
Combination of Levo-Tetrahydropalmatine and Low Dose...
10
1521-0103/357/2/248–257$25.00 http://dx.doi.org/10.1124/jpet.115.229542 THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 357:248–257, May 2016 U.S. Government work not protected by U.S. copyright Combination of Levo-Tetrahydropalmatine and Low Dose Naltrexone: A Promising Treatment for Prevention of Cocaine Relapse Sarah Sushchyk, Zheng-Xiong Xi, and Jia Bei Wang Department of Pharmaceutical Sciences, University of Maryland Baltimore, Baltimore, Maryland (S.S., J.B.W.); Molecular Targets and Medications Discovery Branch, National Institute on Drug Abuse, Intramural Research Program, Baltimore, Maryland (Z.-X.X.) Received October 27, 2015; accepted February 19, 2016 ABSTRACT Relapse to drug use is often cited as the major obstacle in overcoming a drug addiction. Whereas relapse can occur for a myriad of reasons, it is well established that complex neuroadaptations that occur over the course of addiction are major factors. Cocaine, as a potent dopamine transporter blocker, specifically induces alterations in the dopaminergic as well as other monoaminergic neurotransmissions, which lead to cocaine abuse and dependence. Evidence also sug- gests that adaptations in the endogenous opioids play impor- tant roles in pathophysiology of cocaine addiction. Following this evidence, we investigated a combination medication, levo- tetrahydropalmatine (L-THP) and low dose naltrexone (LDN), targeting primarily dopaminergic and endogenous opioid sys- tems as a cocaine-relapse-prevention treatment. In the present study Wistar rats were used to assess the effects of L-THP and LDN on cocaine self-administration, drug-seeking behavior during cocaine reinstatement, spontaneous locomotion, and effects on the endogenous opioid system. We determined that the combination of L-THP and LDN reduces drug-seeking behavior during reinstatement more potently than L-THP alone. Additionally, the combination of L-THP and LDN attenuates the sedative locomotor effect induced by L-THP. Furthermore, we revealed that treatment with the combination of L-THP and LDN has an upregulatory effect on both plasma b-endorphin and hypothalamic POMC that was not observed in L-THP-treated groups. These results suggest that the combination of L-THP and LDN has great potential as an effective and well-tolerated medication for cocaine relapse prevention. Introduction Cocaine is a potent psychostimulant with a high risk for addiction and abuse. Despite the extensive research in the past decades, there remains no FDA-approved treatments for mitigating cocaine addiction (Nestler, 2005; Shorter and Kosten, 2011). Cocaine addiction induces alterations in the mesolimbic dopamine (DA) system, and considerable efforts were placed in the development of DA receptor antagonists as antiaddiction treatments (Dackis and O’Brien, 2001; Platt et al., 2002; Koob and Volkow, 2010). However, DA antago- nists alone are often unsuccessful in clinical trials (Haney et al., 2001). Whereas DA plays a central role in cocaine addiction, many other neurotransmitters take part in the complex changes in the brain over the course of cocaine addiction (Trigo et al., 2010; Yoo et al., 2012). b-Endorphin is emerging as an important player in the development of addiction (Olive et al., 2001; Roth-Deri et al., 2003, 2004, 2008; Simmons and Self, 2009; Nguyen et al., 2012; Dikshtein et al., 2013). Attenuated b-endorphin has been demonstrated to play an important role in relapse to nicotine- or alcohol-seeking behavior (Marinelli et al., 2004; Shaw and al’Absi, 2008; Ueno et al., 2014). Pro-opiomelanocortin (POMC), also a part of the endogenous opioid system, is the precursor to b-endorphin, adrenocorticotropic hormone, or melanocyte-stimulating hormone. POMC is thought to play an important role in reinstatement to drug-seeking behavior, and in a recent study it was observed that animals with higher instances of rein- statement displayed a significant reduction in POMC expres- sion (Zhou et al., 2015). Taken together these data suggest that the endogenous opioid system may be an important target in medication development for treatment of cocaine dependence. Levo-tetrahydropalmatine (L-THP) is a purified alkaloid isolated from the herb Corydalis yanhusuo, native to China. It reduces the reward- and drug-seeking effects of cocaine, oxycodone, and heroin (Jin et al., 1986a,b; Marcenac et al., 1986; Ding, 1987; Mantsch et al., 2007; Liu et al., 2009; Mantsch et al., 2010; Figueroa-Guzman et al., 2011; Wang and Mantsch, 2012), suggesting a potential for L-THP in treatment of drug abuse and addiction. The dopamine receptor–blocking properties of L-THP may substantially contribute to its ability to attenuate drug-seeking behavior. Clinically, a pilot study conducted to assess the utility of L-THP for the treatment of heroin addiction determined that 30 mg L-THP given twice The National Institutes of Health National Institute on Drug Abuse [Grant DA-031401 (J.B.W.)] and the National Institute on Drug Abuse Intramural Research Program supported this work. The authors declare no finical conflict. dx.doi.org/10.1124/jpet.115.229542. ABBREVIATIONS: ANOVA, analysis of variance; DA, dopamine; DAPI, 2-(4-amidinophenyl)-1H-indole-6-carboxamidine; FR, fixed-ratio; LDN, low dose naltrexone; L-THP, levo-tetrahydropalmatine; NTX, naltrexone; POMC, pro-opiomelanocortin; Ubc, Ubiquitin. 248 at ASPET Journals on May 19, 2020 jpet.aspetjournals.org Downloaded from
Transcript of Combination of Levo-Tetrahydropalmatine and Low Dose...

1521-01033572248ndash257$2500 httpdxdoiorg101124jpet115229542THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 357248ndash257 May 2016US Government work not protected by US copyright
Combination of Levo-Tetrahydropalmatine and Low DoseNaltrexone A Promising Treatment for Prevention ofCocaine Relapse
Sarah Sushchyk Zheng-Xiong Xi and Jia Bei WangDepartment of Pharmaceutical Sciences University of Maryland Baltimore Baltimore Maryland (SS JBW) Molecular Targetsand Medications Discovery Branch National Institute on Drug Abuse Intramural Research Program Baltimore Maryland (Z-XX)
Received October 27 2015 accepted February 19 2016
ABSTRACTRelapse to drug use is often cited as the major obstacle inovercoming a drug addiction Whereas relapse can occur fora myriad of reasons it is well established that complexneuroadaptations that occur over the course of addiction aremajor factors Cocaine as a potent dopamine transporterblocker specifically induces alterations in the dopaminergicas well as other monoaminergic neurotransmissions whichlead to cocaine abuse and dependence Evidence also sug-gests that adaptations in the endogenous opioids play impor-tant roles in pathophysiology of cocaine addiction Followingthis evidence we investigated a combination medication levo-tetrahydropalmatine (L-THP) and low dose naltrexone (LDN)targeting primarily dopaminergic and endogenous opioid sys-tems as a cocaine-relapse-prevention treatment In the present
study Wistar rats were used to assess the effects of L-THP andLDN on cocaine self-administration drug-seeking behaviorduring cocaine reinstatement spontaneous locomotion andeffects on the endogenous opioid system We determined thatthe combination of L-THP and LDN reduces drug-seekingbehavior during reinstatement more potently than L-THP aloneAdditionally the combination of L-THP and LDN attenuates thesedative locomotor effect induced by L-THP Furthermore werevealed that treatment with the combination of L-THP and LDNhas an upregulatory effect on both plasma b-endorphin andhypothalamic POMC that was not observed in L-THP-treatedgroups These results suggest that the combination of L-THPand LDN has great potential as an effective and well-toleratedmedication for cocaine relapse prevention
IntroductionCocaine is a potent psychostimulant with a high risk for
addiction and abuse Despite the extensive research in thepast decades there remains no FDA-approved treatments formitigating cocaine addiction (Nestler 2005 Shorter andKosten 2011) Cocaine addiction induces alterations in themesolimbic dopamine (DA) system and considerable effortswere placed in the development of DA receptor antagonists asantiaddiction treatments (Dackis and OrsquoBrien 2001 Plattet al 2002 Koob and Volkow 2010) However DA antago-nists alone are often unsuccessful in clinical trials (Haneyet al 2001) Whereas DA plays a central role in cocaineaddiction many other neurotransmitters take part in thecomplex changes in the brain over the course of cocaineaddiction (Trigo et al 2010 Yoo et al 2012) b-Endorphinis emerging as an important player in the development ofaddiction (Olive et al 2001 Roth-Deri et al 2003 2004 2008Simmons and Self 2009 Nguyen et al 2012 Dikshtein et al2013) Attenuated b-endorphin has been demonstrated to play
an important role in relapse to nicotine- or alcohol-seekingbehavior (Marinelli et al 2004 Shaw and alrsquoAbsi 2008 Uenoet al 2014) Pro-opiomelanocortin (POMC) also a part of theendogenous opioid system is the precursor to b-endorphinadrenocorticotropic hormone or melanocyte-stimulatinghormone POMC is thought to play an important role inreinstatement to drug-seeking behavior and in a recent studyit was observed that animals with higher instances of rein-statement displayed a significant reduction in POMC expres-sion (Zhou et al 2015) Taken together these data suggestthat the endogenous opioid system may be an importanttarget in medication development for treatment of cocainedependenceLevo-tetrahydropalmatine (L-THP) is a purified alkaloid
isolated from the herb Corydalis yanhusuo native to China Itreduces the reward- and drug-seeking effects of cocaineoxycodone and heroin (Jin et al 1986ab Marcenac et al1986 Ding 1987 Mantsch et al 2007 Liu et al 2009Mantsch et al 2010 Figueroa-Guzman et al 2011Wang andMantsch 2012) suggesting a potential for L-THP in treatmentof drug abuse and addiction The dopamine receptorndashblockingproperties of L-THP may substantially contribute to its abilityto attenuate drug-seeking behavior Clinically a pilot studyconducted to assess the utility of L-THP for the treatment ofheroin addiction determined that 30 mg L-THP given twice
The National Institutes of Health National Institute on Drug Abuse [GrantDA-031401 (JBW)] and the National Institute on Drug Abuse IntramuralResearch Program supported this work The authors declare no finical conflict
dxdoiorg101124jpet115229542
ABBREVIATIONS ANOVA analysis of variance DA dopamine DAPI 2-(4-amidinophenyl)-1H-indole-6-carboxamidine FR fixed-ratio LDN lowdose naltrexone L-THP levo-tetrahydropalmatine NTX naltrexone POMC pro-opiomelanocortin Ubc Ubiquitin
248
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
daily increased heroin abstinence over 3months by 63 (Yanget al 2008) However L-THP also produces significantsedative effects in a dose-dependent manner (Xi et al 2007Xu et al 2013) whichmay limit its utility in treatment of drugaddictionNaltrexone primarily a m-opioid receptor antagonist is
FDA-approved for treatment of alcoholism and opioid detox-ification At therapeutic doses naltrexone reduces drug-seeking behavior in rodents but is unsuccessful or ineffectivein most human clinical studies owing to significant un-wanted side effects poor patient compliance and highattrition rates (Schmitz et al 2001 Grassi et al 2007Schmitz et al 2009 Schmitz et al 2014) However thetolerability of naltrexone is significantly improved whengiven at subtherapeutic doses (Zagon et al 2014) Low dosenaltrexone (LDN) defined as 4 mg per day in humans(Brown and Panksepp 2009 Mannelli et al 2010Wee et al2012 Cordery et al 2014) is becoming commonly used as along term treatment of autoimmune disorders (Rahn et al2011 Segal et al 2014) Although the exact mechanismsunderlying these actions are unknown one possibility is thatLDN may functionally upregulate endogenous opioid systemthrough m-opioid receptor antagonismWe therefore propose here that the combination of L-THP
and LDN may be safer than L-THP or LDN alone in pre-vention of cocaine relapse This is on the basis of thefollowing two assumptions First such a combination mayproduce additive therapeutics effects which may signifi-cantly lower each drug dose and therefore attenuate un-wanted side-effects produced by each individual drugs andsecond coadministration of LDN with L-THP may alsoantagonize L-THP-induced sedation by normalization orupregulation of the endogenous opioid system in cocaine-dependent subjectsTo test this hypothesis the present study was designed to
determine whether the combination of L-THP and LDN morepotently reduces relapse to drug-seeking behavior and withless unwanted sedative effect than L-THP alone in rat modelsIn addition we also investigated whether coadministration ofL-THP and LDN alters plasma b-endorphin concentrationsand hypothalamic POMC expression
Materials and MethodsAnimals Experimentally naiumlve male Wistar rats (Charles River
Laboratories Raleigh NC) weighing 250ndash300g were used for allexperiments Animals were housed individually in a reverse lightdark cycle with lights off at 700 and lights on at 1900 Animals were
given ad libitumaccess to food andwater All experimental procedureswere conducted in accordance with the Guide for the Care and Use ofLaboratory Animals of the US National Academy of Sciences andwere approved by the IACUC of University of Maryland and theNational Institutes of Health National Institute of Drug Abuse
Jugular Vein Catheterization The cocaine self-administrationprocedures surgeries and training schedules were the same aspreviously reported (Xi et al 2007) Animals were prepared for self-administration experiments by surgical catheterization of rightexternal jugular vein Venous catheters which were constructed ofMicro-Renathane inner diameter 0012 inches outer diameter 0025inches (Braintree Scientific Braintree MA) was inserted 12 cm intothe right jugular under sodium pentobarbital anesthesia (65 mgkgip) with standard aseptic surgical procedures Catheters exited thejugular vein and passed subcutaneously to the top of the skull wherethey were connected to a modified 24-gauge cannula (Plastics OneRoanoke VA) and mounted to the skull with jewelerrsquos screws anddental acrylic During experimental procedures catheters wereconnected to an infusion pump via tubing encased in a protectivemetal spring from the headmount to the top of the chamber Catheterswere flushed directly prior to and directly following training sessionswith a combination of heparingentamicin (5 IU 1 10 mgml) in 09saline solution to prevent blockage and infections
Apparatus Intravenous self-administration experiments wereconducted in operant test-response chambers 32 25 33cm (MedAssociates Inc St Albans VT) House lights were turned on at thebeginning of each session Every test chamber contained two testlevers one active and one inactive Depression of the active leveractivated infusion pump depression of inactive lever was counted buthad no consequence The house light was turned on at the start of each3-hour test session To aid acquisition and maintenance of drug self-administration behavior each drug infusion was always paired with aconditioned cue-light and a cue-sound (tone) The light and thespeaker were installed 12 cm above the active lever The light andtone were paired (on) during the 46-second drug infusion durationThere was no time-out after the fixed-ratio (FR) 1 or 2 drug infusionAll the animals were first trained on FR1 for approximately seventraining sessions with 1 mgkg per infusion of cocaine (to facilitate theacquisition of self-administered cocaine) and then switched to FR2 for05 mgkg per infusion of cocaine self-administration in the followingexperiments Self-administration tests were completed during FR2conditions Scheduling of experimental events and data collectionwere accomplished using Med Associates software
Cocaine Self-Administration Training Animals were allowedto recover from surgical catheterization After recovery they wereplaced into operant chamber and allowed to lever press for intrave-nous cocaine (1 mgkg per infusion) delivered as 008 ml over anintertrial interval of 46 seconds on an FR1 reinforcement scheduleDuring the intertrial interval additional lever presses were recordedbut did not result in additional infusions After achieving stablecocaine self-administration animals were transitioned to 05 mgkgper infusion on a FR1 schedule and transitioned to the final schedule
TABLE 1l-THP and LDN Treatments During Extinction
Self-Administration Extinction Treatment (Daily) Reinstatement Treatment
Cocaine 05 mgkgper inf FR2
Vehicle (n = 7) Cocaine 10 mgkg ipNone (n = 5) 3 mgkg L-THP and 01 mgkg LDN 30 minutes prior
Cocaine 10 mgkg ipNone (n = 9) 5 mgkg L-THP and 01mgkg LDN
30 minutes priorCocaine 10 mgkg ip
01 mgkg LDN (n = 6) Cocaine 10 mgkg ip3 mgkg L-THP (n = 9) Cocaine 10 mgkg ip5 mgkg L-THP (n = 10) Cocaine 10 mgkg ip3 mgkg L-THP and 01mgkg LDN (n = 10) Cocaine 10 mgkg ip5 mgkg L-THP and 01mgkg LDN (n = 11) Cocaine 10 mgkg ip
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 249
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
of 05 mgkg per infusion on an FR2 schedule Each test session lasted3 hours After stable self-administration pretreatments of 3 mgkg5mgkg or 10mgkg L-THP 01 mgkg LDN and 3mgkg1 01 mgkg5 mgkg 1 01 mgkg or 10 mgkg 1 01 mgkg L-THP 1 LDN wereadministered intraperitoneally 30 minutes prior to the experimentalsession
