
Chronic myelogenous leukemia primitive hematopoietic progenitors demonstrate increased sensitivity...
12
Experimental Hematology 28 (2000) 1401–1412 0301-472X/00 $–see front matter. Copyright © 2000 International Society for Experimental Hematology. Published by Elsevier Science Inc. PII S0301-472X(00)00545-2 Chronic myelogenous leukemia primitive hematopoietic progenitors demonstrate increased sensitivity to growth factor–induced proliferation and maturation Ravi Bhatia a , Heidi A. Munthe a , Andrea D. Williams a , Feiyu Zhang b , Stephen J. Forman a , and Marilyn L. Slovak b a Division of Hematology and Bone Marrow Transplantation, City of Hope National Medical Center, Duarte, Calif., USA and b Cytogenetics Laboratory, Division of Anatomic Pathology, City of Hope National Medical Center, Duarte, Calif., USA (Received 2 May 2000; revised 7 July 2000; accepted 11 August 2000) Objective. We investigated whether primary chronic myelogenous leukemia (CML) hemato- poietic progenitors demonstrated altered proliferation and maturation in response to growth factor (GF) stimulation. Methods. The effect of GF stimulation on proliferation and expansion of committed and prim- itive progenitors (colony forming cells [CFC]) was evaluated. Results. Culture of CML and normal CD34 1 cells with different GF for 7 days resulted in sim- ilar expansion of committed progenitors (CFC). In contrast, GF culture conditions that ex- panded normal primitive progenitors (week-6 long-term culture-initiating cells (LTC-IC)] led to depletion of CML LTC-IC numbers. GF culture also resulted in increased depletion of week-10 extended LTC-IC, which represent an even more primitive progenitor population, from CML compared with normal CD34 1 cells. CML CD34 1 cells enter into cycle more quickly than normal CD34 1 cells and CML CFC expansion was accelerated compared to nor- mal CFC. Evaluation of primitive progenitor proliferation using PKH-26 and single-cell LTC- IC analysis demonstrated that the majority of CML LTC-IC remaining after GF culture orig- inated from divided CD34 1 cells, whereas GF-cultured normal LTC-IC were derived mainly from undivided cells. Depletion of CML primitive progenitor numbers in association with in- creased proliferation suggests increased sensitivity to GF-induced maturation. Conclusions. These studies indicate that CML primitive progenitors have enhanced sensitivity to GF-induced cell division and maturation. Altered GF responsiveness may contribute to ab- normal expansion of malignant myeloid cells in CML. These findings may also be applied to- ward the development of novel approaches to select benign stem cells in CML. © 2000 Inter- national Society for Experimental Hematology. Published by Elsevier Science Inc. Keywords: Chronic myelogenous leukemia—Growth factors—Stem cells—Proliferation— Differentiation Introduction Chronic myelogenous leukemia (CML) is a clonal hemato- poietic disorder resulting from the malignant transformation of a hematopoietic stem cell. Malignant cells in CML con- tain the 9; 22 translocation, which results in the formation of the BCR/ABL fusion oncogene and expression of the p210 BCR/ABL protein [1,2]. In vitro and in vivo experimental evidence clearly indicates that the BCR/ABL chimeric gene plays a critical role in the pathogenesis of CML [3,4]. The initial chronic phase of CML is characterized by a vast ex- pansion of hematopoietic progenitor, precursor, and mature cells, suggesting an increase in progenitor proliferation with retention of differentiation capacity. Other important fea- tures of CML are the presence of increased numbers of cir- culating progenitors and extramedullary hematopoiesis, suggesting abnormalities in progenitor adhesion and/or mi- gration. Although the disease predominantly involves mye- loid cells, especially of neutrophil lineage, the BCR/ABL gene can also be detected in thrombocytic, erythrocytic, dendritic, natural killer, and B cell lineages ( z 30%) and, in some studies, in a fraction of T lymphocytes [5–7]. These observations indicate that the disease arises in a very primi- Offprint requests to: Ravi Bhatia, M.D., Department of Hematology/ BMT, City of Hope National Medical Center, 1500 E Duarte Road, Duarte, CA 91010; E-mail: [email protected]
-
Upload
ravi-bhatia -
Category
Documents
-
view
213 -
download
1
Transcript of Chronic myelogenous leukemia primitive hematopoietic progenitors demonstrate increased sensitivity...

Experimental Hematology 28 (2000) 1401–1412
0301-472X/00 $–see front matter. Copyright © 2000 International Society for Experimental Hematology. Published by Elsevier Science Inc.PII S0301-472X(00)00545-2
Chronic myelogenous leukemia primitivehematopoietic progenitors demonstrate increased
sensitivity to growth factor–induced proliferation and maturation
Ravi Bhatia
a
, Heidi A. Munthe
a
, Andrea D. Williams
a
,Feiyu Zhang
b
, Stephen J. Forman
a
, and Marilyn L. Slovak
b
a
Division of Hematology and Bone Marrow Transplantation, City of Hope National Medical Center, Duarte, Calif., USAand
b
Cytogenetics Laboratory, Division of Anatomic Pathology, City of Hope National Medical Center, Duarte, Calif., USA
(Received 2 May 2000; revised 7 July 2000; accepted 11 August 2000)
Objective.
We investigated whether primary chronic myelogenous leukemia (CML) hemato-poietic progenitors demonstrated altered proliferation and maturation in response to growthfactor (GF) stimulation.
Methods.
The effect of GF stimulation on proliferation and expansion of committed and prim-itive progenitors (colony forming cells [CFC]) was evaluated.
Results.
Culture of CML and normal CD34
1
cells with different GF for 7 days resulted in sim-ilar expansion of committed progenitors (CFC). In contrast, GF culture conditions that ex-panded normal primitive progenitors (week-6 long-term culture-initiating cells (LTC-IC)] ledto depletion of CML LTC-IC numbers. GF culture also resulted in increased depletion ofweek-10 extended LTC-IC, which represent an even more primitive progenitor population,from CML compared with normal CD34
1
cells. CML CD34
1
cells enter into cycle morequickly than normal CD34
1
cells and CML CFC expansion was accelerated compared to nor-mal CFC. Evaluation of primitive progenitor proliferation using PKH-26 and single-cell LTC-IC analysis demonstrated that the majority of CML LTC-IC remaining after GF culture orig-inated from divided CD34
1
cells, whereas GF-cultured normal LTC-IC were derived mainlyfrom undivided cells. Depletion of CML primitive progenitor numbers in association with in-creased proliferation suggests increased sensitivity to GF-induced maturation.
Conclusions.
These studies indicate that CML primitive progenitors have enhanced sensitivityto GF-induced cell division and maturation. Altered GF responsiveness may contribute to ab-normal expansion of malignant myeloid cells in CML. These findings may also be applied to-ward the development of novel approaches to select benign stem cells in CML. © 2000 Inter-national Society for Experimental Hematology. Published by Elsevier Science Inc.
Keywords:
Chronic myelogenous leukemia—Growth factors—Stem cells—Proliferation—Differentiation
Introduction
Chronic myelogenous leukemia (CML) is a clonal hemato-poietic disorder resulting from the malignant transformationof a hematopoietic stem cell. Malignant cells in CML con-tain the 9; 22 translocation, which results in the formation ofthe BCR/ABL fusion oncogene and expression of thep210
BCR/ABL
protein [1,2]. In vitro and in vivo experimentalevidence clearly indicates that the BCR/ABL chimeric geneplays a critical role in the pathogenesis of CML [3,4]. The
initial chronic phase of CML is characterized by a vast ex-pansion of hematopoietic progenitor, precursor, and maturecells, suggesting an increase in progenitor proliferation withretention of differentiation capacity. Other important fea-tures of CML are the presence of increased numbers of cir-culating progenitors and extramedullary hematopoiesis,suggesting abnormalities in progenitor adhesion and/or mi-gration. Although the disease predominantly involves mye-loid cells, especially of neutrophil lineage, the BCR/ABLgene can also be detected in thrombocytic, erythrocytic,
dendritic, natural killer, and B cell lineages (
z
30%) and, insome studies, in a fraction of T lymphocytes [5–7]. Theseobservations indicate that the disease arises in a very primi-
Offprint requests to: Ravi Bhatia, M.D., Department of Hematology/BMT, City of Hope National Medical Center, 1500 E Duarte Road, Duarte,CA 91010; E-mail: [email protected]

1402
R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412
tive, pluripotent hematopoietic cell. The disease invariablyprogresses to a terminal acute leukemic phase after a me-dian of 4 years. Either acute lymphoblastic or myeloblasticleukemia can evolve, providing further evidence that CMLarises in a primitive pluripotent cell. However, it is unclearwhy selective expansion of only myeloid lineage cells oc-curs in chronic phase CML.
