
Ecosystem Modelling Page 1 - meetings.pices.int · Ecosystem Modelling Page 1 Vladimir I. Zvalinsky...
19
Ec o syste m Mo de lling Page 1 Vladimir I. Zvalinsky PICES 2005 Em a il: The ecosystem parameters and its stability. Theoretical consideration V.I.Il'ichev Pacific Oceanological Institute, Far Eastern Branch, Russian Academy of Sciences, Vladivostok, 690041, Russia [email protected]
Transcript of Ecosystem Modelling Page 1 - meetings.pices.int · Ecosystem Modelling Page 1 Vladimir I. Zvalinsky...

Ecosyste m Mode lling Pa ge 1
Vladimir I. Zvalinsky
PICES 2005
Ema il:
The ecosystem parameters and its stability. Theoretical consideration
V.I.Il'ichev Pacific Oceanological Institute, Far Eastern Branch, Russian Academy of Sciences,
Vladivostok, 690041, Russia

Ecosyste m Mode lling Page 2
USUALEc osyste m mod e lling oc curs a s fo llow:- Mo de lling o f e a c h se pa ra te trop hic link;- Co nne ction o f trophic link be twe en e a c h othe r;- Attributing o f some laws for connec tio n o f links.
NEMURO MODEL 2000 is the e xa mple o f such proc edure .In result we ha ve no t a bso lute ly e co system, but c onnec ted amo ng the mse lve s diffe re nt tro phic links.At such ap proa ch e co system pa rame te rs, a s a whole , may not be re ve a le d.
PICES 2005

Ec osyste m mode lling Pa ge 3
ANOTHER WAY
- At first the e c osyste m c once ptua l mo de l is de signe d;- Then the la w o f transition substanc e from o ne trophic link to anothe r (or the c onc eptua l mod e l o f sep ara te link) is de sig ne d;- Formula tion o f full se t of equations;- Red uce the se t o f e qua tio ns to d ime nsionless fo rm a nd a na lysis.
As an example the six compartment e cosyste m mode l is consid ere d.
PICES 2005

Sedi
men
tatio
nLight
In
ZL
I
Mortality
Egestion
Inorganic matter
incorporationG
razing
Grazing
Respiration
Mineralisation
Ph
Exchange
E-cycleTransition
law
Transition law
Transition law
Tran
sition
law
Transition law
Conceptual model of ecosystem as ecological cycleEcosystem represents by 6 compartments. Four of them are livinglinks (Phy, SZ, LZ and Bacterial) and two of them are non living ones(inorganic matter and detritus).
Page 4
PICES 2005
Det
ZS
B
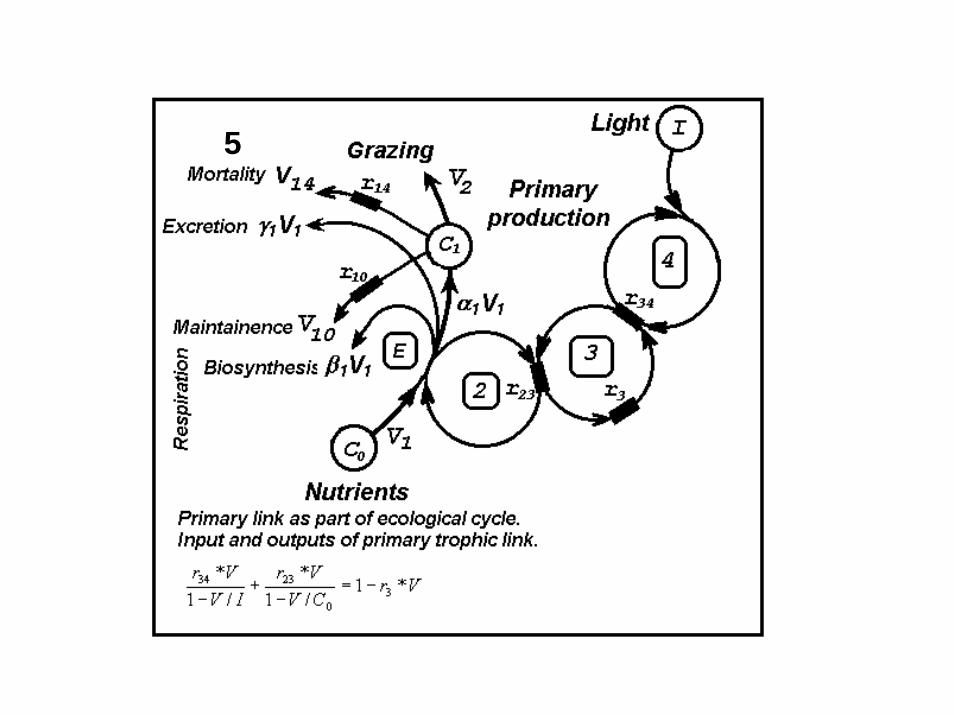
5

