
Differential regulation of the GLUT1 and GLUT3 glucose transporters by growth factors and...
7
Differential regulation of the GLUT1 and GLUT3 glucose transporters by growth factors and pro-inflammatory cytokines in equine articular chondrocytes Toby Phillips, Ivan Ferraz 1 , Susan Bell, Peter D. Clegg, Stuart D. Carter, Ali Mobasheri * Connective Tissue and Molecular Pathogenesis Research Groups, Faculty of Veterinary Science, University of Liverpool, Liverpool L69 7ZJ, UK Accepted 16 January 2004 Abstract Glucose serves as the major energy substrate for articular chondrocytes and as the main precursor for the synthesis of extra- cellular matrix glycosaminoglycans in cartilage. Chondrocytes have been shown to express several glucose transporter (GLUT) isoforms including GLUT1 and GLUT3. The aim of this investigation was to determine the effects of endocrine and cytokine factors on the capacity of equine articular chondrocytes for transporting 2-deoxy-D-[2, 6- 3 H] glucose and on the expression levels of GLUT1 and GLUT3. Chondrocytes maintained in monolayer culture were stimulated for 24 h with TNF-a (100 ng mL 1 ), IL-1b (100 ng mL 1 ), IGF-I (20 ng mL 1 ), TGF-b (20 ng mL 1 ) and insulin (12.5 lg mL 1 ) before measuring uptake of non-metabolizable 2-deoxyglucose in the presence and absence of the glucose transport inhibitor cytochalasin B. Polyclonal antibodies to GLUT1 and GLUT were used to compare GLUT1 and GLUT3 expression in stimulated and un-stimulated alginate encapsulated chondrocytes by Western blotting. Results indicated that 2-deoxyglucose uptake was inhibited by up to 95% in the presence of cytochalasin B suggesting that glucose uptake into equine chondrocytes is GLUT-mediated. Insulin had no effect on glucose uptake, but treatment with IGF-I, TGF-b, IL-1b and TNF-a resulted in a significant increase (>65%) in 2-deoxyglucose uptake compared to control values. GLUT1 was found to be increased in chondrocytes stimulated with all the growth factors and cytokines but GLUT 3 was only upregulated by IGF-I. The data presented support a critical role for glucose in the responses of equine articular chondrocytes to pro-inflam- matory cytokines and anabolic endocrine factors. Ó 2004 Elsevier Ltd. All rights reserved. Keywords: Cartilage; Chondrocyte; Glucose transport; GLUT; Cytokine; Growth factor 1. Introduction Recent research suggests that osteoarticular disorders in humans and veterinary species may be directly linked to obesity and may therefore have nutritional and en- docrine abnormalities at the root of their pathogenesis. Glucose is an essential energy source for mammalian cells and in articular cartilage glucose plays a pivotal role in the physiology of the chondrocytes by driving the extracellular matrix biosynthetic machinery of this un- ique cell type (Wang et al., 1999; Mobasheri et al., 2002b). Glucose is also a major energy source and a precursor for the synthesis of glycosaminoglycans (Mobasheri et al., 2002a). Despite this realisation, there is limited published information about the molecular mechanisms responsible for nutrient transport across the chondrocyte membrane and their regulation by growth factors and pro-inflammatory cytokines. The Veterinary Journal 169 (2005) 216–222 The Veterinary Journal www.elsevier.com/locate/tvjl * Corresponding author. Tel.: +44-151-794-4284; fax: +44-151-794- 4243. E-mail address: [email protected] (A. Mobasheri). 1 Present Address: Servicio de Reumatologia, Hospital Universitario de Canarias, Tenerife, La Cuesta, Santa Cruz de Tenerife and Departamento de Bioqu ımica y Biolog ıa Molecular, Universidad de La Laguna, Tenerife, Spain. 1090-0233/$ - see front matter Ó 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.tvjl.2004.01.026
-
Upload
toby-phillips -
Category
Documents
-
view
213 -
download
0
Transcript of Differential regulation of the GLUT1 and GLUT3 glucose transporters by growth factors and...

The
The Veterinary Journal 169 (2005) 216–222
Veterinary Journalwww.elsevier.com/locate/tvjl
Differential regulation of the GLUT1 and GLUT3 glucosetransporters by growth factors and pro-inflammatory
cytokines in equine articular chondrocytes
Toby Phillips, Ivan Ferraz 1, Susan Bell, Peter D. Clegg,Stuart D. Carter, Ali Mobasheri *
Connective Tissue and Molecular Pathogenesis Research Groups, Faculty of Veterinary Science,
University of Liverpool, Liverpool L69 7ZJ, UK
Accepted 16 January 2004
Abstract
Glucose serves as the major energy substrate for articular chondrocytes and as the main precursor for the synthesis of extra-
cellular matrix glycosaminoglycans in cartilage. Chondrocytes have been shown to express several glucose transporter (GLUT)
isoforms including GLUT1 and GLUT3. The aim of this investigation was to determine the effects of endocrine and cytokine factors
on the capacity of equine articular chondrocytes for transporting 2-deoxy-DD-[2, 6-3H] glucose and on the expression levels of
GLUT1 and GLUT3.
