
Deletion of a dynamic surface loop improves stability and changes kinetic behavior of...
9
Deletion of a Dynamic Surface Loop Improves Stability and Changes Kinetic Behavior of Phosphatidylinositol-Synthesizing Streptomyces Phospholipase D Jasmina Damnjanovi c, Hideo Nakano, Yugo Iwasaki Laboratory of Molecular Biotechnology, Graduate School of Bioagricultural Sciences, Nagoya University, Furo-cho, Chikusa-ku, Nagoya 464-8601, Japan; telephone: þ81-52-789-4143; fax: þ81-52-789-4145; e-mail: [email protected] ABSTRACT: Supplementary phosphatidylinositol (PI) was shown to improve lipid metabolism in animals, thus it is interesting for pharmaceutical and nutritional applications. Homogenous PI can be produced in transphosphatidylation of phosphatidylcholine (PC) with myo-inositol catalyzed by phospholipase D (PLD). Only bacterial enzymes able to catalyze PI synthesis are Streptomyces antibioticus PLD (SaPLD) variants, among which DYR (W187D/Y191Y/Y385R) has the best kinetic profile. Increase in PI yield is possible by providing excess of solvated myo-inositol, which is achievable at high temperatures due to its highly temperature- dependent solubility. However, high-temperature PI synthe- sis requires the thermostable PLD. Previous site-directed combinatorial mutagenesis at the residues of DYR having high B-factor yielded the most improved variant, D40H/ T291Y DYR, obtained by the combination of two selected mutations. D40 and T291 are located within dynamic surface loops, D37-G45 (termed D40 loop) and G273-T313. Thus, in this work, thermostabilization of DYR SaPLD was attempted by rational design based on deletion of the D40 loop, generating two variants, D37-45 DYR and D38-46 DYR PLD. D38-46 DYR showed highest thermostability as its activity half-life at 70 C proved 11.7 and 8.0 times longer than that of the DYR and D37-45 DYR, respectively. Studies on molecular dynamics predicted D38-46 DYR to have the least average RMSD change as temperature dramatically increases. At 60 and 70 C, both mutants synthesized PI in a twofold higher yield compared to the DYR, while at the same time produced less of the hydrolytic side-product, phosphatidic acid. Biotechnol. Bioeng. 2014;111: 674–682. ß 2013 Wiley Periodicals, Inc. KEYWORDS: phospholipase D; flexible loops; stability Introduction Naturally occurring phosphatidylinositol (PI) is a phospho- lipid (PL) present mainly in eukaryotic membrane structures. PI and its metabolites play important roles in glycolipid anchoring of proteins, behave as secondary messengers in signal transduction and participate in mRNA export from the nucleus (Gardocki et al., 2005). Supplementary PI was reported to decrease triacylglycerol levels in serum and liver of model animals, rats, and mice (Yanagita, 2003; Yanagita and Nagao, 2008), and prevents nonalcoholic fatty liver disease in rats (Shirouchi et al., 2008). A recent report claimed that dietary PI influences immune functions in a way that prevents induced pathogenesis and development of the liver injury in mice (Inafuku et al., 2013). Administration of PI to human subjects improved cholesterol metabolism by stimulation of reverse cholesterol transport pathway and increased efflux of cholesterol from peripheral tissues to a high density lipoprotein (Burgess et al., 2005). Thus, the significance of supplementary PI is obvious. As a special concern, PI has been recently shown to have high level of safety regarding oral toxicity and genotoxicity in experiments on rats (Honda et al., 2009). However, PI is not readily available from natural sources. As an example, mature soybean known to contain high level of PLs, has only 287 mg PI/100 g of total weight (Weihrauch and Son, 1983). Furthermore, extraction of PI from natural sources, such as soy lecithin provides moderate yield and heterogeneous PI consisting of species with different fatty acid composition, which may not be suitable for all biotechnological applications. An efficient method of PI production is the enzymatic transphosphatidylation of soy phosphatidylcholine (PC), using phospholipase D (PLD) from non-pathogenic bacteria such as Streptomyces spp. or plants (De Maria et al., 2007; Uesugi and Hatanaka, 2009). Streptomyces PLDs are preferred in the industry as these are easily prepared, show broad substrate specificities and high transphosphatidylation rates (Ulbrich-Hofmann et al., 2005). However, conformation and size of myo-inositol Correspondence to: Y. Iwasaki Contract grant sponsor: Asahi Glass foundation and Japan Society for the Promotion of Science Received 30 July 2013; Revision received 17 October 2013; Accepted 4 November 2013 Accepted manuscript online 13 November 2013; Article first published online 30 November 2013 in Wiley Online Library (http://onlinelibrary.wiley.com/doi/10.1002/bit.25149/abstract). DOI 10.1002/bit.25149 ARTICLE 674 Biotechnology and Bioengineering, Vol. 111, No. 4, April, 2014 ß 2013 Wiley Periodicals, Inc.
Transcript of Deletion of a dynamic surface loop improves stability and changes kinetic behavior of...