Cocaine Reinstatement Following stable self-administrationexperiments seven consecutive training session in which animalsdemonstrated less that 10 change in drug-taking behavior animalswere transitioned to extinction in which depression of the previouslyactive lever resulted in no infusion cue or light signal Infusionsyringes were filled with physiologic saline Animals were randomlyassigned to one of the treatment groups described in Table 1Immediately following each test session animals were weighed andgiven an intraperitoneal treatment injection corresponding to theirassigned group Extinction continued until animals responded with anumber of active lever presses no more than 10 of that achievedduring cocaine self-administration over the 3-hour period Twenty-four hours after extinction was achieved animals were reinstatedusing ip 10 mgkg cocaine (Amen et al 2011) During cocainereinstatement light and environmental cues remained off
Spontaneous Locomotor Activity Locomotor experiments wereconducted in VersaMax open-field test chambers 40 cm 40 cm(Omnitech Electronics Inc Columbus OH) Habituation and baselinewere established over 3 days Animals were weighed prior to eachsession and randomly assigned to treatment groups described inTable 2 During the drug treatment phase animals were placed intoopen-field chambers for 1 hour After the first hour animals wereremoved injected (intraperitoneally) with the assigned treatment andplacedback into the chamber for a secondhour Treatment continued for10 days Ten days was chosen as the typical extinction treatment periodin the reinstatement experiment Locomotion score was calculated bysubtracting baseline average from total movements after injections
Cocaine Forced Abstinence After self-administering cocaine orsucrose for 13 sessions animals were moved to forced abstinenceForced abstinence differed from extinction in that animals remainedin their home cages were handled administered injections every24 hours and did not return to the operant chambers until day 23Groups are described in Table 3 On day 23 animals were placed backinto operant chambers to measure drug craving as determined byactive lever presses Animals were sacrificed 30 minutes followingfinal session and tissue and plasma were harvested
Tissue Sample Collection Thirty-minutes after the final train-ing session animals were rapidly sacrificed via decapitation Brainswere quickly removed and immersed into dry ice-chilled methyl-butane for 10 seconds and stored in Tissue-TekOCTmedium (SakuraTorrance CA) at ndash80degC Blood was collected from the trunk post-decapitation and stored in Lavender Vacutainer Tubes containingEDTA (Becton Dickinson Franklin Lakes NJ) Blood was transferredto centrifuge tubes containing aprotinin Blood samples were centri-fuged at 1600g for 15minutes at 4degC and the plasmawas removed andstored at ndash80degC
Quantification of b-Endorphin Content b-Endorphin wasquantified using a commercially available EIA kit (Phoenix Pharma-ceuticals Hayward CA) Plasma was acidified using equal amounts ofbuffer A and centrifuged at 10000g for 20 minutes at 4degC A SEP-column was equilibrated using washing buffer B followed by washing
buffer A Plasma was loaded onto the column and washed three timeswith buffer A Peptides were eluted using buffer B and the eluent wascollected into a polystyrene tube evaporated and stored at ndash20degCPeptides were reconstituted with 125 ml of assay buffer Reconstitutedsamples were further diluted to 150 with assay buffer Assay wasperformed according to manufacturerrsquos specifications Absorbancewas read at 450 nm
Visualization and Quantification of POMC by RNAscopeBrains were rapidly harvested and snap-frozen 30 minutes after thecompletion of the above behavioral test on day 23 (see Cocaine ForcedAbstinence) Brains were sliced 014-mm thick and mounted to slidesThere were five animals in each of the treatment groups and six sliceswere analyzed from each animal for a total of 30 analyzed slices pergroup OCT-embedded brainsweremounted on cryostat platformwithOCT medium Tissue was sectioned at the arcuate nucleus (Arc)beginning at approximately and ndash17 mm and ndash396 mm bregma asdetermined from the The Rat Brain in Stereotaxic Coordinates(Paxinos and Watson 2006) and 014-mm tissue slices were mountedon Superfrost Plus Gold Slides (FisherScientificThermoFisherScientific Sunnyvale CA)
Tissue sections were fixed in 10 formalin for 15 minutes at 4degCSlides were dehydrated in ethanol baths of 50 70 and 100 atroom temperature Tissues were then incubated in Pretreatment 4from the RNAscope Pretreatment Kit (Advanced Cell DiagnosticsHayward CA) for 15 minutes at room temperature Slides were driedand the probe for POMC was added for a 2-hour incubation in ahybridization oven The hybridization oven was maintained at 40degCfor all incubations Probe hybridization was carried out in thehybridization oven in four amplification steps in alternating 30- and15-minute incubations After each amplification step slides werewashed two times in 1wash buffer for 2 minutes DAPI was added toslides for 10 seconds and cover-slipped with Fluoro-Gel (ElectronMicroscopy Services Hatfield PA)
Wide-field fluorescent images of the arcuate nucleus were cap-tured using Nikon Eclipse Ti-E with an 20 objective Images weredeconvoluted using NIS Elements Software (Nikon InstrumentsMelville NY ) and pseudo-colored (POMC red Ubiquitin (Ubc) greenand DAPI blue) Image Processing and Analysis by Java (ImageJNIH) was used to quantify total mRNA signal
Drugs The drugs used in the present study were as followsCocaine hydrochloride obtained from NIDA was dissolved in phys-iologic saline Levo-tetrahydropalmatine was obtained from thelaboratory of Stephen Hoag at the University of Maryland BaltimoreThe L-THP was dissolved in 01 M H2SO4 brought to a pH of 556 02with 01 M NaOH and brought to correct volume with sterile waterL-THP concentrations of 3 mgkg 5 mgkg and 10 mgkg were used inthe study For L-THP and LDN combinations naltrexone hydrochlo-ride (Sigma-AldrichSigmaMillipore St Louis MO) was dissolved inthe L-THP solution to a concentration of 01 mgkg The 01 M H2SO401MNaOHsterilewater solution pH of 556 02 was used as vehicleand for the 01 mgkg LDN dose Additionally each solution wassyringe-passed through 022-mm filters to ensure the solutions werefree of contaminates
Data Analysis Statistical analyses were performed usingGraphPad Prism 500 (La Jolla CA) for Windows Data are presentedas mean 6 SEM Studentrsquos t test One-way and two-way analyses ofvariance (ANOVAs) followed by Bonferroni analysis were used forcomparisons between groups where appropriate Effect size wascalculated using h2 for ANOVAs and Pearsonrsquos r for correlations andacceptable level of statistical significance for all tests was P 005
TABLE 2l-THP and LDN Treatment on Locomotion
Habituation (Days 1ndash3) Treatment (Days 4ndash13)
No Treatment Vehicle01 mgkg LDN3 mgkg L-THP5 mgkg L-THP3 mgkg L-THP and 01mgkg LDN5 mgkg L-THP and 01mgkg LDN
TABLE 3l-THP and LDN Treatment on Cue Induced Cocaine-Seeking
Self-Administration Forced Abstinence Treatment (Daily)
Cocaine 05 mgkg per inf Vehicle3 mgkg L-THP3 mgkg L-THP and 01 mgkg LDN
250 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
ResultsEffects of L-THP and LDN on Cocaine Self-
Administration In our study setting animals acquiredstable cocaine self-administration over 14 experimental ses-sions (1119 6 3975 lever presses per session) Pretreat-ment with L-THP produced a biphasic drug-taking effect Thelow doses of L-THP produced an initial increase in self-administration whereas the high dose of L-THP produced areduction in cocaine self-administration (Fig 1) One-wayANOVA revealed a significant drug treatment main effect[F(759)5 1156 P 0001] Effect-size calculations suggest amoderate to highly significant practical effect [h2 5 058]Post-hoc Bonferroni analyses revealed a significant reduction
of cocaine self-administration after 10 mgkg L-THP and10 mgkg L-THP and 01 mgkg LDN combination (P 005)Animals administered LDN exhibited no change in self-administration (P 005) Treatment with the combinationL-THP and LDN had no significant effect as with the respec-tive doses of L-THP alone (P 005)Effects of L-THP and LDN on Cocaine-Induced Rein-
statement of Drug-Seeking Behavior After stable self-administration was achieved animals went to extinction untildrug-seeking behaviorwas extinguished defined as active leverresponses less than 10 of the responses achieved over a stablecocaine self-administration experimental sessions (650 6 142lever presses per session) Pretreatment with L-THP andorLDN administered 30 minutes prior to reinstatement testsignificantly attenuated cocaine-induced (10 mgkg ip) rein-statement of drug-seeking behavior [F(263) 5 354 P 005two-wayANOVA] (Fig 2) Effect size however was determinedto be low to moderate [h2 5 038] Bonferroni post-hoc analysesrevealed a significant reduction in drug-seeking after 3 mgkgL-THPand 01mgkgLDN (P 005) and the 5mgkg L-THPand01 mgkg (P 001) compared with the vehicle control groupEffects of Chronic L-THP and LDN Pretreatment on
Cocaine-Induced Reinstatement of Drug-SeekingBehavior Animals in this experiment were exposed to thesame self-administration conditions used in the previousexperiment however during extinction training the animalswere given L-THP andor LDN daily for 10 consecutive daysduring extinction At 24 hours after the last drug injectioncocaine-induced reinstatement of drug-seeking behavior wassignificantly lower in all drug-treatment groups than in thevehicle control group of rats (Fig 3) One-way ANOVA for thesummarized data in Fig 3C revealed a significant treatmentmain effect [F(551)5 9085P 0001] Effect-size calculationsuggests a moderate applied effect [h2 5 049] Bonferronipost-hoc analyses demonstrated a significant reduction incocaine-seeking after 01 mgkg LDN (P 005) 3 mgkgL-THP (P 001) 3 mgkg L-THP and 01 mgkg LDN (P 0001) 5 mgkg L-THP (P 0001) and 5 mgkg L-THP and01 mgkg LDN (P 0001) administration
Fig 1 Effect of pretreatment of L-THP and LDN on cocaine self-administration After animals achieved stable cocaine self-administrationanimals were administered L-THP LDN or L-THP and LDN 30 minutesprior to experimental session Intraperitoneal administration of L-THPproduced a biphasic effect Low doses (3 mgkg and 5 mgkg L-THP n = 7)increased and a high dose (10 mgkg n = 7 and 10 mgkg L-THP and01mgkg LDN n = 7) induced sedation inhibiting cocaine self-administrationP 005 compared with vehicle treatment determined by one-wayANOVA LDN neither altered cocaine self-administration by itself (01 mgkgLDN n = 7) nor altered L-THPrsquos effects on cocaine self-administrationbehavior (3 mgkg L-THP and 01 mgkg LDN n = 7 and 5mgkg L-THP and01 mgkg LDN n = 7) Values are expressed as mean 6 SEM
Fig 2 Effect of single pretreatment ofL-THP and LDN on cocaine reinstatementIn the experimental session followingextinction animals were administeredintraperitoneal injections of L-THP andLDN 30 minutes prior to a priming doseof 10 mgkg cocaine Two-way ANOVAshowed that 3 mgkg L-THP and 01 mgkgLDN (n = 5) and 5 mgkg L-THP and01 mgkg LDN (n = 9) dose-dependentlyand significantly inhibited reinstatementof cocaine-seeking behavior assessed byactive lever presses P 05 P 001respectively compared with vehicle group(n = 7) Values expressed asmean6 SEM
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 251
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Locomotion Animalswere administered L-THP LDN or L-THP1 LDN daily for 10consecutive days The timeline of 10 days was selected on thebasis of the findings in the above reinstatement experiments(see previous section) The baseline was created using theaverage of the spontaneous locomotion measurements takenover the first 3 consecutive days (3381 6 1681 cm averagebaseline) Baseline was subtracted from total movementsfor each day A dose of 3 mgkg or 5 mgkg L-THP alonesignificantly inhibited spontaneous locomotor activities overthe 60 minutes (two-way ANOVA [F(2140) 5 2234 P 0001]) whereas 01 mgkg LDN alone had no effect onspontaneous locomotion (Fig 4A) Both combination doses ofL-THP and LDN did not produce a significant reduction inspontaneous locomotion [F(2195) 5 02794 P 5 02794 two-way ANOVA] (Fig 4B) Effect size was determined to bemoderate to highly significant [h2 5 051]Effects of L-THP and LDN on Cue-Induced Cocaine-
Seeking After stable cocaine self-administration wasachieved animals underwent forced abstinence in their home
cages in the animal facility for 9 days during which 3 mgkgL-THP 3 mgkg L-THP and 01 mgkg LDN or vehicle wasgiven daily to three separate groups of rats (Fig 5A) Animalswere then placed back into the same self-administrationchambers where they obtained cocaine self-administrationbefore on day 23 Re-exposure to cocaine-associated contex-tual cues induced a significant cocaine-seeking behavior(ie active lever presses) in the absence of cocaine orcocaine-associated cues (Fig 5B the vehicle control group)Pretreatment with 3 mgkg L-THP or 3 mgkg L-THP and01 mgkg LDN produced a significant reduction in cue-induced drug-seeking compared with the vehicle controlgroup and the combination group was more effective thenL-THP alone One-way ANOVA for the data shown in Fig 5Brevealed a significant treatmentmain effect [F(319)5 3413P 5 0043] Moreover effect-size calculations revealed amoderate practical effect [h2 5 040] Studentrsquos t testsrevealed a significant reduction in cocaine-seeking after3 mgkg L-THP [t(8) 5 2354 P 5 00464] or 3 mgkg L-THPand 01 mgkg LDN [t(8) 5 3581 P 5 00072]
Fig 3 Effect of 10 days administration of L-THP and LDN on cocaine reinstatement Animals received intraperitoneal injections for 10 days untilextinction was reached In the experimental session following animals were primed with 10 mgkg cocaine and placed back into operant chambers(A and B) Extinction timeline arrows representing administration of L-THP (3 mgkg n = 9 and 5 mgkg n = 10) LDN (01 mgkg n = 6) or L-THP andLDN combinations (3mgkg and 01 mgkg n = 10 and 5mgkg and 01 mgkg n = 11) (C) Significantly inhibited reinstatement of drug-seeking behaviorassessed by active lever responses expressed as mean 6 SEM Bonferroni post-hoc analysis determined that both the combinations of 3 mgkg L-THPand 01 mgkg LDN and 5mgkg L-THP and 01 mgkg LDNwere more efficacious than any of the single doses P 05 P 01 P 0001 P00001 compared with vehicle group (n = 7)
252 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Plasma Concentrationsof b-Endorphin in Rats after Forced Cocaine AbstinenceAfter the completion of the above behavioral test plasmasamples were collected 30 minutes via the implanted in-travenous catheter Coadministration of 3 mgkg L-THP and01 mgkg LDN significantly elevated plasma b-endorphinlevel compared with the vehicle control group A dose of3 mgkg L-THP alone had no effect (Fig 6) One-way ANOVAfor the data shown in Fig 6 revealed a significant treatmentmain effect [F(339) 5 392 P 5 0016] However the effect-size calculations demonstrate a moderate to low significanteffect [h2 5 024]Effects of L-THP and LDN on POMC mRNA Expres-
sion in Rats after Forced Cocaine Abstinence Figure 7Ashows the RNAscope in situ hybridization data illustratingthat coadministration of 3 mgkg L-THP and 01 mgkg LDNsignificantly elevated POMC expression in the hypothala-mus whereas 3 mgkg L-THP alone had no effect One-wayANOVA for the summarized data shown in Fig 7B revealed asignificant treatment main effect [F(3119) 5 6529 P 500004] Effect-size calculations suggest only a minimalpractical effect [h2 5 014] Bonferroni post-hoc analysesindicated a significant increase in POMC expression after3 mgkg L-THP and 01 mgkg LDN (P 005) but not after3 mgkg L-THP alone (P 005) compared with the vehiclegroupCorrelation Between Cocaine-Seeking Behavior
b-Endorphin and POMC Expression Figure 8A showsthe strong significant negative correlation between activelever presses during reinstatement and plasma concentrationsof b-endorphin [r(15)5 ndash0617 P5 00142] Also shown in Fig8B is the weak positive correlation between POMC expressionand plasma concentrations of b-endorphin [r(15) 5 0361
P 5 0187] A correlation was not observed between POMCexpression and active lever presses during reinstatementshown in Fig 8C [r(15) 5 ndash0214 P 5 0440]
DiscussionCombination medications are becoming increasingly com-
monplace in clinical practice and can be especially effectivewhen used to treat individuals suffering from mental disor-ders and who are multisymptomatic (Tohen et al 2003Kupfer et al 2012 Biler et al 2014 Shorter et al 2015) Inthe case of relapse prevention drug cravings may beeffectively managed when medications treat both behavioraland physiologic symptoms However the majority of phar-maceuticals developed for the treatment of substance abusedisorders target one pathway The combination of L-THP andLDN targets both dopaminergic signaling and the regulationof endogenous opioids In this report we provide evidencethat the combination of L-THP and LDN is more potent withfewer unwanted side effects than L-THP alone on the basis ofthe following observations 1) L-THP and LDN has demon-strated significantly greater effect in attenuating drug-seeking behavior than L-THP alone 2) L-THP and LDN whenadministered together do not result in a reduction ofspontaneous locomotion 3) L-THP and LDN appears toincrease peripheral concentrations of b-endorphin 4) L-THPand LDN appears to upregulate expression of POMC in thearcuate nucleus
L-THP has been investigated as a potential treatment ofaddiction by several other groups (Mantsch et al 2007 2010Figueroa-Guzman et al 2011 Wang and Mantsch 2012)Presently we determined that L-THP produced a biphasiceffect and this is consistent with the results of previous study
Fig 4 Effect of L-THP and LDN onspontaneous locomotion Animals estab-lished a baseline over 3 consecutive daysfollowing that they were given intraperi-toneal injections of the assigned treat-ments for 10 days Values are expressedas mean of the locomotor score 6 SEMn = 6 for all groups Two-way ANOVAdetermined treatment had a significanteffect on spontaneous locomotion P00001 (A) L-THP (3 mgkg and 5 mgkg)produced a dose-dependent reduction onlocomotion and LDN (01 mgkg) had noeffect on locomotion (B) Effect of co-administration of L-THP and LDN
