
Neuro Diagnostics Motor Unit Potentials MUPs Motor Unit Potentials MUPs.
Upload
werner-sommerCategory
view
212download
0
ELSEVIER Electroencephalography and clinical Neurophysiology 102 (1997) 192-199
Differential localization of brain systems subserving memory for names and faces in normal subjects with event-related potentials
W e r n e r S o m m e r a'*, E s t h e r K o m o s s b, S t e f a n R. S c h w e i n b e r g e r b
~Psychology Institute, Humboldt University at Berlin, Hausvogteiplatz 5 7, 10117 Berlin. German3' bUniversitv of Konstanz, Konstanz, Germany
Received 13 April 1995; revised version received 1 April 1996; accepted for publication: 30 August 1996
Abstract
Evidence from brain-damaged patients suggests different memory systems for verbal and nonverbal stimuli which also have different neuroanatomical substrates. We explored whether the scalp topographies of event-related potentials (ERPs) might confirm the dis- sociability of these systems. Unfamiliar faces and names were presented in separate conditions, study blocks alternating with recognition blocks. During recognition the familiarity of each stimulus was rated on a 4-point scale. The amplitudes of the ERPs to the study items were monotonically related in size to the mean subsequent familiarity ratings. Memory-related ERP differences (Dm) were calculated by subtracting ERPs recorded during the study phase to items with low subsequent familiarity ratings from items with high familiarity ratings. The Dm for faces and names showed stimulus-specific scalp topographies between 400 and 800 ms. These findings confirm that memory for names and faces is mediated by at least partially different brain systems. © 1997 Elsevier Science Ireland Ltd.
Keywords: Unfamiliar names and faces; Study and recognition; Memory-related ERP differences; Stimulus-specific scalp topographies
1. Introduction
In recent years evidence has accumulated that memory is not unitary, neither in function nor neuroanatomical substrate (Squire, 1987). The case for multiple memory systems is supported by dissociations between different tests of memory in brain-damaged patients. If patients are found that perform normal in one type of memory test but abnormal in a second type, the hypothesis is sup- ported that these tests probe into isolable cognitive sub- systems (Shallice, 1988).
In the domain of short-term or working memory, dis- sociations have been found between impaired auditory- verbal and relatively preserved visual-verbal store (War- rington and Shallice, 1969; Shallice and Vallar, 1990), impaired visual-verbal and preserved visual-spatial store (Kinsbourne and Warrington, 1962) and vice versa (De Renzi and Nichelli, 1975; De Renzi et al., 1977). For the auditory-verbal short-term store detailed analyses of
* Corresponding author.
patient performance have been presented, particularly within the framework of Baddeley's concept of working memory (Caplan and Waters, 1990). Outside of the domain of auditory-verbal short-term store, however, much less is known about isolable subsystems and about the neurophysiological mechanisms and neuroanatomical structures which subserve these functions. A major pro- blem for progress appears to be the rarity of selectively impaired patients and the lack of physiological methods for investigations in normal subjects.
In this paper we explore the usefulness of event-related potentials (ERPs) in differentiating physiological sub- strates of memory systems. We concentrate on a technique which has been introduced by Karis et al. (1984). These authors recorded ERPs to individually presented word sti- muli. Subsequently, these brain responses were sorted and averaged according to whether or not the ERP-eliciting word was recalled in a subsequent test. As it turned out, the ERPs during study were predictive of later recall; elec- trically more positive ERPs were found to recalled than to unrecalled words. As reviewed, for example by Kutas
0013-4694/97/$17.00 © 1997 Elsevier Science Ireland Ltd. All rights reserved PI1 S 0 9 2 1 - 8 8 4 X ( 9 6 ) 9 5 5 7 7 - 0 EEG 95577

196 W. Sommer et al. / Electroencephalog raphy and clinical Neurophysiology 102 (1997) 192-199
/
icantly during the 400-490 and the 500-790 ms segments (F(26, 286) = 3.20 and 4.50; P < 0.01 and 0.001, respec- tively) but not for the 300-390 ms segment (F = 1.27; P > 0.50). The across-time analysis did not indicate any segment-dependent changes of the Dm topography for faces ( F = 2.17; P = 0.07) or names ( F - 2 . 4 7 ; P = 0.21). In sum, we have observed material specific Dm- topographies at least between 400 and 790 ms.
4. Discussion
While previous reports have demonstrated a Dm either for words or for faces, this study performed the first direct comparison for these types of stimuli. Both the face and name Dm started at about 300 ms after stimulus onset and were of considerable duration; during the 800-1380 ms segment the face Dm was still significant and even the smaller name Dm was a statistical trend.