Potential explanations for the enhanced proliferation ofmalignant myeloid progenitors in CML include 1) reducedsensitivity to growth inhibitory stimuli and/or 2) enhancedsensitivity to growth stimulatory factors. CML progenitorshave been reported to be insensitive to inhibition of prolifer-ation by chemokines, such as macrophage inflammatory pro-tein-1
a
and macrophage chemoattractant protein-1, that in-hibit normal progenitor proliferation [8,9]. In addition, CMLprogenitors, although expressing normal levels of
b
1-inte-grin adhesion receptors, demonstrate deficient
b
1-integrin–mediated adhesion and signaling [10]. Since integrin recep-tor engagement leads to inhibition of normal progenitorproliferation, abnormalities in integrin function in CML pro-genitors may contribute to the abnormal expansion of malig-nant cells in CML in addition to their abnormal circulation[11,12]. Introduction of BCR/ABL into factor-dependent he-matopoietic cell lines can result in growth factor indepen-dence for survival and proliferation [13,14]. The cellularcontext and level of BCR/ABL expression play crucial rolesin determining the transformation capacity of BCR/ABL andthe downstream signaling pathways activated [15]. Althoughsome studies have found primary CML hematopoietic pro-genitors to be growth factor independent for proliferationand survival [14,16], this has not been confirmed in otherstudies [17,18]. In addition, some studies suggest that CMLCD34
1
cells may terminally differentiate at concentrationsthat allow normal progenitor proliferation and exit G0/G1more rapidly compared to normal CD34
1
cells followinggrowth factor (GF) stimulation [13,19]. These observationssuggest that CML-committed progenitors may have increasedsensitivity to GF-induced proliferative and maturation sig-naling. Since CML arises in a very primitive multipotent he-matopoietic cell, it is important to determine whether CMLprimitive progenitors also demonstrate altered GF respon-siveness, especially since the GF response may differ de-pending on maturation stage. In a previous study, Petzer etal. made the intriguing observation that GF combinationsthat expanded normal long-term culture-initiating cell (LTC-IC) numbers resulted in depletion of malignant CML LTC-IC [20]. However the mechanism underlying depletion ofCML primitive progenitors following GF culture are un-clear. This effect has been attributed to decreased self-renewing divisions, related to autocrine production of granu-locyte colony-stimulating factor (G-CSF) and interleukin-3(IL-3) by CML CD34
1
CD38
2
cells [21]. However, previousstudies have not directly evaluated whether primitive CMLprogenitors have increased, normal or reduced propensity toproliferate in response to GF stimulation.
The objectives of the present study were to investigatewhether primary primitive CML hematopoietic progenitorsdemonstrated altered proliferation and maturation in re-sponse to growth factor stimulation. We performed a de-tailed analysis of the effect of culture with single GFs andGF combinations, at different concentrations and lengths ofexposure, on CML committed and primitive progenitors. Weconfirmed that GF conditions that expanded normal primi-tive progenitor numbers resulted in depletion of CML primi-tive progenitors. We directly evaluated the proliferative re-sponse of CML progenitors to GF stimulation using cellcycle analysis, single cell assays, and PKH-26 staining. Ourresults indicate that CML primitive, and committed, progen-itors demonstrate increased sensitivity to GF-induced cell di-vision. Since GF stimulation also resulted in reduction innumbers of primitive progenitors, this suggests that CMLprimitive progenitors have increased sensitivity to GF-induced maturation as well. These observations enhance ourunderstanding of mechanisms underlying abnormal myeloidexpansion in CML and have implications for development ofnovel therapeutic approaches to this disease.
Materials and methods
Subjects
Chronic myelogenous leukemia patients (n
5
20) or normalhealthy volunteers (n
5
22) were enrolled to this study after in-formed consent was obtained using guidelines approved by the In-stitutional Review Board of the City of Hope National MedicalCenter. Nineteen CML patients were in chronic phase of disease.One patient had not received any treatment, 19 patients had re-ceived hydroxyurea, and four patients had also received interferon-
a
. Interferon had been discontinued for at least four weeks prior tothe time that marrow samples were obtained.
Selection of CD34
1
progenitors
Heparinized bone marrow samples were obtained from CML pa-tients or normal individuals. Bone marrow mononuclear cells(BMMNC) were isolated by Ficoll-Hypaque (Sigma Diagnostics,St. Louis, MO) density gradient separation (specific gravity 1.077)for 30 minutes at 37
8
C and 400
g.
CD34
1
cell enriched populationswere selected from BMMNC using immunomagnetic column sep-aration (Miltenyi Biotech Inc., Auburn, CA).
Growth factor culture
CD34
1
cells were cultured in 24-well tissue culture plates (BD Fal-con, Franklin Lakes, NJ) in serum-free medium (SFM) (StemPro,GibcoBRL, Gaithersburg, MD) supplemented with one or moregrowth factors (GF). The following GF were used: granulocyte-macrophage colony-stimulating factor (GM-CSF, 10 ng/mL, Immu-nex Biologicals, Seattle, WA), granulocyte colony-stimulating fac-tor (G-CSF, 10 ng/mL, Amgen, Thousand Oaks, CA), interleukin-3(IL-3, 2–50 ng/mL), stem cell factor (SCF, 10–100 ng/mL), Flt-3ligand (FL, 20–200 ng/mL), thrombopoietin (TPO, 2–50 ng/mL) (allfrom Peprotech, Rocky Hill, NJ). Cells were cultured at 37
8
C in ahumidified atmosphere with 5% CO
2
for 1 to 7 days, followingwhich they were harvested and assayed for progenitor content as de-scribed below. The effect of GF stimulation on progenitor number

R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412
1403
was evaluated by comparing numbers of colony forming cells (CFC)and LTC-IC present in the progeny of 1000 initially plated CD34
1
cells after GF culture with the number of CFC and LTC-IC presentin 1000 CD34
1
cells at initiation of culture.
Progenitor assaysA. Colony-forming cells.
CD34
1
cells were plated in semisolidmethylcellulose progenitor culture for 14–18 days and assessed forthe presence of CFU-GM and BFU-E colonies as previously de-scribed [22].
B. Long-term culture-initiating cells (LTC-IC). i. Bulk cultures.
CD34
1
cells were suspended in IMDM (Gibco, Grand Island, NY) supple-mented with 12.5% fetal calf serum (Hyclone Laboratories, Logan,UT), 12.5% horse serum (Stem Cell Technologies, Vancouver, BC,Canada), 2 mM L-glutamine, penicillin (100 u/mL) and streptomy-cin (100 u/mL) (Gibco), and 10
2
6
M hydrocortisone (A-Hydro-cort)(Abbott Laboratories, North Chicago, IL) (long-term bonemarrow culture medium [LTBMC medium]) and plated on M2-10B4 murine fibroblast feeders in 24-well plates as previously de-scribed [23]. Cultures were maintained at 37
8
C in a humidified at-mosphere with 5% CO
2
and fed at weekly intervals by removal ofhalf the medium from the wells and replacement with fresh me-dium. After 6 or 10 weeks all nonadherent and adherent cells wereharvested from stromal cultures, pooled, and plated in methylcellu-lose progenitor culture and the number of CFC evaluated. Thenumber of CFC generated after long-term bone marrow culture(long-term culture-generated CFC [LTC-CFC]) for 6 weeks (week-6 LTC-CFC) and 10 weeks (week-10 LTC-CFC) were enumerated.These were representative of the number of week-6 LTC-IC andweek-10 extended LTC-IC (ELTC-IC) respectively.
ii. Limiting dilution assay.
