
Cloning and developmental expression of the …web.macam.ac.il/~gad100/AAA CV Gad 2010...
10
Communication in Genomics and Proteomics Cloning and developmental expression of the cytochrome P450 aromatase gene (CYP19) in the European eel (Anguilla anguilla) Itai Tzchori a,b, * , Gad Degani b,c , Avshalom Hurvitz b , Boaz Moav a a Department of Zoology, Tel Aviv University, Tel Aviv 69978, Israel b MIGAL, Galilee Technological Center, Kiryat Shmona 10200, Israel c School of Science and Technology, Academic Tel-Hai College, Israel Received 23 February 2004; revised 7 June 2004; accepted 21 June 2004 Available online 13 August 2004 Abstract To characterize the involvement of the aromatase gene during the process of sex determination in the European eel (Anguilla anguilla), the expression of its gonadal form was determined during various developmental stages. The cloned cDNA from the Euro- pean eel gonad (EeCYP19) contains an open reading frame of 1539 bp, encoding a deduced protein of 513 residues. The predicted amino acid sequence shows 97% identity with that of the Japanese eel, and 59–69% of identity with those of the CYP19 gonadal and brain forms of other teleost fish. Two potential initiation sites (ATG) were found downstream of the first ATG codon. A fluorescent- based method of real-time PCR was developed to quantify EeCYP19 expression. The expression levels of EeCYP19 in the gonads of adult males were approximately 12- and 30-fold lower than the levels in adult females and juvenile eels previously treated with E2, respectively. Expression of aromatase was found only in a single specimen in the control group. In contrast, no difference was found among sexes in the aromatase expression in the brain. Treatment with aromatase inhibitor (AI) of juvenile eel resulted in the total loss of aromatase expression in the gonads and brains. The results of this work revealed that AI treatment not only reduces the synthesis of estradiol, but reduces the expression levels of EeCYP19 as well. No evidence for the presence of a distinct extra-gonadal (brain) form of aromatase in the European eel could be provided. Ó 2004 Elsevier Inc. All rights reserved. Keywords: European eel; Sex determination; Estradiol-17b; CYP19; Expression; Aromatase inhibitor; Real-time PCR 1. Introduction Steroids play an important role in gonadal differenti- ation in lower vertebrates (Bogart, 1987; Devlin and Nagahama, 2002; Yamamoto, 1969). Treating fish with exogenous sex steroid hormones during early develop- mental stages may cause sex-inversion in spite of the genotypic sex. It has been demonstrated in many fish species that exogenous androgens generally have a masculinizing effect, while exogenous estrogens have feminizing effects (Yamazaki, 1983). However, the mechanism of action of these steroids on sex differentia- tion is not well understood. Estrogens are C-18 steroids indispensable for normal sexual development of the female, but have been associ- ated with sexual function in male too (Devlin and Naga- hama, 2002). The synthesis of estrogens is dependent on the activity of the steroidogenic enzyme P450 aromatase (CYP19). CYP19 catalyses aromatization of the A ring of C19 steroids to form the phenolic ring characteristic of estrogens, with concomitant loss of the C19 angular methyl group as formic acid. CYP19 activity is an important modulator of E2 concentration and is critical to the regulation processes controlled by E2 (Brodie, 1991). 0016-6480/$ - see front matter Ó 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2004.06.007 * Corresponding author. Present address: Bldg 6B, LMGD/ NICHD, National Institutes of Health, Bethesda, MD, USA. Fax: +301-402-0543. E-mail address: [email protected] (I. Tzchori). www.elsevier.com/locate/ygcen General and Comparative Endocrinology 138 (2004) 271–280 GENERAL AND COMPARATIVE ENDOCRINOLOGY
Transcript of Cloning and developmental expression of the …web.macam.ac.il/~gad100/AAA CV Gad 2010...

www.elsevier.com/locate/ygcen
General and Comparative Endocrinology 138 (2004) 271–280
GENERAL AND COMPARATIVE
ENDOCRINOLOGY
Communication in Genomics and Proteomics
Cloning and developmental expression of the cytochrome P450aromatase gene (CYP19) in the European eel (Anguilla anguilla)
Itai Tzchoria,b,*, Gad Deganib,c, Avshalom Hurvitzb, Boaz Moava
a Department of Zoology, Tel Aviv University, Tel Aviv 69978, Israelb MIGAL, Galilee Technological Center, Kiryat Shmona 10200, Israelc School of Science and Technology, Academic Tel-Hai College, Israel
Received 23 February 2004; revised 7 June 2004; accepted 21 June 2004
Available online 13 August 2004
Abstract
To characterize the involvement of the aromatase gene during the process of sex determination in the European eel (Anguilla
anguilla), the expression of its gonadal form was determined during various developmental stages. The cloned cDNA from the Euro-
pean eel gonad (EeCYP19) contains an open reading frame of 1539bp, encoding a deduced protein of 513 residues. The predicted
amino acid sequence shows 97% identity with that of the Japanese eel, and 59–69% of identity with those of the CYP19 gonadal and
brain forms of other teleost fish. Two potential initiation sites (ATG) were found downstream of the first ATG codon. A fluorescent-
based method of real-time PCR was developed to quantify EeCYP19 expression. The expression levels of EeCYP19 in the gonads of
adult males were approximately 12- and 30-fold lower than the levels in adult females and juvenile eels previously treated with E2,
respectively. Expression of aromatase was found only in a single specimen in the control group. In contrast, no difference was found
among sexes in the aromatase expression in the brain. Treatment with aromatase inhibitor (AI) of juvenile eel resulted in the total
loss of aromatase expression in the gonads and brains. The results of this work revealed that AI treatment not only reduces the
synthesis of estradiol, but reduces the expression levels of EeCYP19 as well. No evidence for the presence of a distinct extra-gonadal
(brain) form of aromatase in the European eel could be provided.
� 2004 Elsevier Inc. All rights reserved.
