
Cloning of a gene encoding the immunodominant surface antigen of Leishmania donovani promastigotes
12
Molecular and Biochemical Parasitology. 23 (1987) 211-222 211 Elsevier MBP 00788 Cloning of a gene encoding the immunodominant surface antigen of Leishmania donovani promastigotes Stephen Heath 1, Michael L. Chance 1, Marcel Hommel: and Julian M. Crampton 3 ~Department of Parasitology. "-Department of Troptcal Medicine. and 3Wolfson Molecular Genetics Untt. Liverpool School of Tropical Medicine. Liverpool. U.K. (Received 19 September 1986; Accepted 3 December 1986) This study describes the characterisation of externally oriented surface peptides of both morphological forms of Leishmania don- ovani, the causative agent of visceral leishmaniasis (kala-azar). Using t-'-~i surface labelling techniques and peptide extraction in the detergents Triton X-I(KI and Triton X-I 1-, t. a major iodinable promastigote peptide at 63 kDa or 65 kDa (depending on de- tergent used) was identified. This peptide was demonstrated to be the immunodominant membrane peptide of L. donovani and ~as strongly recognised by human sera from parasitologicallyconfirmed cases of kala-azar. This peptide was not demonstrated on the surface of tissue amastigotes, although in vitro translations of poly(A*) RNA from both promastigotes and amastigotes dem- onstrated that both forms possessed mRNA that directs the synthesis of a 63 kDa peptide. It is suggested therefore that in amas- tigotes this peptide ma~ be a processed antigen. We also report the isolation of a recombinant cDNA clone in the bacteriophage vector h gtl0 which encodes a 63 kDa polypeptide that is recognised b.~ human kala-azar sera. It is proposed that this surface peptide could be used in a specific immunodiagnostic test for leishmaniasis. Ke.~ words: Visceral leishmaniasis: Leishrnania donovani: Surface antigens; cDNA cloning; mRNA: Hybrid selected translation Introduction Leishmania donovani is a monoflagellate pro- tozoan parasite which has a biphasic life cycle, al- ternating between an intracellular non-motile stage in mononuclear phagocytes in the verte- brate host and an extracellular flagellated form in the insect vector. In man, L. donovani causes visceral leishman- iasis or kala-azar which can be fatal if left un- treated. The differential diagnosis of the disease is not straight-forward and parasitological diag- nosis of visceral leishmaniasis relies on the detec- tion of amastigotes in either bone marrow or Correspondence address: Dr. M.L. Chance, Department of Parasitology, Liverpool School of Tropical Medicine, Pem- broke Place. Liverpool L3 5QA. U.K. Abbreviations: SDS-PAGE, sodium dodecyl sulphate poly- acD'lamide gel electrophoresis: 1135, Tris-buffered saline; TIE. Tris'EDTA; SSC, saline sodium citrate: Denhardt's, PVP. Fi- coil and bovine serum albumin: PIPES, 1,4-piperazinedieth- anesulphonic acid: PMSF, phenylmethylsulphonyl fluoride spleen cell biopsies, both of which require inva- sire procedures which are not without risk to the patient. Serological tests on the other hand are convenient and extremely sensitive, but suffer from a lack of specificity [1]. For example, a re- cent report by Pappas et al. [2] described the use of an improved, extremely sensitive, dot-enzyme linked immunosorbent assay for diagnosing vis- ceral leishmaniasis, but still obtained a 20% false positive background from trypanosomiasis pa- tients and also from healthy Africans. The most important cross-reaction with Leishmania occurs with trypanosomes (African or South American). However, other cross-reactions with Leishmania have been described using sera from patients with leprosy, tuberculosis and malaria [3]. Our long term aim is to improve the specificity of currently used serological tests for visceral leishmaniasis. In this report, we describe prelim- inary results of the characterisation of surface an- tigens of both morphological forms of L. dono- vani and the identity of those surface peptides that 0166-6851,'87'$03,50 ~ 1987 Elsevier Science Publishers B,V. (Biomedical Division)
-
Upload
stephen-heath -
Category
Documents
-
view
215 -
download
1
Transcript of Cloning of a gene encoding the immunodominant surface antigen of Leishmania donovani promastigotes

Molecular and Biochemical Parasitology. 23 (1987) 211-222 211 Elsevier
MBP 00788
Cloning of a gene encoding the immunodominant surface antigen of Leishmania donovani promastigotes
Stephen Heath 1, Michael L. Chance 1, Marcel Hommel: and Julian M. Crampton 3 ~Department of Parasitology. "-Department of Troptcal Medicine. and 3Wolfson Molecular Genetics Untt. Liverpool School of
Tropical Medicine. Liverpool. U.K.
(Received 19 September 1986; Accepted 3 December 1986)
This study describes the characterisation of externally oriented surface peptides of both morphological forms of Leishmania don- ovani, the causative agent of visceral leishmaniasis (kala-azar). Using t-'-~i surface labelling techniques and peptide extraction in the detergents Triton X-I(KI and Triton X-I 1-, t. a major iodinable promastigote peptide at 63 kDa or 65 kDa (depending on de- tergent used) was identified. This peptide was demonstrated to be the immunodominant membrane peptide of L. donovani and ~as strongly recognised by human sera from parasitologically confirmed cases of kala-azar. This peptide was not demonstrated on the surface of tissue amastigotes, although in vitro translations of poly(A*) RNA from both promastigotes and amastigotes dem- onstrated that both forms possessed mRNA that directs the synthesis of a 63 kDa peptide. It is suggested therefore that in amas- tigotes this peptide ma~ be a processed antigen. We also report the isolation of a recombinant cDNA clone in the bacteriophage vector h gtl0 which encodes a 63 kDa polypeptide that is recognised b.~ human kala-azar sera. It is proposed that this surface peptide could be used in a specific immunodiagnostic test for leishmaniasis.
Ke.~ words: Visceral leishmaniasis: Leishrnania donovani: Surface antigens; cDNA cloning; mRNA: Hybrid selected translation
In t roduc t ion
Leishmania donovan i is a monof l age l l a t e p ro- tozoan pa ras i t e which has a b iphas ic life cycle, al- t e rna t ing b e t w e e n an in t race l lu la r non-mot i l e s tage in m o n o n u c l e a r phagocy t e s in the ver te- b ra te host and an ex t r ace l l u l a r f lagel la ted form in the insect vec tor .
