
Cytological studies on triploid Trigonella corniculata L.
8
Genetica (1971) ,t2:469--476 CYTOLOGICAL STUDIES ON TRIPLOID TRIGONELLA CORNICULATA L. AVTAR SINGH and R. G. SAINI Department of Genetics, Punjab Agricultural University Ludhiana, India (Received June z 7, z97z ) Two spontaneous triploid plants of Trigondla corniculata (3n = 24) were observed in a population raised from irradiated seeds. Up to seven trivalents per cell were observed. One of the plants appeared to be a double translocation heterozygote involving two non-homologous chromosomes, and therefore could form a quadrivalent, a ring trivalent and bivalents without corresponding univalents. Introduction Trigonella corniculata L., known as "Kasuri Methi" in India, is used as a vegetable, the other economically important species of Trigonella being T. ]oenum-graecum. Autotetraploids of these two and three other species of this genus were produced and studied by RoY & SINGH (1968) and SINGS & ROY (1971). Crosses between the auto- tetraploid and the diploid of T. ]oenum-graecum were abortive due to incompatibility between the two ploidy levels. Two spontaneous triploid plants which form the subject matter of this paper were, however, observed in a population raised from irradiated seeds of T. corniculata. Material and Methods Seeds of T. corniculata were obtained from the Horticulture De- partment of this university and were irradiated with 45 kr gamma rays at the Indian Agricultural Research Institute, New Delhi. The seeds were sown in the field along with control plants. Young inflo- rescences were fixed in acetic-alcohol (1 : 3) for 24 hours and squashed in acetocarmine for cytological examination. Temporary preparations were used for studies and microphotographs. Pollen fertility was
-
Upload
avtar-singh -
Category
Documents
-
view
214 -
download
2
Transcript of Cytological studies on triploid Trigonella corniculata L.

Genetica (1971) ,t2:469--476
CYTOLOGICAL STUDIES ON T R I P L O I D T R I G O N E L L A C O R N I C U L A T A L.
AVTAR SINGH and R. G. SAINI
Department of Genetics, Punjab Agricultural University Ludhiana, India
(Received June z 7, z97z )
Two spontaneous triploid plants of Trigondla corniculata (3n = 24) were observed in a population raised from irradiated seeds. Up to seven trivalents per cell were observed. One of the plants appeared to be a double translocation heterozygote involving two non-homologous chromosomes, and therefore could form a quadrivalent, a ring trivalent and bivalents without corresponding univalents.
Introduction
Trigonella corniculata L., known as "Kasuri Methi" in India, is used
as a vegetable, the other economically important species of Trigonella
being T. ]oenum-graecum. Autotetraploids of these two and three
other species of this genus were produced and studied by RoY &
SINGH (1968) and SINGS & ROY (1971). Crosses between the auto-
tetraploid and the diploid of T. ]oenum-graecum were abortive due to
incompatibility between the two ploidy levels. Two spontaneous
triploid plants which form the subject matter of this paper were,
however, observed in a population raised from irradiated seeds of
T. corniculata.
Material and Methods
Seeds of T. corniculata were obtained from the Horticulture De-
partment of this university and were irradiated with 45 kr gamma
rays at the Indian Agricultural Research Institute, New Delhi. The
seeds were sown in the field along with control plants. Young inflo-
rescences were fixed in acetic-alcohol (1 : 3) for 24 hours and squashed in acetocarmine for cytological examination. Temporary preparations
were used for studies and microphotographs. Pollen fertility was

470 A V T A R S I N G H A N D R. G. S A I N I
calculated on the basis of stainability with acetocarmine. Only the stained pollen grains were measured. One of the triploid plants was attacked by root rot soon after it came into flower and therefore, the morphological comparisons with the diploid were made from the second plant.
Observations
The triploid plants had stouter stems and larger leaves as compared to the irradiated diploids as well as control plants under field con- ditions. The main stem was longer in the triploids. However, the number of branches per plant was less in the triploids. Guard cells, stomatal aperture and pollen grains were bigger in the triploid as compared to the diploid.
The triploid plants had 3n = 24 and showed different chromosomal
associations (Table l a-b). Metaphase I configurations were analysed in 5 pollen mother cells of the first triploid plant and 50 of the second. Three cells showed the presence of a quadrivalent and 8 cells showed a ring trivalent in the second triploid plant (Fig. 1). Range and mean of univalents and trivalents per cell and chiasma frequency of the diploid and the triploids are given in Table 2. The controls regularly formed eight bivalents.
Anaphase I studied in the second triploid plant was irregular (Fig. 2 and Table 3) and only 2 cells out of the 53 studied showed 12-12
distribution; 13-11 was the most frequent distribution. A few lagging univalents divided at anaphase I (Fig. 3). Telophase U was studied
T A B L E l a
CHROMOSOMAL ASSOCIATIONS AT METAPHASE I OF THE FIRST
TRIPLOID PLANT OF T. corniculata
No. C o n f i g u r a t i o n No. of ce l l s P e r c e n t of ce l l s
]. 7III + 3I l 20
2. 6n[ -}- 2ii -}- 2i I 20
3. 5iIi H- 2ii H- 5i l 20
4. 3I I I + 4II + 7I 1 20
5. l i n + 7 i i + 7x 1 20