Keywords: European eel; Sex determination; Estradiol-17b; CYP19; Expression; Aromatase inhibitor; Real-time PCR
1. Introduction
Steroids play an important role in gonadal differenti-
ation in lower vertebrates (Bogart, 1987; Devlin and
Nagahama, 2002; Yamamoto, 1969). Treating fish with
exogenous sex steroid hormones during early develop-
mental stages may cause sex-inversion in spite of
the genotypic sex. It has been demonstrated in manyfish species that exogenous androgens generally have a
masculinizing effect, while exogenous estrogens have
0016-6480/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygcen.2004.06.007
* Corresponding author. Present address: Bldg 6B, LMGD/
NICHD, National Institutes of Health, Bethesda, MD, USA. Fax:
+301-402-0543.
E-mail address: [email protected] (I. Tzchori).
feminizing effects (Yamazaki, 1983). However, the
mechanism of action of these steroids on sex differentia-
tion is not well understood.
Estrogens are C-18 steroids indispensable for normal
sexual development of the female, but have been associ-
ated with sexual function in male too (Devlin and Naga-
hama, 2002). The synthesis of estrogens is dependent on
the activity of the steroidogenic enzyme P450 aromatase(CYP19). CYP19 catalyses aromatization of the A ring
of C19 steroids to form the phenolic ring characteristic
of estrogens, with concomitant loss of the C19 angular
methyl group as formic acid. CYP19 activity is an
important modulator of E2 concentration and is critical
to the regulation processes controlled by E2 (Brodie,
1991).

272 I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280
In the mammalian brain, the local formation of estro-
gens has been shown to influence the sexual differentia-
tion of neural structure and to modulate neuroendocrine
function and sexual behavior (Lephart, 1996). In lower
vertebrates, such as fish, CYP19 is predominantly ex-
pressed in the gonad and in the brain. It has becomepossible to reduce estrogen biosynthesis by inhibiting
the activity of aromatase, by using nonsteroidal aroma-
tase inhibitors such as fadrozole (CGS 16949A; Steele
et al., 1987). Fadrozole is a reversible competitive inhib-
itor, whereby both substrate and inhibitor compete for
the same site on the enzyme (Steele et al., 1987). In adult
mammalian and non-mammalian vertebrates, disrup-
tion of CYP19 was also shown to impair gonadal devel-opment and alter hormone synthesis (Afonso et al.,
1999). Treatments with aromatase inhibitors during sex-
ual differentiation resulted in skewed sex ratio in mam-
mals (Schieweck et al., 1988; Steele et al., 1987),
reptiles (Wibbles and Crews, 1994), and fish (Afonso
et al., 1999; Guiguen et al., 1999; Ijiri et al., 2003; Kwon
et al., 2000; Piferrer et al., 1994).
The cDNAs encoding CYP19 have been character-ized in a range of species, including mammals (Chen
et al., 1988; Corbin et al., 1988) and birds (McPhaul
et al., 1988). Among teleost fish, the CYP19 cDNA
has been cloned from the ovary of rainbow trout (Tana-
ka et al., 1992), catfish (Trant, 1994), medaka (Fukada
et al., 1996), Nile tilapia (Chang et al., 1997), goldfish
(Gelinas et al., 1998; Tchoudakova and Callard, 1998),
zebrafish (Kishida and Callard, 2001), European seabass (Dalla Valle et al., 2002), and Japanese eel (Ijiri
et al., 2003). Interestingly, teleost fish express two sub-
types of CYP19 cDNA. The isozyme, CYP19B, has
been isolated from the brain of goldfish (Gelinas et al.,
1998), tilapia (Cruz and Canario, 2000; Tsai et al.,
2001), zebrafish (Kishida and Callard, 2001), fathead
minnow (Halm et al., 2001), and trout (Dalla Valle
et al., 2002).European eels show considerable differences of sexual
dimorphism; females gain five times and more body
weight than males. Sex ratios of eel are highly variable
according to environmental sites (D�Ancona, 1959;
Tesch, 1977), and under aquaculture conditions, the
majority of eels differentiate into males (Holmgren and
Mosegaard, 1996). The skewed sex ratio demonstrates
a strong influence of the environment on the processof sex determination in eel. The eel is therefore a good
candidate for the evaluation of aromatase role during
the labile period in a fish with a strong environment-de-
pendent sex determination mechanism.
To evaluate the importance of aromatase expression
during the labile period of sex determination, we iso-
lated and characterized the full length cDNA encoding
CYP19 in European eel. In addition, CYP19 mRNAlevels in the gonads and brains were analyzed, and cor-
related with sex and developmental stages. The existence
of a distinct CYP19 gene in the eel�s brain was examined
as well.
2. Materials and methods
2.1. Animals and treatment
European glass eels (Anguilla anguilla) caught during
their migration in February–March 1999 and April 2000
were purchased from UK (UK Glass). For dry-feed
training they were fed approximately 1 month with com-
mercial eel diet (Trovit, Hendrix, Italy: protein 42.5%,
fats 8%, and ash 11.5%) combined with decreasingamounts of cod raw. Elvers were kept at 23–26 �C, with2–3 changes of total water volume daily and gentle aer-
ation. The appropriate (see below) amount of aromatase
inhibitor (AI: Fadrozole, CGS16949A, Novartis, Sum-
mit, NJ) or estradiol (E2, Sigma–Aldrich, St. Louis,
MO) was dissolved in 99% ethanol. Food for the control
groups was prepared in the same way using ethanol
only. The food was then thoroughly mixed and left ina fume cupboard overnight to allow ethanol evapora-
tion, and then stored at �20 �C. Thawed portions were
supplied daily to the fish.