In man , L. donovan i causes visceral l e i shman- iasis or ka l a - aza r which can be fatal if left un- t r ea t ed . The d i f fe ren t ia l d iagnos i s of the d isease is not s t r a igh t - fo rward and pa ras i to log ica l diag- nosis of visceral l e i shmanias i s re l ies on the de tec- t ion of a m a s t i g o t e s in e i the r bone mar row or
Correspondence address: Dr. M.L. Chance, Department of Parasitology, Liverpool School of Tropical Medicine, Pem- broke Place. Liverpool L3 5QA. U.K.
Abbreviations: SDS-PAGE, sodium dodecyl sulphate poly- acD'lamide gel electrophoresis: 1135, Tris-buffered saline; TIE. Tris'EDTA; SSC, saline sodium citrate: Denhardt's, PVP. Fi- coil and bovine serum albumin: PIPES, 1,4-piperazinedieth- anesulphonic acid: PMSF, phenylmethylsulphonyl fluoride
sp leen cell b iops ies , both of which requi re inva- s i re p roc e du re s which are not wi thout risk to the pa t ien t . Sero logica l tests on the o t h e r hand are conven ien t and e x t r e m e l y sensi t ive , but suffer from a lack of specifici ty [1]. Fo r e x a m p l e , a re- cent r epor t by Pappas et al. [2] desc r ibed the use of an i m p r o v e d , ex t r eme ly sensi t ive , d o t - e n z y m e l inked i m m u n o s o r b e n t assay for d iagnos ing vis- ceral le ishmanias is , but still o b t a i n e d a 20% false posi t ive b a c k g r o u n d from t rypa nosomia s i s pa- t ients and also from hea l thy Afr icans . The most impor t an t c ross - reac t ion with Leishmania occurs with t r y p a n o s o m e s (Af r i can or South A m e r i c a n ) . H o w e v e r , o t h e r c ross - reac t ions with Leishmania have been described using sera from patients with leprosy , tubercu los i s and ma la r i a [3].
O u r long te rm aim is to improve the specif ici ty of cur ren t ly used serologica l tests for visceral le ishmanias is . In this r epor t , we desc r ibe p re l im- inary results of the cha rac te r i sa t ion of surface an- t igens of both morpho log ica l forms of L. dono- vani and the identity of those surface pept ides that
0166-6851,'87'$03,50 ~ 1987 Elsevier Science Publishers B,V. (Biomedical Division)

212
are strongl.~ immunogenic in the course of a nat- ural infection but which do not exhibit an~ sig- nificant cross reaction with heterologous sera. We also report the identification and isolation, using hybrid-selected translation techniques, of a re- combinant clone coding for the immunodominant 63 kDa promastigote surface protein.
Materials and Methods
Parasites. Promastigotes of L. donovani (MHOM, ET,,'67,'H u3, LV9, M H O M,'F R"00,' KA; LV 114, MHOM,'B R,'74,'M2682: LV474, MHOM,'IN,'00,' Naresh-Rai; LV636, MHOM,'SD,'61,'2s; LV710 I were grown in H O - M E M containing 6ok heat in- activated fetal calf serum at 26°C [4]. L. dono- vani amastigotes (LV9) were isolated from in- fected cotton rat (Sigmodon hispidus* spleens and purified using the method of Hart et al. [5].
Sera. Sera were pooled from 15 parasitologicall.~ confirmed cases of kala azar from the surve~ of the Tana river region.
Radiolabelling and detergent extraction of imra - siw6. Late log-phase promastigotes xsere xsashed three time,~ in Tris-buffered saline (TBS: 511 mM Tris-HCI 150 mM NaCI pH 7.4J at 4:'C and the fi- nal pellet taken up in 0.5 ml of TBS. Promasti- gores and purified amastigotes were surface io- dinated via the I O D O G E N technique [6,7] using 1~0 ~Ci of carrier-free ~-'SNaI (Amershaml for lql rain on ice. Parasites xsere washed several times in ice cold TBS to r e m m e unincorporated ~z:I and then kept on ice until detergent extracted. Para- site proteins ~e re extracted ~sith either Triton X- 100 (TX-1001 [8] or Triton X-114 (TX-1141 [9]. All detergent solutions contained a cocktail ot protease inhibitors as follows: 2 mm phen.~lme- th.~lsulphonyl fluoride (PMSFJ, 0.2 mM tosxl-t- phen~lalanine ch[orometh.~l ketone, 0.2 mM to- s.~l-t-I,~sine chloromethyl ketone and 2 mM EDTA. When using TX-I14, a phase separation x~as carried out at 3WC and the detergent phase comaining integral membrane proteins xsa~ sep- arated from h~drophilic components bx centrifu- gation using the method of Bordier et al. [1~]. Detergent extracted proteins xsere separated b~ sodium dodecvl sulphate polyacrylamide gel elec-
trophoresis (SDS-PAGE) [111 under reducing conditions and the gels ssere xacuum dried and autoradiographed at -TWC using Fuji RX film and intensifying screens [12]. Detergent ex- tracted proteins were mixed xsith 15 ~,1 of Kenvan kala-azar sera and immune complexes precipi- tated with protein-A Sepharose [13,14] and ana- lysed bx SDS-PAGE.
Isolation of RNA. Total RNA from both pro- mastigotes and amastigotes of L. donovani (L\'91 ~as isolated using the guanidinium thioc~a- nate,CsCI procedure [151 . PolytAI + RNA was purified from total RNA b~ tx~o passages over an ol igoldTI cellulose column [16] and then ethanol precipitated and taken up in sterile distilled x~ater and stored at -70 :C .
Fractionation ol" tuRN,4 attd in vitro translatton. RNA ~as fractionated on a denaturing 10-411% linear sucrose gradient in 50~ formamide, 10 mM T r i s l mM E D T A {TE) pH 7.5 and I).2% SDS. Total RNA was taken up in denaturing buffer at a concentration of not less than 200 IJ-g ml ~ and la,~ered onto a 15 ml gradient in polyallomer cen- trifuge tubes and centrifuged for 16 h at 1511000 ," g~, at 25"C in a Sor~all SW41 rotor. The gra- dient ~as then fractionated into 0.5 ml fraction,~ and their absorbance at 260 nm ~as monitored. RNA ~as precipitated from each of the fractions ~ith 2.5 vol of ice cold absolute ethanol I -2~1:CI after adding NaCI to 200 raM. The precipitate ~as washed ~ith ice cold 70t'; ethanol, dried under ~acuum and then taken up in 10 p.I o f T E pH 7.5. ,An aliquot of each gradient fraction ~as in-~ itro translated using the rabbit reticulocxte hsate s~-. tem (Amersham) supplemented with [':S] methionine (Amersham) as described b.~ Pelham and Jackson [17]. Translation products were sep- arated on SDS-PAGE gels and visualised b~ fluo- rograph.~ at -70~C after incubating the gels in Amplif3' (Amersham)
Isolation or" DNA. High molecular weight DNA ~as isolated using the method of Bingham et al. [18]. Briefly, parasites were homogenised in 0.5c; TX-100 and centrifuged to obtain a nuclear pel- let. The pellet was lvsed with 2% Sarkosvl and the h'sate centrifuged to equilibrium in CsCI. DNA

was recovered by fractionating the gradient and dialysing the peak fractions extensively against TE pH 7.5.