By considering difference waves between subsequently familiar and unfamiliar items in the topographical analyses we have eliminated all effects that are specific to the sti-
FACES NANES
Fig. 2. Topographical display of grand mean difference waves (Dm) between ERPs to high subsequent familiarity minus low subsequent familiarity, superimposed for faces (thick traces) and names (thin traces). The insert shows the nomenclature for the recording sites used. Positive is plotted upward.
which were performed as follows. First, difference waves, that is Dm, were calculated between ERPs from the study phase, subsequently rated as very familiar ( '4 ' ) and very unfamiliar ( '1 ' ) , respectively (Fig. 2). Then, amplitude measures were calculated for those time segments where both faces and names had shown significant familiarity effects in the preceding ANOVA. Fig. 3 depicts the scalp topographies of these Dm amplitude measures. It shows that for faces the Dm was positive around the ver- tex, slightly shifted to the right, surrounded by negative values. For names, the Dm was positive at anterior sites, shifted somewhat to the right and it was negative at poster- ior sites, shifted somewhat to the left. The Dm amplitudes within each condition and subject were then scaled across all electrodes, with the average distance from the mean, calculated from the grand mean ERPs, as the divisor. These scaled values were submitted to Huynh-Feldt cor- rected ANOVAs with repeated measurements on stimulus condition, time segment (300-390, 400-490, and 500- 790 ms), and electrode site.
This analysis revealed that the topographies of the Dm for faces and names were different (F(26, 2 8 6 ) = 2.0; P = 0.05), The Dm topography was not significantly modulated by the time segment interaction was clarified by Bonferoni-adjusted analyses of simple effects of the stimuli within time segments and within each kind of sti- mulus across time. The within-segment analysis indicated that Dm topographies for faces and names differed signif-
- 3 9 0
- 4 9 0
] 2 j J v
f ~500-790 ..... / , - , r - . r \
Fig. 3. Topographical displays of the scalp distributions of average amplitude measures of the Dm for faces and names at successive time segments.
W. Sommer et al. /Electroencephalography and clinical Neurophysiology 102 (1997) 192 199 195
FACES NAMES
Pz I ~ . b" I I I I I I I ' V "1 I I I I I I
+ D M er~r ~ - I I I I I I
vEOG
HEOG
+ [ zo ,uv
L
~t~ I , I I I 1 J. . . .J
o 1 s
S U B S E Q U E N T F A M I L I A R I T Y
H I G H LOt4
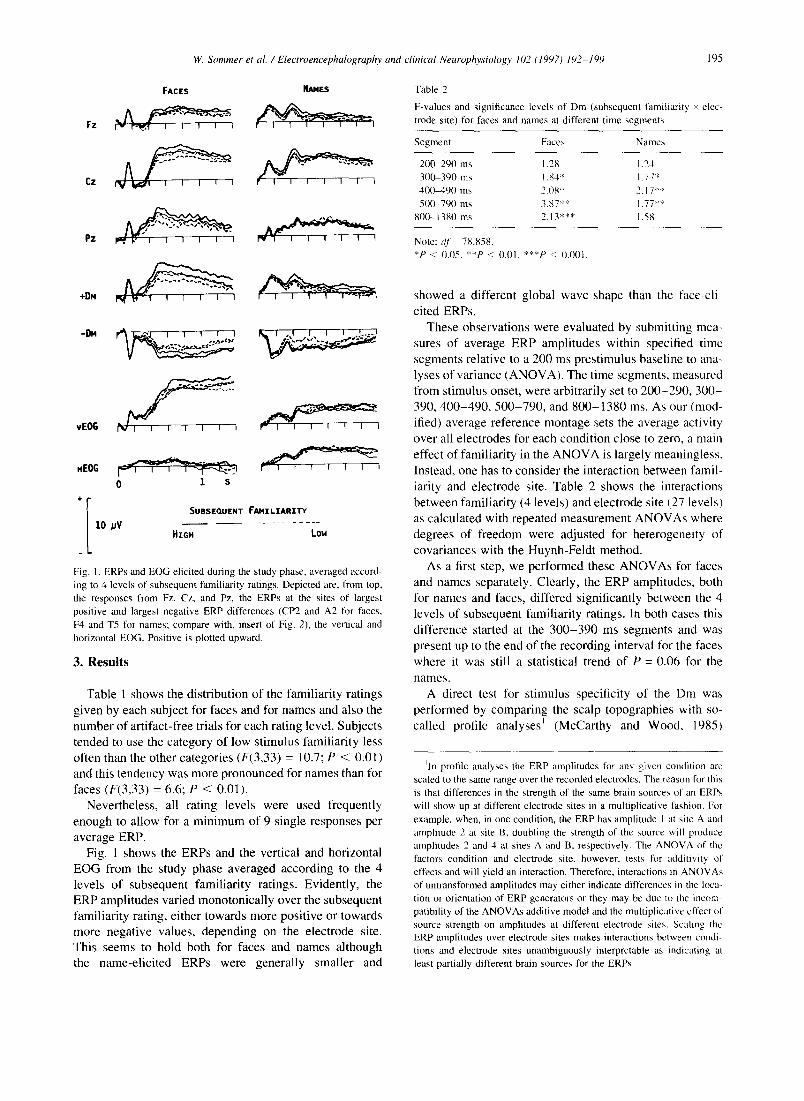
Fig. 1. ERPs and EOG elicited during the study phase, averaged accord- ing to 4 levels of subsequent familiarity ratings. Depicted are, from top, the responses from Fz, Cz, and Pz, the ERPs at the sites of largest positive and largest negative ERP differences (CP2 and A2 for faces, F4 and T5 for names; compare with, insert of Fig. 2), the vertical and horizontal EOG. Positive is plotted upward.