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 253
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
(Xi et al 2007) The increase in cocaine self-administration isattributed to the partial blockade of postsynaptic DA recep-tors which decrease the rewarding properties of cocaine andlead to a compensatory increase in drug-taking Even as thehigher doses of L-THP drastically reduced cocaine self-administration we also noted a strong sedative effectassessed by a decrease in locomotion suggesting that nonspe-cific sedation or locomotion impairment is the main cause forreduced self-administration in the high L-THP dose group Incontrast to L-THP LDN had no effect on cocaine self-administration There was no difference in cocaine self-administration between animals receiving lower doses ofL-THP or L-THP and LDN
We observed that pretreatment of L-THP and LDN signif-icantly attenuated cocaine- or cue-induced reinstatement ofdrug-seeking behavior in a dose-dependent manner This isconsistent with previous findings using L-THP alone (Mantschet al 2007 2010 Figueroa-Guzman et al 2011 Wang andMantsch 2012) suggesting that the combination of L-THP andLDN does not alter therapeutic effects of L-THP In contrastthere is a trend of enhanced therapeutic effects in both cocaineself-administration and reinstatement tests compared withthe effects produced by the same doses of L-THP aloneAdditionally we extensively investigated the effect of a
10-day treatment on drug-seeking behavior during reinstate-ment This dosing schedule was designed to follow a more
Fig 5 Self-administration and drug-seeking behavior (A) Active and inactivelever presses over the course of theexperimental timeline There was nostatistical difference in stable self-administration between the groups (B)Comparison of drug-seeking behavior mea-sured by active lever presses After 13 self-administrations sessions animals weremoved to forced abstinence in which theyreceived treatment in their home cages for10 days On day 23 they were placed backinto the operant chambers with all cuesand rugs removed values are expressed asmean 6 SEM n = 5 for all groups Therewas a significant difference in active leverresponses on day 23 P = 00371 Stu-dentrsquos t test determined a significantdifference between the cocaine-vehiclegroup and the cocaine- 3 mgkg L-THP and01 mgkg LDN group P = 00073 Therewas also less significant difference be-tween cocaine-vehicle group and cocaine3 mgkg L-THP group P = 00464
Fig 6 Mean 6 SEM concentration ofplasma b-endorphin during forced absti-nence Plasma was obtained 30 minutesdirectly following last experimental sessionon day 23 There was a significant differencein b-endorphin concentration between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus vehiclegroup (P 005) as well as between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus 3 mgkgL-THP group P 005 Samples wereanalyzed in duplicates n = 5 for all groups
254 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

daily increased heroin abstinence over 3months by 63 (Yanget al 2008) However L-THP also produces significantsedative effects in a dose-dependent manner (Xi et al 2007Xu et al 2013) whichmay limit its utility in treatment of drugaddictionNaltrexone primarily a m-opioid receptor antagonist is
FDA-approved for treatment of alcoholism and opioid detox-ification At therapeutic doses naltrexone reduces drug-seeking behavior in rodents but is unsuccessful or ineffectivein most human clinical studies owing to significant un-wanted side effects poor patient compliance and highattrition rates (Schmitz et al 2001 Grassi et al 2007Schmitz et al 2009 Schmitz et al 2014) However thetolerability of naltrexone is significantly improved whengiven at subtherapeutic doses (Zagon et al 2014) Low dosenaltrexone (LDN) defined as 4 mg per day in humans(Brown and Panksepp 2009 Mannelli et al 2010Wee et al2012 Cordery et al 2014) is becoming commonly used as along term treatment of autoimmune disorders (Rahn et al2011 Segal et al 2014) Although the exact mechanismsunderlying these actions are unknown one possibility is thatLDN may functionally upregulate endogenous opioid systemthrough m-opioid receptor antagonismWe therefore propose here that the combination of L-THP
and LDN may be safer than L-THP or LDN alone in pre-vention of cocaine relapse This is on the basis of thefollowing two assumptions First such a combination mayproduce additive therapeutics effects which may signifi-cantly lower each drug dose and therefore attenuate un-wanted side-effects produced by each individual drugs andsecond coadministration of LDN with L-THP may alsoantagonize L-THP-induced sedation by normalization orupregulation of the endogenous opioid system in cocaine-dependent subjectsTo test this hypothesis the present study was designed to
determine whether the combination of L-THP and LDN morepotently reduces relapse to drug-seeking behavior and withless unwanted sedative effect than L-THP alone in rat modelsIn addition we also investigated whether coadministration ofL-THP and LDN alters plasma b-endorphin concentrationsand hypothalamic POMC expression
Materials and MethodsAnimals Experimentally naiumlve male Wistar rats (Charles River
Laboratories Raleigh NC) weighing 250ndash300g were used for allexperiments Animals were housed individually in a reverse lightdark cycle with lights off at 700 and lights on at 1900 Animals were
given ad libitumaccess to food andwater All experimental procedureswere conducted in accordance with the Guide for the Care and Use ofLaboratory Animals of the US National Academy of Sciences andwere approved by the IACUC of University of Maryland and theNational Institutes of Health National Institute of Drug Abuse
Jugular Vein Catheterization The cocaine self-administrationprocedures surgeries and training schedules were the same aspreviously reported (Xi et al 2007) Animals were prepared for self-administration experiments by surgical catheterization of rightexternal jugular vein Venous catheters which were constructed ofMicro-Renathane inner diameter 0012 inches outer diameter 0025inches (Braintree Scientific Braintree MA) was inserted 12 cm intothe right jugular under sodium pentobarbital anesthesia (65 mgkgip) with standard aseptic surgical procedures Catheters exited thejugular vein and passed subcutaneously to the top of the skull wherethey were connected to a modified 24-gauge cannula (Plastics OneRoanoke VA) and mounted to the skull with jewelerrsquos screws anddental acrylic During experimental procedures catheters wereconnected to an infusion pump via tubing encased in a protectivemetal spring from the headmount to the top of the chamber Catheterswere flushed directly prior to and directly following training sessionswith a combination of heparingentamicin (5 IU 1 10 mgml) in 09saline solution to prevent blockage and infections
Apparatus Intravenous self-administration experiments wereconducted in operant test-response chambers 32 25 33cm (MedAssociates Inc St Albans VT) House lights were turned on at thebeginning of each session Every test chamber contained two testlevers one active and one inactive Depression of the active leveractivated infusion pump depression of inactive lever was counted buthad no consequence The house light was turned on at the start of each3-hour test session To aid acquisition and maintenance of drug self-administration behavior each drug infusion was always paired with aconditioned cue-light and a cue-sound (tone) The light and thespeaker were installed 12 cm above the active lever The light andtone were paired (on) during the 46-second drug infusion durationThere was no time-out after the fixed-ratio (FR) 1 or 2 drug infusionAll the animals were first trained on FR1 for approximately seventraining sessions with 1 mgkg per infusion of cocaine (to facilitate theacquisition of self-administered cocaine) and then switched to FR2 for05 mgkg per infusion of cocaine self-administration in the followingexperiments Self-administration tests were completed during FR2conditions Scheduling of experimental events and data collectionwere accomplished using Med Associates software
Cocaine Self-Administration Training Animals were allowedto recover from surgical catheterization After recovery they wereplaced into operant chamber and allowed to lever press for intrave-nous cocaine (1 mgkg per infusion) delivered as 008 ml over anintertrial interval of 46 seconds on an FR1 reinforcement scheduleDuring the intertrial interval additional lever presses were recordedbut did not result in additional infusions After achieving stablecocaine self-administration animals were transitioned to 05 mgkgper infusion on a FR1 schedule and transitioned to the final schedule
TABLE 1l-THP and LDN Treatments During Extinction
Self-Administration Extinction Treatment (Daily) Reinstatement Treatment
Cocaine 05 mgkgper inf FR2
Vehicle (n = 7) Cocaine 10 mgkg ipNone (n = 5) 3 mgkg L-THP and 01 mgkg LDN 30 minutes prior
Cocaine 10 mgkg ipNone (n = 9) 5 mgkg L-THP and 01mgkg LDN
30 minutes priorCocaine 10 mgkg ip
01 mgkg LDN (n = 6) Cocaine 10 mgkg ip3 mgkg L-THP (n = 9) Cocaine 10 mgkg ip5 mgkg L-THP (n = 10) Cocaine 10 mgkg ip3 mgkg L-THP and 01mgkg LDN (n = 10) Cocaine 10 mgkg ip5 mgkg L-THP and 01mgkg LDN (n = 11) Cocaine 10 mgkg ip
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 249
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
of 05 mgkg per infusion on an FR2 schedule Each test session lasted3 hours After stable self-administration pretreatments of 3 mgkg5mgkg or 10mgkg L-THP 01 mgkg LDN and 3mgkg1 01 mgkg5 mgkg 1 01 mgkg or 10 mgkg 1 01 mgkg L-THP 1 LDN wereadministered intraperitoneally 30 minutes prior to the experimentalsession
Cocaine Reinstatement Following stable self-administrationexperiments seven consecutive training session in which animalsdemonstrated less that 10 change in drug-taking behavior animalswere transitioned to extinction in which depression of the previouslyactive lever resulted in no infusion cue or light signal Infusionsyringes were filled with physiologic saline Animals were randomlyassigned to one of the treatment groups described in Table 1Immediately following each test session animals were weighed andgiven an intraperitoneal treatment injection corresponding to theirassigned group Extinction continued until animals responded with anumber of active lever presses no more than 10 of that achievedduring cocaine self-administration over the 3-hour period Twenty-four hours after extinction was achieved animals were reinstatedusing ip 10 mgkg cocaine (Amen et al 2011) During cocainereinstatement light and environmental cues remained off
Spontaneous Locomotor Activity Locomotor experiments wereconducted in VersaMax open-field test chambers 40 cm 40 cm(Omnitech Electronics Inc Columbus OH) Habituation and baselinewere established over 3 days Animals were weighed prior to eachsession and randomly assigned to treatment groups described inTable 2 During the drug treatment phase animals were placed intoopen-field chambers for 1 hour After the first hour animals wereremoved injected (intraperitoneally) with the assigned treatment andplacedback into the chamber for a secondhour Treatment continued for10 days Ten days was chosen as the typical extinction treatment periodin the reinstatement experiment Locomotion score was calculated bysubtracting baseline average from total movements after injections
Cocaine Forced Abstinence After self-administering cocaine orsucrose for 13 sessions animals were moved to forced abstinenceForced abstinence differed from extinction in that animals remainedin their home cages were handled administered injections every24 hours and did not return to the operant chambers until day 23Groups are described in Table 3 On day 23 animals were placed backinto operant chambers to measure drug craving as determined byactive lever presses Animals were sacrificed 30 minutes followingfinal session and tissue and plasma were harvested
Tissue Sample Collection Thirty-minutes after the final train-ing session animals were rapidly sacrificed via decapitation Brainswere quickly removed and immersed into dry ice-chilled methyl-butane for 10 seconds and stored in Tissue-TekOCTmedium (SakuraTorrance CA) at ndash80degC Blood was collected from the trunk post-decapitation and stored in Lavender Vacutainer Tubes containingEDTA (Becton Dickinson Franklin Lakes NJ) Blood was transferredto centrifuge tubes containing aprotinin Blood samples were centri-fuged at 1600g for 15minutes at 4degC and the plasmawas removed andstored at ndash80degC
Quantification of b-Endorphin Content b-Endorphin wasquantified using a commercially available EIA kit (Phoenix Pharma-ceuticals Hayward CA) Plasma was acidified using equal amounts ofbuffer A and centrifuged at 10000g for 20 minutes at 4degC A SEP-column was equilibrated using washing buffer B followed by washing
buffer A Plasma was loaded onto the column and washed three timeswith buffer A Peptides were eluted using buffer B and the eluent wascollected into a polystyrene tube evaporated and stored at ndash20degCPeptides were reconstituted with 125 ml of assay buffer Reconstitutedsamples were further diluted to 150 with assay buffer Assay wasperformed according to manufacturerrsquos specifications Absorbancewas read at 450 nm
Visualization and Quantification of POMC by RNAscopeBrains were rapidly harvested and snap-frozen 30 minutes after thecompletion of the above behavioral test on day 23 (see Cocaine ForcedAbstinence) Brains were sliced 014-mm thick and mounted to slidesThere were five animals in each of the treatment groups and six sliceswere analyzed from each animal for a total of 30 analyzed slices pergroup OCT-embedded brainsweremounted on cryostat platformwithOCT medium Tissue was sectioned at the arcuate nucleus (Arc)beginning at approximately and ndash17 mm and ndash396 mm bregma asdetermined from the The Rat Brain in Stereotaxic Coordinates(Paxinos and Watson 2006) and 014-mm tissue slices were mountedon Superfrost Plus Gold Slides (FisherScientificThermoFisherScientific Sunnyvale CA)
Tissue sections were fixed in 10 formalin for 15 minutes at 4degCSlides were dehydrated in ethanol baths of 50 70 and 100 atroom temperature Tissues were then incubated in Pretreatment 4from the RNAscope Pretreatment Kit (Advanced Cell DiagnosticsHayward CA) for 15 minutes at room temperature Slides were driedand the probe for POMC was added for a 2-hour incubation in ahybridization oven The hybridization oven was maintained at 40degCfor all incubations Probe hybridization was carried out in thehybridization oven in four amplification steps in alternating 30- and15-minute incubations After each amplification step slides werewashed two times in 1wash buffer for 2 minutes DAPI was added toslides for 10 seconds and cover-slipped with Fluoro-Gel (ElectronMicroscopy Services Hatfield PA)
Wide-field fluorescent images of the arcuate nucleus were cap-tured using Nikon Eclipse Ti-E with an 20 objective Images weredeconvoluted using NIS Elements Software (Nikon InstrumentsMelville NY ) and pseudo-colored (POMC red Ubiquitin (Ubc) greenand DAPI blue) Image Processing and Analysis by Java (ImageJNIH) was used to quantify total mRNA signal
Drugs The drugs used in the present study were as followsCocaine hydrochloride obtained from NIDA was dissolved in phys-iologic saline Levo-tetrahydropalmatine was obtained from thelaboratory of Stephen Hoag at the University of Maryland BaltimoreThe L-THP was dissolved in 01 M H2SO4 brought to a pH of 556 02with 01 M NaOH and brought to correct volume with sterile waterL-THP concentrations of 3 mgkg 5 mgkg and 10 mgkg were used inthe study For L-THP and LDN combinations naltrexone hydrochlo-ride (Sigma-AldrichSigmaMillipore St Louis MO) was dissolved inthe L-THP solution to a concentration of 01 mgkg The 01 M H2SO401MNaOHsterilewater solution pH of 556 02 was used as vehicleand for the 01 mgkg LDN dose Additionally each solution wassyringe-passed through 022-mm filters to ensure the solutions werefree of contaminates
Data Analysis Statistical analyses were performed usingGraphPad Prism 500 (La Jolla CA) for Windows Data are presentedas mean 6 SEM Studentrsquos t test One-way and two-way analyses ofvariance (ANOVAs) followed by Bonferroni analysis were used forcomparisons between groups where appropriate Effect size wascalculated using h2 for ANOVAs and Pearsonrsquos r for correlations andacceptable level of statistical significance for all tests was P 005
TABLE 2l-THP and LDN Treatment on Locomotion
Habituation (Days 1ndash3) Treatment (Days 4ndash13)
No Treatment Vehicle01 mgkg LDN3 mgkg L-THP5 mgkg L-THP3 mgkg L-THP and 01mgkg LDN5 mgkg L-THP and 01mgkg LDN
TABLE 3l-THP and LDN Treatment on Cue Induced Cocaine-Seeking
Self-Administration Forced Abstinence Treatment (Daily)
Cocaine 05 mgkg per inf Vehicle3 mgkg L-THP3 mgkg L-THP and 01 mgkg LDN
250 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
ResultsEffects of L-THP and LDN on Cocaine Self-
Administration In our study setting animals acquiredstable cocaine self-administration over 14 experimental ses-sions (1119 6 3975 lever presses per session) Pretreat-ment with L-THP produced a biphasic drug-taking effect Thelow doses of L-THP produced an initial increase in self-administration whereas the high dose of L-THP produced areduction in cocaine self-administration (Fig 1) One-wayANOVA revealed a significant drug treatment main effect[F(759)5 1156 P 0001] Effect-size calculations suggest amoderate to highly significant practical effect [h2 5 058]Post-hoc Bonferroni analyses revealed a significant reduction
