
ClC-5 regulates dentin development through TGF- 1...
8
ClC-5 regulates dentin development through TGF-b1 pathway Xiaohong Duan a,1, *, Yong Mao b,1 , Ting Yang a , Xuan Wen a , Huan Wang c , Jin Hou a , Yang Xue d , Rong Zhang e a Department of Oral Biology, School of Stomatology, The Fourth Military Medical University, 145 Changle West Road, Xi’an, Shaanxi 710032, China b Department of Prosthodontics, School of Stomatology, The Fourth Military Medical University, Xi’an, China c Department of Orthodontics, School of Stomatology, The Fourth Military Medical University, Xi’an, China d Department of Oral Surgery, School of Stomatology, The Fourth Military Medical University, Xi’an, China e Department of Endodontics, School of Stomatology, The Fourth Military Medical University, Xi’an, China 1. Introduction ClC-5 is one member of the ClC chloride channel family. 1 The CLCN5 gene mutates in Dent’s disease, an X-linked nephro- pathy associated with low molecular weight proteinuria, hypercalciura, nephrocalcinosis and nephrolithiasis. 2,3 ClC-5 is predominantly expressed in the kidney, 4 but ClC-5 is also present in the liver, brain, lung, testis, small intestine and colon. 2,5–7 In the studies by Wang et al., 8 an animal model for Dent’s disease was developed by studying mice with targeted inactivation of the corresponding murine gene, Clcn5. The authors reported that a small number of mutant mice ‘‘had deformities of the dorsal spine and backward growth of the teeth, but radiologic evidence for osteopenia or rickets was absent’’ 8 ; the mechanism by which abnormal ClC-5 affected tooth development was not elaborated on. Chlorine is one of the abundant elements in teeth. However, in contrast to enormous knowledge regarding chloride channels in other organs, limited data pertaining to chloride channel functions have been demonstrated in tooth archives of oral biology 54 (2009) 1118–1124 article info Article history: Accepted 26 September 2009 Keywords: ClC-5 Dentin Odontoblast TGF-b1 Chloride channel abstract ClC-5 is one of the voltage-dependent chloride channel (ClC) family members. Mutations involving CLCN5 cause an X-linked nephropathy associated with Dent’s disease. Some Clcn5 gene knockout (ClC-5 KO) mice have abnormal growth of the teeth; however, the expression and function of ClC-5 during tooth development is still unknown. Herein we report abnormal dentin structure, decreased DSPP and increased TGF-b1 protein level in ClC-5 KO teeth. In odontoblast-like MDPC-23 cells, the mRNA levels of Tgfb1, Dspp and Dmp- 1 were upregulated with Clcn5 RNAi after 48 h treatment; whilst there was no change in those of TGF-beta receptor Tgfbr1 and Tgfbr2. We suggest that the dentin changes in ClC-5 KO mice might be a result of increasing TGF-b1, and the interplay between ClC-5 and TGF-b1 needs further identified. # 2009 Elsevier Ltd. All rights reserved. * Corresponding author. Tel.: +86 29 84776169; fax: +86 29 83223047. E-mail address: [email protected] (X. Duan). 1 These authors contributed equally to this paper. Abbreviations: ClCN5, Clcn5, chloride channel 5 gene; TGF-b1, tgfb1, transforming growth factor, beta 1; Tgfbr, transforming growth factor beta receptor; DSPP, Dspp, dentin sialophosphoprotein; DMP1, Dmp1, dentin matrix acidic phosphoprotein 1, dentin matrix protein 1. available at www.sciencedirect.com journal homepage: www.intl.elsevierhealth.com/journals/arob 0003–9969/$ – see front matter # 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.archoralbio.2009.09.008
Transcript of ClC-5 regulates dentin development through TGF- 1...

ClC-5 regulates dentin development through TGF-b1pathway
Xiaohong Duan a,1,*, Yong Mao b,1, Ting Yang a, Xuan Wen a, Huan Wang c, Jin Hou a,Yang Xue d, Rong Zhang e
aDepartment of Oral Biology, School of Stomatology, The Fourth Military Medical University, 145 Changle West Road,
Xi’an, Shaanxi 710032, ChinabDepartment of Prosthodontics, School of Stomatology, The Fourth Military Medical University, Xi’an, ChinacDepartment of Orthodontics, School of Stomatology, The Fourth Military Medical University, Xi’an, ChinadDepartment of Oral Surgery, School of Stomatology, The Fourth Military Medical University, Xi’an, ChinaeDepartment of Endodontics, School of Stomatology, The Fourth Military Medical University, Xi’an, China
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 1 8 – 1 1 2 4
a r t i c l e i n f o
Article history:
Accepted 26 September 2009
Keywords:
ClC-5
Dentin
Odontoblast
TGF-b1
Chloride channel
a b s t r a c t
ClC-5 is one of the voltage-dependent chloride channel (ClC) family members. Mutations
involving CLCN5 cause an X-linked nephropathy associated with Dent’s disease. Some
Clcn5 gene knockout (ClC-5 KO) mice have abnormal growth of the teeth; however, the
expression and function of ClC-5 during tooth development is still unknown. Herein we
report abnormal dentin structure, decreased DSPP and increased TGF-b1 protein level in
ClC-5 KO teeth. In odontoblast-like MDPC-23 cells, the mRNA levels of Tgfb1, Dspp and Dmp-
1 were upregulated with Clcn5 RNAi after 48 h treatment; whilst there was no change in
those of TGF-beta receptor Tgfbr1 and Tgfbr2. We suggest that the dentin changes in ClC-5
KO mice might be a result of increasing TGF-b1, and the interplay between ClC-5 and TGF-b1
needs further identified.