3. Results
Table 1 shows the distribution o f the famil iar i ty ratings
g iven by each subject for faces and for names and also the
number of art ifact-free trials for each rating level. Subjects
tended to use the ca tegory o f low stimulus famil iar i ty less
often than the other categories (F(3,33) = 10.7; P < 0.01)
and this tendency was more pronounced for names than for
faces (F(3,33) = 6.6; P < 0.01).
Never theless , all rating levels were used frequent ly
enough to al low for a m i n i m u m of 9 single responses per
average ERP. Fig. 1 shows the ERPs and the vertical and horizontal
E O G from the study phase averaged according to the 4
levels of subsequent famil iar i ty ratings. Evident ly, the
ERP ampli tudes varied monotonica l ly over the subsequent
famil iar i ty rating, either towards more posi t ive or towards
more negat ive values, depending on the electrode site.
This seems to hold both for faces and names al though
the name-el ic i ted ERPs were general ly smaller and
Table 2
F-values and significance levels of Dm (subsequent familiarity × elec- trode site) for faces and names at different time segments
Segment Faces Names
200-290 ms 1.28 1.24 300-390 ins 1.84" 1.77" 400-490 ms 2.08* 2.17** 5(10-790 ms 3.87** 1.77"*
800-1380 ms 2.13"** 1.58
Note: dr= 78,858. *P < 0.05. **P < 0.01. ***P < 0.001.
showed a different global wave-shape than the face-el i-
cited ERPs.
These observat ions were evaluated by submit t ing mea-
sures of average ERP ampli tudes within specified t ime
segments relat ive to a 200 ms prest imulus basel ine to ana-
lyses of var iance (ANOVA) . The t ime segments , measured
f rom stimulus onset, were arbitrarily set to 2 0 0 - 2 9 0 , 3 0 0 -
390, 4 0 0 - 4 9 0 , 5 0 0 - 7 9 0 , and 8 0 0 - 1 3 8 0 ms. As our (mod-
ified) average reference montage sets the average activity
over all e lectrodes for each condi t ion close to zero, a main
effect of famil iar i ty in the A N O V A is largely meaningless.
Instead, one has to consider the interaction be tween famil-
iarity and electrode site. Table 2 shows the interactions
be tween famil iar i ty (4 levels) and electrode site (27 levels)
as calculated with repeated measurement A N O V A s where
degrees o f f reedom were adjusted for heterogenei ty of
covar iances with the Huynh-Feld t method.
As a first step, we per formed these A N O V A s for faces
and names separately. Clearly, the ERP amplitudes, both
for names and faces, differed significantly be tween the 4
levels of subsequent famil iar i ty ratings. In both cases this
difference started at the 3 0 0 - 3 9 0 ms segments and was
present up to the end o f the recording interval for the faces
where it was still a statistical trend of P = 0.06 for the
names.