Construction of cDNA clones derived from Leish- mania donovani RNA. cDNA was constructed from a poly(A) ~- RNA template using AMV re- verse transcriptase (P&S Biochemicals) and an oligo(dT)t2_18 primer following the protocol of Huynh et al. [19]. Briefly, after separating the first strand cDNA from the mRNA template by heat and alkali denaturation, double stranded cDNA was prepared using DNA polymerase I. This re- action used the 3' terminus of the first strand as a primer [20,21]. The double stranded cDNA was then treated with EcoRl methylase to methylate and protect EcoRl cleavage sites within the cDNA. from digestion with EcoRl. After brief treatment of the cDNA with DNA polymerase I to increase the number of flush ended double stranded cDNA molecules [22], EcoRI linkers were ligated onto the ends. cDNA was then li- gated with EcoRI-digested h gtl0 and the ligation mixture was packaged in vitro [23].
Screening of cDNA clones by hybridisation with single stranded [~2P]cDNA. Approximately 10000 clones from a promastigote cDNA library were screened in duplicate with a single stranded [3-'P]cDNA probe prepared from either total L. donovani poly(A) ÷ RNA or fractionated RNA. Unlabelled dCTP was replaced with [3-~P]dCTP in the reverse transcriptase reaction in order to pro- duce a probe of high specific activity. Clones that hybridised strongly to the probe were picked, plaque purified and rescreened [24]. After brief chloroform treatment, phage from each clone was placed in an individual well of a 96-well micro- titre plate. An array of 48 strongly hybridising clones representing highly abundant mRNA spe- cies were stamped out onto an agar plate con- taining host cells (Escherichia coli C600) using an 8×6 replica plating device. The array was repli- cated onto nitrocellulose (Schleicher & Schuell) for further analysis by hybridisation. All hybri- disations were carried out at 42°C in the presence of 50% formamide, 5 × Denhardt 's (100 × Den- hardt's = 2% (w/v) PVP, 2% (w/v) FicoU and 2% (w,,'v) bovine serum albumin). 5 x SSC (20 × SSC
213
= 3 M NaCl/0.3 M Na 3 citrate), 0.1% SDS and I(X) I~g ml -~ sheared salmon sperm DNA. Duplicate nitrocellulose filters were hybridised for 24-48 h and then washed in I×SSC + 0.1c? SDS at 42~C for 30 rain followed by washes of increasing strin- gency at 65°C. Approximately 10 ~ cpm of [3-'P]cDNA per ml of hybridisation buffer ~as used in all hvbridisation reactions.
Hybrid selected translation of mRNA. mRNA se- lection was carried out using a modification of the procedure by Parnes et al. [25]. Briefly, 10 ~g of cloned DNA in TE pH 7.5 was denatured by boiling and addition of alkali and then neutral- ised with 1.4 M NaCI, 0.15 M sodium citrate, 0.25 M Tris-HCl (pH 8.0), 0.25 M HCI. The DNA was immediately bound to nitrocellulose by slow fil- tration (1 ml min -l) through a vacuum minifold (S&S) and baked at 80°C for 2 h. Filters were prehybridised for 10 rain at room temperature in hybridisation buffer (50% formamide, 0.9 M NaCI, 0.2% SDS, 1 mM E D T A and 20 mM 1,4- piperazinediethanesulphonic acid (PIPES) pH 6.4). The filters were then incubated at 37°C for 6 h in hybridisation buffer containing 40 ~g of poly(A) + RNA after which they were washed ex- tensively in 50%. formamide, 20 mM NaCI, 8 mM sodium citrate, 1 mM ED TA and 0 . 5 ~ SDS at 37°C. Hybridised RNA was eluted in 300 ~1 of 1 mM ED TA by boiling for 1 min and then snap freezing in a solid CO,/ethanol bath. After thaw- ing at room temperature the filters were removed and 10 Ixg of calf liver tRNA and sodium acetate to 200 mM were added. Eluted RNA was ex- tracted with phenol/chloroform and then precip- itated with 2.5 vol of ice cold absolute ethanol ( -20°C). RNA was recovered by centrifugation at 12000 × ga, for 10 min, washed twice in 95% ethanol and dried under vacuum. The RNA pel- let was taken up in 5 ~1 of sterile distilled water and in vitro translated.
Southern blotting of L. donovani genomic DNA. L. donovani promastigote total DNA (10 ~g) was digested with 15 units of either BamHI or EcoRl at 37°C for 3 h, separated on a 1% agarose gel and transferred to nitrocellulose using the Southern blot technique [26]. Blots were hybridised with either a [32PJcDNA probe reverse transcribed

214
from an enriched m R N A sucrose gradient frac- tion or cloned D N A inserts 3_'p labelled by nick translation [27].