Deletion of a Dynamic Surface Loop ImprovesStability and Changes Kinetic Behavior ofPhosphatidylinositol-SynthesizingStreptomyces Phospholipase D
Jasmina Damnjanovi�c, Hideo Nakano, Yugo Iwasaki
Laboratory of Molecular Biotechnology, Graduate School of Bioagricultural Sciences,
Nagoya University, Furo-cho, Chikusa-ku, Nagoya 464-8601, Japan;
telephone: þ81-52-789-4143; fax: þ81-52-789-4145; e-mail: [email protected]
ABSTRACT: Supplementary phosphatidylinositol (PI) wasshown to improve lipid metabolism in animals, thus it isinteresting for pharmaceutical and nutritional applications.Homogenous PI can be produced in transphosphatidylationof phosphatidylcholine (PC) with myo-inositol catalyzed byphospholipase D (PLD). Only bacterial enzymes able tocatalyze PI synthesis are Streptomyces antibioticus PLD (SaPLD)variants, among which DYR (W187D/Y191Y/Y385R) has thebest kinetic profile. Increase in PI yield is possible byproviding excess of solvated myo-inositol, which is achievableat high temperatures due to its highly temperature-dependent solubility. However, high-temperature PI synthe-sis requires the thermostable PLD. Previous site-directedcombinatorial mutagenesis at the residues of DYR havinghigh B-factor yielded the most improved variant, D40H/T291Y DYR, obtained by the combination of two selectedmutations. D40 and T291 are located within dynamic surfaceloops, D37-G45 (termed D40 loop) and G273-T313. Thus, inthis work, thermostabilization of DYR SaPLD was attemptedby rational design based on deletion of the D40 loop,generating two variants, D37-45 DYR and D38-46 DYR PLD.D38-46 DYR showed highest thermostability as its activityhalf-life at 70�C proved 11.7 and 8.0 times longer than that ofthe DYR and D37-45 DYR, respectively. Studies on moleculardynamics predicted D38-46 DYR to have the least averageRMSD change as temperature dramatically increases. At 60and 70�C, both mutants synthesized PI in a twofold higheryield compared to the DYR, while at the same time producedless of the hydrolytic side-product, phosphatidic acid.
Biotechnol. Bioeng. 2014;111: 674–682.
� 2013 Wiley Periodicals, Inc.
KEYWORDS: phospholipase D; flexible loops; stability
Introduction
Naturally occurring phosphatidylinositol (PI) is a phospho-lipid (PL) presentmainly in eukaryoticmembrane structures.PI and its metabolites play important roles in glycolipidanchoring of proteins, behave as secondary messengers insignal transduction and participate inmRNA export from thenucleus (Gardocki et al., 2005). Supplementary PI wasreported to decrease triacylglycerol levels in serum and liverof model animals, rats, and mice (Yanagita, 2003; Yanagitaand Nagao, 2008), and prevents nonalcoholic fatty liverdisease in rats (Shirouchi et al., 2008). A recent reportclaimed that dietary PI influences immune functions in a waythat prevents induced pathogenesis and development of theliver injury in mice (Inafuku et al., 2013). Administration ofPI to human subjects improved cholesterol metabolism bystimulation of reverse cholesterol transport pathway andincreased efflux of cholesterol from peripheral tissues to ahigh density lipoprotein (Burgess et al., 2005). Thus, thesignificance of supplementary PI is obvious. As a specialconcern, PI has been recently shown to have high level ofsafety regarding oral toxicity and genotoxicity in experimentson rats (Honda et al., 2009).
However, PI is not readily available from natural sources.As an example, mature soybean known to contain high levelof PLs, has only 287mg PI/100 g of total weight (Weihrauchand Son, 1983). Furthermore, extraction of PI from naturalsources, such as soy lecithin provides moderate yield andheterogeneous PI consisting of species with different fattyacid composition, which may not be suitable for allbiotechnological applications.
An efficient method of PI production is the enzymatictransphosphatidylationof soy phosphatidylcholine (PC), usingphospholipase D (PLD) from non-pathogenic bacteria such asStreptomyces spp. or plants (De Maria et al., 2007; Uesugi andHatanaka, 2009). StreptomycesPLDsarepreferred in the industryas these are easily prepared, show broad substrate specificitiesand high transphosphatidylation rates (Ulbrich-Hofmannet al., 2005). However, conformation and size of myo-inositol
Correspondence to: Y. Iwasaki
Contract grant sponsor: Asahi Glass foundation and Japan Society for the Promotion of
Science
Received 30 July 2013; Revision received 17 October 2013; Accepted 4 November 2013
Accepted manuscript online 13 November 2013;
Article first published online 30 November 2013 in Wiley Online Library
(http://onlinelibrary.wiley.com/doi/10.1002/bit.25149/abstract).
DOI 10.1002/bit.25149
ARTICLE
674 Biotechnology and Bioengineering, Vol. 111, No. 4, April, 2014 � 2013 Wiley Periodicals, Inc.

make it difficult to fit properly into the acceptor binding site ofmost of the PLDs. As a result, only two wild-type plant PLDs,PLD-B from poppy seedlings and spinach leaves have beenreported to catalyze PI synthesis (Mandal et al., 1980;Oblozinsky et al., 2005), but these enzymes are not easilyavailable for commercial applications. Another reason for poorreactivity of PLDs towards myo-inositol is the fact that PLDsprefer primary over secondary alcohols (Damnjanovi�c andIwasaki, 2013 and references therein).A new catalyst with PI-synthesizing ability was previously
created by site-directed saturation mutagenesis of the wild-type Streptomyces antibioticus PLD (SaPLD) at positionssuspected to be involved in substrate recognition and acco-mmodation, W187, Y191, and Y385. High-throughputscreening of �30,000 clones resulted in the isolation of �90PI-synthesizing PLD mutants (Masayama et al., 2008, 2009).Library screening showed that the conservation of Tyr atposition 191 and retention ofArg at position385 are crucial forselectivity in synthesis of 1(3)-PIoverotherPI isomers and thatthe position 187 is essential for affecting 1-PI/3-PI ratio. Asonly 1-PI is naturally abundant and shown to exhibit bio-activity, it was important to conserve 191Yand 385R in futurestudies on PI-synthesizing PLD. However, even with the best-performing variants, the total PI yield remained moderate.One way to improve PI synthesis is through the addition of
excess myo-inositol to the reaction mixture. Since thesolubility of this substrate is highly temperature-dependent,conducting the reaction at elevated temperatures couldincrease the concentration of soluble myo-inositol, thereforeimproving the PI yield. As this approach required thermosta-ble PLD, we previously identified the amino acid residuesaffecting thermostability in one of the PI-synthesizing SaPLDmutants, DYR (W187D/Y191Y/Y385R), chosen for itsfavorable kinetic properties (Damnjanovi�c et al., 2012). TheDYR’s thermostability has been improved by site-directedcombinatorial mutagenesis based on the residues’ B-factor.The most improved variant, D40H/T291Y DYR affordedtwofold increase in PI yield at 60�C. Residue D40 is part of aflexible, nine-residue surface loop of the N-terminal half-domain. The loop is solvent-exposed and highly fluctuatingwith D40 having the highest B-factor. Being distal from theactive site, it is believed that the loop does not have any rolein enzymatic function, however due to its flexibility, itsignificantly contributes to the sensitivity of the structuretowards heat-induced unfolding. A support to this hypothesishas been demonstrated when thermostabilty increasedwithout activity trade-off upon mutation of the loop’s mostflexible residue, D40 to His (Damnjanovi�c et al., 2012). Asreported by Thompson and Eisenberg (1999), such dynamicsurface loop structures are subject of the evolutionary deletiontowards the increase in thermostability. Thermophilicsequences were reported to often have deletions of exposedloop regions usually present in the mesophilic homologs.Based on the above, this study aimed to verify if the
removal of the flexible surface loop, D40 loop, can increasestability of SaPLD. Besides its dynamics, the loop causestwisting of the neighboring b-strand and makes two adjacent
residues to unusually orient on the same side of the b-strand.DYR model structure analysis indicates that the removal ofthe entire loop region (D37-G45 or A38-W46) can reduceflexibility and relieve the local structural tension.