A direct test for st imulus specificity of the Dm was
per formed by compar ing the scalp topographies with so-
cal led profile analyses I (McCarthy and Wood, 1985)
~ln profile analyses the ERP amplitudes for any given condition are scaled to the same range over the recorded electrodes. The reason fl~r this is that differences in the strength of the same brain sources of an ERPs will show up at different electrode sites in a multiplicative fashion. For example, when, in one condition, the ERP has amplitude I at site A and amplitude 2 at site B, doubling the strength of the source will produce amplitudes 2 and 4 at sites A and B, respectively. The ANOVA of the factors condition and electrode site. however, tests for additivity of effects and will yield an interaction. Therefore, interactions in ANOVAs of untransformed amplitudes may either indicate differences in the loca- tion or orientation of ERP generators or they may be due to the incom- patibility of the ANOVAs additive model and the muhiplicativc effect of source strength on amplitudes at different electrode sites. Scaling the ERP amplitudes over electrode sites makes interactions between condi- tions and electrode sites unambiguously interpretable as indicating at least partially different brain sources for the ERPs.

194 W. Sommer et al. / Electroencephalography and clinical Neurophysiology 102 (1997) 192-199
these names were very uncommon to the subjects, who mostly came from the south of Germany, they still appeared name-like, containing name-typical syllables or being related to known names. Examples for the names used are: Aabke, Bajinck, Clamer, Daco, Eilhard, Gedrich, Folbert, Hellmer, Sentje, and Taede. The names were writ- ten in upper and lower case letters and subtended visual angles of 0.29 ° in the vertical and between 1.15 and 2.86 ° in the horizontal direction. Average luminance of the names was 2.2 cd/m 2.
2.3. Procedure
The subjects were seated in an electrically shielded, sound-attenuated chamber where a masking noise of 38 dB(A) was provided by the ventilation. A fixed chin rest provided for a constant viewing distance of 1.0 m. Each trial, during both the study and recognition conditions, began with the presentation of a homogeneously gray area of 1.5 and 8.2 cd/m 2 for the name and face condition, respectively. For faces this was an upright ellipse of the same size and shape as the face stimuli, for names it was a horizontal bar of 73 by 8 mm. The gray area appeared for 200 ms and was replaced for 200 ms by one of the faces or names after which the gray area reappeared for another 1200 ms.
During the recognition blocks a familiarity rating was requested for each stimulus, 500 ms after the disappear- ance of the gray area. A horizontal bar (19.0 by 3.1 cm, 3.0 cd/m 2) was presented on the screen, subdivided by vertical lines into 4 equal rectangles. These rectangles corre- sponded to 4 levels of familiarity ratings from high to low which were selected with the mouse-driven cursor of the Amiga. The cursor was only visible during the rating process.
The order of the face and name conditions was alter- nated over subjects. In both stimulus conditions 21 study blocks alternated with 21 recognition blocks of 10 stimuli each. In each recognition block the same 10 stimuli were shown as in the preceding study block in randomized pre- sentation order. The first pair of blocks in each stimulus condition served practice purposes and was not analyzed. The inter-stimulus interval during study was 6.0 s. During recognition the interval between a rating and the presenta- tion of the next stimulus was 4.8 s. The absolute minimum interval between study and recognition test for a given item was about 10 s with an upper limit of about 2.5 rain.
During study, subjects were instructed to watch each stimulus attentively but not to make any particular efforts to memorize it. During recognition the task was described as judging familiarity of the stimuli after having seen them once, rather than as a test of memory performance. The distractor-free recognition procedure (Wallace, 1982) was introduced in order to minimize elaborative mnemonic strategies and to keep subjects from always rating famil- iarity as high which would be the most sensible thing to do
under performance conditions because only old items were presented. Subjects were advised also to avoid eye move- ments and blinks while the gray area was visible. They were paid DM40.00 for the 4 h experimental session.
2.4. Electrophysiological recordings
The EEG was recorded from 19 electrodes according to the 10-20 system (Jasper, 1958; Fz, Cz, Pz, Fpl, Fp2, F3, F4, C3, C4, P3, P4, O1, 02, F7, F8, T3, T4, T5, T6). Six further electrodes were placed in the center of the quad- rangles formed by Fz, F3, Cz, and C3 ( 'FCI ' ) , by Cz, C3, Pz, and P3 ( 'CPI ' ) , and by C3, T3, P3, and T5 ( 'TPI ' ) and their contralateral homologues 'FC2', 'CP2', and 'TP2'. Finally, 2 electrodes (A1 and A2) were placed on the mastoid processes such that they formed diamond-shaped quadrangles with the T4, T6, and TP2 and with the T3, T5, and TP1 sites, respectively (Fig. 2). All electrodes were made of tin and mounted in an electrode cap (Electrocap Company). Vertical eye movements and eye blinks were recorded with an electrode below the right eye versus Fp2 and horizontal eye movements were monitored with elec- trodes at both outer canthi of the eyes. Initial recordings were referenced to the right mastoid electrode (A2). The EEG and EOG were amplified with a bandpass from 0.016 to 35 Hz and recorded with a sampling rate of 100 Hz for an epoch of 1.6 s, starting 200 ms prior to stimulus onset.