CYTOLOGY OF TRIPLOID TRIGONELLA CORNICULATA L.
T A B L E lb
CHROMOSOMAL ASSOCIATIONS AT METAPttASE I OF THE SECOND TRIPLOID PLANT OF T. corniculata
471
No. Configurat ion No. of ceils Percen t of cells
1. l IV + 2IZI + 41I + 6I 2 4 2. IlV ~ lIII -~ 5II ~- 7I 1 2 3. 7111 + l i i -~- l i 5 10 4. 6III + 2II + 2I 9 18 5. 5111 + 1OIII + 211 + 21 3 6 6. 5III -~- 311 + 31 8 16 7. 4111 + IOlll + 311 -~- 3I 2 4 8. 4111 + 4ii + 41 6 12 9. 3iii -1- 1Oiii _L 4i I ~_ 41 1 2
10. 3ill + 6ii + 31 2 4 1 1. 3 i i i + 511 + 51 2 4 12. 3111 + 411 + 71 1 2 13. 2Ii1 + 1OiiI + 511 + 5i 2 4 14. 2111 + 611 + 61 4 8 IS. 1III -t- 7II + 71 1 2 16. 1III + 811 + 51 1 2
T A B L E 2
RANGE AND MEAN OF UNIVALENTS AND TRIVALENTS AND CHIASMA FREQUENCY IN DIPLOID AND TRIPLOID T, corniculata
Univalents Tr ivalents X t a per Half X t a peI cell per cell cell per
Range Mean Range Mean chromosome
Diploid . . . . . 15.85 ~z 1.08 *) 0.99 Triploid
(First plant) 2-7 3.40 1-7 4.40 14.60 i 2.07 0.61 Triploid
(Second plant) 1-7 3.50 1-7 4.42 16.74 ~ 2.43 0.69
*) S tandard error.
i n t h e s e c o n d t r i p l o i d p l a n t o n l y a n d w a s i r r e g u l a r ; 130 ce l l s w e r e
s t u d i e d a n d 8 8 . 4 6 % w e r e f o u n d t o h a v e l a g g a r d s . P o l l e n g r a i n s in t h e
d i p l o i d w e r e e l l i p s o i d a l w h i l e t h o s e of t r i p l o i d w e r e e i t h e r e l l i p s o i d a l