Chondrocytes maintained in monolayer culture were stimulated for 24 h with TNF-a (100 ngmL�1), IL-1b (100 ngmL�1), IGF-I
(20 ngmL�1), TGF-b (20 ngmL�1) and insulin (12.5 lgmL�1) before measuring uptake of non-metabolizable 2-deoxyglucose in the
presence and absence of the glucose transport inhibitor cytochalasin B. Polyclonal antibodies to GLUT1 and GLUT were used to
compare GLUT1 and GLUT3 expression in stimulated and un-stimulated alginate encapsulated chondrocytes by Western blotting.
Results indicated that 2-deoxyglucose uptake was inhibited by up to 95% in the presence of cytochalasin B suggesting that
glucose uptake into equine chondrocytes is GLUT-mediated. Insulin had no effect on glucose uptake, but treatment with IGF-I,
TGF-b, IL-1b and TNF-a resulted in a significant increase (>65%) in 2-deoxyglucose uptake compared to control values. GLUT1
was found to be increased in chondrocytes stimulated with all the growth factors and cytokines but GLUT 3 was only upregulated
by IGF-I. The data presented support a critical role for glucose in the responses of equine articular chondrocytes to pro-inflam-
matory cytokines and anabolic endocrine factors.
� 2004 Elsevier Ltd. All rights reserved.
Keywords: Cartilage; Chondrocyte; Glucose transport; GLUT; Cytokine; Growth factor
1. Introduction
Recent research suggests that osteoarticular disorders
in humans and veterinary species may be directly linked
to obesity and may therefore have nutritional and en-
* Corresponding author. Tel.: +44-151-794-4284; fax: +44-151-794-
4243.
E-mail address: [email protected] (A. Mobasheri).1 Present Address: Servicio de Reumatologia, Hospital Universitario
de Canarias, Tenerife, La Cuesta, Santa Cruz de Tenerife and
Departamento de Bioqu�ımica y Biolog�ıa Molecular, Universidad de
La Laguna, Tenerife, Spain.
1090-0233/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tvjl.2004.01.026
docrine abnormalities at the root of their pathogenesis.
Glucose is an essential energy source for mammalian
cells and in articular cartilage glucose plays a pivotal
role in the physiology of the chondrocytes by driving the
extracellular matrix biosynthetic machinery of this un-
ique cell type (Wang et al., 1999; Mobasheri et al.,
2002b). Glucose is also a major energy source and aprecursor for the synthesis of glycosaminoglycans
(Mobasheri et al., 2002a). Despite this realisation, there
is limited published information about the molecular
mechanisms responsible for nutrient transport across
the chondrocyte membrane and their regulation by
growth factors and pro-inflammatory cytokines.

T. Phillips et al. / The Veterinary Journal 169 (2005) 216–222 217
The facilitated transport of glucose and glucose-derived compounds across the chondrocyte membrane
represents the rate-limiting step in glucose metabolism
(Shikhman et al., 2001a) and is therefore essential for
chondrogenesis and the functional integrity of articu-
lating joints. Mammalian cells transport glucose across
the plasma membrane by facilitated diffusion through
the glucose transporters (or GLUTs) which belong to
family of over fourteen membrane proteins collectivelyknown as the glucose GLUT/SLC2A family of glucose/
polyol transporters (Joost and Thorens, 2001; Joost
et al., 2002; Wu and Freeze, 2002). Recent reports sug-
gest that chondrocytes express multiple isoforms of
the GLUT/SLC2A family, including the GLUT1 and
GLUT3 proteins (Ohara et al., 2001; Shikhman et al.,
2001a; Mobasheri et al., 2002a). Anabolic endocrine
factors such as insulin-like growth factor I (IGF-I)stimulate glucose uptake in human chondrocytes (Rich-
ardson et al., 2003).
Studies from our group (Richardson et al., 2003) and
one other laboratory (Shikhman et al., 2001a) suggest
that net glucose transport in human chondrocytes is also
stimulated by pro-inflammatory cytokines such as inter-
leukin 1 beta (IL-1b), interleukin 6 (IL-6) and tumour
necrosis factor alpha (TNF-a). There is also evidence tosuggest that the GLUT1 protein is a stress inducible
glucose transporter in human chondrocytes and is sig-
nificantly upregulated in cells under the influence of pro-
inflammatory cytokines (Shikhman et al., 2001a).
Therefore, enhanced glucose transport and glycolysis by
articular chondrocytes may be important pre-requisites
for cartilage degradation in degenerative diseases of ar-
ticulating joints. However, it is not yet known if the in-dividual GLUT isoforms expressed in equine articular
chondrocytes respond to growth factors and cytokines in
a similar way to human chondrocytes. It is also not
known if the stimulated glucose uptake in response to
these physiologically relevant mediators is the result of de
novo synthesis and upregulation of glucose transporters.