Materials and Methods
Materials
Oligonucleotides were synthesized by Greiner Bio-One,Japan. Restriction enzymes and Pyrobest DNA polymerasewere purchased from Takara Bio., Japan. Dioleoylphospha-tidylcholine (DOPC), dioleoylphosphatidylserine (DOPS),and dioleoylphosphatidylglycerol (DOPG) were purchasedfrom NOF, Japan. Soy lecithin (SLP-PC 70) was provided byTsuji Oil Mill, Japan. 10-Acetyl-3,7-dihydroxyphenoxazine(Amplex Red reagent) was purchased from CaymanChemical, USA. Choline oxidase and peroxidase werepurchased from Toyobo, Japan. Organic solvents and otherchemicals were purchased from Wako, Japan.
Construction of the Deletion Mutants
Two dyr pld mutants having deletions of the nine amino acidresidues in the D40 loop region were constructed byoverlapping polymerase chain reaction (PCR; Fig. S1).New mutant genes were cut with SpeI and XhoI restrictionenzymes and ligated to a vector plasmid, pETKmS1-Term(Mishima et al., 1997) previously cut with SpeI and SalIrestriction enzymes, using commercial DNA ligation kit“Mighty Mix” (Takara Bio., Japan). The scheme of theplasmid construction is given in Supplementary Figure S2.The resultant ligation mixture was introduced into Escherichiacoli DH5a cells and positive clones selected on LB agar platessupplemented with 50mg/mL of chloramphenicol (Cm) and50mg/mL of kanamycin (Km). Overnight colonies weresuspended in LB liquid broth, cultivated for 16–18 h at 37�Cand used for plasmid preparation by conventional alkaline-sodium dodecyl sulfate method. Gene sequence wasconfirmed by DNA sequencing. Correct plasmid constructswere introduced into E. coli BL21 (DE3) cells used for PLDexpression. Oligonucleotide primers used for each PCR andfor DNA sequencing are listed in Supplementary Table SI.
Large-Scale Expression and Purification
BL21 (DE3) cells transformed with the pld-expressingplasmids containing the dyr, D37-45 dyr or D38-46 dyr pldgenes were used for large-scale PLD expression by auto-induction (Studier, 2005). Synthetic expression medium(Iwasaki et al., 1995) supplemented with 0.6% (v/v) glycerol,0.05% (w/v) glucose, 0.2% (w/v) lactose and 150mg/mL ofKm was inoculated with 1% of the overnight pre-cultureobtained by growing single colonies in LB/Km liquidmedium. Expression by auto-induction was carried out at30�C for 72 hwith shaking. Enzymes were recovered from theculture supernatants by (NH4)2SO4 (60%) precipitation. The
Damnjanovi�c et al.: Loop Deletion Improves Stability of PLD 675
Biotechnology and Bioengineering

crude enzyme solutions were first dialyzed against 50mMCH3COOH/CH3COONa buffer (pH 4.5) and centrifuged toremove the precipitated contaminant proteins and thendialyzed against 20mM Tris–HCl buffer (pH 8.0). The firststep of the purification was carried out using AKTA explorer(Amersham Pharmacia Biotech AB, Sweden) liquid chroma-tography system equipped with a Resource Q (6mL) anion-exchange column (GE Healthcare, UK). 0.5M NaCl in20mM Tris–HCl buffer (pH 8.0) was used as an eluent in alinear gradient against the same buffer. The second step of thepurification was carried out using the same equipmenthowever, with different column [cation-exchange POROS1
20 CM (PerSeptive Biosystems, USA)] and buffer system[0.5M NaCl in 20mM CH3COOH/CH3COONa (pH 5.0) ina linear gradient against the same buffer]. Protein concen-tration was measured using Bio-Rad protein assay kit (Bio-Rad) with bovine serum albumin as a standard.
PLD-Catalyzed PI Synthesis
PLD-catalyzed PI synthesis was performed using CaSO4-assisted aqueous suspension reaction system (Iwasaki et al.,2003)under thepreviouslydescribedconditions (Damnjanovi�cet al., 2012). Briefly, the reaction mixture composed of soylecithin SLP-PC 70 (50mg), myo-inositol (126mg at 40�C,163mg at 50�C, 205mg at 60�C, and 264mg at 70�C),CaSO4·2H2O (36mg) and purified PLD (15mg) was con-tained in 500mL of 50mM CH3COOH/CH3COONa buffer(pH5.6).After incubationat40, 50,60,or70�Cwithshaking for24h, the reaction was quenched with 2N HCl, PLs extractedwith chloroform/methanol (2/1) and analyzed by HPLC.