Limiting dilution analyses wereperformed by plating cells on M2-10B4 feeders, subcultured in 96-well plates. Cells were suspended in LTBMC medium and platedat four different concentrations with 22 replicates for each concen-tration as previously described [24]. Cultures were maintained at37
8
C in a humidified atmosphere with 5% CO
2
with weekly half-medium exchanges with fresh medium for 6 weeks. Subsequently,80% of the medium was carefully removed without agitation andwells were overlaid with CFC growth-supporting medium. Al-though some nonadherent cells may be removed, we have foundthat most cells are retained. However, it is possible that there maybe some loss of nonadherent progenitors, which may lead to under-estimation of LTC-IC frequency. Wells were scored as positive ornegative for the presence of CFC after 14 days. The frequency oflong-term culture-initiating cells was calculated using L-CALCsoftware (Stem Cell Technologies) based on the reciprocal of theconcentration of test cells that gave 37% negative wells.
Single-cell analysis
CD34
1
selected populations were labeled with anti-CD34-FITC andanti-CD38
2
PE antibodies (Becton-Dickinson Biosciences, San Jose,CA). An automated cell deposition unit of a MoFlow (Cytomation,Fort Collins, CO) flow cytometer was used to deposit singleCD34
1
CD38
2
cells into wells of 96-well plates. Cells were eitherdeposited directly onto M210-B4 feeder layers or deposited in wellswithout feeder layers containing SFM with FL (50 ng/mL)
1
SCF(20 ng/mL) and IL-3 (10 ng/mL). Cells deposited in GF containingwells were cultured for 7 days, then divided equally into four sepa-rate wells containing M210-B4 feeder layers. After 6 weeks of stro-
mal culture, wells were overlaid with medium supporting CFCgrowth and evaluated for colony growth after 14 days. The LTC-ICfrequency in single CD34
1
CD38
2
cells, before and after GF culture,was compared. LTC-IC self-replication was determined to have oc-curred during GF culture if two or more of four wells generatedfrom the contents of a single well contained LTC-IC.
Cell cycle analysis
CD34
1
selected cells were cultured in SFM with FL (50 ng/mL)
1
SCF (20 ng/mL) and IL-3 (10 ng/mL) for 1 to 4 days. Cells weresubsequently harvested, washed with phosphate buffered saline(PBS)
1
0.3% BSA, fixed, and permeabilized with ice-cold 70%ethanol. Cells were stored at
2
20
8
C for 1–7 days, washed, and re-suspended in PBS (GibcoBRL) containing propidium iodide (5
m
g/mL)(Sigma, St. Louis, MO) and RNAse (0.2 mg/mL)(Sigma)overnight prior to evaluation by FACS.
PKH-26 labeling
CD34
1
selected cells were labeled with membrane labeling dyePKH-26 (Sigma). CD34
1
cells within a narrow range of PKH fluo-rescence intensity were selected using flow cytometry prior to es-tablishment of culture to allow better resolution of cell generations.The selected cells were cultured in SFM with FL (20 ng/mL)
1
SCF(5 ng/mL) and IL-3 (2 ng/mL) for 7 days. A fraction of the cellswere fixed with 4% paraformaldehyde (Mallinkrodt Inc., Paris,KY) prior to culture to determine the fluorescence intensity of un-divided cells. Cultured cells were analyzed by FACS and PKH-26-bright (undivided), PKH-26-intermediate (1–3 divisions), andPKH-26-dim (
.
3 divisions) populations were selected by FACS.These different PKH-26 fractions were assayed for CFC and LTC-IC frequency and the percentage of progenitors present in the dif-ferent fractions was calculated.
Fluorescent in situ hybridization (FISH) analysis
Cells (5000 to 50,000) were resuspended in 1 mL of 0.4% KCl hy-potonic solution, and incubated at 37
8
C for 20 minutes. Cells werepelleted at 3000 rpm for four minutes using an Eppendorf micro-centrifuge and fixed using 500
m
l of Carnoy’s fixative (1 aceticacid: 3 methanol), on ice for one hour. Just prior to slide prepara-tion, fixed cells were resuspended in 5–10
m
l of Carnoy’s fixative.Hybridization using the LSI dual labeled BCR/ABL DNA probefollowed manufacturer’s instruction (Vysis, Inc., Downers Grove,IL). To identify male from female cells in sex mismatched sam-ples, the CEP Y satellite III centromere probe labeled with Spec-trum Aqua (Vysis, Inc.) was hybridized simultaneously with theLSI BCR/ABL probe. Lymphocytes from a phenotypically normalmale were used as a BCR/ABL negative, male control; SD1 cellline, derived from a female ALL patient, served as BCR/ABL pos-itive, female control. Two hundred nuclei were scored for eachsample. A percentage less than or equal to 6% of cells with a BCR/ABL fusion signal is considered within background limits basedon laboratory validation studies (sensitivity, 95.2%, with 95% con-fidence limits of 76.2–99.9%, and specificity, 100.0%, with 95%confidence limits of 82.4–100.0%).
Statistics
Results of data obtained from multiple experiments were reportedas the mean
6
1 SEM. Significance levels were determined byStudent’s
t
-test analysis.

1404
R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412
Results
Growth factor stimulation is associated with depletion of CML primitive progenitors
The effect of culture of CML and normal CD34
1
cells in thepresence of different growth factors (GF) in serum-free me-dium on the numbers of committed progenitors (colony-forming cells or CFC) was evaluated. The GFs studied in-cluded G-CSF, GM-CSF, IL-3, SCF, FL, and TPO andcombinations of FL
1
SCF
1
IL-3 and FL
1
SCF
1
TPO.The CFC frequency in CML and normal CD34
1
cells priorto initiation of GF culture were not significantly different.CML and normal CFC numbers following culture in the ab-sence of any GF were not significantly different (data notshown). Increasing GF concentrations resulted in a dose-dependent increase in expansion of normal CFC numbers.There was no significant difference in the expansion ofCML vs normal committed progenitor numbers following 7days culture with IL-3, SCF, FL, and TPO (Fig. 1) orG-CSF and GM-CSF (data not shown). However, with thecombinations of FL
1
SCF
1
IL-3, CML CFC expansionwas blunted at higher concentrations and was significantlyless than normal CFC expansion at the highest GF concen-tration studied (
p
5
0.05). Similar trends were seen with theFL
1
SCF
1
TPO combination.We evaluated the effects of the same growth factors (GF)
on CML and normal primitive myeloid progenitors. Theweek-6 LTC-IC frequency in CML and normal CD34
1
cellsprior to initiation of GF culture were not significantly dif-ferent. Whereas culture for 7 days with IL-3, SCF, G-CSF,GM-CSF, and TPO as individual GF did not result in expan-sion of normal LTC-IC (measured as week-6 LTC-CFC),culture with FL alone or the combinations of FL
1
SCF
1
IL-3 and FL
1
SCF
1
TPO resulted in expansion of normalweek-6 LTC-IC numbers (Fig. 2 and data not shown). Incontrast, CML LTC-CFC numbers were not increased afterculture with GFs (FL, FL
1
SCF
1
IL-3 and FL
1
SCF
1
TPO) that expanded normal LTC-CFC numbers (Fig. 2). Adose-dependent increase in expansion of normal LTC-CFCnumbers was seen with increasing GF concentrations butwas not seen for CML LTC-CFC. Enumeration of LTC-ICfrequency by limiting dilution analysis confirmed that cul-ture with FL
1
IL3
1
SCF resulted in increased numbers ofnormal week-6 LTC-IC but reduced numbers of CMLweek-6 LTC-IC (Fig. 3).
CML CD34
1
cells contain both BCR/ABL positive andnegative LTC-IC [25,26]. The effects of culture of CMLCD34
1
cells in GF for 7 days on the relative growth ofBCR/ABL negative and positive LTC-IC were compared.There was significant variation in the number of BCR/ABLnegative LTC-IC amongst CML patients. As shown in Ta-ble 1, GF culture was associated with increased outgrowthof BCR/ABL negative LTC-IC in some but not all CML pa-tients. Selective expansion of BCR/ABL negative LTC-ICtended to be seen in samples in which a higher proportion of
BCR/ABL negative LTC-IC were present prior to GF cul-ture. These experiments suggest that GF stimulation canlead to preferential outgrowth of BCR/ABL negative LTC-IC in some CML patients.
Week-10 extended LTC-IC (ELTC-IC) are progenitorscapable of generating CFC after 10 weeks of long-term cul-ture) and are an even more primitive progenitor population.In contrast to normal week-6 LTC-IC, normal week-10LTC-IC were reduced in number after culture with FL
1
IL-3
1
SCF for 7 days (Fig. 4). However, these culture con-ditions resulted in significantly greater depletion of CMLweek-10 LTC-CFC compared with normal week-10 LTC-IC (
p
5
0.02).