The primary objective of this study was to test a novel
hypothesis that may implicate chondrocyte glucosetransporters in osteoarticular disorders thus establishing
a link between chondrocyte bioenergetics, nutrient status
and joint disease. The hypothesis tested was that glucose
transporters in equine chondrocytes are differentially
regulated by growth factors and pro-inflammatory cy-
tokines whose pathophysiological and bioenergetic ef-
fects may exacerbate the phenotypic and molecular
alterations observed in degenerative joint disease. Ac-cordingly, the aim of the investigation was to determine
the effects of IL-1b, TNF-a, IGF-I and transforming
growth factor beta (TGF-b) on (1) the net glucose uptake
capacity of equine articular chondrocytes in monolayer
culture and (2) the expression of the GLUT1 andGLUT3
glucose transporter protein isoforms in equine articular
chondrocytes in alginate culture.
2. Materials and methods
2.1. Chemicals
All chemicals used in this study were molecular biol-
ogy grade and purchased from Sigma/Aldrich. 2-Deoxy-
DD-[2, 6-3H] glucose was from Amersham Biosciences,
protein assay kits were obtained from Bio-Rad and an-
tibodies were purchased from Chemicon International.
2.2. Cartilage source
Normal equine articular cartilage was obtained from
the stifle joints of three horses (age range: 5–21 years)
euthanased for clinical reasons at the Philip Leverhulme
Large Animal Hospital, University of Liverpool. The
study was conducted with ethical approval in strict ac-cordance with local guidelines (none of these three
horses were euthanased for the purpose of this study).
2.3. Chondrocyte isolation and culture
Equine cartilage shavings were rinsed with phos-
phate-buffered saline (PBS), cut into small slices and
incubated overnight with type I collagenase (EC 3.4.24.3from Clostridium histolyticum) (approximately 100 col-
lagen digestion units mL�1) in Dulbecco’s modified
Eagles medium (DMEM) supplemented with 1000
mgL�1 glucose, 10% fetal calf serum and 1% antibiotic/
antimycotic solution. The filtered cell suspension was
washed three times in fresh DMEM and the cells
counted on a haemocytometer; cell viability was deter-
mined by trypan blue dye exclusion and was usually 95%or higher. The cells (2� 106 cells/mL) were cultured in
alginate beads as previously described (Mobasheri et al.,
2002a; Richardson et al., 2003).
Two different culture protocols were used in this
study for the following reason: 2-deoxy-DD-[2,6-3H] glu-
cose uptake experiments can only be performed with
monolayer cultured chondrocytes as the final washes in
non-radioactive medium cannot be performed rapidlywith alginate beads. Cells used for uptake studies were
not passaged more than three times to prevent chon-
drocyte dedifferentiation and phenotypic instability.
Chondrocytes encapsulated and maintained in alginate
culture were used for Western blot analyses as this
method allows investigators to maintain equal cell
numbers thus ensuring equal protein loading on SDS–
PAGE.
2.4. Deoxy-DD-[2,6-3H] glucose uptake
Net glucose transport was determined by measuring
the uptake of non-metabolizable 2-deoxy-DD-[2,6-3H]
glucose into equine chondrocytes in 24-well plates in the
presence and absence of the glucose transport inhibitor

218 T. Phillips et al. / The Veterinary Journal 169 (2005) 216–222
cytochalasin B (1 lM). Most of the uptake experimentsdescribed were performed with chondrocytes stimulated
for 24 h with TNF-a (100 ngmL�1), IL-1b (100
ngmL�1), IGF-I (20 ngmL�1), TGF-b (20 ngmL�1)
and insulin (12.5 lgmL�1). Selected experiments were
carried to study the short-term effect of insulin on 2-
deoxyglucose uptake by pre-incubating chondrocytes
for 15 min with insulin (12.5 lgmL�1). The insulin used
in this study was derived from the bovine species. All thecytokines and growth factors used in this study were
human recombinants.
Prior to the commencement of the assay, the chon-
drocyte cultures were rinsed three times with PBS and the
uptake of the radiolabelled 2-deoxyglucose (1 lCi mL�1
culture medium) was assayed for 35 min at 20 �C in
modified DMEM lacking glucose, pyruvate and serum.
The 24-well plates were washed three times with ice-coldPBS and the chondrocytes lysed with a cell lysis solution
consisting of 0.5% sodium dodecyl sulphate and 0.5%
Triton X-100 in PBS. Aliquots (450 lL) of the cell lysis
solution were mixed with 3.55 mL of NACS104 aqueous
scintillation cocktail (Amersham Biosciences) and
counted in a scintillation counter. The remaining 50 lLaliquots of the cell lysis solution were used to determine
total cell protein content using a Bio-Rad detergentcompatible (DC) protein assay kit and 2-deoxyglucose
uptake was normalized to total cell protein. All uptake
experiments were carried out in triplicate and repeated
under identical conditions at least three times and the
data are presented as percentage change in total 2-deoxy-
DD-[2,6-3H] glucose uptake.
