Distribution and hormone regulation of estrogen receptor immunoreactive cells in the hippocampus of...
10
Distribution and Hormone Regulation of Estrogen Receptor Immunoreactive Cells in the Hippocampus of Male and Female Rats N.G. WEILAND, 1 * C. ORIKASA, 2 S. HAYASHI, 3 AND B.S. MCEWEN 1 1 Laboratory of Neuroendocrinology, Rockefeller University, NewYork, NewYork 10021 2 Department of Physiology, Nippon Medical School, Tokyo 113, Japan 3 Department ofAnatomy and Embryology, Tokyo Metropolitan Institute for Neuroscience, Tokyo 183, Japan ABSTRACT Estrogen regulates the synaptic plasticity and physiology of the hippocampus as well as learning behaviors that are mediated by the hippocampus. The density of dendritic spines and synapses, the number of N-methyl-D-aspartate (NMDA) binding sites, the levels of NMDA receptor subunit NR1 protein, muscimol binding to the g-amino butyric acid (GABA) A receptor, and levels of glutamic acid decarboxylase message in the CA1 region of the hippocampus are altered with estrogen treatment. In addition, some of these parameters exhibit sex differences in their response to estrogen treatment. To establish that estrogen can have a direct effect on the hippocampus and to determine whether or not sex differences in estrogen responsiveness are due to sex differences in estrogen receptor (ER) levels, we used immunocytochemistry with the AS409 antibody to map the location of ER-immunoreactive (ER-ir) cells in the hippocampus of male and female rats. We found that (1) the ERs appear to be in interneurons rather than pyramidal or granule cell neurons, (2) ER-ir cells are located in greatest concentration in the hilus of the dentate gyrus and the stratum radiatum of the CA1 region, (3) the density of ER-ir cells exhibits a rostral to caudal gradient in the hilus and the CA1 regions, (4) there are no sex differences in either the number or immunostaining intensity of ER-ir cells in the hippocampus, (5) the ER levels are down-regulated by estrogen in both male and female rats, and (6) the mean intensity of staining for the ER-ir cells in the hippocampus is about 25% of that in the ER-ir cells of the hypothalamus. From this, we can conclude that estrogen can have a direct effect on hippocampal neurons and that any sex differences in estrogen responsiveness is due to something other than sex differences in ER levels or function in the hippocampus. J. Comp. Neurol. 388:603–612, 1997. r 1997 Wiley-Liss, Inc. Indexing terms: dentate gyrus; CA1; cortex; progesterone receptor; synaptic plasticity Estrogen has been shown to influence several processes associated with hippocampal function, including altering certain types of learning behavior (Phillips and Sherwin, 1992a, 1992b; O’Neil et al., 1996), decreasing the threshold to seizure activity (Terasawa and Timiras, 1968; Back- strom, 1976), and altering the course of Alzheimer’s dis- ease (Fillit et al., 1986). Prior work has established several possible mechanisms for the effect of estrogen on the hippocampus. Estrogen increases the density of dendritic spines (Gould et al., 1990) and synapses (Woolley and McEwen, 1992) on CA1 pyramidal neurons, and this increase is correlated with an increase in the synthesis of the N-methyl-D-aspartate (NMDA) receptor subunit NR1 protein (Gazzaley et al., 1996) resulting in an increase in agonist binding to the NMDA receptor (Weiland, 1992a). Evidence suggests that the effect of estrogen on CA1 pyramidal cells is indirect. The estrogen induction of dendritic spines requires activation of the NMDA receptor Grant sponsor: National Institutes of Health; Grant numbers: NS30105, NS07080. *Correspondence to: Dr. Nancy G. Weiland, Laboratory of Neuroendocri- nology, Rockefeller University, 1230 York Avenue, New York, NY 10021. E-mail: [email protected] Received 18 December 1996; Revised 20 June 1997; Accepted 24 June 1997 THE JOURNAL OF COMPARATIVE NEUROLOGY 388:603–612 (1997) r 1997 WILEY-LISS, INC.
Transcript of Distribution and hormone regulation of estrogen receptor immunoreactive cells in the hippocampus of...

Distribution and Hormone Regulationof Estrogen Receptor ImmunoreactiveCells in the Hippocampus of Male and
Female Rats
N.G. WEILAND,1* C. ORIKASA,2 S. HAYASHI,3 AND B.S. MCEWEN1
1Laboratory of Neuroendocrinology, Rockefeller University, New York, New York 100212Department of Physiology, Nippon Medical School, Tokyo 113, Japan
3Department of Anatomy and Embryology, Tokyo Metropolitan Institute for Neuroscience,Tokyo 183, Japan
ABSTRACTEstrogen regulates the synaptic plasticity and physiology of the hippocampus as well as
learning behaviors that are mediated by the hippocampus. The density of dendritic spines andsynapses, the number of N-methyl-D-aspartate (NMDA) binding sites, the levels of NMDAreceptor subunit NR1 protein, muscimol binding to the g-amino butyric acid (GABA)Areceptor, and levels of glutamic acid decarboxylase message in the CA1 region of thehippocampus are altered with estrogen treatment. In addition, some of these parametersexhibit sex differences in their response to estrogen treatment. To establish that estrogen canhave a direct effect on the hippocampus and to determine whether or not sex differences inestrogen responsiveness are due to sex differences in estrogen receptor (ER) levels, we usedimmunocytochemistry with the AS409 antibody to map the location of ER-immunoreactive(ER-ir) cells in the hippocampus of male and female rats. We found that (1) the ERs appear tobe in interneurons rather than pyramidal or granule cell neurons, (2) ER-ir cells are located ingreatest concentration in the hilus of the dentate gyrus and the stratum radiatum of the CA1region, (3) the density of ER-ir cells exhibits a rostral to caudal gradient in the hilus and theCA1 regions, (4) there are no sex differences in either the number or immunostaining intensityof ER-ir cells in the hippocampus, (5) the ER levels are down-regulated by estrogen in bothmale and female rats, and (6) the mean intensity of staining for the ER-ir cells in thehippocampus is about 25% of that in the ER-ir cells of the hypothalamus. From this, we canconclude that estrogen can have a direct effect on hippocampal neurons and that any sexdifferences in estrogen responsiveness is due to something other than sex differences in ERlevels or function in the hippocampus. J. Comp. Neurol. 388:603–612, 1997.r 1997 Wiley-Liss, Inc.
Indexing terms: dentate gyrus; CA1; cortex; progesterone receptor; synaptic plasticity
Estrogen has been shown to influence several processesassociated with hippocampal function, including alteringcertain types of learning behavior (Phillips and Sherwin,1992a, 1992b; O’Neil et al., 1996), decreasing the thresholdto seizure activity (Terasawa and Timiras, 1968; Back-strom, 1976), and altering the course of Alzheimer’s dis-ease (Fillit et al., 1986). Prior work has established severalpossible mechanisms for the effect of estrogen on thehippocampus. Estrogen increases the density of dendriticspines (Gould et al., 1990) and synapses (Woolley andMcEwen, 1992) on CA1 pyramidal neurons, and thisincrease is correlated with an increase in the synthesis ofthe N-methyl-D-aspartate (NMDA) receptor subunit NR1
protein (Gazzaley et al., 1996) resulting in an increase inagonist binding to the NMDA receptor (Weiland, 1992a).Evidence suggests that the effect of estrogen on CA1pyramidal cells is indirect. The estrogen induction ofdendritic spines requires activation of the NMDA receptor
Grant sponsor: National Institutes of Health; Grant numbers: NS30105,NS07080.