Each EEG and EOG channel was screened for activity ranges greater than I00 #V during the recording interval and for channel blockings. Only artifact-free ERPs to the faces and names from the study phase were analyzed. After averaging, the waveforms were digitally low-pass filtered at 8.8 Hz (-3 dB (Ruchkin and Glaser, 1978)) and recalculated to an average reference (Lehmann, 1987). In order to evenly represent scalp area, the value of the average reference was based on the 19 locations of the 10-20 system only, without the 8 non-standard electro- des. These values were then used to calculate average referenced activities for all 27 channels.
Table 1
Mean frequencies of familiarity ratings for faces and names and number of artifact-free trials in the study phase for the corresponding rating category (SDs in parenthesis)
Familiarity rating
1 (Low) 2 3 4 (High)
Faces 30.8 (9.4) 55.8 (12.6) 58.8 (9.9) 54.8 (21.1) a 22.5 (11.3) 42.5 (10.6) 48.6 (12.4) 51.7 (20.6) b Names 24.8 (8.5) 44.8 (16.3) 61.3 (11.3) 69.2 (27.0) 22.3 (9.2) 38.4 (12.8) 48.9 (11.5) 54.6 (26.8)
aAll trials.
bArtifact-free trials.

W. Sommer et al. / Electroencephalography and clinical Neurophysiology 102 (1997) 192 199 193
(1988) and Rugg (1995) this basic finding has been repli- cated many times. Pallet et al. (1987) have suggested the general term 'Dm' for any differences between ERPs eli- cited by study items dependent on subsequent memory performance, which we will adopt here also.
A number of findings indicate that the Dm is in fact sensitive to memory-specific processes. The Dm depends in its latency and scalp topography on whether subjects use simple rote or elaborative learning strategies (Karis et al., 1984; Fabiani et al., 1986). Sommer et al. (1991) showed that a Dm to pictures of unfamiliar faces was independent of fluctuations in attentional and perceptual factors during the study phase. Sommer et al. (1995) confirmed the Dm for faces and provided support for the idea that the Dm during encoding may be intimately related to facial dis- tinctiveness. It is well-known that distinctive faces are better recognized than typical faces (Bruce, 1988). Som- mer et al. (1995) found a remarkable similarity between the topographies of the Dm and those aspects in the face- elicited ERP that were related to the difference in high and low distinctiveness ratings. Scalp topography is not only important in distinguishing ERP components (Gaillard, 1988); differences in topography are also an unambiguous indicator for at least partially distinct underlying brain sources (McCarthy and Wood, 1985). The similarity of the topographies of the Dm and of the distinctiveness effect in the Sommer et al. study suggests that the Dm during encoding may be intimately related to facial distinctiveness. Pallet (1990) provided some evidence that the Dm is sensitive to established differences in mem- ory systems. He compared the Dm during intentional learning and explicit subsequent retrieval with the Dm elicited during incidental learning and implicit retrieval. In the former condition he obtained a frontal-positive Dm while in the latter condition the Dm seemed to be distrib- uted more posteriorly. Therefore, Paller's results also indi- cate that different brain sources may be involved in implicit and explicit memory, which in turn confirms the dissociations found between these aspects of memory with other approaches (Squire et al., 1993). Finally, Besson and Kutas (1993) found the Dm elicited by the final word of a sentence to vary with linguistic context and word meaning which also modulated the repetition effect in the ERPs. In addition, these authors found a similar scalp topography for Dm and the repetition effect, indicating similar under- lying processes.
The dissociations between verbal and spatial short-term store mentioned above suggest that different cognitive subsystems are involved in these tasks (Shallice, 1988) and that they are supported by different anatomical sub- strates. A type of stimulus which is often considered to be prototypically nonverbal are pictures of unfamiliar faces because it is extremely difficult to memorize or recognize them on the basis of verbal labels. There are also a number of studies which have shown that memory for faces can be dissociated from memory for verbal stimuli. While
impaired verbal memory is typically found after left- brain damage, impairments in recognition of unfamiliar faces are often found after right-brain damage (Milner, 1968; Vilkki, 1987; Warrington, 1984). Therefore, by comparing verbal stimuli and faces one might expect to maximize differences between a putative visual-verbal and a visual-spatial or nonverbal memory system.
