107 CFU/mL, in order to acquire high enough count rates.
2.8.2. Flow cytometry assay of Cbf-14 affinity to the bacterial cellmembrane
E. coli BL21 (DE3)-NDM-1 and S. aureus were suspended at1 � 107 CFU/ml in PBS and incubated in separate suspensions withFITC-Cbf-14 (32 mg/ml) for 30 min at 37 �C. The efficiency of Cbf-14adherence to the bacterial cell surface was quantified by flowcytometry.
2.8.3. TEM of Cbf-14-treated bacteriaBacteria from mid-exponential phase cultures (OD600 ¼ 0.5)
were incubated with Cbf-14 for 4 h to determine the morphologicalchanges using TEM as previously described [15]. The sliced sampleswere double stained with uranyl acetate and lead citrate, observedand imaged to investigate the changes in the bacterial cellmembrane.
2.8.4. Outer membrane permeabilityPeptide-induced outer membrane permeability was measured
as previously described [16]. Briefly, log-phase cultures of E. coliBL21 (DE3)-NDM-1 cells were collected, washed and then sus-pended to an OD600 of 0.25 in 5 mM HEPES buffer (pH 7.4), andplated in 96-well blackmicroplates towhich NPN (10 mM) dissolvedin 95% ethanol was added. Thereafter, peptide (final concentrationsof 0.25, 0.5, 1, 2, 4 � MIC) or 0.1% Triton X-100, were respectivelyadded to each well. The time-dependent effect of Cbf-14 on NPNfluorescence was measured at the excitation/emissionwavelengthsof 350/420 nm, respectively.
2.8.5. Inner membrane permeabilityThe ability of Cbf-14 to permeate the inner membrane of E. coli
BL21 (DE3)-NDM-1 was assessed using LIVE/DEAD BacLight™ Bac-terial Viability kit L7012. Briefly, mid-log phase E. coli cells wereharvested and re-suspended in 0.85% (w/v) NaCl to yield1 � 106 CFU/ml. Each sample was incubated with Cbf-14 (0.25, 0.5,1, 2, 4 � MIC) at 37 �C for 1 h, then washed and re-suspended in0.85% (w/v) NaCl. Following the addition of SYTO 9/PI dye mixture,
Table 1The antimicrobial activity of Cbf-14 in vitro
Bacterial strain MIC (mg/ml) MBC (mg/ml)
Cbf-14 Penicillin Cbf-14 Penicillin
G� E. coli BL21 (DE3))-NDM-1(þ) 8 >256 16 >256E. coli BL21 (DE3))-NDM-1(�) 8 >256 16 >256E. coli BL21 (DE3) 16 4 32 16E. colia 16 >256 32 >256E. coli ATCC25922 16 4 32 16K. pneumoniaa 32 >256 128 >256P. aeruginosaa 16 >256 64 >256P. aeruginosa ATCC27853 8 4 32 32
Gþ S. aureusa 64 >256 128 >256S. aureus ATCC25923 32 1 64 4S. epidermidisa 32 >256 128 >256
(þ)NDM-1 carrying E. coli BL21 (DE3) with IPTG (1.0 mM) induction for 4 h.(�)NDM-1 carrying E. coli BL21 (DE3) without IPTG induction.
a Clinical penicillin-resistant strains were isolates from human clinical specimens
the samples were incubated for 15min at room temperature in darkand then analysed by a flow cytometer. The additional bacterialsamples for staining with SYTO 9 alone and with PI alone wereprepared. A bacterial suspension without peptide treatment wasserved as the negative control.
2.8.6. Calcein release assayCbf-14 at final concentrations of 0.25�, 0.5�, 1�, 2� and
4 � MIC was adding to calcein-loaded liposomes, which wereprepared by thin film hydration as described previously with DPPE/DPPG at a ratio of 0.8:0.2 (mol:mol) [17,18]. Real-time fluorescenceintensity was monitored at 530 nm (excitation at 490 nm). Theextent of calcein efflux was calculated as (Ft � F0)/(Fmax � F0),where Ft is the fluorescence of a peptide liposome/calcein solutionat time t, and F0 and Fmax are the initial fluorescence and thefluorescence after complete liposomal disruption by the detergentTriton X-100 (final concentration, 10% w/w), respectively.
2.9. Cbf-14 effects on genomic DNA and plasmid-based replicationand expression of NDM-1
2.9.1. Growth curve of E. coli BL21 (DE3)-NDM-1 in response to sub-MIC Cbf-14
The assay was conducted according to Pal's method with minormodifications [19]. Bacterial suspensions (50 ml; OD600 ¼ 0.5) wereinoculated into a series of duplicate tubes and cultured with con-stant shaking in the absence and presence of different concentra-tions of Cbf-14 (1, 2, or 4 mg/ml). Bacterial suspensions (500 ml) werecollected at different time points (0, 4, 8 and 16 h) to count thebacteria. The experiment was performed in triplicate, and the re-sults were averaged.
2.9.2. Confocal laser scanning microscopyThe localization of sub-MIC Cbf-14 on E. coli BL21 (DE3)-NDM-1
was determined by confocal laser scanning microscopy. Bacteriawere cultured as described for electron microscopy and respec-tively incubated with FITC-Cbf-14 (4 or 8 mg/ml) for 15, 30 and60 min, washed twice with PBS and fixed in 4% polyformaldehyde/PBS for 10min at room temperature. Followed by the centrifugationat 8000 rpm for 3 min, the sediment was re-suspended in DAPI/PBS(DAPI 0.1 mg/ml) and stained for 10 min in dark. After washing DAPIdye, bacteria pellets were re-suspended in 0.1 ml PBS and analysedwith a LSM 700 confocal laser scanning microscope (Carl ZeissMeditec, Oberkochen, Germany).
2.9.3. Cbf-14 DNA binding assayThe DNA binding assay was performed by agarose gel electro-
phoresis. E. coli BL21 (DE3)-NDM-1 cells were incubated at 37 �C for8 h, with a 4 h induction with 1.0 mM IPTG; then, the cells wereobtained by centrifugation. Bacterial plasmid DNA was extractedusing a SanPrep Column Plasmid Mini-Prep Kit and quantified withan ELISA reader at 280 nm. Then, the DNA samples were incubatedwith an equal volume of Cbf-14 (8, 16, 32, or 64 mg/ml) at 25 �C for10 min. DNA migration was detected on a 1% agarose gel [20].
2.9.4. In vivo replication assayMid-log-phase E. coli BL21 (DE3)-NDM-1 bearing plasmid pET-
28(a) which constructed NDM-1 gene was centrifuged, washedand re-suspended in PBS supplemented with 1% TSB, adjusted toOD600 ¼ 0.2, and supplemented with 170 mg/ml chloramphenicol.Each sample was respectively treated with peptide or kept un-treated, and then taken out at the 0, 60, 120, 180, 240 min of co-incubation. Meanwhile, the bacteria pellets treated with differentconcentrations of peptide (1, 2, 4, 8 mg/ml) were also collected after60 min co-incubation. Before the subsequent analysis of PCR, the

L. Ma et al. / Biomaterials 87 (2016) 32e45 37
samples were heated at 100 �C for 30 min and immediately frozeat �80 �C for 1 h, and so repeated three times. The supernatant(1 ml) was used as template in a 25 ml reaction solution including2.5 mM dNTPs mixture, 10 pM each primer and 0.25 U Ex Taq or PfuDNA polymerase for 30 or 35 cycles. The sequences of primer pairfor the NDM-1 gene were 50-CGCGGATCCATGGAATTGCCCAA-TATTATGCACCCGG-30 (forward) and 50-CCGGAATTCTCAGCG-CAGCTTGTCGGCCAT-30 (reverse) with an annealing temperature of56 �C. The plasmid replication of pET-28(a) was monitored overtime with the primer as 50-TAATACGACTCACTATAGGG-30 (T7 pro-moter) and 50-TGCTAGTTATTGCTCAGCGG-30 (T7 terminator). Ascontrol for stalled chromosomal DNA replication, the 16S rRNAgene was amplified. The PCR samples were subsequently loaded ona 1.5% agarose gel containing 0.5 mg/ml ethidium bromide and runin a 1 � TAE buffer system. Bands were visualized under UV light.