*Correspondence to: Dr. Nancy G. Weiland, Laboratory of Neuroendocri-nology, Rockefeller University, 1230 York Avenue, New York, NY 10021.E-mail: [email protected]
Received 18 December 1996; Revised 20 June 1997; Accepted 24 June1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 388:603–612 (1997)
r 1997 WILEY-LISS, INC.

(Woolley and McEwen, 1994), and early autoradiographicstudies suggest that interneurons and not pyramidalneurons concentrate [3H]estradiol (Loy et al., 1988). Theactivity of g-amino-butyric acid (GABA) interneurons,determined by measuring expression of the GABA syn-thetic enzyme glutamate decarboxylase, is also altered bygonadal steroids in the hippocampus (Weiland, 1992b)indicating that the site of action of estrogen may be theGABA neuron.
We hypothesized that estrogen regulates pyramidalcells by acting directly on interneurons. Each interneuroncontacts multiple pyramidal cells (Sik et al., 1994) andregulates their responses to activation from CA1 Schaffercollaterals from CA3 or from commissural associationinputs. To test this hypothesis, we needed to establishwhether or not pyramidal cells or interneurons expressfunctional estrogen receptors. By using a polyclonal anti-body to the rat estrogen receptor from Okamura et al.(1992), we mapped the location of estrogen receptors in thehippocampus and some regions of the cortex. Preliminarystudies indicated that this antibody, combined with theinclusion of acrolein in the perfusion fixative, has lownonspecific staining and greater sensitivity than the H222or ER21 antibodies. In addition, we determined (1) whetherestrogen receptor immunoreactive (ER-ir) cells are presentat equivalent levels in male rats, (2) whether estrogentreatment results in a down-regulation of ER receptors ineither sex, and (3) whether estrogen-induction of theprogestin receptor occurs in the hippocampus.
MATERIALS AND METHODS
Animals
Young adult male and female Sprague-Dawley rats(Charles River Laboratories, Wilmington, MA) were main-tained in a temperature controlled environment on a 14:10light-dark cycle with free access to laboratory chow andtap water. All of the animals were treated in accord withthe principles and procedures of the National Institutes ofHealth Guide for the Care and Use of Laboratory Animals,and our protocols were approved by the Animal Care andUse Committee of Rockefeller University. Surgeries wereperformed under Metofane anesthesia. For the first experi-ment, six male and six female rats were gonadectomized 1week before perfusion. For the second experiments, 13female (experiment II) and 12 male (experiment III) ratswere gonadectomized. After 1 week, half of the animalswere implanted with Silastic capsules containing 17b-estradiol (200 µg/ml sesame oil), and the other half weresham treated for 56 hours before perfusion. This treatmentparadigm induces estradiol levels of 25–30 pg/ml serum.All the animals were killed 2.5 to 3.5 hours before lightsoff. The animals were deeply anesthetized with Metofaneand transcardially perfused with 3% acrolein in 2% para-formaldehyde. Preliminary studies showed that, althoughestrogen receptor immunostaining was present with 4%paraformaldehyde fixation, staining was more intensewith the addition of acrolein. The brains were removed,post-fixed for 2 hours in the perfusion fixative, post-fixedovernight in 2% paraformaldehyde, and transferred to 0.1M phosphate buffered saline (PBS). Coronal sections werecut at 40 µm on a Leica VT1000E and stored in cryopro-tectant at 220°C.
Immunocytochemistry
Immunocytochemistry was performed on every sixthsection through the hippocampus by using standard pub-lished procedures (Berghorn et al., 1994) with minormodifications. For detection of the estrogen receptor (ER),a polyclonal antibody (AS409), generated in rabbit againsta peptide containing amino acid 61 to the carboxyl termi-nus of the native rat ER and containing the DNA andligand binding domains, was obtained from Okamura et al.(1992). A mouse monoclonal antibody to amino acids533-547 of the human progesterone receptor peptide (PR;Traish and Wotiz, 1990; Affinity Bioreagents, NeshanicStation, NJ) was used for detection of the PR. In addition,comparison staining was performed with the H222 andER21 antibodies for the estrogen receptor, generouslyprovided by Dr. G. Greene (1984). To determine whetherthe ER are in neurons or astrocytes, double label immuno-cytochemistry was performed for ER and glial fibrillaryacidic protein (GFAP; monoclonal; Boehringer Mannheim,Indianapolis, IN). The sections were removed from cryopro-tectant, rinsed in 0.1 M PBS, incubated in 1% sodiumborohydride in PBS to neutralize the acrolein, rinsed inPBS, and blocked in 2% normal goat serum (ER) or normalhorse serum (PR) and 0.1% Triton X-100 in PBS. Sectionswere incubated at 4°C for 7 days with the estrogenreceptor antiserum diluted 1:40,000 or for 2 to 3 days withPR antiserum diluted 1:100 in 0.1% Triton X-100 in PBS.The AS409 antibody recognizes both the occupied andunoccupied forms of the estrogen receptor (Okamura et al.,1992). Primary antibody was omitted from control sec-tions, and no immunoreactivity was observed. Sectionswere rinsed in PBS, incubated in biotinylated goat-anti-rabbit (ER) or horse anti-mouse (PR) IgG (Vector Laborato-ries, Burlingame, CA) for 1 hour, rinsed in PBS, trans-ferred to AB solution (avidin-biotin complex; ABC ‘‘Elite’’kit, Vector) for 45 minutes, rinsed in PBS, and incubated incobalt/nickel enhanced 3,38-diaminobenzidine tetrahydro-chloride (DAB; Pierce, Rockford, IL) for 3 minutes. Forcolocalization of ER with GFAP, sections were rinsed inPBS, incubated overnight in with the monoclonal antibodyto GFAP (1:50). The sections were then treated as for thePR except for the use of DAB for the chromagen. Thesections were mounted on gelatin-coated slides, driedovernight, counterstained with methyl green (omittedwith double label), dehydrated, and coverslipped withPermount. Because initial attempts failed to visual PR inthe hippocampus, a variety of fixatives (paraformalde-hyde, glutaraldehyde, and acrolein in various concentra-tions), antibody concentrations, and assay conditions weretried, in addition to those cited above.
Specificity of antibody
Absorption studies were performed to demonstrate speci-ficity of the AS409 antibody. AS409, diluted 1:400 in 0.1 Mphosphate buffer (pH 7.4) containing 1% bovine serumalbumin (BSA), was mixed with Escherichia coli cell lysateexpressing estrogen-receptor-b-galactosidase fusion pro-tein (Okamura et al., 1992) in approximately 1:40 proteinratio. After incubation overnight at 4°C, the mixture wascentrifuged at 15,000 3 g for 15 minutes. The supernatantwas considered as preadsorbed serum. Brain sections wereincubated with the prepared serum at dilutions of 1:5,000–20,000 for 7 days at 4°C and then processed for ABC-DABmethod as described previously (Orikasa et al., 1994). For
604 N.G. WEILAND ET AL.

negative adsorption control, cell lysate of the E. coliexpressing b-galactosidase but not fusion protein was usedfor preincubation. No estrogen receptor immunoreactive(ER-ir) signals were detected in the sections incubatedwith the preadsorbed serum, whereas clear signals weredetected in sections with the negative control serum(Fig. 1).