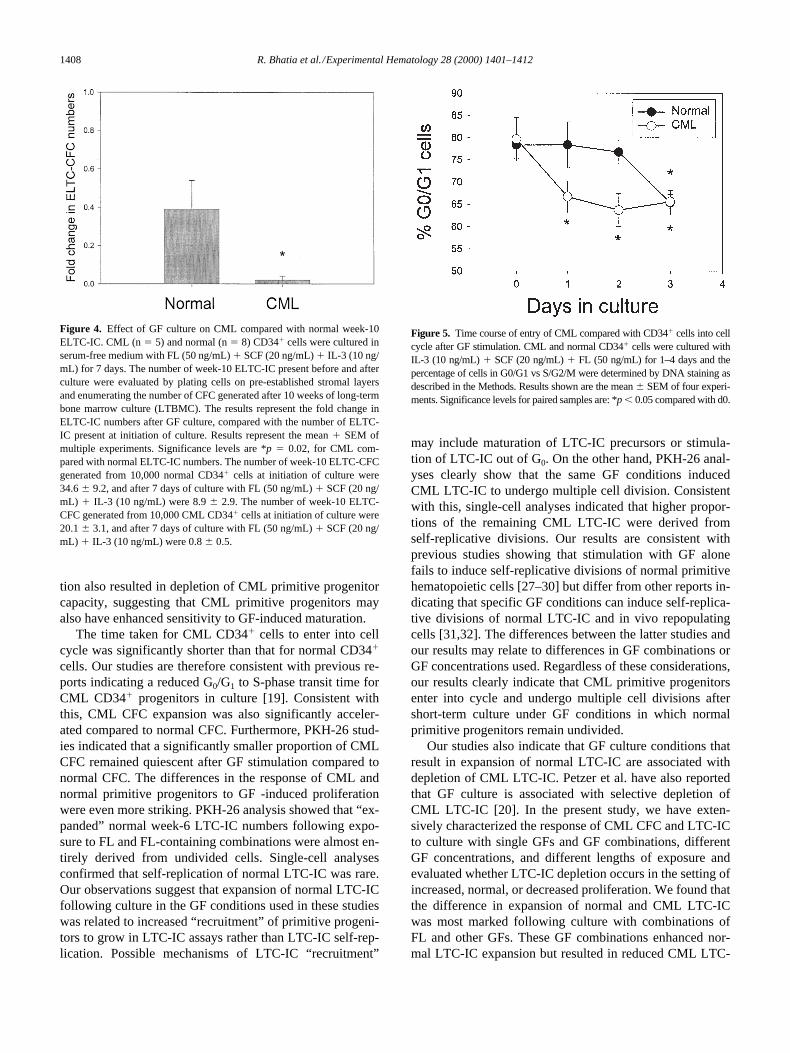
GF stimulation is associated with increasedproliferation of CML committed and primitive progenitors
The kinetics of entry of CML and normal CD34
1
cells intocell cycle following GF stimulation was evaluated. The per-centage of CML and normal CD34
1
cells in G0/G1 vs S/G2/M was not significantly different on isolation, prior tothe initiation of GF culture. However, CML CD34
1
cellsentered into cell cycle much more rapidly than normalCD34
1
cells. Maximal entry into cell cycle of normalCD34
1
cells required
$
3 days of GF stimulation whereasmaximal cycling was observed 1 day after GF stimulationfor CML CD34
1
cells (Fig. 5). Consistent with the earlierentry of CML CD34
1
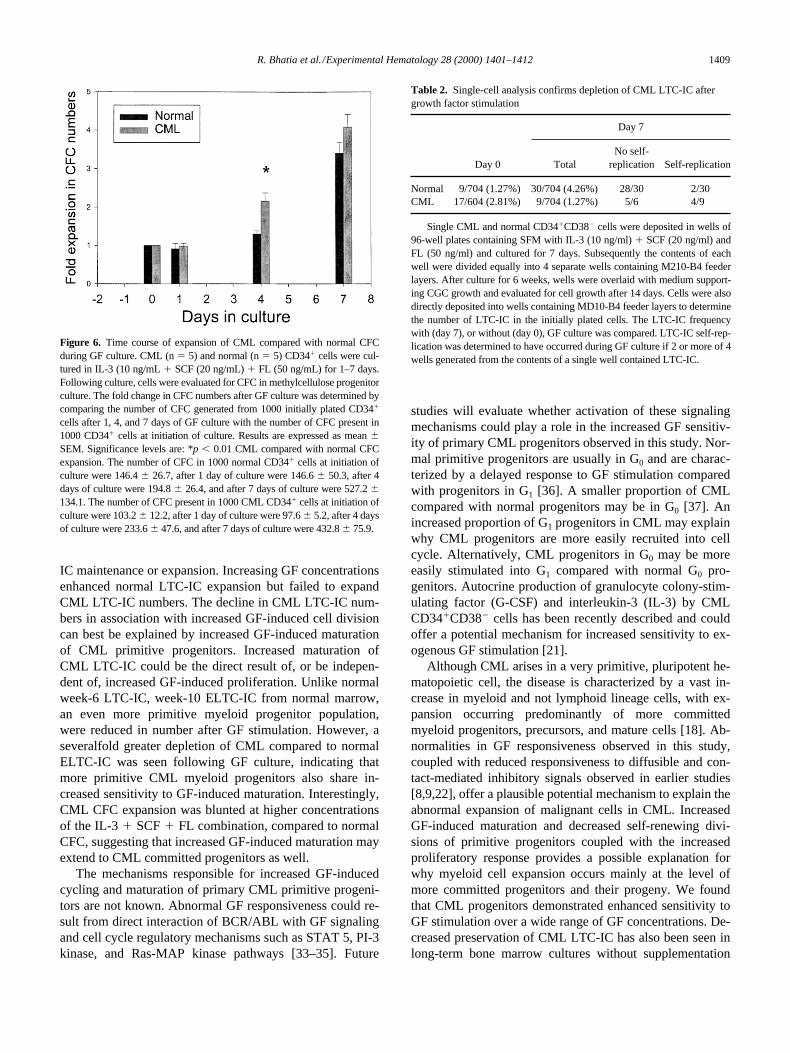
cells into cell cycle, CML CFC ex-pansion occurred more rapidly than normal CFC expansion,with significantly increased CML CFC expansion seen after4 days of GF culture (
p
,
0.01). However, this differencewas no longer significant after 7 days in culture (Fig. 6).These observations suggest that CML committed progeni-tors are more sensitive to GF-induced proliferative signal-ing than their normal counterparts. Evaluation of LTC-ICexpansion over time revealed significantly reduced expan-sion of CML LTC-IC compared with normal LTC-IC afterboth 4 and 7 days of GF exposure (data not shown).
We evaluated the effect of GF stimulation on LTC-IC fre-quency and LTC-IC self-replication at the single-cell level.Single CD34
1
CD38
2
cells were sorted into wells of 96-wellplates using flow cytometry and cultured for 7 days with FL(20 ng/mL)
1
SCF (10 ng/mL)
1
IL3 (2 ng/mL). The con-tents of each well were then divided between four wells con-taining preestablished stromal feeder layers and long-termculture established. The LTC-IC frequency was determinedby enumerating the number of wells positive for LTC-ICgrowth and expressing this as a percentage of the total num-ber of wells that were initially seeded. The LTC-IC fre-quency in the initiating CD34
1
CD382 population withoutGF culture was determined by depositing single cells di-rectly on stromal feeder layers. The LTC-IC frequency after7 days of GF culture was compared with LTC-IC frequencywithout GF culture. GF culture resulted in an increased fre-quency of week-6 LTC-IC in normal CD341CD382 cellscompared with cells sorted directly onto stroma. Presence of

R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412 1405
Figure 1. Effect of culture with GF on CML compared with normal CFC numbers. CML (n 5 4–9) and normal (n 5 5–10) CD341 cells were cultured inserum-free medium in the presence of increasing GF concentrations as shown for 7 days. Following culture, cells were evaluated for CFC in methylcelluloseprogenitor culture. The fold change in CFC numbers after GF culture was determined by comparing the number of CFC generated from 1000 initially platedCD341 cells after GF culture with the number of CFC present in 1000 CD341 cells at initiation of culture. Results are expressed as the mean 6 SEM. Signifi-cance levels are *p # 0.05, for CML compared with normal CFC expansion. The number of CFC present in 1000 normal CD341 cells at initiation of culturewere 117.7 6 16.3, and after 7 days of culture were 117 6 30.2 for IL-3 (10 ng/mL), 118.8 6 20.8 for SCF (20 ng/mL), 197.6 6 36.5 for FL (50 ng/mL), 57 63.9 for TPO (10 ng/mL), 447.8 6 69.7 for the combination of FL (50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) and 289 6 18 for FL (50 ng/mL) 1 SCF(20 ng/mL) 1 TPO (10 ng/mL). The number of CFC present in 1000 CML CD341 cells at initiation of culture were 99.2 6 7.8, and after 7 days of culture were154.4 6 40.8 for IL-3 (10 ng/mL), 166.7 6 36.9 for SCF (20 ng/mL), 175.3 6 27.6 for FL (50 ng/mL), 69.3 6 1.3 for TPO (10 ng/mL), 362.7 6 51.7 for thecombination of FL (50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) and 222.7 6 34.7 for FL (50 ng/mL) 1 SCF (20 ng/mL) 1 TPO (10 ng/mL).