Results
Identification or" 12Sl-/abelled membrane-associ- ated peptides. Peptide profiles of TX-100 ex- tracted L~sI-labelled L. donovani (LV9) promas- tigotes and amastigotes revealed the presence of a major protein at 65 kDa in promastigotes which appeared to be absent from the surface of amas- tigotes (Fig. la). Immunoprecipi ta t ion of surface associated proteins by human kala-azar sera demonstra ted that the promastigote 65 kDa and a 57 kDa protein from amastigotes were immu- nogenic in the course of a natural infection (Fig. lb). Extraction of different geographic isolates of L. donovani promast igotes in TX-114, revealed strain specific differences in the surface expres- sion of integral membrane proteins (Fig. 2A). Significant differences in integral membrane pro-
LV9 L V 9 p A
,4 ,g
'q45
'125
,q17
~12
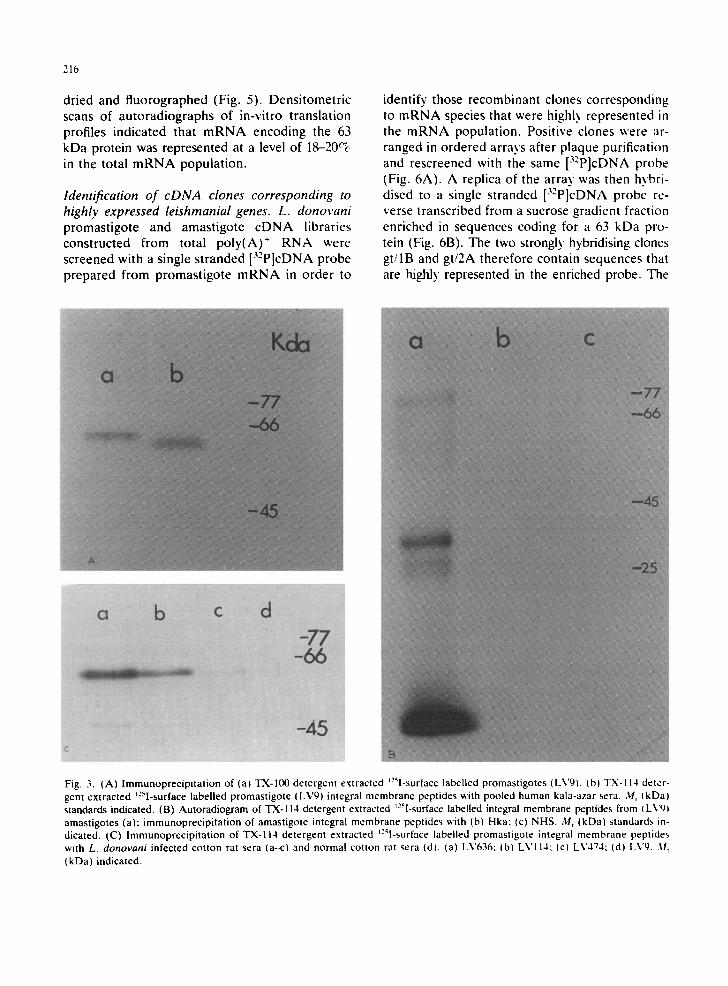
rein profiles of Indian (LV636), Sudanese (LV710), South American (LV474) and Mediter- ranean isolates ( L V I I 4 ) were demonstrated. However , Mediterranean and South American profiles ~ere identical to each other. All isolates possessed a 63 kDa protein, but this was shown to be only a minor protein in Sudanese isolate. Reaction of TX-114 detergent extracts with hu- man kala-azar sera precipitated the major 63 kDa integral membrane protein and a minor compo- nent at 45 kDa, although no proteins were recog- nised from the Sudanese isolate (Fig. 2B). The difference in the rate of migration of TX-100 and TX-I14 extracted promastigote membrane pep- tides in polyacrylamide gels x~as demonstrated b~ running the different samples side b~ side (Fig. 3A). This difference was found to be approxi- mately 2 kDa.
Comparison of amastigote TX-1IX) (Fig. la) and TX-114 peptide profiles (Fig. 3B) revealed that the immunodominant amastigote membrane-as- sociated protein at 57 kDa does not partition into
9Pr
77_
9am 9 Pt 9am
W
Hka H~ NHS NHS
Fig. 1 la) Autoradnogram of SDS-PAGE ~Z~l-surIace la- belled pepude profile of promastigotes I P) and amasugotes (Am) of L. donovant (LV9) extracted with the detergent TX- 100. M, {kDa) standards indicated on the ie[t. (b) Immuno- precipitation of TX-I(~] detergent extracted L'~I-labe[[ed sur- face peptides of L donovani (iX'q) promastigotes (P~ and amastigotes (Am) by pooled human kala-azar sera (Hka) and normal human sera INHS).

the organic phase of TX-114, lmmunoprecipi ta- tion of TX-114 detergent extracted amastigote in- tregral membrane proteins with human kala-azar sera, did not demonst ra te any recognition of membrane components .
We have also demonst ra ted that sera from cot- ton rats initially infected with only the amastigote stage of L. donovani also recognised the promas- tigote 63 kDa integral membrane protein (Fig. 3C), suggesting that the 63 kDa promastigote and the 57 kDa amastigote protein either have com- mon antigenic determinants or are the same pro- tein. We have also not been able to demonstrate any significant recognition of immunodominant amastigote and promastigote membrane compo- nents by human sera from clinically diagnosed cases of tuberculosis, leprosy, syphilis and ma- laria even when SDS gels had been autoradi- ographed for extended periods (i.e. >4 weeks).
Fractionation and in-vitro translation o f m R N A . A prerequisite to the preparat ion of c D N A which
215
is representative of the total m R N A population is the isolation of biologically active mRNA. Poly(A) + RNA from both promastigotes and amastigotes of L. donovani (LV9) was isolated, in-vitro translated and the products analysed on SDS-PAGE gels (Fig. 4A). Qualitatively, there was shown to be very little difference in transla- tion profiles of both morphological forms of L. donovani. Surprisingly, when amastigote m R N A was in-vitro translated a protein at 63 kDa was observed. This was also recognised along with a promastigote 63 kDa protein, by pooled human kala-azar sera (Fig. 4B). In order to prepare [3-'P]cDNA probes enriched in sequences of in- terest, m R N A was fractionated on denaturing su- crose gradients. The fraction(s) containing the 63 kDa m R N A was identified by its ability to direct the synthesis of a 63 kDa protein which could be immunoprecipi tated by pooled human kala-azar sera. Aliquots of fractions between 9 and 30 s were in-vitro translated and the resulting proteins characterised on SDS-PAGE gels which were
kd
o77
-25
i14 474 636 710
kd
qp77
aa56
!14 474 636 710
~ l l m ~ ~ . . . , -~_~ -~ ~
Fig. 2. (A) Autoradiogram of SDS-PAGE 12Sl-surface labelled integral membrane peptide profile of different geographic isolates of L. donovani promastigotes. 114=Mediterranean: 474=South American: 636= Indian: 710=Sudanese. Integral membrane pep- tides were extracted into the organic phase of the detergent TX-114. 3,1, (kDa) standards indicated. IB) Immunoprecipitation of TX- 114 detergent extracted ~zSI-labelled integral membrane peptides of different geographnc isolates of L donovani promastigotes with pooled human kala-azar sera. 114= Mediterranean isolate: 474= South American isolate: 636= Indian isolate: 710= Su- danese isolate. M~ (kDa) standards indicated.
