
Complement Factor H Inhibits CD47-Mediated Resolution of ...

Effect of Rheumatoid Factor on Complement-MediatedPhagocytosis
F. C. McDuffie, H. W. Brumfield
J Clin Invest. 1972;51(12):3007-3014. https://doi.org/10.1172/JCI107128.
The frequency and amount of IgM rheumatoid factor (RF) in the blood of patients with rheumatoid arthritis (RA) correlatewith the severity of the disease and the number of complications. Though previous studies of RF in subacute bacterialendocarditis have shown that RF inhibits phagocytosis of microorganisms by granulocytes, the presence of low levels ofcomplement (C) in blood and synovial fluid of patients with the highest titers of RF suggests that an interaction betweenRF and C may contribute to the inflammatory process in RA. We thus employed a quantitative methodology to examinethe effect of RF on complement-dependent phagocytosis of sheep erythrocytes by rabbit granulocytes. Addition of 2500molecules of IgM RF to sheep cells heavily coated with IgG antibody (195,000 molecules per cell) resulted in virtuallycomplete inhibition of uptake of C3 (β1c) and prevention of phagocytosis, an effect resulting from inhibition of uptake ofC1 by the cells. When erythrocytes coated with only 34,000 molecules of IgG antibody were employed, phagocytosis wassimilarly inhibited. However the effect of RF on such cells was shown to be primarily mediated through inhibition of C4rather than C1 uptake. Although the results do not exclude the participation of an IgM RF of higher avidity, present only inthe tissues in rheumatoid inflammation, circulating IgM RF probably does not play […]
Research Article
Find the latest version:
https://jci.me/107128/pdf

Effect of Rheumatoid Factor on
Complement-Mediated Phagocytosis
F. C. McDUFFIE and H. W. BRUMFIELD
From the Departments of Medicine and Microbiology, Mayo Clinic and Mayo
Foundation, Rochester Minnesota 55901
A B S T R A C T The frequency and amount of IgM rheu-matoid factor (RF) in the blood of patients with rheu-matoid arthritis (RA) correlate with the severity ofthe disease and the number of complications. Thoughprevious studies of RF in subacute bacterial endocarditishave shown that RF inhibits phagocytosis of micro-organisms by granulocytes, the presence of low levels ofcomplement (C) in blood and synovial fluid of patientswith the highest titers of RF suggests that an interactionbetween RF and C may contribute to the -inflammatoryprocess in RA. We thus employed a quantitative metho-dology to examine the effect of RF on complement-de-pendent phagocytosis of sheep erythrocytes by rabbitgranulocytes. Addition of 2500 molecules of IgM RF tosheep cells heavily coated with IgG antibody (195,000molecules per cell) resulted in virtually complete in-hibition of uptake of C3 (flue) and prevention of phago-cytosis, an effect resulting from inhibition of uptake ofCl by the cells. When erythrocytes coated with only34,000 molecules of IgG antibody were employed, phag-ocytosis was similarly inhib.ited. However the effect ofRF on such cells was shown to be primarily mediatedthrough inhibition of C4 rather than Cl uptake. Al-though the results do not exclude the participation of anIgM RF of higher avidity, present only in the tissuesin rheumatoid inflammation, circulating IgM RF prob-ably does not play a potentiating role in rheumatoidinflammation.
INTRODUCTIONWhether rheuxnatoid factor (RF)1 (anti-IgG antibodiespresent in the sera of most patients with rheumatoid ar-
This work was presented in part at the 17th Interim Sci-entific Session of the American Rheumatism Association,10 December 1971.
Received for publicationt 27 March 1972 and in revisedform 25 July 1972.
'Abbreviations used in this paper: C, complement; EM,
thritis [RA] ) contributes to the pathogenesis of the in-flammation of synovium or other tissues in this disease isnot known. Though the exper.iments of Hollander andhis collaborators (1, 2) (who found that injection ofautologous IgG into quiescent joints could produce ex-acerbations of synovitis) were interpreted on the basisof an immune complex reaction between injected IgG andRF, this work has not been successfully repeated (3).Davis has postulated that RF may actually inhibit inflam-mation produced by immune complexes (4). In supportof this latter concept Messner and his associates (5)found that IgM RF isolated from patients with subacutebacterial endocarditis was able to inhibit polymorpho-nuclear leukocyte phagocytosis of the infecting organismscoated with 7S IgG antibody. The inhibitory effect ob-served seemed to be primarily directed at the phagocy-tosis-promoting effect of fresh normal serum.
In the present study we have pursued the question ofthe effect of IgM RF on complement-mediated phagocy-tosis employing a quantitative methodology aimed at de-fining the effect with respect to quantities of antibodyand RF involved as well as seeking to identify the point inthe complement sequence at which RF acts. The resultsshow that IgM RF does inhibit complement-mediatedphagocytosis by blocking fixation of both Cl and C4.Interference with Cl fixation predominates at high levelsof IgG antibody whereas interference with C4 fixationplays an important role at lower levels of antibody.
METHODSAntibodies and immunoglobulins. IgM RF was pre-
pared from a pool of five sera containing high titers of RF.All five donors to this pool had active RA at the time ofbleeding and RF titers by latex fixation tests were from1: 2,500 to 1: 10,000. After ultracentrifugation for 60 hr at200,000 g at a density of 1.250 (a-hieved by addition of so-dium bromide) to remove lipoprotein, the lower layer wasdialyzed against pH 7.0 0.01 M phosphate buffer to pre-
effective molecule; RA, rheumatoid arthritis; RF, rheuma-toid factor.