Glass eels (0.5g) or elvers (5–7g) were fed 300mg/kg
dry feed AI for 90 days, or 10mg/kg dry feed of E2 for
150 days, or 10mg/kg bw of E2 for 150 days followed by
300mg/kg AI treatment for 90 days. The feed was given
ad libitum twice per day.To increase the percentage of females, untreated glass
eels (0.5g) were kept at a low density (100 fish/tank)
during the labile period in tanks equipped with shelters
(Degani and Kushnirov, 1992). The macroscopic
classification of the gonadal sex (male, females, and
intersexes) was done according to Beullens et al.
(1997). In case of doubt, samples were fixed in 4% form-
aldehyde for 24h, then in 15% sucrose in phosphate buf-fer saline (PBS) for 24h, and frozen sections (5lm)
stained with methylene-blue were observed under the
microscope to verify the sex. Data for sex ratio of two
replicate tanks for each treatment were pooled by treat-
ments for 50 fish.
2.2. Isolation of total RNA
Ovaries and brains samples were collected from eels
at different developmental stages, or following E2 and
AI treatments. The samples were frozen in liquid nitro-
gen and stored at �80 �C until used for RNA extraction.
RNA for polymerase chain reaction (PCR) was isolated
from 100mg (or less) ovaries and brain samples by the
acid guanidinium thiocyanate–phenol–chloroform
extraction method (Chomczynski and Sacchi, 1987)using EZ-RNA (Biological Industries, Israel). Oli-
go(dT)-primed double stranded cDNA was synthesized

I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280 273
using Promega cDNA synthesis system according to the
manufacturer�s recommendation.
2.3. Isolation and PCR amplification of CYP19 cDNA
fragment
To obtain a CYP19 cDNA fragment of European eel,
5 0 and 3 0 primers for PCR were designed from amino
acid sequences, which are highly conserved among spe-
cies. Ear-1 5 0 primer, 5 0-ATATTCGCTCAGAACC
AC-3 0 and Ear-2 3 0 primer, 5 0-CATACGCCCAACAT
TAAG-3 0 were designed. The positions at which they
hybridize are shown in Fig. 1. PCR was carried out in
25ll reaction mixture consisting of 10mM Tris–HCl,pH 8.3, 50lM MgCl2, 12.5lM of each primer, 1nM
each of dATP, dCTP, dGTP, and dTTP, 0.75U DyNA-
zyme II DNA polymerase (Finnzyme), and 10–100ng
cDNA. Thirty-five amplification cycles were carried
out under the following conditions: denaturation at
93 �C for 45s, annealing at 52 �C for 1min, and exten-
sion at 72 �C for 2min.
The PCR products were separated through 1% aga-rose gel and purified by using the QIAquick Gel Extrac-
tion kit (QIAGEN, Germany). The product was cloned
into the pGEM-T easy plasmid vector (Promega, Milan,
Italy) and sequenced using the BigDye Terminator Cycle
Sequencing Core kit (Applied Biosystem, Monza, Italy).
2.4. 5 0 and 3 0 RACE-PCR
To obtain the 5 0-end of the CYP19 sequence, three
gene specific antisenses, Ear-3R (5 0-GTCCACCAC
TGGATGATAGCGTAG-3 0), Ear-4R (5 0-CTTTCTT
GTCACCGATCACAGTGT-3 0), and Ear-5R (5 0-AC
CCTGAGGAACATCTGGTTGGAGA-3 0) were de-
signed to replicate regions in the open reading frame
of the CYP19 cDNA of the eel.
Fig. 1. Representative samples of specific RT-PCR products for
aromatase and b-actin in the gonads of European eel. The molecular
size marker (M) was the 100bp DNA ladder. The b-actin is a product
of 192bp and the aromatase is 145bp lengths.
Using the SMART RACE cDNA amplification kit
(BD Biosciences, Palo Alto), first strand synthesis was
carried out with 500ng of total RNA and 100nM of
Ear-3R. Two consecutive PCRs were performed, using
100nM of Ear-4R and Ear-5R in nested PCR. Amplifi-
cation by touchdown PCR was carried out for 30 cyclesof 30s at 94 �C, 5 cycles of 50s at 72 �C, and 25 cycles of
50s at 68 �C. PCR products were cloned, and four clones
containing the 5 0-end of European eel CYP19 were se-
quenced as described above. To obtain the 3 0-end of
the CYP19 cDNA sequences, a gene specific sense pri-
mer Ear-6R (5 0-AACAGCTGGCTGAGAAAGACA-
3 0) was designed. First strand synthesis and RT-PCR
were carried out using dT-anchor (Clontech). Amplifica-tion was performed using touchdown PCR as described
above, with range of Ta temperature from 50 to 60 �C.
2.5. Computer analysis
Sequence alignments and comparisons of homology
between various aromatase sequences were performed
using the Vector NTI Advance software (Informax).Phylogenetic trees were constructed by maximum-
parsimony analysis with the computer program PAUP
4 (b7). Bootstrap proportions were used to assess the
robustness of tree with 1000 bootstrap replications.
2.6. Real-time quantitative RT-PCR analysis
For the quantitative real-time PCR analysis of Ee-CYP19 mRNA expression levels, the LightCycler sys-
tem (Roche Molecular Biochemicals, Indianapolis) and
a LightCycler DNA master SYBR green I kit were used.
For hot start PCR, LightCycler FastStart DNA Master
SYBR green I containing the FastStart Taq DNA poly-
merase was used. Preliminary experiments were per-
formed to determine the optimal conditions for PCR
(annealing temperature and final Mg2+ concentration)on each pair of primers.
An aliquot of the RNA (1lg) was reversed tran-
scribed as described above and 1/100th of this reaction
was used in duplicate quantitative fluorescent PCR to
determine transcript abundance.
Primer sets were selected so that the amplified cDNA
would span two exon/intron boundaries of EeCYP19
gene, thus eliminating amplification of genomic DNA.The primer set for CYP19 was Ear-7 5 0-GAACC
CAGAGGTAGAGC-3 0 (forward) and Ear-8 5 0-ACCT
TGTAGCCTTCGAT-3 0 (reverse). The b-actin was used
as an internal control to normalize abundance, with pri-
mer set of 5 0-ACCACCTTCAACTCCATC-3 0 (for-
ward), 5 0-TCCTGCTTGCTAATCCAC-3 0 (reverse).