216
dried and f luorographed (Fig. 5). Dens i tometr ic scans of au to rad iographs of in-vitro translation profiles indicated that m R N A encoding the 63 kDa protein was represented at a level of 18-20% in the total m R N A popula t ion.
Identification of cDNA clones corresponding to highb' expressed leishmanial genes. L. donovani promast igote and amast igote c D N A libraries const ructed from total po ly (A) ÷ R N A were screened with a single s t randed [3Zp]cDNA probe prepared from promast igote m R N A in o rder to
identify those recombinant clones cor responding to m R N A species that were highly represented in the m R N A populat ion. Positive clones were ar- ranged in ordered arrays after plaque purification and rescreened with the same [3~p]cDNA probe (Fig. 6A). A replica of the array was then hvbri- dised to a single s t randed [32P]cDNA probe re- verse transcribed from a sucrose gradient fraction enriched in sequences coding for a 63 kDa pro- tein (Fig. 6B). The two strongly hybridising clones gt,"lB and gt/2A therefore contain sequences that are highly represented in the enriched probe. The
a b c d
-77 -66
-45
Fig. 3. (A) lmmunoprecipltation of (a) TX-100 detergent extracted ~:5l-surface labelled promastigotes (LV9). (b) TX-114 deter- gent extracted t:51-surface labelled promastigote (LV9I integral membrane peptides with pooled human kala-azar sera. Mr (kDa) standards indicated. (B) Autoradiogram of 1"X-114 detergent extracted t-'sI-surface labelled integral membrane peptides from I LVqj amastigotes (a) immunoprecipitation of amastigote integral membrane peptides with (b¿ Hka; (c) NHS. M, (kDa) standards in- dicated. (C) lmmunoprecipitation of TX-II4 detergent extracted t:Sl-surface labelled promastigote integral membrane peptides with L. donovani infected cotton rat sera (a--c) and normal cotton rat sera (d). (a) LV636; (b) LVI14: (c) LV4"74; (dl Lvg. ),I T (kDa) indicated.

217
Kd
77- 66 -
4 5 -
2 0 -
Kd
7?- 68-
2 0 -
I~.lS I'lKa
06 0-6 G2 G6 ~2 06
. . . . 7
I~ B Aim I~ ~ ~ I~ i~.
Fig. 4. (A) SDS-PAGE profile of :'SS-labelled in-vitro translation products of polylA)* RNA from L. donovant (LV9) amastigotes (Am) and promastigotes (Pr). Figures above each track represent the amount of input pol}(A) ~ RNA in i.tg. 3.IT (kDa) standards indicated. (B) immunoprecipitation of -~SS-labelled in-vitro translation products of poly(A)" RNA from L. donovant (LV91 amas- tigotes (Am) and promastigotes I Pr) with pooled human kala-azar sera (Hkal and normal human sera (NHS) M, (k Da) standards indicated•
Kd
77-
4 6 -
Fig. 5. SDS-PAGE profile of 35S-labelled in-vitro translation products of sucrose gradient fractionated promasugote [ LV9) poly(A)* RNA. The top part of the gradient is indicated (Top) along with Mr (kDa) standards.
insert from gt,,'2A was cut out using EcoR1 and 32p labe l led by nick t rans la t ion . W h e n this p robe was hybr id i sed to the 8 x 6 a r ray , it also hybri- d ised to clone gt.,'lB (resul t not shown) indica t ing that the two r ecombinan t c lones con t a ined simi- lar sequences . C lone gt, '2A, which con ta ined an insert of 1.3 kb, was t he re fo re used for fur ther s tudies . In o r d e r to e l imina te the poss ibi l i ty that these two c lones con ta ined sequences corre- spond ing to tubul in m R N A species , the same ar- ray was hybr id i sed to an a lpha plus be ta tubul in p r o b e der ived f rom Trypanosoma cruzi ( suppl ied by J .M. C r a m p t o n ) . None of the c lones in the ar- ray hybr id i sed to this p robe (resul t not shown) . Sou the rn blot analysis of the 63 k D a enr i ched single s t r anded [32P]cDNA probe to d iges ted L. donovani genomic D N A (Fig. 7), d e m o n s t r a t e d that it hybr id i sed s t rongly to two d iscre te E c o R I f ragments at 5.9 kb and 8.3 kb.

218
A
0
j,
•
° I1
t i l
Fig. 6. (A) An 8x6 array of recombinant phage plaques picked from a primary screen of a promastigote L. donovant cDNA libraD, in h gtl0 hybridised with a [L'P]cDNA probe prepared from promastigote total pol.~lA) * RNA. (B) An 8x6 array of plaque purified phage clones from a L. donovani promastigote cDNA librar} hybridised with a [3-'P]cDNA probe enriched for sequences coding for the 63 kDa promastigote peptide. Clone gr2A was used in hybrid selected translation experiments.
Identification of proteins encoded by cDNA clones. In order to confirm the identity of proteins en- coded by cDNA clones, DNA was isolated from individual clones and used to select out mRNA species from total RNA. Selected mRNAs were then in-vitro translated in the rabbit reticulocyte system, separated on SDS-PAGE gels and fluo- rographed as previously described (Fig. 8). Clone gt/2A, which hybridised strongly to the cDNA probe enriched in mRNA sequences coding for the 63 kDa protein, selected out mRNA which directed the synthesis of a protein that had an ap- parent molecular mass of 63 kDa and was im- munoprecipitated by pooled human kala-azar sera.
Discussion
This stud}' set out to identify and characterise the surface antigens of promastigotes and amas- tigotes of L. donovani which are recognised by the host during a natural infection and which do not cross-react with heterologous sera. Previous re- ports have produced a confusing array of peptide profiles for the promastigote stage of the parasite
[7,28.29]. The differences observed between lab- oratories has undoubtedly been due to the method of detergent extraction, the actual detergent used and the way in which the parasites were handled (i.e. temperature and the presence of protease inhibitors). In extreme cases [28], the surface peptide profiles have appeared identical to the patterns obtained from limited proteolysis of de- tergent extracted surface components [30].
