Cytoplasmic and nuclear protein synthesis during in vitro differentiation of murine ICM and...
13
DEVELOPMENTAL BIOLOGY 74,351-363 (1980) Cytoplasmic and Nuclear Protein Synthesis during in Vitro Differentiation of Murine ICM and Embryonal Carcinoma Cells CHIN C.HOWE,RUDOLFGM~~R,ANDDAVORSOLTER The Wistar Institute of Anatomy and Biology, 36th Street at Spruce, Philadelphia, Pennsylvania 19104 Received December 13, 1978; accepted in revised form June 18, 1979 Protein syntheses during in vitro differentiation of inner cell masses (ICM) isolated from mouse blastocysts and of pluripotent embryonal carcinoma cells (ECC) were compared by two-dimen- sional electrophoretic analysis of [%]methionine-labeled cells. While most of the polypeptides found in ICM, ECC, and embryoid bodies (EB) derived from them were common to all four preparations, some distinct differences were noted. More polypeptides changed in intensity during the differentiation of ICM than during the differentiation of ECC. Analysis of ECC prior to differentiation revealed that only some of the polypeptides abundant in ICM were present, while at the same time, some of the polypeptides abundant in ICM-EB were being synthesized. These data indicate that ECC represent cells further advanced in development than the cells of ICM isolated from 4-day-old blastocysts. The EB derived from ECC also differ from those from ICM. Comparison of EB derived from ICM and ECC with cells of the parietal yolk sac line, PYS, indicates that all three synthesize two polypeptides abundant in EB. These two polypeptides can, therefore, be used as biochemical markers of parietal entoderm differentiation. Pluripotent ECC synthesize small amounts of characteristic EB proteins and the lo-nm filament protein (also found in PYS cells but not in EB). This indicates that small numbers of differentiated or differentiating cells are present in pluripotent ECC cultures. INTRODUCTION Spontaneous and experimentally induced teratocarcinomas are tumors composed of various haphazardly mixed, differentiated adult tissues and stem cells called em- bryonal carcinoma cells (ECC). ECC are remarkably similar to cells of the early em- bryo as evidenced by their ability, after injection into a mouse blastocyst, to partic- ipate along with the embryonic cells in the formation of a normal adult mouse (Brins- ter, 1974; Papaioannou et al., 1975; Mintz and Illmensee, 1975; Ilhnensee and Mintz, 1976; Dewey et al., 1977). This similarity between ECC and early embryos has been described at the ultrastructural (Damjanov et al., 1971a; Pierce et al., 1967), biochemi- cal (Damjanov et al., 1971b; Bernstine et al., 1973; Sherman et al., 1976; Strickland et al., 1976; Linney and Levinson, 1977), and cell surface antigenic (Artzt et al., 1973; Stern et al., 1975; Solter and Knowles, 1978) levels. Indeed, analysis of the patterns of morphologic changes during the develop- ment of embryoid bodies (EB) from both ECC and inner cell masses (ICM) isolated from blastocysts shows striking similarities which parallel the early postimplantation in uiuo development of mouse embryos. First an outer layer of entodermal cells is formed around an inner cell core, then the core develops into ectoderm and mesoderm (Martin et al., 1977; Wiley et al., 1978; Hogan and Tilly, 1978). It is known that changes in the pattern of protein synthesis occur during the preim- plantation development of mouse embryos (Epstein and Smith, 1974; Van Blerkom and Brockway, 1975; Levinson et al., 1978; Van Blerkom et al., 1976; Howe and Solter, 1979) and rabbit embryos (Van Blerkom and Manes, 1974; Van Blerkom and Mc- Gaughey, 1978; Tucker and Schultz, 1978). Thus, changes in protein synthesis must 351 0012-1606/80/020351-13$02.00/O Copyright 0 1980 by Academic Press, Inc All rights of reproduction in any form reserved.
Transcript of Cytoplasmic and nuclear protein synthesis during in vitro differentiation of murine ICM and...

DEVELOPMENTAL BIOLOGY 74,351-363 (1980)
Cytoplasmic and Nuclear Protein Synthesis during in Vitro Differentiation of Murine ICM and Embryonal Carcinoma Cells
CHIN C.HOWE,RUDOLFGM~~R,ANDDAVORSOLTER
The Wistar Institute of Anatomy and Biology, 36th Street at Spruce, Philadelphia, Pennsylvania 19104
Received December 13, 1978; accepted in revised form June 18, 1979
Protein syntheses during in vitro differentiation of inner cell masses (ICM) isolated from mouse blastocysts and of pluripotent embryonal carcinoma cells (ECC) were compared by two-dimen- sional electrophoretic analysis of [%]methionine-labeled cells. While most of the polypeptides found in ICM, ECC, and embryoid bodies (EB) derived from them were common to all four preparations, some distinct differences were noted. More polypeptides changed in intensity during the differentiation of ICM than during the differentiation of ECC. Analysis of ECC prior to differentiation revealed that only some of the polypeptides abundant in ICM were present, while at the same time, some of the polypeptides abundant in ICM-EB were being synthesized. These data indicate that ECC represent cells further advanced in development than the cells of ICM isolated from 4-day-old blastocysts. The EB derived from ECC also differ from those from ICM. Comparison of EB derived from ICM and ECC with cells of the parietal yolk sac line, PYS, indicates that all three synthesize two polypeptides abundant in EB. These two polypeptides can, therefore, be used as biochemical markers of parietal entoderm differentiation. Pluripotent ECC synthesize small amounts of characteristic EB proteins and the lo-nm filament protein (also found in PYS cells but not in EB). This indicates that small numbers of differentiated or differentiating cells are present in pluripotent ECC cultures.