2.5. SDS–PAGE and western blot analysis
Briefly, chondrocytes from alginate cultures were
rinsed in PBS and proteins were extracted with lysis
buffer (50 mM Tris/HCl, pH 7.2, 150 mM NaCl, l% (v/v)
Triton X-100, 1 mM sodium orthovanadate, 50 mM
sodium pyrophosphate, 100 mM sodium fluoride, 0.01%
(v/v) aprotinin, 4 lg/mL pepstatin A, 10 lg/mL leupep-
tin, 1 mM PMSF) on ice for 30 min. After adjustment oftotal protein concentration using a Bio-Rad DC protein
assay kit, samples were separated by SDS–PAGE on
12% minigels under reducing conditions. Separated
proteins were transferred onto nitrocellulose membranes
(Sigma) and incubated in blocking buffer (5% (w/v)
skimmed milk powder in PBS/0.1% Tween 20) for 1 h at
room temperature (RT). Membranes were incubated
with polyclonal primary antibodies to GLUT1 andGLUT3 (Chemicon International) diluted 1 in 1000 in
blocking buffer for 1 h at RT, washed three times with
blocking buffer, and incubated with secondary goat anti-
rabbit antibody conjugated with alkaline phosphatase
for 30 min at RT. Membranes were rinsed in blocking
buffer and washed three times in 0.1 M Tris pH 9.5,
containing 0.05 M MgCl2 and 0.1 M NaCl. Specific an-
tigen-antibody complexes were visualised by incubatingwith premixed nitro blue tetrazolium (NBT) in con-
junction with 5-bromo-4-chloro-3-indolyl phosphate
(BCIP) as precipitating agent for alkaline phosphatase.
2.6. Statistical analysis
The results are expressed as the means� SE of a
representative experiment performed in triplicate. Themeans were compared using Student’s t test assuming
equal variances. P < 0:05 was considered statistically
significant.
2.7. Image analysis
Quantification of Western blots was carried out using
Scion Image for Windows (version 4.0.2 http://www.scioncorp.com/) based on NIH Image for Macintosh.
The GLUT1 and GLUT3 immunoblots were scanned in
128 shades of grey. Thresholding was used to segment
scanned blots into objects of interest displayed in black
(i.e. GLUT bands) and background displayed in white
based on the grey levels in the original blot.
3. Results
We found that phenotypically stable equine articular
chondrocytes were capable of transporting 2-deoxy-DD-
[2,6-3H] glucose, which is consistent with our previous
observations in human chondrocytes and provides, for
the first time evidence for the presence of functional
glucose transporters in equine articular chondrocytes.
3.1. 2-Deoxy-DD-[2,6-3H] glucose uptake is not signifi-
cantly influenced by insulin
Equine chondrocytes in 24-well plates were stimulated
with bovine insulin for up to 15 min (short term) at room
temperature or at least 24 h (long term) at 37 �C before
measuring net 2-deoxy-DD-[2,6-3H] glucose uptake. Up-take into untreated (control) chondrocytes was consid-
ered to be 100%. The specificity of uptake was determined
by addition of cytochalasin B at the point of assay initi-
ation.We found that short-term or long-term stimulation
with bovine insulin did not affect 2-deoxyglucose uptake
in a statistically significant manner (Fig. 1). Cytochalasin
B inhibited 2-deoxyglucose transport in control and in-
sulin stimulated cultures (P < 0:025). Short-term stimu-lation with insulin (15 min) resulted in a small but
statistically insignificant (P ¼ 0:22) increase in net 2-
deoxy-DD-[2,6-3H] glucose uptake. Long-term incubation
with insulin (24 h) also had no stimulatory effect on
glucose uptake (P ¼ 0:15). These observations suggest
that the insulin-responsive GLUT4 isoform is either not
expressed in equine articular chondrocytes or that its

**
*
P = 0.22
P = 0.15%
of
2-D
eoxy
-D-[
2,6-
3 H]
Glu
cose
Up
take
0
25
50
75
100
125
150
175
Control Control + Cyt B Insulin(Short Term)
Insulin(Short Term)
+ Cyt B
Insulin(Long Term)
Insulin(Long Term)
+ Cyt B
Fig. 1. 2-Deoxy-DD-[2,6-3H] glucose uptake is not significantly influenced by insulin stimulation of equine chondrocytes. Cells were stimulated with
insulin for 15 min (short term) at room temperature or 24 h (long term) at 37 �C. 2-deoxyglucose uptake into untreated (control) cells was considered
to be 100%. Specificity of uptake was determined cytochalasin B treatment during the assay period. Neither short-term nor long-term stimulation
with insulin resulted in a significant change in 2-deoxyglucose uptake. There was no significant increase in 2-deoxyglucose uptake following short
term insulin stimulation (P ¼ 0:22). Long-term stimulation with insulin resulted in a decrease in 2-deoxyglucose uptake (P ¼ 0:15). * Denotes a
significant difference (P < 0:025) between control and experimental groups.
T. Phillips et al. / The Veterinary Journal 169 (2005) 216–222 219
plasma membrane abundance is too low to affect netglucose uptake in response to insulin treatment in equine
articular chondrocytes.