Each reaction (20ll) contained 2ll of the respective
cDNA dilution, primers at 0.2 mM, and Mg2+ at2mM. Amplification by touchdown PCR was carried
out for 30s at 94 �C, 5 cycles of 5s at 68 �C, 40 cycles

274 I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280
of 5s at 65 �C for the b-actin, and by 30s at 94 �C, 5 cy-
cles of 5s at 65 �C, 40 cycles of 5s at 63 �C for the
CYP19. Unspecific PCR products, e.g., primer dimers,
melt at the elevated temperature, the non-specific fluo-
rescence signal is eliminated, and accurate quantification
of the desired product is ensured (Fig. 1).To ensure the fidelity of mRNA extraction and re-
verse transcription, all samples were subjected to PCR
amplification with oligonucleotide primers specific for
the constitutively gene expressed b-actin. Transcript
abundance of the CYP19 was normalized to the abun-
dance of b-actin, and reported as a relative transcripts
abundance. Constructing a standard curve with serial
dilutions of known template concentration is essentialto quantify the concentration of unknown templates,
thus we used the serial dilution (1:1–1:108) of cDNA.
2.7. Southern blot
European eel genomic DNA was extracted from liver,
digested (10lg) with BglII or EcoRI, and fractionated
by electrophoresis on 0.8% agarose gel. Then it was blot-ted onto a nylon membrane (NYTRAN, Schleicher and
Schuell) and then incubated overnight at 42 �C in
hybridization buffer containing the 32P-labeled cDNA
probe, 6· SSC, 10· Denhard solution, 0.1% SDS, 50%
formamide, and 50lg/ml of calf thymus DNA. The
membrane was washed in 2· SSC, 0.5% SDS, then in
1· SSC, 0.1% SDS. After a final wash with 0.5· SSC,
0.1% SDS at 42 �C, the membrane was exposed on aPhosphorimager (Molecular Dynamics, CA) at room
temperature for 24h. The probe was a 192bp fragment
that coding for 64 amino acids of the European eel aro-
matase peptide.
2.8. Statistical analysis
The relative mRNA expression levels were calculatedfrom each set of quantified data. All data are shown as
means±SEM, although where appropriate they were
subject to logarithmic transformation before being ana-
lyzed by one way ANOVA followed by Tukey�s multiple
range test. A probability level of P<0.05 was used as the
criterion for significance of difference between groups.
The significance of difference in sex ratio between con-
trol and treated groups was tested by Fisher exact test.
3. Results
3.1. Isolation and sequence of European eel CYP19
cDNA
The gonadal form of EeCYP19 was cloned using RT-PCR and RACE-PCR. The full length of the open read-
ing frame (ORF) of CYP19 (EeCYP19) was 1539bp
encoding a protein of 513 amino acids (Fig. 2). This
cDNA sequence showed 97% identity to that of the A.
japonica. The EeCYP19 amino acid showed 97% iden-
tity with that of A. japonica, 59–66% with teleost gona-
dal isoforms, and 56–69% with teleost brain isoforms
(Figs. 3 and 4).Two other potential initiations signals situated 9 and
33bp downstream of the first ATG codon were
identified.
3.2. Tissue specific expression of EeCYP19 and sex ratio
Juvenile eels from the E2 treatment group displayed a
predominately female population (88%), while only a5% female presence was detected in the control un-
treated juvenile group at density of 1000 fish/1m3 (Table
1). Reducing the density of fish (100 fish/1m3) and the
social stress by providing shelters increased significantly
the female percentage to 36%.
The expression of EeCYP19 gene was investigated in
tissues of juvenile eels (0.5g), elvers (5–7g), adult males
(80–120g), and adult females (100–200g), using RT-PCR. The abundance of EeCYP19 transcripts indicated
that the gene was expressed in both the gonads and the
brain. Expression of EeCYP19 in the adult male�sgonads was 12-fold lower when compared to the adult
female group, and 30-fold lowers when compared to
the juvenile E2 treated group. In the control untreated
juvenile fish, transcripts were detected in only one
sample (1/11) (Fig. 5).The abundance of EeCYP19 transcripts in the brains
of elvers previously treated with E2, adult females,
adult males, and control were not significantly different
(Fig. 5).
3.3. Effects of aromatase inhibitor on mRNA levels and
sex ratio
The abundance of EeCYP19 transcripts after AI
treatments was quantified. EeCYP19 was not detected
in the gonads of AI treated fish and the treatment re-
sulted in a complete masculinization of all fish (Table 1).
EeCYP19 mRNA average levels did not differ be-
tween E2 and E2+AI treated fish due to the high vari-
ance, although only 50% (5/10) of the fish were
detected with mRNA levels in the E2+AI as comparedto 80% (8/10) in the E2 treated group (Fig. 6). In the
control untreated fish that were held under low density
conditions, the EeCYP19 mRNA levels were signifi-
cantly lower as compared to the two treatment groups,
and only 36% (4/11) of the fish had detectable EeCYP19
mRNA levels.
The abundance of EeCYP19 transcripts in the brains
was extremely high in the E2 treated group (Fig. 6).Treatment with AI resulted in a strong reduction of
the EeCYP19 mRNA expression in the E2+AI groups.

Fig. 2. The full length of nucleotide and the deduced amino acid sequences of the European eel aromatase form of CYP19 (EeCYP19). Open reading
frame of 1539bp encodes a protein of 513 residues. The numbers on the right refers to the position of the nucleotides and the amino acids. The
positions of primers used at real-time PCR and PCR are indicated with arrow under the sequence. The initiation codons (ATG) and the stop codon
(TAA) have been marked in bold font.