This study demonstrated that externally dis- posed integral membrane peptide profiles of a number of different geographic isolates of L. donovani promastigotes were significantly differ- ent. Indian and Sudanese isolates were shown to be clearly different to the South American and Mediterranean strains and also different to each other. However, there appeared to be no differ- ence in profiles between South American and Mediterranean strains. A major common feature of all strains studied was the presence of a mem- brane protein which had an apparent molecular mass of 63 or 65 kDa. A surface protein with this M r has been shown to be present in all leishman- ias studied so far [7,9,13,29]. This protein was re- cognised by pooled human kala-azar sera in all

21.2-
m
n
4-B-
" - : s -
219
a p p a r e n t mo lecu l a r mass of the i m m u n o d o m i - nant m e m b r a n e p ro te in was 65 k D a and was also recognised by human ka la -aza r sera . W e suggest that this d i f fe rence in the o b s e r v e d ra te of migra- tion of the m a j o r p roma s t i go t e m e m b r a n e pro-
a b c d
Fig. 7. BamHl and EcoRl digests of total DNA extracted from L. donovam (LV9) promastigotes. Digests were run in dupli- cate, Southern blotted and probed with single stranded [~2P]cDNA reverse transcribed from an enriched mRNA su- crose gradient fraction directing the synthesis of a 63 kDa peptide. Size markers are EcoRI digested. DNA in kb pairs.
s t ra ins s tud ied , with the excep t ion of a Sudanese isolate (LV710). The reason for this apparen t lack of recognit ion is as yet still unresolved, but it may be that in this pa r t i cu l a r s t ra in the p ro te in occurs in a more heavi ly g lycosy la t ed form which may shield the p e p t i d e ep i t ope ( s ) . The poss ib i l i ty of the m a j o r p ro t e in possess ing d i f fe ren t sacchar- ides is cons i s ten t with ou r f indings that there are signif icant d i f f e rences in the level of b ind ing of concanava l in A to the 63 k D a m e m b r a n e p ro te in be t ween d i f fe ren t g e o g r a p h i c i sola tes of L. don- ovani ( resul ts not shown) . The i m m u n o d o m i n a n t na tu re of the 63 k D a m e m b r a n e p ro te in de- scribed in this s tudy is in agreement with the work of Etges et al. [9], who, in add i t i on , d e m o n - s t ra ted s t ruc tura l s imi la r i t i es in this surface mem- b rane p ro t e in b e t w e e n cer ta in species . We also d e m o n s t r a t e d that when ex t r ac t ion of p romas t i - gotes was achieved using the de tergent TX-100 the
Fig. 8. Identification of a recombinant clone containing se- quences complementary to mRNA that directs the synthesis of a 63 kDa peptide. Recombinant bacteriophage DNA was denatured and bound to nitrocellulose. The DNA filters were hybridised to promastigote (LV9) poly(A) ~ RNA, washed and the selected RNA was translated in a reticulocyte I.vsate. Tracks on the autoradiogram are (a) ~sS in-vitro translation profile from 40 ~g of promastigote poly(A)" RNA (this aas used in hybrid selected translations): (b) ~sS in-vitro transla- tion profile from 4 I.tg of promastlgote poly(A) ÷ RNA: (c) ~S- labelled in-vitro translated hybrid selected mRNA: (dl im- munoprecipitatlon of ~SS-labelled in-vitro translated products from (c) ~ith pooled human kala-azar sera.

220
tein in SDS gels under reducing conditions could be due to the possession of a hydrophilic com- ponent which does not partition into the organic phase of the detergent TX-114. A recent report by Bouvier et al. [31] showed that a hydrophilic form of the 63 kDa membrane protein could be generated during a purification procedure ~hich had an apparent molecular mass slightly greater than that observed for the amphiphilic protein. In this stud}' the 63 kDa promastigote membrane protein was shown to be absent from TX-I14 amastigote protein profiles. In addition, immu- noprecipitation of TX-114 extracted amastigotes with human kala-azar sera demonstrated that none of the integral membrane proteins were re- cognised. Results presented here also demon- strate from TX-100 detergent extracts of amasti- gotes of L. donovani that the 65 kDa protein was not present on the surface of amastigotes. This result is contrary to the findings of Colomer- Gould et al. [32]. A recent report by Lawrence and Rober t -Gero [33] demonstrated the presence of heat shock proteins at the surface of promas- tigotes x~hen in-vitro cultures are temperature shifted from 26 to 37°C. They also suggest that this may work in reverse (i.e. when amastigotes are subjected to temperatures approaching that used for in-vitro culture of promastigotes) and haxe demonstrated that temperature shifts can alter the protein biosynthesis pattern of tubulin in amas- tigotes. In this study immunoprecipitation of TX- 100 detergent extracts of amastigotes with human kala azar sera precipitated a single protein with an apparent molecular mass of 57 kDa. Results from immunoprecipitation experiments using in- fected cotton rat sera may suggest that the pro- mastigote 63 kDa and amastigote 57 kDa surface membrane proteins have cross-reactive epitopes. Bouvier et al. [31] demonstrated that complete deglycosylation of the purified promastigote 63 kDa protein with trifluoromethanosulfonic acid increased the migration rate of this protein in SDS-PAGE gels which then ran with apparent molecular mass of 57 kDa. The single identity of the two proteins in this study was confirmed us- ing limited proteolysis. Also, a recent report b~ Chang and Chang [34] demonstrated an increase in electrophoretic mobility on SDS-PAGE gels of a 63 kDa Leishmania surface membrane protein
after treatment with endoglycosidase H. In consideration of our work and the findings
of Bouvier et al. [31], we suggest therefore that the promastigote 63 kDa protein ma.~ be present in a completely deglycosylated form on the sur- face of amastigotes. Our results with TX-114 ex- tracted amastigotes would also suggest that the deglycosylated form is a hydrophilic membrane protein and as such does not partition into the or- ganic phase of TX-114. This would also explain the lack of recognition of amastigote TX-114 ex- tracted integral membrane proteins by human kala-azar sera. Recently, Etges et al. [35] dem- onstrated in L. major that the immunodominant surface protein is a protease which has the ability to clip off the lipid anchor of the amphiphilic 63 kDa protein producing a soluble form of this molecule.
The dominant nature of the 63 kDa protein was also indicated from in vitro translations of poly(A) * RNA from promastigote and amasti- gotes. Surprisingly little difference was demon- strated in translation profiles between the two morphological forms. The fact that the 63 kDa protein was translated from amastigote mRNA and immunoprecipitated by human kala-azar sera may indicate that in amastigotes this protein undergoes some processing upon being inserted into the outer membrane. By analog,,' ~ith other membrane glycoproteins it is possible that nas- cent polypeptides are co-translationall~ glycos}- lated and this would explain the similar migration rate of the promastigote 63 kDa protein in in-~i- tro translation products and integral membrane peptide profiles.