of cocaine self-administration after 10 mgkg L-THP and10 mgkg L-THP and 01 mgkg LDN combination (P 005)Animals administered LDN exhibited no change in self-administration (P 005) Treatment with the combinationL-THP and LDN had no significant effect as with the respec-tive doses of L-THP alone (P 005)Effects of L-THP and LDN on Cocaine-Induced Rein-
statement of Drug-Seeking Behavior After stable self-administration was achieved animals went to extinction untildrug-seeking behaviorwas extinguished defined as active leverresponses less than 10 of the responses achieved over a stablecocaine self-administration experimental sessions (650 6 142lever presses per session) Pretreatment with L-THP andorLDN administered 30 minutes prior to reinstatement testsignificantly attenuated cocaine-induced (10 mgkg ip) rein-statement of drug-seeking behavior [F(263) 5 354 P 005two-wayANOVA] (Fig 2) Effect size however was determinedto be low to moderate [h2 5 038] Bonferroni post-hoc analysesrevealed a significant reduction in drug-seeking after 3 mgkgL-THPand 01mgkgLDN (P 005) and the 5mgkg L-THPand01 mgkg (P 001) compared with the vehicle control groupEffects of Chronic L-THP and LDN Pretreatment on
Cocaine-Induced Reinstatement of Drug-SeekingBehavior Animals in this experiment were exposed to thesame self-administration conditions used in the previousexperiment however during extinction training the animalswere given L-THP andor LDN daily for 10 consecutive daysduring extinction At 24 hours after the last drug injectioncocaine-induced reinstatement of drug-seeking behavior wassignificantly lower in all drug-treatment groups than in thevehicle control group of rats (Fig 3) One-way ANOVA for thesummarized data in Fig 3C revealed a significant treatmentmain effect [F(551)5 9085P 0001] Effect-size calculationsuggests a moderate applied effect [h2 5 049] Bonferronipost-hoc analyses demonstrated a significant reduction incocaine-seeking after 01 mgkg LDN (P 005) 3 mgkgL-THP (P 001) 3 mgkg L-THP and 01 mgkg LDN (P 0001) 5 mgkg L-THP (P 0001) and 5 mgkg L-THP and01 mgkg LDN (P 0001) administration
Fig 1 Effect of pretreatment of L-THP and LDN on cocaine self-administration After animals achieved stable cocaine self-administrationanimals were administered L-THP LDN or L-THP and LDN 30 minutesprior to experimental session Intraperitoneal administration of L-THPproduced a biphasic effect Low doses (3 mgkg and 5 mgkg L-THP n = 7)increased and a high dose (10 mgkg n = 7 and 10 mgkg L-THP and01mgkg LDN n = 7) induced sedation inhibiting cocaine self-administrationP 005 compared with vehicle treatment determined by one-wayANOVA LDN neither altered cocaine self-administration by itself (01 mgkgLDN n = 7) nor altered L-THPrsquos effects on cocaine self-administrationbehavior (3 mgkg L-THP and 01 mgkg LDN n = 7 and 5mgkg L-THP and01 mgkg LDN n = 7) Values are expressed as mean 6 SEM
Fig 2 Effect of single pretreatment ofL-THP and LDN on cocaine reinstatementIn the experimental session followingextinction animals were administeredintraperitoneal injections of L-THP andLDN 30 minutes prior to a priming doseof 10 mgkg cocaine Two-way ANOVAshowed that 3 mgkg L-THP and 01 mgkgLDN (n = 5) and 5 mgkg L-THP and01 mgkg LDN (n = 9) dose-dependentlyand significantly inhibited reinstatementof cocaine-seeking behavior assessed byactive lever presses P 05 P 001respectively compared with vehicle group(n = 7) Values expressed asmean6 SEM
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 251
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Locomotion Animalswere administered L-THP LDN or L-THP1 LDN daily for 10consecutive days The timeline of 10 days was selected on thebasis of the findings in the above reinstatement experiments(see previous section) The baseline was created using theaverage of the spontaneous locomotion measurements takenover the first 3 consecutive days (3381 6 1681 cm averagebaseline) Baseline was subtracted from total movementsfor each day A dose of 3 mgkg or 5 mgkg L-THP alonesignificantly inhibited spontaneous locomotor activities overthe 60 minutes (two-way ANOVA [F(2140) 5 2234 P 0001]) whereas 01 mgkg LDN alone had no effect onspontaneous locomotion (Fig 4A) Both combination doses ofL-THP and LDN did not produce a significant reduction inspontaneous locomotion [F(2195) 5 02794 P 5 02794 two-way ANOVA] (Fig 4B) Effect size was determined to bemoderate to highly significant [h2 5 051]Effects of L-THP and LDN on Cue-Induced Cocaine-
Seeking After stable cocaine self-administration wasachieved animals underwent forced abstinence in their home
cages in the animal facility for 9 days during which 3 mgkgL-THP 3 mgkg L-THP and 01 mgkg LDN or vehicle wasgiven daily to three separate groups of rats (Fig 5A) Animalswere then placed back into the same self-administrationchambers where they obtained cocaine self-administrationbefore on day 23 Re-exposure to cocaine-associated contex-tual cues induced a significant cocaine-seeking behavior(ie active lever presses) in the absence of cocaine orcocaine-associated cues (Fig 5B the vehicle control group)Pretreatment with 3 mgkg L-THP or 3 mgkg L-THP and01 mgkg LDN produced a significant reduction in cue-induced drug-seeking compared with the vehicle controlgroup and the combination group was more effective thenL-THP alone One-way ANOVA for the data shown in Fig 5Brevealed a significant treatmentmain effect [F(319)5 3413P 5 0043] Moreover effect-size calculations revealed amoderate practical effect [h2 5 040] Studentrsquos t testsrevealed a significant reduction in cocaine-seeking after3 mgkg L-THP [t(8) 5 2354 P 5 00464] or 3 mgkg L-THPand 01 mgkg LDN [t(8) 5 3581 P 5 00072]
Fig 3 Effect of 10 days administration of L-THP and LDN on cocaine reinstatement Animals received intraperitoneal injections for 10 days untilextinction was reached In the experimental session following animals were primed with 10 mgkg cocaine and placed back into operant chambers(A and B) Extinction timeline arrows representing administration of L-THP (3 mgkg n = 9 and 5 mgkg n = 10) LDN (01 mgkg n = 6) or L-THP andLDN combinations (3mgkg and 01 mgkg n = 10 and 5mgkg and 01 mgkg n = 11) (C) Significantly inhibited reinstatement of drug-seeking behaviorassessed by active lever responses expressed as mean 6 SEM Bonferroni post-hoc analysis determined that both the combinations of 3 mgkg L-THPand 01 mgkg LDN and 5mgkg L-THP and 01 mgkg LDNwere more efficacious than any of the single doses P 05 P 01 P 0001 P00001 compared with vehicle group (n = 7)
252 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Plasma Concentrationsof b-Endorphin in Rats after Forced Cocaine AbstinenceAfter the completion of the above behavioral test plasmasamples were collected 30 minutes via the implanted in-travenous catheter Coadministration of 3 mgkg L-THP and01 mgkg LDN significantly elevated plasma b-endorphinlevel compared with the vehicle control group A dose of3 mgkg L-THP alone had no effect (Fig 6) One-way ANOVAfor the data shown in Fig 6 revealed a significant treatmentmain effect [F(339) 5 392 P 5 0016] However the effect-size calculations demonstrate a moderate to low significanteffect [h2 5 024]Effects of L-THP and LDN on POMC mRNA Expres-
sion in Rats after Forced Cocaine Abstinence Figure 7Ashows the RNAscope in situ hybridization data illustratingthat coadministration of 3 mgkg L-THP and 01 mgkg LDNsignificantly elevated POMC expression in the hypothala-mus whereas 3 mgkg L-THP alone had no effect One-wayANOVA for the summarized data shown in Fig 7B revealed asignificant treatment main effect [F(3119) 5 6529 P 500004] Effect-size calculations suggest only a minimalpractical effect [h2 5 014] Bonferroni post-hoc analysesindicated a significant increase in POMC expression after3 mgkg L-THP and 01 mgkg LDN (P 005) but not after3 mgkg L-THP alone (P 005) compared with the vehiclegroupCorrelation Between Cocaine-Seeking Behavior
b-Endorphin and POMC Expression Figure 8A showsthe strong significant negative correlation between activelever presses during reinstatement and plasma concentrationsof b-endorphin [r(15)5 ndash0617 P5 00142] Also shown in Fig8B is the weak positive correlation between POMC expressionand plasma concentrations of b-endorphin [r(15) 5 0361
P 5 0187] A correlation was not observed between POMCexpression and active lever presses during reinstatementshown in Fig 8C [r(15) 5 ndash0214 P 5 0440]
DiscussionCombination medications are becoming increasingly com-
monplace in clinical practice and can be especially effectivewhen used to treat individuals suffering from mental disor-ders and who are multisymptomatic (Tohen et al 2003Kupfer et al 2012 Biler et al 2014 Shorter et al 2015) Inthe case of relapse prevention drug cravings may beeffectively managed when medications treat both behavioraland physiologic symptoms However the majority of phar-maceuticals developed for the treatment of substance abusedisorders target one pathway The combination of L-THP andLDN targets both dopaminergic signaling and the regulationof endogenous opioids In this report we provide evidencethat the combination of L-THP and LDN is more potent withfewer unwanted side effects than L-THP alone on the basis ofthe following observations 1) L-THP and LDN has demon-strated significantly greater effect in attenuating drug-seeking behavior than L-THP alone 2) L-THP and LDN whenadministered together do not result in a reduction ofspontaneous locomotion 3) L-THP and LDN appears toincrease peripheral concentrations of b-endorphin 4) L-THPand LDN appears to upregulate expression of POMC in thearcuate nucleus
L-THP has been investigated as a potential treatment ofaddiction by several other groups (Mantsch et al 2007 2010Figueroa-Guzman et al 2011 Wang and Mantsch 2012)Presently we determined that L-THP produced a biphasiceffect and this is consistent with the results of previous study
Fig 4 Effect of L-THP and LDN onspontaneous locomotion Animals estab-lished a baseline over 3 consecutive daysfollowing that they were given intraperi-toneal injections of the assigned treat-ments for 10 days Values are expressedas mean of the locomotor score 6 SEMn = 6 for all groups Two-way ANOVAdetermined treatment had a significanteffect on spontaneous locomotion P00001 (A) L-THP (3 mgkg and 5 mgkg)produced a dose-dependent reduction onlocomotion and LDN (01 mgkg) had noeffect on locomotion (B) Effect of co-administration of L-THP and LDN
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 253
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
(Xi et al 2007) The increase in cocaine self-administration isattributed to the partial blockade of postsynaptic DA recep-tors which decrease the rewarding properties of cocaine andlead to a compensatory increase in drug-taking Even as thehigher doses of L-THP drastically reduced cocaine self-administration we also noted a strong sedative effectassessed by a decrease in locomotion suggesting that nonspe-cific sedation or locomotion impairment is the main cause forreduced self-administration in the high L-THP dose group Incontrast to L-THP LDN had no effect on cocaine self-administration There was no difference in cocaine self-administration between animals receiving lower doses ofL-THP or L-THP and LDN
We observed that pretreatment of L-THP and LDN signif-icantly attenuated cocaine- or cue-induced reinstatement ofdrug-seeking behavior in a dose-dependent manner This isconsistent with previous findings using L-THP alone (Mantschet al 2007 2010 Figueroa-Guzman et al 2011 Wang andMantsch 2012) suggesting that the combination of L-THP andLDN does not alter therapeutic effects of L-THP In contrastthere is a trend of enhanced therapeutic effects in both cocaineself-administration and reinstatement tests compared withthe effects produced by the same doses of L-THP aloneAdditionally we extensively investigated the effect of a
10-day treatment on drug-seeking behavior during reinstate-ment This dosing schedule was designed to follow a more
Fig 5 Self-administration and drug-seeking behavior (A) Active and inactivelever presses over the course of theexperimental timeline There was nostatistical difference in stable self-administration between the groups (B)Comparison of drug-seeking behavior mea-sured by active lever presses After 13 self-administrations sessions animals weremoved to forced abstinence in which theyreceived treatment in their home cages for10 days On day 23 they were placed backinto the operant chambers with all cuesand rugs removed values are expressed asmean 6 SEM n = 5 for all groups Therewas a significant difference in active leverresponses on day 23 P = 00371 Stu-dentrsquos t test determined a significantdifference between the cocaine-vehiclegroup and the cocaine- 3 mgkg L-THP and01 mgkg LDN group P = 00073 Therewas also less significant difference be-tween cocaine-vehicle group and cocaine3 mgkg L-THP group P = 00464
Fig 6 Mean 6 SEM concentration ofplasma b-endorphin during forced absti-nence Plasma was obtained 30 minutesdirectly following last experimental sessionon day 23 There was a significant differencein b-endorphin concentration between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus vehiclegroup (P 005) as well as between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus 3 mgkgL-THP group P 005 Samples wereanalyzed in duplicates n = 5 for all groups
254 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

of 05 mgkg per infusion on an FR2 schedule Each test session lasted3 hours After stable self-administration pretreatments of 3 mgkg5mgkg or 10mgkg L-THP 01 mgkg LDN and 3mgkg1 01 mgkg5 mgkg 1 01 mgkg or 10 mgkg 1 01 mgkg L-THP 1 LDN wereadministered intraperitoneally 30 minutes prior to the experimentalsession
Cocaine Reinstatement Following stable self-administrationexperiments seven consecutive training session in which animalsdemonstrated less that 10 change in drug-taking behavior animalswere transitioned to extinction in which depression of the previouslyactive lever resulted in no infusion cue or light signal Infusionsyringes were filled with physiologic saline Animals were randomlyassigned to one of the treatment groups described in Table 1Immediately following each test session animals were weighed andgiven an intraperitoneal treatment injection corresponding to theirassigned group Extinction continued until animals responded with anumber of active lever presses no more than 10 of that achievedduring cocaine self-administration over the 3-hour period Twenty-four hours after extinction was achieved animals were reinstatedusing ip 10 mgkg cocaine (Amen et al 2011) During cocainereinstatement light and environmental cues remained off
Spontaneous Locomotor Activity Locomotor experiments wereconducted in VersaMax open-field test chambers 40 cm 40 cm(Omnitech Electronics Inc Columbus OH) Habituation and baselinewere established over 3 days Animals were weighed prior to eachsession and randomly assigned to treatment groups described inTable 2 During the drug treatment phase animals were placed intoopen-field chambers for 1 hour After the first hour animals wereremoved injected (intraperitoneally) with the assigned treatment andplacedback into the chamber for a secondhour Treatment continued for10 days Ten days was chosen as the typical extinction treatment periodin the reinstatement experiment Locomotion score was calculated bysubtracting baseline average from total movements after injections
Cocaine Forced Abstinence After self-administering cocaine orsucrose for 13 sessions animals were moved to forced abstinenceForced abstinence differed from extinction in that animals remainedin their home cages were handled administered injections every24 hours and did not return to the operant chambers until day 23Groups are described in Table 3 On day 23 animals were placed backinto operant chambers to measure drug craving as determined byactive lever presses Animals were sacrificed 30 minutes followingfinal session and tissue and plasma were harvested
Tissue Sample Collection Thirty-minutes after the final train-ing session animals were rapidly sacrificed via decapitation Brainswere quickly removed and immersed into dry ice-chilled methyl-butane for 10 seconds and stored in Tissue-TekOCTmedium (SakuraTorrance CA) at ndash80degC Blood was collected from the trunk post-decapitation and stored in Lavender Vacutainer Tubes containingEDTA (Becton Dickinson Franklin Lakes NJ) Blood was transferredto centrifuge tubes containing aprotinin Blood samples were centri-fuged at 1600g for 15minutes at 4degC and the plasmawas removed andstored at ndash80degC
Quantification of b-Endorphin Content b-Endorphin wasquantified using a commercially available EIA kit (Phoenix Pharma-ceuticals Hayward CA) Plasma was acidified using equal amounts ofbuffer A and centrifuged at 10000g for 20 minutes at 4degC A SEP-column was equilibrated using washing buffer B followed by washing
buffer A Plasma was loaded onto the column and washed three timeswith buffer A Peptides were eluted using buffer B and the eluent wascollected into a polystyrene tube evaporated and stored at ndash20degCPeptides were reconstituted with 125 ml of assay buffer Reconstitutedsamples were further diluted to 150 with assay buffer Assay wasperformed according to manufacturerrsquos specifications Absorbancewas read at 450 nm
Visualization and Quantification of POMC by RNAscopeBrains were rapidly harvested and snap-frozen 30 minutes after thecompletion of the above behavioral test on day 23 (see Cocaine ForcedAbstinence) Brains were sliced 014-mm thick and mounted to slidesThere were five animals in each of the treatment groups and six sliceswere analyzed from each animal for a total of 30 analyzed slices pergroup OCT-embedded brainsweremounted on cryostat platformwithOCT medium Tissue was sectioned at the arcuate nucleus (Arc)beginning at approximately and ndash17 mm and ndash396 mm bregma asdetermined from the The Rat Brain in Stereotaxic Coordinates(Paxinos and Watson 2006) and 014-mm tissue slices were mountedon Superfrost Plus Gold Slides (FisherScientificThermoFisherScientific Sunnyvale CA)