The Jouirnal of Clinical Itnvestigationi Voltumle 51 December 1972 33007
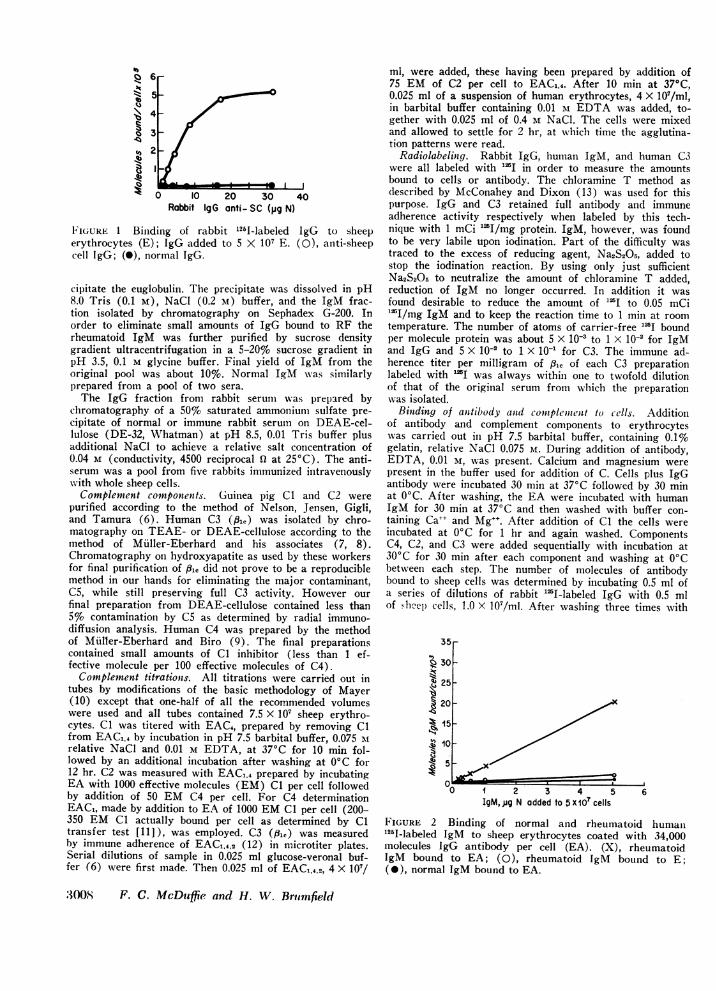
0O 10 20 30 40Rabbit IgG anti- SC (pg N)
FIGURE 1 Binding of rabbit l26l-labeled IgG to sheeperythrocytes (E); IgG added to 5 X 107 E. (0), anti-sheepcell IgG; (0), normal IgG.
cipitate the euglobulin. The precipitate was dissolved in pH8.0 Tris (0.1 M), NaCl (0.2 M) buffer, and the IgM frac-tion isolated by chromatography on Sephadex G-200. Inorder to eliminate small amounts of IgG bound to RF therheumatoid IgM was further purified by sucrose densitygradient ultracentrifugation in a 5-20% sucrose gradient inpH 3.5, 0.1 M glycine buffer. Final yield of IgM from theoriginal pool was about 10%o. Normal IgM was similarlyprepared from a pool of two sera.
The IgG fraction from rabbit seruml was prepared bychromatography of a 50%o saturated ammoniutmi sulfate pre-cipitate of normal or immune rabbit serum on DEAE-cel-lulose (DE-32, Whatman) at pH 8.5, 0.01 Tris buffer plusadditional NaCl to achieve a relative salt concentration of0.04 M (conductivity, 4500 reciprocal Q at 25°C). The anti-serum was a pool from five rabbits immunized intravetnouslywith whole sheep cells.
Complement compontents. Guinea pig Cl and C2 werepurified according to the method of Nelson, Jensen, Gigli,and Tamura (6). Human C3 (,ic) was isolated by chro-matography on TEAE- or DEAE-cellulose according to themethod of Muller-Eberhard and his associates (7, 8).Chromatography oni hydroxyapatite as used by these workersfor final purification of #,B1 did not prove to be a reproduciblemethod in our hands for eliminating the major contaminant,C5, while still preserving full C3 activity. However ourfinal preparation from DEAE-cellulose contained less than5%s contamination by C5 as determined by radial immuno-diffusion analysis. Human C4 was prepared by the methodof Muller-Eberhard and Biro (9). The final preparationscontained small amounts of Cl inhibitor (less than 1 ef-fective molecule per 100 effective molecules of C4).
Complement titrations. All titrations were carried out intubes by modifications of the basic methodology of Mayer(10) except that one-half of all the recommended volumeswere used and all tubes contained 7.5 X 107 sheep erythro-cytes. Cl was titered with EACG, prepared by removing Clfrom EAC1,4 by incubation in pH 7.5 barbital buffer, 0.075 Mirelative NaCl and 0.01 M EDTA, at 370C for 10 min fol-lowed by an additional incubation after washing at 0°C for12 hr. C2 was measured with EAC1,4 prepared by incubatingEA with 1000 effective molecules (EM) Cl per cell followedby addition of 50 EM C4 per cell. For C4 determinationEAC1, made by addition to EA of 1000 EMCl per cell (200-350 EM Cl actually bound per cell as determined by Cltransfer test [11]), was employed. C3 (#,,e) was measuredby immune adherence of EAC1,4,2 (12) in microtiter plates.Serial dilutions of sample in 0.025 ml glucose-veronal buf-fer (6) vere first made. Then 0.025 ml of EAC1,4,2, 4 X 1OV
ml, were added, these having been prepared by addition of75 EM of C2 per cell to EAC1,4. After 10 min at 37°C,0.025 ml of a suspension of human erythrocytes, 4 X 107/ml,in barbital buffer containing 0.01 Ai EDTA was added, to-gether with 0.025 ml of 0.4 M NaCl. The cells were mixedand allowed to settle for 2 hr, at which time the agglutina-tion patterns were read.