# 2009 Elsevier Ltd. All rights reserved.
avai lable at www.sc iencedi rec t .com
journal homepage: www.intl.elsevierhealth.com/journals/arob
1. Introduction
ClC-5 is one member of the ClC chloride channel family.1 The
CLCN5 gene mutates in Dent’s disease, an X-linked nephro-
pathy associated with low molecular weight proteinuria,
hypercalciura, nephrocalcinosis and nephrolithiasis.2,3
ClC-5 is predominantly expressed in the kidney,4 but ClC-5
is also present in the liver, brain, lung, testis, small intestine
and colon.2,5–7 In the studies by Wang et al.,8 an animal model
for Dent’s disease was developed by studying mice with
* Corresponding author. Tel.: +86 29 84776169; fax: +86 29 83223047.E-mail address: [email protected] (X. Duan).
1 These authors contributed equally to this paper.Abbreviations: ClCN5, Clcn5, chloride channel 5 gene; TGF-b1, tgfb
factor beta receptor; DSPP, Dspp, dentin sialophosphoprotein; DMP1, D1.0003–9969/$ – see front matter # 2009 Elsevier Ltd. All rights reservedoi:10.1016/j.archoralbio.2009.09.008
targeted inactivation of the corresponding murine gene, Clcn5.
The authors reported that a small number of mutant mice
‘‘had deformities of the dorsal spine and backward growth of
the teeth, but radiologic evidence for osteopenia or rickets was
absent’’8; the mechanism by which abnormal ClC-5 affected
tooth development was not elaborated on.
Chlorine is one of the abundant elements in teeth.
However, in contrast to enormous knowledge regarding
chloride channels in other organs, limited data pertaining to
chloride channel functions have been demonstrated in tooth
1, transforming growth factor, beta 1; Tgfbr, transforming growthmp1, dentin matrix acidic phosphoprotein 1, dentin matrix protein
d.

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 1 8 – 1 1 2 4 1119
development, with the exception of CFTR.9,10 Recently we
showed abundant ClC channels in tooth germ and odonto-
blast-like cells. Those chloride channels might regulate cell
proliferation and the cell cycle of odontoblast.11 In our
previous data, we also found the spatial and temporal
distribution of ClC-5 during rat tooth germ development.12,13
ClC-5 emerged in epithelial thickening stage, and mainly
located in ameloblasts and odontoblast during the bell stage,
and then disappeared in the late apposition stage.
To further explore the biological role of ClC-5 during tooth
development, herein we detected the morphological and
molecular characteristic of tooth germ and odontoblasts with
Clcn5 gene knockout (ClC-5 KO) mice and ClC-5 RNAi
technique. We tested the hypothesis that ClC-5 regulates
tooth growth, especially dentin formation, through the TGF-b1
pathway.
2. Materials and methods
2.1. Animals, cell lines and antibodies
Tooth germs samples of ClC-5 KO mice8 and wild-type C57BL/6
mice were kindly provided by Dr. Sandra E. Guggino from the
core store of Hopkins Digestive Disease Basic Research
Development Center (Johns Hopkins University, Baltimore,
MD, USA). Three litters of newborn wild-type C57BL/6 mice
from the Animal Center of the Fourth Military Medical
University (Xi’an, China) were used in the western analysis.
The animal studies were approved by the Animal Care and Use
Committee at the Fourth Military Medical University. The
mouse odontoblast-like cell line (MDPC-23) was provided by
Prof. C.T. Hanks and Dr. Jacques E. Nor at the University of
Michigan (MI, USA).
The antibodies used in this study were as follows: mouse
anti-TGF-b1 antibody (R&D Systems, Minneapolis, MN, USA);
monoclonal anti-b-actin antibody (A-5441; Sigma, St. Louis,
MO, USA), HRP-conjugated sheep anti-mouse antibodies
(Amersham Biosciences, Piscataway, NJ, USA); mouse anti-
DSPP antibody (provided by Dr. Rong Zhang in School of
Stomatology of the Fourth Military Medical University);
fluorescein (FITC) affini-pure donkey anti-mouse IgG (Jackson
ImmunoResearch, 715-095-1500); Hoechst 33342 (Sigma).
2.2. Histological studies
The appearance of incisors and molars were compared
between wild-type and gene knockout mice by general
observation among 20 adult ClC-5 KO/wild-type mice.
Three-day-old pups of the wild-type and knockout mice
were over-anaesthetized with halothane. The heads of the
mice were fixed in 4% paraformaldehyde in a 0.1 mol/L
phosphate buffer (pH 7.4). After fixation, the tissues were
processed by decalcification in 5% ethylene-diamine tetra-
acetic acid (EDTA; pH 7.4) for 5–10 days at 4 8C. The specimens
were dehydrated through a graded series of ethanol and
embedded in paraffin. Five mm sections were stained with H&E
and immunofluoresent staining. The ultrastructure of the
incisor samples from wild-type and ClC-5 KO mice were
compared with transmission electron microscopy (TEM,
200 kV, JEM2000, JEOL). Four pups of wild-type or ClC-5 KO
mice were respectively performed with HE staining and TEM
examinations. Genotyping was performed at the same time.