2.9.5. NDM-1 expression of E. coli BL21 (DE3)-NDM-1 in responseto sub-MIC Cbf-14
After an overnight culture, a 3 ml suspension was removed asthe normal control, and IPTG at a final concentration of 1.0 mMwasadded to the residual culture. After adding Cbf-14 at a final con-centration of 1, 2, or 4 mg/ml, the mixtures were shaking continu-ously for 4 h. Protein was obtained by sonicating the bacterial cellsafter centrifugation. SDS-PAGE was performed with a 12% sepa-rating gel and a 5% stacking gel to investigate NDM-1 expression inE. coli BL21 (DE3)-NDM-1 cells treated with Cbf-14.
3. Results
3.1. Effective in vitro antimicrobial activity of Cbf-14, derived from acathelin-like domain, against penicillin-resistant and NDM-1-carrying bacteria
Cbf-14 was designed and synthesized based on its parent pep-tide, Cbf-K16, in which a highly a-helical sequence RLLR wasintroduced in the N-terminus, and inactive amino acids in the C-terminus were deleted (Fig.1A and B). Its amino acids sequencewasH-Arg-Leu-Leu-Arg-Lys-Phe-Phe-Arg-Lys-Leu-Lys-Lys-Ser-Val-OHand key physicochemical properties were shown in Fig. 1B. The fat-soluble index and hydrophobicity increased from 71.33 to 104.29,and from 0.430 to 0.805 in Cbf-14, respectively. Meanwhile, its a-helical content in 500 mMMLVswas increased compared to 50% TFE(7.1% versus 57.0%), as shown in Fig. 1C. These data suggested thatCbf-14 could possess superior membrane permeation ability due toincreased hydrophobicity and a-helical content.
The MIC and MBC of Cbf-14 against the penicillin resistant-strains are summarized in Table 1. Cbf-14 possessed potent anti-bacterial activity against clinical E. coli, P. aeruginosa (MIC: 16 mg/ml), S. epidermidis and K. pneumonia (MIC: 16 mg/ml) as well asNDM-1-carrying recombinant E. coli cells (MIC: 8 mg/ml). Penicillindid not inhibit these selected bacteria (MIC > 256 mg/ml).
The timeekill curves showed that Cbf-14 (4 � MIC) exertedkilling effects on both gram-negative and gram-positive bacteria,thoroughly eliminating the bacteria by 4 h after co-incubation(Fig. 1D, E). Interestingly, Cbf-14 exhibited impressive antimicro-bial effects against gram-negative strains, especially NDM-1-carrying E. coli with the MIC of 8 mg/ml (Fig. 1F).
Our results indicated that Cbf-14 had powerful and broad-spectrum antibacterial activity against penicillin-resistant strainsin vitro. Similarly, as prior literature reported, most cathelicidins areproteolytically cleaved to release the cathelin domain and C-ter-minal mature peptides with antimicrobial activities, like dCATH, aa-helical peptide identified from ducks, has a strong antimicrobialactivity against a broad range of bacteria in vitro [21].
3.2. NDM-1 expression in E. coli BL21 (DE3)-NDM-1-infected miceafter IPTG induction
To establish an NDM-1-carrying bacterial infection model toevaluate the antimicrobial activity of Cbf-14 in vivo, we firstdetermined the metabolic parameters of IPTG and NDM-1expression in the mice. IPTG was quickly eliminated from theblood, with t1/2a and t1/2b values of 5 min and 203.6 min, respec-tively (Fig. 2A). According to themetabolic parameters after a singleinjection, Phoenix WinNonlin 6.0 calculated that if the IPTG in-duction dose was increased to 1.8 g/kg, blood IPTG levels would bemaintained at �1.0 mM for longer than 2 h. Thus, in a follow-upexperiment, mice were injected with IPTG for six times (1.8 g/kgeach time) at an interval of 2 h. As the drug concentrationetimecurve of the multiple injections shows in Fig. 2B, blood levels ofIPTG could be maintained at an efficient induction concentration,which was consistent with in vitro reports, and no symptoms oftoxicity were observed in the mice. Therefore, 1.8 g/kg IPTG was anefficient and safe induction dose.
After culturing agar plates coated with liver or lung homogenatefor 18 h, there was no obvious bacteriostatic ring around the imi-penem disks, but a significant IZD surrounded the imipenem diskscontaining EDTA (Fig. 2D). Compared with the control group, theliver and lung NDM-1 activity in the infected mice were elevated12.6- and 16.2-fold, respectively (Fig. 2E).
These data indicated that our induction method could success-fully establish an NDM-1-carrying bacterial-infected mice model.
3.3. Effective antimicrobial activity of Cbf-14 in E. coli BL21 (DE3)-NDM-1-infected mice
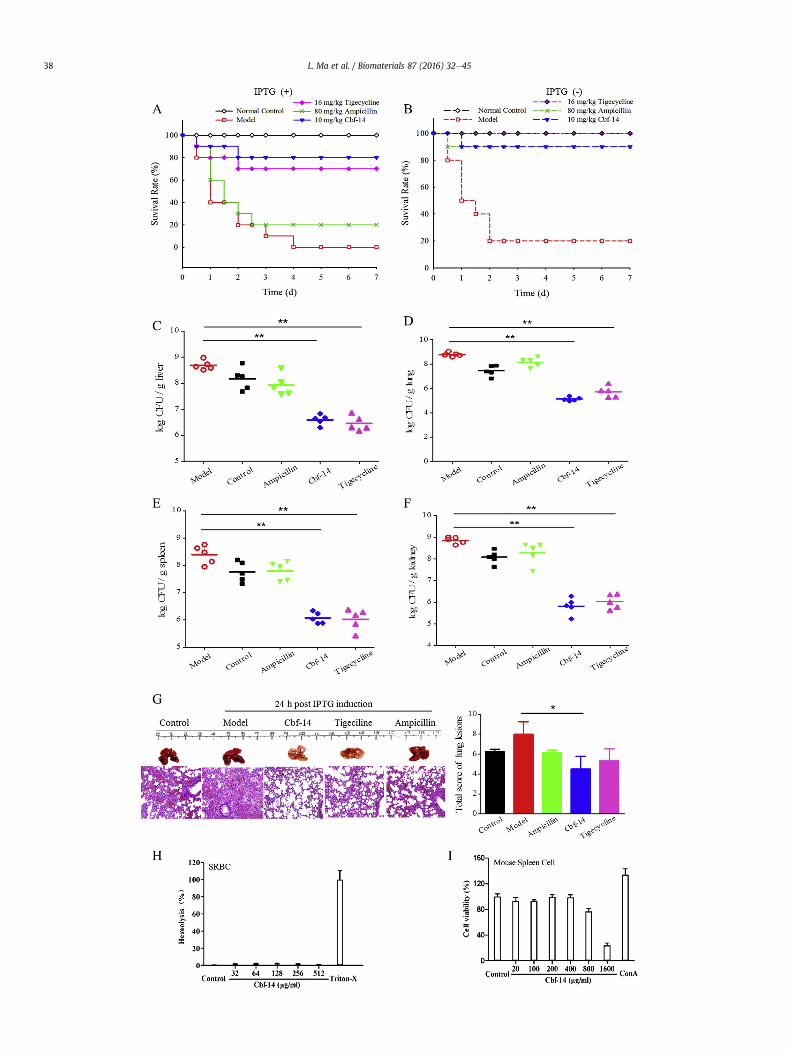
The in vivo antimicrobial activity was mainly determined basedon animal survival, tissue bacterial titres and attenuation of lunginjury. The administration of Cbf-14 (10 mg/kg) in both the IPTG-induced and non-IPTG-induced groups significantly improved thesurvival rate, from 20% to 0% in the saline-control groups to 90% and80%, respectively. However, ampicillin (80 mg/kg) was only effec-tive in mice without IPTG induction, not in induced mice (Fig. 3Aand B). According to the bacteria enumeration results, the survivalimprovements were associated with decreased bacterial titres inthe tissues (liver, lung, spleen, and kidney) of infected mice. Aver-agely, there was more than a 2.5 log unit reduction of CFU in theliver, lung, spleen and kidney in Cbf-14-treated mice (Fig. 3CeF).