Radiolabelinzg. Rabbit IgG, humain IgM, and human C3were all labeled with "I in order to measure the amountsbound to cells or antibody. The chloramine T method asdescribed by McConahey and Dixon (13) was used for thispurpose. IgG and C3 retained full antibody and immuneadherence activity respectively when labeled by this tech-nique with 1 mCi 'I/mg protein. IgM, however, was foundto be very labile upon iodination. Part of the difficulty wastraced to the excess of reducing agent, Na2S205, added tostop the iodination reaction. By using only just sufficientNa2S205 to neutralize the amount of chloramine T added,reduction of IgM no longer occurred. In addition it wasfound desirable to reduce the amount of "I to 0.05 mCi"I/mg IgM and to keep the reaction time to 1 min at room
temperature. The number of atoms of carrier-free "I boundper molecule protein was about 5 X 10-3 to 1 X 10-2 for IgMand IgG and 5 x 10-2 to 1 X 101- for C3. The immune ad-herence titer per milligram of j#% of each C3 preparationlabeled with '1I was always within one to twofold dilutionof that of the original serum from which the preparationwas isolated.
Binding of anttiboldy a)ld comnplenent to c-ells. Additionof antibody and complement components to erythrocyteswas carried out in pH 7.5 barbital buffer, containing 0.1%gelatin, relative NaCl 0.075 AM. During addition of antibody,EDTA, 0.01 M, was present. Calcium and magnesium werepresent in the buffer used for addition of C. Cells plus IgGantibody were incubated 30 min at 37°C followed by 30 minat 0°C. After washing, the EA were incubated with humanIgM for 30 min at 37°C and then washed with buffer con-taining Ca++ and Mg'+. After addition of Cl the cells wereincubated at 0'C for 1 hr and again washed. ComponentsC4, C2, and C3 were added sequentially with incubation at30°C for 30 min after each component and washing at 0°Cbetween each step. The number of molecules of antibodybound to sheep cells was determined by incubating 0.5 ml ofa series of dilutions of rabbit "SI-labeled IgG with 0.5 mlof bchep) cells, 1.0 X 107/ml. After washing three times with
1 2 3 4 5IgM,pug N added to 5 x 107 cells
FIGURE 2 Binding of normal and rheumatoid humai1251-labeled IgM to sheep erythrocytes coated with 34,000molecules IgG antibody per cell (EA). (X), rheumatoidIgM bound to EA; (0), rheumatoid IgM bound to E;( 0), normal IgM bound to EA.
3008 F. C. McDuiffie and H. W. Brum field
--L ~ ~ ~ ~

the buffer the cells were counted in a Packard autogammascintillation counter (Packard Instrument Co., DownersGrove, Ill.). Counts in tubes not containing cells were sub-tracted. These "glass binding" counts were always lessthan 5% of the counts added. The determinations of boundRF and C3 were made by the same technique except that thebuffer used for C3 did not contain EDTA. Appropriatecontrols for nonspecific binding by E, EA, or EAC1,4 wereperformed as well as controls of normal, nonimmune rabbitIgG, or human IgM. Binding of Cl to EACwas determinedby the Cl transfer test of Borsos and Rapp (11).
Phagocytosis. We employed the method of Gigli andNelson (14) in which sheep erythrocytes coated with anti-body and complement were phagocytized by rabbit neutro-phils. Nonphagocytized cells were measured by lysis withdistilled water since the erythrocytes within granulocyteswvere protected. A portion of appropriately coated sheeperythrocytes, 0.5 ml containing 2.5 x 102 cells in 0.075 mrelative NaCl barbital buffer, was incubated with 3.0 ml ofrabbit granulocytes containing 2-3 X 10' cells in Hanks'solution, and the mixture tumbled for 1 hr at 37°C. Then5.0 ml of distilled water was added, the cells centrifuged,and the OD of the supernates determined at 415 nm in aspectrophotometer. Appropriate confols of granulocytes aloneand uncoated erythrocytes plus granulocytes were preparedas well. The number of cells phagocytized was calculated as:
OD (test mixture) - OD (granulocytes)- OD(EA, spontaneous lysis) X 1
OD (EA + H20) - OD (EA, spontaneous lysis)
where1
FODof
t F-
RESULTSBinding of antibody and RF to cells. Figs. 1 and 2
show representative examples of the binding data ob-tained for rabbit IgG antibody to E and rheumatoid IgMto EA. The very low blank values obtained for normalIgG and IgM with E and EA respectively establish thevalidity of the method for the measurement of anti-sheep cell antibody and RF. In subsequent experimentsreference curves such as those shown were used in cal-culating the amount of unlabeled antibody or RF boundto cells.