2.3. Western blot analysis
All the incisor and molar germs from three to four newborn
mice were dissected and collected as one group. There were
about 24–32 mixed incisors and molar germs in one group.
There were three ClC-5 KO groups and three wild-type groups
in each experiment. Dissected tooth germs from newborn
mice were collected at 4 8C in lysis buffer (1% NP-40, 0.5%
sodium deoxycholate and 0.1% sodium dodecyl sulphate [SDS]
in 0.01 mM phosphate-buffered saline [PBS]) containing
CompleteTM protease inhibitor (Roche Molecular Biochem-
icals, Mannheim, Germany). The protein samples were
separated on 12% SDS-polyacrylamide gel electrophoresis
(PAGE) and transferred to polyvinylidine difluoride (PVDF)
membranes (Bio-Rad). The western blot was performed with
mouse monoclonal anti-TGF-b1 antibody (1:50,000) and mouse
monoclonal anti-b-actin antibody (1:3000) respectively. The
secondary antibodies were HRP-conjugated sheep anti-mouse
antibodies (1:10,000). The blots were visualized using Western
Lightning Chemiluminescence Reagent Plus (Perkin Elmer Life
Sciences, Boston, MA, USA). Pixels on blots from three ClC-5
KO groups and three wild-type groups were measured using
Image J software (free from NIH website) for morphometric
analysis.
2.4. Immunofluoresent staining of tooth germ
In order to detect the protein level of DSPP and TGF-b1 in tooth
germ, 5 mm sections from tooth germs of wild-type and ClC-5
KO mice were stained with mouse anti-DSPP and mouse anti-
TGF-b1 antibody respectively, followed by staining with
fluorescein (FITC) affini-pure donkey anti-mouse IgG and
Hoechst 33342. A FluoView FV1000 confocal microscope was
used to capture pictures.
2.5. Clcn5 RNA interference
Mouse odontoblast-like cells (MDPC-23) were cultured in a-
MEM medium (Gibco, Grand Island, NY, USA) supplemented
with 10% fetal bovine serum (FBS) (Gibco), 2 mmol/L L-
glutamine, 50 mg/mL of vitamin C (Sigma), 100 IU/mL of
penicillin and 100 mg/mL of streptomycin in a humidified
atmosphere containing 5% CO2 in an incubator at 37 8C.11
The pENTR-Clcn5 27 was designed to target positions 27–47
of the complete mouse Clcn5 nucleotide sequence (Genbank
accession # NM_016691) with pENTRTM/U6 (Invitrogen), the
related short hairpin loop sequences were as follows:
NM_shrna_27_top CAC CGC TGT CAA GCC GTG TTC TAG ACG
AAT CTA GAA CAC GGC TTG ACA GC; and NM _shrna_27_-
bottom AAA AGC TGT CAA GCC GTG TTC TAG ATT CGT CTA
GAA CAC GGC TTG ACA GC. Another good construct pENTR-
Clcn5 2638 (targeting positions 2638–2658 of the complete
mouse Clcn5 nucleotide sequence, NM_016691) also showed a
