
AQUATIC MACROPHYTES IN THE TROPICS: ECOLOGY...
28
AQUATIC MACROPHYTES IN THE TROPICS: ECOLOGY OF POPULATIONS AND COMMUNITIES, IMPACTS OF INVASIONS AND USE BY MAN S. M. Thomaz Department of Biology/Nupélia, Maringá State University, Paraná, 87020-900, Brazil F. A. Esteves Department of Ecology/Nupem, Federal University of Rio de Janeiro, Brazil K. J. Murphy Division of Environmental and Evolutionary Biology, Institute of Biomedical and Life Sciences, Graham Kerr Building, University of Glasgow, Glasgow G12 8QQ, UK A. M. dos Santos State University of Montes Claros, Minas Gerais, Brazil A. Caliman Department of Ecology, Federal University of Rio de Janeiro, Brazil R. D. Guariento Department of Ecology, Federal University of Rio de Janeiro, Brazil Key-words Aquatic biodiversity, nutrient cycling, food webs, nuisance species, assemblages Summary 1. Introduction The term ‘aquatic macrophytes’ refers to large plants visible to the naked eye and having at least their vegetative parts growing in permanently or periodically aquatic habitats. These plants colonize a variety of aquatic habitats and can be divided into the following life forms: rooted submerged – plants that grow completely submerged and are rooted into the sediment (e.g. elodea, Elodea canadensis); free-floating – plants that float on or under the water surface (e.g. water hyacinth, Eichhornia crassipes); emergent – plants rooted in the sediment with foliage extending into the air (e.g. cattail, Typha domingensis); and floating-leaved – plants rooted in the sediment with leaves floating on the water surface (e.g. water lilies, Nymphaea spp). An additional two life forms have been proposed: epiphytes – plants growing over other aquatic macrophytes (e.g. Oxycarium cubense); and amphibious – plants that live most of their life in saturated soils, but not necessarily in water (e.g. Polygonum spp).
-
Upload
nguyenhuong -
Category
Documents
-
view
216 -
download
0
Transcript of AQUATIC MACROPHYTES IN THE TROPICS: ECOLOGY...

AQUATIC MACROPHYTES IN THE TROPICS: ECOLOGY OF POPULATIONS AND COMMUNITIES, IMPACTS OF INVASIONS
AND USE BY MAN S. M. Thomaz Department of Biology/Nupélia, Maringá State University, Paraná, 87020-900, Brazil F. A. Esteves Department of Ecology/Nupem, Federal University of Rio de Janeiro, Brazil K. J. Murphy Division of Environmental and Evolutionary Biology, Institute of Biomedical and Life Sciences, Graham Kerr Building, University of Glasgow, Glasgow G12 8QQ, UK A. M. dos Santos State University of Montes Claros, Minas Gerais, Brazil A. Caliman Department of Ecology, Federal University of Rio de Janeiro, Brazil R. D. Guariento Department of Ecology, Federal University of Rio de Janeiro, Brazil Key-words Aquatic biodiversity, nutrient cycling, food webs, nuisance species, assemblages Summary 1. Introduction
The term ‘aquatic macrophytes’ refers to large plants visible to the naked eye and
having at least their vegetative parts growing in permanently or periodically aquatic habitats. These plants colonize a variety of aquatic habitats and can be divided into the following life forms: rooted submerged – plants that grow completely submerged and are rooted into the sediment (e.g. elodea, Elodea canadensis); free-floating – plants that float on or under the water surface (e.g. water hyacinth, Eichhornia crassipes); emergent – plants rooted in the sediment with foliage extending into the air (e.g. cattail, Typha domingensis); and floating-leaved – plants rooted in the sediment with leaves floating on the water surface (e.g. water lilies, Nymphaea spp). An additional two life forms have been proposed: epiphytes – plants growing over other aquatic macrophytes (e.g. Oxycarium cubense); and amphibious – plants that live most of their life in saturated soils, but not necessarily in water (e.g. Polygonum spp).