Data analysis
To ensure reliable and reproducible results when quanti-fying the ER immunointensity, caution was taken to treatall sections exactly the same: same fixative, time ofpost-fix, washes, exposure to antibodies and chromagen,temperature, and dehydration. Therefore, any shrinkageor distortion of the tissue section should be equivalent inall sections. Brain sections from each animal were matchedfor anatomical localization according to Swanson (1992),and ER-ir cells were counted in regions of the hippocam-pus and retrosplenial and parietal cortex. Cell counts fromthe right and left hemispheres were averaged, and datafrom sections within a specified rostral-caudal expanse,defined as distance from bregma, were combined. The datawere analyzed by using a two-way analysis of variance forrepeated measures for effects of distance from bregma andsex (Exp. I), or hormone treatment (Exp. II and III). Inaddition, the intensity of the immunoreactivity of the
ER-ir cells from select brain regions was measured byusing an image analysis system (Imaging Research, Inc.,Saint Catherines, Ontario, Canada). The slides were codedand immunodetection was performed by using a blindprotocol. Under high power (3 400) magnification, the cellnuclei were circled and relative optical density (ROD) wasmeasured. ROD was based on a curve generated withoptical density standards to account for any uncontrolledvariability in light source. All labeled cells were measuredin any given region of the hippocampus or cortex. Forcomparison, ROD measurements of staining intensity wasalso done for the preoptic area (POA) and the ventrolateralpart of the ventral medial nucleus (VMN). Because of thehigh density of immunoreactive cells in the POA and VMN,a defined field within the center of the region was digitizedunder lower power magnification (3 100), and the cellswere defined by using threshold ranges of density toeliminated density contribution by spaces between cells,and the optical density of thresholded area was deter-mined. Because of changes in the light requirements withthe change in magnification, individual cells had a twofoldgreater intensity at 3400 vs. 3100, and thus a 23conversion factor was used to graph data from the VMNand POA on the same axis with the hippocampal andcortex data. Immunointensity data were analyzed by usinga t test for effect of sex (Exp. I) or hormone (Exp. II and III).
RESULTS
Localization of ER immunoreactivity
As has been previously reported, cells immunopositivefor the ER were prominent in those areas related to sexualbehaviors and gonadotropin release (Blaustein, 1992;Clancy and Michael, 1994) such as the VMN and POA(Figs. 1, 2) and were also observed in other brain regionsnot traditionally associated with reproductive function,including the hippocampus (Fig. 2) and some regions ofcortex (Fig. 3). The metal-enhanced DAB reaction productwas distinctly located in high density in the nucleus, witha minority of cells exhibiting some chromogen in thecytoplasm. Background levels of staining were very lowfacilitating the identification of labeled cells. No stainingwas observed with omission of the primary antibody. Withthe H222 and ER21 antibodies, a similar pattern but lessintense level of staining was observed in the VMN andPOA; however, very little staining was present in thehippocampal regions, and background levels were higherthan observed with the #AS409 antibody (data not shown).
In the hippocampus of both male and female rats,neurons immunoreactive for the estrogen receptor werescattered in low frequency throughout most regions withthe highest frequency in the hilus and stratum radiatum ofCA1 (Table I). The density of ER-ir cells also exhibited arostral-caudal gradient that was prominent in both thehilus and CA1 radiatum (Fig. 3). ER-ir cells were greatestin number in the hilus in the very rostral, dorsal sections(22.2 to 23.0 from bregma; Fig. 4). In addition, ER-ir cellswere relatively concentrated in the rostral ventral hilus(23.9 to 24.5; Table II), whereas very few ER-ir cells wereobserved in the areas of hilus located more caudally orlaterally between these two areas. In CA1 radiatum, ER-ircells were more dispersed and in greatest concentrationdorsally between 23.0 and 24.8 from bregma (Fig. 5). Inaddition, in CA1 radiatum the ER-ir cells were moreabundant adjacent to the lacunosum moleculare rather
Fig. 1. Photomicrograph of estrogen receptor immunoreactive(ER-ir) cells in the ventromedial (VMN) and arcuate nuclei (AN) in themedial basal hypothalamus showing the preadsorption test. Thesections were incubated with AS409 preincubated with the E. coli celllysate expressing the estrogen-receptor-b-galactosidase fusion protein(B) or preincubated with that expressing the b-galactosidase (A). NoER-ir signals were visible in the section preadsorbed with the former.Scale bar 5 200 µm in B (applies to A,B).
HIPPOCAMPUS ESTROGEN RECEPTOR IMMUNOREACTIVE CELLS 605

than close to the pyramidal cell layer. As seen in the hilus,ER-ir were also observed in ventral CA1; however, in thisregion (bregma 24.2 to 24.8), ER-ir cells were scatteredmost densely in the pyramidal cell layer with fewer labeled
cells in the strata radiatum and oriens (Table II). ER-ircells were also observed in all layers of the CA2 and CA3regions, but they were few in number, and therefore thenumber of cells from all three layers (radiatum, pyrami-
Fig. 2. Photomicrographs of estrogen receptor immunoreactivecells in the ventral medial nucleus (A), hilus of the dentate gyrus indorsal hippocampus (B; bregma 22.5), stratum radiatum of CA1 (C;bregma 23.7), and in the dentate gyrus in ventral hippocampus [D;bregma 24.45 (see Fig. 4)]. The immunoreactive products were
visualized with metal/enhanced DAB. DGlb, lateral blade of the dentalgyrus; g, granule cell layer; h, hilus; lm, lacunosum moleculare; m,molecular layer; p, pyramidal cell layer; po, polymorph layer; r,stratum radiatum. Scale bar 5 40 µm.
606 N.G. WEILAND ET AL.

dale, and oriens) were pooled (Table I). Based on their sizeand location, the ER-ir cells appear to be interneuronswith the exception of a very limited region of the ventraldentate gyrus located 23.7 from bregma, where the mostlateral dentate granule cells appear to express ER at low
levels. In the ventral part of the retrosplenial cortex (RSP),ER-ir cells were fairly abundant and confined to layers 2and 3. In addition, the numbers of ER-ir cells were greaterin regions 24.8 to 26.3 from bregma compared with morerostral regions (21.9 to 24.2) (Fig. 6). ER-ir cells inparietal cortex were counted from the RSP cortex to therhinal fissure, where they were found predominantly inlayers 2 and 3 but in a lower density than in RSP cortex.No rostral caudal gradient in the number of ER-ir cellswere observed in parietal cortex (Fig. 7). No colocalizationof ER-ir was observed within GFAP staining cells indicat-ing that the ER-ir cells are not astrocytes.
Sex differences
No sex differences in the overall number of ER-ir cellswere observed in any region examined in gonadectomizedrats (Figs. 4–8), although some minor rostral-caudal distri-bution differences were observed in the dentate hilus (Fig.4). Male and female rats have equivalent numbers of ER-ircells in the hippocampus and cortex. In regions of thehippocampus and cortex, the intensity of immunostainingfor the ER was equivalent in male and female rats;however, the intensity of staining was lower in the VMN ofthe male animals (Fig. 8).