similar reduction of Clcn5 mRNA level as pENTR-Clcn5 27.
Finally we chose pENTR-Clcn5 27 as the best RNAi construct in
this experiment. pENTR-LacZ was made as the RNAi control.
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 1 8 – 1 1 2 41120
The pENTR-Clcn5 27 was transfected into MDPC-23 cells with
Lipofectin 2000. The mRNA levels of Clcn5 and other genes were
detected by real-time PCR after a 48 h transfection.
2.6. Quantitative reverse-transcriptase/polymerasechain-reaction (RT-PCR)
Total RNAs of cells were isolated and a M-MLV reverse
transcriptase cDNA synthesis Kit (Takala) was used to synthe-
size cDNA. The primers for detecting Clcn5, Tgfb1, Tgfbr1,
Tgfbr2, Dspp, Dmp1 and Gapdh are shown in Table 1. Real-time
reverse-transcriptase-PCR (RT-PCR) analyses were performed
with SBrilliant1 SYBR1 Green QPCR Master Mix (Takala)
according to the company’s suggested procedures. PCR condi-
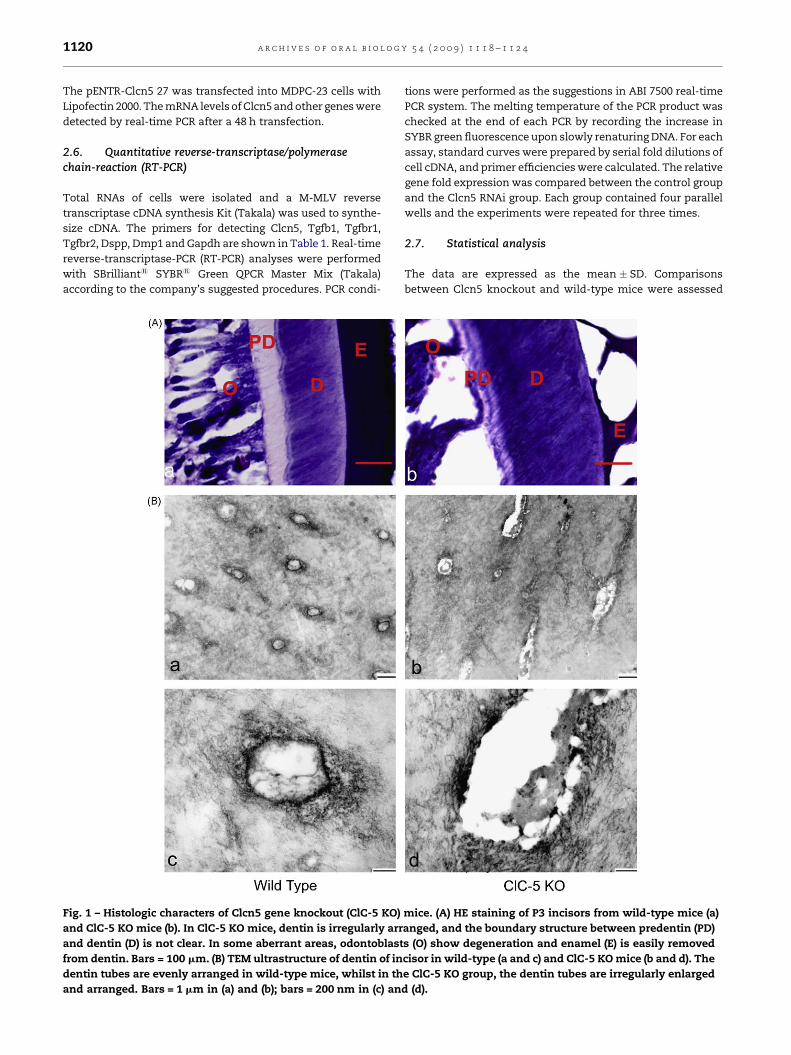
Fig. 1 – Histologic characters of Clcn5 gene knockout (ClC-5 KO)
and ClC-5 KO mice (b). In ClC-5 KO mice, dentin is irregularly arr
and dentin (D) is not clear. In some aberrant areas, odontoblast
from dentin. Bars = 100 mm. (B) TEM ultrastructure of dentin of in
dentin tubes are evenly arranged in wild-type mice, whilst in th
and arranged. Bars = 1 mm in (a) and (b); bars = 200 nm in (c) an
tions were performed as the suggestions in ABI 7500 real-time
PCR system. The melting temperature of the PCR product was
checked at the end of each PCR by recording the increase in
SYBR green fluorescence upon slowly renaturing DNA. For each
assay, standard curves were prepared by serial fold dilutions of
cell cDNA, and primer efficiencies were calculated. The relative
gene fold expression was compared between the control group
and the Clcn5 RNAi group. Each group contained four parallel
wells and the experiments were repeated for three times.
2.7. Statistical analysis
The data are expressed as the mean � SD. Comparisons
between Clcn5 knockout and wild-type mice were assessed
mice. (A) HE staining of P3 incisors from wild-type mice (a)
anged, and the boundary structure between predentin (PD)
s (O) show degeneration and enamel (E) is easily removed
cisor in wild-type (a and c) and ClC-5 KO mice (b and d). The
e ClC-5 KO group, the dentin tubes are irregularly enlarged
d (d).
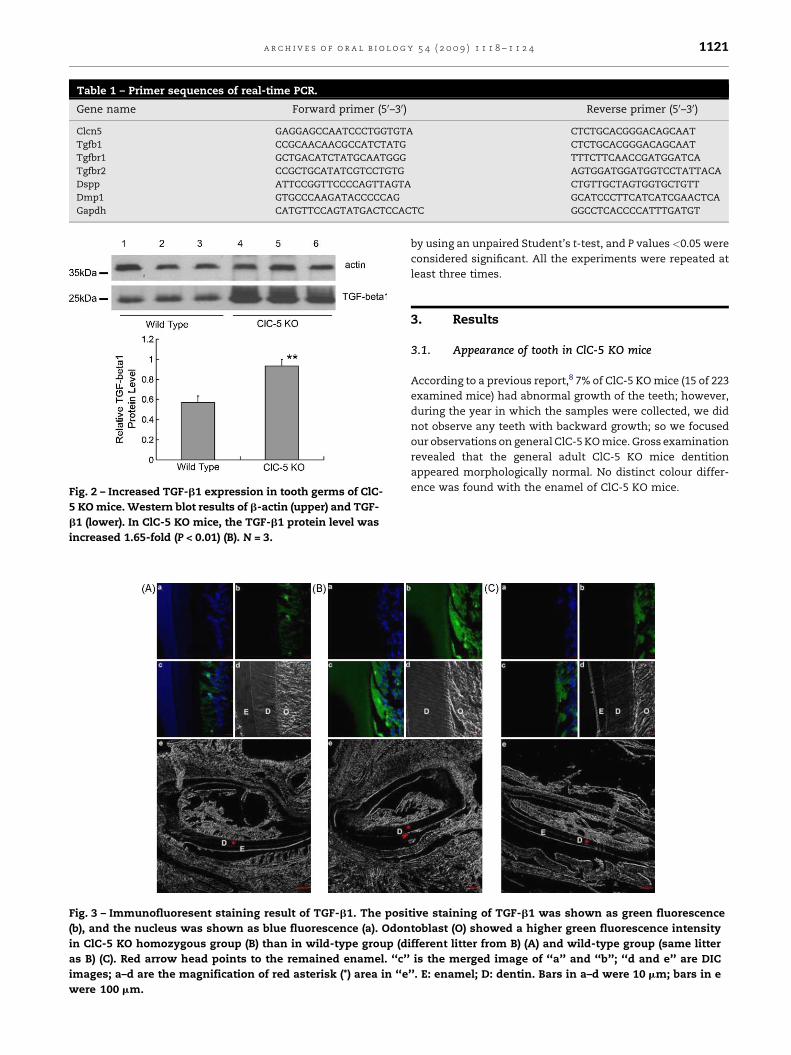
Fig. 2 – Increased TGF-b1 expression in tooth germs of ClC-
5 KO mice. Western blot results of b-actin (upper) and TGF-
b1 (lower). In ClC-5 KO mice, the TGF-b1 protein level was
increased 1.65-fold (P < 0.01) (B). N = 3.