Kinetic Analysis
Kinetic analysis of PC hydrolysis was carried out in a bi-phasic system consisted of equal volumes of 50mMCH3COOH/CH3COONa buffer (pH 5.6) and ethyl acetate.Fifty microliter of ethyl acetate containing DOPC (0.50–10mM) was mixed with 50mL of 50mM CH3COOH/CH3COONa buffer (pH 5.6) phase containing 15mg of eitherDYR, D37-45 DYR or D38-46 DYR PLD. The mixtures wereincubated at 30�C for 1 h with shaking followed by reactionquenching with 50mL of 2N HCl and PL extraction with140mL of chloroform/methanol (2/1). Phosphatidic acid(PA) and PC contents were determined by HPLC. Based onthe PA yield, the reaction rate was calculated and subjected tonon-linear fitting to Hill equation using GraphPad Prism 5.Kinetic constants were calculated from the derived fit.
Specific Activity and Thermal Tolerance Assay
Specific activities of PLDs were determined using two-stepAmplex Red PLD Assay, as described previously(Damnjanovi�c et al., 2012). The first step of the assay isPLD-catalyzed hydrolysis of DOPC generating choline andPA. In the second step, the choline product is oxidized bycholine oxidase to yield betaine and H2O2. As H2O2 is being
generated, Amplex Red is converted to resorufin byhorseradish peroxidase in an H2O2-dependent oxidation.PLD (5mg/mL)-catalyzed reaction was conducted in 50mMCH3COOH/CH3COONa buffer (pH 5.6) containing 0.5mMDOPC and 0.25% (w/v) Triton X-100. After incubation at30�C for 20min, PLD was inactivated by heat (10min at95�C) and mixture cooled down. Cooled mixture was addedto the working solution containing 100mM Amplex Red,0.6 U/mL choline oxidase and 2U/mL horseradish peroxi-dase in 0.4M Tris–HCl buffer (pH 8.0). Fluorescence wasread at Ex/Em¼ 540/590 nm using Fusion fluorescence platereader (Packard BioScience, USA). Although based onindirect activity measurement, the assay is fast andconvenient for comparison between parent and the mutantenzymes in specific activities and thermal tolerance.
For the thermal tolerance assay, purified enzyme solutions(50mg/mL in 50mM CH3COOH/CH3COONa buffer pH5.6) were heat treated for 10, 20, 30, 40, 50, and 60min at 65and 70�C, cooled down and assayed for the remaininghydrolytic activity. Logarithmic values of the residualactivities were plotted against length of the heat treatmentand fitted into linear regression equation used to calculateactivity half-lives (t1/2) of the PLDs at 65 and 70�C.
MD Simulations
Models of the DYR and DYRD40 loop deletionmutants wereconstructed using YASARA Structure software. DYR model wasprepared using X-ray structure of DYR. As the side chains of11 residues (D40, R74, T162, N185, D187, Q250, S323, V332,L416, S424, and L485) in the structure of DYR contained twodifferent rotamers with occupancy values of 0.5, one of therotamers in each of those residues, except for D187, waschosen as adequate so that the structure becomes similar tothat of SaPLD. As for D187, one of the DYR mutation sites,one rotamer that can form a hydrogen bond with a nearbyresidue (D189) was chosen. As the internal residues L289-T291, T295-K296, and V380-S382 of the DYR X-raystructure were not resolved, the missing parts were graftedfrom 2ZE4. Model structures of the DYR D40 loop deletionmutants were created by deletion of the correspondingresidues (D37-G45 or A38-W46) in the DYRmodel structureand introduction of the peptide bond between residues L36and W46 or D37 and L47 followed by energy minimization(Wang et al., 2000). The MD simulations were performedusing Amber 03 force field parameters at pH 5.6, temperature40, 50, 60, and 70�C or 80, 100, 150, 200, and 250�C with thecorresponding solvent density (water) adjustment. Themultiple trajectories were analyzed and several parameterswere calculated. Final simulation employed temperature of250�C and simulation time of 20 ns.
Other Methods
PL quantification by HPLC was performed as describedpreviously (Damnjanovi�c et al., 2012, 2013). Proteinsequence alignment was done using ClustalW software(http://www.genome.jp/tools/clustalw).
676 Biotechnology and Bioengineering, Vol. 111, No. 4, April, 2014

Results
Structural Analysis of the D40 Loop Region and Design ofthe Deletion Mutants
The D40 loop region refers to the nine amino acid residues(D37-G45 or A38-W46) located within the N-terminal half-domain of SaPLD (Fig. 1A and B). This region is one of thepreviously identified thermostability “hot-spots,” with D40having B-factor of 44.3, located at the loop’s most exposedpart (Damnjanovi�c et al., 2012). Analysis of the three-dimensional SaPLD structure (2ZE4 and 2ZE9) showed thatthe loop is placed between two shortb-strands, N34-L36, andW46-Q49, seemingly interrupting the formation of one longstrand. This steric tension is further aggravated byuncommonly oriented W46 located at the same side of theb-strand as the neighboring L47. In SaPLD, N- and C-terminal half-domains show high degree of secondarystructure and conformational similarity and can be alignedwith low RMSD although the sequence similarity is onlyapprox. 26.3%. b-sheets of the two domains show remark-ably similar structure (Fig. 1B) except in the b-strand where
D40 loop is located. Further on, protein sequence alignmentof the 10 Streptomyces spp. PLDs showed that the D40 loopregion is present in all enzymes, however with a non-conserved sequence (Fig. 1C). Lack of the crystal structures ofmore than two Streptomyces spp. PLDs did not allow foraccurate conformational analysis of the region. Still, the sameloop conformation was observed in SaPLD and Streptomycessp. strain PMF PLD (1VOS; Fig. 1D). Based on the structuralanalysis of the loop region, two possible deletion patternswere designed (Fig. 2): (1) delete D37-G45 which wouldallow W46 to orient to the opposite direction and, (2) deleteA38-W46 which would eliminate W46.
