CD3 TAA-BiTE -CD3 complex TAA - Amgen Oncology · a range of tumors: one arm engages CD3 to...
16
BiTE ® antibody TAA-BiTE ® -CD3 complex TAA CD3 BiTE ® Antibodies Designed to Bridge T Cells to Cancer Cells EDUCATIONAL SLIDE KIT INCLUDED
Transcript of CD3 TAA-BiTE -CD3 complex TAA - Amgen Oncology · a range of tumors: one arm engages CD3 to...

BiTE® antibody
TAA-BiTE®-CD3 complex
TAA
CD3
BiTE® AntibodiesDesigned to Bridge T Cells to Cancer Cells
EDUCATIONAL SLIDE KIT INCLUDED

Naive T cell
Cytotoxic T Lymphocytes (CTLs) Are Highly Specific and Potent Effector Cells
The immune system represents an important part of the host anti-tumor defense system. Cytotoxic T lymphocytes, or CTLs, are principal components of immune function. CTLs are highly specific and potent effector cells that mediate directed lysis of target cells.1 Acquisition of the T cell receptor (TCR) is an important feature of initial T cell differentiation. The TCR is a complex of integral membrane proteins that participates in the activation of T-cells in response to the presentation of antigen.The glycoprotein CD3 is an essential component of the TCR responsible for signal transduction. Because it is capable of significant signal amplification within CTLs, only a small number of antigen binding events involving CD3 are required to initiate effector function.1,2
CTLs cannot interact with free antigens.1 Cells in the body present antigens via class I major histocompatibility complex (MHC I) molecules. CTL activation leads to expansion and differentiation of the antigen-specific T cell clones. Activated CTLs enlarge, become polarized, and synthesize unique lysosomal granules necessary for effector function. Upon recognizing an antigen, CTLs engage target cells, initiating the cell lysis process.

Naive T cell
Cancer cells
Cancer Evasion From T Cell Recognition and Destruction
CTLs play a critical role in identification and destruction of viral-infected host cells.3 Since viruses replicate exclusively within cells, their destruction depends on the elimination of the cells they’ve infected. Viral antigens presented by MHC molecules on target cells are specifically recognized by CTLs. Upon CTL binding of the target cell, apoptosis, or programmed cell death, is rapidly induced. CTLs represent a particularly effective form of host defense against viruses, intracellular bacteria, and some protozoan infections.
CTLs are also well known to have the capacity to detect cancer cells, and many investigators have sought to exploit the capacity of T cells to lyse target cells as anticancer therapy.4 Nonetheless, endogenous T cell responses are observed to be generally ineffective in promoting targeted lysis of cancer cells. Individuals with cancer have been shown to demonstrate low but detectable cellular responses to tumor-associated antigens.5 Studies have shown that circulating T cells recognize tumor-associated antigens in patients with advanced cancer, and some are able to generate an immune response in vitro.6 Peripheral T cell responsiveness did not affect survival among patients with cancer. The presence of intratumoral T cells correlates with improved clinical outcomes among patients with a certain type of cancer and is associated with molecular evidence of activation of antitumor mechanisms.7 These findings indicate that while T cells are able to detect cancer cells, downstream killing mechanisms are not fully functional.8
Activated cytotoxic T cell

Strategies That Harness the Cytotoxic Activity of CTLs
CTLs bind to target cells, initiating lysis via mechanisms that facilitate necrosis and apoptosis.1 This observation has led to the development of various strategies that harness the cytotoxic activity of CTLs to target cancer cells.2,9,10 One such approach is the use of vaccines where antigen presenting cells (APCs) are exposed to tumor specific molecules in order to activate CTLs.2 The use of adoptive T cell transfer is also in development. Tumor specific T cells are rapidly expanded ex vivo and then re-infused into the patient. These strategies however have produced variable response rates, most likely due to the cancer cell’s ability to develop various mechanisms to evade T cell-recognition and prevent an antigen-specific T cell-response.9,11

Cancer Cells Evade Host Immune Mechanisms
Cancer cells develop various mechanisms that prevent an antigen-specific CTL response and evade T cell-recognition.2,4,11 Tolerance is a principal mechanism of immune escape whereby CD4+ T regulatory cells limit inflammation and suppress immune response. Peripheral tolerance includes deletion, in which T cells expressing a TCR with high affinity for a specific tumor antigen are deleted; ignorance, in which reactive T cells persist but are not activated by the antigen; and anergy, in which T cells are unresponsive after TCR binding in the absence of co-stimulation.4
The local tumor environment may also act to suppress immune recognition of cancer cells. The cancer cells themselves produce inhibitory cytokines that recruit T regs and inhibit maturation of APCs.4 Tumor stroma acts as a physical barrier to the incursion of immune cells, and produces cytokines that stimulate cancer cell growth. As cancer cells continue to proliferate and acquire additional mutations, they gain additional rescue mechanisms that act to further impede immune surveillance.
Abnormalities in antigen presentation are also common; these include downregulation of MHC I molecules related to loss of heterozygosity in 6p21, mutated ß2-microglobulin, abnormalities in transcription regulation, and DNA hypermethylation.11 These all further contribute to the ability of cancer cells to evade immune recognition.