Hormone regulation
In GDX rats, the mean level of ER immunoreactivity isapproximately fourfold greater in the cells of the POA andVMN than in the hippocampus and cortex (Fig. 9). Treat-ment with estrogen decreased the levels of ER immunore-activity per cell to the same extent in both male and femalerats in all brain regions examined (Fig. 9; data not shownfor male rats). The number or distribution of ER-ir cellsdid not change with estrogen treatment in the dorsal hilus(Fig. 10), the CA1 stratum radiatum, or the retrosplenialcortex (cell counts were not done for POA and VMN).
Progesterone receptor
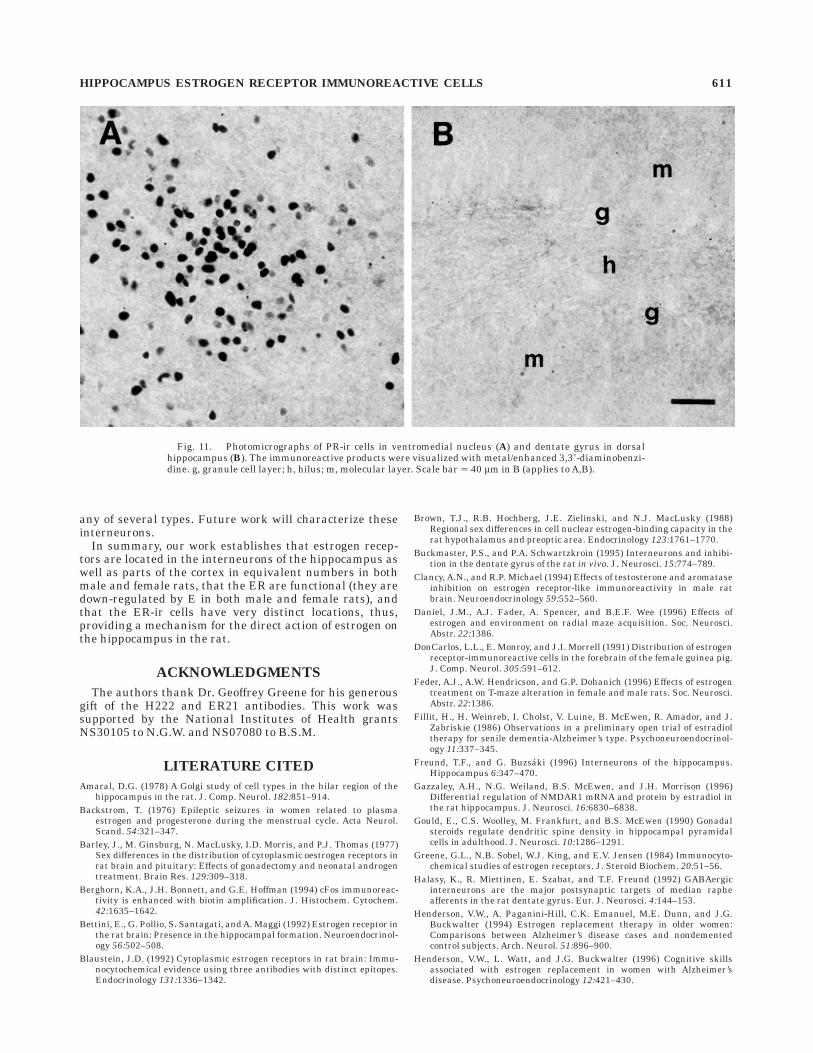
Although progesterone receptor immunoreactivity wasdistinctly present in regions associated with reproductivefunction, the POA and VMN (Fig. 11A), as has beenpreviously reported by others, none was observed in anyregion of the hippocampus under any of the immunocyto-chemical conditions used (Fig. 11B).
DISCUSSION
We have demonstrated that ER-ir cells are present inthe hippocampus of adult rats, are not located in pyrami-dal neurons or astrocytes, are in interneurons scatteredthroughout the hippocampus with the greatest density inthe stratum radiatum and dentate hilus, and are equiva-lent in both the number and intensity of immunostainingin male and female rats. Recently, many investigatorshave demonstrated effects of ovarian steroids on severalaspects of hippocampal physiology, including synaptogen-esis (Gould et al., 1990; Woolley and McEwen, 1992),long-term potentiation (Warren et al., 1995), and GABAA(McCarthy et al., 1992; Weiland and Orchinik, 1995) andNMDA receptor function (Weiland, 1992a; Woolley andMcEwen, 1994; Gazzaley et al., 1996; Woolley et al., 1996).These observations have prompted a surge of interest inexamining estrogen regulation of hippocampal-dependentlearning and memory. E treatment of OVX females hasbeen reported to improve acquisition on a radial arm maze
Fig. 3. Drawings are modified from L.W. Swanson, Brain Maps:Computer Graphics Files (version 1.0), and represent coronal sectionsfrom three different levels of brain measured from bregma. Each dotrepresents one estrogen receptor immunoreactive cell (total number ismean from male and female rats). DGlb, lateral blade of the dentategyrus; ENT, entorhinal cortex; fc, fasciola cinerea; fi, fimbria; hf, hilarfissure; mo, molecular layer of the dentate gyrus; PAR, parietal cortex;po, polymorph layer (hilus) of the dentate; RSP, retrosplenial cortex;sg, stratum granulosum; so, stratum oriens; sp, stratum pyramidale;sr, stratum radiatum; SUB, subiculum; v3, third ventricle; vip, veluminterpositum; vl, lateral ventricle.
HIPPOCAMPUS ESTROGEN RECEPTOR IMMUNOREACTIVE CELLS 607

(Daniel et al., 1996; Luine et al., 1996) as well as in areinforced T-maze alternation task (Feder et al., 1996) andimproves performance in a working memory task (O’Neilet al., 1996). In addition, E treatment has been reported topromote a shift in strategy that female rats use to solve atwo choice discrimination by using a response as opposedto a spatial strategy (Korol et al., 1996). And finally, agingfemale rats with low plasma levels of E were reported toperform significantly worse in a Morris water maze thanrats with high E levels (Juraska and Warren, 1996). Theaffects of E on learning and memory are not confined torodents. E replacement therapy has been shown to restorecognitive deficits in postmenopausal women (Sherwin,1988; Phillips and Sherwin, 1992a) and in women estrogensuppressed with GnRH agonist (Sherwin and Tulandi,1996). In addition, E treatment can delay the onset andalleviate some of the symptoms of Alzheimer’s disease inwomen (Fillit et al., 1986; Henderson et al., 1994, 1996;
Paganini-Hill and Henderson, 1994; Tang et al., 1996).Thus, it is important to establish the location and mecha-nism of the effect of E on the hippocampus.