In the present study we investigated whether the topo- graphy of the Dm is sensitive to differences in memory processes for unfamiliar faces and names. The Dm studies which used verbal stimuli have usually found a unipolar positive Din. Although this contrasts with the topography of the Dm for faces reported by Sommer et al. (1991), this cannot be taken as conclusive evidence for different brain sources underlying the word and face Dm. Apart from numerous other methodological differences, Sommer et al. have used an average reference electrode montage, while most other studies have used mastoids or earlobes as common reference sites. We therefore decided to directly compare face and name Dm with as many other factors held constant as possible. If Dm topography is specific for the types of stimuli used, this would confirm the existence of different neuroanatomical substrates and validate the use of event-related potentials as a toot in distinguishing memory systems in normal subjects.
2. Method
2.1. Subjects
The data of 12 subjects (6 males, mean age 25.9 years) could be used in the analysis. Four further subjects were lost because of technical problems and one had to be dropped because his ratings were not variable enough tbr calculating several rating-dependent ERPs. The remaining subjects were all strongly right-handed according to the Edinburgh Handedness Inventory (Oldfield, 1971), had right-handed first-degree relatives and had normal or cor- rected-to-normal visual acuity.
2.2. Stimuli
Two-hundred-ten photographs of male college students with neutral facial expression and without peculiar fea- tures like beards or spectacles were video-digitized from 1949 and 1950 year-books. All faces were turned slightly to the viewer's right and were fit into oval frames just above the hairline and below the chin. For examples of 'unframed' faces see Sommer et al. (1991). The frames subtended visual angles of 6.5 x 6.0 °. The faces were pre- sented with an average luminance of 8.2 cd/m 2 on the screen of a Commodore Amiga 2000 microcomputer.
For the verbal condition a list of 210 extremely unfami- liar male names with 4-10 characters were selected from a compilation of 10th to 16th century northern Germany (Friesland) parish registers (Brons, 1877). Although

W. Sommer et al. /Electroencephalography and clinical Neurophysiology 102 (1997) 192-199
Faces Names
197
[ ] + 5 2 ~ + 3 0 + 2 7
+ 2 } {} +2 i
U +i ~:: I + I a I r a + l ! [ ] +(~ g
[ ] +!~ ~,
[ ] +2 % ? + 2 2 +28 + ! ?
{{} + i i
B +~.,4 Im +gZ
i go [ ] -0
n .o 7;:,~
Fig. 4. Same as for the 4 0 0 - 4 9 0 ms segment but with a spline
muli but do not relate to memory. Thus, we have elimi- nated all effects in the ERPs that may relate to overall differences between faces and words in stimulus shape, intensity, contrast and so forth. By scaling the difference waves we have also excluded any effects that may relate to differences in the overall amplitudes between faces and words. All that is left are those effects in the ERP that have to do with the recognition process.
From the topographical comparisons of these famili- arity-dependent difference waves, 2 conclusions can be drawn. First, it reveals that the Dm for faces is a separate component from the Dm observed for words. This adds a further variety to the ERP components that have been found to predict memory performance. We note here that our name Dm resembles the Dm previously found to words. If the A1 and A2 electrodes are taken as reference points, as has been done in most other studies, then the negative aspects of the Dm at posterior sites would vanish and the remaining positive Dm would culminate at frontal sites (Fig. 4). The same procedure applied to the face Dm would leave the (positive) maximum over parietal sites. Also the present face Dm is quite compatible with the ones observed by Sommer et al. (1991) and Sommer et al. (1995). The more anterior border between the positive and negative areas in those studies is probably due to the fact that fewer electrodes had been used over frontal scalp sites where the face Dm is negative. The average reference balances positive and negative voltages over all electrodes not only for each ERP but also for the difference between any 2 ERPs. The absence of several electrodes with nega-
interpolat ion and recalculated to a c o m m o n linked masto id reference.
tive Dm in the studies of Sommer and coworkers may have caused a more negative reference-level and thus an enlar- gement of the scalp area with positive Dm.
The second and most important conclusion from the profile analysis of the face and name Dm is that they originate from different sources in the brain. As efforts had been made to control for stimulus meaning, frequency, gender, and category level, presentation and response modes, it is unlikely that the topographical differences relate to these factors. The present results therefore corro- borate neuropsychological findings that memory for non- verbal and, particularly, face stimuli relates to different brain structures than does memory for verbal stimuli. The differences in the intracranial sources of the Dm are most clear between 500 and 790 ms after stimulus onset but they may emerge as early as 300 ms after stimulus onset. During the time range for which we have analyzed the Dm there was no indication of changes in topographies over time. Fig. 3 even shows a remarkable stability of the stimulus-specific Dm topographies. Therefore, it is plausi- ble, albeit not logically compelling, to assume that the invariance of the Dm during this interval indicates time invariance of the specific intracranial sources.