3.2. Cytokine and endocrine stimulation of 2-deoxy-[2,6-3H] glucose uptake
Uptake of 2-deoxy-[2,6-3H] glucose was significantly
increased in equine articular chondrocytes pre-stimu-lated with IGF-I, TGF-b, TNF-a and IL-1b (Fig. 2). In
these experiments baseline 2-deoxyglucose uptake was
measured in un-stimulated (control) chondrocytes and
Fig. 2. Uptake of 2-deoxy-[2,6-3H] glucose by equine articular chondrocytes
glucose uptake in un-stimulated chondrocytes (control cells) was considere
* Denotes a significant difference between control and experimental groups
the value represented as 100%. Treatment with IGF-I,TGF-b, IL-1b and TNF-a resulted in increased 2-de-
oxyglucose uptake by over 65% compared to control
values (P < 0:025).
3.3. GLUT1 expression is upregulated by pro-inflamma-
tory cytokines and growth factors but only IGF-I up-
regulates GLUT3
We observed that the effects of growth factors and
pro-inflammatory cytokines on the individual GLUT
protein isoforms (i.e. GLUT1 and GLUT3) was not the
stimulated with IGF-I, TGF-b, TNF-a and IL-1b. Baseline 2-deoxy-
d as 100%. Error bars indicate standard errors of the means (n ¼ 3).
(P < 0:025).

220 T. Phillips et al. / The Veterinary Journal 169 (2005) 216–222
same despite the fact that the uptake of 2-deoxy-[2,6-3H]glucose was significantly increased in equine articular
chondrocytes pre-stimulated with IGF-I, TGF-b, TNF-
a and IL-1b. Western blot analysis of equine chondro-
cyte cell lysates demonstrated that equine chondrocytes
expressed the GLUT1 and GLUT3 proteins with ap-
parent molecular weights of between 55 and 60 kDa.
Previous studies have revealed similar molecular weights
for GLUT1 and GLUT3 in human chondrocytes.Stimulation of alginate cultures of equine chondrocytes
with IGF-I, TGF-b, TNF-a and IL-1b all resulted in
upregulation of GLUT1 (P < 0:05). In the case of
GLUT3, a similar upregulation was measured for cul-
tures exposed to IGF-I (P < 0:05). However TGF-bappeared to reduce the levels of GLUT3 protein and
TNF-a and IL-1b had no significant effect on the
abundance of GLUT3. Note that the same cells exhib-ited a large increase in 2-deoxyglucose uptake in re-
sponse to TGF-b treatment which suggests that while
GLUT3 was down-regulated in response to this growth
factor, GLUT1 (and possibly other GLUT isoforms)
may have been activated or significantly up-regulated,
resulting in an increase in net 2-deoxyglucose transport.
4. Discussion
The major findings of this study are summarized as
follows: (1) equine chondrocytes express functional glu-
cose transporters; (2) the uptake of 2-deoxy-DD-[2,6-3H]
glucose into equine chondrocytes is consistent with a fa-
cilitated, GLUT-mediated process for glucose transport;
(3) short-term insulin treatment resulted in a statisticallyinsignificant increase in the uptake of 2-deoxyglucose but
long-term insulin treatment had no effect on 2-deoxy-
glucose uptake; (4) IGF-I, IL-1b, TGF-b and TNF-a all
resulted in a significant (over 65%) increase in glucose
uptake compared to basal levels; (5) IGF-I stimulation of
equine chondrocytes over a period of 24 h resulted in
upregulation ofGLUT1andGLUT3proteins; (6) TGF-bapparently reduced the levels of GLUT3; (7) TNF-aand IL-1b stimulation upregulated GLUT1 expression
but down-regulated GLUT3 supporting the hypothesis
that growth factors and pro-inflammatory cytokines dif-
ferentially regulate the GLUT1 and GLUT3 glucose
transporters.
Articular cartilage is an avascular tissue in which
oxygen tensions are generally very low (Otte, 1991;
Mobasheri et al., 2002b). In the absence of oxidativephosphorylation, chondrocytes endure hypoxic condi-
tions by generating ATP by glycolytic breakdown of
glucose (Mobasheri et al., 2002a). Hypoxia generally in-
duces expression of several important genes including
vascular endothelial growth factor (VEGF), glucose
transporter-1 (GLUT1; SLC2A1) and glucose trans-
porter-3 (GLUT3; SLC2A3) (Semenza, 1999). Evidence
from other cell types in the literature suggests that ele-vated glucose uptake stimulated by pro-inflammatory
cytokines is accompanied by upregulation of GLUT
transporter proteins. In response to pro-inflammatory
cytokines a number of genes are activated including
VEGF and GLUT1 (Pufe et al., 2001; Shikhman et al.,
2001a; Richardson et al., 2003) resulting in the upregu-
lation of the proteins encoded by these genes in chon-
drocytes. These proteins are key mediators of nutrientand energy provision in degenerative conditions such as
osteoarthritis, which is particularly relevant to the equine
species; the glucose transporters provide the means for
accelerated glucose transport and enhanced substrate
utilization in response to stimulation by pro-inflamma-
tory, catabolic cytokines, including TNF-a, IL-6 and IL-
1b (Shikhman et al., 2001a; Shikhman et al., 2001b;
Richardson et al., 2003) and to chemokines such as con-nective tissue activating peptide III (Ku Tai et al., 1992).