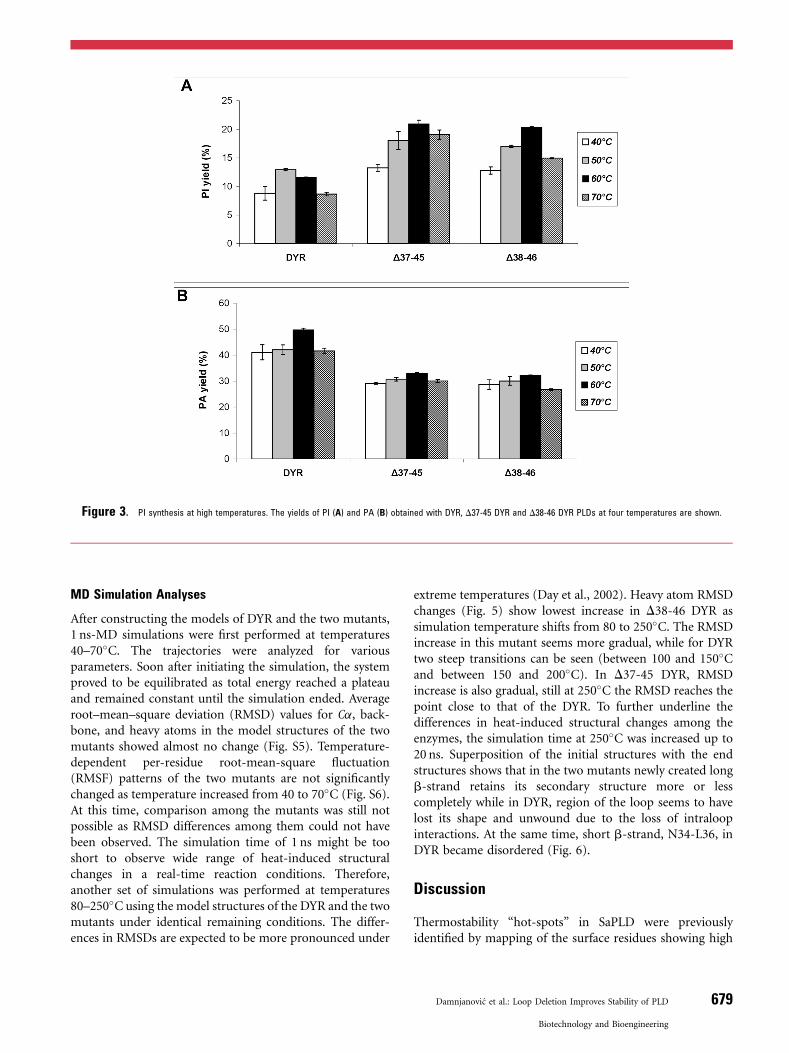
High-Temperature PI Synthesis
Purified DYR, D37-45DYR and D38-46DYR PLDs weretested in PI synthesis at increased temperatures (Fig. 3).Within the applied temperature interval (40–70�C) themutants showed better performance than the parent DYR,producing higher PI yields. Rise in yield of PI produced by themutants continued until 60�C, when the mutants achieved
Figure 1. Structure of Streptomyces PLD. (A) Overview of DYR SaPLD. The D40 loop is colored blue, (B) Structures of N- and C-terminal half-domains of SaPLD, showing
conformational similarity. b-strands are colored light blue with the D40 loop in blue. (C) Amino acid sequence alignment of 10 Streptomyces PLDs. The regions corresponding to the
D40 loop are highlighted in gray. Abbreviations refer to the strain names of the PLD producers; San: S. antibioticus (UniProt: Q53728), Sra: S. racemochromogenes strain 10-3
(E0D7I2), K1: S. halstedii K1 (Q8VW86), P821: Streptomyces sp. P821 (Q6YI41), YU100: Streptomyces sp. YU100 (B0FZD9), K6: S. halstedii subsp. scabies K6 (Q8KIK9), Scin:
Streptoverticillium cinnamoneum (Q9Z4Q0), Ssom: S. somaliensis (Q68VC2), PMF: Streptomyces sp. PMF (P84147), TH2: S. septatus TH2 (Q93HV1). Asterisks indicate identical
residues among all PLDs. Colons and periods indicate conservation between groups of strongly and weakly similar properties, respectively. The residue numbers on the top row are
of SaPLD. (D) Superposition of the D40 loop in SaPLD (magenta; PDB code 2ZE4) and Streptomyces sp. strain PMF PLD (gray; PDB code 1VOS).
Damnjanovi�c et al.: Loop Deletion Improves Stability of PLD 677
Biotechnology and Bioengineering

twofold higher PI yield than the DYR. Surprisingly, yield ofthe side-product, PA, obtained in the reaction with D37-45DYR and D38-46 DYR PLDs at 60�C was only 66.3 and64.5%, respectively, of that of the DYR (Fig. 3). At 70�C,mutants still retained >70% of the PI yield shown at 60�C.
Thermal Tolerance Assay
Residual activities after the heat treatment at 65 and 70�C forthe time interval of 10–60min were determined by theAmplex Red Phospholipase D Assay (Fig. 4). After the heattreatment at 65 and 70�C, D38-46 DYR PLD showed thehighest residual activity (Fig. 4A and C). After 10 and 60min-treatment at 65�C D38-46 DYR had 1.3 and 1.7 times,respectively, higher residual activity thanDYR and 1.2 and 1.4times, respectively, higher residual activity thanD37-45 DYR.After the treatment at 70�C, the difference between D38-46DYR and the other two enzymes was even more pronounced.After 10 and 60min-treatment at 70�C,D38-46 DYR retained3.1 and 4.1 times, respectively, higher residual activity thanDYR with similar differences observed between D38-46 DYRand D37-45 DYR. As shown in Figure 4B and D and Table I,the two mutants were measured to have enhanced thermaltolerance as evaluated by the activity half-life (t1/2) at both, 65and 70�C. Difference in thermal tolerance between D38-46DYR and DYR was more pronounced at 70�C, when D38-46DYR had 11.7 times longer t1/2, while t1/2 of D37-45 DYR was
only 1.5 times longer than that of the DYR. At 65�C, D37-45DYR had 1.3 times longer t1/2 than the DYR, while t1/2 ofD38-46 DYR proved nearly 2.0 times longer.