Effect of IgM RF on phagocytosis. Fig. 3 shows theeffect of rheumatoid IgM on phagocytosis of cells coatedwith 195,000 molecules IgG antibody per cell, an amountwhich represents the optimum needed for complementfixation. Human IgM was added to the cells afterantibody and before Cl. Little inhibition of phagocytosiswas produced by normal IgM, even at a concentration of10 lAg protein N. Rheumatoid IgM on the other hand, ata concentration as low as 0.5 ltg N (resulting in 2500molecules bound per cell), produced striking inhibitionof phagocytosis to the base line level. When the amountof IgG antibody was reduced by 85% to 34,000 mole-cules bound per cell, a similar inhibitory effect was seen
II
I0
9
8
7
6
'k5a4
3
2 F-
_ EAC1,4.2.3
EAC1,4, 2
I I I 10 2 4 6 8 IO
pug N IgM added to EA 5 x107
FIGURE 3 Inhibition of phagocytosis by rheumatoid IgMof EA coated with 195,000 molecules IgG antibody per cell.Complement components used per cell were Cl, 100 EM;C2, 75 EM; C4, 100 EM; C3 (Bl,) 280 molecules. (0),normal IgM; (0), rheumatoid IgM.
as shown in Fig. 4. About twice as much rheumatoidIgM, 1.0 ,ug N, was needed to produce comparable inhi-bition, but this amount added actually resulted in thesame amount of bound RF, 2400 molecules per cell. Theresults shown were obtained with 9500 molecules of C3( per cell, a larger amount than was employed inFig. 3. However, very similar results were obtained whena smaller amount of C3 (B1c) was employed, 280 mole-cules per cell.
Role of complement components in inhibition of phago-cytosis (high input of IgG antibody). In order to de-termine whether the inhibitory effect of rheumatoid IgMon phagocytosis which we had observed was mediatedthrough an effect on the complement system, we ex-amined the fixation of complement components to cellscoated with 195,000 molecules of antibody per cell andtreated with rheumatoid IgM before addition of comple-ment components. Fig. 5 demonstrates that inhibition ofphagocytosis (Fig. 3) was almost exactly paralleled byinhibition of binding of 'I-labeled C3 (Fig. 5). To an-swer the question of whether inhibition of C3 bindingreflected a direct effect on C3 binding or inhibition ofan earlier step in the complement sequence, Cl transfertests on EAC1 made with normal and rheumatoid IgMwere carried out. Fig. 6 shows that when a high inputof rabbit IgG antibody was employed (195,000 moleculesper cell) Cl uptake was almost completely inhibited byas little as 0.5 ,g N of rheumatoid IgM, the same amountrequired to block complement-dependent phagocytosisand C3 uptake completely (Figs. 3 and 5).
Effect of Rheumatoid Factor on Complement-Mediated Phagocytosis
I am -- - -T 1. -
to
-
3009
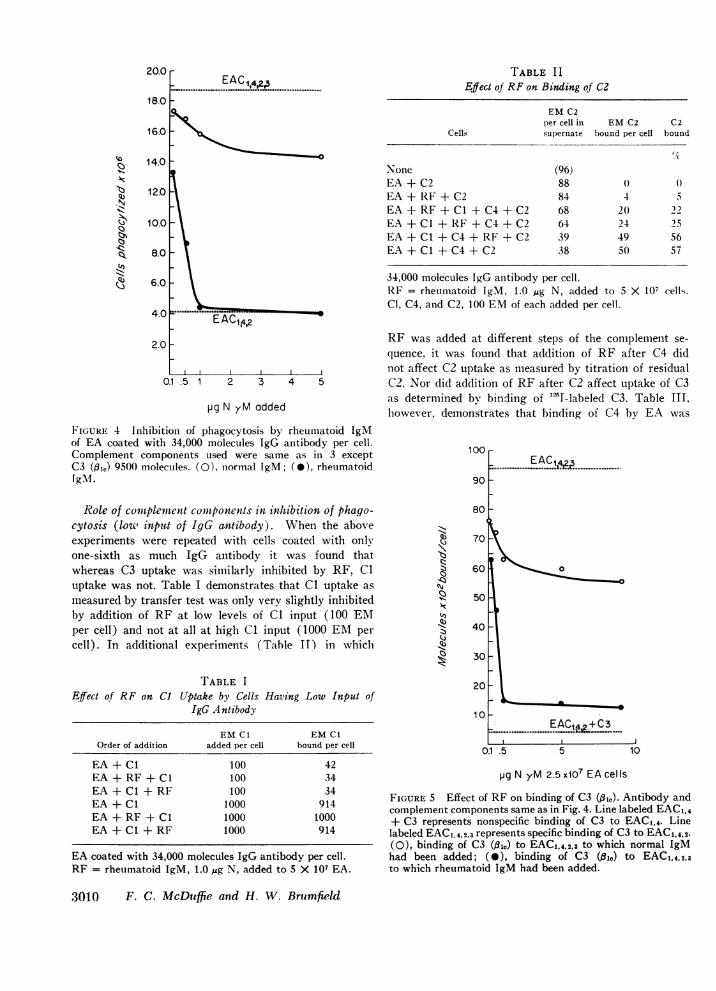
20.0
18.0
16.0
~IZi
('rI(.)
14.0
12.0
10.0
8.0
6.0
2.0
0.1 .5 1 2 3 4 5
pg N rM added
FIGURE 4 Inhibitioin of phagocytosis by rheumatoid IgMof EA coated with 34,000 molecules IgG antibody per cell.Complement components used were same as in 3 exceptC3 (#,,) 9500 molecules. (0), normal IgM; (0), rheumatoidIgMN.