The presence of ER protein and mRNA in the hippocam-pus of rats has been demonstrated by using binding assays(O’Keefe et al., 1993), Western blots (Maggi et al., 1989),Northern blots (Bettini et al., 1992; O’Keefe et al., 1995),and in situ hybridization techniques (Pelletier et al., 1988;Simerly et al., 1990; O’Keefe et al., 1995). Anatomicallocalization of the ER is not possible with Western andNorthern blots. Although greater cellular resolution ispossible with in situ hybridization techniques, the findingof ER mRNA in the pyramidal cells vs. interneurons seemsto depend on the DNA sequences to which the probes weremade (Pelletier et al., 1988; Simerly et al., 1990; O’Keefe etal., 1995). Additionally, the presence of truncated forms ofthe ER mRNA (Skipper et al., 1993), which may berecognized by some probes and which do not generatefunctional protein (Pfeffer et al., 1993; Koehorst et al.,1994), may confound the interpretation of the in situresults. Recent work with a newly cloned b form of the ER(Kuiper et al., 1996) has established the presence of ERbmRNA in brain (Kuiper et al., 1997). Preliminary workindicates that the H-222 antibody does not cross-react theERb protein on immunoblots (Kuiper et al., 1997). Whetheror not the AS409 antibody cross-reacts with the ERbremains to be established. Results from earlier workutilizing [3H]estradiol uptake (Pfaff and Keiner, 1973; Loyet al., 1988) indicated the presence of E-concentrating cellsin the hippocampus, however poor histology due to thelong exposure time did not permit cellular localization.ER-ir (H222) cells have been observed previously in thehippocampus and cingulate cortex of the guinea pig (Don-Carlos et al., 1991). The greater sensitivity and lowbackground staining with the AS409 antibody comparedwith the H222 or ER21 have permitted visualization ofER-ir cells in regions of brain that appear to have lowernumbers of ER.
The lack of sex differences in the number and distribu-tion of ER in gonadectomized rats is not surprising. Maleand female gonadectomized rats have equivalent ER bind-ing capacity in whole hypothalamus, and sex differencesappear only in microdissected regions or in response tovariations in circulating steroids (Barley et al., 1977;Brown et al., 1988; Yuan et al., 1995). No sex differences inthe intensity of ER immunoreactivity in gonadectomizedanimals were observed in the hippocampus, retrosplenial
TABLE 1. Number of Cells Immunoreactive for the Estrogen Receptor/Section/Hemisphere in the Dorsal Hippocampus and Retrosplenial (RSP) andParietal (PAR) Cortex in Male and Female Rats1
Distancefrom bregma Sex
CA1radiatum
CA1pyramidale
CA1oriens CA2 CA3
Dentatehilus
Fasciolacinerea RSP cortex PAR cortex
21.9 to 22.2 Female 4.5 6 1.2 4.7 6 1.4 3.7 6 0.5 19.8 6 1.9Male 4.7 6 1.5 4.5 6 0.8 3.0 6 0.2 21.5 6 1.6
22.2 to 23.0 Female 2.9 6 0.6 0.8 6 0.4 0.2 6 0.1 1.4 6 0.1 2.8 6 1.3 20.1 6 0.8 0.8 6 0.2 4.9 6 0.5 20.1 6 1.8Male 2.4 6 0.9 0.7 6 0.2 0.1 6 0.1 0.8 6 0.1 1.1 6 0.3 18.0 6 2.5 1.1 6 0.3 3.6 6 0.2 22.1 6 2.7
23.0 to 23.8 Female 6.6 6 1.8 0.6 6 0.3 0.1 6 0.1 1.3 6 0.6 1.4 6 0.3 7.4 6 1.2 2.2 6 0.6 4.5 6 0.9 20.2 6 2.4Male 5.3 6 0.6 1.5 6 0.4 0.2 6 0.1 1.3 6 0.3 1.1 6 0.5 7.7 6 1.4 1.4 6 0.4 4.1 6 0.7 24.0 6 0.5
23.8 to 24.8 Female 7.3 6 0.9 1.4 6 0.5 0.2 6 0.1 1.1 6 0.2 1.3 6 0.5 7.0 6 1.1 1.6 6 0.4 4.1 6 0.4 18.0 6 2.4Male 6.5 6 1.5 0.8 6 0.2 0.5 6 0.2 1.2 6 0.2 0.6 6 0.3 8.7 6 1.6 0.8 6 0.3 4.3 6 0.5 19.9 6 1.1
24.2 to 24.8 Female 8.6 6 0.7 1.9 6 0.6 0.5 6 0.4 0.8 6 0.3 2.2 6 0.6 5.6 6 0.9 1.3 6 0.1 4.2 6 0.3 20.1 6 2.6Male 9.1 6 1.6 2.1 6 0.5 0.5 6 0.2 1.0 6 0.3 2.8 6 0.6 8.6 6 0.9 0.8 6 0.3 4.8 6 0.6 22.8 6 2.1
24.8 to 25.5 Female 4.0 6 1.0 1.7 6 0.2 0.6 6 0.4 1.6 6 0.6 4.4 6 1.4 4.9 6 0.8 0.2 6 0.2 6.5 6 1.0 19.4 6 2.1Male 4.4 6 1.6 1.4 6 0.6 0.7 6 0.3 1.2 6 0.6 4.6 6 1.3 4.9 6 1.0 0.8 6 0.2 3.7 6 0.9 18.6 6 2.0
25.5 to 26.3 Female 4.5 6 1.2 1.6 6 0.3 0.9 6 0.4 6.6 6 2.3 7.1 6 1.3 4.7 6 0.9 21.6 6 3.6Male 3.6 6 1.3 3.8 6 1.2 0.9 6 0.4 1.3 6 1.2 11.6 6 1.1 8.5 6 0.3 18.2 6 0.2
1One to three sections per animal were averaged to obtain a mean for each animal. Five to six animals/group were averaged to obtain the final mean 6 SEM for each region.
Fig. 4. The distribution of estrogen receptor immunoreactive(ER-ir) cells in the dorsal hilus measured from bregma in male andfemale rats. The greatest density of ER-ir cells was between 22.2 and22.3 from bregma. No significant overall sex difference in the numberof ER-ir cells was detected; however, small differences in distributionwere observed.
608 N.G. WEILAND ET AL.

cortex, or preoptic area; however, slightly lower levels ofimmunoreactivity were observed in the VMN of male rats.A reduced nuclear estrogen binding capacity (Brown et al.,1988) is also found in microdissected VMN in male rats.The sex differences in ER levels may contribute to the sexdifferences in the VMN in the response to the estrogen-induction of PR (Rainbow et al., 1982), [3H]muscimolbinding to the GABAA receptor (McCarthy et al., 1992),and dendritic spines (Segarra and McEwen, 1991). Similarsex differences in [3H]muscimol binding (McCarthy et al.,1992) and dendritic spine induction (Lewis et al., 1995) arefound in the hippocampus; however, in this region thebasis for the sex differences must be due to somethingother than differences in the ER. Treatment with estrogendecreased ER immunointensity to equivalent levels inboth male and female gonadectomized rats. ER levels inintact animals will depend, in females, on the day of theestrous cycle and the resulting serum estrogen levels and,in males, on the level of testosterone aromatized to estro-gen in the hippocampus.