The experimental design used here with its retention intervals between 10 s and 2.5 min is not uncommon in the study of memory-related brain potentials. However, it does not allow an unambiguous interpretation of the results in terms of short-term versus long-term memory. The interval used is short enough to allow for effects of short-term storage but there may be contributions of

198 W. Sommer et al. / Electroencephalograph 3, and clinical Neurophysiology 102 (1997) 192-199
long-term memory also. It will be one of the issues of future research to clarify the differential contribution of short- and long-term memory to the memory-related brain potentials.
Although the profile analyses demonstrate differential brain sources for the face and name Dm, they do not loca- lize these sources. As it is well known, there is no unique solution to the inverse problem of finding the intracranial source(s) that give rise to a potential distribution at the scalp without additional limiting assumptions, derived for example from animal experiments or studies of brain-damaged patients. As there appear to be no suffi- ciently strong limiting assumptions for the conditions of this study, we refrain from speculating on the localization of the generators of the Dm.
A remarkable feature of the present findings is the monotonicity of the relationship between the ERP ampli- tudes and the subsequent familiarity over 4 rating levels for both stimulus types. Incidentally, the very small ampli- tude differences between the intermediate rating cate- gories (Fig. 1) indicate that 3 point scales for the fam- iliarity ratings might have been sufficient. It is also note- worthy that at least for the faces and when considering the extremes of the familiarity ratings we have obtained Dm amplitudes of up to 5 #V (adding maximal positive and negative amplitudes) which is among the largest Dm effects reported to date. As our previous study with faces (Sommer et al., 1991) yielded sizable Dms between 3 and 4/~V even under more difficult conditions (there were 20 faces in each study block and an additional rating was required) and by merely dichotomizing the stimuli, it appears that unfamiliar faces are fairly powerful stimuli for eliciting Dm.
Although no distractor stimuli were used during the recognition phase, one can still expect the subjects to give honest ratings about stimulus familiarity (Ley and Long, 1987; Wallace, 1982). The present results also demonstrate that distractor items are not necessary in order to obtain Dm in recognition tasks. Therefore, dis- tractor-free recognition may be an interesting task variant for future Dm studies which is parsimonious as regards experimental time and stimulus material. The present find- ings validate the Dm as a tool for testing the differential localization of memory systems. One might now use the Dm topography in order to explore differential localization of memory-related brain processes specific, for example, to more subtle differences in stimulus material, retention intervals, encoding strategies and so on (Ellis and Young, 1989). In general, topographical dissociations indicate isolable underlying brain systems and may provide useful evidence in constructing taxonomies for multiple memory systems. In certain cases, we may also use the topography of the Dm in order to test hypotheses about the brain structures that relate to particular memory disorders, for example of the auditory-verbal short-term store (Caplan and Waters, 1990; Warrington et al., 1971 ).
Acknowledgements
We thank Juliana Matt and Hartmut Leuthold for their helpful comments on earlier versions of this manuscript.
References
Besson, M. and Kutas, M. The many facets of repetition: a cued-recall and event-related potential analysis of repeating words in same versus different sentence contexts. J. Exp. Psychol.: Learn., Mem., Cognit.. 1993 19: 1115-1133.
Brons, B. Jr. (1877). Friesischc Namen und Mittheilungen dartiber. Emden: W. Heynel. Reprinted 1972. Walluf: M. Sgndig.
Bruce, V. 11988). Recognising Faces. Hove and London: LEA. Caplan, D. and Waters, G. Short-term memory and language comprehen-
sion: a critical review of the neuropsychological literature. In: G. Vallar and T. Shallice (Eds.I, Neuropsychological Impairments of Short-term Memory. Cambridge University Press, Cambridge. MA. 1990, pp. 337-389.
De Renzi, E. and Nichelli, P. Verbal and non-verbal short-term memory impairment following hemispheric damage. Cortex, 1975, I1:341 354.
De Renzi. E., Faglioni, P. and Previdi. P. Spatial memory and hemi- spheric locus of lesion. Cortex. 1977, 13: 424-433.
Fabiani, M., Karis, D. and Donchin. E. P300 and recall in an incidental memory paradigm, Psychophysiology, 1986, 23: 298-308.
Galliard, A.W.K. Problems and paradigms in ERP research. Biol. Psy- chol., 1988, 26: 91-109.