Granzyme
Perforin
T cell
CTLs Effect Cell Lysis
To effect target cell lysis, CTLs require cell-to-cell contact; the TCR on the CTL must recognize and engage the appropriately MHC-presented antigen on the target cell.1 This initiates a signaling cascade within the CTL, causing T cell activation, driving production of a variety of cytokines, and leading to the creation of a cytolytic synapse between the CTL and its target.
Perforin and granzymes are highly toxic molecules that are stored in preformed granules that reside in activated CTLs.1,9 The cytoplasmic granules migrate toward the CTL membrane, to ultimately fuse with it, releasing their contents in directed fashion into the cytolytic synapse, toward the target cancer cell.1,9,12 Perforin, a 67 kDa protein, oligomerizes to form a ring-like pore structure within the membrane of the target cell, disrupting target cell plasma membrane stability.13,14
The pore created by perforin acts as a port of entry for granzymes. Granzymes are a family of serine proteases that are packaged in cytoplasmic granules in CTLs that induce apoptosis of target cells.15 Granzymes may promote cell death via activation of caspases, or by different mechanisms.14-16

Fused granule
Peptide
MHC1
CD3TCR
Formation of perforin pore
Cancer cell
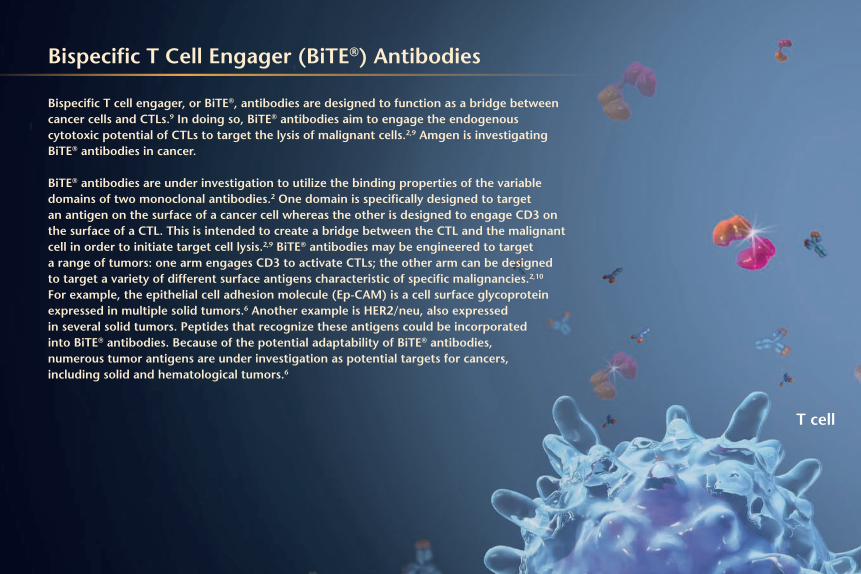
Bispecific T Cell Engager (BiTE®) Antibodies
Bispecific T cell engager, or BiTE®, antibodies are designed to function as a bridge between cancer cells and CTLs.9 In doing so, BiTE® antibodies aim to engage the endogenous cytotoxic potential of CTLs to target the lysis of malignant cells.2,9 Amgen is investigating BiTE® antibodies in cancer.
BiTE® antibodies are under investigation to utilize the binding properties of the variable domains of two monoclonal antibodies.2 One domain is specifically designed to target an antigen on the surface of a cancer cell whereas the other is designed to engage CD3 on the surface of a CTL. This is intended to create a bridge between the CTL and the malignant cell in order to initiate target cell lysis.2,9 BiTE® antibodies may be engineered to target a range of tumors: one arm engages CD3 to activate CTLs; the other arm can be designed to target a variety of different surface antigens characteristic of specific malignancies.2,10 For example, the epithelial cell adhesion molecule (Ep-CAM) is a cell surface glycoprotein expressed in multiple solid tumors.6 Another example is HER2/neu, also expressed in several solid tumors. Peptides that recognize these antigens could be incorporated into BiTE® antibodies. Because of the potential adaptability of BiTE® antibodies, numerous tumor antigens are under investigation as potential targets for cancers, including solid and hematological tumors.6
T cell

CD3-specific domain
BiTE® antibody
Cancer cells
Investigational BiTE® antibodies utilize the binding properties of the variable domains of two distinct monoclonal antibodies.

T cellBiTE® Antibodies Are Designed to Bridge Cancer Cells and CTLs to Enable Cancer Cell Lysis
Investigational BiTE® antibodies may circumvent cancer cell escape mechanisms and other modes of drug resistance.2,10,11,17-19 BiTE® antibodies are designed to enhance CTL response by eliminating the need for antigen processing, MHCI/ß2-microglobulin and peptide antigens as well as co-stimulatory molecules.2 BiTE® antibodies target specific antigens expressed on the surface of malignant cells.9
Upon binding of both arms of BiTE® antibodies at their specific receptors, CTLs and cancer cells are forced into close proximity with one another.9 The tumor antigen specific arm of the BiTE® antibody targets a cancer cell, and the CD3 arm engages a CTL, creating a link between the cells. This allows for initiation of the signaling cascade mediated by the T cell receptor.9 As a result, a cytolytic synapse is created between the effector CTL and the target cancer cell, perforin and granzymes are released, and target cell lysis ensues.