Macrophytes include macroalgae of the divisions Chlorophyta (green algae), Xanthophyta (yellow-green algae) and Rhodophyta (red algae) and the “blue-green algae” (more correctly known as Cyanobacteria); Bryophyta (mosses and liverworts); Pteridophyta (ferns); and Spermatophyta (seed-bearing plants). However, most of the literature devoted to freshwater macrophytes has investigated three major groups: the Charales (an order of Chlorophyta comprisingy large – up to 2 m – and relatively complex multicellular algae), together with the vascular plant groups, Pteridophyta and Spermatophyta.
Macrophytes colonize virtually all freshwater habitats, from the tiny “living ponds” provided by Bromeliaceae (e.g. Utricularia spp), to thermal springs (e.g. Najas tequefolia) and waterfalls (e.g. members of the Podostemaceae colonize even the giant Iguaçu Falls, Brazil/Argentina). Most rivers, lakes, lagoons and reservoirs are colonized to differing degrees by macrophytes, whilst wetlands are characterized as areas where macrophytes dominate.
Studies on aquatic macrophytes, and especially their ecology, were few in number before the 1960s. The reasons are historical because the science of limnology primarily originated in north-temperate countries, where deep lakes are characteristic: such freshwater systems are amongst the least favorable of habitats to support aquatic macrophytes. Consequently, phytoplankton was considered (correctly) as the main primary producer and pelagic food webs were prioritized in those studies. A great increase in the literature concerning macrophytes occurred after 1960, caused probably by the recognition that a great number, if not most, aquatic ecosystems were in fact shallow, with extensive littoral regions favorable for supporting aquatic macrophyte communities. A second factor was increasing recognition of the role played by macrophytes in the biodiversity-support functioning of freshwater systems: vital for many animal communities, such as aquatic invertebrates, fish and aquatic birds.
In this article it is not possible to cover all relevant topics in depth: the literature on tropical macrophyte ecology and management is too large for this to be possible. We utilise Neotropical ecosystems (which support the highest macrophyte diversity) for many of our examples but also include data from tropical and sub-tropical Australasia, Africa and Asia. Following the publication guidelines for this book, we cited only the 20 most used literature items for the article. However, we also used numerous other references which are provided in a separate table (see Appendix 1). The link of these references to each specific topic considered in our article will be provided by the first author ([email protected]) upon request.
2. General features of macrophytes
2.1. Evolution Although still controversial, the origin of terrestrial plants is generally agreed to be
from green algae of the order Charales, known as stoneworts. After colonizing the land, representatives of numerous different families returned to water, colonizing both freshwater and marine ecosystems, with good evidence for at least 211 (but probably more) independent colonization events of this nature having occurred.
It is interesting to note that angiosperms began the return to water very early in their evolutionary history. An analysis of the angiosperm phylogenetic tree shows the terrestrial

shrub Amborella trichopoda as the first diverging lineage from the main branch of the angiosperm phylogenetic tree, but the families Cabombaceae, Nymphaeaceae and Hydatellaceae, which comprise only aquatic species, occupy the second basal lineage. Fossil material collected in the Vale de Agua locality (in a complex of clay pits situated in the Beira Littoral, Portuguese Basin) and in Crato (Northeast Brazil) confirms that water lilies have colonized this region since the Early Cretaceous (125-115 Mya). Thus, some adaptations found in extant submerged species, like aerial pollination and presence of stomata (see below), are interpreted only under an evolutionary perspective.
2.2. Main adaptations to life in water The aquatic habitat imposes strong pressures on plant survival. Although all life
forms of macrophytes face a limiting environment, pressures on survival are strongest for submerged plants, the ones that have most fully completed the evolutionary return of angiosperms to the aquatic habit. Water has greater density and viscosity compared to air and thus gases (including CO2, needed for photosynthesis) diffuse at extremely low rates in water, compared to in air. This is even more prominent in lentic ecosystems, where the absence of flow means that large boundary-layers may surround leaves, leading to rapid depletion of CO2 near plant surfaces. Aquatic ecosystems also have often-anoxic sediments which cause problems for root survival. In addition, light may be strongly reduced beneath the water surface not only by absorption of light energy by water molecules, but also by the presence of biogenic (e.g. algae) and abiogenic (e.g. silt and clay) suspended matter, as well as dissolved organic matter (usually humic substances). Again, light limitation primarily affects mainly rooted-submerged plants, which have consequently evolved a number of adaptations to cope with light limitation and other pressures on plant survival in water.