Meanwhile, the gross appearance revealed obvious pathologicaldamage with abscesses and blood extravasation in infected micelung. Furthermore, alveolar interstitial congestion and oedema,pulmonary blood vessel dilatation and substantial inflammatorycell infiltration were also observed in the mice model, as evidencedby H&E staining. The administration of Cbf-14 could alleviate theselesions to some degree, but no clear improvement was observed inthe ampicillin-treated group. Similarly, Cbf-14 decreased the totallung lesion scores by approximately 43.8%, and its ability to alle-viate lung injury was greater than that of tigecycline (Fig. 3G).These data indicated that Cbf-14 could reverse the lung damagecaused by bacterial infection. Similarly with this, Paula has foundthat LL-37 could elevate the bacterial clearance in mice lung andenhance host defence against pulmonary infection in vivo [22]. Thedata obtained suggests that Cbf-14 possesses significant antibac-terial activity against NDM-1-carrying E. coli in vivo.
Haemolytic activity is an important parameter to evaluate theeukaryotic cell toxicity of peptide. As shown in Fig. 3H, Cbf-14exhibited a low haemolytic activity (HC50 > 512 mg/ml) againstthe SRBC, caused no obvious proliferation inhibition to mousespleen cells at the concentration up to 800 mg/ml which was muchhigher than effective antimicrobial dose (Fig. 3I), suggesting that
L. Ma et al. / Biomaterials 87 (2016) 32e4538

L. Ma et al. / Biomaterials 87 (2016) 32e45 39
this peptide has no apparent toxicity against mammalian cells.
3.4. Effective anti-inflammatory activity of Cbf-14 in E. coli BL21(DE3)-NDM-1 -infected mice
The anti-inflammatory effect of Cbf-14 was evaluated bydetermining the serum levels of IL-6, TNF-a, IL-4 and IL-10 and theexpression of caspase-1 and NF-kB p65. IL-6 and TNF-a level in themodel group significantly increased at 12 h post-infection, but Cbf-14 treatment decreased the levels of these two pro-inflammatoryfactors, especially those of TNF-a, by approximately 50%. Ampi-cillin had no obvious effect on IL-6 or TNF-a production. Interest-ingly, this peptide also increased serum IL-4 and IL-10 levels to agreater extent than did ampicillin (Fig. 4AeD). Actually, similarfindings were obtained by Lee JK et al. for HPA3P2, a helix-PXXP-helix peptide [23] and by Paula et al. for LL-37 [22]. Chae et al.have demonstrated that caspase-1 activation is regulated by Pyrin.Pyrin is cleaved by caspase-1 and the cleaved N-terminal fragmenttranslocates to nucleus and enhances caspase-recruitment domain-independent NF-kB activation through interactions with p65 NF-kBand IkB-a [24]. NF-kB is of central importance in immune and in-flammatory responses, and its activation related to Pyrin regulated-caspase-1 activation [25]. Our results indicated that caspase-1 andNF-kB p65 immunohistochemistry on model mice lung tissuesshowed strong positive staining, but this staining was significantlyreduced by Cbf-14 (Fig. 4E). Furthermore, the expression ofcaspase-1 and NF-kB p65 as detected by western blotting (Fig. 4F)was consistent with the immunohistochemistry results. Our resultsdemonstrated that Cbf-14 possessed effective anti-inflammatoryactivity in NDM-1-carrying E. coli-infected mice.
3.5. Cbf-14 suppressed LPS-induced inflammation anddisassociated LPS aggregation
LPS aggregation has been found to be important for inflamma-tion triggering [26]. Peptide-induced LPS aggregate disruption canpotentially be coupled to anti-inflammatory effects. Here, ELISAmethod was used to confirm the effect of Cbf-14 on the productionof pro-inflammatory cytokines induced by LPS. As shown in Fig. 5A,LPS (18 mg/kg, i.p.) alone induced the production of TNF-a and IL-6for about 948 and 360 ng/L, respectively. But in Cbf-14 (10 mg/kg)treated group, mice serum levels of IL-6 and TNF-a significantlydecreased, while IL-10 and IL-4 were much higher than that of LPSalone group, which was consistent with the anti-inflammatoryactivity of Cbf-14 against NDM-1-carrying E. coli infection model.Similarly, Artur Schmidtchen et al. also found that pro-inflammatory cytokines IL-6, TNF-a, and MCP-1 were reduced inKYE28-treated mice, whereas the anti-inflammatory IL-10 wastransiently increased [27]. Hu X et al. demonstrated that TRIAPderived decoy peptide (TR6) significantly inhibited LPS-inducedproduction of TNF-a, IL-1b and IL-6 [28].
LPS is a major component of E. coli outer membrane andcontribute to the negative zeta potential of bacterial surface. In ourpresent study, the results of zeta potential measurements indicated
Fig. 3. Antibacterial activity of Cbf-14 in E. coli BL21 (DE3)-NDM-1-infected mice. (AeB) Surv(1 � 1010 CFU/ml) (A) with repeated IPTG induction or (B) without induction and treated witof normal saline. (CeF) Colonization of E. coli BL21 (DE3)-NDM-1 inoculum in the liver, splee14 on acute lung damage in infected mice. The gross appearance and H&E images (�40) of lumanner by a certified pathologist according to a semi-quantitative scoring rank (right) (*pactivity of peptide against the SRBC pellets. Peptide (final concentration at 32, 64, 128, 256control: 0.1% Triton X-100, negative control: PBS. (I) The cytotoxicity of Cbf-14 to mouse spleone representative experiment were shown.
that pure LPS solution and the suspension of E. coli cells exhibitedaverage negative zeta potential of �28.0 mV and �33.2 mV(Fig. 5C), respectively, but the potential shifted towards neutralitywere observed as the addition of Cbf-14 (16 mg/ml), suggesting thispeptide effectively neutralizes the negative charge of LPS andbacterial surface. As we know, LPS forms micellar aggregates inwater and FITC fluorescence is highly quenched in FITC-LPS mi-celles [29]. Interactions of some AMPs with LPS may cause anenhancement of the FITC fluorescence because of dequenching,which indicates the dissociation of large LPS aggregates intosmaller sizes [30]. Here, the dynamic-light-scattering studiesshowed that LPS alone produced micellar aggregates with adiameter centred at 10360 nm, but a dramatic shift of the size of LPStoward a lower value (175 nm)was observed in the presence of Cbf-14 (16 mg/ml), and the aggregation centred at 10360 nm totallydisappeared (Fig. 5B). Additionally, Cbf-14 had a strong effect on theFITC-conjugated LPS aggregates, and the addition of peptide causeda dose-dependent increase of FITC-conjugated LPS fluorescence.When the concentration of Cbf-14 was higher than 10 mg/ml, thefluorescence intensity increased more than 60% after 1 h incuba-tion, as shown in Fig. 5D. Similar findings were also obtained by SunY et al. for chensinin-1b [17] and by Bhunia et al. forfowlicidin-1fragments [31], the fluorescence intensity was increased as theaddition of these peptides in a dose-depended manner.
The data presented above demonstrated that Cbf-14 exertedanti-inflammatory by inducing the LPS aggregate disintegration.