6

C5Solar light
Herbi-vores
Carnivores
Bacte-rium
Detritus
Phyto-plank- ton
Primaryproduction4
3
C1
C2C
C
C0
I
Respiration
Sedimentation
Vertical
exchange
MortalityEgestion
PP
ZS
ZL
B
rEB rEP
rES
rEL
Vij
C5
Ecosystem as a system of coupled cyclical processesEcosystem parameters
rP2rP1
rS1rS2
rB2
rL2rL1
rB1
rP
B2rrB1+R =B
rS2rS1 +R =ZS
rL2rL1 +R =ZL
r rP2 PrP1 + +R =PP
E
RBRZS R ZLRPP, , ,
C +0C =tot C +3 C5C +1 C +2 C +4
- total resistance of each trophic link
- total concentration of limiting nutrient
rEPrEB rES rEL, , , - coupling resistance between ecosystem cycle and each trophic link
7

There are three Type of Ecosystem Parameters
RBRZS R ZLRPP, , ,
E
mmR = P /Pij P ij
- total resistance of corresponding trophic link, which is inversely proportional to maximum specific rate
- total concentration of limiting nutrient
- coupling resistance between ecosystem cycle and each trophic link
[Ctot]=[C0]+[C1]+[C2]+[C3]+[C4]+[C5]
1.
2.
3.
KCi = Pim/{kCi*[ECi]} - substrate constant (Michaelis
constant?) of corresponding link
rEP, rES, rEL, rEB
rEi = KCi/Σ[Ci] = KCI/[Ctot]
Page 8
PICES 2005
‘
The task is: How the Ecosystem stability depends on ecosystem parameters

Page 9
PICES 2005
0 2 4 6Substrate concentration, S=[S]/KC
0
0.2
0.4
0.6
0.8
1
Proc
ess
rela
tive
rate
, V=P
/Pm
M-M
g=0.80.95BL
tanh
exp
KC
Different kinetic curves for description of biological process what most often are using.Change of the factor hyperbola non-rectangularity "g" from "0.0" up to "1.0" changes the curve shape from "M-M" variant up to "Blackman" one.
g=0.0g=0.
999
0 2 4 6Substrate concentration, S=[S]/KC
0
0.2
0.4
0.6
0.8
1
M-M
0.95BL
KC
We used two kinetic curves: M-M curve (g=0.0) and curve with factor g=0.95 (which close to BL and close to experimental kinetic curves.
g=0.0g=0.
999

Page 10Parameters combination used for execution of
ecosystem behavior
Parameters I II III IV
RPP 1.0 1.0 1.0 1.0
RZS 2.0 2.0 4.0 4.0
RZL 4.0 4.0 8.0 8.0
Factor "g" 0.0 0.0 0.95 0.95
rEP=rES=rEL V a r i a b l e s

Time course of primary link at different rE Page 11
g=0.0 (M-M kinetics); rPP=1; rZS=2; rZL=4
0 100 200 300 400 500Time, in 1/Pm (=days)
0
0.2
0.4
0.6
0.8Co
ncen
trat
ion
C Ph
Nemuro, g=0.0 (M-M); rPP=1; rZS=2; rZL=4
rE=1
rE=0.5
rE=2
rE=KC/[Ctot]L
Decreasing of rE (increasing of [Ctot]L) causes oscillations
rE=0.4
At Ctot=2*KC (rE=0.5) - undamped Osc.