Studies in chondrocytes from the growth plates of
rodents (Wang et al., 1999; Ohara et al., 2001) and in
human Meckel’s cartilage (Ishizeki et al., 2002) have
shown that the insulin-responsive GLUT4 isoform is
expressed and functional in these cells. However, we and
others have previously reported on the absence of in-
sulin-responsive glucose transporters in mature humanarticular chondrocytes (Shikhman et al., 2001a; Moba-
sheri et al., 2002a). The results presented in this study
(Fig. 1) support our earlier observations in human
chondrocytes (Richardson et al., 2003) and confirm that
insulin-responsive glucose transporters are not ex-
pressed in fully developed equine articular chondro-
cytes, and that insulin does not acutely increase glucose
transport in these cells. In the presence of high levels ofserum, chondrocytes from Meckel’s cartilage have been
shown to change phenotype and differentiate into
adipocytes (Ishizeki et al., 2002). There are no published
reports that suggest articular chondrocytes are capable
of undergoing a similar transdifferentiation.
The cytokines and growth factors used in this study
were human recombinants. This was due to the absence
of suitable equine counterparts. The observed differ-ences in the magnitude of cytokine and growth factor
stimulated 2-deoxyglucose transport (Fig. 2) is likely to
be due to the lower biological activity of human re-
combinant cytokines and growth factors on equine
chondrocytes. The magnitude of the changes we ob-
served in glucose uptake in response to IGF-I, IL-1band TNF-a was similar to the values previously reported
by our group (Richardson et al., 2003). However, theaugmented 2-deoxyglucose uptake was lower than val-
ues reported by another group for human chondrocytes
(Shikhman et al., 2001a).
In response to long-term TGF-b stimulation we
observed increased 2-deoxyglucose transport and up-
regulation of GLUT1 but GLUT3 appeared to be
down-regulated (Fig. 3). Studies in the kidney have
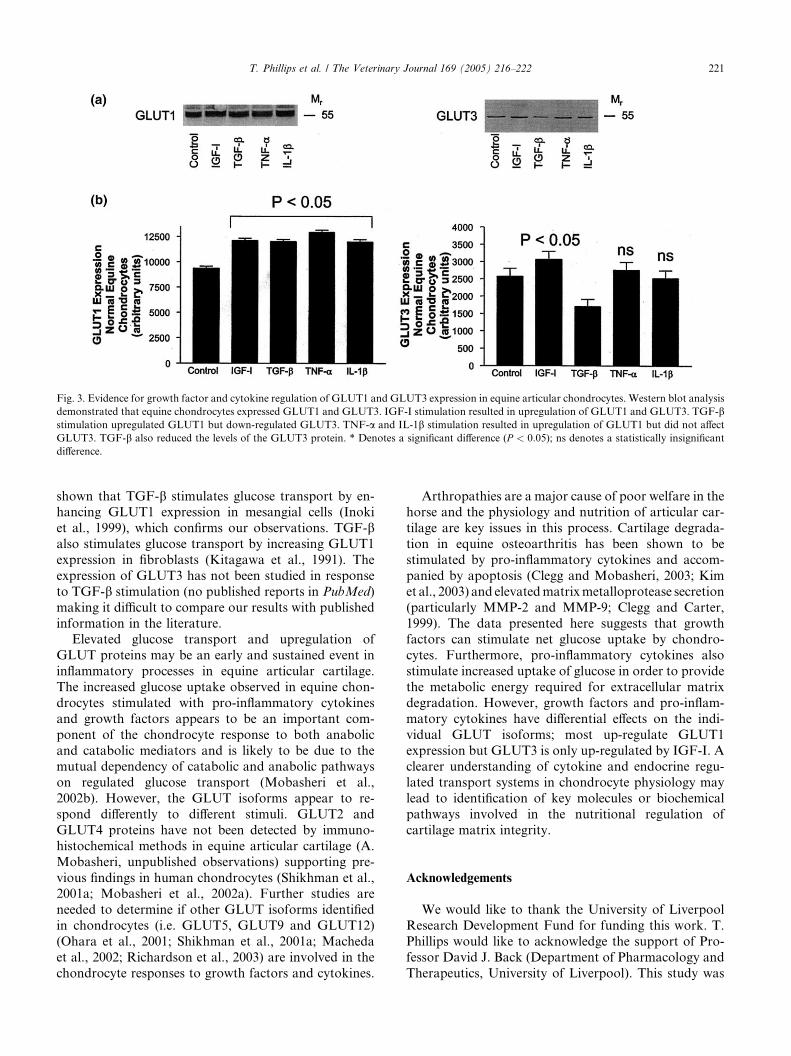
Fig. 3. Evidence for growth factor and cytokine regulation of GLUT1 and GLUT3 expression in equine articular chondrocytes. Western blot analysis
demonstrated that equine chondrocytes expressed GLUT1 and GLUT3. IGF-I stimulation resulted in upregulation of GLUT1 and GLUT3. TGF-bstimulation upregulated GLUT1 but down-regulated GLUT3. TNF-a and IL-1b stimulation resulted in upregulation of GLUT1 but did not affect
GLUT3. TGF-b also reduced the levels of the GLUT3 protein. * Denotes a significant difference (P < 0:05); ns denotes a statistically insignificant
difference.