Concerning CO2 acquisition, submerged macrophytes display an array of physiological and exploitation strategies to ameliorate the carbon constraints within the aquatic medium. Probably one of the most common strategies is the use of the ion bicarbonate (HCO3
-), the concentration of which in the ocean and in most fresh-waters (except soft water ecosystems) are high compared to dissolved CO2. However, CO2 is still the preferred form of inorganic carbon used by most aquatic plants, since the exploitation of bicarbonate involves the expensive synthesis of a complex of enzymes, such as carbonic anhydrase, thereby elevating the energetic cost of photosynthesis. Even so, this mechanism is efficient in waters with high pH values (>8.0), where CO2 is scarce or even absent, providing a competitive advantage to species able to assimilate bicarbonate. About 50% of species so far tested show evidence of bicarbonate use (attesting the efficiency of this strategy in carbon acquisition). Examples of species that use bicarbonate are Egeria densa, Egeria najas (both native to South America), Elodea canadensis and Potamogeton spp.
Other physiological strategies to overcome carbon limitation involve the use of C4 enzymes or crassulacean acid metabolism (CAM). Examples of species with C4 - like metabolism include Egeria densa, Elodea nuttallii and Hydrilla verticillata while CAM metabolism is found in Isoetes bolanderi, Crassula, Littorella, and Sagittaria, amongst others.
Exploitation strategies to overcome CO2 limitation involve morphological and anatomical adaptations that allow plants to obtain alternative sources of carbon, in addition to that present in the water medium. Floating or aerial leaves are common in several species (e.g. Cabomba furcata, Myriophyllum brasiliense and Potamogeton amplifolius) and they

allow plants to absorb CO2 directly from the atmosphere. Other species use their roots to exploit the high concentrations of CO2 found in sediments, usually well above the concentrations found in water. Species using CO2 from sediments have modified transport vessels to permit movement of CO2 from the roots to leaves and high root:shoot weight ratios (0.5 - 2.0). This strategy is found mainly in isoetids (e.g. Littorella and Lobelia), providing a very specialized adaptation for this group whose species mainly colonize oligotrophic softwater habitats, where CO2 is usually scarce in the water. Highly dissected and thinner leaves, compared to terrestrial angiosperms, found for example in Cabomba and Ceratophyllum, increase leaf surface area and also thereby increase carbon acquisition rates.
The problems with anoxic sediment faced by aquatic macrophytes have led many macrophytes to aerenchyma, a tissue containing gas spaces linking roots to leaves. Oxygen is transported from leaves toward roots by this system. In addition, some species (e.g., Ludwigia adscendens) develop air roots that may be interpreted as short circuits to the atmosphere, allowing greater transport of oxygen to the submerged and underground organs.
Concerning light limitation under water, most plants cope with this in part by increasing pigment content in leaves and locating chloroplast-containing cells in the superficial epidermis. Most submerged macrophytes are considered “shade plants”, since they are very efficient photosynthesisers at low light levels. This is usually achieved by reduced respiration rates, and by reducing the thickness of their leaves. Several submerged freshwater macrophytes also elongate shoots and concentrate their photosynthetic tissues close to the water surface, in a strategy known as “canopy forming” (e.g. Egeria densa and Hydrilla verticillata). This strategy renders these species (known as elodeids) a great competitive ability and once they colonize a habitat, they may eliminate others colonizing only the deeper parts of the water column (especially the isoetids, such as Isoetes).
Submerged plants typically have conspicuously reduced cuticule, useful to minimize water loss in terrestrial habitats but superfluous under water. Vascular tissues, such as xylem, and structural tissue such as lignin are also characteristically reduced. Although important for terrestrial plants, these tissues lose their function under water, since this medium furnishes support for plants. By reducing the need to synthesize such tissues aquatic macrophytes further reduce energetic costs.