Fig. 3 – Immunofluoresent staining result of TGF-b1. The posi
(b), and the nucleus was shown as blue fluorescence (a). Odon
in ClC-5 KO homozygous group (B) than in wild-type group (d
as B) (C). Red arrow head points to the remained enamel. ‘‘c’’
images; a–d are the magnification of red asterisk (*) area in ‘‘e
were 100 mm.
Table 1 – Primer sequences of real-time PCR.
Gene name Forward primer (50–30) Reverse primer (50–30)
Clcn5 GAGGAGCCAATCCCTGGTGTA CTCTGCACGGGACAGCAAT
Tgfb1 CCGCAACAACGCCATCTATG CTCTGCACGGGACAGCAAT
Tgfbr1 GCTGACATCTATGCAATGGG TTTCTTCAACCGATGGATCA
Tgfbr2 CCGCTGCATATCGTCCTGTG AGTGGATGGATGGTCCTATTACA
Dspp ATTCCGGTTCCCCAGTTAGTA CTGTTGCTAGTGGTGCTGTT
Dmp1 GTGCCCAAGATACCCCCAG GCATCCCTTCATCATCGAACTCA
Gapdh CATGTTCCAGTATGACTCCACTC GGCCTCACCCCATTTGATGT
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 1 8 – 1 1 2 4 1121
by using an unpaired Student’s t-test, and P values <0.05 were
considered significant. All the experiments were repeated at
least three times.
3. Results
3.1. Appearance of tooth in ClC-5 KO mice
According to a previous report,8 7% of ClC-5 KO mice (15 of 223
examined mice) had abnormal growth of the teeth; however,
during the year in which the samples were collected, we did
not observe any teeth with backward growth; so we focused
our observations on general ClC-5 KO mice. Gross examination
revealed that the general adult ClC-5 KO mice dentition
appeared morphologically normal. No distinct colour differ-
ence was found with the enamel of ClC-5 KO mice.
tive staining of TGF-b1 was shown as green fluorescence
toblast (O) showed a higher green fluorescence intensity
ifferent litter from B) (A) and wild-type group (same litter
is the merged image of ‘‘a’’ and ‘‘b’’; ‘‘d and e’’ are DIC
’’. E: enamel; D: dentin. Bars in a–d were 10 mm; bars in e

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 1 8 – 1 1 2 41122
3.2. Abnormal dentin in ClC-5 KO tooth germs
Histological evaluation of both incisors and associated
enamel showed no apparent difference between ClC-5 KO
mice and normal mice, whilst the dentin demonstrated
structural abnormalities in the ClC-5 KO mice. In wild-type
mice, the mantle dentin layer was continuous with a
narrow submantle band of tubules. The dentin zone ended
abruptly in a broad, undulating zone parallel to the dentin–
enamel junction (DEJ). ClC-5 KO mice presented an unclear
boundary between dentin and predentin. The tubular
structures were ill-defined. Enamel was easily removed
from dentin in some severe cases (Fig. 1A). The abnormal
extent of enamel and dentin varied a little among different
ClC-5 KO teeth. Transmission electron microscopy (TEM)
results showed the enlarged channel-like structures in the
dentin of ClC-5 KO mice. The shape of dentin tubules was
irregular (Fig. 1B).
We found that enamel was easily removed from dentin
during tissue section preparing in ClC-5 KO tooth germ
(Figs. 3Be and 4Be). In order to avoid that enamel’s peeling
off was due to the histological artefact, we compared tooth
germs of wild-type mice and ClC-5 KO homozygous mice in
the same litter. These two groups followed the exact same
histological procedure. The detachment of enamel was only
found in ClC-5 KO teeth.
Fig. 4 – Immunofluoresent staining result of DSPP. The positive
(b), and the nucleus was shown as blue immunofluorescence (a
intensity in ClC-5 KO homozygous group (B) than in wild-type g
litter as B) (C). Red arrow head points to the detached enamel. ‘‘c’
a–d are the magnification of red asterisk (*) area in ‘‘e’’. E: ename
3.3. Increased TGF-b1 and decreased DSPP in ClC-5 KOtooth germs
To understand the mechanism by which ClC-5 regulates
tooth development, we used western blot to detect
TGF-b1 in tooth germs. Three-day-old ClC-5 KO newborn
mice had much greater amounts of TGF-b1 (1.65-fold,
P < 0.01) in total tooth germ lysates compared with 3-day-
old wild-type mice (Fig. 2A and B). Immunofluoresent
staining results showed that odontoblasts in ClC-5 KO
tooth germ showed higher expression of TGF-b1 and
lower expression of DSPP than in wild-type group (Figs. 3
and 4).