T. Phillips et al. / The Veterinary Journal 169 (2005) 216–222 221
shown that TGF-b stimulates glucose transport by en-hancing GLUT1 expression in mesangial cells (Inoki
et al., 1999), which confirms our observations. TGF-balso stimulates glucose transport by increasing GLUT1
expression in fibroblasts (Kitagawa et al., 1991). The
expression of GLUT3 has not been studied in response
to TGF-b stimulation (no published reports in PubMed)
making it difficult to compare our results with published
information in the literature.Elevated glucose transport and upregulation of
GLUT proteins may be an early and sustained event in
inflammatory processes in equine articular cartilage.
The increased glucose uptake observed in equine chon-
drocytes stimulated with pro-inflammatory cytokines
and growth factors appears to be an important com-
ponent of the chondrocyte response to both anabolic
and catabolic mediators and is likely to be due to themutual dependency of catabolic and anabolic pathways
on regulated glucose transport (Mobasheri et al.,
2002b). However, the GLUT isoforms appear to re-
spond differently to different stimuli. GLUT2 and
GLUT4 proteins have not been detected by immuno-
histochemical methods in equine articular cartilage (A.
Mobasheri, unpublished observations) supporting pre-
vious findings in human chondrocytes (Shikhman et al.,2001a; Mobasheri et al., 2002a). Further studies are
needed to determine if other GLUT isoforms identified
in chondrocytes (i.e. GLUT5, GLUT9 and GLUT12)
(Ohara et al., 2001; Shikhman et al., 2001a; Macheda
et al., 2002; Richardson et al., 2003) are involved in the
chondrocyte responses to growth factors and cytokines.
Arthropathies are a major cause of poor welfare in thehorse and the physiology and nutrition of articular car-
tilage are key issues in this process. Cartilage degrada-
tion in equine osteoarthritis has been shown to be
stimulated by pro-inflammatory cytokines and accom-
panied by apoptosis (Clegg and Mobasheri, 2003; Kim
et al., 2003) and elevatedmatrixmetalloprotease secretion
(particularly MMP-2 and MMP-9; Clegg and Carter,
1999). The data presented here suggests that growthfactors can stimulate net glucose uptake by chondro-
cytes. Furthermore, pro-inflammatory cytokines also
stimulate increased uptake of glucose in order to provide
the metabolic energy required for extracellular matrix
degradation. However, growth factors and pro-inflam-
matory cytokines have differential effects on the indi-
vidual GLUT isoforms; most up-regulate GLUT1
expression but GLUT3 is only up-regulated by IGF-I. Aclearer understanding of cytokine and endocrine regu-
lated transport systems in chondrocyte physiology may
lead to identification of key molecules or biochemical
pathways involved in the nutritional regulation of
cartilage matrix integrity.
Acknowledgements
We would like to thank the University of Liverpool
Research Development Fund for funding this work. T.
Phillips would like to acknowledge the support of Pro-
fessor David J. Back (Department of Pharmacology and
Therapeutics, University of Liverpool). This study was

222 T. Phillips et al. / The Veterinary Journal 169 (2005) 216–222
also funded by a short-term fellowship granted to I.Ferraz from the Servicio de Reumatologia, Hospital
Universitario de Canarias, in Santa Cruz de Tenerife.
References
Clegg, P.D., Carter, S.D., 1999. Matrix metalloproteinase-2 and -9 are
activated in joint diseases. Equine Veterinary Journal 31, 324–330.
Clegg, P.D., Mobasheri, A., 2003. Chondrocyte apoptosis, inflamma-
tory mediators and equine osteoarthritis. The Veterinary Journal
166, 3–4.
Inoki, K., Haneda, M., Maeda, S., Koya, D., Kikkawa, R., 1999.
TGF-beta 1 stimulates glucose uptake by enhancing GLUT1
expression in mesangial cells. Kidney International 55, 1704–1712.
Ishizeki, K., Takahashi, N., Nawa, T., 2002. Phenotypic characteristics
of adipocytes generated from Meckel’s chondrocytes in response to
chick serum in vitro. Cell and Tissue Research 309, 251–260.
Joost, H.G., Thorens, B., 2001. The extended GLUT-family of sugar/
polyol transport facilitators: nomenclature, sequence characteris-
tics, and potential function of its novel members (review).
Molecular Membrane Biology 18, 247–256.