Finally, most submerged species still depend on the aerial medium for reproduction, giving that pollination is primarily by insects. Flowers of these species are usually positioned above the water surface. The flowers of the carnivorous Utricularia foliosa, for example, reach up to 10 cm above the water surface, while Vallisneria spp. produce long peduncles that may reach more than 1 meter and raise the female flowers into the air. However, some species of Callitriche, Najas and Ceratophyllum, among others, have developed hydrophilic pollination, thereby completing the adaptation of their sexual reproduction to the aquatic environment. 3. Importance of macrophytes for ecosystem structure and functioning
Macrophytes affect aquatic ecosystems in a variety of ways, especially the shallower
ones where they colonize large areas. These plants change the water and sediment physicochemistry, influence nutrient cycling, may serve as food for invertebrates and

vertebrates, both as leaves and dead biomass (detritus) and, in particular, change the spatial structure of the waterscape by increasing habitat complexity. These roles of aquatic plants have been extensively shown in temperate regions but they also occur in tropical habitats.
Aquatic plants are not inert objects, but active organisms whose metabolism affects the water medium. The littoral region usually differs from the limnetic one in terms of the thermal regime, gases, concentrations of ions (including the most limiting nutrients nitrogen and phosphorus), pH and dissolved carbon, among other features. Under the higher temperatures of tropical waters, oxygen is usually super-saturated, CO2 is non-detectable and pH values may easily reach 9.5 inside stands of submerged plants at noon.
Nutrients are released rapidly during decomposition contributing to the inorganic and organic nutrient pools in the water. The majority of phosphorus, calcium, magnesium and other ions are released from decomposing detritus in the first week. Giving the high stocks of nutrients contained in macrophytes, their release through decomposition strongly affects the water column. In a neotropical floodplain lagoon (Mogi, Brazil), 71% of nitrogen and phosphorus were found in the biomass of Eichhornia azurea and Scirpus cubensis, and only 29% were in the water column. High stocks of nutrients were also measured in a Neotropical reservoir (Lobo, Brazil): the biomass of Nymphoides indica and Pontederia cordata together had 12 and 5 times more nitrogen, and 18 and 24 times more phosphorus, than the water of the littoral and limnetic regions, respectively. It is interesting to note that part of these nutrients were locked up in sediments, but through macrophyte activity they are made available for periphyton and plankton.
In temperate ecosystems nutrients are released through decomposition during fall, when macrophyte shoots die back, but in tropical, more temperature-constant aquatic habitats, release can occur thorough the year. However, even in the tropics, several aquatic ecosystems do have seasons (e.g., rainy and dry) and may have periods of greater nutrient inputs by plant decomposition. For example, together with decomposition of the amphibious vegetation, that covers most of river-floodplain habitats, the decomposition of aquatic macrophytes contributes to increase nutrients and reduces oxygen in these ecosystems during high water periods, when those plant species unable to tolerate inundation die and decompose. In higher-latitude tropical areas also seasonal differences in temperature, also affects decomposition. Significant effects of temperature in tropical areas have been experimentally shown: an increase of temperature from 17 to 27o C (a range easily found in several tropical aquatic ecosystems), increased Egeria najas decomposition rates threefold. In addition, water oxygen decreased and ions increased from 3 to 5 times faster at 27o C than at 17 o C.
Macrophytes also contribute indirectly to nutrient cycling by releasing dissolved organic matter that, in turn, supports the activity of nitrogen-fixing bacteria. This semi-symbiotic relationship, in which the heterotrophic bacteria are favored by organic compounds released by macrophytes and in turn furnishes nitrogen to these plants, has been shown for several species of macrophytes (e.g. Utricularia sp, Eichhornia crassipes, Nymphoides indica). In general, nitrogen fixation is higher close to the rhizosphere and thus, concentrations of nutrient in sediments may be increased by this means, with the surplus being released to the water column. In addition to nutrient amendments in sediment, organic matter is also increased in this compartment under macrophyte colonization.
Giving the high production and great colonization of waterbodies by macrophytes, it is tempting to suggest that the exceptional biomass produced by these plants enters food webs, either directly by grazing (herbivory food webs) as well as through detritus

(detritivorous food webs). In fact, macrophytes have been shown to compose the diet of several fish species in a variety of tropical freshwater ecosystems. Maybe the most studied example of macrophyte herbivorous in tropics is the grass carp, which feeds on several species of macrophytes. Invertebrates also use several species of tropical macrophyte directly as food. For example, in the northeastern Argentina, at least 23 species of invertebrates were found feeding on 13 species of aquatic macrophytes. Not only leaves are eaten, but pollen has also been shown to compose the main diet of bees. This was found for several species of the emergent Ludwigia in wetlands of South America.