INTRODUCTION Spontaneous and experimentally induced
teratocarcinomas are tumors composed of various haphazardly mixed, differentiated adult tissues and stem cells called em- bryonal carcinoma cells (ECC). ECC are remarkably similar to cells of the early em- bryo as evidenced by their ability, after injection into a mouse blastocyst, to partic- ipate along with the embryonic cells in the formation of a normal adult mouse (Brins- ter, 1974; Papaioannou et al., 1975; Mintz and Illmensee, 1975; Ilhnensee and Mintz, 1976; Dewey et al., 1977). This similarity between ECC and early embryos has been described at the ultrastructural (Damjanov et al., 1971a; Pierce et al., 1967), biochemi- cal (Damjanov et al., 1971b; Bernstine et al., 1973; Sherman et al., 1976; Strickland et al., 1976; Linney and Levinson, 1977), and cell surface antigenic (Artzt et al., 1973; Stern et al., 1975; Solter and Knowles, 1978)
levels. Indeed, analysis of the patterns of morphologic changes during the develop- ment of embryoid bodies (EB) from both ECC and inner cell masses (ICM) isolated from blastocysts shows striking similarities which parallel the early postimplantation in uiuo development of mouse embryos. First an outer layer of entodermal cells is formed around an inner cell core, then the core develops into ectoderm and mesoderm (Martin et al., 1977; Wiley et al., 1978; Hogan and Tilly, 1978).
It is known that changes in the pattern of protein synthesis occur during the preim- plantation development of mouse embryos (Epstein and Smith, 1974; Van Blerkom and Brockway, 1975; Levinson et al., 1978; Van Blerkom et al., 1976; Howe and Solter, 1979) and rabbit embryos (Van Blerkom and Manes, 1974; Van Blerkom and Mc- Gaughey, 1978; Tucker and Schultz, 1978). Thus, changes in protein synthesis must
351 0012-1606/80/020351-13$02.00/O Copyright 0 1980 by Academic Press, Inc All rights of reproduction in any form reserved.

352 DEVELOPMENTAL BIOLOGY VOLUME 74,1980
also occur as the ICM or ECC differentiates into an EB containing primitive entoderm and pluripotent ectoderm and mesoderm. Recent studies have employed both one- and two-dimensional gel electrophoresis to study similarities and differences between ICM, ECC, and their EB derivatives (Dewey et al., 1978; Martin et al., 1978).
This study reports on the changes in the pattern of protein synthesis that occur dur- ing differentiation of ICM in vitro and com- pares them with the changes that occur during differentiation of ECC. Our purpose was to ascertain how closely differentiation of teratocarcinoma cells reflects differentia- tion of normal embryos. We also compared pluripotent and nullipotent ECC lines to see whether common protein patterns exist for each of these ECC types and how each of them compares with normal embryonic cells from different developmental stages.
MATERIALS AND METHODS
Preparation and Culture of ICM
ICM were isolated by immunosurgery (Solter and Knowles, 1975) from 4-day-old blastocysts obtained from ICR random- bred mice. Individual ICM were grown in drops of Dulbecco’s modified Eagle’s me- dium (DME) supplemented with 15% fetal bovine serum (FBS) under paraffin oil in bacteriological petri dishes (Falcon, 1007) in a COz incubator at 37°C. Under these conditions, ICM cannot attach to the sub-
strate and, within 2 to 3 days, form EB with a clearly delineated outer layer of cells. Within 5 days most ICM-derived EB are composed of several cell layers with one or two cavities (Hogan and Tilly, 1978; Wiley et al., 1978; our unpublished observations).
Cell Cultures
All cells were cultured in DME supple- mented with 15% FBS. The mouse cell lines employed in this study are listed in Table 1.
Pluripotent ECC lines, PSA and NG2, were maintained in the undifferentiated state by subculturing the cells on a feeder layer as described (Martin and Evans, 1975; Martin et al., 1977). To obtain undifferen- tiated ECC for radiolabeling, the ECC were seeded without feeder cells in a tissue cul- ture dish (Martin et al., 1977). When ECC started to form tightly rounded clumps, they were carefully detached from the re- maining feeder cells by gently flushing with a stream of medium from a pipet. The clumps were then transferred to a conical tube and underlayed with fresh culture me- dium. The detached clumps were allowed to settle through the underlayed fresh me- dium for a few minutes to facilitate sepa- ration of undifferentiated cell clumps from the contaminating feeder cells which re- mained in the upper medium as the clumps settled to the bottom. The medium and feeder cells were removed by pipetting.
Cell line
TABLE 1
CELLLINESDERIVEDFROMVARIOUSTERATOCARCINOMAS Cell type Teratocarci- Source of Genotype
noma tumor Reference
PSA Pluripotent ECC NG2 Pluripotent ECC SCCI Nullipotent ECC
PCC4 F9 MH-15 FA-25 OTT6050f PYS
Pluripotent ECC Nullipotent ECC Nullipotent ECC Nullipotent ECC Fibroblasts Parietal entoder-
ma1 cells
OTT-5584 OTT-5584 402-A-11
OTT-6050 OTT-6050 MH-15 tumor FA-25 tumor OTT-6050 OTT-6050
3-day embryo 3-day embryo Spontaneous
testicular 6-day embryo 6-day embryo 7-day embryo ‘I-day embryo 6-day embryo 6-day embryo
129/sv 129/sv 129/sv
129/sv 129/sv BALB/cJ AKR/J 129/sv 129/sv
Martin et al. (1977) Dewey et al. (1977) Martin and Evans (1975)
Jakob et al. (1973) Bernstine et ccl. (1973) Solter and Knowles (1978) Solter and Knowles (1978) Solter and Knowles (1978) Lehman et al. (1974)

HOWE, GM~~R, AND SOLTER Proteins of Embryos and Teratocarcinoma Cells
To provide differentiated cultures (i.e., EB) for radiolabeling, the undifferentiated ECC clumps were cultured in suspension in bacteriological petri dishes (Martin et al., 1977). Within 2 days, an outer entodermal cell layer surrounding the inner cell core became clearly apparent (our observations; Martin and Evans, 1975; Martin et al., 1977). EB were cultured for 7 more days and the medium was changed daily.