3.6. Cbf-14 effectively enhanced the membrane permeation ofpenicillin-resistant bacteria
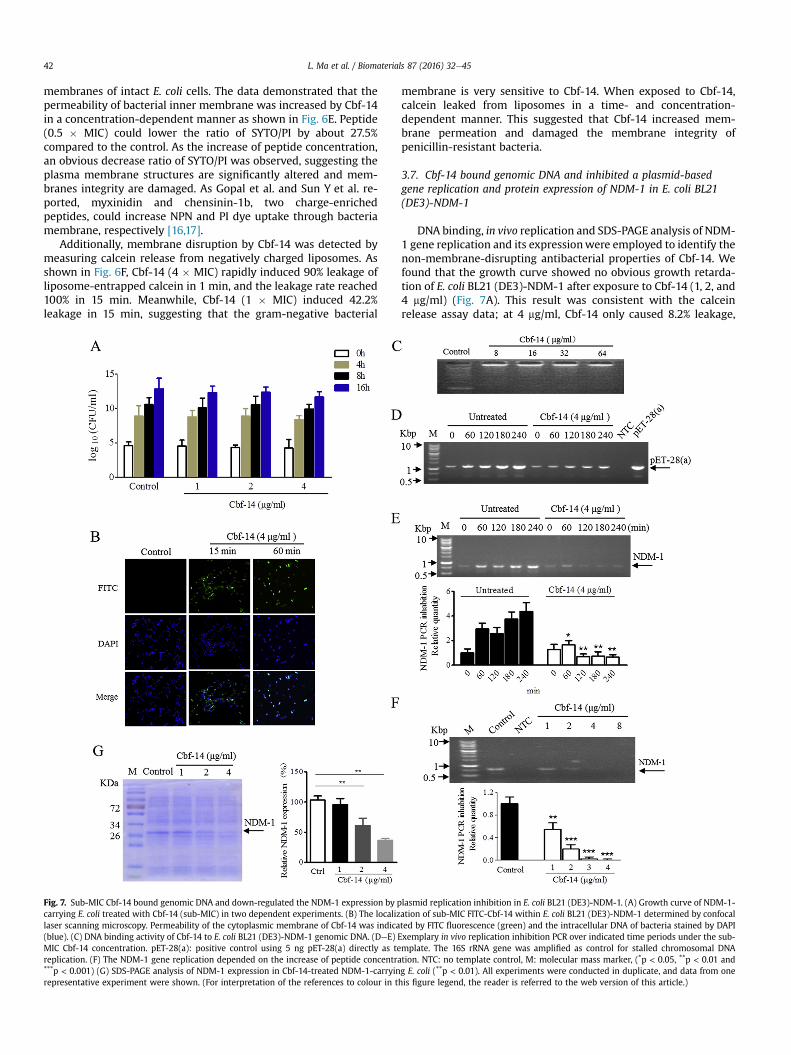
Zeta-potential was used to test the interaction of Cbf-14 withE. coli which presents a negative charge (�33.15 mV). By addingpeptide, there was a concentration-dependent zeta-potential in-crease from �19.49 to �2.49 mV (Fig. 6A), suggesting Cbf-14 couldeffectively neutralize the negative charge of gram-negative bacte-rial surface.
Affinity plays a vital role in peptide-membrane interactions thatenable membrane disruption [32]. The affinity of Cbf-14 for themembranes of penicillin-resistant E. coli and S. aureus was evalu-ated using FITC-labelled Cbf-14. Cbf-14 (1�MIC) had a high affinityfor the bacterial cell membrane after 30 min of co-incubation; theadherence rates were 97.9% and 91.1% for E. coli and S. aureus,respectively (Fig. 6B).
To further explore whether Cbf-14 could interfere with bacterialcell membrane integrity, morphological changes in E. coli andS. aureus isolates were determined by TEM 4 h after treatment withCbf-14 (80 mg/ml). As shown in Fig. 6C, compared with the smooth,clearly visible structure and continuous double membrane of theuntreated bacteria, the cytoplasmic membrane of Cbf-14-treatedcells was almost completely ruptured. Nevertheless, the cell wallbackbone remained recognizable. Simultaneously, a large amountof cellular debris was observed in the visual field, and numerousbleb-like structures were present, indicating that the outer walls ofthe bacterial cells had been separated layer-by-layer from the cell
ival plot of mice inoculated via an intraperitoneal injection with NDM-1-carrying E. colih Cbf-14 (10 mg/kg), ampicillin (80 mg/kg), tigecycline (16 mg/kg) or the same amountn, lung and kidney of infected mice induced with IPTG. (G) The protective effect of Cbf-ng lesions after infection (left). The total lung lesion scores were generated in a blinded< 0.05 and **p < 0.01). The numbers of mice in each group were 10. (H) Haemolyticand 512 mg/ml) was respectively mixed with SRBC for 4 h incubation at 37 �C. Positiveen cells after 48 h incubation. The studies were conducted in duplicate, and data from
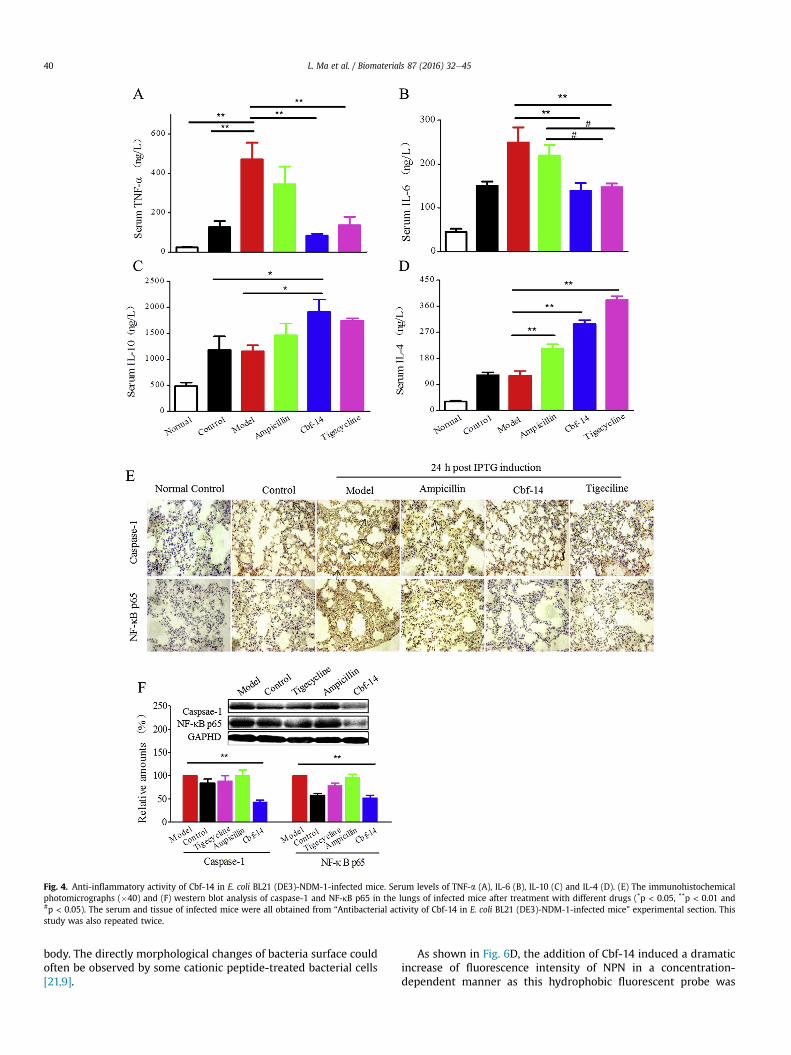
Fig. 4. Anti-inflammatory activity of Cbf-14 in E. coli BL21 (DE3)-NDM-1-infected mice. Serum levels of TNF-a (A), IL-6 (B), IL-10 (C) and IL-4 (D). (E) The immunohistochemicalphotomicrographs (�40) and (F) western blot analysis of caspase-1 and NF-kB p65 in the lungs of infected mice after treatment with different drugs (*p < 0.05, **p < 0.01 and#p < 0.05). The serum and tissue of infected mice were all obtained from “Antibacterial activity of Cbf-14 in E. coli BL21 (DE3)-NDM-1-infected mice” experimental section. Thisstudy was also repeated twice.
L. Ma et al. / Biomaterials 87 (2016) 32e4540
body. The directly morphological changes of bacteria surface couldoften be observed by some cationic peptide-treated bacterial cells[21,9].