472 AVTAR SINGH AND R. G. SAINI
or round (Fig. 4-5). However , the el l ipsoidal pollen grains of the t r ip lo id
were b roader t han those of the diploid and t ended to be round. Pol len
fe r t i l i ty was 99.47 and 57.8% in the d iploid and the second t r ip lo id
respect ively .
TABLE 3
DISTRIBUTION OF CHROMOSOMES AT A N A P H A S E I IN THE
SECOND TRIPLOID PLANT OF T. corniculata
Distribution No. of cells Percent of cells
15-0-9 5 9.25 15-1-8 1 1.89 14-0-10 7 13.20 13-0-11 10 18.87 13-1-10 3 5.66 13-2-9 2 3.77 12-0-12 2 3.77 12-1-11 5 9.25 12-2-10 7 13.20 12-3-9 2 3.77 11-2-11 4 7.85 11-3-10 2 3.77 1 1 - 4 - 9 1 1 . 8 9
10-5--9 1 1.89 10-6-8 l 1.89
T h i r t y a p p a r e n t l y good seeds were produced b y one t r ip lo id and
60 b y the second. There were also some shrivel led seeds.
D i s c u s s i o n
The t r ip lo id p lan t s of T. corniculata, though found in a popula t ion
raised from i r r ad ia t ed seeds, are not the consequence thereof. Doubl ing
of chromosome number in the root cells has been observed to resul t
f rom i r rad ia t ion which also found expression in the pollen mo the r
cells (SINGH & ROY, in press). There seems to be no plausible mecha-
nism for the origin of a t r ip lo id b y i r rad ia t ion of seeds. These t r ip lo ids
might have been produced b y the fusion of a diploid and a no rma l
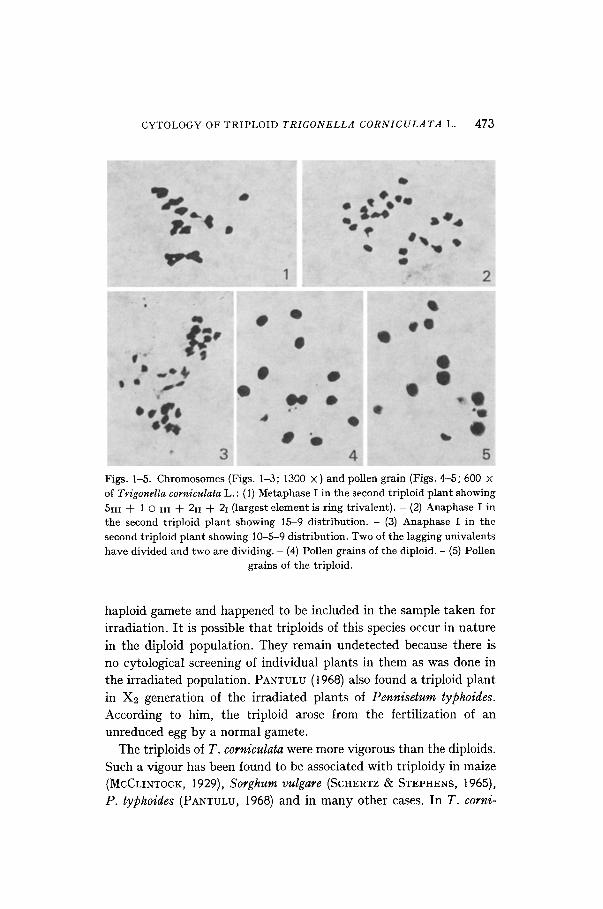
CYTOLOGY OF TRIPLOID TRIGONELLA CORNICULATA L. 473
Figs. 1-5. Chromosomes (Figs. 1-3; 1300 ×)and pollen grain (Figs. 4-5; 600 × of Trigonella corniculata L. : (1) Metaphase I in the second triploid plant showing 5111 + 10 iii + 2ii + 21 (largest element is ring trivalent). - (2) Anaphase I in the second triploid plant showing 15-9 distribution. - (3) Anaphase I in the second triploid plant showing 10-5-9 distribution. Two of the lagging univalents have divided and two are dividing. - (4) Pollen grains of the diploid. - (5) Pollen
grains of the triploid.
haploid gamete and happened to be included in the sample taken for
irradiation. I t is possible tha t triploids of this species occur in nature
in the diploid population. They remain undetected because there is
no cytological screening of individual plants in them as was done in
the irradiated population. PANTULU (1968) also found a triploid plant
in X2 generation of the irradiated plants of Pennisetum typhoides. According to him, the triploid arose from the fertilization of an
unreduced egg by a normal gamete.
The triploids of T. corniculata were more vigorous than the diploids.
Such a vigour has been found to be associated with triploidy in maize (McCLINTOCK, 1929), Sorghum vulgare (SCHERTZ & STEPHENS, 1965),
P. typhoides (PANTULU, 1968) and in m a n y other cases. In T. corni-

474 AVTAR SINGH AND R. G. SAINI
A C C
B B I B D
A A
it. reciprocal translocat ion T
reciprocal ] translocation D
C I I I
A A A C B C
B C D B D D Fig. 6. lViode of a l t e ra t ion in t he two se ts of non -homologous c h r o m o s o m e s b y
two reciprocal translocations in the second triploid plant.
culata, the autotetraploid was shorter and morphologically inferior to the diploid whereas autotetraploids of all other species of Trigonella studied show gigantism (Roy & SINGH, 1968 and SINGH & ROY, 1971).
No pollen mother cell having all the chromosomes paired was observed in the triploids of T. corniculata. In triploid P. typhoides (3n • 21), 50% of the cells studied showed 7 trivalents (PANTOLU, 1968). GILL et al. (1970) found only 2 cells with 7 trivalents out of 347 in triploid P. typhoides. According to these authors, the cause of the low frequency of trivalents was that the triploids were produced by natural cross-pollination of tetraploid and diploid plants and the pollen parent was different from the tetraploid plant. A naturally derived triploid of Secale cereale (3n ~ 21) showed 7 trivalents in only 6 cells out of 200 studied (KISHIKAWA, 1966). In the triploids of T. corniculata, the modal configuration was 6111 + 211 + 21 and 54.4% of cells examined had 5-7 trivalents.
Presence of a quadrivalent, a ring trivalent and more bivalents than expected corresponding univalents was an interesting character- istic of the second triploid plant of T. corniculata. Quadrivalents were observed in 5 out of 347 cells of triploid P. typhoides (GILL et al., 1970). They attributed it to pairing of non-homologous chromosomes.