[3”S]Methionine Labeling of ICM and Cell Cultures
Freshly isolated ICM and ICM after 5 days of culture were labeled at 37°C for 5 hr in methionine-free Eagle’s minimal es- sential medium (MEM) containing 10% di- alyzed FBS and 500 pCi/ml of [35S]methi- onine (New England Nuclear, specific activ- ity > 400 Ci/mmole). In the case of cell cultures, cells (one 6-cm petri dish per cell line) were washed three times with methi- onine-free MEM containing 10% dialyzed FBS. They were then labeled at 37°C for 5 hr in methionine-free MEM containing IO% dialyzed FBS and 100 &i/ml of [35S]me- thionine.
Separation of Proteins into Cytoplasmic and Nuclear Fractions
We found that nuclei of ICM and ECC were more fragile than those of other tissue culture cells, e.g., fibroblastic or epithelial cells. To avoid rupture of nuclei, we have modified the procedures of Berkowitz et al. (1969) to minimize the handling of nuclei. Briefly, radiolabeled cells were washed three times with protein-free Whitten’s me- dium (for embryos) or with Dulbecco’s phosphate-buffered saline (for cell lines) and resuspended in a buffer containing 0.01 M NaCl, 0.01 M Tris-HCl, pH 7.4,1% NP40, and 2 mM phenylmethylsulfonylfluoride (PMSF) as a protease inhibitor. The sus- pensions were kept on ice for 1 hr with occasional vortexing and centrifuged through a cushion of 0.32 M sucrose, 3 mM MgCL, and 0.01 M Tris-HCl, pH 7.5, at
2000 rpm for 20 min in an International centrifuge. The resulting nuclei were vir- tually free of unbroken cells and cytoplas- mic components as examined by phase-con- trast or electron microscopy. The superna- tant, which contained the cytoplasmic pro- teins, was lyophilized. The lyophilized cy- toplasmic proteins and the nuclear proteins (the pellet) were prepared for two-dimen- sional gel electrophoresis according to the procedures described by Peterson and McConkey (1976a). The resulting solutions were then centrifuged to remove any insol- uble proteins and immediately electropho- resed or stored at -70°C and used for elec- trophoresis as soon as possible. In no case were the proteins stored as lyophilized pow- ders since artifactual heterogeneity is known to result (O’Farrell, 1975).
Two-Dimensional Gel Electrophoresis of Proteins Proteins were separated by two-dimen-
sional polyacrylamide gel electrophoresis according to the method of O’Farrell(1975). The ampholine compositions used in the first-dimension isoelectrofocusing were 1.6% of pH 5-8, 0.1% of pH 2.5-4, and 0.1% of pH 4-6. This ampholine composition gave a pH range of 4.5-6.7. This narrow range was chosen to achieve uniform distri- bution and hence clear resolution of pro- teins. The pH gradient was determined as described by O’Farrell (1975).
RESULTS
Cytoplasmic Protein Synthesis during in Vitro Differentiation of ICM and ECC
The PSA cell line was chosen as a rep- resentative ECC line because these cells and isolated ICM were shown to differen- tiate in a morphologically similar manner (Martin et al., 1977; Wiley et al., 1978). In addition, PSA cells are pluripotent, since following injection into a blastocyst the cells derived from the PSA line participate in the formation of an adult mouse (Dewey et al., 1977).

354 DEVELOPMENTAL BIOLOGY VOLUME 74,198O
FTC. 1. Autoradiograms of two-dimensional gels of cytoplasmic proteins from (a) ICM, (b) EB derived from in vitro differentiation of ICM, (c) PSA cells, and (d) EB derived from in vitro differentiation of PSA cells. The approximate pH range is given at the top, and the approximate molecular weight is noted at the far left. The molecular weight scale was based on the positions of the following marker proteins: myosin heavy chain (200,600 daltons), phosphorylase A (94,000 daltons), bovine serum albumin (68,000 daltons), actin (45,000 daltons), and carbonic anhydrase (29,660 daltons). In this figure, actin (three forms) is indicated by the letter “A” and tubulins (55,000 and 53,000 daltons) by the letter “T”. The actin and tubulin were identified by electrophoresis of purified actin and tubulin from mouse cells. Approximately 300,000 TCA-precipitable cpm in less than 20 pg of proteins was applied to each first-dimension gel. The duration of X-ray film exposure was about 2 weeks. The proteins prepared in the presence of the protease inhibitor (PMSF) gave gel patterns indistinguishable from those of controls without PMSF.
When the two-dimensional gel profiles of in each. However, the amounts of several 35S-labeled proteins from ICM, ECC, and proteins synthesized changed during differ- their EB derivatives (Fig. 1) were com- entiation. In the case of ICM differentia- pared, it was apparent that most of the tion, six spots (1, 2, and 5-8 in Fig. la) several hundred detectable spots not only clearly decreased in intensity and two spots were common to all four preparations but (a and b in Fig. lb) clearly increased in were synthesized in roughly equal amounts intensity. Closer examination revealed

HOWE,GM~~R,ANDSOLTER Proteins of Embryos and Teratocarcinoma Cells 355
some additional spots undergoing minor changes in intensity, with two spots (3 and 4 in Fig. la) decreasing and five spots (c-g in Fig. lb) increasing. Throughout the rest of this paper, spots 1-8 will be referred to as ICM-abundant polypeptides, and spots a-g as EB-abundant polypeptides. In con- trast to the differentiation of ICM, fewer proteins changed in intensity during the differentiation of ECC. Only two spots (a and b in Fig. Id) increased significantly, while spots c and d (Fig. Id) increased and spots 1 and 5 (Fig. lc) decreased, but only marginally.