As shown in Fig. 6D, the addition of Cbf-14 induced a dramaticincrease of fluorescence intensity of NPN in a concentration-dependent manner as this hydrophobic fluorescent probe was
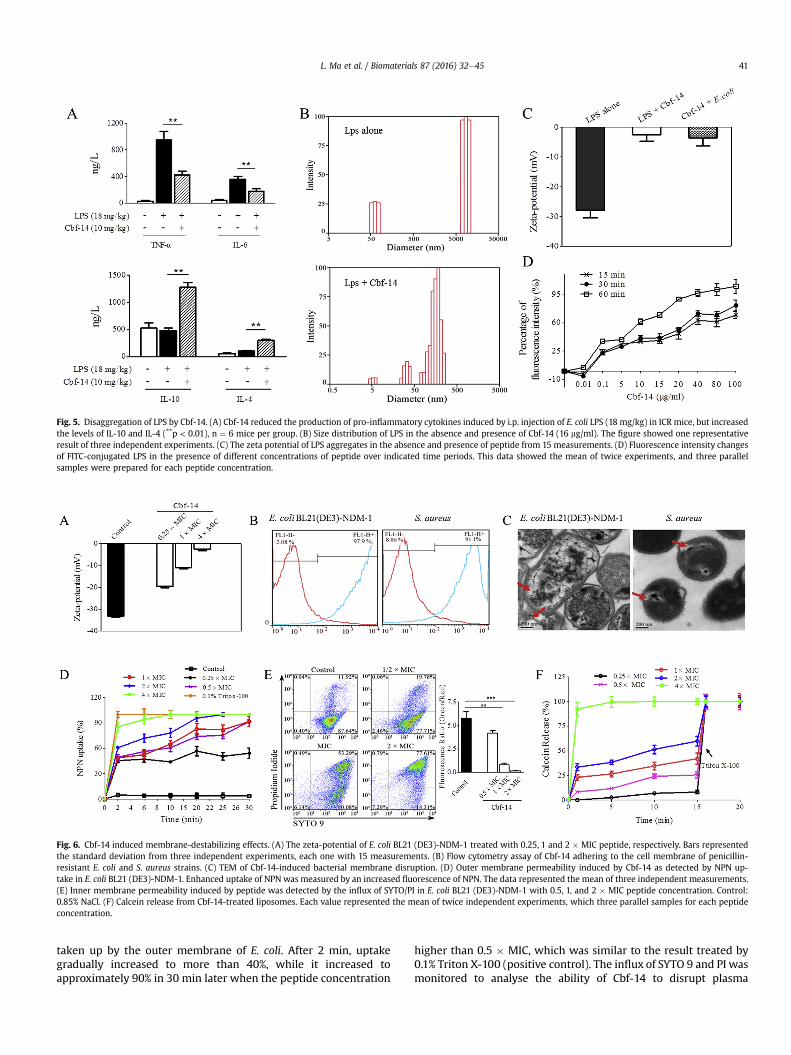
Fig. 5. Disaggregation of LPS by Cbf-14. (A) Cbf-14 reduced the production of pro-inflammatory cytokines induced by i.p. injection of E. coli LPS (18 mg/kg) in ICR mice, but increasedthe levels of IL-10 and IL-4 (**p < 0.01), n ¼ 6 mice per group. (B) Size distribution of LPS in the absence and presence of Cbf-14 (16 mg/ml). The figure showed one representativeresult of three independent experiments. (C) The zeta potential of LPS aggregates in the absence and presence of peptide from 15 measurements. (D) Fluorescence intensity changesof FITC-conjugated LPS in the presence of different concentrations of peptide over indicated time periods. This data showed the mean of twice experiments, and three parallelsamples were prepared for each peptide concentration.
Fig. 6. Cbf-14 induced membrane-destabilizing effects. (A) The zeta-potential of E. coli BL21 (DE3)-NDM-1 treated with 0.25, 1 and 2 � MIC peptide, respectively. Bars representedthe standard deviation from three independent experiments, each one with 15 measurements. (B) Flow cytometry assay of Cbf-14 adhering to the cell membrane of penicillin-resistant E. coli and S. aureus strains. (C) TEM of Cbf-14-induced bacterial membrane disruption. (D) Outer membrane permeability induced by Cbf-14 as detected by NPN up-take in E. coli BL21 (DE3)-NDM-1. Enhanced uptake of NPN was measured by an increased fluorescence of NPN. The data represented the mean of three independent measurements.(E) Inner membrane permeability induced by peptide was detected by the influx of SYTO/PI in E. coli BL21 (DE3)-NDM-1 with 0.5, 1, and 2 � MIC peptide concentration. Control:0.85% NaCl. (F) Calcein release from Cbf-14-treated liposomes. Each value represented the mean of twice independent experiments, which three parallel samples for each peptideconcentration.
L. Ma et al. / Biomaterials 87 (2016) 32e45 41
taken up by the outer membrane of E. coli. After 2 min, uptakegradually increased to more than 40%, while it increased toapproximately 90% in 30 min later when the peptide concentration
higher than 0.5 � MIC, which was similar to the result treated by0.1% Triton X-100 (positive control). The influx of SYTO 9 and PI wasmonitored to analyse the ability of Cbf-14 to disrupt plasma
L. Ma et al. / Biomaterials 87 (2016) 32e4542
membranes of intact E. coli cells. The data demonstrated that thepermeability of bacterial inner membrane was increased by Cbf-14in a concentration-dependent manner as shown in Fig. 6E. Peptide(0.5 � MIC) could lower the ratio of SYTO/PI by about 27.5%compared to the control. As the increase of peptide concentration,an obvious decrease ratio of SYTO/PI was observed, suggesting theplasma membrane structures are significantly altered and mem-branes integrity are damaged. As Gopal et al. and Sun Y et al. re-ported, myxinidin and chensinin-1b, two charge-enrichedpeptides, could increase NPN and PI dye uptake through bacteriamembrane, respectively [16,17].
Additionally, membrane disruption by Cbf-14 was detected bymeasuring calcein release from negatively charged liposomes. Asshown in Fig. 6F, Cbf-14 (4 � MIC) rapidly induced 90% leakage ofliposome-entrapped calcein in 1 min, and the leakage rate reached100% in 15 min. Meanwhile, Cbf-14 (1 � MIC) induced 42.2%leakage in 15 min, suggesting that the gram-negative bacterial
Fig. 7. Sub-MIC Cbf-14 bound genomic DNA and down-regulated the NDM-1 expression by pcarrying E. coli treated with Cbf-14 (sub-MIC) in two dependent experiments. (B) The localizlaser scanning microscopy. Permeability of the cytoplasmic membrane of Cbf-14 was indica(blue). (C) DNA binding activity of Cbf-14 to E. coli BL21 (DE3)-NDM-1 genomic DNA. (DeE) EMIC Cbf-14 concentration. pET-28(a): positive control using 5 ng pET-28(a) directly as tereplication. (F) The NDM-1 gene replication depended on the increase of peptide concentra***p < 0.001) (G) SDS-PAGE analysis of NDM-1 expression in Cbf-14-treated NDM-1-carryinrepresentative experiment were shown. (For interpretation of the references to colour in t
membrane is very sensitive to Cbf-14. When exposed to Cbf-14,calcein leaked from liposomes in a time- and concentration-dependent manner. This suggested that Cbf-14 increased mem-brane permeation and damaged the membrane integrity ofpenicillin-resistant bacteria.
3.7. Cbf-14 bound genomic DNA and inhibited a plasmid-basedgene replication and protein expression of NDM-1 in E. coli BL21(DE3)-NDM-1
DNA binding, in vivo replication and SDS-PAGE analysis of NDM-1 gene replication and its expressionwere employed to identify thenon-membrane-disrupting antibacterial properties of Cbf-14. Wefound that the growth curve showed no obvious growth retarda-tion of E. coli BL21 (DE3)-NDM-1 after exposure to Cbf-14 (1, 2, and4 mg/ml) (Fig. 7A). This result was consistent with the calceinrelease assay data; at 4 mg/ml, Cbf-14 only caused 8.2% leakage,
lasmid replication inhibition in E. coli BL21 (DE3)-NDM-1. (A) Growth curve of NDM-1-ation of sub-MIC FITC-Cbf-14 within E. coli BL21 (DE3)-NDM-1 determined by confocalted by FITC fluorescence (green) and the intracellular DNA of bacteria stained by DAPIxemplary in vivo replication inhibition PCR over indicated time periods under the sub-mplate. The 16S rRNA gene was amplified as control for stalled chromosomal DNAtion. NTC: no template control, M: molecular mass marker, (*p < 0.05, **p < 0.01 andg E. coli (**p < 0.01). All experiments were conducted in duplicate, and data from onehis figure legend, the reader is referred to the web version of this article.)