3.4. Expression of Tgfb1, Tgfbr1, Tgfbr2, Dspp and Dmp1with Clcn5 in the MDPC-23 cell line
We confirmed the expression of Clcn5 mRNA in MDPC-23
cells.11 The RNAi method was used to knockdown the
expression of Clcn5. The mRNA of Tgfb1, Tgfbr1, Tgfbr2, Dspp
and Dmp1 were analyzed by real-time PCR. We found that the
Clcn5 RNAi construct reduced the expression of Clcn5 mRNA
expression about 81.01% in MDPC-23 cells (P < 0.01), and at the
same time, the mRNA level of Tgfb1 was upregulated 1.87-fold
(P < 0.05). To our surprise, the mRNA level of Dspp and Dmp1
was also increased in RNAi treatment group (P < 0.01). No
staining of DSPP was shown as green immunofluorescence
). Odontoblast (O) showed a lower green fluorescence
roup (different litter from B) (A) and wild-type group (same
’ is the merged image of ‘‘a’’ and ‘‘b’’; ‘‘d, e’’ are DIC images;
l; D: dentin. Bars in a–d were 10 mm; bars in e were 100 mm.
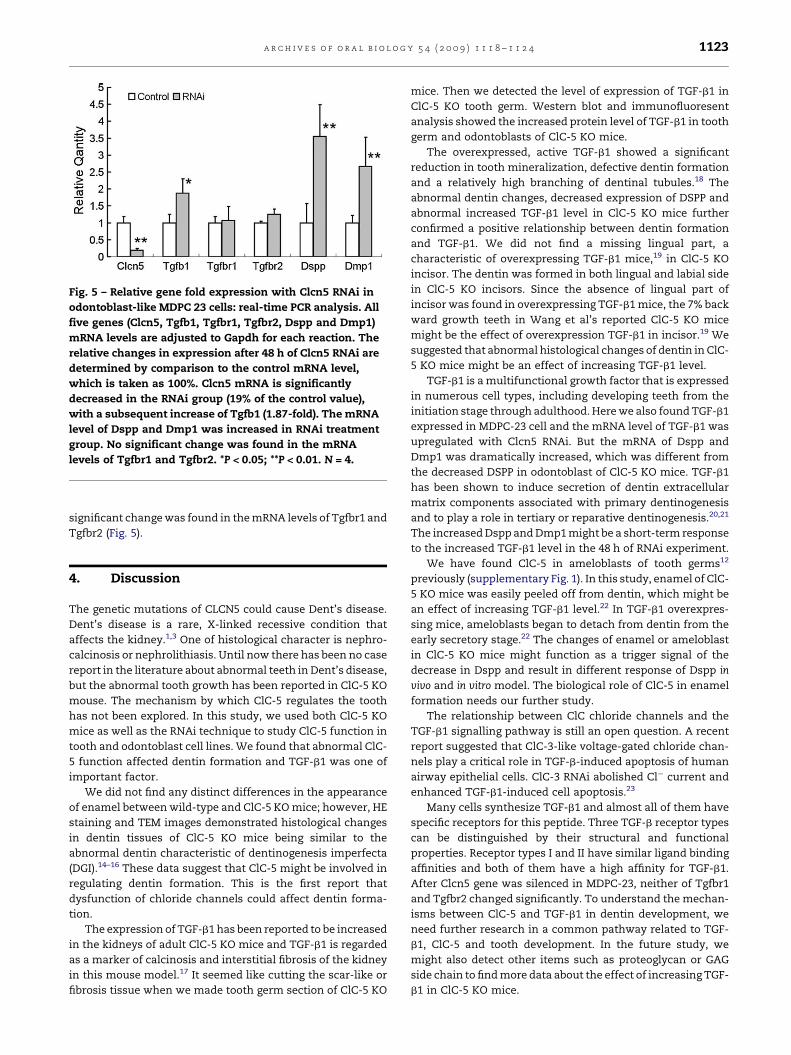
Fig. 5 – Relative gene fold expression with Clcn5 RNAi in
odontoblast-like MDPC 23 cells: real-time PCR analysis. All
five genes (Clcn5, Tgfb1, Tgfbr1, Tgfbr2, Dspp and Dmp1)
mRNA levels are adjusted to Gapdh for each reaction. The
relative changes in expression after 48 h of Clcn5 RNAi are
determined by comparison to the control mRNA level,
which is taken as 100%. Clcn5 mRNA is significantly
decreased in the RNAi group (19% of the control value),
with a subsequent increase of Tgfb1 (1.87-fold). The mRNA
level of Dspp and Dmp1 was increased in RNAi treatment
group. No significant change was found in the mRNA
levels of Tgfbr1 and Tgfbr2. *P < 0.05; **P < 0.01. N = 4.
a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 1 8 – 1 1 2 4 1123
significant change was found in the mRNA levels of Tgfbr1 and
Tgfbr2 (Fig. 5).
4. Discussion
The genetic mutations of CLCN5 could cause Dent’s disease.
Dent’s disease is a rare, X-linked recessive condition that
affects the kidney.1,3 One of histological character is nephro-
calcinosis or nephrolithiasis. Until now there has been no case
report in the literature about abnormal teeth in Dent’s disease,
but the abnormal tooth growth has been reported in ClC-5 KO
mouse. The mechanism by which ClC-5 regulates the tooth
has not been explored. In this study, we used both ClC-5 KO
mice as well as the RNAi technique to study ClC-5 function in
tooth and odontoblast cell lines. We found that abnormal ClC-
5 function affected dentin formation and TGF-b1 was one of
important factor.
We did not find any distinct differences in the appearance
of enamel between wild-type and ClC-5 KO mice; however, HE
staining and TEM images demonstrated histological changes
in dentin tissues of ClC-5 KO mice being similar to the
abnormal dentin characteristic of dentinogenesis imperfecta
(DGI).14–16 These data suggest that ClC-5 might be involved in
regulating dentin formation. This is the first report that
dysfunction of chloride channels could affect dentin forma-
tion.