Role of complemtent comnponents in inhibition of phago-cytosis (louw in.pzt of IgG antibody). When the aboveexperiments were repeated with cells coated with onlyone-sixth as much IgG antibody it was found thatwhereas C3 uptake was similarly inhibited by RF, Cluptake was not. Table I de.monstrates that Cl uptake asmeasured by transfer test was only very slightly inhibitedby addition of RF at low levels of Cl input (100 EMper cell) and not at all at high Cl input (1000 EMpercell). In additional experiments (Table II) in whicl
TABLE IEffect of RF on Cl Uptake by Cells Having Low Input of
IgG Antibody
EMCl EMClOrder of addition added per cell hound per cell
EA + C1 100 42EA+RF+Cl 100 34EA+C1+RF 100 34EA + Cl 1000 914EA+RF+C1 1000 1000EA + C1 + RF 1000 914
EA coated with 34,000 molecules IgG antibody per cell.RF = rheumatoid IgM, 1.0 ,ug N, added to 5 X 107 EA.
TABLE I IEffect of RF on Binding of CZ
EMC2per cell in EMC2 C2
Cells supernate bound per cell bound
CkNone (96)EA + C2 88 0 0EA+IF +C2 84 4 5EA + RF + C1 + C4 + C2 68 20 2EA + C1+ RF + C4 + C2 64 24 25EA + C1 + C4 + RF + C2 39 49 56EA + C1 + C4 + C2 38 50 57
34,000 molecules IgG anitibody per cell.RF = rheumatoid IgM, 1.0 ,ug N, added to 5 X 10( cells.Cl, C4, and C2, 100 EMof each added per cell.
RF was added at different steps of the complemiient se-quence, it was found that addition of RF after C4 didnot affect C2 uptake as measured by titration of residualC2. Nor did addition of RF after C2 affect uptake of C3as determined by binding of "I-labeled C3. Table III,however, demonstrates that binding of C4 by EA was
100
90
80
rz
CZj
0
(\3
0~
70
60
50
40
30
20
10
pg N yM 2.5 X107 EA cel Is
FIGURE 5 Effect of RF on binding of C3 ( Antibody andcomplement components same as in Fig. 4. Line labeled EAC1.4+ C3 represents nonspecific binding of C3 to EAC1,4. Linelabeled EAC1,42.,3 represents specific binding of C3 to EAC1.4.2.(0), binding of C3 (03(,) to EAC1.4,2.3 to which normal IgMhad been added; (0), binding of C3 (#,(,) to EAC1,4,.,3to which rheumatoid IgM had been added.
3010 F. C. McDuffie and H. W. Bruimfield

impaired, even when RF was added after Cl. Thus al-though EAC, not treated with IgM, bound 66 moleculesC4 per cell, addition of RF after Cl reduced C4 bindingto 26 molecules per cell, a result suggesting interferenceby RF of binding of C4 to antibody.
Effect of human Cl. The above experiments wereperformed with partially purified guinea pig Cl as asource of the first component of complement. Zvaiflerand Bloch (26) found that RF inhibited uptake of guineapig complement at 40 and 37°C yet was able to fix humancomplement at 370 but not 4°C. We therefore examinedthe effect of RF on phagocytosis of cells first coated withhuman Cl at 370C. The results of Table IV show thatRF added before or after human Cl inhibited phagocy-tosis of such cells about equally as well as cells madewith guinea pig Cl (compare Fig. 4).
DISCUSSIONPhagocytosis of microorganisms, erythrocytes, or otherparticles by polymorphonuclear granulocytes is markedlyenhanced by the addition of antibody and complement.Indeed whether antibody alone in the absence of com-plement is capable of promoting phagocytosis is notentirely certain. Although a specific binding site for IgGantibodies has been clearly demonstrated on the surfaceof the macrophage (15) it has been more difficult toestablish the presence of such a site on granulocytes.Messner and Jelinek have presented evidence for such aspecific receptor site for the Fc portion of vG-globulinon the surface of human neutrophils (16). However theseauthors were only able to achieve binding of Rh-positiveerythrocytes to neutrophils by using a particular hyper-immune complement-fixing anti-Rh (Ripley) and notwith ordinary anti-Rh sera. Even when purified IgGand thoroughly washed granulocytes are used in phago-cytic assays it is difficult to exclude participation of smallamounts of complement bound to leukocytes. Golden andMcDuffie (17), for example, found that inhibition oflupus erythematosus cell formation could be achieved withanti-complement antiserum in a phagocytic system inwhich complement could not be detected by ordinarymeans. Although factors other than complement may beresponsible for the enhancing effect of fresh serum onphagocytosis none has as yet been unequivocally identi-fied. Gigli and Nelson (14) showed that addition of Cl,C2, or C4 had no effect on phagocytosis whereas additionof C3 produced a marked enhancement, not affected byaddition of the subsequently reacting components, C5-C9.
Thus it is not surprising that IgM RF which blockscomplefnemnt fixation by IgG antibodies is able to inhibitphagocytosis of particles coated with antibody and com-plement. Messner, Lasdal, Quie, and Williams (5) havepreviously shown that IgM RF isolated from patientswith subacute bacterial endocarditis (SBE) was able to
:zz93ci
'tI Q.lc-
.k-
Z31U)-ttil
-91)i
EAC,.________.
N-o
0.5pg N IgM
1.0
FIGURE 6 Effect of RF on binding of Cl to EA coated with195,000 molecules of antibody per cell. 100 EMCl added percell. (0), normal IgM; (0), rheumatoid IgM.
inhibit ingestion by polymorphonuclear leukocytes ofbacteria coated with IgG opsonin. If RF was added toantibody-coated bacteria before addition of fresh normalserum the effect was more pronounced than if it wasadded after fresh serum. The present experiments clearlydemonstrate that this effect is mediated through inhibi-tion of Cl and C4 uptake by antigen-antibody complexes.Presumably the inhibition of Cl uptake is the result ofcompetition between RF and Clq for the same site or ad-jacent sites on the Fc portion of IgG (18, 19).