L. Ma et al. / Biomaterials 87 (2016) 32e45 43
which represented the maximum percentage of calcein leakage atthis concentration (Fig. 6E). Furthermore, confocal laser scanningmicroscopy was used to investigate the location of sub-MIC Cbf-14within bacterial cells. As shown in Fig. 7B, Cbf-14 (4 mg/ml) wasable to permeate bacterial membrane and then bind nuclear acidafter 1 h co-incubation. In addition, the electrophoretic gelmobilityshift assay showed that Cbf-14 bound genomic DNA in E. coli BL21(DE3)-NDM-1, and the electrophoretic movement of DNA wasalmost completely inhibited by Cbf-14 (8 mg/ml). Compared withthe non-complexed DNA control, an obvious retardation wasobserved, and the DNA migration at different concentrations ofCbf-14 was similar (Fig. 7C). This DNA binding effect, like that ofTachyplesin and Buforin, could interfere with the biological func-tions of bacteria [33]. Furthermore, a plasmid-based replicationassay was developed to test the effect of Cbf-14 on bacterialreplication in vivo. As Fig. 7D showed, in bacteria treated solelywith chloramphenicol, plasmid concentration increased over time.But addition of Cbf-14 at concentration of its sub-MIC value couldimpede plasmid replication. Meanwhile, we found that the NDM-1gene replication was also significantly blocked by Cbf-14 overindicated time periods compared to the bacteria treated solely withchloramphenicol, and this inhibitory in a dose-depended manner(Fig. 7EeF). Similarly, Hansmann et al. found that FLG2, a skin-derived C-terminal Filaggrin-2 Fragments, interfered with thebacterial replication in vitro and in vivo [34]. Notably, NDM-1expression in E. coli BL21 (DE3)-NDM-1 induced with IPTG wasinhibited by different concentrations of Cbf-14 (1, 2, and 4 mg/ml) ina concentration-dependent manner after 4 h exposure, as shown inFig. 7G.
Specially, Cbf-14 (4 mg/ml) evoked a greater than 64% reductionin NDM-1 expression. These suggested that at low concentrations(sub-MIC), Cbf-14 achieved antibacterial effects against NDM-1-carrying bacteria mainly by binding the genomic DNA, inhibitingplasmid replication and finally down-regulating NDM-1 expressioninstead of disrupting the bacterial membrane.
4. Discussion
Antimicrobial resistance represents a general public healthconcern; certain multi-resistant organisms not only exhibit resis-tance to a single agent but are frequently resistant tomost availableantimicrobial agents [35]. Cathelicidin-like peptides exert potentantimicrobial activity against clinical drug-resistant strains. In thepresent study, a cationic peptide, Cbf-14, was designed based on thestructure-activity analysis of an analogue of the cathelicidin-likepolypeptide BF-30, Cbf-K16. Cbf-14 contains 14 amino acids, anti-microbial residues 1 to 10 of Cbf-K16 and an RLLR a-helical motif,and exhibits a high fat-soluble index, hydrophobicity, and a-helicalcontent in hydrophobic environments. As CD spectroscopy shown,its a-helical content was increased sharply from 0% in water to57.0% in 500 mM MLVs, It has reported that an increased antimi-crobial activity of a linear a-helical peptide is close correlated withan increase of a-helical secondary structure [36]. To investigatewhether this designwas benefit to its antimicrobial activity, clinicalpenicillin-resistant strains as well as NDM-1-carrying E. coli BL21(DE3)-NDM-1 cells and a correspondingly infected mice model wasused. The in vitro antimicrobial activity assay showed that Cbf-14displays potent antimicrobial activity against penicillin-resistantbacteria with an MIC of 8e64 mg/ml; and kills almost all bacteriawithin 240min. Furthermore, Cbf-14-treatedmice achieved an 80%survival rate, a greater than 2.5 log unit reduction in CFU in tissuesand a 43.8% reduction in lung injury, suggesting that Cbf-14 exertseffective antimicrobial activity in vivo, especially against NDM-1-carrying bacteria. It also indicates that Cbf-14, which combinesthe antibacterial sequence of Cbf-K16 and the RLLR sequence, has
powerful and broad-spectrum antibacterial activity againstpenicillin-resistant strains. Meanwhile, this peptide exhibits lowhaemolytic activity and cytotoxicity toward mammalian cells.
However, the antimicrobial activity of Cbf-14 against gramnegative bacteria was higher than gram positive bacteria (Table 1),such as S. aureus, suggesting its more competitive potentials.Similar results also appeared from the membrane disruption data(Fig. 6C). As previously reported [7,9], AMPs of cathelicidins familyhave direct antimicrobial effects against a broad spectrum of mi-crobes, but exhibits a more prominent activity against gram-negative bacteria than gram-positive bacteria, such as Cbf-K16and BF-30, which range of MICs for gram-negative bacteria andgram-positive bacteria were 2e32 mg/ml and 16e32 mg/ml,respectively.
In our previous study, NDM-1-carrying E. coli with high NDM-1activity were generated by inserting the K. pneumoniae NDM-1gene (GenBank accession no. HQ328085) into a pET28a vectorand transforming it into E. coli BL21 (DE3) cells, because the robustreproductive ability, rapid transmission speed and limited sourcemake it difficult to perform numerous experiments by NDM-1-carrying isolates. Excitingly, this recombinant E. coli BL21 (DE3)-NDM-1 cell has been utilized in in vivo studies with IPTG inductionat the optimal concentration of 1.0 mM for 4 h [8]. Therefore, in thisstudy, we attempted to use this strain to duplicate a safe and stableinfection model in mice to investigate the antimicrobial activity ofCbf-14 in vivo. Herein, mice were injected with IPTG for six times(1.8 g/kg each time) at an interval of 2 h to induce the in vivoexpression of NDM-1. As a result, higher NDM-1 expression and alower survival rate were observed in IPTG-induced mice. Thissuggests that the mice model induced by NDM-1-carrying E. coliwas duplicated successfully and it could be widely and safely usedfor the screening of antibacterial agents.
In many cases, the current therapy for infection involves anti-bacterial and ant-inflammatory drugs [37]. Here, the antimicrobialand anti-inflammatory activities of Cbf-14 were evaluated by thismode. We found that this peptide significantly improved the sur-vival rate to approximately 80% and alleviated the lung injury inE. coli infected mice, and this protective effect was similar to LL-37,which could elevate the bacterial clearance in tissues and enhancehost defence against pulmonary infection [22]. Meanwhile, theseresults are consistent with the in vitro bacterial susceptibility as-says, and they demonstrate that Cbf-14 exerts strong antibacterialactivity against penicillin-resistant bacteria.
LPS plays a crucial role in the pathophysiology of inflammationsepsis and shock caused by gram-negative bacteria [38]. Oncereleased into the blood system, it will cause monocytes andphagocytic cells to produce large amounts of cytokines such as TNF-a, IL-6, and interleukin-8 (IL-8). Because of a growing number ofstudies on peptide-LPS interaction, we investigated the conse-quence of their interaction for cytokine generation in this study.Bezzerri et al. [39] and Liu Y et al. [40] have reported that thepeptide TB-KKG6A and the synthetic hybrid peptides LB-PG/CA-PGcan down-regulate the cytokines IL-8, IL-6 and TNF-a and inhibitthe expression of LPS-induced proinflammatory cytokines.