Despite being used by fish and invertebrates, as revealed by stomach content analyses, the first studies carried in tropical wetlands using stable isotopes raised doubts about the importance of macrophytes as a source of energy for higher trophic levels. An early investigation carried with detritivorous fish in the 1980s in the Amazonian floodplain suggested that phytoplankton, instead of higher plants, composed the base of food webs. Although detecting a varying importance of macrophytes for higher trophic levels, the predominance of algae (both phytoplanktonic and periphytic) in tropical floodplains as primary sources of energy have been confirmed by several other more recent studies. These results led to a paradox: the higher biomass and faster production of macrophytes in tropical waters are apparently not significant as source of energy for higher trophic levels. However, recent studies have shown different trends. Investigations carried in an Amazonian floodplain lake indicated that C4 macrophytes may contribute up to 59% of the carbon for two species of fish. In another study carried in Rio Grande (Mexico), fish larvae obtained carbon predominately from algal production in early summer, but used organic carbon derived from emergent macrophytes as river discharge decreased in mid-summer. Thus, this area is still open, since it seems that macrophytes do play a major role as energy sources at least in specific ecosystems or habitats, or during part of the seasonal cycle. In addition, the massive quantity of detritus produced during macrophyte decomposition releases dissolved organic matter which, together with particulate matter, sustains microbial food webs. Thus, independently of being important as a basic resource for food chains composed of large organisms, the “burning” of organic matter originated from macrophytes may drive nutrient cycling in aquatic ecosystems. This aspect is poorly known in tropical waters where microorganism activity is believed to be much faster than in temperate waters.
In addition to aquatic organisms, there are several species of terrestrial animals such as birds and mammals, that use regularly macrophytes as food in the tropics. Good examples are manatee (Trichechus inunguis), deer (Blastocerus dichoromus) and capybara (Hydrochoerus hydrochaeris) in South America; hippopotamus (Hippopotamus amphibious) in Africa; and goose (Anseranus semipalmata) in Australia. Terrestrial invertebrates may feed heavily on macrophytes: the combined effects of the coleopteran (adults) Neochetina bruchi and N. eichhorniae, together with the larvae of the dipteran Thrypticus sp. may cause extensive damage to natural populations of water hyacinth in the Neotropics. These and other herbivorous insect species are regularly used in the biological control of water hyacinth, even in other continents.
If the role of macrophytes as energy source is still a matter of debate, there is a consensus that this vegetation plays a major role by increasing the habitat complexity of waterscape, at different scales. Increasing complexity (also known as habitat heterogeneity) has direct and positive impacts upon the aquatic biota by increasing food at small scales and by providing refuge for aquatic invertebrates, small sized, and young fish, that use the

littoral zones to feed, escape from predators or as nesting sites. Together, these mechanisms lead to the maintenance of the high diversity found in littoral habitats (Fig. 1).
The positive effects of increased complexity provided by macrophytes have been widely suggested in tropical streams, lakes and reservoirs. Studies in Neotropical reservoirs show that the benefits for fish diversity and densities are recognized at different spatial scales: at a single stand of macrophytes, in different arms of a same reservoir and among reservoirs of different basins. The importance of aquatic plants as key components of aquatic ecosystems raises the possibility of using macrophytes as a tool to manipulate aquatic habitats, aiming at increasing the fish densities and diversity in tropical reservoirs, since in many cases these man-made ecosystems are poorly structured.
AQUATIC MACROPHYTES
INCREASEHABITAT
COMPLEXITY
INCREASE THESURFACE FOR
COLONIZATION BYMICROORGANISMS
INCREASEREFUGIA
AVAILABILITY
COLONIZATIONBY PERIPHYTON
INCREASEFOOD FOR
INVERTEBRATEAND SMALL
FISH
ATRACTION OF INVERTEBRATES,
JUVENILE FISHAND SMALLBODY-SIZED
FISH
INCREASE SPECIES
RICHNESS
Figure 1. Some possible mechanisms lading to higher species richness in littoral habitats.