We cannot explain why inhibition of Cl fixation byRF was so much more pronounced when cells heavilycoated with antibody were used. Aho, Harboe, and Lei-kola (20) found that the agglutination by RF of sheepcells coated with small amounts of IgG could be inhibited
TABLE I IIEffect of RF on C4 Uptake by Cells Having Low Input of
IgG Antibody
100 EMC4 added per cell
EMC4per cell in EMC4 C4
Order of addition supernate bound per cell bound
EA 82 0 0EA+C4 82 0 0EA + RF+C4 82 0 0EA+ C1 + C4 16 66 80EA + C1 + RF + C4 56 26 32EA + RF + C1 + C4 74 8 10
EA coated with 34,000 molecules IgG antibody per cell.RF = rheumatoid IgM, 1.0 ,ug N, added to 5 X 107 cells.Cl, 100 EM per cell. 100 EM C4 added per cell and aftercentrifugation residual C4 determined in the supernate byhemolytic titration. Bound C4 determined by subtraction ofappropriate controls.
Efect of Rheumatoid Factor on Complement-Mediated Phagocytosis 3011

TABLE IVEffect of RF on Phagocytosis (Human C1)
Cellsplhagocytized Cells
Order of addition X 106 phagocytized
EAC1.4. 2 1.25 5EACI, 4.2.3 12.0 48EA + N IgM + C1,4,2,3 9.3 37EAC1 + N IgM + C4,2,3 9.8 39EA + RF + C1,4,2,3 2.5 10EAC1 + RF + C4,2,3 7.0 28
34,000 molecules IgG antibody per cell. EMI added per cellwere 100 Cl, 75 C4, and 50 C2. 200 molecules C3 (i,3l) addedper cell.1.0 ,ug N RF or normal IgM per cell.
by native IgG in serum, but agglutination of heavilycoated cells could only be inhibited with aggregated IgGsuch as that in an immune precipitate. Whether thegreater affinity of RF to heavily coated cells reflectsonly the involvement of more binding sites (out of apossible total of five on the IgM molecule) or a con-figurational change occurring on the IgG antibody afterits reaction with red cell antigens cannot be answered.Clq is also multivalent, and Cl binds to EA morestrongly as the number of IgG molecules on the cell sur-face is increased (21). A similar mechanism of competi-tion may account for the inhibition of C4 uptake whichwe observed at lower levels of antibody input, since C4is known to bind to an as yet unidentified site on anti-body (22). Since C2 binds directly to C4 to form a com-plex which acts in turn to activate or bind C3, failure ofRF to inhibit binding of C2 when added after C4 is notsurprising (23). Although C3 or j%c may bind directlyto immunoglobulin (24) it may also attach to the surfaceof sensitized erythrocytes (25), a phenomenon whichmay explain failure of RF to inhibit C3 binding whenadded to EAC1,4,a.
IgM RF has previously been shown to interfere withfixation of complement by IgG antibodies (26, 27).Zvaifler and Schur (28) found that the complex formedbetween RF and mercaptoethanol-treated aggregated IgG(which reacts with RFbut does not itself fix complement)did not consume human or guinea pig complement whenthe complex was added to fresh rheumatoid serum. Largeamounts (> 1.0 mg) or preformed complexes of RF andmercaptoethanol-treated IgG were, however, able to fixhuman but not guinea pig complement. In our experimentswe found RF equally inhibited phagocvtosis of cellscoated with either human or guinea pig Cl. Schmid,Roitt, and Rocha (29) found tlhat, whereas RF inhibitedcomplement fixation of sheep cells optimally sensitizedwith IgG antibody, some complement fixation by RF
occurred when cells coated with suboptimal amounts ofantibody were used. In the present experiments, how-ever, reduction in the amount of IgG antibody used tocoat sheep cells to 15% of the optimal amount did notsignificantly reduce the complement uptake nor theamount of phagocytosis observed. The complement-fixingactivity of RF must be Nweak at best, even under care-fully controlled conditions.
The mechanism by which RF inhibits complement fixa-tionl remainis obscure. No significant structural differ-ences between IgMl RF and other human IgM globulinshave so far been detected (30, 31). Yet IgAM antibodiesof most species fix complement and promote phagocyto-sis more efficiently than IgG antibodies (32-34). AMac-Kenzie, Creevy, and Heh (35) have reported thlat somieIgM myeloma globulins bind Clq to a muclh lesser extentthan normal. Possibly RF contains a high proportion ofsuch antibodies since we found that inihibitioin of CI fixa-tion was virtually conmplete wvhen optimally sensitizeed cellswere pretreated with RF. The explaniationi for the inllibi-tory effect on C4 binding is not apparent from our (lataaltlhough it lhas been founid that IgAM antibodies are lessefficient thani IgG antibodies in the titration of C4 (36).Human Igl\I antinuclear antibodies in our experiencealso appear to be deficient in their ability to fix comple-menit and to promote phagocytosis (37). Most of theprevious work on inhibition of complement fixation byRF has employed inhibitioni of immune hemolysis as anindclicator so that no one has excluded the possibility thatRF may fix complement at a position sufficiently distantfrom the cell surface so that, in spite of activation of thecomponents, cell lysis may not occur. The present ex-periments clearly show that activation and binding of thefirst four steps of the complement sequence are actuallvp)revented by RF.