Tissue sections were fixed in 10 formalin for 15 minutes at 4degCSlides were dehydrated in ethanol baths of 50 70 and 100 atroom temperature Tissues were then incubated in Pretreatment 4from the RNAscope Pretreatment Kit (Advanced Cell DiagnosticsHayward CA) for 15 minutes at room temperature Slides were driedand the probe for POMC was added for a 2-hour incubation in ahybridization oven The hybridization oven was maintained at 40degCfor all incubations Probe hybridization was carried out in thehybridization oven in four amplification steps in alternating 30- and15-minute incubations After each amplification step slides werewashed two times in 1wash buffer for 2 minutes DAPI was added toslides for 10 seconds and cover-slipped with Fluoro-Gel (ElectronMicroscopy Services Hatfield PA)
Wide-field fluorescent images of the arcuate nucleus were cap-tured using Nikon Eclipse Ti-E with an 20 objective Images weredeconvoluted using NIS Elements Software (Nikon InstrumentsMelville NY ) and pseudo-colored (POMC red Ubiquitin (Ubc) greenand DAPI blue) Image Processing and Analysis by Java (ImageJNIH) was used to quantify total mRNA signal
Drugs The drugs used in the present study were as followsCocaine hydrochloride obtained from NIDA was dissolved in phys-iologic saline Levo-tetrahydropalmatine was obtained from thelaboratory of Stephen Hoag at the University of Maryland BaltimoreThe L-THP was dissolved in 01 M H2SO4 brought to a pH of 556 02with 01 M NaOH and brought to correct volume with sterile waterL-THP concentrations of 3 mgkg 5 mgkg and 10 mgkg were used inthe study For L-THP and LDN combinations naltrexone hydrochlo-ride (Sigma-AldrichSigmaMillipore St Louis MO) was dissolved inthe L-THP solution to a concentration of 01 mgkg The 01 M H2SO401MNaOHsterilewater solution pH of 556 02 was used as vehicleand for the 01 mgkg LDN dose Additionally each solution wassyringe-passed through 022-mm filters to ensure the solutions werefree of contaminates
Data Analysis Statistical analyses were performed usingGraphPad Prism 500 (La Jolla CA) for Windows Data are presentedas mean 6 SEM Studentrsquos t test One-way and two-way analyses ofvariance (ANOVAs) followed by Bonferroni analysis were used forcomparisons between groups where appropriate Effect size wascalculated using h2 for ANOVAs and Pearsonrsquos r for correlations andacceptable level of statistical significance for all tests was P 005
TABLE 2l-THP and LDN Treatment on Locomotion
Habituation (Days 1ndash3) Treatment (Days 4ndash13)
No Treatment Vehicle01 mgkg LDN3 mgkg L-THP5 mgkg L-THP3 mgkg L-THP and 01mgkg LDN5 mgkg L-THP and 01mgkg LDN
TABLE 3l-THP and LDN Treatment on Cue Induced Cocaine-Seeking
Self-Administration Forced Abstinence Treatment (Daily)
Cocaine 05 mgkg per inf Vehicle3 mgkg L-THP3 mgkg L-THP and 01 mgkg LDN
250 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
ResultsEffects of L-THP and LDN on Cocaine Self-
Administration In our study setting animals acquiredstable cocaine self-administration over 14 experimental ses-sions (1119 6 3975 lever presses per session) Pretreat-ment with L-THP produced a biphasic drug-taking effect Thelow doses of L-THP produced an initial increase in self-administration whereas the high dose of L-THP produced areduction in cocaine self-administration (Fig 1) One-wayANOVA revealed a significant drug treatment main effect[F(759)5 1156 P 0001] Effect-size calculations suggest amoderate to highly significant practical effect [h2 5 058]Post-hoc Bonferroni analyses revealed a significant reduction
of cocaine self-administration after 10 mgkg L-THP and10 mgkg L-THP and 01 mgkg LDN combination (P 005)Animals administered LDN exhibited no change in self-administration (P 005) Treatment with the combinationL-THP and LDN had no significant effect as with the respec-tive doses of L-THP alone (P 005)Effects of L-THP and LDN on Cocaine-Induced Rein-
statement of Drug-Seeking Behavior After stable self-administration was achieved animals went to extinction untildrug-seeking behaviorwas extinguished defined as active leverresponses less than 10 of the responses achieved over a stablecocaine self-administration experimental sessions (650 6 142lever presses per session) Pretreatment with L-THP andorLDN administered 30 minutes prior to reinstatement testsignificantly attenuated cocaine-induced (10 mgkg ip) rein-statement of drug-seeking behavior [F(263) 5 354 P 005two-wayANOVA] (Fig 2) Effect size however was determinedto be low to moderate [h2 5 038] Bonferroni post-hoc analysesrevealed a significant reduction in drug-seeking after 3 mgkgL-THPand 01mgkgLDN (P 005) and the 5mgkg L-THPand01 mgkg (P 001) compared with the vehicle control groupEffects of Chronic L-THP and LDN Pretreatment on
Cocaine-Induced Reinstatement of Drug-SeekingBehavior Animals in this experiment were exposed to thesame self-administration conditions used in the previousexperiment however during extinction training the animalswere given L-THP andor LDN daily for 10 consecutive daysduring extinction At 24 hours after the last drug injectioncocaine-induced reinstatement of drug-seeking behavior wassignificantly lower in all drug-treatment groups than in thevehicle control group of rats (Fig 3) One-way ANOVA for thesummarized data in Fig 3C revealed a significant treatmentmain effect [F(551)5 9085P 0001] Effect-size calculationsuggests a moderate applied effect [h2 5 049] Bonferronipost-hoc analyses demonstrated a significant reduction incocaine-seeking after 01 mgkg LDN (P 005) 3 mgkgL-THP (P 001) 3 mgkg L-THP and 01 mgkg LDN (P 0001) 5 mgkg L-THP (P 0001) and 5 mgkg L-THP and01 mgkg LDN (P 0001) administration
Fig 1 Effect of pretreatment of L-THP and LDN on cocaine self-administration After animals achieved stable cocaine self-administrationanimals were administered L-THP LDN or L-THP and LDN 30 minutesprior to experimental session Intraperitoneal administration of L-THPproduced a biphasic effect Low doses (3 mgkg and 5 mgkg L-THP n = 7)increased and a high dose (10 mgkg n = 7 and 10 mgkg L-THP and01mgkg LDN n = 7) induced sedation inhibiting cocaine self-administrationP 005 compared with vehicle treatment determined by one-wayANOVA LDN neither altered cocaine self-administration by itself (01 mgkgLDN n = 7) nor altered L-THPrsquos effects on cocaine self-administrationbehavior (3 mgkg L-THP and 01 mgkg LDN n = 7 and 5mgkg L-THP and01 mgkg LDN n = 7) Values are expressed as mean 6 SEM
Fig 2 Effect of single pretreatment ofL-THP and LDN on cocaine reinstatementIn the experimental session followingextinction animals were administeredintraperitoneal injections of L-THP andLDN 30 minutes prior to a priming doseof 10 mgkg cocaine Two-way ANOVAshowed that 3 mgkg L-THP and 01 mgkgLDN (n = 5) and 5 mgkg L-THP and01 mgkg LDN (n = 9) dose-dependentlyand significantly inhibited reinstatementof cocaine-seeking behavior assessed byactive lever presses P 05 P 001respectively compared with vehicle group(n = 7) Values expressed asmean6 SEM
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 251
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Locomotion Animalswere administered L-THP LDN or L-THP1 LDN daily for 10consecutive days The timeline of 10 days was selected on thebasis of the findings in the above reinstatement experiments(see previous section) The baseline was created using theaverage of the spontaneous locomotion measurements takenover the first 3 consecutive days (3381 6 1681 cm averagebaseline) Baseline was subtracted from total movementsfor each day A dose of 3 mgkg or 5 mgkg L-THP alonesignificantly inhibited spontaneous locomotor activities overthe 60 minutes (two-way ANOVA [F(2140) 5 2234 P 0001]) whereas 01 mgkg LDN alone had no effect onspontaneous locomotion (Fig 4A) Both combination doses ofL-THP and LDN did not produce a significant reduction inspontaneous locomotion [F(2195) 5 02794 P 5 02794 two-way ANOVA] (Fig 4B) Effect size was determined to bemoderate to highly significant [h2 5 051]Effects of L-THP and LDN on Cue-Induced Cocaine-
Seeking After stable cocaine self-administration wasachieved animals underwent forced abstinence in their home
cages in the animal facility for 9 days during which 3 mgkgL-THP 3 mgkg L-THP and 01 mgkg LDN or vehicle wasgiven daily to three separate groups of rats (Fig 5A) Animalswere then placed back into the same self-administrationchambers where they obtained cocaine self-administrationbefore on day 23 Re-exposure to cocaine-associated contex-tual cues induced a significant cocaine-seeking behavior(ie active lever presses) in the absence of cocaine orcocaine-associated cues (Fig 5B the vehicle control group)Pretreatment with 3 mgkg L-THP or 3 mgkg L-THP and01 mgkg LDN produced a significant reduction in cue-induced drug-seeking compared with the vehicle controlgroup and the combination group was more effective thenL-THP alone One-way ANOVA for the data shown in Fig 5Brevealed a significant treatmentmain effect [F(319)5 3413P 5 0043] Moreover effect-size calculations revealed amoderate practical effect [h2 5 040] Studentrsquos t testsrevealed a significant reduction in cocaine-seeking after3 mgkg L-THP [t(8) 5 2354 P 5 00464] or 3 mgkg L-THPand 01 mgkg LDN [t(8) 5 3581 P 5 00072]
Fig 3 Effect of 10 days administration of L-THP and LDN on cocaine reinstatement Animals received intraperitoneal injections for 10 days untilextinction was reached In the experimental session following animals were primed with 10 mgkg cocaine and placed back into operant chambers(A and B) Extinction timeline arrows representing administration of L-THP (3 mgkg n = 9 and 5 mgkg n = 10) LDN (01 mgkg n = 6) or L-THP andLDN combinations (3mgkg and 01 mgkg n = 10 and 5mgkg and 01 mgkg n = 11) (C) Significantly inhibited reinstatement of drug-seeking behaviorassessed by active lever responses expressed as mean 6 SEM Bonferroni post-hoc analysis determined that both the combinations of 3 mgkg L-THPand 01 mgkg LDN and 5mgkg L-THP and 01 mgkg LDNwere more efficacious than any of the single doses P 05 P 01 P 0001 P00001 compared with vehicle group (n = 7)
252 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Plasma Concentrationsof b-Endorphin in Rats after Forced Cocaine AbstinenceAfter the completion of the above behavioral test plasmasamples were collected 30 minutes via the implanted in-travenous catheter Coadministration of 3 mgkg L-THP and01 mgkg LDN significantly elevated plasma b-endorphinlevel compared with the vehicle control group A dose of3 mgkg L-THP alone had no effect (Fig 6) One-way ANOVAfor the data shown in Fig 6 revealed a significant treatmentmain effect [F(339) 5 392 P 5 0016] However the effect-size calculations demonstrate a moderate to low significanteffect [h2 5 024]Effects of L-THP and LDN on POMC mRNA Expres-
sion in Rats after Forced Cocaine Abstinence Figure 7Ashows the RNAscope in situ hybridization data illustratingthat coadministration of 3 mgkg L-THP and 01 mgkg LDNsignificantly elevated POMC expression in the hypothala-mus whereas 3 mgkg L-THP alone had no effect One-wayANOVA for the summarized data shown in Fig 7B revealed asignificant treatment main effect [F(3119) 5 6529 P 500004] Effect-size calculations suggest only a minimalpractical effect [h2 5 014] Bonferroni post-hoc analysesindicated a significant increase in POMC expression after3 mgkg L-THP and 01 mgkg LDN (P 005) but not after3 mgkg L-THP alone (P 005) compared with the vehiclegroupCorrelation Between Cocaine-Seeking Behavior
b-Endorphin and POMC Expression Figure 8A showsthe strong significant negative correlation between activelever presses during reinstatement and plasma concentrationsof b-endorphin [r(15)5 ndash0617 P5 00142] Also shown in Fig8B is the weak positive correlation between POMC expressionand plasma concentrations of b-endorphin [r(15) 5 0361
P 5 0187] A correlation was not observed between POMCexpression and active lever presses during reinstatementshown in Fig 8C [r(15) 5 ndash0214 P 5 0440]
DiscussionCombination medications are becoming increasingly com-
monplace in clinical practice and can be especially effectivewhen used to treat individuals suffering from mental disor-ders and who are multisymptomatic (Tohen et al 2003Kupfer et al 2012 Biler et al 2014 Shorter et al 2015) Inthe case of relapse prevention drug cravings may beeffectively managed when medications treat both behavioraland physiologic symptoms However the majority of phar-maceuticals developed for the treatment of substance abusedisorders target one pathway The combination of L-THP andLDN targets both dopaminergic signaling and the regulationof endogenous opioids In this report we provide evidencethat the combination of L-THP and LDN is more potent withfewer unwanted side effects than L-THP alone on the basis ofthe following observations 1) L-THP and LDN has demon-strated significantly greater effect in attenuating drug-seeking behavior than L-THP alone 2) L-THP and LDN whenadministered together do not result in a reduction ofspontaneous locomotion 3) L-THP and LDN appears toincrease peripheral concentrations of b-endorphin 4) L-THPand LDN appears to upregulate expression of POMC in thearcuate nucleus
L-THP has been investigated as a potential treatment ofaddiction by several other groups (Mantsch et al 2007 2010Figueroa-Guzman et al 2011 Wang and Mantsch 2012)Presently we determined that L-THP produced a biphasiceffect and this is consistent with the results of previous study
Fig 4 Effect of L-THP and LDN onspontaneous locomotion Animals estab-lished a baseline over 3 consecutive daysfollowing that they were given intraperi-toneal injections of the assigned treat-ments for 10 days Values are expressedas mean of the locomotor score 6 SEMn = 6 for all groups Two-way ANOVAdetermined treatment had a significanteffect on spontaneous locomotion P00001 (A) L-THP (3 mgkg and 5 mgkg)produced a dose-dependent reduction onlocomotion and LDN (01 mgkg) had noeffect on locomotion (B) Effect of co-administration of L-THP and LDN
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 253
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
(Xi et al 2007) The increase in cocaine self-administration isattributed to the partial blockade of postsynaptic DA recep-tors which decrease the rewarding properties of cocaine andlead to a compensatory increase in drug-taking Even as thehigher doses of L-THP drastically reduced cocaine self-administration we also noted a strong sedative effectassessed by a decrease in locomotion suggesting that nonspe-cific sedation or locomotion impairment is the main cause forreduced self-administration in the high L-THP dose group Incontrast to L-THP LDN had no effect on cocaine self-administration There was no difference in cocaine self-administration between animals receiving lower doses ofL-THP or L-THP and LDN
We observed that pretreatment of L-THP and LDN signif-icantly attenuated cocaine- or cue-induced reinstatement ofdrug-seeking behavior in a dose-dependent manner This isconsistent with previous findings using L-THP alone (Mantschet al 2007 2010 Figueroa-Guzman et al 2011 Wang andMantsch 2012) suggesting that the combination of L-THP andLDN does not alter therapeutic effects of L-THP In contrastthere is a trend of enhanced therapeutic effects in both cocaineself-administration and reinstatement tests compared withthe effects produced by the same doses of L-THP aloneAdditionally we extensively investigated the effect of a
10-day treatment on drug-seeking behavior during reinstate-ment This dosing schedule was designed to follow a more
Fig 5 Self-administration and drug-seeking behavior (A) Active and inactivelever presses over the course of theexperimental timeline There was nostatistical difference in stable self-administration between the groups (B)Comparison of drug-seeking behavior mea-sured by active lever presses After 13 self-administrations sessions animals weremoved to forced abstinence in which theyreceived treatment in their home cages for10 days On day 23 they were placed backinto the operant chambers with all cuesand rugs removed values are expressed asmean 6 SEM n = 5 for all groups Therewas a significant difference in active leverresponses on day 23 P = 00371 Stu-dentrsquos t test determined a significantdifference between the cocaine-vehiclegroup and the cocaine- 3 mgkg L-THP and01 mgkg LDN group P = 00073 Therewas also less significant difference be-tween cocaine-vehicle group and cocaine3 mgkg L-THP group P = 00464
Fig 6 Mean 6 SEM concentration ofplasma b-endorphin during forced absti-nence Plasma was obtained 30 minutesdirectly following last experimental sessionon day 23 There was a significant differencein b-endorphin concentration between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus vehiclegroup (P 005) as well as between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus 3 mgkgL-THP group P 005 Samples wereanalyzed in duplicates n = 5 for all groups
254 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
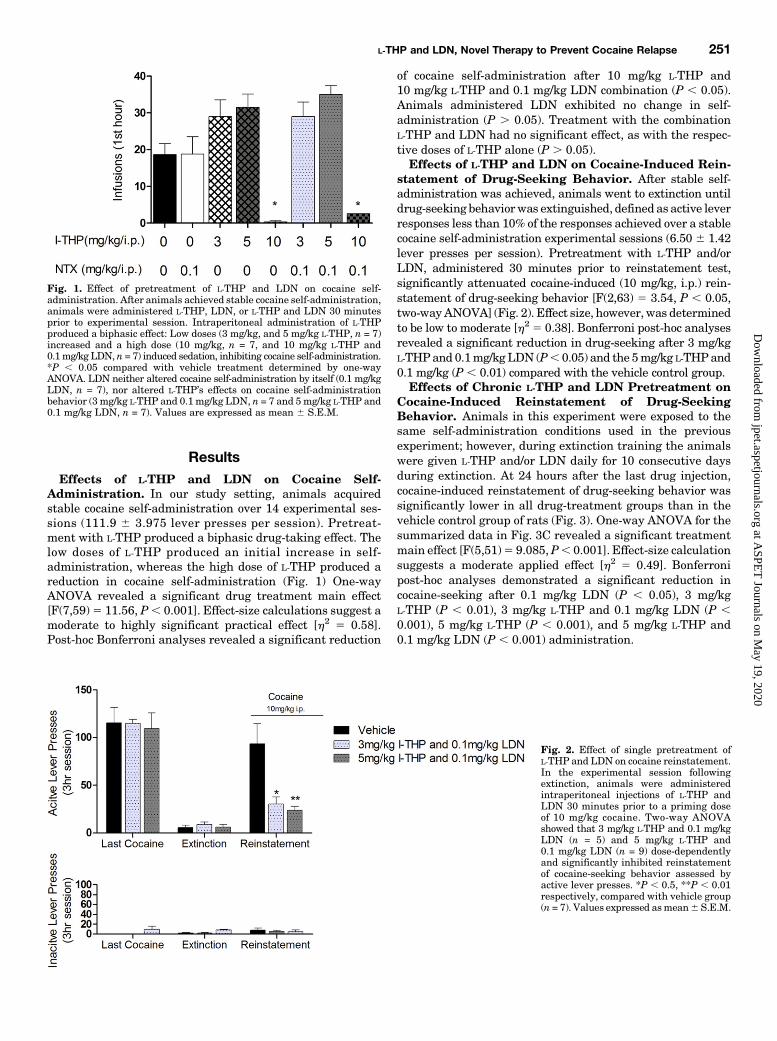
ResultsEffects of L-THP and LDN on Cocaine Self-
Administration In our study setting animals acquiredstable cocaine self-administration over 14 experimental ses-sions (1119 6 3975 lever presses per session) Pretreat-ment with L-THP produced a biphasic drug-taking effect Thelow doses of L-THP produced an initial increase in self-administration whereas the high dose of L-THP produced areduction in cocaine self-administration (Fig 1) One-wayANOVA revealed a significant drug treatment main effect[F(759)5 1156 P 0001] Effect-size calculations suggest amoderate to highly significant practical effect [h2 5 058]Post-hoc Bonferroni analyses revealed a significant reduction