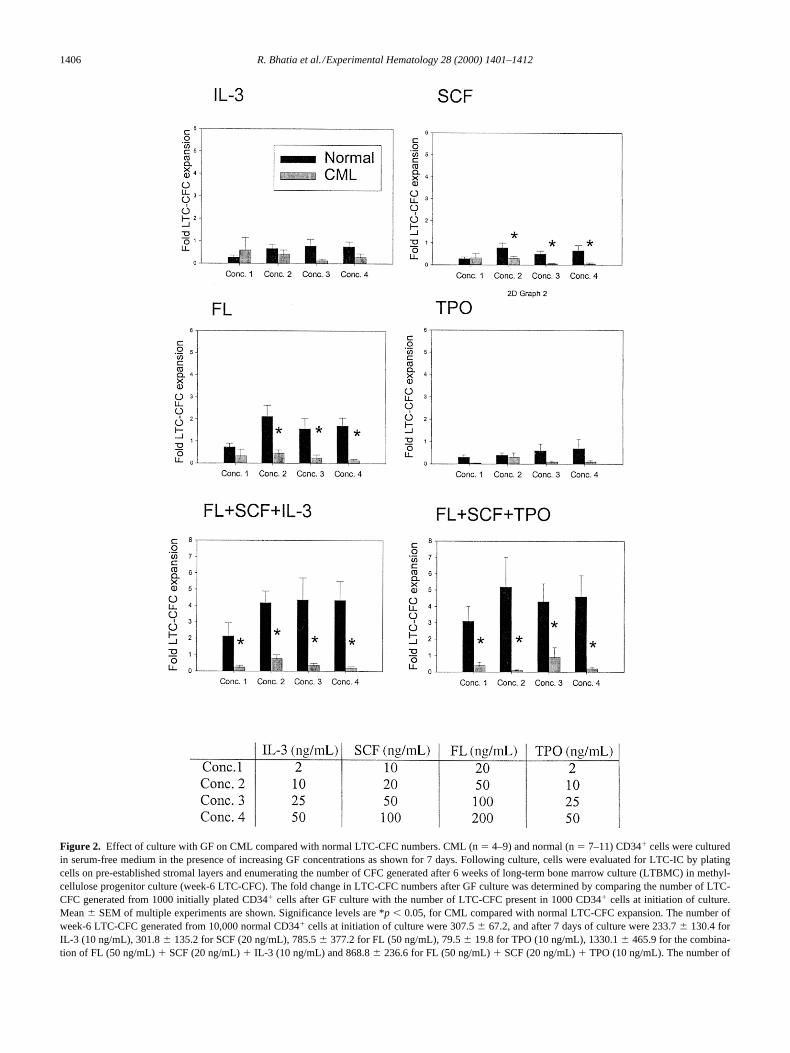
1406 R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412
Figure 2. Effect of culture with GF on CML compared with normal LTC-CFC numbers. CML (n 5 4–9) and normal (n 5 7–11) CD341 cells were culturedin serum-free medium in the presence of increasing GF concentrations as shown for 7 days. Following culture, cells were evaluated for LTC-IC by platingcells on pre-established stromal layers and enumerating the number of CFC generated after 6 weeks of long-term bone marrow culture (LTBMC) in methyl-cellulose progenitor culture (week-6 LTC-CFC). The fold change in LTC-CFC numbers after GF culture was determined by comparing the number of LTC-CFC generated from 1000 initially plated CD341 cells after GF culture with the number of LTC-CFC present in 1000 CD341 cells at initiation of culture.Mean 6 SEM of multiple experiments are shown. Significance levels are *p , 0.05, for CML compared with normal LTC-CFC expansion. The number ofweek-6 LTC-CFC generated from 10,000 normal CD341 cells at initiation of culture were 307.5 6 67.2, and after 7 days of culture were 233.7 6 130.4 forIL-3 (10 ng/mL), 301.8 6 135.2 for SCF (20 ng/mL), 785.5 6 377.2 for FL (50 ng/mL), 79.5 6 19.8 for TPO (10 ng/mL), 1330.1 6 465.9 for the combina-tion of FL (50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) and 868.8 6 236.6 for FL (50 ng/mL) 1 SCF (20 ng/mL) 1 TPO (10 ng/mL). The number of

R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412 1407
LTC-IC in two or more of the four wells derived from divid-ing the contents of a single well indicated that LTC-IC self-replication had occurred. Although there was expansion ofnormal LTC-IC after 7 days of GF culture, it was rare for asingle cell to give rise to more than one LTC-IC, indicatingthat increased normal LTC-IC numbers did not result fromLTC-IC self-replication, but rather represented recruitmentof primitive hematopoietic cells to grow in LTC-IC assays.On the other hand, GF culture was associated with reducedLTC-IC frequency in CML CD341CD382 cells (Table 2).Interestingly, although CML LTC-IC were depleted follow-ing GF culture, the remaining LTC-IC were more likely tobe derived from self-replicative divisions.
CD341 cells were also labeled with PKH-26 dye prior toGF culture to track cell divisions during GF culture. Eachcell division results in reduction in PKH-26 intensity by half.
After 7 days of culture in FL 1 SCF 1 IL-3 the proportionof PKH-26-bright (undivided) and PKH-26-intermediate and-dim (1–3 divisions and .3 divisions) cells were similar forcultures initiated with CML and normal CD341 cells (Fig.7A and B). Analysis of populations after 7 days of GF cul-ture sorted by PKH-26 intensity indicated that the majorityof normal and CML CFC present were derived from dividedcells (Fig. 7B). However, significantly fewer CML com-pared with normal CFC remained undivided (1.0 6 0.7% vs9.5 6 2.9%, p , 0.05). On the other hand, the majority ofnormal LTC-IC were derived from undivided cells and only7.3 6 5.6% were derived from the divided cells (Fig. 7C).This observation is consistent with the results of single-cellanalysis (Table 2) and suggests that GF-induced expansionin normal LTC-IC numbers is related to increased LTC-IC“recruitment” rather than LTC-IC self-replication. In con-trast, the majority of CML LTC-IC were derived from di-vided CD341 cells with a large fraction from cells that hadundergone .3 divisions. These studies indicate that CMLprimitive progenitors, although depleted in GF culture, un-dergo significantly more cell divisions after GF stimulationthan normal CD341 cells. These results also suggest thatCML primitive progenitors are more sensitive to GF-inducedmaturation, since there is reduction in LTC-IC numbers inspite of increased cell division.
DiscussionIn this study we have shown that primitive and committedhematopoietic progenitors from CML bone marrow have anincreased proliferative response to GF stimulation com-pared with their normal counterparts. However, GF stimula-
week-6 LTC-CFC generated from 10,000 CML CD341 cells at initiation of culture were 202.1 6 43.4, and after 7 days of culture were 42.6 6 14.6 for IL-3(10 ng/mL), 26.1 6 8.5 for SCF (20 ng/mL), 70.7 6 24.0 for FL (50 ng/mL), 47 6 8.6 for TPO (10 ng/mL), 132.3 6 53 for the combination of FL (50 ng/mL)1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) and 42.6 6 18.9 for FL (50 ng/mL) 1 SCF (20 ng/mL) 1 TPO (10 ng/mL).