The expression of TGF-b1 has been reported to be increased
in the kidneys of adult ClC-5 KO mice and TGF-b1 is regarded
as a marker of calcinosis and interstitial fibrosis of the kidney
in this mouse model.17 It seemed like cutting the scar-like or
fibrosis tissue when we made tooth germ section of ClC-5 KO
mice. Then we detected the level of expression of TGF-b1 in
ClC-5 KO tooth germ. Western blot and immunofluoresent
analysis showed the increased protein level of TGF-b1 in tooth
germ and odontoblasts of ClC-5 KO mice.
The overexpressed, active TGF-b1 showed a significant
reduction in tooth mineralization, defective dentin formation
and a relatively high branching of dentinal tubules.18 The
abnormal dentin changes, decreased expression of DSPP and
abnormal increased TGF-b1 level in ClC-5 KO mice further
confirmed a positive relationship between dentin formation
and TGF-b1. We did not find a missing lingual part, a
characteristic of overexpressing TGF-b1 mice,19 in ClC-5 KO
incisor. The dentin was formed in both lingual and labial side
in ClC-5 KO incisors. Since the absence of lingual part of
incisor was found in overexpressing TGF-b1 mice, the 7% back
ward growth teeth in Wang et al’s reported ClC-5 KO mice
might be the effect of overexpression TGF-b1 in incisor.19 We
suggested that abnormal histological changes of dentin in ClC-
5 KO mice might be an effect of increasing TGF-b1 level.
TGF-b1 is a multifunctional growth factor that is expressed
in numerous cell types, including developing teeth from the
initiation stage through adulthood. Here we also found TGF-b1
expressed in MDPC-23 cell and the mRNA level of TGF-b1 was
upregulated with Clcn5 RNAi. But the mRNA of Dspp and
Dmp1 was dramatically increased, which was different from
the decreased DSPP in odontoblast of ClC-5 KO mice. TGF-b1
has been shown to induce secretion of dentin extracellular
matrix components associated with primary dentinogenesis
and to play a role in tertiary or reparative dentinogenesis.20,21
The increased Dspp and Dmp1 might be a short-term response
to the increased TGF-b1 level in the 48 h of RNAi experiment.
We have found ClC-5 in ameloblasts of tooth germs12
previously (supplementary Fig. 1). In this study, enamel of ClC-
5 KO mice was easily peeled off from dentin, which might be
an effect of increasing TGF-b1 level.22 In TGF-b1 overexpres-
sing mice, ameloblasts began to detach from dentin from the
early secretory stage.22 The changes of enamel or ameloblast
in ClC-5 KO mice might function as a trigger signal of the
decrease in Dspp and result in different response of Dspp in
vivo and in vitro model. The biological role of ClC-5 in enamel
formation needs our further study.
The relationship between ClC chloride channels and the
TGF-b1 signalling pathway is still an open question. A recent
report suggested that ClC-3-like voltage-gated chloride chan-
nels play a critical role in TGF-b-induced apoptosis of human
airway epithelial cells. ClC-3 RNAi abolished Cl� current and
enhanced TGF-b1-induced cell apoptosis.23
Many cells synthesize TGF-b1 and almost all of them have
specific receptors for this peptide. Three TGF-b receptor types
can be distinguished by their structural and functional
properties. Receptor types I and II have similar ligand binding
affinities and both of them have a high affinity for TGF-b1.
After Clcn5 gene was silenced in MDPC-23, neither of Tgfbr1
and Tgfbr2 changed significantly. To understand the mechan-
isms between ClC-5 and TGF-b1 in dentin development, we
need further research in a common pathway related to TGF-
b1, ClC-5 and tooth development. In the future study, we
might also detect other items such as proteoglycan or GAG
side chain to find more data about the effect of increasing TGF-
b1 in ClC-5 KO mice.

a r c h i v e s o f o r a l b i o l o g y 5 4 ( 2 0 0 9 ) 1 1 1 8 – 1 1 2 41124
In summary, here we showed abnormal dentin structure in
ClC-5 KO mice and we suggest the abnormal ClC-5 function
affects dentin formation through TGF-b1 pathway.
Acknowledgements
We thank Sandra E. Guggino of Johns Hopkins University (JHU)
for providing samples of Clcn5 gene knockout mice, William B.
Guggino (JHU) for helpful suggestions, as well as Jennifer Sipes
(JHU) for technical support. We are also thankful for support
from the National Natural Science Foundation of China
(30371540).
Funding: National Natural Science Foundation of China
(30371540).
Competing interests: None declared.
Ethical approval: Not required.
Appendix A. Supplementary data
Supplementary data associated with this article can be
found, in the online version, at doi:10.1016/j.archoral-
bio.2009.09.008.
r e f e r e n c e s
1. Jentsch TJ, Stein V, Weinreich F, Zdebik AA. Molecularstructure and physiological function of chloride channels.Physiol Rev 2002;82(2):503–68.
2. Edmonds RD, Silva IV, Guggino WB, Butler RB, Zeitlin PL,Blaisdell CJ. ClC-5: ontogeny of an alternative chloridechannel in respiratory epithelia. Am J Physiol Lung Cell MolPhysiol 2002;282(3):L501–7.