Joost, H.G., Bell, G.I., Best, J.D., Birnbaum, M.J., Charron, M.J.,
Chen, Y.T., Doege, H., James, D.E., Lodish, H.F., Moley, K.H.,
Moley, J.F., Mueckler, M., Rogers, S., Schurmann, A., Seino, S.,
Thorens, B., 2002. Nomenclature of the GLUT/SLC2A family of
sugar/polyol transport facilitators. American Journal of Physiology
Endocrinology and Metabolism 282, E974–E976.
Kim, D.Y., Taylor, H.W., Moore, R.M., Paulsen, D.B., Cho, D.Y.,
2003. Articular chondrocyte apoptosis in equine osteoarthritis. The
Veterinary Journal 166, 52–57.
Kitagawa, T., Masumi, A., Akamatsu, Y., 1991. Transforming growth
factor-beta 1 stimulates glucose uptake and the expression of
glucose transporter mRNA in quiescent Swiss mouse 3T3 cells.
Journal of Biological Chemistry 266, 18066–18071.
Ku Tai, P.-K., Liao, J.-F., Hossler, P.A., Castor, C.W., Carter-Su, C.,
1992. Regulation of glucose transporters by connective tissue
activating peptide-III isoforms. Journal of Biological Chemistry
267, 19579–19586.
Macheda, M.L., Kelly, D.J., Best, J.D., Rogers, S., 2002. Expression
during rat fetal development of GLUT12-a member of the class III
hexose transporter family. Anatomy and Embryology (Berlin) 205,
441–452.
Mobasheri, A., Neama, G., Bell, S., Richardson, S., Carter, S.D.,
2002a. Human articular chondrocytes express three facilitative
glucose transporter isoforms: GLUT1, GLUT3 and GLUT9. Cell
Biology International 26, 297–300.
Mobasheri, A., Vannucci, S.J., Bondy, C.A., Carter, S.D., Innes, J.F.,
Arteaga, M.F., Trujillo, E., Ferraz, I., Shakibaei, M., Martin-
Vasallo, P., 2002b. Glucose transport and metabolism in chondro-
cytes: a key to understanding chondrogenesis, skeletal development
and cartilage degradation in osteoarthritis. Histology and Histo-
pathology 17, 1239–1267.
Ohara, H., Tamayama, T., Maemura, K., Kanbara, K., Hayasaki, H.,
Abe, M., Watanabe, M., 2001. Immunocytochemical demonstra-
tion of glucose transporters in epiphyseal growth plate chondro-
cytes of young rats in correlation with autoradiographic
distribution of 2-deoxyglucose in chondrocytes of mice. Acta
Histochemica 103, 365–378.
Otte, P., 1991. Basic cell metabolism of articular cartilage. Manometric
studies. Zeitschrift f€ur Rheumatologie 50, 304–312.
Pufe, T., Petersen, W., Tillmann, B., Mentlein, R., 2001. The splice
variants VEGF121 and VEGF189 of the angiogenic peptide
vascular endothelial growth factor are expressed in osteoarthritic
cartilage. Arthritis and Rheumatism 44, 1082–1088.
Richardson, S., Neama, G., Phillips, T., Bell, S., Carter, S.D., Moley,
K.H., Moley, J.F., Vannucci, S.J., Mobasheri, A., 2003. Molecular
characterization and partial cDNA cloning of facilitative glucose
transporters expressed in human articular chondrocytes; stimula-
tion of 2-deoxyglucose uptake by IGF-I and elevated MMP-2
secretion by glucose deprivation. Osteoarthritis and Cartilage 11,
92–101.
Semenza, G.L., 1999. Regulation of mammalian O2 homeostasis by
hypoxia-inducible factor 1. Annual Review of Cell and Develop-
mental Biology 15, 551–578.
Shikhman, A.R., Brinson, D.C., Valbracht, J., Lotz, M.K., 2001a.
Cytokine regulation of facilitated glucose transport in human
articular chondrocytes. Journal of Immunology 167, 7001–7008.
Shikhman, A.R., Kuhn, K., Alaaeddine, N., Lotz, M., 2001b. N-
acetylglucosamine prevents IL-1 beta-mediated activation of
human chondrocytes. Journal of Immunology 166, 5155–5160.
Wang, J., Zhou, J., Bondy, C.A., 1999. Igf1 promotes longitudinal
bone growth by insulin-like actions augmenting chondrocyte
hypertrophy. FASEB Journal 13, 1985–1990.
Wu, X., Freeze, H.H., 2002. GLUT14, a duplicon of GLUT3, is
specifically expressed in testis as alternative splice forms. Genomics
80, 553–557.
















![Review Open Access€¦ · transporter 1 enzyme (GLUT1). GLUT1 improves the uptake of glucose[17] and induces glycolytic enzymes such as phosphoglycerate kinase[18]. In turn, phosphoglycerate](https://static.fdocuments.in/doc/165x107/5fa20fb8c4d32a0d83370841/review-open-access-transporter-1-enzyme-glut1-glut1-improves-the-uptake-of-glucose17.jpg)