The density of ER-ir cells, although greatest in the hilusand the stratum radiatum of CA1, is still rather sparse incomparison with hypothalamic regions such as the VMN.An in situ hybridization study by using film demonstrated
a similar low distribution of ER mRNA in the hippocam-pus, although cellular localization was not determined(Simerly et al., 1990). Not only are the ER-ir cells moresparse, but the average cellular intensity of immunostain-ing is about fourfold lower in the hippocampus and corticalregions than in hypothalamic areas. Although the densityof immunostaining does not necessarily have a one to onerelationship to the level of ER protein, the mean amountsof ER immunoreactivity per cell varied very little fromanimal to animal within the same group or from experi-ment to experiment within the same treatment groups,indicating that the relative comparisons of ER levelsbetween regions and treatment groups are valid. Thesereceptors are physiologically functional, in that they canbind estradiol and be down-regulated as we have demon-strated here in the hippocampus and has been shown byothers with Western blotting in hypothalamic as well asperipheral tissue such as the uterus (Zhou et al., 1995).The influence of each of the ER-ir neurons may be veryextensive (Buckmaster and Schwartzkroin, 1995). TheER-ir cells appear to be interneurons based on theirlocation and lack of colocalization with GFAP. Some inter-neurons have been shown to contact as many as 1,500pyramidal neurons as well as 40–60 other interneurons(Sik et al., 1995). Distinct types of interneurons have been
TABLE 2. The Number of Cells Immunoreactive for the Estrogen Receptor/Section/Hemisphere in the Dorsal and Ventral Subiculum and the VentralHippocampus in Male and Female Rats1
Distancefrombregma Sex
CA1radiatum
CA1pyramidal
CA1oriens CA3
Dentatehilus
ENTcortex
Dorsalsubiculum
Ventralsubiculum
24.2 to 24.8 Female 2.9 6 0.9 9.9 6 4.2 2.0 6 0.3 7.9 6 1.5 17.3 6 3.8 1.6 6 1.2 2.6 6 0.2 4.7 6 1.0Male 2.5 6 1.5 9.1 6 2.4 2.2 6 0.6 5.5 6 2.5 14.4 6 4.3 1.1 6 0.7 2.0 6 0.4 5.0 6 1.4
24.8 to 25.5 Female 3.4 6 0.9 9.0 6 2.1 1.9 6 0.4 2.0 6 0.6 6.4 6 1.0 1.6 6 1.0 4.3 6 0.9 5.6 6 1.0Male 2.8 6 0.5 11.6 6 2.3 3.4 6 0.5 4.6 6 1.3 9.5 6 2.4 0.8 6 0.4 3.4 6 0.9 5.2 6 1.0
1One to three sections per animal were averaged to obtain a mean for each animal. Five to six animals/group were averaged to obtain the final mean 6 SEM for each region. ENT,entorhinal cortex.
Fig. 5. The distribution of estrogen receptor immunoreactive(ER-ir) cells in dorsal CA1 radiatum increased from rostral to caudaluntil 24.8 from bregma and the dropped to low levels. No sexdifferences were observed in the number or distribution of ER-ir cells.
Fig. 6. The distribution of estrogen receptor immunoreactive(ER-ir) cells in the retrosplenial cortex (RSP) increased caudally inboth male and female rats. The increase occurred earlier in the female(24.8 to 25.5) than in the male (25.5 to 26.3) rats; however, no overallsex difference in ER-ir cell number was detected.
HIPPOCAMPUS ESTROGEN RECEPTOR IMMUNOREACTIVE CELLS 609

described and classified according to their location, mor-phology, firing characteristics, and peptide content (Ama-ral, 1978; Walaas, 1983; Kosaka et al., 1987; Lacaille et al.,1987; Sloviter and Nilaver, 1987; Sloviter, 1989; Halasy etal., 1992; Samulack et al., 1993; for review see Freund andBuzsaki, 1996). At present, we have only classified theER-ir cells based on their location. In the CA1 region of thehippocampus, most of the ER-ir cells are in the stratumradiatum adjacent to the lancunosum moleculare, whereasonly a few cells are associated with the pyramidal cell
layer or stratum oriens. Therefore, most of these cells arenot basket cells or O-LM cells as described by Sik et al. (Siket al., 1995). In the dentate hilus, most of the ER-ir cellswere adjacent to the granule cell layer, where they could be
Fig. 7. No sex or distribution differences were observed in thenumber of estrogen receptor immunoreactive cells in the parietalcortex.
Fig. 8. The intensity of immunostaining was equivalent in gonad-ectomized male and female rats in regions of the hippocampus andpreoptic areas (POA). Levels of immunostaining in the ventromedialnucleus (VMN) were significantly lower in male rats (P , 0.05).
Fig. 9. Treatment with estrogen decreased the intensity of immu-nostaining /cell in ovariectomized (OVX) female rats in all regionsexamined. The intensity of immunostaining was approximately four-fold greater in the hypothalamic regions (POA and VMN) than in thehippocampus (CA1 and Hilus) or cortex (RSP). *P , 0.05 for ovariecto-mized plus estrogen (OVX 1 E) compared with OVX. ER, estrogenreceptor; POA, preoptic area; VMN, ventromedial nucleus; RSP,retrosplenial cortex; ROD, relative optical density.
Fig. 10. Treatment of ovariectomized (OVX) rats with estrogen didnot change the number or distribution of estrogen receptor immunore-active (ER-ir) cells in the dorsal hilus of the dentate gyrus.
610 N.G. WEILAND ET AL.
any of several types. Future work will characterize theseinterneurons.
In summary, our work establishes that estrogen recep-tors are located in the interneurons of the hippocampus aswell as parts of the cortex in equivalent numbers in bothmale and female rats, that the ER are functional (they aredown-regulated by E in both male and female rats), andthat the ER-ir cells have very distinct locations, thus,providing a mechanism for the direct action of estrogen onthe hippocampus in the rat.
ACKNOWLEDGMENTS
The authors thank Dr. Geoffrey Greene for his generousgift of the H222 and ER21 antibodies. This work wassupported by the National Institutes of Health grantsNS30105 to N.G.W. and NS07080 to B.S.M.
LITERATURE CITED
Amaral, D.G. (1978) A Golgi study of cell types in the hilar region of thehippocampus in the rat. J. Comp. Neurol. 182:851–914.
Backstrom, T. (1976) Epileptic seizures in women related to plasmaestrogen and progesterone during the menstrual cycle. Acta Neurol.Scand. 54:321–347.
Barley, J., M. Ginsburg, N. MacLusky, I.D. Morris, and P.J. Thomas (1977)Sex differences in the distribution of cytoplasmic oestrogen receptors inrat brain and pituitary: Effects of gonadectomy and neonatal androgentreatment. Brain Res. 129:309–318.
Berghorn, K.A., J.H. Bonnett, and G.E. Hoffman (1994) cFos immunoreac-tivity is enhanced with biotin amplification. J. Histochem. Cytochem.42:1635–1642.
Bettini, E., G. Pollio, S. Santagati, and A. Maggi (1992) Estrogen receptor inthe rat brain: Presence in the hippocampal formation. Neuroendocrinol-ogy 56:502–508.
Blaustein, J.D. (1992) Cytoplasmic estrogen receptors in rat brain: Immu-nocytochemical evidence using three antibodies with distinct epitopes.Endocrinology 131:1336–1342.
Brown, T.J., R.B. Hochberg, J.E. Zielinski, and N.J. MacLusky (1988)Regional sex differences in cell nuclear estrogen-binding capacity in therat hypothalamus and preoptic area. Endocrinology 123:1761–1770.