Kinetic Analysis
Although DYR and the mutants showed similar specificactivities (Table I), in PI synthesis variant enzymes producedsignificantly less PA compared to the DYR and previouslyobtained single, double, and triple DYR mutants(Damnjanovi�c et al., 2012). Therefore, the kinetic analysiswas carried out to quantitatively describe how kineticbehavior of the variant enzymes changed regarding PChydrolysis. Changes in the reaction rate depending on thesubstrate concentration in PC hydrolysis catalyzed by DYR,D37-45 DYR and D38-46 DYR are shown in Figure S4. Thedata points proved to best fit sigmoidal curve defined by theHill equation as it was also previously suggested for wild-type SaPLD and its mutants (Masayama et al., 2008). Suchkinetic behavior is generally observed with PLDs. Thesigmoidal character of the rate curve can be well observed ifonly first four data points are plotted (Fig. S4B). After non-linear fitting to Hill equation, kinetic constants, KM and Vmax,were calculated (Table I). Vmax of D37-45 DYR and D38-46DYR was 55.9 and 47.3%, respectively, of that of the DYR. Inthe same time KM also decreased to 80.3% and 59.8% of thatof the DYR in D37-45 and in D38-46 DYR PLDs respectively.
Figure 2. Deletion of the D40 loop. (A) Removal of the residues 37–45 or 38–46 from the parent enzyme (DYR) connects the two interrupted b-strands, resulting in the variants
D37-45 DYR or D38-46 DYR, respectively. (B) Enlarged view of the panel (A) with the zoom on W46 and L47.
678 Biotechnology and Bioengineering, Vol. 111, No. 4, April, 2014
MD Simulation Analyses
After constructing the models of DYR and the two mutants,1 ns-MD simulations were first performed at temperatures40–70�C. The trajectories were analyzed for variousparameters. Soon after initiating the simulation, the systemproved to be equilibrated as total energy reached a plateauand remained constant until the simulation ended. Averageroot–mean–square deviation (RMSD) values for Ca, back-bone, and heavy atoms in the model structures of the twomutants showed almost no change (Fig. S5). Temperature-dependent per-residue root-mean-square fluctuation(RMSF) patterns of the two mutants are not significantlychanged as temperature increased from 40 to 70�C (Fig. S6).At this time, comparison among the mutants was still notpossible as RMSD differences among them could not havebeen observed. The simulation time of 1 ns might be tooshort to observe wide range of heat-induced structuralchanges in a real-time reaction conditions. Therefore,another set of simulations was performed at temperatures80–250�C using the model structures of the DYR and the twomutants under identical remaining conditions. The differ-ences in RMSDs are expected to be more pronounced under
extreme temperatures (Day et al., 2002). Heavy atom RMSDchanges (Fig. 5) show lowest increase in D38-46 DYR assimulation temperature shifts from 80 to 250�C. The RMSDincrease in this mutant seems more gradual, while for DYRtwo steep transitions can be seen (between 100 and 150�Cand between 150 and 200�C). In D37-45 DYR, RMSDincrease is also gradual, still at 250�C the RMSD reaches thepoint close to that of the DYR. To further underline thedifferences in heat-induced structural changes among theenzymes, the simulation time at 250�C was increased up to20 ns. Superposition of the initial structures with the endstructures shows that in the two mutants newly created longb-strand retains its secondary structure more or lesscompletely while in DYR, region of the loop seems to havelost its shape and unwound due to the loss of intraloopinteractions. At the same time, short b-strand, N34-L36, inDYR became disordered (Fig. 6).
Discussion
Thermostability “hot-spots” in SaPLD were previouslyidentified by mapping of the surface residues showing high
Figure 3. PI synthesis at high temperatures. The yields of PI (A) and PA (B) obtained with DYR, D37-45 DYR and D38-46 DYR PLDs at four temperatures are shown.
Damnjanovi�c et al.: Loop Deletion Improves Stability of PLD 679
Biotechnology and Bioengineering

flexibility as evaluated by the B-factor value (Damnjanovi�cet al., 2012). One of the seven selected residues was D40, partof the dynamic nine-amino acid loop placed between the twoshort b-strands of the N-terminal half-domain. The loop,termed D40 loop (D37-G45), is oriented towards the outerenvironment and highly exposed (Fig. 1A). The interloopinteractions, two hydrophobic and four hydrogen bonds,maintain its shape. By its position, the loop preventsformation of one long instead of two short b-strands,N34-L36, and W46-Q49, while causing the second strand totwist. As a result, W46 is uncommonly oriented at the sameside of the b-strand as the neighboring L47 (Fig. 2B).Furthermore, the similar loop is absent from the corre-sponding part of the C-terminal half-domain (Fig. 1B). Thusthe following questions have arisen. Is this loop necessary tomaintain the structure? If not, is it possible to remove theunstable loop and make the local structure more rigid? Howwould this affect the overall stability? Producing one
continuous b-strand instead of two short ones coulddramatically reduce the local structural strain. As modelingstudies have predicted, trimming of the loop residues D37-G45 or A38-W46 could produce one continuous long b-strand, untwisting of the W46-Q49 region, thus, removal orreorientation of W46.
As the mutants were expressed as secreted active enzymesin E. coli (Fig. S3A and B), it was understood that the loop isnot necessary for proper folding or structure maintenance inPLD enzymes.
In the high-temperature PI synthesis, the mutant enzymesbehaved similarly to the previously selected, most stablevariant of DYR SaPLD, D40H/T291Y (Damnjanovi�c et al.,2012), achieving maximum of twofold higher PI yield at 60�Ccompared to the parent DYR. Further improvement of the PIyield was attempted but without significant success. Oneprobable reason is the accumulation of choline confirmed toinactivate PLD (Juneja et al., 1992). Effective removal of
Figure 4. Heat-inactivation of DYR, D37-45 DYR and D38-46 DYR PLDs at 65 and 70�C. (A) Residual activity vs. length of the heat treatment at 65�C; (B) Semi-logarithmic plot ofresidual activity vs. length of the heat treatment at 65�C; (C) Residual activity vs. length of the heat treatment at 70�C; (D) Semi-logarithmic plot of residual activity vs. length of theheat treatment at 70�C; Data at (A) and (C) present average of four measurements with SD< 10%. Symbols: open square DYR, closed triangleD37-45 DYR, closed squareD38-46 DYR.