From our surface labelling studies, we dem- onstrated that the promastigote 63 kDa protein was not significantly recognised by sera from pa- tients with leprosy, tuberculosis, syphilis and ma- laria. We suggested therefore that this membrane protein may be a good candidate for use in a spe- cific anti-leishmanial antibody detection system, particularly when differential diagnosis from other febrile infections with marked hepatosplenomeg- al.v is requited, e.g. malaria.
In order to identify and clone the gene coding for the 63 kDa protein our approach was to ini- tially construct a cDNA library in the bacterio- phage vector gtl0 and select out those clones

which strongly hybridised to a single stranded [3:P]cDNA probe. Our mRNA in-~itro transla- tions had demonstrated that promastigote poly(A) + RNA directing the synthesis of the 63 kDa protein was highly represented in the total RNA population. The library consisted of 50000 clones of which approximately 30% gave good hybridisation signals to a single stranded [3-'P]cDNA probe. Further studies using gt.'2A in- vol~ing hybrid selected translation and immtmo- precipitation experiments demonstrated unequi- vocall~ that this clone contained sequences corresponding to the 63 kDa gene. This approach to identifying specific cloned genes was used bx Hoeijmakers et al. [36] to isolate cDNA clones complementary to trypanosome variant surface glycoprotein. This procedure has recently been scaled up to enable large numbers of clones to be screened successfully using a minimum number of in-vitro translations [37]. In a recent report, Sheppard and Dwyer [38] successfully isolated recombinant h g t l l clones expressing leishmanial surface proteins (;'fir 4000 to 22000). However, in contrast to this report, the identitx of the pro- teins encoded by the leishmanial inserts x~as not ascertained.
The ultimate aim of these cloning experiments is to obtain expression of the gene coding for the 63 kDa protein in E. colt. Since a direct immu- nological screen of an expression library would
References
1 Voller. A. and De Savigny, D. (1981) Diagnostic serolog.,, of tropical parasitic diseases. J. lmmunol. Methods 46, 1-29.
2 Pappas, M.G., Hajkowskh R., Tang, D.B. and Hock- meyer, W.T. (1985) Reduced false positive reactions m the dot-enzyme-linked immunoadsorbent assay for human vis- ceral leishmaniasis. Clin, lmmunol. Immunopathol. 34. 392-390.
3 Ranque, P., Quilici, M., Dunan, S. and Assadourian. Y. (1969) Etudes des parent6s antig~,niques entre Leishmania tropwa. Leiahmania donovani et divers autres flagellds. Eighth Int. Congr. Trop. Med. Malaria. Teheran. pp. 1178--1182.
4 Berens, R.L.. Brun, R. and Krassner, S.M. (197~) A sim- ple monophasic medium for axenic culture of haemofla- gellates. J. Parasitol. 62,300-365.
5 Hart. D.T., Vickerman. K. and Coombs. G.H. (1981) A quick simple method for purifying Letshmania mericana amastigotes in large numbers. Parasitolog} 82,345--355.
221
require a positive expressor to have an insert in the correct orientation and translational reading frame, it is highly likely that a primary screen of this nature would overlook a large number of clones containing the specific gene.
The insert from this clone is now being used in our laboratory as a probe to isolate larger gen- omic clones from a library constructed in h gt.wes. We are also in the process of subcloning this in- sert into the expression vector h g t l l which will enable us to screen directly for expression using either human kala-azar sera or a rabbit anti-pro- mastigote serum. An.,,' expressed fusion proteins will be evaluated for use in an antibody detection system using human kala-azar sera. As the 63 kDa protein has been shown to be present in Leish- mania, sera from other clinical manifestations will also be investigated for specific antibody. Also, expressed proteins will be assessed for their abil- it}' to produce a protective immune response in experimental animals.
Acknowledgements
We are very grateful to Dr. E. Devaney for useful suggestions in the preparation of this man- uscript and to Dr. D. Smith for supplying serum. This work was supported by a Medical Research Council project grant awarded to M.L.C. . M.H. and J.M.C.
6 Fraker, P.J. and Speck, J.C. (1978l Protein and cell mem- brane iodination with a sparingl) soluble chloramide, 1,3,4.6-tetrachloro-3a-6a-diphen.~ Igl} couril. Biochem. Biophys. Res. Commun. 80, 849-857.
7 Gardiner, P.R. and Dw)er, D.M. (1983) Radloiodination and identification of externally disposed membrane com- ponents of Leishmania tropica. Mol. Biochem. Parasitol. 8, 283--295.
8 Ho,~ard, R.J.. Kaushal, D.C. and Carter. R. (1982) Ra- dio.odination of parasite antigens with 1,3.4,6-tetrachloro- 3a-6a-diphenylgl),couril (IODOGEN)-studles with zygotes of Plasmodium galhnaecium. J. Protozool. 29. 114-117.
9 Etges. R.J., Bouvier, J., Hoffman. R. and Bordier, C. (1985') Evidence that the major surface proteins of three Leishmania species are structurally related. Mol. Biochem. Parasitol. 14, 141-149.
10 Bordier. C. ( 1981 ) Phase separation of integral membrane proteins in Triton XI14 solution. J. Biol. Chem. 256(4). 1 b04-10(17

11 Laemmli . U.K. (19701 Cleavage of structural proteins during assembl.~ of the head of bacteriophage T4. Nature 227,680-685.
12 Swanstrom, R. and Shank. P.R. (1978) X-ray intensif,,ing screens greatly enhance the detection b,,, autoradiograph~ of the radioactive isotopes -~-~P and ~'--~I..,M~al. Biochem. 86, 184-192.
13 Gardiner , P .R. , Jaffe, E.L. and Dw)er , D.M. 119841 Identification of cross-reactive promast igote cell surface antigens of leishmanial stocks by t-51 labelhng and .mmu- noprecipitation. Infect. l mmun . 43,637-643.
14 Kessler, S.W (19751 Rap~d isolauon of antigens from cells with a Staphylococcal protein-A antibod3 adsorbent. Pa- rameters of the interaction of ant igcn-ant ibod) complexes x~ith protein-A. J. lmmunol . 115(61, 1617-1624.