Buckmaster, P.S., and P.A. Schwartzkroin (1995) Interneurons and inhibi-tion in the dentate gyrus of the rat in vivo. J. Neurosci. 15:774–789.
Clancy, A.N., and R.P. Michael (1994) Effects of testosterone and aromataseinhibition on estrogen receptor-like immunoreactivity in male ratbrain. Neuroendocrinology 59:552–560.
Daniel, J.M., A.J. Fader, A. Spencer, and B.E.F. Wee (1996) Effects ofestrogen and environment on radial maze acquisition. Soc. Neurosci.Abstr. 22:1386.
DonCarlos, L.L., E. Monroy, and J.I. Morrell (1991) Distribution of estrogenreceptor-immunoreactive cells in the forebrain of the female guinea pig.J. Comp. Neurol. 305:591–612.
Feder, A.J., A.W. Hendricson, and G.P. Dohanich (1996) Effects of estrogentreatment on T-maze alteration in female and male rats. Soc. Neurosci.Abstr. 22:1386.
Fillit, H., H. Weinreb, I. Cholst, V. Luine, B. McEwen, R. Amador, and J.Zabriskie (1986) Observations in a preliminary open trial of estradioltherapy for senile dementia-Alzheimer’s type. Psychoneuroendocrinol-ogy 11:337–345.
Freund, T.F., and G. Buzsaki (1996) Interneurons of the hippocampus.Hippocampus 6:347–470.
Gazzaley, A.H., N.G. Weiland, B.S. McEwen, and J.H. Morrison (1996)Differential regulation of NMDAR1 mRNA and protein by estradiol inthe rat hippocampus. J. Neurosci. 16:6830–6838.
Gould, E., C.S. Woolley, M. Frankfurt, and B.S. McEwen (1990) Gonadalsteroids regulate dendritic spine density in hippocampal pyramidalcells in adulthood. J. Neurosci. 10:1286–1291.
Greene, G.L., N.B. Sobel, W.J. King, and E.V. Jensen (1984) Immunocyto-chemical studies of estrogen receptors. J. Steroid Biochem. 20:51–56.
Halasy, K., R. Miettinen, E. Szabat, and T.F. Freund (1992) GABAergicinterneurons are the major postsynaptic targets of median rapheafferents in the rat dentate gyrus. Eur. J. Neurosci. 4:144–153.
Henderson, V.W., A. Paganini-Hill, C.K. Emanuel, M.E. Dunn, and J.G.Buckwalter (1994) Estrogen replacement therapy in older women:Comparisons between Alzheimer’s disease cases and nondementedcontrol subjects. Arch. Neurol. 51:896–900.
Henderson, V.W., L. Watt, and J.G. Buckwalter (1996) Cognitive skillsassociated with estrogen replacement in women with Alzheimer’sdisease. Psychoneuroendocrinology 12:421–430.
Fig. 11. Photomicrographs of PR-ir cells in ventromedial nucleus (A) and dentate gyrus in dorsalhippocampus (B). The immunoreactive products were visualized with metal/enhanced 3,38-diaminobenzi-dine. g, granule cell layer; h, hilus; m, molecular layer. Scale bar 5 40 µm in B (applies to A,B).
HIPPOCAMPUS ESTROGEN RECEPTOR IMMUNOREACTIVE CELLS 611

Juraska, J.M., and S.G. Warren (1996) Spatial memory decline in aged,noncycling female rats varies with the phase of estropause. Soc.Neurosci. Abstr. 22:1387.
Koehorst, S.G.A., J.J. Cox, G.H. Donker, S. Lopes da Silva, J.P.H. Burbach,J.H.H. Thijssen, and M.A. Blankenstein (1994) Functional analysis ofan alternatively spliced estrogen receptor lacking exon 4 isolated fromMCF-7 breast cancer cells and meningioma tissue. Mol. Cell. Endocri-nol. 101:237–245.
Korol, D.L., J.M. Couper, C.K. McIntyre, and P.E. Gold (1996) Strategies forlearning across the estrous cycle in female rats. Soc. Neurosci. Abstr.22:1386.
Kosaka, T., H. Katsumaru, K. Hama, J.-Y. Wu, and C.W. Heizmann (1987)GABAergic neurons containing the Ca21-binding protein parvalbuminin the rat hippocampus and dentate gyrus. Brain Res. 419:119–130.
Kuiper, G.G.J.M., E. Enmark, M. Pelto-Huikko, S. Nilsson, and J.A.Gustafsson (1996) Cloning of a novel estrogen receptor expressed in ratprostate and ovary. Proc. Natl. Acad. Sci. USA 93:5925–5930.
Kuiper, G.G.J.M., B. Carlsson, K. Graddien, E. Enmark, J. Haggblad, S.Nilsson, and J.-A. Gustafsson (1997) Comparison of the ligand bindingspecificity and transcript tissue distribution of estrogen receptors a andb. Endocrinology 138:863–870.
Lacaille, J.-C., A.L. Mueller, D.D. Kunkel, and P.A. Schwartzkroin (1987)Local circuit interactions between oriens/alveus interneurons and CA1pyramidal cells in hippocampal slices: Electrophysiology and morphol-ogy. J. Neurosci. 7:1979–1993.
Lewis, C., B.S. McEwen, and M. Frankfurt (1995) Estrogen-induction ofdendritic spines in ventromedial hypothalamus and hippocampus: Effects ofneonatal aromatase blockade and adult GDX. Dev. Brain Res. 87:91–95.
Loy, R., J.L. Gerlach, and B.S. McEwen (1988) Autoradiographic localiza-tion of estradiol-binding neurons in the rat hippocampal formation andentorhinal cortex. Dev. Brain Res. 39:245–251.
Luine, V.N., J. Renatas, L. Sterbank, and K. Beck (1996) Estradiol effects onrat spatial memory. Soc. Neurosci. Abstr. 22:1387.
Maggi, A., L. Susanna, E. Bettini, G. Mantero, and I. Zucchi (1989)Hippocampus: A target for estrogen action in mammalian brain. Mol.Endocrinol. 3:1165–1170.
McCarthy, M.M., H. Coirini, M. Schumacher, A.E. Johnson, D.W. Pfaff, S.Schwartz-Giblin, and B.S. McEwen (1992) Steroid regulation and sexdifferences in [3H]muscimol binding in hippocampus, hypothalamusand midbrain in rats. J. Neuroendocrinol. 4:393–399.
O’Keefe, J.A., E.B. Pedersen, A.J. Castro, and R.J. Handa (1993) Theontogeny of estrogen receptors in heterochronic hippocampal andneocortical transplants demonstrates an intrinsic development pro-gram. Dev. Brain Res. 75:105–112.
O’Keefe, J.A., Y. Li, L.H. Burgess, and R.J. Handa (1995) Estrogen receptormRNA alterations in the developing rat hippocampus. Mol. Brain Res.30:115–124.
O’Neil, M.F., L.W. Means, M.C. Poole, and R.J. Hamm (1996) Estrogenaffects performance of ovariectomized rats in a two-choice water-escapeworking memory task. Psychoneuroendocrinology 21:51–65.
Okamura, H., K. Yamamoto, S. Hayashi, A. Kuroiwa, and M. Muramatsu(1992) Apolyclonal antibody to the rat oestrogen receptor expressed inEscherichia coli: Characterization and application to immunohistochem-istry. J. Endocrinol. 135:333–341.