I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280 275
Moreover, the low-density control fish showed low
expression levels of EeCYP19 as well.
3.4. Southern blot analysis of eel genomic DNA
European eel DNA was digested by two restrictionenzymes (BglII and EcoRI) and hybridized with Ee-
CYP19 specific probe, as shown in Fig. 7. Hybridization
of the fractionated DNA to the labeled probe resulted in
a singled band corresponding to approximately 2.1kb
EeCYP19 cDNA.
4. Discussion
In this study, a cDNA encoding the cytochrome P450
aromatase (EeCYP19) was cloned and sequenced from
the ovary of the European eel, and the importance of

Fig. 3. Alignment of the amino acid sequence of European eel P450arom (CYP19) with that of the Japanese eel, tilapia (ovary and brain), medaka
(ovary and brain), goldfish (ovary and brain), fathead minnow (brain), stingray, chicken (ovary), and human placenta. The amino acid sequences
identical to the European eel CYP19 are marked by asterisks and dots.
276 I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280

Fig. 4. Phylogenetic tree of CYP19 amino acid. The tree was
constructed by using program PAUP 4. Comparisons were made with
the deduced amino acid sequences of CYP19 forms in chicken ovary,
human placenta, fathead minnow, goldfish (brain and ovary), tilapia
(brain and ovary), medaka (brain and ovary), and stingray. Length of
lines indicates genetic distance. The values are bootstrap probabilities
estimated by 1000 replications.
Table 1
Effects of treatments on percentage of females in European eel
Treatment n F (%)
High density (1000/1m3) 100 5ab
Low density (100/1m3) 50 36c
E2 50 88d
AI 50 0a
E2+AI 50 8b
Eels were randomly sampled from each group for macroscopic clas-
sification of the gonad according to Beullens et al. (1997). The groups
were high-density (1000fish/1m3) and low-density (100fish/1m3) tanks;
E2: fish fed estradiol for feminization for 150 days. AI: 300mg/kg bw
AI treatment for 90 days; E2+AI: estradiol treated group for 150 days
with 10mg/kg bw, followed by 300mg/kg bw AI treatment for 90 days.
Female�s percentages marked by the same letter do not differ signifi-
cantly from each other (p>0.05).
I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280 277
its expression and activity during sex determination was
demonstrated. The cloned cDNA of EeCYP19 from the
European eel gonad contains an open reading frame of
1539bp, encoding a deduced protein of 513 residues.
The predicted amino acid sequence shows 97% identity
with that of the Japanese eel. Interestingly, the compar-
ison of the EeCYP19 to other teleosts CYP19 gene re-
vealed that in some fish the higher identity was to thebrain form (medaka, tilapia, and trout), while in others
it was more similar to the ovarian form (goldfish, zebra-
fish). Two potential initiation sites (ATG) were found
downstream of the first ATG codon. These results
are similar to multiple initiation sites reported in other
teleosts (Dalla Valle et al., 2002; Halm et al., 2001; Ijiri
et al., 2003).
Quantitative real-time PCR analysis revealed that Ee-
CYP19 transcripts expressed in the gonads of females
were significantly higher than those reported for the
male counterparts. These differences are more evidentin juvenile eels previously subjected to E2 treatments
for feminization, whereas in the untreated juveniles
CYP19 expression, which was not detected. These re-
sults indicate a sexual differential expression of aroma-
tase in European eel gonads, favoring the female sex.
This trend is not unexpected considering the strong syn-
thesis of estrogen in the ovary, compared to limited
estrogen synthesis in testis.In contrast to the situation in the gonads, no signifi-
cant difference in expression levels was found in the
brain of eels at various developmental stages. It is
important to note that these values were determined in
eels that weigh more than 15g.; therefore, it is possible
that sexual differentiation may have already occurred
at this particular stage. In teleosts, similar studies aimed
to characterize the involvement of CYP19 gene/s expres-sion during sex differentiation indicate its complexity. In
the Nile tilapia (Oreochromis niloticus) (Kwon et al.,
2001) and in the European sea bass (Dicentrarchus lab-
rax) (Dalla Valle et al., 2002), high expression levels
of the gonadal form of CYP19 were found in the gonad
but not in the brain. In contrast, the initial differences in
the CYP19 expression in the zebrafish could be first de-
tected in the brain-form of CYP19 in the brain (Trantet al., 2001).
Treatment of juvenile eels with AI resulted in total
absence of aromatase expression. Since high levels of
CYP19 were found in the gonads after E2 treatment,
the decline in CYP19 levels in some fish after exposure
to AI in the combined treatment (E2+AI) can be due
to the inhibitory affect of AI. The heterogeneity of the
expression of this group can be due to the fact that somefish, which received E2 previously were already differen-
tiated into females, and therefore the gonads were no
longer sensitive to the AI treatments. The strong inhibi-
tion of CYP19 expression was found in the brain as well.
AI treatment of elvers, with or without previous E2
treatment, resulted in dramatic decreases in aromatase
expression in the brain of all treated fish.