Undifferentiated ECC (Fig. lc) differed significantly from ICM (Fig. la). All eight ICM-abundant polypeptides (l-8) were synthesized in smaller amounts than in ICM. Of these, six (2, 3, 4, 6, 7, and 8) were equal in intensity to those in ICM-EB (or ECC-EB). Only two (1 and 5) were greater than both EB. Hence, only two (1 and 5) of the eight ICM-abundant polypeptides were found to be ECC abundant. In addition, ECC synthesized four polypeptides more abundantly than ICM: a new polypeptide (9, Fig. lc) barely detectable in ICM and three EB-abundant polypeptides (e, f, and g, Fig. lc). Protein 9 was identified as the major polypeptide -of 10-m filaments by electrophoresis of purified protein from mouse tibroblasts. Although ECC differed significantly from ICM, the differentiated derivatives of ECC were similar to the dif- ferentiated derivatives of ICM (Figs. lb and d). The same seven EB-abundant polypep- tides (a-g) and low levels of ICM-abundant polypeptides (1-8) were present in both EB.
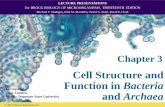
To determine whether the differences we observed between PSA-ECC and ICM were general to all ECC, protein patterns of the ECC lines listed in Table 1 were compared to those of ICM and ICM-EB. ECC lines isolated from tumors of widely different origins (see Figs. 2a-f), including nullipo- tent cells (Figs. 2c-f), were similar to PSA cells by the following criteria: (1) The same two ICM-abundant polypeptides (1 and 5) were synthesized more abundantly in every
ECC tested than in the ICM-EB or PSA- EB; (2) the same six ICM-abundant poly- peptides (2,3,4,6, 7, and 8) were present in small amounts in each ECC line; and (3) the same three EB-abundant polypeptides (e, f, and g) were found in all of the ECC lines. However, pluripotent ECC differed slightly from nullipotent ECC. Pluripotent ECC contained more of the EB-abundant polypeptides a and b than n&potent ECC. In addition, protein 9, or the major protein of lo-nm filaments, was detectable in plu- ripotent ECC but not significantly in nulli- potent ECC. In addition to the differences between pluripotent and nullipotent ECC, differences among various pluripotent ECC lines were observed.
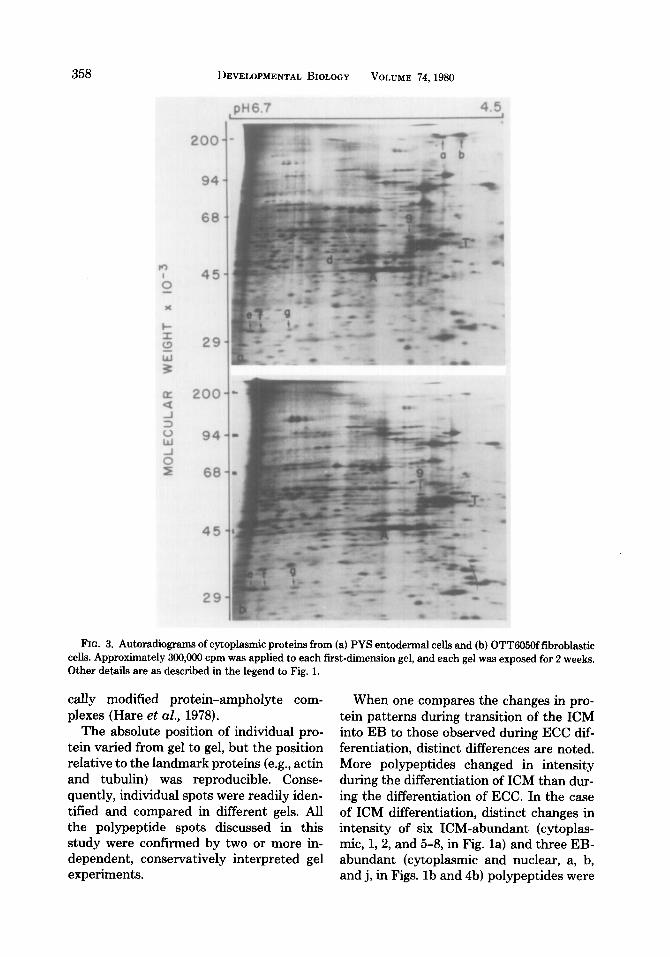
The entodermal cell is one of the major differentiated cell types associated with EB development. It was, therefore, of interest to determine whether the PYS entodermal cell line expresses ICM-abundant and EB- abundant polypeptides. As a control, a fi- broblastic cell line (OTT605Of) was also analyzed. PYS cells (Fig. 3a) synthesized very low levels of the ICM-abundant poly- peptides (l-8)) but did synthesize six of the seven EB-abundant polypeptides (a, b, and d-g), as well as lo-nm filament protein (9). Like PYS cells, the OTT6050f fibroblasts (Fig. 3b) synthesized lo-nm filament pro- tein (9) and very low levels of ICM-abun- dant proteins (l-8). However, unlike PYS cells, OTT6050f fibroblasts synthesized very little of EB-abundant proteins (a-d).