Linear fitting: (B) y¼�0.0093xþ 1.9087, R2¼ 0.93 (DYR, solid line), y¼�0.0076xþ 1.9219, R2¼ 0.93 (D37-45 DYR, dotted line), and y¼�0.0058xþ 1.9540, R2¼ 0.95 (D38-46 DYR,
dash-dotted line); (D) y¼�0.0220xþ 1.7357, R2¼ 0.90 (DYR, solid line), y¼�0.0250xþ 1.7596, R2¼ 0.91 (D37-45 DYR, dotted line), and y¼�0.0136xþ 1.9643, R2¼ 0.96 (D38-46 DYR,
dash-dotted line).
Table I. Characterization of the DYR, D37-45 DYR and D38-46 DYR PLDs.
Specific activitya in PChydrolysis, nmolmg�1min�1 t651=2
a, min t701=2a, min
KMb in PC
hydrolysis, mMVmax
b in PChydrolysis, mmolmg�1min�1
Vmax/KMb in
PC hydrolysis Hill coefficienta
DYR 22.53� 0.58 22.6 1.67 5.90� 1.82 2.45� 0.74 0.42 1.01� 0.22D37-45DYR 17.13� 1.23 29.3 2.43 4.74� 1.51 1.37� 0.36 0.29 1.16� 0.28D38-46DYR 16.18� 1.02 44.0 19.5 3.53� 0.68 1.16� 0.14 0.33 1.21� 0.17
aSpecific activities,t651=2and t701=2were determined using Amplex red PLD assay, based on quantification of released choline in a twostep enzymatic reaction.bKinetic constants were determined in the assay based on direct quantification of PA, the product of PC hydrolysis.
680 Biotechnology and Bioengineering, Vol. 111, No. 4, April, 2014

choline by the coupled choline oxidase and catalase reactionimproved the conversion of PC to PS for more than 20%. Weattempted applying the same coupled enzymatic system forcholine removal during synthesis of PI, however, the PI yielddid not improve. Further attempts to change the substratefromDOPC to DOPG or DOPS did not result in increased PIyield either. For this purpose, a novel strategy has to beadopted.Surprisingly the mutants synthesized PA in drastically
lower quantity than the parent DYR and its single, double,and triple mutants. This phenomenon could be attributed toacquired structural rigidity, which might have influenced theflexibility of the residues around the active site promotingtransphosphatidylation rather than the hydrolysis. Thechange in the reaction specificity as a consequence of theloop removal was quantified by kinetic analysis of PChydrolysis. Vmax of the deletion mutants proved to beapproximately half of that of the DYR, accompanied by amilder reduction of KM. The change in Vmax had adominating influence on the change of the kinetic behavior
of the two variant enzymes, as seen by the reduction of Vmax/KM ratio. The reason for the moderate improvement of thesubstrate affinity reflected by the KM and in the same timelower Vmax may be sought in the change of structural rigidity.While the substrate binding can be facilitated, furthercatalytic steps probably require certain flexibility.Thermal tolerance of the mutants measured after different
length of incubation at 65 and 70�C was highest for D38-46DYR PLD. Significantly prolonged t1/2 of this mutant at bothassayed temperatures defines D38-46 DYR as the more stablevariant, unlike the D37-45 DYR whose thermal tolerance at70�C was not very distinctive from DYR’s. The t1/2 of D38-46DYR PLD at 65�C is also 9.4min longer compared to the t1/2
Figure 5. Heavy atom RMSD patterns of DYR, D37-45 DYR and D38-46 DYR PLDs
at 80, 100, 150, 200, and 250�C. At each temperature, MD trajectory was
computationally analyzed and average RMSDs of the heavy atoms in the whole
structure were calculated for every time step. Reference structure for comparisonwas
the initial model structure.
Figure 6. Local structure around the D40 loop region after 20 ns simulation at
250�C. Initial structures (blue) are superimposed with the structures at the simulation
end (yellow). The D40 loop region is encircled.
Damnjanovi�c et al.: Loop Deletion Improves Stability of PLD 681
Biotechnology and Bioengineering

of the most thermostable DYR variant, D40H/T291Y,obtained in the previous study (Damnjanovi�c et al., 2012).
Analysis of intramolecular interactions executed by PICserver (Tina et al., 2007) revealed that the acquired stability ofD38-46 DYR can be largely attributed to the formation of tennew hydrogen bonds between the surface exposed residues(main chain-main chain: 3, main chain-side chain: 6 and sidechain-side chain: 1). The difference in RMSD at varyingtemperatures indicated the extent of the conformationchange under the target conditions compared to the referencestructure. The mutants were predicted to have insignificantlylow or no change in RMSDs of Ca carbons, backbone, andheavy atoms at 40–70�C. When simulations were performedat 80–250�C, the least change was predicted for D38-46 DYR.Even after extension of the simulation time until 20 ns at thehighest temperature, D38-46 DYR almost completelyconserved the secondary structure of the newly formed b-strand, while DYR suffered drastic loss of the original loopshape and part of the secondary structure in shorterb-strand.
In summary, applied rational design by flexible surfaceloop deletion proved as a useful strategy for stability increasewhile having no negative effects on folding or function of thenew enzymes. While both, D37-45 DYR and D38-46 DYRPLDs performed better than the DYR in high-temperature PIsynthesis providing similar PI yield, D38-46 DYR PLDacquired higher thermostability improvement as confirmedby thermal tolerance test and computational analyses. Theobserved remarkable differences between D37-45 DYR andD38-46 DYR PLDs indicated the importance of the correctchoice of the residues to delete.