Figure 3. Results of limiting dilution analyses to evaluate effect of GFculture on CML and normal LTC-IC numbers. CML (n 5 3–4) and normal(n 5 4–6) CD341 cells were cultured in serum-free medium in the pres-ence of increasing GF concentrations for 7 days. Following culture, cellswere evaluated for week-6 LTC-IC numbers using limiting dilution analy-ses, as described in the Methods section. The fold change in LTC-IC num-bers after GF culture was determined by comparing the number of LTC-ICpresent in 1000 cells after GF culture with the number of LTC-IC presentin 1000 CD341 cells at initiation of culture. Results shown represent themean 6 SEM of multiple experiments. Differences between CML and nor-mal LTC-IC expansion were significant for all concentrations (p , 0.05).The number of week-6 LTC-IC present in 10,000 normal CD341 cells atinitiation of culture were 32.5 6 7.5, and after 7 days of culture with FL(50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) were 80.3 6 26.2. Thenumber of week-6 LTC-IC present in 10,000 CML CD341 cells at initia-tion of culture were 16.4 6 5.1, and after 7 days of culture with FL (50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) were 3.1 6 2.0.
Table 1. Effect of culture with GF for 7 days on the proportion of BCR/ABL negative LTC-IC
Subjectnumber
Disease stage/timefrom diagnosis
% BCR/ABL2ve LTC-IC
Before GF culture
After 7 daysof GF culture
11 CP/1m 17 1922 CP/6m 44.5 8332 CP/9m 11.5 841 CP/7m 77.5 78.552 CP/1.5y 10 1562 CP/2y 62.5 9371 AP/3.5y 8 8
CML CD341 cells were plated in LTC-IC assays after culture with IL-31 SCF 1 FL for 7 days, as described in the methods. The proportion ofBCR/ABL1 cells LTC-IC before and after FG culture was compared.1FISH performed on all cells generated from long-term culture. 2FISH per-formed on CGC generated from long-term culture. CP, chronic phase; AP,accelerated phase
1408 R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412
tion also resulted in depletion of CML primitive progenitorcapacity, suggesting that CML primitive progenitors mayalso have enhanced sensitivity to GF-induced maturation.
The time taken for CML CD341 cells to enter into cellcycle was significantly shorter than that for normal CD341
cells. Our studies are therefore consistent with previous re-ports indicating a reduced G0/G1 to S-phase transit time forCML CD341 progenitors in culture [19]. Consistent withthis, CML CFC expansion was also significantly acceler-ated compared to normal CFC. Furthermore, PKH-26 stud-ies indicated that a significantly smaller proportion of CMLCFC remained quiescent after GF stimulation compared tonormal CFC. The differences in the response of CML andnormal primitive progenitors to GF -induced proliferationwere even more striking. PKH-26 analysis showed that “ex-panded” normal week-6 LTC-IC numbers following expo-sure to FL and FL-containing combinations were almost en-tirely derived from undivided cells. Single-cell analysesconfirmed that self-replication of normal LTC-IC was rare.Our observations suggest that expansion of normal LTC-ICfollowing culture in the GF conditions used in these studieswas related to increased “recruitment” of primitive progeni-tors to grow in LTC-IC assays rather than LTC-IC self-rep-lication. Possible mechanisms of LTC-IC “recruitment”
may include maturation of LTC-IC precursors or stimula-tion of LTC-IC out of G0. On the other hand, PKH-26 anal-yses clearly show that the same GF conditions inducedCML LTC-IC to undergo multiple cell division. Consistentwith this, single-cell analyses indicated that higher propor-tions of the remaining CML LTC-IC were derived fromself-replicative divisions. Our results are consistent withprevious studies showing that stimulation with GF alonefails to induce self-replicative divisions of normal primitivehematopoietic cells [27–30] but differ from other reports in-dicating that specific GF conditions can induce self-replica-tive divisions of normal LTC-IC and in vivo repopulatingcells [31,32]. The differences between the latter studies andour results may relate to differences in GF combinations orGF concentrations used. Regardless of these considerations,our results clearly indicate that CML primitive progenitorsenter into cycle and undergo multiple cell divisions aftershort-term culture under GF conditions in which normalprimitive progenitors remain undivided.
Our studies also indicate that GF culture conditions thatresult in expansion of normal LTC-IC are associated withdepletion of CML LTC-IC. Petzer et al. have also reportedthat GF culture is associated with selective depletion ofCML LTC-IC [20]. In the present study, we have exten-sively characterized the response of CML CFC and LTC-ICto culture with single GFs and GF combinations, differentGF concentrations, and different lengths of exposure andevaluated whether LTC-IC depletion occurs in the setting ofincreased, normal, or decreased proliferation. We found thatthe difference in expansion of normal and CML LTC-ICwas most marked following culture with combinations ofFL and other GFs. These GF combinations enhanced nor-mal LTC-IC expansion but resulted in reduced CML LTC-
Figure 4. Effect of GF culture on CML compared with normal week-10ELTC-IC. CML (n 5 5) and normal (n 5 8) CD341 cells were cultured inserum-free medium with FL (50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) for 7 days. The number of week-10 ELTC-IC present before and afterculture were evaluated by plating cells on pre-established stromal layersand enumerating the number of CFC generated after 10 weeks of long-termbone marrow culture (LTBMC). The results represent the fold change inELTC-IC numbers after GF culture, compared with the number of ELTC-IC present at initiation of culture. Results represent the mean 1 SEM ofmultiple experiments. Significance levels are *p 5 0.02, for CML com-pared with normal ELTC-IC numbers. The number of week-10 ELTC-CFCgenerated from 10,000 normal CD341 cells at initiation of culture were34.6 6 9.2, and after 7 days of culture with FL (50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) were 8.9 6 2.9. The number of week-10 ELTC-CFC generated from 10,000 CML CD341 cells at initiation of culture were20.1 6 3.1, and after 7 days of culture with FL (50 ng/mL) 1 SCF (20 ng/mL) 1 IL-3 (10 ng/mL) were 0.8 6 0.5.
Figure 5. Time course of entry of CML compared with CD341 cells into cellcycle after GF stimulation. CML and normal CD341 cells were cultured withIL-3 (10 ng/mL) 1 SCF (20 ng/mL) 1 FL (50 ng/mL) for 1–4 days and thepercentage of cells in G0/G1 vs S/G2/M were determined by DNA staining asdescribed in the Methods. Results shown are the mean 6 SEM of four experi-ments. Significance levels for paired samples are: *p , 0.05 compared with d0.
R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412 1409
IC maintenance or expansion. Increasing GF concentrationsenhanced normal LTC-IC expansion but failed to expandCML LTC-IC numbers. The decline in CML LTC-IC num-bers in association with increased GF-induced cell divisioncan best be explained by increased GF-induced maturationof CML primitive progenitors. Increased maturation ofCML LTC-IC could be the direct result of, or be indepen-dent of, increased GF-induced proliferation. Unlike normalweek-6 LTC-IC, week-10 ELTC-IC from normal marrow,an even more primitive myeloid progenitor population,were reduced in number after GF stimulation. However, aseveralfold greater depletion of CML compared to normalELTC-IC was seen following GF culture, indicating thatmore primitive CML myeloid progenitors also share in-creased sensitivity to GF-induced maturation. Interestingly,CML CFC expansion was blunted at higher concentrationsof the IL-3 1 SCF 1 FL combination, compared to normalCFC, suggesting that increased GF-induced maturation mayextend to CML committed progenitors as well.
The mechanisms responsible for increased GF-inducedcycling and maturation of primary CML primitive progeni-tors are not known. Abnormal GF responsiveness could re-sult from direct interaction of BCR/ABL with GF signalingand cell cycle regulatory mechanisms such as STAT 5, PI-3kinase, and Ras-MAP kinase pathways [33–35]. Future
studies will evaluate whether activation of these signalingmechanisms could play a role in the increased GF sensitiv-ity of primary CML progenitors observed in this study. Nor-mal primitive progenitors are usually in G0 and are charac-terized by a delayed response to GF stimulation comparedwith progenitors in G1 [36]. A smaller proportion of CMLcompared with normal progenitors may be in G0 [37]. Anincreased proportion of G1 progenitors in CML may explainwhy CML progenitors are more easily recruited into cellcycle. Alternatively, CML progenitors in G0 may be moreeasily stimulated into G1 compared with normal G0 pro-genitors. Autocrine production of granulocyte colony-stim-ulating factor (G-CSF) and interleukin-3 (IL-3) by CMLCD341CD382 cells has been recently described and couldoffer a potential mechanism for increased sensitivity to ex-ogenous GF stimulation [21].