Nuclear Protein Synthesis during in Vitro Differentiation of ECC and ICM
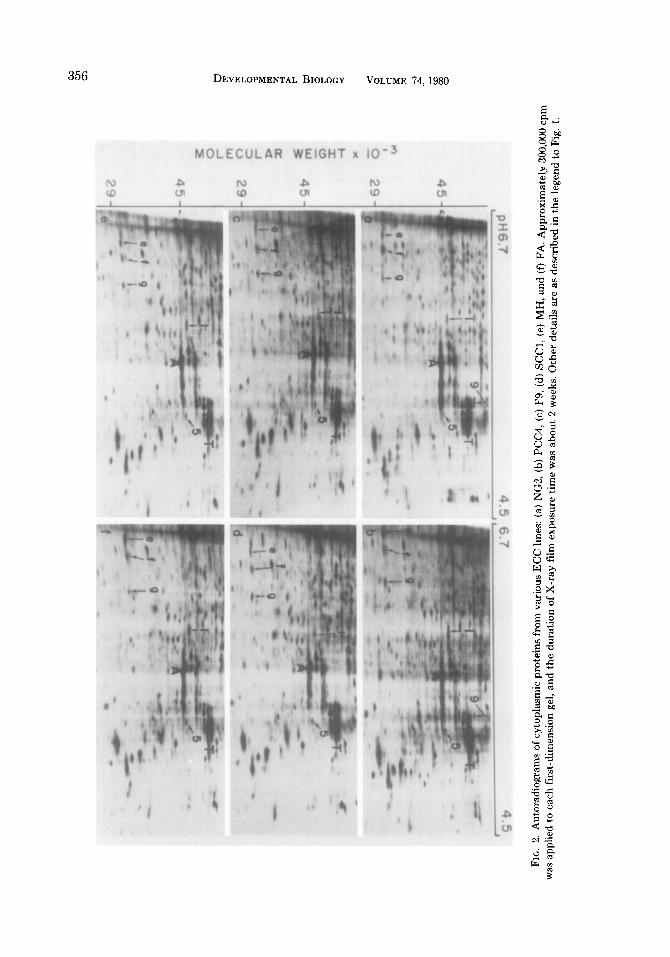
Autoradiographic patterns of nuclear proteins from ICM, PSA-ECC, and the EB derivatives of each are presented in Fig. 4. More than one hundred spots were de- tected. Many of them were clearly more abundant in the nuclear fractions than in the cytoplasmic fractions and are referred to as nuclear-abundant polypeptides (indi- cated by unnumbered arrows, Figs. 4a-d). The most striking changes occurring during the differentiation of ICM or ECC are the
356 DEVELOPMENTAL BIOLOGY VOLUME PI,1980

HOWE, GM~~R, AND SOLTER Proteins ofEmbryos and Teratocarcinoma Cells 357
increases in the intensity of three spots (a, b, and j, Figs. 4b and d) in the EB deriva- tives (hereafter referred to as EB abun- dant). Less noticeable during differentia- tion of ICM are the minor increases in two spots (h and i, Fig. 4b) and the decrease in six spots (5, 6, and 10-13, Fig. 4a, referred to as ICM abundant).
PSA (Fig. 4c) or any of the other ECC tested differed from JCM in that they con- tained low levels of the ICM-abundant nu- clear polypeptides (5, 6, and 10-13). On the other hand, nuclear preparations from PSA-ECC contained more protein 9, the lo-nm filament protein, than that from ICM. In general more lo-nm filament pro- tein was found in the nuclear fraction of pluripotent ECC than in that of nullipotent ECC (results not shown). The EB derived from PSA cells (Fig. 4d) also differed from those from ICM. The EB-abundant nuclear polypeptides h and i found in the ICM-EB were not synthesized in large quantities in the PSA-EB nuclear preparation. However, both ICM- and PSA-EB synthesized large amounts of the EB-abundant nuclear poly- peptides a, b, and j. The EB-abundant nu- clear polypeptides a and b were identical to those detected in the corresponding cyto- plasmic fractions of EB. Consistent with the observation from the cytoplasmic pro- teins, these two polypeptides were again present more abundantly in the nuclear fraction of PYS cells (Fig. 5a) than in that of OTT6050f cells (Fig. 5b). On the other hand, the EB-abundant nuclear protein j was more abundant in the nuclear fraction of OTT6050f than in that of PYS. It is of interest to note that the lo-nm filament protein is found in the nuclear fractions of both PYS and OTT6050f cells.
DISCUSSION
In this study we sought to compare the polypeptides synthesized by ICM and ECC during differentiation into EB. This step, in u&o, mimics the first detectable differen- tiation events in the embryo proper in uiuo,
namely, the formation of distinct layers of cells both of which are determined (Diwan and Stevens, 1976). Study of changes in polypeptide patterns and identification of proteins synthesized during this crucial pe- riod might provide clues as to how differ- entiation occurs. To facilitate the analysis of the large number of proteins present in whole cells, cells were separated into cyto- plasmic and nuclear fractions. NP40 was used in the fractionation to minimize con- tamination of the nuclear fraction by cyto- plasmic proteins (Berkowitz et al., 1969; Howe et al., 1975). Phase-contrast and elec- tron microscopic examination of the nu- clear preparations confirmed that contam- ination was minimal. Treatment of cells with NP40 in the absence of sucrose is known to make the nucleus leak certain freely diffusable nuclear proteins (Keller- mayer et al., 1976; Loeb et al., 1976). Thus, with our fractionation procedures, the cy- toplasmic fraction consists of cytoplasmic and nucleoplasmic proteins, whereas the nuclear fraction consists of nuclear proteins that are not freely diffusable or are tightly bound to the chromosomes as well as NP40- insoluble extranuclear proteins. We have observed many proteins to be more abun- dant in cytoplasmic fractions, and others in the nuclear fractions. The distinctly differ- ent electrophoretic patterns of the two frac- tions along with the microscopic evidence make it very clear that cytoplasmic con- tamination of nuclei was not significant. Our observation that nuclear and cytoplas- mic fractions share many of the same pro- teins is in agreement with reported results (Peterson and McConkey, 1976a,b: Howe and Solter, 1979).