Granzymes flowing through perforin pore
Activated caspase
Inactive caspase
Fused granule
TAA-BiTE-CD3 complex
TAA
CD3T cell
Cancer cell

After target cell destruction through lysis, BiTE® antibodies are designed to move through the local environment and locate additional target cells and initiate cell death.
BiTE®-Activated T Cells and Serial Lysis of Multiple Target Cells In Vitro
It is important to recognize that CTLs are not consumed during lysis of target cells; therefore, after causing lytic destruction of one target cell, an activated CTL can move on through the local environment toward other target cells, engage target antigen binding regions, and initiate additional cell lysis.20 A single CTL may be capable of causing lysis in multiple target cells. In vitro studies utilizing video-assisted microscopy indicate that individual activated CTLs may be capable of causing lyses of multiple target cells. The rate of serial lysis of cancer cells is mostly a function of CTL movement and target cell apoptosis.
Furthermore, in addition to lysing target cells, activated CTLs may release proinflammatory cytokines and produce even more perforin and granzymes, supporting subsequent interactions to engage target cells.2,9 Importantly, the sustained activation of CTLs during cell lysis forces T cells into the cell cycle, resulting in enhanced local T cell expansion.
Taken together, these two mechanisms lead to enhanced effects; as the population of CTLs increases locally in the target tissue and the effector-to-target ratio improves; a more complete elimination of the target cell population can be achieved than with each mechanism alone.


References
1. Groscurth P, Filgueira L. News Physiol Sci. 1998;13:17-21.
2. Baeuerle PA, Kufer P, Bargou R. Curr Opin Mol Ther. 2009;11:22-30.
3. Janeway Jr CA, Traveres P, Walport M et al. 5th ed. New York. Garland Science; 2001. NCBI bookshelf ID: NBK27101.
4. Disis ML, Bernhard H, Jaffee EM. Lancet. 2009.373;673-683.
5. Biragyn A, Longo DL. Semin Cancer Biol. 2012;22:50-59.
6. Nagorsen D, Keilholz U, Rivoltini L, et al. Cancer Res. 2000;60:4850-4854.
7. Zhang L, Conejo-Garcia JR, Katsaros D, et al. N Engl J Med. 2003;348:203-213.
8. Nagorsen D, Scheibenbogen C, Letsch A, et al. J Translational Med. 2005;3:doi:10.1186/1479-5876-3-3.
9. Nagorsen D, Baeuerle PA. Exp Cell Res. 2011;317:1255-1260.
10. Choi BD, Cai M, Bigner DD, et al. Expert Opin Biol Ther. 2001;11:843-853.
11. Töpfer K, Kempe S, Müller N, et al. J Biomed Biotech. 2011. Article 918471.
12. Haas C, Krinner E, Brischwein K, et al. Immunobiol. 2009;214:441-453.
13. Duke RC, Persechini PM, Chang S, et al. J Exp Med. 1989;170:1451-1456.
14. Anthony DA, Andrews DM, Watt SV, et al. Immunol Rev. 2010;235:73-92.
15. Ewen CL, Kane KP, Bleackley RC. Cell Death Differen. 2012;19:28-35.
16. Talanian RV, Yang XH, Turboc J, et al. J Exp Med. 1997;186:1323-1331.
17. Offner S, Hofmeister R, Romaniuk A, et al. Mol Immunol. 2006;43:763-771.
18. Baeuerle PA and Reinhardt C. Cancer Res. 2009;69:4941-4944.
19. Bargou R, Leo E, Zugmaier G, et al. Science. 2008;321:974-977.
20. Hoffmann P, Hofmeister R, Brischwein K, et al. Int J Cancer. 2005;115:98-104.

BiTE® Antibodies: Designed to Bridge T Cells to Cancer Cells An Educational Slide Deck
Provided as an educational resource. Do not copy or distribute.©2013 Amgen Inc. All rights reserved.73597-R1-V1

This booklet contains forward-looking statements that are based on Amgen’s current expectations and beliefs and are subject to a number of risks, uncertainties, and assumptions that could cause actual results to differ materially from those described. All statements, other than
statements of historical fact, are statements that could be deemed forward-looking statements. Forward-looking statements involve significant risks and uncertainties, including those more fully described in the Risk Factors found in the most recent Annual Report on Form 10-K and
periodic reports on Form 10-Q and Form 8-K filed by Amgen with the U.S. Securities and Exchange Commission, and actual results may vary materially. Except where otherwise indicated, Amgen is providing this information as of March 29, 2013 and does not undertake any obligation to update any forward-looking statements contained in this booklet as a result of new information, future events, or otherwise.
Provided as an educational resource. Do not copy or distribute. ©2013 Amgen Inc. All rights reserved.G73617-R1-V1 73596-R1-V1