of cocaine self-administration after 10 mgkg L-THP and10 mgkg L-THP and 01 mgkg LDN combination (P 005)Animals administered LDN exhibited no change in self-administration (P 005) Treatment with the combinationL-THP and LDN had no significant effect as with the respec-tive doses of L-THP alone (P 005)Effects of L-THP and LDN on Cocaine-Induced Rein-
statement of Drug-Seeking Behavior After stable self-administration was achieved animals went to extinction untildrug-seeking behaviorwas extinguished defined as active leverresponses less than 10 of the responses achieved over a stablecocaine self-administration experimental sessions (650 6 142lever presses per session) Pretreatment with L-THP andorLDN administered 30 minutes prior to reinstatement testsignificantly attenuated cocaine-induced (10 mgkg ip) rein-statement of drug-seeking behavior [F(263) 5 354 P 005two-wayANOVA] (Fig 2) Effect size however was determinedto be low to moderate [h2 5 038] Bonferroni post-hoc analysesrevealed a significant reduction in drug-seeking after 3 mgkgL-THPand 01mgkgLDN (P 005) and the 5mgkg L-THPand01 mgkg (P 001) compared with the vehicle control groupEffects of Chronic L-THP and LDN Pretreatment on
Cocaine-Induced Reinstatement of Drug-SeekingBehavior Animals in this experiment were exposed to thesame self-administration conditions used in the previousexperiment however during extinction training the animalswere given L-THP andor LDN daily for 10 consecutive daysduring extinction At 24 hours after the last drug injectioncocaine-induced reinstatement of drug-seeking behavior wassignificantly lower in all drug-treatment groups than in thevehicle control group of rats (Fig 3) One-way ANOVA for thesummarized data in Fig 3C revealed a significant treatmentmain effect [F(551)5 9085P 0001] Effect-size calculationsuggests a moderate applied effect [h2 5 049] Bonferronipost-hoc analyses demonstrated a significant reduction incocaine-seeking after 01 mgkg LDN (P 005) 3 mgkgL-THP (P 001) 3 mgkg L-THP and 01 mgkg LDN (P 0001) 5 mgkg L-THP (P 0001) and 5 mgkg L-THP and01 mgkg LDN (P 0001) administration
Fig 1 Effect of pretreatment of L-THP and LDN on cocaine self-administration After animals achieved stable cocaine self-administrationanimals were administered L-THP LDN or L-THP and LDN 30 minutesprior to experimental session Intraperitoneal administration of L-THPproduced a biphasic effect Low doses (3 mgkg and 5 mgkg L-THP n = 7)increased and a high dose (10 mgkg n = 7 and 10 mgkg L-THP and01mgkg LDN n = 7) induced sedation inhibiting cocaine self-administrationP 005 compared with vehicle treatment determined by one-wayANOVA LDN neither altered cocaine self-administration by itself (01 mgkgLDN n = 7) nor altered L-THPrsquos effects on cocaine self-administrationbehavior (3 mgkg L-THP and 01 mgkg LDN n = 7 and 5mgkg L-THP and01 mgkg LDN n = 7) Values are expressed as mean 6 SEM
Fig 2 Effect of single pretreatment ofL-THP and LDN on cocaine reinstatementIn the experimental session followingextinction animals were administeredintraperitoneal injections of L-THP andLDN 30 minutes prior to a priming doseof 10 mgkg cocaine Two-way ANOVAshowed that 3 mgkg L-THP and 01 mgkgLDN (n = 5) and 5 mgkg L-THP and01 mgkg LDN (n = 9) dose-dependentlyand significantly inhibited reinstatementof cocaine-seeking behavior assessed byactive lever presses P 05 P 001respectively compared with vehicle group(n = 7) Values expressed asmean6 SEM
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 251
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Locomotion Animalswere administered L-THP LDN or L-THP1 LDN daily for 10consecutive days The timeline of 10 days was selected on thebasis of the findings in the above reinstatement experiments(see previous section) The baseline was created using theaverage of the spontaneous locomotion measurements takenover the first 3 consecutive days (3381 6 1681 cm averagebaseline) Baseline was subtracted from total movementsfor each day A dose of 3 mgkg or 5 mgkg L-THP alonesignificantly inhibited spontaneous locomotor activities overthe 60 minutes (two-way ANOVA [F(2140) 5 2234 P 0001]) whereas 01 mgkg LDN alone had no effect onspontaneous locomotion (Fig 4A) Both combination doses ofL-THP and LDN did not produce a significant reduction inspontaneous locomotion [F(2195) 5 02794 P 5 02794 two-way ANOVA] (Fig 4B) Effect size was determined to bemoderate to highly significant [h2 5 051]Effects of L-THP and LDN on Cue-Induced Cocaine-
Seeking After stable cocaine self-administration wasachieved animals underwent forced abstinence in their home
cages in the animal facility for 9 days during which 3 mgkgL-THP 3 mgkg L-THP and 01 mgkg LDN or vehicle wasgiven daily to three separate groups of rats (Fig 5A) Animalswere then placed back into the same self-administrationchambers where they obtained cocaine self-administrationbefore on day 23 Re-exposure to cocaine-associated contex-tual cues induced a significant cocaine-seeking behavior(ie active lever presses) in the absence of cocaine orcocaine-associated cues (Fig 5B the vehicle control group)Pretreatment with 3 mgkg L-THP or 3 mgkg L-THP and01 mgkg LDN produced a significant reduction in cue-induced drug-seeking compared with the vehicle controlgroup and the combination group was more effective thenL-THP alone One-way ANOVA for the data shown in Fig 5Brevealed a significant treatmentmain effect [F(319)5 3413P 5 0043] Moreover effect-size calculations revealed amoderate practical effect [h2 5 040] Studentrsquos t testsrevealed a significant reduction in cocaine-seeking after3 mgkg L-THP [t(8) 5 2354 P 5 00464] or 3 mgkg L-THPand 01 mgkg LDN [t(8) 5 3581 P 5 00072]
Fig 3 Effect of 10 days administration of L-THP and LDN on cocaine reinstatement Animals received intraperitoneal injections for 10 days untilextinction was reached In the experimental session following animals were primed with 10 mgkg cocaine and placed back into operant chambers(A and B) Extinction timeline arrows representing administration of L-THP (3 mgkg n = 9 and 5 mgkg n = 10) LDN (01 mgkg n = 6) or L-THP andLDN combinations (3mgkg and 01 mgkg n = 10 and 5mgkg and 01 mgkg n = 11) (C) Significantly inhibited reinstatement of drug-seeking behaviorassessed by active lever responses expressed as mean 6 SEM Bonferroni post-hoc analysis determined that both the combinations of 3 mgkg L-THPand 01 mgkg LDN and 5mgkg L-THP and 01 mgkg LDNwere more efficacious than any of the single doses P 05 P 01 P 0001 P00001 compared with vehicle group (n = 7)
252 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Plasma Concentrationsof b-Endorphin in Rats after Forced Cocaine AbstinenceAfter the completion of the above behavioral test plasmasamples were collected 30 minutes via the implanted in-travenous catheter Coadministration of 3 mgkg L-THP and01 mgkg LDN significantly elevated plasma b-endorphinlevel compared with the vehicle control group A dose of3 mgkg L-THP alone had no effect (Fig 6) One-way ANOVAfor the data shown in Fig 6 revealed a significant treatmentmain effect [F(339) 5 392 P 5 0016] However the effect-size calculations demonstrate a moderate to low significanteffect [h2 5 024]Effects of L-THP and LDN on POMC mRNA Expres-
sion in Rats after Forced Cocaine Abstinence Figure 7Ashows the RNAscope in situ hybridization data illustratingthat coadministration of 3 mgkg L-THP and 01 mgkg LDNsignificantly elevated POMC expression in the hypothala-mus whereas 3 mgkg L-THP alone had no effect One-wayANOVA for the summarized data shown in Fig 7B revealed asignificant treatment main effect [F(3119) 5 6529 P 500004] Effect-size calculations suggest only a minimalpractical effect [h2 5 014] Bonferroni post-hoc analysesindicated a significant increase in POMC expression after3 mgkg L-THP and 01 mgkg LDN (P 005) but not after3 mgkg L-THP alone (P 005) compared with the vehiclegroupCorrelation Between Cocaine-Seeking Behavior
b-Endorphin and POMC Expression Figure 8A showsthe strong significant negative correlation between activelever presses during reinstatement and plasma concentrationsof b-endorphin [r(15)5 ndash0617 P5 00142] Also shown in Fig8B is the weak positive correlation between POMC expressionand plasma concentrations of b-endorphin [r(15) 5 0361
P 5 0187] A correlation was not observed between POMCexpression and active lever presses during reinstatementshown in Fig 8C [r(15) 5 ndash0214 P 5 0440]
DiscussionCombination medications are becoming increasingly com-
monplace in clinical practice and can be especially effectivewhen used to treat individuals suffering from mental disor-ders and who are multisymptomatic (Tohen et al 2003Kupfer et al 2012 Biler et al 2014 Shorter et al 2015) Inthe case of relapse prevention drug cravings may beeffectively managed when medications treat both behavioraland physiologic symptoms However the majority of phar-maceuticals developed for the treatment of substance abusedisorders target one pathway The combination of L-THP andLDN targets both dopaminergic signaling and the regulationof endogenous opioids In this report we provide evidencethat the combination of L-THP and LDN is more potent withfewer unwanted side effects than L-THP alone on the basis ofthe following observations 1) L-THP and LDN has demon-strated significantly greater effect in attenuating drug-seeking behavior than L-THP alone 2) L-THP and LDN whenadministered together do not result in a reduction ofspontaneous locomotion 3) L-THP and LDN appears toincrease peripheral concentrations of b-endorphin 4) L-THPand LDN appears to upregulate expression of POMC in thearcuate nucleus
L-THP has been investigated as a potential treatment ofaddiction by several other groups (Mantsch et al 2007 2010Figueroa-Guzman et al 2011 Wang and Mantsch 2012)Presently we determined that L-THP produced a biphasiceffect and this is consistent with the results of previous study
Fig 4 Effect of L-THP and LDN onspontaneous locomotion Animals estab-lished a baseline over 3 consecutive daysfollowing that they were given intraperi-toneal injections of the assigned treat-ments for 10 days Values are expressedas mean of the locomotor score 6 SEMn = 6 for all groups Two-way ANOVAdetermined treatment had a significanteffect on spontaneous locomotion P00001 (A) L-THP (3 mgkg and 5 mgkg)produced a dose-dependent reduction onlocomotion and LDN (01 mgkg) had noeffect on locomotion (B) Effect of co-administration of L-THP and LDN
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 253
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
(Xi et al 2007) The increase in cocaine self-administration isattributed to the partial blockade of postsynaptic DA recep-tors which decrease the rewarding properties of cocaine andlead to a compensatory increase in drug-taking Even as thehigher doses of L-THP drastically reduced cocaine self-administration we also noted a strong sedative effectassessed by a decrease in locomotion suggesting that nonspe-cific sedation or locomotion impairment is the main cause forreduced self-administration in the high L-THP dose group Incontrast to L-THP LDN had no effect on cocaine self-administration There was no difference in cocaine self-administration between animals receiving lower doses ofL-THP or L-THP and LDN
We observed that pretreatment of L-THP and LDN signif-icantly attenuated cocaine- or cue-induced reinstatement ofdrug-seeking behavior in a dose-dependent manner This isconsistent with previous findings using L-THP alone (Mantschet al 2007 2010 Figueroa-Guzman et al 2011 Wang andMantsch 2012) suggesting that the combination of L-THP andLDN does not alter therapeutic effects of L-THP In contrastthere is a trend of enhanced therapeutic effects in both cocaineself-administration and reinstatement tests compared withthe effects produced by the same doses of L-THP aloneAdditionally we extensively investigated the effect of a
10-day treatment on drug-seeking behavior during reinstate-ment This dosing schedule was designed to follow a more
Fig 5 Self-administration and drug-seeking behavior (A) Active and inactivelever presses over the course of theexperimental timeline There was nostatistical difference in stable self-administration between the groups (B)Comparison of drug-seeking behavior mea-sured by active lever presses After 13 self-administrations sessions animals weremoved to forced abstinence in which theyreceived treatment in their home cages for10 days On day 23 they were placed backinto the operant chambers with all cuesand rugs removed values are expressed asmean 6 SEM n = 5 for all groups Therewas a significant difference in active leverresponses on day 23 P = 00371 Stu-dentrsquos t test determined a significantdifference between the cocaine-vehiclegroup and the cocaine- 3 mgkg L-THP and01 mgkg LDN group P = 00073 Therewas also less significant difference be-tween cocaine-vehicle group and cocaine3 mgkg L-THP group P = 00464
Fig 6 Mean 6 SEM concentration ofplasma b-endorphin during forced absti-nence Plasma was obtained 30 minutesdirectly following last experimental sessionon day 23 There was a significant differencein b-endorphin concentration between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus vehiclegroup (P 005) as well as between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus 3 mgkgL-THP group P 005 Samples wereanalyzed in duplicates n = 5 for all groups
254 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

Effects of L-THP and LDN on Locomotion Animalswere administered L-THP LDN or L-THP1 LDN daily for 10consecutive days The timeline of 10 days was selected on thebasis of the findings in the above reinstatement experiments(see previous section) The baseline was created using theaverage of the spontaneous locomotion measurements takenover the first 3 consecutive days (3381 6 1681 cm averagebaseline) Baseline was subtracted from total movementsfor each day A dose of 3 mgkg or 5 mgkg L-THP alonesignificantly inhibited spontaneous locomotor activities overthe 60 minutes (two-way ANOVA [F(2140) 5 2234 P 0001]) whereas 01 mgkg LDN alone had no effect onspontaneous locomotion (Fig 4A) Both combination doses ofL-THP and LDN did not produce a significant reduction inspontaneous locomotion [F(2195) 5 02794 P 5 02794 two-way ANOVA] (Fig 4B) Effect size was determined to bemoderate to highly significant [h2 5 051]Effects of L-THP and LDN on Cue-Induced Cocaine-
Seeking After stable cocaine self-administration wasachieved animals underwent forced abstinence in their home
cages in the animal facility for 9 days during which 3 mgkgL-THP 3 mgkg L-THP and 01 mgkg LDN or vehicle wasgiven daily to three separate groups of rats (Fig 5A) Animalswere then placed back into the same self-administrationchambers where they obtained cocaine self-administrationbefore on day 23 Re-exposure to cocaine-associated contex-tual cues induced a significant cocaine-seeking behavior(ie active lever presses) in the absence of cocaine orcocaine-associated cues (Fig 5B the vehicle control group)Pretreatment with 3 mgkg L-THP or 3 mgkg L-THP and01 mgkg LDN produced a significant reduction in cue-induced drug-seeking compared with the vehicle controlgroup and the combination group was more effective thenL-THP alone One-way ANOVA for the data shown in Fig 5Brevealed a significant treatmentmain effect [F(319)5 3413P 5 0043] Moreover effect-size calculations revealed amoderate practical effect [h2 5 040] Studentrsquos t testsrevealed a significant reduction in cocaine-seeking after3 mgkg L-THP [t(8) 5 2354 P 5 00464] or 3 mgkg L-THPand 01 mgkg LDN [t(8) 5 3581 P 5 00072]
Fig 3 Effect of 10 days administration of L-THP and LDN on cocaine reinstatement Animals received intraperitoneal injections for 10 days untilextinction was reached In the experimental session following animals were primed with 10 mgkg cocaine and placed back into operant chambers(A and B) Extinction timeline arrows representing administration of L-THP (3 mgkg n = 9 and 5 mgkg n = 10) LDN (01 mgkg n = 6) or L-THP andLDN combinations (3mgkg and 01 mgkg n = 10 and 5mgkg and 01 mgkg n = 11) (C) Significantly inhibited reinstatement of drug-seeking behaviorassessed by active lever responses expressed as mean 6 SEM Bonferroni post-hoc analysis determined that both the combinations of 3 mgkg L-THPand 01 mgkg LDN and 5mgkg L-THP and 01 mgkg LDNwere more efficacious than any of the single doses P 05 P 01 P 0001 P00001 compared with vehicle group (n = 7)
252 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
Effects of L-THP and LDN on Plasma Concentrationsof b-Endorphin in Rats after Forced Cocaine AbstinenceAfter the completion of the above behavioral test plasmasamples were collected 30 minutes via the implanted in-travenous catheter Coadministration of 3 mgkg L-THP and01 mgkg LDN significantly elevated plasma b-endorphinlevel compared with the vehicle control group A dose of3 mgkg L-THP alone had no effect (Fig 6) One-way ANOVAfor the data shown in Fig 6 revealed a significant treatmentmain effect [F(339) 5 392 P 5 0016] However the effect-size calculations demonstrate a moderate to low significanteffect [h2 5 024]Effects of L-THP and LDN on POMC mRNA Expres-
sion in Rats after Forced Cocaine Abstinence Figure 7Ashows the RNAscope in situ hybridization data illustratingthat coadministration of 3 mgkg L-THP and 01 mgkg LDNsignificantly elevated POMC expression in the hypothala-mus whereas 3 mgkg L-THP alone had no effect One-wayANOVA for the summarized data shown in Fig 7B revealed asignificant treatment main effect [F(3119) 5 6529 P 500004] Effect-size calculations suggest only a minimalpractical effect [h2 5 014] Bonferroni post-hoc analysesindicated a significant increase in POMC expression after3 mgkg L-THP and 01 mgkg LDN (P 005) but not after3 mgkg L-THP alone (P 005) compared with the vehiclegroupCorrelation Between Cocaine-Seeking Behavior