Although CML arises in a very primitive, pluripotent he-matopoietic cell, the disease is characterized by a vast in-crease in myeloid and not lymphoid lineage cells, with ex-pansion occurring predominantly of more committedmyeloid progenitors, precursors, and mature cells [18]. Ab-normalities in GF responsiveness observed in this study,coupled with reduced responsiveness to diffusible and con-tact-mediated inhibitory signals observed in earlier studies[8,9,22], offer a plausible potential mechanism to explain theabnormal expansion of malignant cells in CML. IncreasedGF-induced maturation and decreased self-renewing divi-sions of primitive progenitors coupled with the increasedproliferatory response provides a possible explanation forwhy myeloid cell expansion occurs mainly at the level ofmore committed progenitors and their progeny. We foundthat CML progenitors demonstrated enhanced sensitivity toGF stimulation over a wide range of GF concentrations. De-creased preservation of CML LTC-IC has also been seen inlong-term bone marrow cultures without supplementation
Figure 6. Time course of expansion of CML compared with normal CFCduring GF culture. CML (n 5 5) and normal (n 5 5) CD341 cells were cul-tured in IL-3 (10 ng/mL 1 SCF (20 ng/mL) 1 FL (50 ng/mL) for 1–7 days.Following culture, cells were evaluated for CFC in methylcellulose progenitorculture. The fold change in CFC numbers after GF culture was determined bycomparing the number of CFC generated from 1000 initially plated CD341
cells after 1, 4, and 7 days of GF culture with the number of CFC present in1000 CD341 cells at initiation of culture. Results are expressed as mean 6SEM. Significance levels are: *p , 0.01 CML compared with normal CFCexpansion. The number of CFC in 1000 normal CD341 cells at initiation ofculture were 146.4 6 26.7, after 1 day of culture were 146.6 6 50.3, after 4days of culture were 194.8 6 26.4, and after 7 days of culture were 527.2 6134.1. The number of CFC present in 1000 CML CD341 cells at initiation ofculture were 103.2 6 12.2, after 1 day of culture were 97.6 6 5.2, after 4 daysof culture were 233.6 6 47.6, and after 7 days of culture were 432.8 6 75.9.
Table 2. Single-cell analysis confirms depletion of CML LTC-IC after growth factor stimulation
Day 7
Day 0 TotalNo self-
replication Self-replication
Normal 9/704 (1.27%) 30/704 (4.26%) 28/30 2/30CML 17/604 (2.81%) 9/704 (1.27%) 5/6 4/9
Single CML and normal CD341CD382 cells were deposited in wells of96-well plates containing SFM with IL-3 (10 ng/ml) 1 SCF (20 ng/ml) andFL (50 ng/ml) and cultured for 7 days. Subsequently the contents of eachwell were divided equally into 4 separate wells containing M210-B4 feederlayers. After culture for 6 weeks, wells were overlaid with medium support-ing CGC growth and evaluated for cell growth after 14 days. Cells were alsodirectly deposited into wells containing MD10-B4 feeder layers to determinethe number of LTC-IC in the initially plated cells. The LTC-IC frequencywith (day 7), or without (day 0), GF culture was compared. LTC-IC self-rep-lication was determined to have occurred during GF culture if 2 or more of 4wells generated from the contents of a single well contained LTC-IC.

1410 R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412
with additional GF supplementation and may occur throughsimilar pathophysiological mechanisms [38].
It may be possible to exploit differences in the responseof CML and normal primitive progenitors to growth factorsignals to develop novel therapeutic strategies for selectionof BCR/ABL negative primitive progenitors. Variable pro-portions of BCR/ABL negative primitive progenitors persistin the marrow of CML patients [25,26,39]. Based on our re-sults, GF stimulation may preferentially deplete BCR/ABLpositive primitive myeloid progenitors while retaining nor-mal BCR/ABL negative progenitors. This would allow forapplications such as ex vivo purging of grafts for autolo-gous transplantation or in vivo administration of GF as a
therapeutic strategy. We show that GF culture resulted in anincrease in the proportion of BCR/ABL negative LTC-IC insome CML subjects who had increased proportions of BCR/ABL negative LTC-IC at initiation of culture. The low num-bers of BCR/ABL negative primitive progenitors in somepatients could limit the utility of this approach. However,optimal expansion of residual BCR/ABL-veQ6 LTC-IC islikely to require additional diffusible and contact mediatedstromal factors and optimization of conditions for selectiveoutgrowth of BCR/ABL negative stem cells will be evalu-ated in future studies [27–29,40].
In conclusion, our results show that CML primitive andcommitted progenitors demonstrate increased proliferation
Figure 7. PKH-26 analysis of CML compared with normal CD341 cells, CFC, and week-6 LTC-IC cell division during GF culture. CML (n 5 3) and normal(n 5 5) CD341 cells were labeled with PKH-26 and cultured in SFM with FL (20 ng/mL) 1 SCF (5 ng/mL) and IL-3 (2 ng/mL) for 7 days as described in theMethods. The cultured cells were analyzed by FACS, and PKH-26-bright (undivided), PKH-26-intermediate (1–3 divisions), and PKH-26-dim (.3 divisions)populations were selected by FACS sorting. The different PKH-26 fractions were assayed for CFC and LTC-IC frequency and the percentage of these progen-itors present in the different fractions was calculated. Results represent the mean 6 SEM of aggregate experiments. *p , 0.001 CML compared with normalLTC-IC, s p , 0.05 CML compared with normal CFC.

R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412 1411
and maturation in response to GF stimulation. These abnor-malities may contribute to the abnormal expansion of differ-entiated malignant myeloid cells in CML. Future studieswill evaluate the mechanisms underlying these abnormali-ties and explore their application towards the developmentof novel strategies to target malignant cells and select be-nign stem cells in this disease.
AcknowledgmentsWe are grateful for the excellent technical support of Lucy Brownand Jim Bolen in the Analytical Cytometry Core, who performedthe FACS sorting and analysis. We would also like to express ourgratitude to the following nurse coordinators: Allison Ahlers, Jean-ine Stevenson, Pilar Fonbuena, and Rodrigo Nunez, and all thephysicians in Department of Hematology/BMT for their invalu-able help with these studies.
This work was supported in part by the Public Health Servicesgrants CA74455 from the National Cancer Institute, AmericanCancer Society grant RPG-99-202-01-LBC, and The Leukemiaand Lymphoma Society Translational Research Grant 6468 toRavi Bhatia and Public Health Services grants CA30206 andCA33572 from the National Cancer Institute to Marilyn L. Slovak.