Some spots showing charge heterogene- ity were observed in all of the gels exam- ined. Since all precautions as recommended by O’Farrell (1975) were taken to avoid modification of proteins, these proteins probably represent in viuo modified glyco- proteins, phosphorylated proteins, acetyla- ted proteins (O’Farrell, 1975), and chemi-
358 DEVELOPMENTAL BIOLOGY VOLUME 74,198O
FIG. 3. Autoradiograms of cytoplasmic proteins from (a) PYS entodermal cells and (b) OTT605Offibroblastic cells. Approximately 300,000 cpm was applied to each first-dimension gel, and each gel was exposed for 2 weeks. Other details are as described in the legend to Fig. 1.
calIy modified protein-ampholyte com- plexes (Hare et al., 1978).
The absolute position of individual pro- tein varied from gel to gel, but the position relative to the landmark proteins (e.g., actin and tubulin) was reproducible. Conse- quently, individual spots were readily iden- tified and compared in different gels. All the polypeptide spots discussed in this study were confirmed by two or more in- dependent, conservatively interpreted gel experiments.
When one compares the changes in pro- tein patterns during transition of the ICM into EB to those observed during ECC dif- ferentiation, distinct differences are noted. More polypeptides changed in intensity during the differentiation of ICM than dur- ing the differentiation of ECC. In the case of ICM differentiation, distinct changes in intensity of six ICM-abundant (cytoplas- mic, 1,2, and 5-8, in Fig. la) and three EB- abundant (cytoplasmic and nuclear, a, b, and j, in Figs. lb and 4b) polypeptides were

HOWE, GM~R, AND SOLTER Proteins of Embryos and Teratocarcinoma Cells 359
detected, with some additional spots show- ing minor changes in intensity. However, in the case of ECC differentiation, only three EB-abundant polypeptides (cytoplasmic and nuclear, a, b, and j, in Figs. Id and 4d) showed distinct changes in intensity. Our finding that the protein pattern of ECC differs from those of ECC-EB is consistent with a study using an OTT6050 retrans- plantable teratocarcinoma maintained by serial intraperitoneal transplant in mice (Dewey et al., 1978).
It is also of interest to examine the
changes in protein patterns during ICM differentiation in light of earlier results. The ICM-abundant polypeptides (1,2, and 5-8, in Fig. la) which are present in large amounts in ICM and in decreasing quanti- ties as ICM differentiate were found to appear in large quantities in embryos before the 4- to B-cell stage (Howe and Solter, 1979). Although we observed many poly- peptides of which the synthesis increased during ICM differentiation, we did not de- tect the new 55,000-dalton basic protein observed by Martin et al. (1978), presum-
FIG. 4. Autoradiographs of nuclear proteins from (a) ICM, (b) EB derived from ICM, (c) PSA cells, and (d) EB derived from PSA cells. Approximately 300,OOO cpm was applied to each first-dimension gel, and each gel was exposed for 2 weeks. Other details are as described in the legend to Fig. 1.

360 DEVELOPMENTAL BIOLOGY VOLUME 74, 1980
FIG. 5. Autoradiographs of nuclear proteins from (a) PYS entodermal cells and (b) OTT6050f fibroblastic cells. Approximately 300,000 cpm was applied to each first-dimension gel, and each gel was exposed for 2 weeks. Other details are as described in the legend to Fig. 1.
ably because it falls outside the pH range used in our study.
Comparison of the PSA protein pattern of the present study with the patterns of preimplantation embryos at various stages of development (Levinson et al., 1978; Howe and Solter, 1979) shows that PSA- ECC most closely resemble the ICM. How- ever, the presence in PSA cells of three EB- abundant cytoplasmic polypeptides (e, f, and g) and of a reduced number of ICM- abundant cytoplasmic or nuclear polypep- tides (Figs. la, c, and 4a, c) indicate that
PSA-ECC represent cells at a later stage of development than ICM. This finding con- firms the recent report of Martin et al. (1978), who studied these systems using one-dimensional gel electrophoresis, how- ever, the use of two-dimensional electro- phoresis has allowed us to detect many more differences between ICM and ECC. It should also be pointed out that the differ- ences between ICM and ECC described in the present study are not limited to cell lines cultured in vitro. Dewey et al. (1978) have found that ECC isolated from ascites

HOWE, GM~~R, AND SOLTER Proteins of Embryos and Teratocarcinoma Cells 361
also differ from ICM. Of the EB-abundant polypeptides we
have been able to identify two (a and b) in both the cytoplasmic and the nuclear frac- tions of ICM- and PSA-EB (Figs. lb, d, and 4b, d). These two polypeptides are parietal entoderm specific: They are clearly present in PYS entodermal cells but barely detect- able in OTT6050f fibroblastic cells (Figs. 3 and 5), and have been identified as base- ment membrane proteins, a parietal ento- derm product (manuscript in preparation). These two basement membrane polypep- tides are found in the nuclear fraction be- cause of their insolubility in NP40. Thus, these two polypeptides should be good markers for parietal entoderm differentia- tion. Our observation of the appearance of basement membrane proteins in EB by electrophoresis parallels similar observa- tions in morphological studies (Martin et al., 1977; Wiley et al., 1978; Hogan and Tilly, 1978).