This work was supported in part by Asahi Glass Foundation to Y.I.and by a Grant-in-Aid for scientific research C from Japan Society forthe Promotion of Science to Y.I. [25425830]. The financial support toJ.D. by Ministry of Education, Culture, Sports, Science andTechnology (MEXT) of Japan is deeply appreciated.
References
Burgess JW, Neville TA-M, Rouillard P, Harder Z, Beanlands DS, Sparks DL.2005. Phosphatidylinositol increases HDL-C levels in humans. J LipidRes 46:350–355.
Damnjanovi�c J, Iwasaki Y. 2013. Phospholipase D as a catalyst: Application inphospholipid synthesis, molecular structure and protein engineering. JBiosci Bioeng 116:271–280.
Damnjanovi�c J, Takahashi R, Suzuki A, Nakano H, Iwasaki Y. 2012.Improving thermostability of phosphatidylinositol-synthesizing Strepto-myces phospholipase D. Protein Eng Des Sel 25:415–424.
Damnjanovi�c J, Nakano H, Iwasaki Y. 2013. Simple and efficient profiling ofphospholipids in phospholipase D-modified soy lecithin by HPLC withcharged aerosol detection. J Am Oil Chem Soc 90:951–957.
Day R, Bennion BJ, Ham S, Daggett V. 2002. Increasing temperatureaccelerates protein unfolding without changing the pathway ofunfolding. J Mol Biol 322:189–203.
De Maria L, Vind J, Oxenbøll K, Svendsen A, Patkar S. 2007. Phospholipasesand their industrial applications. Appl Microbiol Biotechnol 74:290–300.
Gardocki ME, Jani N, Lopes JM. 2005. Phosphatidylinositol biosynthesis:Biochemistry and regulation. Biochim Biophys Acta 1735:89–100.
Honda K, Enoshima T, Oshikata T, Kamiya K, Hamamura M, Yamaguchi N,Nakamura K, Oguma Y, Fujiwara S, Takabe M, et al. 2009. Toxicitystudies of Asahi Kasei PI, purified phosphatidylinositol from soy lecithin.J Toxicol Sci 34:265–280.
Inafuku M, Nagao K, Inafuku A, Yanagita T, Taira N, Toda T, Oku H. 2013.Dietary phosphatidylinositol protects C57BL/6 mice from concanavalinA-induced liver injury by modulating immune cell functions. Mol NutrFood Res 57:1671–1679.
Iwasaki Y, Mishima N, Mizumoto K, Nakano H, Yamane T. 1995.Extracellular production of phospholipase D of Streptomyces antibioticususing recombinant Escherichia coli. J Ferment Bioeng 79:417–421.
Iwasaki Y, Mizumoto Y, Okada T, Yamamoto T, Tsutsumi K, Yamane T. 2003.An aqueous suspension system for phospholipase D-mediated synthesisof PS without toxic organic solvent. J Am Oil Chem Soc 80:653–657.
Juneja LR, Taniguchi E, Shimizu S, Yamane T. 1992. Increasingproductivity by removing choline in conversion of phosphatidylcho-line to phosphatidylserine by phospholipase D. J Ferment Bioeng73:357–361.
Mandal SB, Sen PC, Chakrabarti P. 1980. In vitro synthesis of phosphati-dylinositol and phosphatidylcholine by phospholipase D. Phytochemis-try 19:1661–1663.
Masayama A, Takahashi T, Tsukada K, Nishikawa S, Takahashi R, Adachi M,Koga K, Suzuki A, Yamane T, Nakano H, et al. 2008. Streptomycesphospholipase D mutants with altered substrate specificity capable ofphosphatidylinositol synthesis. ChemBioChem 9:974–981.
Masayama A, Tsukada K, Ikeda C, Nakano H, Iwasaki Y. 2009. Isolation ofphospholipase D mutants having phosphatidylinositol-synthesizingactivity with positional specificity on myo-inositol. ChemBioChem10:559–564.
Oblozinsky M, Ulbrich-Hofmann R, Bezakova L. 2005. Head groupspecificity of phospholipase D isoenzymes from poppy seedlings(Papaver somniferum L.) Biotechnol Lett 27:181–185.
Shirouchi B, Nagao K, Inoue N, Furuya K, Koga S, Matsumoto H, Yanagita T.2008. Dietary phosphatidylinositol prevents the development ofnonalcoholic fatty liver disease in Zucker (fa/fa) rats. J Agric FoodChem 56:2375–2379.
Studier FW. 2005. Protein production by auto-induction in high-densityshaking cultures. Protein Expr Purif 41:207–234.
Thompson MJ, Eisenberg D. 1999. Transproteomic evidence of a loop-deletion mechanism for enhancing protein thermostability. J Mol Biol290:595–604.
Tina KG, Bhadra R, Srinivasan N. 2007. PIC: Protein interactions calculator.Nucleic Acids Res 35(suppl 2):W473–W476.
Uesugi Y, Hatanaka T. 2009. Phospholipase D mechanism using StreptomycesPLD. Biochim Biophys Acta 1791:962–969.
Ulbrich-Hofmann R, Lerchner A, Oblozinsky M, Bezakova L. 2005.Phospholipase D and its application in biocatalysis. Biotechnol Lett27:535–544.
Wang J, Cieplak P, Kollman PA. 2000. How well does a restrainedelectrostatic potential (RESP) model perform in calculating conforma-tional energies of organic and biological molecules? J Comput Chem21:1049–1074.
Weihrauch J, Son Y-S. 1983. Phospholipid content of foods. J Am Oil ChemSoc 60:1971–1978.
Yanagita T. 2003. Nutritional functions of dietary phosphatidylinositol.Inform 14:64–66.
Yanagita T, Nagao K. 2008. Functional lipids and the prevention of themetabolic syndrome. Asia Pac J Clin Nutr 17(Suppl 1):189–191.
Supporting Information
Additional supporting information may be found in theonline version of this article.
682 Biotechnology and Bioengineering, Vol. 111, No. 4, April, 2014


