15 Chirgwin. J .M, , Przbyla, A .E . , MacDonald. R.J. and Rutter . W.J. i19791 Isolation of biologicall3 active ribon- ucleic acid from sources enriched in ribonuclease. B~o- chemistry 18, 5294-5299.
16 Aviv, H. and Leder, P. (19721 Purification of b~ologicall) active globm messenger R N A by chromatography on oh- gothymidylic acid cellulose. Proc. Natl. Acad. Sci. LiSA 69, 1408-1412.
17 Pelham, H.R.B. and Jackson, R.J. (19'7,61 An effic|ent m R N A - d e p e n d e n t translation system for reticuloc.vte lys- ares. Eur. J. Biochem. 67. 247-256.
18 Bingham, P.M., Le~is. R. and Rubin, G.M. (19811 Clon- ing of D N A sequences from the ~ht te locus of D. melan- ogaster by a novel and general method. Cell 25.69,1-71).-1.
19 Huynh, T.V., Young, R.A. and Da,,rs, R.W. I19851 DNA cloning. Vol. 1: A Practtcal Approach. pp. 49-78. IRL Press, Oxford, Washington
20 Efstratiadis, A. , Kafatos, F.C. , Maxam. A M. and Man- .atis, T. ( 19761 Enzymatic in vitro s.vnthes~s of globm genes. Cell 7 ,279-288.
21 Wickens, M . P , Buell, G.N and Sch.mke. R T. (19781 Synthesis of double s t randed DNA complementar ) to k - sozyme, o, ,omucoid and ovalbumin m R N A ' s . J Biol Chem. 253,248,!-2494.
22 Seeburg, P H . , Shine. J., Martial, J .A. . Baxter, J .D. and Goodman , H.M. (19771 Nucleotide sequence and ampli- fication in bacteria of structural gene for rat growth hor- mone. Nature 270,486~194.
23 Holn, B. (1979) in vitro packaging of Lambda and Cos- mid DNA. Methods Enz}mol. 68, 299-309.
24 Benton, W.D. and Da~,is. R.W. (19771 Screemng gt re- combinant clones by hybridisation to single plaques in-situ. Science 196, 18(>182
25 Parnes. J .R. . \ ' e l an , B., Felsenfeld. A. . Ramana than , L , Ferrim. U. , Appela , E. and Seidman, J .G. ( 1981 ) blouse b:-microglobulin eDNA clones: A screening procedure for
e D N A c lone~correspondmg to rare m R N A ' x Proc Natl Acad. SOL USA "7,~, 2253-2257
2~, Southern. E.b,| 119~51 Detect ion ol ~pec~fic -,equence,, among. D N A fragments ,,eparated b.~ gel electrophore,q, J. Mol . Biol. 98. 5(13-51 ~
27 Rigb) . P.\V,J.. Dieekmann, b.l , Rhode,,. C and Berg.. P c197 ~) Label l ing deox.,,ribonucletc dctd to h~h ,,pectfic ac- tixit~ m ',itro b~, nick t r anqauon s~th DNA pol~mera,e I J. Mol. Btol. 113. 237,-251.
2,'q Lemesre. J.L.. Riz,,i, F.. Afeham. D.. Sad~g.arsk~,. N1 Capron, A. and Santoro, F 11~1851 Sub,,pecLe~ ,,peclfic ~urface antigen,, ol proma~,tlg.otes of the Letshmania don- o~ ant complex. Inlect. lmmun 5LL 13h-I-ll.
29 Lepa.,,, D A. . Nogueira, N. and Ct/hn. Z. ( lu,',:31 Surface ,mtigen~- ot Letshmant¢a dottt)t.¢llll proma ,ugo te , J Exp Med. 1: :,~, 1562-1572
3tt Handman . E.. Mttchell. G F and Godmg. J \ \ t l~,':,lD [denufication .rod characterization ot protein anugen , ,,t Lei~hmama troptca L-,olate~. J lmmunol . 12h. 511,'¢-512
31 Bou~ter. J.. Etge,,. R J. and Bordter. C q 1'4,'.,51 Identda- cation and purification of membrane and ,,oluble torm,, ol the malor surface protein o[ Lel$httltltlia promd,,llgOte,,. J Biol. Chem, 2t~cL [551)J,-15511'4
32 Co lomer -Gou ld , V , Oumtao. L.G . Ketthl~. J. and No- guelra. N 119851 A common m,iior ~urlace antigen on amdMig.ote-., and promastlgolex ol Letsllmania ~pecle,, J Exp. Med. 162. q()"-t416.
33 La~rence . F and Rober t -Gero, M. (1'4,',;51 Induction ,~1 heat shock and stre,~_, protein,, in promast igote , o| three Let~hmtttttd ~pecte',. Proc Natl. Acad Sei USA ',2. 4 4 1 ~ - t l ~
34 Chang. C S and (hang. . K P I lq,N61 Monoclonal ,mtl- bod 3 affintt\ punf icauon ot a Lei~hmanttt membrane g.l~,- coprotem and its inhibition ot Lev,_hmama-macroph,ge binding Proc. Natl Acad. Sc~ LiSA S3. l()tl-14.-l.
35 Etges. R.J. , Bou~]er. J. and Bord~er, C I lq~hl The rnul,~r surface protein ot Lt'l$hltltlttla proma-,tlgotes i,, a prote,~,e J. B~ol ( ' 'hem 261. '~0t~,";-ql~H
3~ Hoel lmakers . J H.J . Bor-,t, P.. \ an Den Burg. J.. \k e~,,,,- mann, C and C'ro,~. G .A .M. I lq~t)l 1"he ~olatton ot pla,,- raids containing DNA complementar~ to me,,,-enger RNA lot ~.arlant ,urlace gl)eoproteln.-, ot ]-r~piltzosomll hrttct~l. Gene S. 391-41"
3" Cordmgle.,.. J.S.. Haddo',,.. \~, J . Nene. V and Ta'.h,r D. \k . ( 1~S;61 Identification b) me~sage ,e lecuon ,,f eDNA clone,, encoding antigens of .~C]tlSlO~Ot?ltl t~ltltl~Otll ~k1~l B~ochem. Parasttol. 1,R 73-,~8.
3,':, Sheppard. H.W. and D ~ e r . D.M. I lt4,R6l Cloning - t LetshmatHa ¢ltmol alll gene,, encoding anugen , recogm~ed during human ~sceral le~shmania,,is Mol Biochem P,.r- a,qtol 1o. 34-43.


