4. Macrophytes in populations
Tropical ecosystems were seen as different from temperate ones not only because of
their diversity (see “Biodiversity and Endemism” in this chapter), but also as places with small seasonal variation. This assumption was largely based on the relatively constant temperatures during the year, but it is not sustained when we consider the great variations in rainfall. Besides producing direct effects upon aquatic ecosystems, seasonal rains cause strong and predictable water level fluctuations, which lead aquatic populations to respond with morphological, physiological and behavioral features. Macrophytes are no exception, and populations are also strongly affected by such environmental changes often showing conspicuous seasonal alterations.
The responses of the emergent Eleocharis interstincta to annual drawdowns, caused by a combination of low rain and sand bar breaching, were studied in a Brazilian coastal lagoon. Both stem height (R2 = 0.90; p< 0.05) and stem biomass (R2 = 0.65; p< 0.05) were positively affected by water level (Fig. 2a). The adaptations of E. interstincta for solving
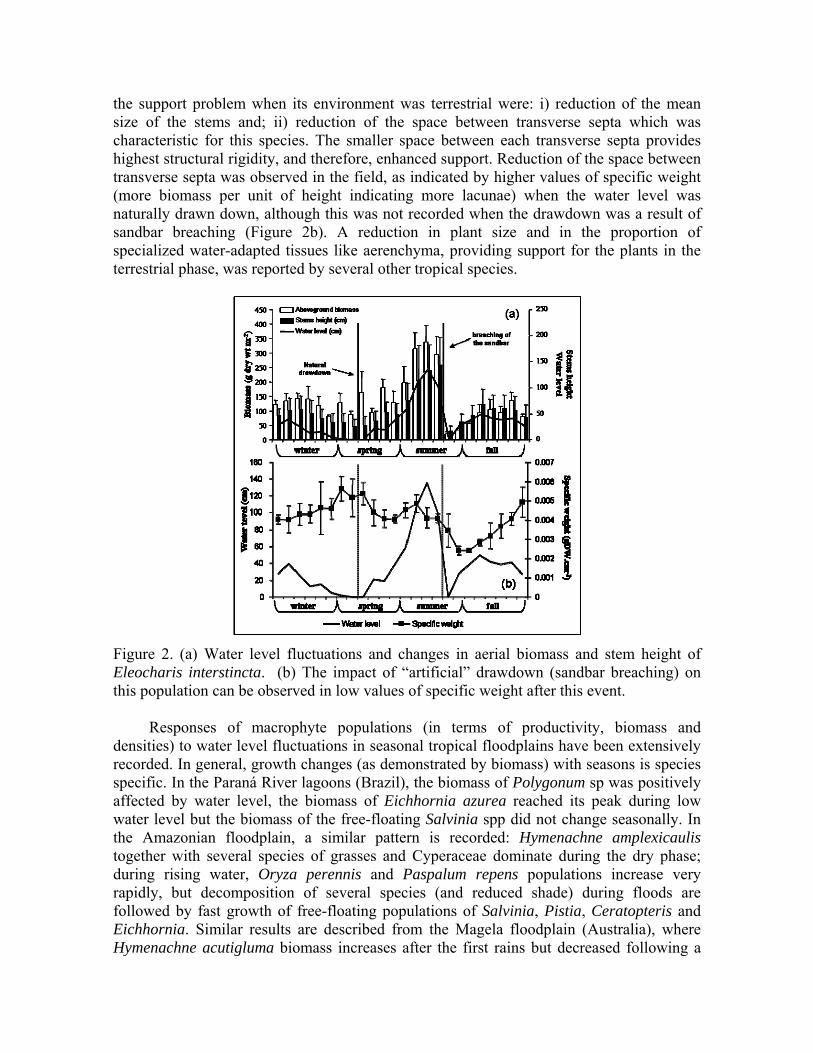
the support problem when its environment was terrestrial were: i) reduction of the mean size of the stems and; ii) reduction of the space between transverse septa which was characteristic for this species. The smaller space between each transverse septa provides highest structural rigidity, and therefore, enhanced support. Reduction of the space between transverse septa was observed in the field, as indicated by higher values of specific weight (more biomass per unit of height indicating more lacunae) when the water level was naturally drawn down, although this was not recorded when the drawdown was a result of sandbar breaching (Figure 2b). A reduction in plant size and in the proportion of specialized water-adapted tissues like aerenchyma, providing support for the plants in the terrestrial phase, was reported by several other tropical species.