The expression levels of EeCYP19 were 40-fold high-er in the E2 treated group compared to the AI treated
group. The fact that high levels of CYP19 were associ-
ated with high percentage of female, whereas AI treat-
ment causing full inhibition of CYP19 expression
resulted in the masculinization of all treated fish, indi-
cating an importance for high CYP19 levels during fe-
male differentiation. This is in line with the results of
(Kwon et al., 2002) in Nile tilapia, which showed thatlow aromatase activity during the crucial period of sex
differentiation always resulted in masculinization,
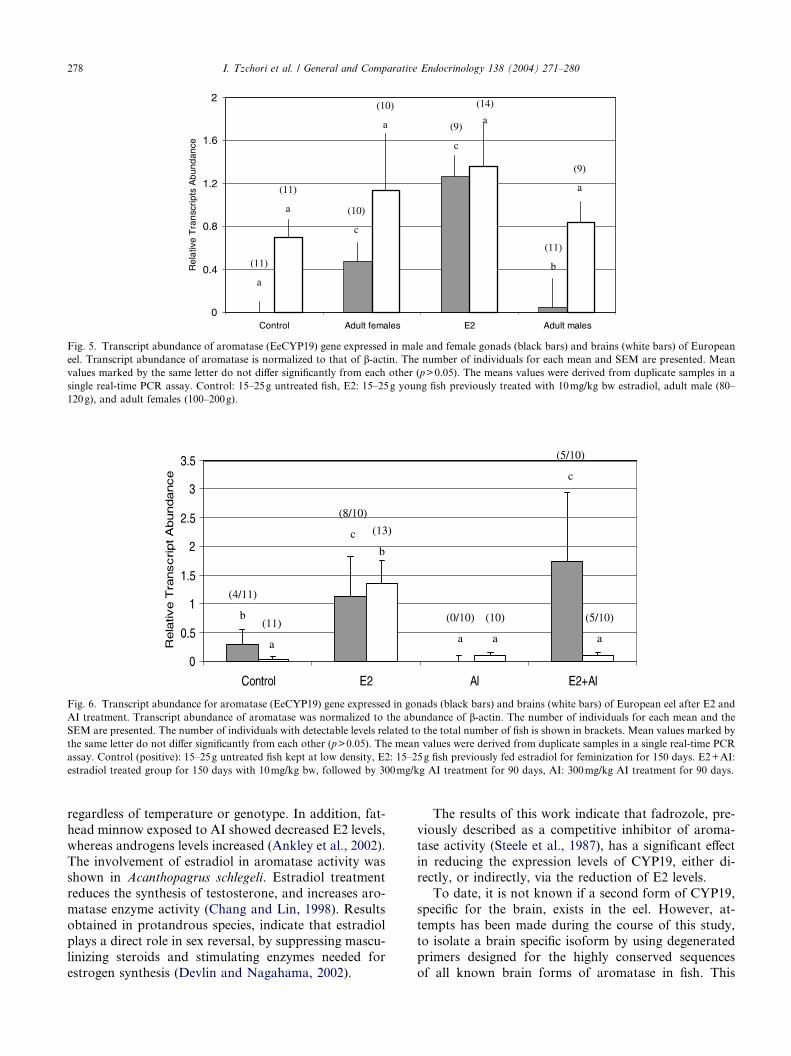
Fig. 5. Transcript abundance of aromatase (EeCYP19) gene expressed in male and female gonads (black bars) and brains (white bars) of European
eel. Transcript abundance of aromatase is normalized to that of b-actin. The number of individuals for each mean and SEM are presented. Mean
values marked by the same letter do not differ significantly from each other (p>0.05). The means values were derived from duplicate samples in a
single real-time PCR assay. Control: 15–25g untreated fish, E2: 15–25g young fish previously treated with 10mg/kg bw estradiol, adult male (80–
120g), and adult females (100–200g).
Fig. 6. Transcript abundance for aromatase (EeCYP19) gene expressed in gonads (black bars) and brains (white bars) of European eel after E2 and
AI treatment. Transcript abundance of aromatase was normalized to the abundance of b-actin. The number of individuals for each mean and the
SEM are presented. The number of individuals with detectable levels related to the total number of fish is shown in brackets. Mean values marked by
the same letter do not differ significantly from each other (p>0.05). The mean values were derived from duplicate samples in a single real-time PCR
assay. Control (positive): 15–25g untreated fish kept at low density, E2: 15–25g fish previously fed estradiol for feminization for 150 days. E2+AI:
estradiol treated group for 150 days with 10mg/kg bw, followed by 300mg/kg AI treatment for 90 days, AI: 300mg/kg AI treatment for 90 days.
278 I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280
regardless of temperature or genotype. In addition, fat-
head minnow exposed to AI showed decreased E2 levels,
whereas androgens levels increased (Ankley et al., 2002).
The involvement of estradiol in aromatase activity wasshown in Acanthopagrus schlegeli. Estradiol treatment
reduces the synthesis of testosterone, and increases aro-
matase enzyme activity (Chang and Lin, 1998). Results
obtained in protandrous species, indicate that estradiol
plays a direct role in sex reversal, by suppressing mascu-
linizing steroids and stimulating enzymes needed for
estrogen synthesis (Devlin and Nagahama, 2002).
The results of this work indicate that fadrozole, pre-
viously described as a competitive inhibitor of aroma-
tase activity (Steele et al., 1987), has a significant effect
in reducing the expression levels of CYP19, either di-rectly, or indirectly, via the reduction of E2 levels.
To date, it is not known if a second form of CYP19,
specific for the brain, exists in the eel. However, at-
tempts has been made during the course of this study,
to isolate a brain specific isoform by using degenerated
primers designed for the highly conserved sequences
of all known brain forms of aromatase in fish. This

Fig. 7. Southern blot analysis of European eel genomic DNA. DNA
prepared from eel liver was fractionated by electrophoresis on 0.8%
agarose gel after digestion with BglII and EcoRI. The fractionated
DNA was blotted onto NYTRAN membrane and hybridized with a32P-labeled cDNA probe coding for 64 amino acids of the eel
aromatase peptide. Sizes in kilobases are indicated in the right margin.
I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280 279
technique was unable to distinguish an extra-gonadal
isoform, revealing only a single form of CYP19, namely
the gonadal form. Furthermore, the results from South-
ern blot analysis show only a single band in the entire
genome. These results are consistent with those in thecartilaginous fish, the stingray. The transcripts from
the extra-gonadal form encodes a protein identical to
the ovarian form (Ijiri et al., 2000). Moreover, Northern
blot analysis detected only a single aromatase form in
the Japanese eel (Ijiri et al., 2003). The possible absence
of a second form of aromatase in eel may be explained
by a theory that duplication in the fish genome took
place after the Anguilliform fish diverged from the mainstock of the Euteleostei (Robinson-Rechavi et al., 2003).