These results imply that circulating IgM RF does notplay a significant role in the inflammation of vessels andtissues in RA, since the phlogistic effects of IgAM anti-bodies are probably mediated largely througlh the com-plement system. Thouglh immune complexes appear to beresponsible for the low levels of complement in synovialfluid (38), pleural fluid (39), and serum2 of rheumatoidpatients, IgM RF itself does not seem to be the majorantibody in these complexes. WVinchester, Agnello, andKunkel (38) fotund only IgG and not IgAl in the highnmolecular weight material in rheumatoid synovial fluidwhich they were able to relate to complement depletion.Horwitz, Garrett, and Davis (40) foumd that rheama-toid IgM\ inhibited rather than enlhanced release of com-plement-derived chemotactic activity. In experimentswith passive immune synovitis in the rabbit knee jointwve have not found anly consistent enhancing effect of
aHunder, G. G., and F. C. McDuffie. Hypocomplementemiain rheumatoid arthritis. Unpublished observations.
3012 F. C. McDuffie and H. W. Brumfield

RF on the inflammatory reaction, slight inhibition orno effect being the usual result.3 On the other hand Tesar,Schmid, and Suarey (41) have pointed out that RF varyin avidity for antigen and that those of high avidity fixcomplement better than those of low avidity. Thus it ispossible that a small proportion of RF of particularlyhigh avidity may be contributing to tissue damage.
ACKNOWLEDGMENTSThis study was supported by the National Institute of Ar-thritis and Metabolic Diseases, Research Grant AM11630.
REFERENCES1. Hollander, J. L., D. J. McCarty, Jr., G. Astorga, and E.
Castro-Murillo. 1965. Studies on the pathogenesis ofrheumatoid joint inflammation. I. The "RA cell" anda working hypothesis. Ann. Intern. Med. 62: 271.
2. Restifo, R. A., A. J. Lussier, A. J. Rawson, J. H.Rockey, and J. L. Hollander. 1965. Studies on the patho-genesis of rheumatoid joint inflammation. III. The ex-perimental production of arthritis by the intra-articularinjection of purified 7S gamma globulin. Ann. Intern.Med. 62: 285.
3. Sliwinski, A. J., and N. J. Zvaifler. 1969. The removalof aggregated and nonaggregated autologous gammaglobulin from rheumatoid joints. Arthritis Rheum. 12:504.
4. Davis, J. S., IV. 1966. A hypothetical common mecha-nism in systemic lupus erythematosus and rheumatoidarthritis. Arthritis Rheum. 9: 631.
5. Messner, R. P., T. Ladal, P. G. Quie, and R. C. Wil-liams, Jr. 1968. Serum opsonin, bacteria, and polymor-phonuclear leukocyte interactions in subacute bacterialendocarditis. Anti-'y-globulin factors and their interactionwith specific opsonins. J. Clin. Invest. 47: 1109.
6. Nelson, R. A., J. Jensen, I. Gigli, and N. Tamura. 1966.Methods for the separation, purification and measure-ment of nine components of hemolytic complement inguinea-pig serum. Immunochemistry. 3: 111.
7. Miiller-Eberhard, H. J., U. Nilsson, and T. Aronsson.1960. Isolation and characterization of two p1-glycopro-teins of human serum. J. Exp. Med. 111: 201.
8. Nilsson, U. R., and H. J. Miuller-Eberhard. 1965. Iso-lation of pi,t-globulin from human serum and its charac-terization as the fifth component of complement. J. Exp.Med. 122: 277.
9. Miiller-Eberhard, H. J., and C. E. Biro. 1963. Isolationand description of the fourth component of human com-plement. J. Exp. Med. 118: 447.
10. Mayer, M. M. 1961. Complement and complement fixa-tion. In Kabat and Mayer's Experimental Immunochem-istry. Charles C. Thomas, Publisher, Springfield, Ill.2nd edition. 133.
11. Borsos, T., and H. J. Rapp. 1965. Hemolysin titrationbased on fixation of the activated first component ofcomplement: evidence that one molecule of hemolysinsuffices to sensitize an erythrocyte. J. Immunol. 95: 559.
12. Nelson, R. A., Jr. 1953. The immune-adherence phe-nomenon. An immunologically specific reaction betweenmicroorganisms and erythrocytes leading to enhancedphagocytosis. Science (Wash. D. C.). 118: 733.
3Ramirez-Peredo, J., and F. C. McDuffie. Unpublishedobservations.
13. McConahey, P. J., and F. J. Dixon. 1966. A methodof trace iodination of proteins for immunologic studies.Imtt. Arch. Allergy Appl. Immunol. 29: 185.
14. Gigli, I., and R. A. Nelson, Jr. 1968. Complement de-pendent immune phagocytosis. I. Requirements for C'l,C'4, C'2 and C'3. Exp. Cell Res. 51: 45.
15. Berken, A., and B. Benacerraf. 1966. Properties of anti-bodies cytophilic for macrophages. J. Exp. Med. 123:119.
16. Messner, R. P., and J. Jelinek. 1970. Receptors forhuman yG globulin on human neutrophils. J. Clin. In-vest. 49: 2165.
17. Golden, H. E., and F. C. McDuffie. 1967. Role of lupuserythematosus factor and accessory serum factors inproduction of extracellular nuclear material. Ann. In-tern. Med. 67: 780.
18. Schubart, A. F., and R. W. Ewald. 1965. Kinetic studiesof serum complement components creating inhibition ofthe agglutination activating activity of the rheumatoidfactor. Arthritis Rheum. 8: 467. (Abstr.)