Figure 2. (a) Water level fluctuations and changes in aerial biomass and stem height of Eleocharis interstincta. (b) The impact of “artificial” drawdown (sandbar breaching) on this population can be observed in low values of specific weight after this event.
Responses of macrophyte populations (in terms of productivity, biomass and densities) to water level fluctuations in seasonal tropical floodplains have been extensively recorded. In general, growth changes (as demonstrated by biomass) with seasons is species specific. In the Paraná River lagoons (Brazil), the biomass of Polygonum sp was positively affected by water level, the biomass of Eichhornia azurea reached its peak during low water level but the biomass of the free-floating Salvinia spp did not change seasonally. In the Amazonian floodplain, a similar pattern is recorded: Hymenachne amplexicaulis together with several species of grasses and Cyperaceae dominate during the dry phase; during rising water, Oryza perennis and Paspalum repens populations increase very rapidly, but decomposition of several species (and reduced shade) during floods are followed by fast growth of free-floating populations of Salvinia, Pistia, Ceratopteris and Eichhornia. Similar results are described from the Magela floodplain (Australia), where Hymenachne acutigluma biomass increases after the first rains but decreased following a

large increase in water level, whilst Oryza meridionalis germinates after the first rains and continues to grow as the plain fills with water.
Changes attributed to rain are even stronger in tropical temporary habitats, common in arid and semi-arid regions. Investigations of the resistance and resilience of Najas marina, a submerged species, to disturbances caused by flash floods in a permanent fluvial pool of a Brazilian semiarid intermittent stream, showed that decreases in macrophyte biomass were positively correlated with flood magnitude, varying from 25 to 53% when discharges were lower than or equal to 0.5 m3sec-1 and between 70 and 100% when discharges were higher than 1.0 m3sec-1. Macrophyte resilience was greater after floods of low magnitude. After floods of 0.5 m3sec-1, three weeks were necessary to re-establish 88 percent of biomass lost, and after a flood of 1.4 m3sec-1, six months were needed to initiate N. marina regrowth.
In addition to water levels and rain, light intensities, temperature and concentration of dissolved inorganic carbon (DIC) are important variables that control the primary production and population attributes of submerged aquatic macrophytes. Evidence about the importance of these variables for primary production of three submerged species (Utricularia foliosa, Egeria densa and Cabomba furcata) was found in Brazilian coastal plain rivers where the low values of PAR, temperature and DIC in winter were limiting to primary production of U. foliosa, and lower values of PAR, in winter, appeared to limit the production of E. densa. On the other hand the higher values of PAR, and lower values of DIC in winter and spring limited the production of C. furcata. The most productive species in rivers of the target area was U. foliosa, a submerged non-rooted species. Its carnivorous habit is an important additional source of nutrients for this species and probably for this reason the gross primary production is not limited by the low total nitrogen and total phosphorus concentration in water.
Despite being less productive than other life forms, submerged plants may also reach high growth rates in tropical waters, especially when underwater light radiation is high. Measurements taken in situ indicate that the Neotropical submerged species Egeria najas may double its biomass at rates varying from 8.5 to 31.5 days. However, growth rates of submerged plant populations are much more affected by underwater radiation, rather than nutrients, which affect much more the free-floating species.
Indeed, fast growth rates fueled by nutrient inputs and high temperatures are usually found for populations of emergent grasses and free-floating species. Extremely high production has been found for populations of C4 grasses (especially Echinochloa polystachya) in the Amazonian floodplain, where this species may reach c. 9 kg DM m-2
year-1. These results were recorded with two different methods, namely biomass changes and CO2 flux measurements, and they are comparable with productivity of fertilized maize fields in warm temperate conditions in Canada and the United States.