b-Endorphin and POMC Expression Figure 8A showsthe strong significant negative correlation between activelever presses during reinstatement and plasma concentrationsof b-endorphin [r(15)5 ndash0617 P5 00142] Also shown in Fig8B is the weak positive correlation between POMC expressionand plasma concentrations of b-endorphin [r(15) 5 0361
P 5 0187] A correlation was not observed between POMCexpression and active lever presses during reinstatementshown in Fig 8C [r(15) 5 ndash0214 P 5 0440]
DiscussionCombination medications are becoming increasingly com-
monplace in clinical practice and can be especially effectivewhen used to treat individuals suffering from mental disor-ders and who are multisymptomatic (Tohen et al 2003Kupfer et al 2012 Biler et al 2014 Shorter et al 2015) Inthe case of relapse prevention drug cravings may beeffectively managed when medications treat both behavioraland physiologic symptoms However the majority of phar-maceuticals developed for the treatment of substance abusedisorders target one pathway The combination of L-THP andLDN targets both dopaminergic signaling and the regulationof endogenous opioids In this report we provide evidencethat the combination of L-THP and LDN is more potent withfewer unwanted side effects than L-THP alone on the basis ofthe following observations 1) L-THP and LDN has demon-strated significantly greater effect in attenuating drug-seeking behavior than L-THP alone 2) L-THP and LDN whenadministered together do not result in a reduction ofspontaneous locomotion 3) L-THP and LDN appears toincrease peripheral concentrations of b-endorphin 4) L-THPand LDN appears to upregulate expression of POMC in thearcuate nucleus
L-THP has been investigated as a potential treatment ofaddiction by several other groups (Mantsch et al 2007 2010Figueroa-Guzman et al 2011 Wang and Mantsch 2012)Presently we determined that L-THP produced a biphasiceffect and this is consistent with the results of previous study
Fig 4 Effect of L-THP and LDN onspontaneous locomotion Animals estab-lished a baseline over 3 consecutive daysfollowing that they were given intraperi-toneal injections of the assigned treat-ments for 10 days Values are expressedas mean of the locomotor score 6 SEMn = 6 for all groups Two-way ANOVAdetermined treatment had a significanteffect on spontaneous locomotion P00001 (A) L-THP (3 mgkg and 5 mgkg)produced a dose-dependent reduction onlocomotion and LDN (01 mgkg) had noeffect on locomotion (B) Effect of co-administration of L-THP and LDN
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 253
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
(Xi et al 2007) The increase in cocaine self-administration isattributed to the partial blockade of postsynaptic DA recep-tors which decrease the rewarding properties of cocaine andlead to a compensatory increase in drug-taking Even as thehigher doses of L-THP drastically reduced cocaine self-administration we also noted a strong sedative effectassessed by a decrease in locomotion suggesting that nonspe-cific sedation or locomotion impairment is the main cause forreduced self-administration in the high L-THP dose group Incontrast to L-THP LDN had no effect on cocaine self-administration There was no difference in cocaine self-administration between animals receiving lower doses ofL-THP or L-THP and LDN
We observed that pretreatment of L-THP and LDN signif-icantly attenuated cocaine- or cue-induced reinstatement ofdrug-seeking behavior in a dose-dependent manner This isconsistent with previous findings using L-THP alone (Mantschet al 2007 2010 Figueroa-Guzman et al 2011 Wang andMantsch 2012) suggesting that the combination of L-THP andLDN does not alter therapeutic effects of L-THP In contrastthere is a trend of enhanced therapeutic effects in both cocaineself-administration and reinstatement tests compared withthe effects produced by the same doses of L-THP aloneAdditionally we extensively investigated the effect of a
10-day treatment on drug-seeking behavior during reinstate-ment This dosing schedule was designed to follow a more
Fig 5 Self-administration and drug-seeking behavior (A) Active and inactivelever presses over the course of theexperimental timeline There was nostatistical difference in stable self-administration between the groups (B)Comparison of drug-seeking behavior mea-sured by active lever presses After 13 self-administrations sessions animals weremoved to forced abstinence in which theyreceived treatment in their home cages for10 days On day 23 they were placed backinto the operant chambers with all cuesand rugs removed values are expressed asmean 6 SEM n = 5 for all groups Therewas a significant difference in active leverresponses on day 23 P = 00371 Stu-dentrsquos t test determined a significantdifference between the cocaine-vehiclegroup and the cocaine- 3 mgkg L-THP and01 mgkg LDN group P = 00073 Therewas also less significant difference be-tween cocaine-vehicle group and cocaine3 mgkg L-THP group P = 00464
Fig 6 Mean 6 SEM concentration ofplasma b-endorphin during forced absti-nence Plasma was obtained 30 minutesdirectly following last experimental sessionon day 23 There was a significant differencein b-endorphin concentration between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus vehiclegroup (P 005) as well as between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus 3 mgkgL-THP group P 005 Samples wereanalyzed in duplicates n = 5 for all groups
254 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

Effects of L-THP and LDN on Plasma Concentrationsof b-Endorphin in Rats after Forced Cocaine AbstinenceAfter the completion of the above behavioral test plasmasamples were collected 30 minutes via the implanted in-travenous catheter Coadministration of 3 mgkg L-THP and01 mgkg LDN significantly elevated plasma b-endorphinlevel compared with the vehicle control group A dose of3 mgkg L-THP alone had no effect (Fig 6) One-way ANOVAfor the data shown in Fig 6 revealed a significant treatmentmain effect [F(339) 5 392 P 5 0016] However the effect-size calculations demonstrate a moderate to low significanteffect [h2 5 024]Effects of L-THP and LDN on POMC mRNA Expres-
sion in Rats after Forced Cocaine Abstinence Figure 7Ashows the RNAscope in situ hybridization data illustratingthat coadministration of 3 mgkg L-THP and 01 mgkg LDNsignificantly elevated POMC expression in the hypothala-mus whereas 3 mgkg L-THP alone had no effect One-wayANOVA for the summarized data shown in Fig 7B revealed asignificant treatment main effect [F(3119) 5 6529 P 500004] Effect-size calculations suggest only a minimalpractical effect [h2 5 014] Bonferroni post-hoc analysesindicated a significant increase in POMC expression after3 mgkg L-THP and 01 mgkg LDN (P 005) but not after3 mgkg L-THP alone (P 005) compared with the vehiclegroupCorrelation Between Cocaine-Seeking Behavior
b-Endorphin and POMC Expression Figure 8A showsthe strong significant negative correlation between activelever presses during reinstatement and plasma concentrationsof b-endorphin [r(15)5 ndash0617 P5 00142] Also shown in Fig8B is the weak positive correlation between POMC expressionand plasma concentrations of b-endorphin [r(15) 5 0361
P 5 0187] A correlation was not observed between POMCexpression and active lever presses during reinstatementshown in Fig 8C [r(15) 5 ndash0214 P 5 0440]
DiscussionCombination medications are becoming increasingly com-
monplace in clinical practice and can be especially effectivewhen used to treat individuals suffering from mental disor-ders and who are multisymptomatic (Tohen et al 2003Kupfer et al 2012 Biler et al 2014 Shorter et al 2015) Inthe case of relapse prevention drug cravings may beeffectively managed when medications treat both behavioraland physiologic symptoms However the majority of phar-maceuticals developed for the treatment of substance abusedisorders target one pathway The combination of L-THP andLDN targets both dopaminergic signaling and the regulationof endogenous opioids In this report we provide evidencethat the combination of L-THP and LDN is more potent withfewer unwanted side effects than L-THP alone on the basis ofthe following observations 1) L-THP and LDN has demon-strated significantly greater effect in attenuating drug-seeking behavior than L-THP alone 2) L-THP and LDN whenadministered together do not result in a reduction ofspontaneous locomotion 3) L-THP and LDN appears toincrease peripheral concentrations of b-endorphin 4) L-THPand LDN appears to upregulate expression of POMC in thearcuate nucleus
L-THP has been investigated as a potential treatment ofaddiction by several other groups (Mantsch et al 2007 2010Figueroa-Guzman et al 2011 Wang and Mantsch 2012)Presently we determined that L-THP produced a biphasiceffect and this is consistent with the results of previous study
Fig 4 Effect of L-THP and LDN onspontaneous locomotion Animals estab-lished a baseline over 3 consecutive daysfollowing that they were given intraperi-toneal injections of the assigned treat-ments for 10 days Values are expressedas mean of the locomotor score 6 SEMn = 6 for all groups Two-way ANOVAdetermined treatment had a significanteffect on spontaneous locomotion P00001 (A) L-THP (3 mgkg and 5 mgkg)produced a dose-dependent reduction onlocomotion and LDN (01 mgkg) had noeffect on locomotion (B) Effect of co-administration of L-THP and LDN
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 253
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
(Xi et al 2007) The increase in cocaine self-administration isattributed to the partial blockade of postsynaptic DA recep-tors which decrease the rewarding properties of cocaine andlead to a compensatory increase in drug-taking Even as thehigher doses of L-THP drastically reduced cocaine self-administration we also noted a strong sedative effectassessed by a decrease in locomotion suggesting that nonspe-cific sedation or locomotion impairment is the main cause forreduced self-administration in the high L-THP dose group Incontrast to L-THP LDN had no effect on cocaine self-administration There was no difference in cocaine self-administration between animals receiving lower doses ofL-THP or L-THP and LDN
We observed that pretreatment of L-THP and LDN signif-icantly attenuated cocaine- or cue-induced reinstatement ofdrug-seeking behavior in a dose-dependent manner This isconsistent with previous findings using L-THP alone (Mantschet al 2007 2010 Figueroa-Guzman et al 2011 Wang andMantsch 2012) suggesting that the combination of L-THP andLDN does not alter therapeutic effects of L-THP In contrastthere is a trend of enhanced therapeutic effects in both cocaineself-administration and reinstatement tests compared withthe effects produced by the same doses of L-THP aloneAdditionally we extensively investigated the effect of a
10-day treatment on drug-seeking behavior during reinstate-ment This dosing schedule was designed to follow a more
Fig 5 Self-administration and drug-seeking behavior (A) Active and inactivelever presses over the course of theexperimental timeline There was nostatistical difference in stable self-administration between the groups (B)Comparison of drug-seeking behavior mea-sured by active lever presses After 13 self-administrations sessions animals weremoved to forced abstinence in which theyreceived treatment in their home cages for10 days On day 23 they were placed backinto the operant chambers with all cuesand rugs removed values are expressed asmean 6 SEM n = 5 for all groups Therewas a significant difference in active leverresponses on day 23 P = 00371 Stu-dentrsquos t test determined a significantdifference between the cocaine-vehiclegroup and the cocaine- 3 mgkg L-THP and01 mgkg LDN group P = 00073 Therewas also less significant difference be-tween cocaine-vehicle group and cocaine3 mgkg L-THP group P = 00464
Fig 6 Mean 6 SEM concentration ofplasma b-endorphin during forced absti-nence Plasma was obtained 30 minutesdirectly following last experimental sessionon day 23 There was a significant differencein b-endorphin concentration between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus vehiclegroup (P 005) as well as between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus 3 mgkgL-THP group P 005 Samples wereanalyzed in duplicates n = 5 for all groups
254 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

(Xi et al 2007) The increase in cocaine self-administration isattributed to the partial blockade of postsynaptic DA recep-tors which decrease the rewarding properties of cocaine andlead to a compensatory increase in drug-taking Even as thehigher doses of L-THP drastically reduced cocaine self-administration we also noted a strong sedative effectassessed by a decrease in locomotion suggesting that nonspe-cific sedation or locomotion impairment is the main cause forreduced self-administration in the high L-THP dose group Incontrast to L-THP LDN had no effect on cocaine self-administration There was no difference in cocaine self-administration between animals receiving lower doses ofL-THP or L-THP and LDN
We observed that pretreatment of L-THP and LDN signif-icantly attenuated cocaine- or cue-induced reinstatement ofdrug-seeking behavior in a dose-dependent manner This isconsistent with previous findings using L-THP alone (Mantschet al 2007 2010 Figueroa-Guzman et al 2011 Wang andMantsch 2012) suggesting that the combination of L-THP andLDN does not alter therapeutic effects of L-THP In contrastthere is a trend of enhanced therapeutic effects in both cocaineself-administration and reinstatement tests compared withthe effects produced by the same doses of L-THP aloneAdditionally we extensively investigated the effect of a
10-day treatment on drug-seeking behavior during reinstate-ment This dosing schedule was designed to follow a more
Fig 5 Self-administration and drug-seeking behavior (A) Active and inactivelever presses over the course of theexperimental timeline There was nostatistical difference in stable self-administration between the groups (B)Comparison of drug-seeking behavior mea-sured by active lever presses After 13 self-administrations sessions animals weremoved to forced abstinence in which theyreceived treatment in their home cages for10 days On day 23 they were placed backinto the operant chambers with all cuesand rugs removed values are expressed asmean 6 SEM n = 5 for all groups Therewas a significant difference in active leverresponses on day 23 P = 00371 Stu-dentrsquos t test determined a significantdifference between the cocaine-vehiclegroup and the cocaine- 3 mgkg L-THP and01 mgkg LDN group P = 00073 Therewas also less significant difference be-tween cocaine-vehicle group and cocaine3 mgkg L-THP group P = 00464
Fig 6 Mean 6 SEM concentration ofplasma b-endorphin during forced absti-nence Plasma was obtained 30 minutesdirectly following last experimental sessionon day 23 There was a significant differencein b-endorphin concentration between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus vehiclegroup (P 005) as well as between thecocaine plus 3 mgkg L-THP and 01 mgkgLDN group and the cocaine plus 3 mgkgL-THP group P 005 Samples wereanalyzed in duplicates n = 5 for all groups
254 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

clinically relevant timeline as often a repeated daily treat-ment plan is used as opposed to single treatment prior to arelapse (Katz andHiggins 2003 Epstein et al 2006 Yahyavi-Firouz-Abadi and See 2009 Bailey and Husbands 2014)
Cocaine reinstatement tests were performed about18ndash20 hours after the final dosing of L-THP and LDN Half-lives of L-THP andNTX are 10 hours and 4 hours respectivelyleaving less than 25 and 3125 of the bolus doses of L-THP
Fig 7 Fluorescence intensity of POMC mRNA duringforced abstinence (A) POMC mRNA in red detectedby RNAscope Brains were harvested and snap-frozen30 minutes following the last experimental session on day23 Six 014-mm coronal sections were taken from eachanimal (n = 5 for all groups total of 30 sections per group)and analyzed for total POMC mRNA fluorescence be-tween ndash2 mm and ndash35 mm bregma 3V third ventriclescale bar 100 mm (B) Fluorescence intensity of POMCmRNA expressed as mean 6 SEM Treatment had asignificant effect on POMC mRNA intensity P = 00004Bonferroni post-hoc analysis revealed a significant differ-ence between the cocaine plus vehicle and the cocaine plus3 mgkg L-THP and 01 mgkg LDN group P 001between cocaine plus 3 mgkg L-THP and cocaine plus3 mgkg L-THP and 01 mgkg LDN P 001
Fig 8 Correlation between cocaine-seeking behavior b-endorphin and POMC expression (A) Strong negative correlation between plasmaconcentrations of b-endorphin and cocaine-seeking behavior during reinstatement r = ndash0617 P 005 (B) Weak positive correlation butnonsignificant between plasma concentrations of b-endorphin and POMC expression r = 361P = 0187 Finally (C) demonstrates the lack of correlationbetween cocaine-seeking behavior during reinstatement and POMC expression r = 0214 P = 0440 n = 15 for all correlation analysis
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 255
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

and LDN biologically available at the time of reinstatementtest (Dunbar et al 2006 Slawson et al 2007Wu et al 2013)suggesting that repeated drug treatment produced aprolonged therapeutic effect in prevention of relapse to drug-seeking behavior To our knowledge we are the first to reportthat the combination of L-THP and LDN as well as L-THPalone administered on this schedule significantly attenuatesdrug-seeking behavior A previous study conducted by Gerritset al (2005) observed 3 mgkg NTX attenuates cocainereinstatement only when given repeatedly the authorssuggested that the endogenous opioid system specificallyb-endorphin may be involved in relapse to cocaine-seekingbehaviorThe 10-day administration of L-THP reduced spontaneous