Small amounts of these two parietal en- toderm-specific polypeptides, a and b, were consistently detected in pluripotent cells such as PSA (Figs. lc and 4c), NG2, and PCC4 cells, but only in trace amounts in nullipotent cells. These results support our morphological observations; small numbers of differentiated entodermal cells are pres- ent in these nominally undifferentiated cul- tures of pluripotent cells. Our results are also consistent with the observation by Sherman and Miller (1978) of a very small number of differentiated entodermal cells in the culture of nullipotent ECC (F9). The lo-nm filament protein (protein 9) is also synthesized in trace amounts by nullipotent ECC but in larger amounts by pluripotent PSA, NG2, and PCC4 and still larger amounts by PYS and OTT6050f cells. Again, the presence in pluripotent ECC cultures of small numbers of entodermal cells would explain the presence of IO-nm filament protein in these cultures. Surpris- ingly, the EB that contain a layer of ento- dermal cells do not synthesize lo-nm fila-
ment protein. One of the many biological functions of lo-nm filaments is the spread- ing and attachment of cells to substratum (Goldman and Follet, 1970). Since cells must detach from the substratum and form aggregates in suspension before ECC can differentiate into EB, it is possible that the entodermal cells do not synthesize lo-nm filament protein because they are not at- tached to the substratum. In addition to the differences between pluripotent and nullipotent ECC, we have consistently ob- served differences even among various plu- ripotent ECC lines as previously noted by Dewey et al. (1978).
This study presents a preliminary at- tempt to identify the biochemical events associated with differentiation of pluripo- tent cells. These events are associated with the narrowing of developmental potentials open to the embryonic stem cells. We have been able to identify some of the products which are synthesized by the differentiated cells. It would also be of interest to identify the proteins which disappear, i.e., short- term proteins which appear in embryos be- fore the 4- to B-cell stage and subsequently disappear. Perhaps the two cytoplasmic polypeptides, 1 and 5 (the only ICM-abun- dant proteins shared by the ECC), are in this respect most fascinating.
This work was supported by USPHS Research Grants CA 10815, CA 17546, and CA 21069 from the National Cancer Institute, HD 12487 from NICHHD, PCM 78-16177 from the National Science Foundation, and l-695 from the National Foundation-March of Dimes. H.G. was supported by a Fellowship from the Swiss National Science Foundation.
We thank David Lugg for his excellent assistance in some of these experiments.
REFERENCES
ARTZT, K., DLJBOIS, P., BENNETT, D., CONDAMINE, H., BABINET, C., and JACOB, F. (1973). Surface antigens common to mouse cleavage embryos and primitive teratocarcinoma cells in culture. Proc. Nat. Acad. Sci. USA 70,2988-2992.
BERKOWITZ, D. M., KAKEFUDA, T., and SPORN, M. B. (1969). Simple and rapid method for the isolation of enzymatically active HeLa cell nuclei. .J. Cell Biol. 42,851-855.

362 DEVELOPMENTAL BIOLOGY VOLUME 74,198O
BERNSTINE, E. G., HOOPER, M. L., GRANDCHAMP, S., and EPHRUSSI, B. (1973). Alkaline phosphatase ac- tivity in mouse teratoma. Proc. Nat. Acad. Sci. USA 70,3899-3903.
BRINSTER, R. L. (1974). The effect of cells transferred into the mouse blastocyst on subsequent develop- ment. J. Exp. Med. 140, 1049-1056.
DAMJANOV, I., SOLTER, D., BELICZA, M., and SKREB, N. (1971a). Teratomas obtained through extrauter- me growth of seven-day mouse embryos. J. Nat. Cancer Inst. 46,471-480.
DAMJANOV, I., SOLTER, D., and SKREB, N. (1971b). Enzyme histochemistry of experimental embryo-de- rived teratocarcinomas. Z. Krebsforsch. 76,249-256.
DEWEY, M. J., MARTIN, D. W., JR., MARTIN, G. R., and MINTZ, B. (1977). Mosaic mice with teratocar- cinema-derived mutant cells deficient in hypoxan- thine phosphoribosyltransferase. Proc. Nat. Acad. Sci. USA 74,5564-5568.
DEWEY, M. J., FILLER, R., and MINTZ, B. (1978). Protein patterns of developmentally totipotent mouse teratocarcinoma cells and normal early em- bryo cells. Develop. Biol. 66, 171-182.
DIWAN, S. B., and STEVENS, L. C. (1976). Develop- ment of teratomas from the ectoderm of mouse egg cylinders. J. Nat. Cancer Inst. 57,937-942.
EPSTEIN, C. J., and SMITH, S. A. (1974). Electropho- retie analysis of proteins synthesized by preimplan- tation mouse embryos. Develop. Biol. 40, 233-244.
GOLDMAN, R., and FOLLET, E. (1970). Birefringent filamentous organelle in BHK-21 cella and ita pos- sible role in cell spreading and motility. Science 169, 286-288.
HARE, D. L., STIMPSON, D. I., and CANN, J. R. (1978). Multiple bands produced by interaction of a single macromolecule with carrier ampholytes during iso- electric focusing. Arch. Biochem. Biophys. 187,274- 275.
HOGAN, B., and TILLY, R. (1978). In vitro development of inner cell masses isolated immunosurgicslly from mouse blastocysta. I. Inner cell masses from 3.5-day p.c. blastocysta incubated for 24 hr before immuno- surgery. J. Embryol. Exp. Morphol. 45,93-105.
HOWE, C. C., and SOLTER, D. (1979). Cytoplasmic and nuclear protein synthesis in preimplantation mouse embryos. J. Embryol. Exp. Morphol. 52, 209-225.
HOWE, C., TAN, K. B., and SOKOL, F. (1975). Fate of parental simian virus 40 DNA in permissive monkey kidney cells. J. Gen. Virol. 27, 11-24.
ILLMENSEE, K., and MINTZ, B. (1976). Totipotency and normal differentiation of single teratocarcinoma cells cloned by injection into blastocysts. Proc. Nat. Acad. Sci. USA 73,549-553.
JAKOB, H., BOON, T., GAILLARD, J., NICOLAS, J. F., and JACOB, F. (1973). Teratocarcinome de la souris: Isolement, culture et proprietes de cellules a poten- tialites multiples. Ann. Microbial. (Inst. Pasteur) 124B, 269-282.