This theory is supported by the sequence analysis of
EeCYP19, indicating a similarity to the brain form in
some fish and to the gonadal form in others.
In summary, we have cloned a CYP19 cDNA from
the ovary of the European eel. High amino acid identi-
ties were found with the gonadal form in some fish
and to the brain form in other fish. Transcripts ofEeCYP19 were found in both the ovary and brain tis-
sues; however, EeCYP19 levels were higher in ovarian
than in testicular tissues. Treatment with AI resulted
in a strong reduction of the EeCYP19 mRNA expres-
sion. No evidence could be provided for the presence
of an extra-gonadal (brain) form of aromatase in eel.
Acknowledgments
We thank Prof. Zvi Yaron and Dr. Hanna Rosenfeld
for helpful comments and critical reading the manu-
script. This work was supported by the Israeli Ministry
of Agriculture (Grant 99-0169-596), and by the Israeli
Ministry of Science, Culture and Sport (Grant 1296-
199). Fadrozole was kindly supplied by Novartis.
References
Afonso, L.O.B., Iwama, G.K., Smith, J., Donaldson, E.M., 1999.
Effects of the aromatase inhibitor fadrozole on plasma sex steroid
secretion and oocyte maturation in female coho salmon (Oncorhyn-
chus kisutch) during vitellogenesis. Fish Physiol. Biochem. 20, 231–
241.
Ankley, G.T., Kahl, M.D., Jensen, K.M., Hornung, M.W., Korte, J.J.,
Makynen, E.A., Leino, R.L., 2002. Evaluation of the aromatase
inhibitor fadrozole in a short-term reproduction assay with the
fathead minnow (Pimephales promelas). Toxicol. Sci. 67, 121–130.
Beullens, K., Eding, E.H., Gilson, P., Ollevier, F., Komen, J., 1997.
Gonadal differentiation, intersexuality and sex ratio of European
eel (Anguilla anguilla L.) kept in captivity. Aquaculture 153, 135–
150.
Bogart, M.H., 1987. Sex determination: A hypothesis based on steroid
ratios. J. Theor. Biol. 128, 349–357.
Brodie, A., 1991. Aromatase and its inhibitors—an overview. J.
Steroid Biochem. Mol. Biol. 40, 255–261.
Chang, X.T., Kobayashi, T., Kajiura, H., Nakamura, M., Nagahama,
Y., 1997. Isolation and characterization of the cDNA encoding the
tilapia (Oreochromis niloticus) cytochrome P450 aromatase
(P450arom): changes in P450arom mRNA, protein and enzyme
activity in ovarian follicles during oogenesis. J. Mol. Endocrinol.
18, 57–66.
Chang, C.F., Lin, B.Y., 1998. Estradiol-17-beta stimulates aromatase
activity and reversible sex change in protandrous black porgy,
Acanthopagrus schlegeli. J. Exp. Zool. 280, 165–173.
Chen, S., Besman, M.J., Sparkes, R.S., Zollman, S., Klisak, I.,
Mohandas, T., Hall, P.F., Shively, J.E., 1988. Human aromatase:
cDNA cloning, southern blot-analysis, and assignment of the gene
to chromosome 15. DNA 7, 27–38.
Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA
isolation by acid guanidinium thiocyanate phanol chloroform
extraction. Anal. Biochem. 162, 156–159.
Corbin, C.J., Graham-Lorence, S., McPhaul, M., Mason, M., Men-
delson, C.R., Simpson, E.R., 1988. Isolation of a full-length cDNA
insert encoding human aromatase system cytochrome P-450 and its
expression in nonsteroidogenic cells. Proc. Natl. Acad. Sci. USA
85, 8948–8952.
Cruz, M., and Canario, A.V.M., 2000. cDNA cloning of brain and
ovary aromatase in tilapia, Oreochromis mossambicus. In: Proceed-
ings of the Sixth International Symposium of Reproductive
Physiology of Fish. Bergen, Norway, July 4–9, 1999, p. 193.
D�Ancona, U., 1959. Distribution of the sexes and environmental
influence in the European eel. Archives d�Anatomie microscopique
et de Morphologie expe�rimentale 48, 61–70.
Dalla Valle, L., Lunardi, L., Colombo, L., Belvedere, P., 2002.
European sea bass (Dicentrarchus labrax L.) cytochrome
p450arom: cDNA cloning, expression and genomic organization.
J. Steroid Biochem. Mol. Biol. 80, 25–34.
Degani, G., Kushnirov, D., 1992. Effects of 17b-estradiol and grouping
on sex determination of European eels. Prog. Fish Cult. 54, 88–91.
Devlin, H., Nagahama, Y., 2002. Sex determination and sex differen-
tiation in fish: an overview of genetic, physiological, and environ-
mental influences. Aquaculture 208, 191–364.
Fukada, S., Tanaka, M., Matsuyama, M., Kobayashi, D., Nagahama,
Y., 1996. Isolation, characterization, and expression of cDNAs
encoding the medaka (Oryzias latipes) ovarian follicle cytochrome
P-450 aromatase. Mol. Reprod. Dev. 45, 285–290.
Gelinas, D., Pitoc, G.A., Callard, G.V., 1998. Isolation of a goldfish
brain cytochrome P450 aromatase cDNA: mRNA expression
during the seasonal cycle and after steroid treatment. Mol. Cell.
Endocrinol. 138, 81–93.
Guiguen, Y., Baroiller, J.F., Ricordel, M.J., Iseki, K., McMeel, O.M.,
Martin, S.A.M., Fostier, A., 1999. Involvement of estrogens in the

280 I. Tzchori et al. / General and Comparative Endocrinology 138 (2004) 271–280
process of sex differentiation in two fish species: the rainbow trout
(Oncorhynchus mykiss) and the Tilapia (Orechromis niloticus). Mol.
Reprod. Dev. 54, 154–162.