Orikasa, C., H. Okamura, and S. Hayashi (1994) Estrogen receptor found inthe facial nucleus of the newborn rat is suppressed by exogenousestrogen: Immuno- and in situ hybridization histochemical studies.Dev. Brain Res. 82:9–17.
Paganini-Hill, A., and V.W. Henderson (1994) Estrogen deficiency and riskof Alzheimer’s disease in women. Am. J. Epidemiol. 3:3–16.
Pelletier, G., N. Liao, N. Follea, and M.V. Govindan (1988) Mapping ofestrogen receptor-producing cells in the rat brain by in situ hybridiza-tion. Neurosci. Lett. 94:23–28.
Pfaff, D., and M. Keiner (1973) Atlas of estradiol-concentrating cells in thecentral nervous system of the female rat. J. Comp. Neurol. 151:121–157.
Pfeffer, U., E. Fecarotta, L. Castagnetta, and G. Vidali (1993) Estrogenreceptor variant messenger RNA lacking exon 4 in estrogen-responsivehuman breast cancer cell lines. Cancer Res. 53:741–743.
Phillips, S.M., and B.B. Sherwin (1992a) Effects of estrogen on memoryfunction in surgically menopausal women. Psychoneuroendocrinology17:485–495.
Phillips, S.M., and B.B. Sherwin (1992b) Variations in memory functionand sex steroid hormones across the menstrual cycle. Psychoneuroendo-crinology 17:497–506.
Rainbow, T.C., B. Parsons, and B.S. McEwen (1982) Sex differences in ratbrain oestrogen and progestin receptors. Nature 300:648–649.
Samulack, D.D., S. Williams, and J. Lacaille (1993) Hyperpolarizing synapticpotentials evoked in CA1 pyramidal cells by glutamate stimulation ofinterneurons from the oriens/alveus border of rat hippocampal slices: I.Electrophysiological response properties. Hippocampus 3:331–344.
Segarra, A.C., and B.S. McEwen (1991) Estrogen increases spine density inventromedial hypothalamic neurons of peripubertal rats. Neuroendocri-nology 54:365–372.
Sherwin, B.B. (1988) Estrogen and/or androgen replacement therapy andcognitive functioning in surgically menopausal women. Psychoneuroen-docrinology 13:345–357.
Sherwin, B.B., and T. Tulandi (1996) ‘‘Add-back’’ estrogen reverses cogni-tive deficits induced by a gonadotropin-releasing agonist in women withleiomyomata uteri. J. Clin. Endocrinol. Metab. 81:2545–2549.
Sik, A., A. Ylinin, M. Penttonen, and G. Buzsaki (1994) Inhibitory CA1-CA3-hilar region feedback in the hippocampus. Science 265:1722–1724.
Sik, A., M. Penttonen, A. Ylinen, and G. Buzsaki (1995) Hippocampal CA1interneurons: An in vivo intracellular labeling study. J. Neurosci.15:6651–6665.
Simerly, R.B., C. Chang, M. Muramatsu, and L.W. Swanson (1990) Distribu-tion of androgen and estrogen receptor mRNA-containing cells in therat brain: An in situ hybridization study. J. Comp. Neurol. 294:76–95.
Skipper, J.K., L.J. Young, J.M. Bergeron, M.T. Tetzlaff, C.T. Osborn, and D.Crews (1993) Identification of an isoform of the estrogen receptormessenger RNA lacking exon four and present in the brain. Proc. Natl.Acad. Sci. USA 90:7172–7175.
Sloviter, R.S. (1989) Chemically defined hippocampal interneurons andtheir possible relationship to seizure mechanisms. In V. Chan-Palayand C. Kohler (eds): Neurology and Neurobiology, Vol. 52. New York:Alan R. Liss, Inc. pp. 443–461.
Sloviter, R.S., and G. Nilaver (1987) Immunocytochemical localization ofGABA-, cholecystokinin-, vasoactive intestinal polypeptide-, and somato-statin-like immunoreactivity in the area dentata and hippocampus ofthe rat. J. Comp. Neurol. 256:42–60.
Swanson, L.W. (1992) Brain Maps: Structure of the Rat Brain. New York:Elsevier.
Tang, M.X., D. Jacobs,Y. Stern, K. Marder, P. Schofield, B. Gurland, H.Andrews,and R. Mayeux (1996) Effect of oestrogen during menopause on risk andage of onset of Alzheimer’s disease. Endocrinology 115:1412–1417.
Terasawa, E., and P.S. Timiras (1968) Electrical activity during the estrouscycle of the rat: Cyclic changes in limbic structures. Endocrinology83:207–216.
Traish, A.B., and H.H. Wotiz (1990) Monoclonal and polyclonal antibodiesto human progesterone receptor peptide-(533-547) recognize a specificsite in unactivated (8s) and activated (4S) progesterone receptor anddistinguish between intact and proteolyzed receptors. Endocrinology127:1167–1175.
Walaas, I. (1983) The hippocampus. In P.C. Emson (ed): Clinical Neuro-anatomy. New York: Raven Press, pp. 337–358.
Warren, S.G., A.G. Humphreys, J.M. Juraska, and W.T. Greenough (1995)LTP varies across the estrous cycle: Enhanced synaptic plasticity inproestrus rats. Brain Res. 703:26–30.
Weiland, N.G. (1992a) Glutamic acid decarboxylase messenger ribonucleicacid is regulated by estradiol and progesterone in the hippocampus.Endocrinology 131:2697–2702.
Weiland, N.G. (1992b) Estradiol selectively regulates agonist binding siteson the N-methyl-D-aspartate receptor complex in the CA1 region of thehippocampus. Endocrinology 131:662–668.
Weiland, N.G., and M. Orchinik (1995) Specific subunit mRNAs of theGABAA receptor are regulated by progesterone in subfields of thehippocampus. Mol. Brain Res. 32:271–278.
Woolley, C.S., N.G. Weiland, B.S. McEwen, and P.A. Schwartzkroin (1997)Estradiol increases the sensitivity of hippocampal CA1 pyramidal cellsto N-methyl-D-aspartate receptor-mediated synaptic input: Correlationwith dendritic spine density. J. Neurosci. 17:1848–1859.
Woolley, C.S., and B.S. McEwen (1992) Estradiol mediates fluctuation inhippocampal synapse density during the estrous cycle in the adult rat.J. Neurosci. 12:2549–2554.
Woolley, C.S., and B.S. McEwen (1994) Estradiol regulates hippocampaldendritic spine density via an N-methyl-D-aspartate receptor-depen-dent mechanism. J. Neurosci. 14:7680–7687.
Yuan, H., D.A. Bowlby, T.J. Brown, R.B. Hochberg, and N.J. MacLusky(1995) Distribution of occupied and unoccupied estrogen receptors inthe rat brain: Effects of physiological gonadal steroid exposure. Endocri-nology 136:96–105.
Zhou, Y., P.J. Shughrue, and D.M. Dorsa (1995) Estrogen receptor protein isdifferentially regulated in the preoptic area of the brain and in theuterus during the rat estrous cycle. Neuroendocrinology 61:276–283.
612 N.G. WEILAND ET AL.