CYTOLOGY OF TRIPLOID TRIGONELLA C O R N I C U L A T A L. 475
ERICHSON (1963) found more bivalents than univalents in 3 out of 141
cells of triploid Sorghum and suggested the same reason for it. In the present case non-homologous pairing does not seem to be a valid explanation for the formation of quadrivalent, a ring trivalent and a higher number of bivalents than of univalents for the following
reasons : 1. No species of Trigonella has so far been found to have a somatic
number less than 16 and 8 has been suggested as its pr imary basic
number (SINGH & ROY, 1970). 2. In the 3 cells showing a quadrivalent, always two extra univalents
were present. This situation may be explained if two reciprocal translocations are
assumed to have taken place in two sets of non-homologous chromo-
somes as a result of irradiation as shown in Figure 6. These 6 chromo- somes, out of which 4 are altered, may produce the following con-
figurations :
(a) l iv + 2i (b) 1QIII -~- l I I -~- l I
(C) 1OIII -~- 3I
(d) [ o I I I ~- l I I I
(e) 3ii Configurations 1 and 2 of Table lb are explicable by (a), the ring
trivalent by either (b) or (c) or (d) and the presence of extra bivalents
as in configurations l0 and 16 by (e). Confirmation of this assumption may come from the crosses of the progeny that might carry trans- located chromosomes with normal diploid plants.
Progeny of these triploids may be expected to have chromosome numbers varying from 16 to 30 as seen from the anaphase I distri- bution (Table 3). But the laggards and dividing univalents are factors which may modify the theoretical expectation. Moreover, it remains
to be seen if male gametes with chromosome numbers other than 8 would succeed in fertilizing the egg in competition with the possible normal gametes. Nevertheless, aneuploid numbers are expected to be transmit ted from the ovular side which is not subject to gametic
competition. I t may, therefore, be possible ult imately to produce a trisomic series in T. corniculata.
The authors are grateful to Dr. J. L. MINOCHA and Professor 13. S. GILL for their interest in the present investigation.

476 AVTAR SINGH AND R. G. SAINI
REFERENCES
ERICHSON, A. W. ~: J. G. Ross (1963}. A tr iploid from a selfed Sorghum plant . Crop Sci. 3 : 99-100.
GILL, B. S., J. L. MINOCHA, M. K. GEORGE ~; D. S. BRAR (1970). Cytological s tudies in diploid, autotr iploid and auto te t rap lo id pearl millet. J. Res., Punjab Agr. Univ. 7: 417-422.
KISmKAWA, H. (1966). Cytological s tudy on tf iploid rye wi th four accessory chromosomes. Jap. J. Genet. 41 : 427-437.
McCLINTOCK, t3. (1929). A cytological and genetical s tudy of tr iploid maize. Genetics 14: 180-222.
DANTULU, J. V. (1908). Meiosis in an autotr iploid pearl millet. Caryologia 21: 11-15.
RoY, R. P. & A. SINGH (1968). Cytomorphological s tudies of the colchicine- induced te t raploid Trigonella [oenum-graecum L. Genet. 1ber. 20: 37-54.
SCHERTZ, K. F. & J. G. STEPHENS (1965). Origin and occurrence of tr iploids of Sorghum vulgare Pers. and their chromosomal and morphological charac- teristics. Crop. Sci. 5: 514-516.
SING~, A. & R. P. R o y (1970). Karyological s tudies in Trigonella, Ind,go/era and Phaseolus. Nucleus 13: 41-54.
SINGH, A. & R. I ~. RoY (1971). Studies on colchiploids of four species of Tri- gonella. Cytologia 36 : 133-142.
SINGH, A. and R. P. RoY. X-ir radia t ion studies on Trigonella /oenum-graecum L. Genet. Iber. (In press).


