Similarly, our peptide also reduced serum IL-6 and TNF-a levelsbut increased IL-4 and IL-10 levels in not only bacteria infectedmice but also LPS alone induced-mice, suggesting Cbf-14 exertsanti-inflammatory activity through interfering with peptide-LPSinteraction. As LPS aggregation has been found to be importantfor inflammation triggering, we investigated whether Cbf-14 couldinduce LPS aggregate disruption, and demonstrated that this pep-tide, similar with anti-inflammatory peptide LL-37, could disasso-ciate LPS aggregates. Meanwhile, a down-regulation of NF-kBactivation was observed in our study, but the anti-inflammatorymechanism involved in NF-kB pathway deserves future

L. Ma et al. / Biomaterials 87 (2016) 32e4544
investigation.For AMPs, which can bind to negatively charged anionic phos-
pholipids on LPS of gram-negative bacteria [41], damages the cellmembrane and kills bacteria by membranolytic effects afterachieving a threshold concentration on bacteria surface [42]. Here,Cbf-14 accumulated on bacteria surface, and neutralised thenegative charges, thereby increased the permeability of cellmembrane. Consequently, the bacterial membrane structure wasaltered; pores were formed as the concentrations increase, leadingto the leakage of important cytoplasm. In our study, Cbf-14(1 � MIC) had a high affinity for penicillin-resistant E. coli isolateswith adherence rates of 97.9%, increased membrane uptake to NPNand PI in a dose-dependent manner, evoked effective cytoplasmicmembrane disruption after 4 h treatment with 80 mg/ml peptide.Furthermore, peptide induced 42.2% leakage of liposome-entrapped calcein in 15 min, but the leakage rapidly increased to90% at 1 min and 100% at 15 min as the concentration of Cbf-14increased to 4 � MIC (Fig. 6F), suggesting obvious membranepermeation.
Meanwhile, it has been reported that non-membrane-permeabilizing AMPs can bind and inhibit DNA, RNA or proteinsynthesis processes [43]. LL37, a cathelicidin family cationic pep-tide, displays binding affinities to DNA, thereby affecting replicationevents [44]. Interestingly, in the present study, Cbf-14 could bindgenomic DNA of E. coli BL21 (DE3)-NDM-1, disrupt plasmid DNAreplication, including drug-resistance gene expression. To monitorthe effect of Cbf-14 on in vivo replication, an assay was developedby addition of the translation inhibitor chloramphenicol to blockthe replication of the bacterial chromosome [45]. Results showedthat the replication of NDM-1 gene transformed by recombinantplasmid was increased over indicated time points, but it wassignificantly reduced upon Cbf-14 treatment in a dose-dependentmanner in E. coli (Fig. 7 EeF). Meanwhile, a decreased NDM-1protein expression was also observed in sub-MIC peptide treatedE. coli. All these data suggest Cbf-14 exerts antibacterial activityagainst penicillin-resistant strains potentially by binding genomicDNA, inhibiting plasmid replication and finally down-reguating theplasmid-based gene replication and protein expression of the drug-resistant genes. However, from the data obtained, it is still unclearwhether Cbf-14 could bind to drug-resistant gene specifically, suchas NDM-1, which needs to be further clarified.
Together with above, Cbf-14 has potent antimicrobial activityin vitro with low MICs value and effective killing activity againstpenicillin-resistant bacteria, especially NDM-1-carrying E. coli,mainly due to the membrane rupture induced by its high fat-soluble index, hydrophobicity and a-helical content resulted fromthe structure design. Further, through binding genomic DNA,inhibiting plasmid replication and finally interfering with theplasmid-based replication and expression of drug-resistant gene isthe non-lytic modes of action for its antimicrobial activity againstpenicillin-resistant bacteria. Meanwhile, this peptide, displays LPSaggregates disruption, thereby affecting LPS-induced inflammationwith improved animal survival and alleviated organic injury inE. coli BL21 (DE3)-NDM-1-infected mice, suggesting its potentialclinical application.
5. Conclusions
The antibacterial activity of Cbf-14 against penicillin-resistantbacteria is retained, or even strengthened, in vivo and in vitrowith the interception of the antibacterial sequence and introduc-tion of the RLLR sequence within the parent peptide. Meanwhile,Cbf-14 combines an anti-inflammatory effect with antibacterialactivity in mice infected by NDM-1-carrying bacteria, and theseactivities are responsible for the peptide induced LPS aggregates
disruption. For its antimicrobial mechanism, high concentration ofCbf-14 (�MIC) could directly increase the permeability and evendisrupt the membrane of bacterial cells; Cbf-14 at concentration ofsub-MIC potentially by binding genomic DNA, inhibiting plasmidreplication and finally down-regulating the plasmid-based genereplication and protein expression of drug-resistant genes.
Acknowledgements
This work was supported by the Scientific and TechnologicalSupport & Social Development Plan of Jiangsu Province(BE2012743), and the Priority Academic Programme Development(PAPD) of Jiangsu Higher Education Institution.
References
[1] A.J. Huh, Y.J. Kwon, “Nanoantibiotics”: a new paradigm for treating infectiousdiseases using nanomaterials in the antibiotics resistant era, J. Control. Release156 (2011) 128e145.
[2] T.R. Walsh, J. Weeks, D.M. Livermore, M.A. Toleman, Dissemination of NDM-1positive bacteria in the New Delhi environment and its implications for hu-man health: an environmental point prevalence study, The. Lancet Infect Dis.11 (2011) 355e362.
[3] S. Padhi, New Delhi metallo-beta-lactamase: a weapon for the newlyemerging drug-resistant bacteria, Indian J. Med. Sci. 65 (2011) 317e320.
[4] J.F. Sanchez, F. Wojcik, Y.S. Yang, M.P. Strub, J.M. Strub, A. Van Dorsselaer, etal., Overexpression and structural study of the cathelicidin motif of theprotegrin-3 precursor, Biochemistry 41 (2002) 21e30.
[5] L.T. Nguyen, E.F. Haney, H.J. Vogel, The expanding scope of antimicrobialpeptide structures and their modes of action, Trends Biotechnol. 29 (2011)464e472.
[6] Y.P. Wang, J. Hong, X.H. Liu, H.L. Yang, R. Liu, J. Wu, et al., Snake cathelicidinfrom Bungarus fasciatus is a potent peptide antibiotics, PLoS One (2008),http://dx.doi.org/10.1371/Journal.Pone.0003217.
[7] H.M. Zhou, J. Dou, J. Wang, L.L. Chen, H. Wang, W.D. Zhou, et al., The anti-bacterial activity of BF-30 in vitro and in infected burned rats is throughinterference with cytoplasmic membrane integrity, Peptides 32 (2011)1131e1138.
[8] Y.W. Tian, H. Wang, B. Li, M.Y. Ke, J. Wang, J. Dou, C.L. Zhou, The cathelicidin-BF Lys16 mutant Cbf-K16 selectively inhibits non-small cell lung cancer pro-liferation in vitro, Oncol. Rep. 30 (2013) 2502e2510.
[9] Q.R. Hao, H. Wang, J. Wang, J. Dou, M. Zhang, W.D. Zhou, C.L. Zhou, Effectiveantimicrobial activity of Cbf-K16 and Cbf-A7A13 against NDM-1-carryingEscherichia coli by DNA binding after penetrating the cytoplasmic mem-brane in vitro, J. Pept. Sci. 193 (2013) 173e180.
[10] I. Wiegand, K. Hilpert, R.E. Hancock, Agar and broth dilution methods todetermine the minimal inhibitory concentration (MIC) of antimicrobial sub-stances, Nat. Protoc. 3 (2008) 163e175.
[11] J.R. Zgoda, J.R. Porter, A convenient microdilution method screening naturalproducts against bacteria and fungi, Pharm. Biol. 3 (2001) 221e225.
[12] W. Huang, Z. Beharry, Z. Zhang, T. Palzkill, A broad-spectrum peptide inhibitorof b-lactamase identified using phage display and peptide arrays, Protein Eng.16 (2003) 853e860.
[13] B.A. Fomda, A. Khan, D. Zahoor, NDM-1 (New Delhi metallo beta lactamase-1)producing gram-negative bacilli: emergence & clinical implications, Indian J.Med. Res. 140 (2014) 672e678.
[14] M.M. Domingues, P.S. Santiago, M.A.R.B. Castanho, N.C. Santos, What can lightscattering spectroscopy do for membrane-active peptide studies? J. Pept. Sci.14 (2008) 394e400.
[15] E. de Leeuw, C. Li, P. Zeng, M. Diepeveen-de Buin, W.Y. Lu, E. Breukink, W. Lu,Functional interaction of human neutrophil peptide-1 with the cell wallprecursor lipid II, FEBS Lett. 584 (2010) 1543e1548.