Halm, S., Rand-Weaver, M., Sumpter, J.P., Tyler, C.R., 2001. Cloning
and molecular characterization of an ovarian-derived (brain-like)
P450 aromatase cDNA and development of a competitive RT-PCR
assay to quantify its expression in the fathead minnow (Pimephales
promelas). Fish Physiol. Biochem. 24, 49–62.
Holmgren, K., Mosegaard, H., 1996. Implications of individual
growth status on the future sex of the European eel. J. Fish Biol.
49, 910–925.
Ijiri, S., Berard, C., Trant, J., 2000. Characterization of gonadal and
extra-gonadal forms of the cDNA encoding the Atlantic stingray
(Dasyatis sabina) cytochrome P450 aromatase (CYP19). Mol. Cell.
Endocrinol. 164, 169–181.
Ijiri, S., Kazeto, Y., Lokman, M., Adachi, S., Yamauchi, K., 2003.
Characterization of a cDNA encoding P-450 aromatase (CYP19)
from Japanese eel ovary and its expression in ovarian follicles
during induced ovarian development. Gen. Comp. Endocrinol.
130, 193–203.
Kishida, M., Callard, G.V., 2001. Distinct cytochrome P450 aromatase
isoforms in zebrafish (Danio rerio) brain and ovary are differentially
programmed and estrogen regulated during early development.
Endocrinology 142, 740–750.
Kwon, J.Y., McAndrew, B.J., Penman, D.J., 2001. Cloning of brain
aromatase gene and expression of brain and ovarian aromatase
genes during sexual differentiation in genetic male and female Nile
tilapia Oreochromis niloticus. Mol. Reprod. Dev. 59, 359–370.
Kwon, J.Y., McAndrew, B.J., Penman, D.J., 2002. Treatment with an
aromatase inhibitor suppresses high-temperature feminization of
genetic male (YY) Nile tilapia. J. Fish Biol. 60, 625–636.
Kwon, J.Y., Haghpanah, V., Kogson-Hurtado, L.M., McAndrew,
B.J., Penman, D.J., 2000. Masculinization of genetic female Nile
tilapia (Oreochromis niloticus) by dietary administration of an
aromatase inhibitor during sexual differentiation. J. Exp. Zool. 287,
46–53.
Lephart, E.D., 1996. A review of brain aromatase cytochrome P450.
Brain Res. Brain Res. Rev. 22, 1–26.
McPhaul, M.J., Noble, J.F., Simpson, E.R., Mendelson, C.R., Wilson,
J.D., 1988. The expression of a functional cDNA encoding the
chicken cytochrome P-450arom (aromatase) that catalyzes the
formation of estrogen from androgen. J. Biol. Chem. 263, 16358–
16363.
Piferrer, F., Zanuy, S., Carrillo, M., Solar II, Devkubm, R.H.,
Donaldson, E.M., 1994. Brief treatment with an aromatase
inhibitor during sex differentiation causes chromosomally female
salmon to develop as normal, functional males. J. Exp. Zool. 270,
255–262.
Robinson-Rechavi, M., Marchand, O., Esriva, H., Bardet, P.-L.,
Zelus, D., Hughes, S., Luadet, V., 2003. Euteleost fish genome are
characterized by expansion of gene families. Gen. Res. 11, 781–788.
Schieweck, K., Bhatnagar, A.S., Matter, A., 1988. CGS 16949A, a new
nonsteroidal aromatase inhibitor: effects on hormone-dependent
and independent tumors in vivo. Cancer Res. 48, 834–838.
Steele, R.E., Mellor, L.B., Sawyer, W.K., Wasvary, J.M., Browne,
L.J., 1987. In vitro and in vivo studies demonstrating potent and
selective estrogen inhibition by the nonsteroidal aromatase inhib-
itor CGS 16949A. Steroids 50, 147–161.
Tanaka, M., Telecky, T.M., Fukada, S., Adachi, S., Chen, S.,
Nagahama, Y., 1992. Cloning and sequence analysis of the cDNA
encoding P-450 aromatase (P450arom) from rainbow trout
(Oncorhynchus mykiss) ovary; relationship between the amount of
P450arom mRNA and the production of oestradiol-17b in the
ovary. J. Mol. Endocrinol. 8, 53–61.
Tchoudakova, A., Callard, G.V., 1998. Identification of multiple CYP
19 genes encoding different cytochrome P450 aromatase isozymes
in brain and ovary. Endocrinology 139, 2179–2189.
Tesch, F.W., 1977. The Eel. Biology and Management of Anguillid
Eels. Chapman & Hall, London.
Trant, J.M., 1994. Isolation and characterization of the cDNA
encoding the channel catfish (Ictalurus punctatus) form of cyto-
chrome P450arom. Gen. Comp. Endocrinol. 95, 155–168.
Trant, J.M., Gavasso, S., Ackers, J., Chung, B., Place, A.R., 2001.
Development expression of cytochrome P450 aromatase genes
(CYP19a and CYP19b) in zebrafish fry (Danio rerio). J. Exp. Zool.
290, 475–483.
Tsai, C.-L., Wang, L.-H., Fang, L.-S., 2001. Estradiol and para-
chlorophenylalanine downregulate the expression of brain aroma-
tase and estrogen receptor-a mRNA during the critical period of
feminization in tilapia, Oreochromis mossambicus. Neuroendocri-
nology 74, 325–334.
Wibbles, T., Crews, D., 1994. Putative aromatase inhibitor induces
male sex determination in a female unisexual lizard and in turtle
with temperature-dependent sex determination. J. Endocrinology
141, 295–299.
Yamamoto, T., 1969. Sex differentiation. In: Hoar, W.S., Randall, D.J.
(Eds.), Fish Physiology, vol. III. Academic press, New York, pp.
117–175.
Yamazaki, F., 1983. Sex control and manipulation in fish. Aquaculture
33, 329–355.


