locomotion at both 3 mgkg and 5 mgkg doses Previousexperiments reported observing sedation at slightly higherdoses however we attributed the observed differences tochanges in the locomotion experimental design (Xi et al 2007Xu et al 2013) Coadministration of L-THP with LDNprevented the reduction of locomotion produced by L-THPalone yet LDN alone did not alter locomotor behavior at sucha low dose suggesting that LDN coadministration may blockthe sedative effects of L-THP This effect may be mediatedthrough brief opioid receptor antagonism leading to a tran-sient b-endorphin release (Corder et al 2013) In turn theb-endorphin induces the release of DA in the nucleusaccumbens via activation of m-opioid receptors on GABAergicneurons that inhibits ventral tegmental area GABAergicneurons (Simmons and Self 2009 Spanagel et al1991)resulting in the overall increase of spontaneous locomotionTo test this b-endorphin hypothesis we further investigated
the effect of L-THP and LDN on plasma b-endorphin and onthe expression of the precursor polypeptide POMC andmeasured both b-endorphin and POMC in rats after 10 daysof withdrawal from the last cocaine self-administrationsession in the absence or presence of L-THP and LDN pre-treatment The 3-mgkg L-THP and 01-mgkg LDN treatmentsignificantly increased plasmas b-endorphin concentrationssupporting our hypothesis that treatment with L-THP andLDN in rats during forced abstinence may initially increasebrain b-endorphin release resulting in an increase in periph-eral concentrations of b-endorphin We found a significantreduction of POMC gene expression in the cocaine self-administration group compared with the sucrose self-administration group Pretreatment with L-THP and LDNmight renormalize (or increase) brain POMC expression Wedo however acknowledge that there could be several othermediators at play whose activation could also elicit a similareffect to the one seen in the present study POMC is also theprecursor to adrenocorticotropic hormone and melanocyte-stimulating hormone and therefore the effect on POMC couldbe unrelated to the result of one the factors outside the scope ofthese experimentOur findings solidify a connection between cocaine-seeking
behavior and the endogenous opioids and corroborate thefindings of many other groups (Olive et al 2001 Kiefer et al2002 Marinelli et al 2003 2004 Roth-Deri et al 2003 2008Shaw and alrsquoAbsi 2008 Seo et al 2009 Nguyen et al 2012Dikshtein et al 2013 Zhou et al 2013) b-endorphin is knownto play an important role in addictions In recovering smokersattenuated concentrations of b-endorphin directly contributeto smoking relapse (Marinelli et al 2003 Roth-Deri et al
2008 Shaw and alrsquoAbsi 2008) In addition to being acontributing factor in cocaine nicotine and heroin craving ab-endorphin deficit is hypothesized as a contributing factor inrelapse of alcohol use (Roth-Deri et al 2004 Dikshtein et al2013) Hypothalamic POMC expression is significantlyreduced in animals that return to heroin-seeking behaviorduring relapse (Zhou Y et al 2015)We believe the evidence presented supports a b-endorphin-
deficit hypothesis Other groups have demonstrated a corre-lation between attenuated b-endorphin release in the brainand cocaine-seeking behavior (Dikshtein et al 2013) how-ever to our knowledge we are the first to demonstrate a strongnegative correlationbetween reinstatementplasmab-endorphinFrom a more meaningful and practical point of view post-hocanalysis demonstrated weak positive correlation betweenplasma levels of b-endorphin and brain POMC expressionAlthough this correlation was not significant we believe thatthis result was limited by of our relatively small sample sizeand hypothesize that future experiments conducted with alarger sample size would demonstrate a significant positivecorrelation between plasma b-endorphin and POMC expres-sion Alternatively the weak correlation may be the result ofother modulations of POMC expression which are beyond thescope of the present studyLDN is used as an off-label treatment of autoimmune
disorders however there is no conclusive study examiningits mechanism (Brown and Panksepp 2009) As L-THP is welldocumented not to possess affinity for opioid receptors thechanges in b-endorphin and POMC expression are believed tobe the result of treatment with LDN However the completemechanism by which L-THP or the combination of L-THP andLDN exert their effects remains elusive L-THP is reported tofacilitate DA release in the nucleus accumbens normalizingDA signaling DA changes combined with increase of hypo-thalamic POMC and b-endorphin during forced abstinencepresumably reduce drug cravings attenuating drug-seekingbehavior Furthermore the b-endorphin release increasesspontaneous locomotion in rodents which explains spon-taneous locomotion observed between animals treatedwith L-THP and L-THP and LDN (Spanagel et al 1991Marquez et al 2008)In summary the data presented is evidence of the thera-
peutic potential of the combination of L-THP and LDN as anefficacious treatment for prevention of cocaine relapse Wehypothesize L-THP and LDN mediates release of endogenousopioids and dopamine allowing dopaminergic signaling in thebrain to approach preaddiction homeostasis This treats boththe behavioral symptoms and physiologic symptoms observedduring recovery and reduces drug cravings and relapsesFuture studies should explore the proposed mechanism ofLDN as well as validate efficacy of L-THP and LDN in humantrials
Acknowledgments
The authors would like to thank Guo-Hua Bi Haying Zhang andXiao-Fei Wang for technical assistance
Authorship Contributions
Participated in research design Xi Wang SushchykConducted experiments and preformed data analysis SushchykWrote or contributed to writing of the manuscript Xi Wang
Sushchyk
256 Sushchyk et al
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from
References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from

References
Amen SL Piacentine LB Ahmad ME Li SJ Mantsch JR Risinger RC and BakerDA (2011) Repeated N-acetyl cysteine reduces cocaine seeking in rodents andcraving in cocaine-dependent humans Neuropsychopharmacology 36871ndash878
Bailey CP and Husbands SM (2014) Novel approaches for the treatment ofpsychostimulant and opioid abuse - focus on opioid receptor-based therapiesExpert Opin Drug Discov 91333ndash1344
Brown N and Panksepp J (2009) Low-dose naltrexone for disease prevention andquality of life Med Hypotheses 72333ndash337
Corder G Doolen S Donahue RR Winter MK Jutras BL He Y Hu X Wieskopf JSMogil JS and Storm DR et al (2013) Constitutive m-opioid receptor activity leadsto long-term endogenous analgesia and dependence Science 3411394ndash1399
Cordery SF Taverner A Ridzwan IE Guy RH Delgado-Charro MB Husbands SMand Bailey CP (2014) A non-rewarding non-aversive buprenorphinenaltrexonecombination attenuates drug-primed reinstatement to cocaine and morphine inrats in a conditioned place preference paradigm Addict Biol 19575ndash586
Dackis CA and OrsquoBrien CP (2001) Cocaine dependence a disease of the brainrsquosreward centers J Subst Abuse Treat 21111ndash117
Dikshtein Y Barnea R Kronfeld N Lax E Roth-Deri I Friedman A and Yadid G(2013) Beta-endorphin via the delta opioid receptor is a major factor in the incubationof cocaine craving Neuropsychopharmacology 382508ndash2514 101038npp2013155
Ding GS (1987) Important Chinese herbal remedies Clin Ther 9345ndash357Dunbar JL Turncliff RZ Dong Q Silverman BL Ehrich EW and Lasseter KC (2006)Single- and multiple-dose pharmacokinetics of long-acting injectable naltrexoneAlcohol Clin Exp Res 30480ndash490
Epstein DH Preston KL Stewart J and Shaham Y (2006) Toward a model of drugrelapse an assessment of the validity of the reinstatement procedure Psycho-pharmacology (Berl) 1891ndash16
Figueroa-Guzman Y Mueller C Vranjkovic O Wisniewski S Yang Z Li SJ Bohr CGraf EN Baker DA and Mantsch JR (2011) Oral administration of levo-tetrahydropalmatine attenuates reinstatement of extinguished cocaine seekingby cocaine stress or drug-associated cues in rats Drug Alcohol Depend 11672ndash79
Gerrits M A Kuzmin A V and van Ree J M (2005) Reinstatement of cocaine-seeking behavior in rats is attenuated following repeated treatment with the opioidreceptor antagonist naltrexone European neuropsychopharmacology 15(3)297ndash303
Grassi MC Cioce AM Giudici FD Antonilli L and Nencini P (2007) Short-termefficacy of Disulfiram or Naltrexone in reducing positive urinalysis for both cocaineand cocaethylene in cocaine abusers a pilot study Pharmacol Res 55117ndash121
Haney M Ward AS Foltin RW and Fischman MW (2001) Effects of ecopipam aselective dopamine D1 antagonist on smoked cocaine self-administration byhumans Psychopharmacology (Berl) 155330ndash337
Jin GZ Wang XL and Shi WX (1986a) Tetrahydroprotoberberinendasha new chemicaltype of antagonist of dopamine receptors Scientia Sinica Series B 29527ndash534
Katz JL and Higgins ST (2003) The validity of the reinstatement model of cravingand relapse to drug use Psychopharmacology (Berl) 16821ndash30
Kiefer F Horntrich M Jahn H and Wiedemann K (2002) Is withdrawaL-inducedanxiety in alcoholism based on beta-endorphin deficiency Psychopharmacology(Berl) 162433ndash437
Koob GF and Volkow ND (2010) Neurocircuitry of addictionNeuropsychopharmacology35217ndash238
Liu YL Yan LD Zhou PL Wu CF and Gong ZH (2009) Levo-tetrahydropalmatineattenuates oxycodone-induced conditioned place preference in rats Eur J Pharmacol602321ndash327
Mannelli P Peindl K Patkar A A Wu L T Pae C U and Gorelick D A (2010)Reduced cannabis use after low-dose naltrexone addition to opioid detoxificationJ Clin Psychopharmacol 30476-478 DOI101097JCP0b013e3181e5c168
Mantsch JR Li SJ Risinger R Awad S Katz E Baker DA and Yang Z (2007) Levo-tetrahydropalmatine attenuates cocaine self-administration and cocaine-inducedreinstatement in rats Psychopharmacology (Berl) 192581ndash591
Mantsch JR Wisniewski S Vranjkovic O Peters C Becker A Valentine A Li SJBaker DA and Yang Z (2010) Levo-tetrahydropalmatine attenuates cocaine self-administration under a progressive-ratio schedule and cocaine discrimination inrats Pharmacol Biochem Behav 97310ndash316
Marcenac F Jin GZ and Gonon F (1986) Effect of L-tetrahydropalmatine on dopa-mine release and metabolism in the rat striatum Psychopharmacology (Berl) 8989ndash93
Marinelli PW Quirion R and Gianoulakis C (2003) A microdialysis profile of beta-endorphin and catecholamines in the rat nucleus accumbens following alcoholadministration Psychopharmacology (Berl) 16960ndash67
Marinelli PW Quirion R and Gianoulakis C (2004) An in vivo profile of beta-endorphin release in the arcuate nucleus and nucleus accumbens following expo-sure to stress or alcohol Neuroscience 127777ndash784
Marquez P Baliram R Dabaja I Gajawada N and Lutfy K (2008) The role of beta-endorphin in the acute motor stimulatory and rewarding actions of cocaine in micePsychopharmacology (Berl) 197443ndash448
Nestler EJ (2005) The neurobiology of cocaine addiction Sci Pract 34ndash10Nguyen AT Marquez P Hamid A Kieffer B Friedman TC and Lutfy K (2012) Therewarding action of acute cocaine is reduced in b-endorphin deficient but notin m opioid receptor knockout mice Eur J Pharmacol 68650ndash54
Olive MF Koenig HN Nannini MA and Hodge CW (2001) Stimulation of endorphinneurotransmission in the nucleus accumbens by ethanol cocaine and amphet-amine J Neurosci 21RC184
Paxinos G Watson C (2006) The Rat Brain in Stereotaxic Coordinates (6th ed)Cambridge MA Academic Press
Platt DM Rowlett JK and Spealman RD (2002) Behavioral effects of cocaine anddopaminergic strategies for preclinical medication development Psychopharma-cology (Berl) 163265ndash282
Rahn KA McLaughlin PJ and Zagon IS (2011) Prevention and diminished expres-sion of experimental autoimmune encephalomyelitis by low dose naltrexone (LDN)or opioid growth factor (OGF) for an extended period Therapeutic implications formultiple sclerosis Brain Res 1381243ndash253
Roth-Deri I Zangen A Aleli M Goelman RG Pelled G Nakash R Gispan-Herman IGreen T Shaham Y and Yadid G (2003) Effect of experimenter-delivered and self-administered cocaine on extracellular beta-endorphin levels in the nucleusaccumbens J Neurochem 84930ndash938
Roth-Deri I Schindler CJ and Yadid G (2004) A critical role for beta-endorphin incocaine-seeking behavior Neuroreport 15519ndash521
Roth-Deri I Green-Sadan T and Yadid G (2008) Beta-endorphin and drug-inducedreward and reinforcement Prog Neurobiol 861ndash21
Schmitz JM Stotts AL Rhoades HM and Grabowski J (2001) Naltrexone and relapseprevention treatment for cocaine-dependent patients Addict Behav 26167ndash180
Schmitz JM Lindsay JA Green CE Herin DV Stotts AL and Moeller FG (2009)High-dose naltrexone therapy for cocaine-alcohol dependence Am J Addict 18356ndash362
Schmitz JM Green CE Stotts AL Lindsay JA Rathnayaka NS Grabowski Jand Moeller FG (2014) A two-phased screening paradigm for evaluating candidatemedications for cocaine cessation or relapse prevention modafinil levodopa-carbidopa naltrexone Drug Alcohol Depend 136100ndash107
Segal D Macdonald JK and Chande N (2014) Low dose naltrexone for induction ofremission in Crohnrsquos disease Cochrane Database Syst Rev 2CD010410
Seo YJ Kwon MS Choi SM Lee JK Park SH Jung JS Sim YB and Suh HW (2009)Possible involvement of the hypothalamic pro-opiomelanocortin gene and beta-endorphin expression on acute morphine withdrawal development Brain Res Bull80359ndash370
Shaw D and alrsquoAbsi M (2008) Attenuated beta endorphin response to acute stressis associated with smoking relapse Pharmacol Biochem Behav 90357ndash362DOI101016jpbb200803020
Shorter D and Kosten TR (2011) Novel pharmacotherapeutic treatments for cocaineaddiction BMC Med 9119
Simmons D and Self DW (2009) Role of mu- and delta-opioid receptors in the nucleusaccumbens in cocaine-seeking behavior Neuropsychopharmacology 341946ndash1957
Slawson MH Chen M Moody D Comer SD Nuwayser ES Fang WB and Foltz RL(2007) Quantitative analysis of naltrexone and 6beta-naltrexol in human rat andrabbit plasma by liquid chromatography-electrospray ionization tandem massspectrometry with application to the pharmacokinetics of Depotrex in rabbitsJ Anal Toxicol 31453ndash461
Spanagel R Herz A Bals-Kubik R and Shippenberg TS (1991) Beta-endorphin-inducedlocomotor stimulation and reinforcement are associated with an increase in dopaminerelease in the nucleus accumbens Psychopharmacology (Berl) 10451ndash56
Trigo JM Martin-Garciacutea E Berrendero F Robledo P and Maldonado R (2010) Theendogenous opioid system a common substrate in drug addiction Drug AlcoholDepend 108183ndash194
Ueno K Kiguchi N Kobayashi Y Saika F Wakida N Yamamoto C Maeda T OzakiM Kishioka S (2014) Possible involvement of endogenous opioid system locateddownstream of a7 nicotinic acetylcholine receptor in mice with physicaldependence on nicotine J Pharmacol Sci 12447ndash53
Wang JB and Mantsch JR (2012) L-tetrahydropalamatine a potential new medica-tion for the treatment of cocaine addiction Future Med Chem 4177ndash186
Wee S Vendruscolo LF Misra KK Schlosburg JE and Koob GF (2012) A combi-nation of buprenorphine and naltrexone blocks compulsive cocaine intake in ro-dents without producing dependence Sci Translat MedI 4146ra110 DOI 101126scitranslmed3003948
Wu X Luo Y Xu J Gu S Luo X Wu H and Zhao G (2013) Determination of levo-tetrahydropalmatine in rat plasma by HPLC and its application to pharmacoki-netics studies Afr J Pharm Pharmacol 7545ndash551
Xi ZX Yang Z Li SJ Li X Dillon C Peng XQ Spiller K and Gardner EL (2007) Levo-tetrahydropalmatine inhibits cocainersquos rewarding effects experiments with self-administration and brain-stimulation reward in rats Neuropharmacology 53771ndash782
Xu W Wang Y Ma Z Chiu YT Huang P Rasakham K Unterwald E Lee DYand Liu-Chen LY (2013) L-isocorypalmine reduces behavioral sensitization andrewarding effects of cocaine in mice by acting on dopamine receptors Drug AlcoholDepend 133693ndash703
Yang Z Shao YC Li SJ Qi JL Zhang MJ Hao W and Jin GZ (2008) Medication ofL-tetrahydropalmatine significantly ameliorates opiate craving and in-creases the abstinence rate in heroin users a pilot study Acta Pharmacol Sin 29781ndash788
Yoo JH Kitchen I and Bailey A (2012) The endogenous opioid system in cocaineaddiction what lessons have opioid peptide and receptor knockout mice taught usBr J Pharmacol 1661993ndash2014
Zagon I Hammer L and McLaughlin P (2014) Low dose naltrexone inhibits theprogression of clinical disease in established relapse-remitting experimental au-toimmune encephalomyelitismdasha model for multiple sclerosis (6513) FASEB J 28(1 Supplement) 6513
Zhou Y Leri F Cummins E and Kreek MJ (2015) Individual differences in geneexpression of vasopressin D2 receptor POMC and orexin vulnerability to relapseto heroin-seeking in rats Physiol Behav 139127ndash135
Zhou Y Colombo G Niikura K Carai MA Femeniacutea T Garciacutea-Gutieacuterrez MS ManzanaresJ Ho A Gessa GL and Kreek MJ (2013) Voluntary alcohol drinking enhancesproopiomelanocortin gene expression in nucleus accumbens shell and hypothalamus ofSardinian alcohoL-preferring rats Alcohol Clin Exp Res 37 (Suppl 1)E131ndashE140
Address correspondence to Dr Jia Bei Wang Department of Pharmacol-ogy University of Maryland School of Pharmacy 20 N Pine St Room N527Baltimore MD 21201 jwangrxumarylandedu
L-THP and LDN Novel Therapy to Prevent Cocaine Relapse 257
at ASPE
T Journals on M
ay 19 2020jpetaspetjournalsorg
Dow
nloaded from


