[16] M.H. Hyo, G. Ramamourthy, P. Yoonkyung, Design and membrane-disruptionmechanism of charge-enriched AMPs exhibiting cell selectivity, high-saltresistance, and anti-biofilm properties, Amino Acids (2015), http://dx.doi.org/10.1007/s00726-015-2104-0.
[17] Y. Sun, W.B. Dong, L. Sun, L.J. Ma, D.J. Shang, Insights into the membraneinteraction mechanism and antibacterial properties of chensinin-1b, Bio-materials 37 (2015) 299e311.
[18] A. Oszl�anczi, A. B�ota, E. Klumpp, Influence of aminoglycoside antibiotics onthe thermal behaviour and structural features of DPPE-DPPG model mem-branes, Colloids Surf. B Biointerfaces 75 (2010) 141e148.
[19] S. Pal, Y.K. Tak, J.M. Song, Does the antibacterial activity of silver nanoparticlesdepend on the shape of the nanoparticle? A study of the gram-negativebacterium Escherichia coli, Appl. Environ. Microbiol. 73 (2007) 1712e1720.
[20] Y.Q. Wang, J. Su, F. Wu, P. Lu, L.F. Yuan, W.E. Yuan, J. Sheng, T. Jin, et al.,Biscarbamate cross-linked polyethylenimine derivative with low molecularweight, low cytotoxicity, and high efficiency for gene delivery, Int. J.Nanomed. 7 (2012) 693e704.
[21] W. Gao, L. Xing, P. Qu, T. Tan, N. Yang, D. Li, H. Chen, X. Feng, Identification of a

L. Ma et al. / Biomaterials 87 (2016) 32e45 45
novel cathelicidin antimicrobial peptide from ducks and determination of itsfunctional activity and antibacterial mechanism, Sci. Rep. (2015), http://dx.doi.org/10.1038/srep17260.
[22] P.E. Beaumont, B. McHugh, E. Gwyer Findlay, A. Mackellar, K.J. Mackenzie,R.L. Gallo, J.R. Govan, A.J. Simpson, D.J. Davidson, Cathelicidin host defencepeptide augments clearance of pulmonary Pseudomonas aeruginosa infectionby its influence on neutrophil function in vivo, PLoS One 9 (2014) e99029.
[23] J.K. Lee, S.C. Park, K.S. Hahm, Y. Park, A helix-PXXP-helix peptide with anti-bacterial activity without cytotoxicity against MDRPA-infected mice, Bio-materials 35 (2014) 1025e1039.
[24] J.J. Chae, I. Aksentijevich, D.L. Kastner, Advances in the understanding of fa-milial Mediterranean fever and possibilities for targeted therapy, Br. J. Hae-matol. 146 (2009) 467e478.
[25] C.E. Hellweg, The nuclear factor kB pathway: a link to the immune system inthe radiation response, Cancer Lett. 368 (2015) 275e289.
[26] M. Mueller, B. Lindner, S. Kusumoto, K. Fukase, A.B. Schromm, U. Seydel,Aggregates are the biologically active units of endotoxin, J. Biol. Chem. 279(2004) 26307e26313.
[27] A. Schmidtchen, M. Malmsten, (Lipo)polysaccharide interactions of antimi-crobial peptides, J. Colloid Interface Sci. 449 (2015) 136e142.
[28] X. Hu, Y. Fu, Y. Tian, Z. Zhang, W. Zhang, X. Gao, X. Lu, Y. Cao, N. Zhang, Theanti-inflammatory effect of TR6 on LPS-induced mastitis in mice, Int. Immu-nopharmacol. 30 (2016) 150e156.
[29] J.K. Kim, E. Lee, S. Shin, K.W. Jeong, J.Y. Lee, S.Y. Bae, S.H. Kim, J. Lee, S.R. Kim,D.G. Lee, J.S. Hwang, Y. Kim, Structure and function of papiliocin with anti-microbial and anti-inflammatory activities isolated from the swallowtailbutterfly, Papilio xuthus, J. Biol. Chem. 286 (2011) 41296e41311.
[30] Y. Rosenfeld, N. Papo, Y. Shai, Endotoxin (lipopolysaccharide) neutralizationby innate immunity host-defense peptides. Peptide properties and plausiblemodes of action, J. Biol. Chem. 281 (2006) 1636e1643.
[31] A. Bhunia, H. Mohanram, S. Bhattacharjya, Lipopolysaccharide bound struc-tures of the active fragments of fowlicidin-1, a cathelicidin family of antimi-crobial and antiendotoxic peptide from chicken, determined by transferrednuclear overhauser effect spectroscopy, Biopolymers 92 (2009) 9e22.
[32] M. Wu, R.E. Hancock, Interaction of the cyclic antimicrobial cationic peptidebactenecin with the outer and cytoplasmic membrane, J. Biol. Chem. 274(1999) 29e35.
[33] E.T. Uyterhoeven, C.H. Butler, D. Ko, D.E. Elmore, Investigating the nucleic acidinteractions and antimicrobial mechanism of buforin II, FEBS Lett. 582 (2008)
1715e1718.[34] B. Hansmann, J.M. Schr€oder, U. Gerstel, Skin-derived C-terminal filaggrin-2
fragments are Pseudomonas aeruginosa-directed antimicrobials targetingbacterial replication, PLoS Pathog. 11 (2015) e1005159.
[35] C.S. Zinn, H. Westh, V.T. Rosdahl, An international multicenter study of anti-microbial resistance and typing of hospital Staphylococcus aureus isolatesfrom 21 laboratories in 19 countries or states, Microb. Drug Resist 10 (2004)160e168.
[36] N. Pathak, R. Salas-Auvert, G. Ruche, M.H. Janna, D. Mccarthy, R.G. Harrison,Comparison of the effects of hydrophobicity, amphiphilicity, and alpha-helicity on the activities of antimicrobial peptides, Proteins 22 (1995)182e186.
[37] R. Bucki, K. Leszczynska, F.J. Byfield, D.E. Fein, E. Won, K. Cruz, et al., Combinedantibacterial and anti-inflammatory activity in a cationic disubstituteddexamethasone spermine conjugate (D2S), Antimicrob. Agents Chemother. 54(2010) 2525e2533.
[38] H. Breithaupt, The new antibiotics, Nat. Biotechnol. 17 (2000) 1165e1169.[39] V. Bezzerri, C. Avitabile, M.C. Dechecchi, I. Lampronti, M. Borgatti,
G. Montagner, et al., Antibacterial and anti-inflammatory activity of a tem-porin B peptide analogue on an in vitro model of cystic fibrosis, J. Pept. Sci. 20(2014) 822e830.
[40] Y. Liu, X. Xia, L. Xu, Y. Wang, Design of hybrid beta-hairpin peptides withenhanced cell specificity and potent anti-inflammatory activity, Biomaterials34 (2013) 237e250.
[41] E.M. Tytler, G.M. Anantharamaiah, D.E. Walker, V.K. Mishra,M.N. Palgunachari, J.P. Segrest, Molecular basis for prokaryotic specificity ofmagainin-induced lysis, Biochemistry 34 (1995) 4393e4401.
[42] J.L. Narayana, H.N. Huang, C.J. Wu, J.Y. Chen, Epinecidin-1 antimicrobial ac-tivity: In vitro membrane lysis and In vivo efficacy against Helicobacter pyloriinfection in a mouse model, Biomaterials 61 (2015) 41e51.
[43] M. Scocchi, M. Mardirossian, G. Runti, M. Benincasa, Non-membrane per-meabilizing modes of action of antimicrobial peptides on bacteria, Curr. Top.Med. Chem. 16 (2016) 76e88.
[44] R. Lande, J. Gregorio, V. Facchinetti, B. Chatterjee, Y.-H. Wang, B. Homey, et al.,Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial pep-tide, Nature 449 (2007) 564e569.
[45] K. Skarstad, E. Boye, H.B. Steen, Timing of initiation of chromosome replicationin individual Escherichia coli cells, EMBO J. 5 (1986) 1711e1717.


