19. Ewald, R. W., and A. F. Schubart. 1966. Agglutinatingactivity of the complement component C'lq in the F-IIlatex fixation test. J. Immunol. 97: 100.
20. Aho, K., M. Harboe, and J. Leikola. 1964. Studies ofthe antibody nature of the rheumatoid factor. Reactionof the rheumatoid factor with sheep erythrocytes sensi-tized with human anti-sheep cell antibodies and with 0Rh positive cells sensitized with incomplete anti-Rh anti-bodies. Immunology. 7: 403.
21. Linscott, W. D. 1970. Complement fixation: the effectsof IgG and IgM antibody concentration of Cl-bindingaffinity. J. Immunol. 105: 1013.
22. Willoughby, W. F., and M. M. Mayer. 1965. Antibody-complement complexes. Science (Wash. D. C.). 150:907.
23. Miiller-Eberhard, H. J., M. J. Polley, and M. A. Cal-cott. 1967. Formation and functional significance ofa molecular complex derived from the second and thefourth component of human complement. J. Exp. Med.125: 359.
24. Leddy, J. P., R. F. Bakemeier, and J. H. Vaughan.1965. Fixation of complement components to autoanti-body eluted from human RBC. J. Clin. Invest. 44: 1066.(Abstr.)
25. Miiller-Eberhard, H. J., A. P. Dalmasso, and M. A.Calcott. 1966. The reaction mechanism of p,10-globulin(C'3) in immune hemolysis. J. Exp. Med. 123: 33.
26. Zvaifler, N. J., and K. J. Bloch. 1962. Rheumatoid fac-tor-an inhibitor of the complement fixation reaction.Arthritis Rheum. 5: 127. (Abstr.)
27. Heimer, R., F. M. Levin, and M. F. Kahn. 1963. In-hibition of complement fixation by human serum. II.The activity of a y--m globulin and rheumatoid factoron complement fixation reactions. J. Immunol. 91: 866.
28. Zvaifler, N. J., and P. Schur. 1968. Reactions of aggre-gated mercaptoethanol treated gamma globulin withrheumatoid factor-precipitin and complement fixationstudies. Arthritis Rheum. 11: 523.
29. Schmid, F. R., I. M. Roitt, and M. J. Rocha. 1970.Complement fixation by a two-component antibody sys-tem: immunoglobulin G and immunoglobulin M anti-globulin (rheumatoid factor). J. Exp. Med. 132: 673.
30. Kunkel, H. G., E. C. Franklin, and H. J. Muller-Eber-
Effect of Rheumatoid Factor on Complement-Mediated Phagocytosis 3013

hard. 1959. Studies on the isolation and characterizationof the "rheumatoid factor." J. Clin. Invest. 38: 424.
31. Heimer, R., K. R. Woods, and R. L. Engle, Jr. 1962.Amino acid analyses of rheumatoid factors and normalgammaglobulins. Proc. Soc. Exp. Biol. Med. 110: 496.
32. Onoue, K., N. Tanigaki, Y. Yagi, and D. Pressman.1965. IgM and IgG anti-hapten antibody: hemolytichemagglutinating and precipitating activity. Proc. Soc.Exp. Biol. AMed. 120: 340.
33. Robbins, J. B., K. Kenny, and E. Suter. 1965. The iso-lation and biological activities of rabbit yM- and 'yG-anti-Salmonella typhimurium antibodies. J. Exp. Med.122: 385.
34. Rowley, D., and K. J. Turner. 1966. Number of mole-cules of antibody required to promote phagocytosis ofone bacterium. Nature (Lond.). 210: 496.
35. MacKenzie, M. R., N. Creevy, and M. Heh. 1971. Theinteraction of human IgM and Clq. J. Inimunol. 106:65.
36. Frank, M. M., and T. Gaither. 1970. Evidence that rab-bit yG-haemolysin is capable of utilizing guinea-pig
complement more efficiently than rabbit 'yM-haemolysin.Immunology. 19: 975.
37. Blondin, C., aid F. C. McDuffie. 1970. Role of IgGand IgM antinuclear antibodies in formation of lupuserythematosus cells and extracellular material. ArthritisRheum. 13: 786.
38. Winchester, R. J., V. Agnello, and H. G. Kunkel. 1970.Gammaglobulin complexes in synovial fluids of patientswith rheumatoid arthritis. Partial characterization andrelationship to lowered complement levels. Clin. Exp.Immunol. 6: 689.
39. Hunder, G. G., F. C. McDuffie, and N. G. G. Hepper.1972. Pleural fluid complement in systemic lupus erythe-matosus and rheumatoid arthritis. Antn. Intern. Med.76: 357.
40. Horwitz, D. A., M. A. Garrett, and J. S. Davis, IV.1971. Modification of leukocyte chemotaxis by rheuma-toid factor. Arthritis Rheum. 14: 388. (Abstr.)
41. Tesar, J. T., F. R. Schmid, and A. Suarey. 1972. Asso-ciation of avidity of rheumatoid factor (RF) with com-plement fixation. Arthritis Rheumn. 15: 128. (Abstr.)
:3014 F. C. McDuffie and H. W. Brumfield















![Complement-Mediated Neutralization of Dengue Virus ... · fever/dengue shock syndrome [DHF/DSS]) (2). Although the pathogenesisofDENVinfectionremainscontroversial,antibody-dependent](https://static.fdocuments.in/doc/165x107/5cb4737a88c99310568bc0bf/complement-mediated-neutralization-of-dengue-virus-feverdengue-shock-syndrome.jpg)