The same time course, but g=0.95 Page 12(BL kinetics); at rE < 0.5 - undamped oscillation At such kinetics of each process less amplitude oscillations
0 100 200 300 400 500Time, in 1/Pm (=days)
0
0.2
0.4
0.6
0.8
Conc
entr
atio
n C P
h
rE=1
rE=0.3
rE=0.5
rE=2
rE=0.19
rE=0.17rE=KC/[Ctot]L

Time course of primary link at different rE Page 13g=0.0 (M-M kinetics); rPP=1; but rZS=4; rZL=8
More amlitude oscillations, more period
0 200 400 600 800 1000Time, in 1/Pm (=days)
0
0.2
0.4
0.6
0.8
Conc
entr
atio
n C P
h
rE=1
rE=0.5
rE=2
rE=0.4
At Ctot=KC (rE=1.0) - undamped Osc.

The same time course, but g=0.95 Page 14(BL kinetics); at rE < 0.5 - undamped oscillation
At such kinetics of each process less amplitude oscillations
0 200 400 600 800 1000Time, in 1/Pm (=days)
0
0.2
0.4
0.6
0.8
Conc
entr
atio
n C P
h
rE=0.5
rE=2
rE=1(4:8)
rE=0.37

Page 15
• Michaelis-Menten not steep kinetics of each biological process couses higher amplitude oscillation at less total concentration ecosystem limiting nutrient
• Steep kinetics (close to Blackman’s broken line) couses less amplitude oscillation at higher total concentration ecosystem limiting nutrient
• Higher difference between specific rate growth of neighboring trophic links couses higher amplitude of oscillations with higher period

Very high period of oscillations shows the non biological nature of oscillations. The Czs equal to 0.0 for a long time? We think that this is impossible in Nature. It must be some Threshold for min C. We introduce threshold.
Page 16
0 100 200 300 400 500Time, in 1/Pm (=days)
0
0.2
0.4
0.6
0.8
Con
cent
ratio
n C
Ph, C
ZS, C
ZL
rE=0.5 CPh
CZS
CZL
It is very high period of oscillations
g=0.0 (M-M); rPP=1; rZS=2; rZL=4

Dinamics of the same ecosystem at introducing Page 17of Threshold = 0.1*rE = 0.1*Kc. No oscillations. Real time relaxation.
More steep kinetic curve (g=0.95) - less time relaxation
0 40 80 120 160Time, in 1/Pm (=days)
0
0.2
0.4
0.6
Con
cent
ratio
n C
Ph, C
ZS, C
ZL
CPh
CZS
g=0.0
g=0.95
g=0.95 g=0.0

Whith threshold of Ci at M-M kinetic curve (g=0.0) Page 18it is possible to find special ratio of rEi, when oscillations take place: rEP=0.7; rES=0.13; rEL=0.9. But at steep kinetic curve (g=0.95) there are no oscillation at any rEi.
0 40 80 120 160 200Time, in 1/Pm (=days)
0
0.2
0.4
Conc
entr
atio
n C P
h
Tresh=0.05
g=0.
95 g=0.
0
g=0.
0Tr
esh=
0.1

Page 19Conclusion
• In order to construct the predictive ecosystem model we must to understand the essential inner attributes of marine ecosystem.
• For the ecosystem modelling it is necessary to reveal mathematical couses of ecosystem stability/unstability from the biological and ecological ones and divide them from each other.
• On PICES 2001 Ken Denman said: «Stability of simple Box Model is ‘Fragile’»
• Our knowlidge about marine ecosystem is not less ‘Fragile’


