KELLERMAYER, M., TALAS, M., WONG, C., BUSCH, H., and BUTEL, J. S. (1976). The solubility characteris- tics of SV40 tumor antigen. Exp. Cell Res. 99,456- 460.
LEHMAN, J. M., SPEERS, W. C., SWARTZENDRUBER, D. E., and PIERCE, G. B. (1974). Neoplastic differ- entiation: Characteristics of cell lines derived from a murine teratocarcinoma. J. Cell. Physiol. 84, 13- 28.
LEVINSON, J., GOODFELLOW, P., VADEBONCOEUR, M., and MCDEVITT, H. (1978). Identification of stage- specific polypeptides synthesized during murine preimplantation development. Proc. Nat. Acad. Sci. USA 75,3332-3336.
LINNEY, E., and LEVINSON, B. (1977). Teratocarci- noma differentiation: Plasminogen activator activity associated with embryoid body formation. Cell 10, 297-304.
LOEB, J. E., RITZ, E., CREUZET, C., and JAMI, J. (1976). Comparison of chromosomal proteins of mouse primitive teratocarcinoma, liver and L cells. Exp. Cell Res. 103,450-454.
MARTIN, G. R., and EVANS, M. J. (1975). Differentia- tion of clonal lines of teratocarcinoma cells: For- mation of embryoid bodies in vitro. Proc. Nat. Acad. Sci. USA 72,1441-1445.
MARTIN, G. R., WILEY, L. M., and DAMJANOV, I. (1977). The development of cystic embryoid bodies in vitro from clonal teratocarcinoma stem cells. De- velop. Biol. 61, 230-244.
MARTIN, G. R., SMITH, S., and EPSTEIN, C. J. (1978). Protein synthetic patterns in teratocarcinoma stem cells and mouse embryos at early stages of devel- opment. Develop. Biol. 66, 8-16.
MINTZ, B., and ILLMENSEE, K. (1975). Normal genet- ically mosaic mice produced from malignant tera- tocarcinoma cells. Proc. Nat. Acad. Sci. USA 72, 3585-3589. .
C’FARRELL, P. H. (1975). High resolution two-dimen- sional electrophoresis of proteins. J. Biol. Chem. 250,4007-4021.
PAPAIOANNOU, V. E., MCBURNEY, M. W., GARDNER, R. L., and EVANS, M. J. (1975). Fate of teratocarci- noma cells injected into early mouse embryos. Na- ture (London) 258,70-73.
PETERSON, J. L., and MCCONKEY, E. H. (1976a). Non- hi&one chromosomal proteins from HeLa cells: A survey by high resolution, two-dimensional electro- phoresis. J. Biol. Chem. 251, 548-554.
PETERSON, J. L., and MCCONKEY, E. H. (1976b). Pro- teins of Friend leukemia cells Comparison of he- moglobin-synthesizing and noninduced populations. J. Biol. Chem. 251,555-558.
PIERCE, G. B., STEVENS, L. C., and NAKANE, P. K. (1967). Ultrastructural analysis of the early devel- opment of teratocarcinomas. J. Nat. Cancer Inst. 39.755-773.
SHERMAN, M. I., and MILLER, R. A. (1978). F9 em-

HOWE, GM~~R, AND SOLTER Proteins of Embryos and Teratocarcinoma Cells 363
bryonal carcinoma cells can differentiate into en- doderm-like cells. Develop. Biol. 63, 27-37.
SHERMAN, M. I., STRICKLAND, S., and REICH, E. (1976). Differentiation of early mouse embryonic and teratocarcinoma cells in vitro: Plasminogen ac- tivator production. Cancer Res. 36,4208-4216.
SOLTER, D., and KNOWLES, B. B. (1975). Immunosur- gery of mouse blastocyst. Proc. Nat. Acad. Sci. USA 72.5099-5102.
SOLTER, D., and KNOWLES, B. B. (1978). Monoclonal antibody defining a stage-specific mouse embryonic antigen (SSEA-1). Proc. Nat. Acad. Sci. USA 75, 5565-5569.
STERN, P. L., MARTIN, G. R., and EVANS, M. J. (1975). Cell surface antigens of clonal teratocarcinoma cells at various stages of differentiation. Cell 6.455-465.
STRICKLAND, S., REICH, E., and SHERMAN, M. I. (1976). Plasminogen activator in early embryogen- esis: Enzyme production by trophoblast and parietal endoderm. Cell 9,231-240.
TUCKER, E. B., and SCHULTZ, G. A. (1978). Two-di- mensional electrophoretic analysis of proteins syn-
thesized during the early cleavage period in rabbit embryos. Exp. Cell Res. 114,438-443.
VAN BLERKOM, J., and MANES, C. (1974). Develop- ment of preimplantation rabbit embryos in vivo and in vitro. II. A comparison of qualitative aspects of protein synthesis. Develop. Biol. 40, 40-51.
VAN BLERKOM, J., and BROCKWAY, G. 0. (1975). Qual- itative patterns of protein synthesis in the preim- plantation mouse embryo. I. Normal pregnancy. Develop. Biol. 44, 148-157.
VAN BLERKOM, J., BARTON, S. C., and JOHNSON, M. H. (1976). Molecular differentiation in the preim- plantation mouse embryo. Nature 259,319-321.
VAN BLERKOM, J., and MCGAUGHEY, R. W. (1978). Molecular differentiation of the rabbit ovum. II. During the preimplantation development of in vivo and in vitro matured oocytes. Develop. Biol. 63, 151-164.
WILEY, L. M., SPINDLE, A. I., and PEDERSEN, R. A. (1978). Morphology of isolated mouse inner cell masses developing in vitro. Develop. Biol. 63, I-10.