3. Lloyd SE, Pearce SH, Fisher SE, Steinmeyer K, Schwappach B,Scheinman SJ, et al. A common molecular basis for threeinherited kidney stone diseases. Nature 1996;379(6564):445–9.
4. Takeuchi Y, Uchida S, Marumo F, Sasaki S. Cloning, tissuedistribution, and intrarenal localization of ClC chloridechannels in human kidney. Kidney Int 1995;48(5):1497–503.
5. Cornejo I, Niemeyer MI, Sepulveda FV, Cid LP. Cloning,cellular distribution and functional expression of smallintestinal epithelium guinea pig ClC-5 chloride channel.Biochim Biophys Acta 2001;1512(2):367–74.
6. Scherer CR, Linz W, Busch AE, Steinmeyer K. Geneexpression profiles of CLC chloride channels in animalmodels with different cardiovascular diseases. Cell PhysiolBiochem 2001;11(6):321–30.
7. Vandewalle A, Cluzeaud F, Peng KC, Bens M, Luchow A,Gunther W, et al. Tissue distribution and subcellularlocalization of the ClC-5 chloride channel in rat intestinalcells. Am J Physiol Cell Physiol 2001;280(2):C373–81.
8. Wang SS, Devuyst O, Courtoy PJ, Wang XT, Wang H, Wang Y,et al. Mice lacking renal chloride channel CLC-5, are a modelfor Dent’s disease, a nephrolithiasis disorder associated
with defective receptor-mediated endocytosis. Hum MolGenet 2000;9(20):2937–45.
9. Wright JT, Kiefer CL, Hall KI, Grubb BR. Abnormal enameldevelopment in a cystic fibrosis transgenic mouse model. JDent Res 1996;75(4):966–73.
10. Wright JT, Hall KI, Grubb BR. Enamel mineral composition ofnormal and cystic fibrosis transgenic mice. Adv Dent Res1996;10(2):270–4.
11. Hou J, Situ Z, Duan X. ClC chloride channels in tooth germand odontoblast-like MDPC-23 cells. Arch Oral Biol2008;53(9):874–8.
12. Duan XH. Spatial–temporal distribution of CLC-5 in rat toothgerm development. J Dent Res 2004;83(special issue A):2741.
13. Hou J, Duan XH, Si-Tu ZQ. Expression of CIC-5 in rat toothgerm development. Hua Xi Kou Qiang Yi Xue Za Zhi2007;25(5):444–6. 453.
14. Lindau B, Dietz W, Lundgren T, Storhaug K, Noren JG.Discrimination of morphological findings in dentine fromosteogenesis imperfecta patients using combinations ofpolarized light microscopy, microradiography andscanning electron microscopy. Int J Paediatr Dent 1999;9(4):253–61.
15. Malmgren B, Lundberg M, Lindskog S. Dentinogenesisimperfecta in a six-generation family A clinical,radiographic and histologic comparison of twobranches through three generations. Swed Dent J1988;12(3):73–84.
16. Siar CH. Quantitative histological analysis of the humancoronal dentine in dentinogenesis imperfecta types I and II.Arch Oral Biol 1986;31(6):387–90.
17. Cebotaru V, Kaul S, Devuyst O, Cai H, Racusen L, GugginoWB, et al. High citrate diet delays progression of renalinsufficiency in the ClC-5 knockout mouse model of Dent’sdisease. Kidney Int 2005;68(2):642–52.
18. Thyagarajan T, Sreenath T, Cho A, Wright JT, Kulkarni AB.Reduced expression of dentin sialophosphoprotein isassociated with dysplastic dentin in mice overexpressingtransforming growth factor-beta 1 in teeth. J Biol Chem2001;276(14):11016–20.
19. Opsahl S, Septier D, Aubin I, Guenet JL, Sreenath T, KulkarniA, et al. Is the lingual forming part of the incisor a structuralentity? Evidences from the fragilitas ossium (fro/fro) mousemutation and the TGFbeta1 overexpressing transgenicstrain. Arch Oral Biol 2005;50(2):279–86.
20. Unterbrink A, O’Sullivan M, Chen S, MacDougall M. TGFbeta-1 downregulates DMP-1 and DSPP in odontoblasts.Connect Tissue Res 2002;43(2–3):354–8.
21. Magne D, Bluteau G, Lopez-Cazaux S, Weiss P, Pilet P,Ritchie HH, et al. Development of an odontoblast in vitromodel to study dentin mineralization. Connect Tissue Res2004;45(2):101–8.
22. Haruyama N, Thyagarajan T, Skobe Z, Wright JT, Septier D,Sreenath TL, et al. Overexpression of transforminggrowth factor-beta1 in teeth results in detachment ofameloblasts and enamel defects. Eur J Oral Sci 2006;114(Suppl 1):30–4.
23. Cheng G, Shao Z, Chaudhari B, Agrawal DK. Involvement ofchloride channels in TGF-beta1-induced apoptosis ofhuman bronchial epithelial cells. Am J Physiol Lung Cell MolPhysiol 2007;293(5):L1339–47.

本文献由“学霸图书馆-文献云下载”收集自网络,仅供学习交流使用。
学霸图书馆(www.xuebalib.com)是一个“整合众多图书馆数据库资源,
提供一站式文献检索和下载服务”的24 小时在线不限IP
图书馆。
图书馆致力于便利、促进学习与科研,提供最强文献下载服务。
图书馆导航:
图书馆首页 文献云下载 图书馆入口 外文数据库大全 疑难文献辅助工具


