Jasper. H.H. Report of the committee on methods of clinical examination in electroencephalography. Electroenceph. clin. Neurophysiol., 1958, 10: 370-375.
Karis, D., Fabiani, M. and Donchin, E. 'P300' and memory: individual differences in the von Restorff effect. Cognit. Psychol., 1984, 16: 177 216.
Kinsbourne, M. and Warrington, E.K. A disorder of simultaneous form perception. Brain. 1962, 85: 461-486.
Kutas. M. Review of event-related potential studies of memory. In: M.S. Gazzaniga (Ed.), Perspectives in Memory Research. MIT Press, Cam- bridge, MA, 1988, pp. 181-217.
Lehmann. D. Principles of spatial analysis. In: A.S. Gevins and A. Remond (Eds.), Handbook of Etectroencephalography and Clinical Neurophysiology, Vol. I, Methods of Analysis of Brain Electrical and Magnetic Signals. Elsevier, Amsterdam, 1987, pp. 309-354.
Ley, R. and Long, K. A distractor-li'ee test of recognition and false recognition. Bull. Psychonom. Soc., 1987, 25:411-414.
McCarthy, G. and Wood, C.C. Scalp distributions of event-related poten- tials: an ambiguity associated with analysis of variance models. Elec- troenceph. Clin. Neurophysiol., 1985, 62. 203-208.
Milner. B. Visual recognition and recall after right temporal-lobe exci- sion in man. Neuropsychologia, 1968.6: 191-209.
Oldfield, R.C. The assessmcnt and analysis of handedness: The Edin- burgh Inventory. Neuropsychologia. 1971, 9: 97-I13,
Paller, K. Recall and stem-complefion priming have different electro- physiological correlates and are modified differentially by directed forgetting. J. Exp. Psychol.: Learn.. Mere., Cognit., 1990, 16, 1021 1032.
Pallet, K.A., Kutas, M. and Mayes, A.R. Neural correlates of encoding in an incidental learning paradigm. Electroenceph. clin. Neurophysiol.. 1987, 67: 360-371.
Ruchkin, D. and Glaser, E.M. Simple digital filters for examining CNV and P300 on a single trial basis. In: D.A. Otto (Ed.) Multidisciplinary Perspectives in Event-Related Brain Potential Research. US Environ- mental Protection Agency, Washington, DC, SPA-600/9-77-043, 1978, pp. 579-581.
Rugg, M.D. ERP studies of memory. In: M.D. Rugg and M.G.H. Coles

W. Sommer et al. /Electroencephalography and clinical Neurophysiology 102 (1997) 192-199
(Ed~.), Electrophysiology of Mind. Event-Related Potentials and Cog- nition, Oxford University Press, Cambridge, MA, 1995, pp. 132-170.
Shallice, T. From neuropsychology to mental structure. Cambridge Uni- versity Press, Cambridge, MA, 1988.
Shallice, T. and Vallar, G. The impairment of auditory-verbal short-term storage. In G. Vallar and T. Shallice (Eds.), Neuropsychological Im- pairments of Short-Term Memory. Cambridge University Press, Cam- bridge, MA, 1990, pp. 11-53.
Sommer, W., Schweinberger, S.R. and Matt, J. Human brain potential correlates of face encoding into memory. Electroenceph. clin. Neuro- physiol., 1991, 79: 457-463.
Sommer, W., Heinz, A., Leuthold, H., Matt, J. and Schweinberger, S.R. Metamemory, distinctiveness, and event-related potentials in recogni- tion memory for faces. Mem. Cognit., 1995, 23, 1-11.
199
Squire, L.R. Memory and Brain. Oxford University Press, Cambridge, MA, 1987.
Squire, L.R., Knowlton, B.J., and Musen, G. The structure and organiza- tion of memory. Annu. Rev. Psychol., 1993, 44: 453-495.
Vilkki, J. Incidental and deliberate memory for words and faces after focal cerebral lesions. Neuropsychologia, 1987, 25, 221-230.
Wallace, W.P. Distractor-free recognition tests of memory. Am. J. Psy- chol., 1982: 421-440.
Warrington, E.K. Recognition Memory Test. Windsor: Nelson, 1984. Warrington, E.K. and Shallice, T. The selective impairment of auditory
verbal short-term memory. Brain, 1969, 92: 885-896. Warrington, E.K., Logue, V. and Pratt, R.T.C. The anatomical localiza-
tion of selective impairment of auditory verbal short-term memory. Neuropsychologia, 1971, 9: 377-387.


