References1. Rowley JD (1973) A new consistent chromosome abnormality in
chronic myelogenous leukemia identified by quinacrine fluorescenceand Giemsa staining. Nature 243:209
2. DeKlein A, Van Kessel AG, Grosveld G, et al. (1982) A cellular onco-gene is translocated to the Philadelphia chromosome in chronic myelo-cytic leukemia. Nature 300:765
3. Daley GQ, Van Etten RA, Baltimore D (1990) Induction of chronicmyelogenous leukemia in mice by the p210 bcr/abl gene of the Phila-delphia chromosome. Science 247:824
4. Daley GQ, Baltimore D (1988) Transformation of an interleukin-3-dependent hematopoietic cell line by the chronic leukemia-specificP210 bcr/abl protein. Proc Natl Acad Sci U S A 85:9312
5. al-Amin A, Lennartz K, Runde V, Seeber S, Opalka B, Kloke O (1998)Frequency of clonal B lymphocytes in chronic myelogenous leukemiaevaluated by fluorescence in situ hybridization. Cancer Genet Cytoge-net 104(1):45
6. Haferlach T, Winkemann M, Nickenig C, et al. (1997) Which com-partments are involved in Philadelphia-chromosome positive chronicmyeloid leukaemia? An answer at the single cell level by combiningMay-Grunwald-Giemsa staining and fluorescence in situ hybridizationtechniques (see comments). Br J Haematol 97(1):99
7. Takahashi N, Miura I, Saitoh K, Miura AB (1998) Lineage involve-ment of stem cells bearing the philadelphia chromosome in chronicmyeloid leukemia in the chronic phase as shown by a combination offluorescence-activated cell sorting and fluorescence in situ hybridiza-tion. Blood 92(12):4758
8. Eaves CJ, Cashman JD, Wolpe SD, et al. (1993) Unresponsiveness ofprimitive chronic myeloid leukemia cells to macrophage inflammatoryprotein 1a, an inhibitor of primitive normal hematopoietic cells. ProcNatl Acad Sci U S A 90(24):12015
9. Cashman JD, Eaves CJ, Sarris AH, Eaves AC (1998) MCP-1, notMIP-1a, is the endogenous chemokine that cooperates with TGF-b toinhibit the cycling of primitive normal but not leukemic (CML) pro-genitors in long-term human marrow cultures. Blood 92(7):2338
10. Verfaillie CM, McCarthy JB, McGlave PB (1992) Mechanisms under-lying abnormal trafficking of malignant progenitors in chronic myel-
ogenous leukemia: decreased adhesion to stroma and fibronectin butincreased adhesion to the basement membrane components lamininand collagen type IV. J Clin Invest 90:1232
11. Bhatia R, McCarthy JB, Verfaillie CM (1996) Interferon-a restoresnormal b 1 integrin-mediated inhibition of hematopoietic progenitorproliferation by the marrow microenvironment in chronic myeloge-nous leukemia. Blood 87(9):3883
12. Hurley RW, McCarthy JB, Verfaillie CM (1995) Direct adhesion tobone marrow stroma via fibronectin receptors inhibits hematopoieticprogenitor proliferation. J Clin Invest 96(1):511
13. Bedi A, Griffin CA, Barber JP, et al. (1994) Growth factor–mediatedterminal differentiation of chronic myeloid leukemia. Cancer Res54:5535
14. Bedi A, Zehnbauer BA, Barber JP, Sharkis SJ, Jones RJ (1994) Inhibi-tion of apoptosis by BCR-ABL in chronic myeloid leukemia. Blood83:2038
15. Ghaffari S, Daley GQ, Lodish HF (1999) Growth factor independenceand BCR/ABL transformation: promise and pitfalls of murine modelsystems and assays. Leukemia 13(8):1200
16. Maguer-Satta V, Burl S, Liu L, et al. (1998) BCR-ABL acceleratesC2-ceramide-induced apoptosis. Oncogene 16(2):237
17. Amos TA, Lewis JL, Grand FH, Gooding RP, Goldman JM, GordonMY (1995) Apoptosis in chronic myeloid leukaemia: normal re-sponses by progenitor cells to growth factor deprivation, X-irradiationand glucocorticoids. Br J Haematol 91(2):387
18. Clarkson BD, Strife A, Wisniewski D, Lambek C, Carpino N (1997)New understanding of the pathogenesis of CML: a prototype of earlyneoplasia. Leukemia 11:1404
19. Traycoff CM, Halstead B, Rice S, McMahel J, Srour EF, Cornetta K(1998) Chronic myelogenous leukaemia CD341 cells exit G0/G1phases of cell cycle more rapidly than normal marrow CD341 cells. BrJ Haematol 102(3):759
20. Petzer AL, Eaves CJ, Barnett MJ, Eaves AC (1997) Selective expan-sion of primitive normal hematopoietic cells in cytokine-supplementedcultures of purified cells from patients with chronic myeloid leukemia.Blood 90(1):64
21. Jiang X, Lopez A, Holyoake T, Eaves A, Eaves C (1999) Autocrine pro-duction and action of IL-3 and granulocyte colony-stimulating factor inchronic myeloid leukemia. Proc Natl Acad Sci U S A 96(22):12804
22. Bhatia R, Wayner EA, McGlave PB, Verfaillie CM (1994) Interferon-arestores normal adhesion of chronic myelogenous leukemia hemato-poietic progenitors to bone marrow stroma by correcting impaired b 1integrin receptor function (see comments). J Clin Invest 94(1):384
23. Bhatia R, McGlave PB, Dewald GW, Blazar BR, Verfaillie CM (1995)Abnormal function of the bone marrow microenvironment in chronicmyelogenous leukemia: role of malignant stromal macrophages. Blood85(12):3636
24. Bhatia R, McGlave PB, Miller JS, Wissink S, Lin WN, Verfaillie CM(1997) A clinically suitable ex vivo expansion culture system for LTC-IC and CFC using stroma-conditioned medium. Exp Hematol 25(9):980
25. Maguer-Satta V, Petzer AL, Eaves AC, Eaves CJ (1996) BCR-ABLexpression in different subpopulations of functionally characterizedPh1 CD341 cells from patients with chronic myeloid leukemia. Blood88(5):1796
26. Verfaillie CM, Bhatia R, Miller W, et al. (1996) BCR/ABL negativeprimitive progenitors suitable for transplantation can be selected fromthe marrow of most early-chronic phase but not accelerated-phasechronic myelogenous leukemia patients. Blood 87(11):4770
27. Bennaceur-Griscelli A, Tourino C, Izac B, Vainchenker W, CoulombelL (1999) Murine stromal cells counteract the loss of long-term culture-initiating cell potential induced by cytokines in CD34(1)CD38(low/neg) human bone marrow cells. Blood 94(2):529
28. Breems DA, Blokland EAW, Siebel KE, Mayen AEM, Engels LJA,Ploemacher R (1998) Stroma-contact prevents loss of hematopoieticstem cell quality during ex vivo expansion of CD341 mobilized pe-ripheral blood stem cells. Blood 91(1):111

1412 R. Bhatia et al./Experimental Hematology 28 (2000) 1401–1412
29. Verfaillie CM (1993) Soluble factor(s) produced by human bone mar-row stroma increase cytokine-induced proliferation and maturation ofprimitive hematopoietic progenitors while preventing their terminaldifferentiation. Blood 82(7):2045
30. Peters SO, Kittler EL, Ramshaw HS, Quesenberry PJ (1996) Ex vivoexpansion of murine marrow cells with interleukin-3 (IL-3), IL- 6, IL-11, and stem cell factor leads to impaired engraftment in irradiatedhosts. Blood 87(1):30
31. Petzer AL, Hogge DE, Lansdorp PM, Reid DS, Eaves CJ (1996) Self-renewal of primitive human hematopoietic cells (long-term culture-ini-tiating cells) in vitro and their expansion in defined medium. Proc NatlAcad Sci U S A 93(4):1470
32. Glimm H, Eaves CJ (1999) Direct evidence for multiple self-renewaldivisions of human in vivo repopulating hematopoietic cells in short-term culture. Blood 94(7):2161
33. Goga A, McLaughlin J, Afar DEH, Saffran DC, Witte ON (1995) Al-ternative signals to RAS for hematopoietic transformation by theBCR-ABL oncogene. Cell 82:981
34. Skorski T, Kanakaraj P, Nieborowska-Skorska M, et al. (1995) Phosphati-dylinositol-3 kinase activity is regulated by BCR/ABL and is required forthe growth of Philadelphia chromosome-positive cells. Blood 86:726
35. Ilaria RL, Jr, Van Etten RA (1996) P210 and P190(BCR/ABL) inducethe tyrosine phosphorylation and DNA binding activity of multiplespecific STAT family members. J Biol Chem 271(49):31704
36. Gothot A, Pyatt R, McMahel J, Rice S, Srour EF (1997) Functionalheterogeneity of human CD34(1) cells isolated in subcompartmentsof the G0/G1 phase of the cell cycle. Blood 90(11):4384
37. Holyoake T, Jiang X, Eaves C, Eaves A (1999) Isolation of a highlyquiescent subpopulation of primitive leukemic cells in chronic mye-loid leukemia. Blood 94(6):2056
38. Udomsakdi C, Eaves CJ, Swolin B, Reid DS, Barnett MJ, Eaves AC(1992) Rapid decline of chronic myeloid leukemic cells in long-termculture due to a defect at the leukemic stem cell level. Proc Natl AcadSci U S A 89:6192
39. Petzer AL, Eaves CJ, Lansdorp PM, Ponchio L, Barnett MJ, Eaves AC(1996) Characterization of primitive subpopulations of normal andleukemic cells present in the blood of patients with newly diagnosed aswell as established chronic myeloid leukemia. Blood 88(6):2162
40. Dao MA, Hashino K, Kato I, Nolta JA (1998) Adhesion to fibronectinmaintains regenerative capacity during ex vivo culture and transduc-tion of human hematopoietic stem and progenitor cells. Blood92(12):4612


