The fast growth of free-floating plants makes species belonging to this life form among the most troublesome macrophyte especially in the tropics (see “Macrophytes as Weeds” in this chapter). Eichhornia crassipes is probably the floating species with the highest competitive ability and it is has been shown that it displaces other free-floating plants when occurring together. This conclusion comes from both laboratory experiment and field evidence. Experiments testing the ecological interactions of E. crassipes and Pistia stratiotes, two free-floating macrophytes, showed an aggressive competitive behavior of the former, which even improved its biomass, inhibiting the growth and establishment of the latter. Similar results were recorded at the Itaipu Reservoir (Brazil/Paraguay), where occasional explosive population growths of free-floating species are recorded. In of these

events Salvinia herzogii (together with P. stratiotes and E. crassipes) covered large areas in c. 3 weeks, but three months later, E. crassipes dominated the stands, dislodging the other two species. The architecture of E. crassipes, which is taller and better able to capture light, probably explains its dominance upon other species that develop more horizontally, such as Salvinia spp.
The fast population growth of free floating species has been recorded in several tropical waters: doubling times for biomass from 3 to 5 days have been recorded for E. crassipes and Salvinia spp in Africa and South America, under near-optimal conditions, although under in situ favorable conditions these rates vary from 8 to 15 days. Especially Salvinia spp, which grows very fast horizontally, may double its colonized areas in c. 2 - 3 days under favorable conditions, transforming it into a nuisance very quickly. Together with high temperatures and stable water table, explosive growths usually occur with inputs of nutrients, especially phosphorus. However, the initial plant density is also an important determinant of growth rates and time to colonize specific habitats. Simulations of the effects of nutrients and initial plant densities upon floating plant growth under tropical conditions showed that under similar initial populations but in waters with 5, 15 and 35 μg P/L, the growth rates were of 0.4%, 1.0 and 1.6% day-1, respectively. When the simulation considered a constant phosphorus concentration (35 μg P/L), and an ecosystem with 10km2 in area, increasing 10 times the initial population size, reducing the time for total ecosystem cover by 0.43 year (c. 5.2 months). Simulations like this are lacking in tropics but they are extremely important due to widespread problems caused by floating leaved species in several tropical countries.
Finally, it is interesting to note that although free floating species such as E. crassipes, Salvinia spp and P. stratiotes are the very common plants in Neotropical river-floodplain systems, where they are native, they rarely cause problems in these ecosystems. This is because their populations are naturally controlled by water level fluctuations, together with damage caused by native insect and fungus species. The lack of these controlling factors contributes to the success of these species in other continents and/or other ecosystems.
Despite the general fast growth and decomposition of macrophytes in tropics, when compared to temperate ecosystems, caution is necessary to generalize this conclusion especially if we consider high altitudes. In a high altitude reservoir in Colombia, for example, it was suggested that the submerged macrophyte Egeria densa developed extremely high biomasses (the highest biomass for this species recorded ever since was found in this study), as a result of a combination of low decay rates and continuous growth throughout the year. This situation was possible because the reservoir experiences high light income together with low water temperature throughout the year. Thus, it is tempting to suggest that contrary to what was believed by the first ecologists who explored the tropics, namely that tropical ecosystems were physically stable and with high temperatures round the year, in fact this is rarely the case, with the few exceptions perhaps being high altitude aquatic ecosystems where temperatures approach those of temperate regions. 5. Macrophyte communities
5.1. The organization of macrophyte assemblages

Macrophytes rarely occur as monospecific populations in freshwater systems but tend to form recognizable assemblages composed of several species belonging to different life forms (e.g., free-floating, submerged, emergent and floating-leaved). As in temperate lakes, tropical macrophyte assemblages are commonly organized along depth gradients often forming easily-distinguishable zones in relation to water depth. In general emergent species dominate in shallow areas while the submerged ones colonize deeper sites within a littoral transect, with floating-leaved species commonly intermediate between the two primary depth zones.
As relevant to tropical macrophytes as to any other group of organisms is the question: “are communities naturally organized in space and time or they are produced by a random assemblage of species”? One of the ways to answer this question is to analyze plant distribution along gradients (like depth zonation in macrophytes). It is interesting that although macrophytes offer an excellent opportunity to test this central issue in community ecology, relatively few studies using macrophytes have directly addressed this question. In several temperate wetlands, gradient analyses tested against null models have provided evidence that macrophytes are organized in clusters along depth gradients.
In a first attempt to test this question in tropical floodplain lagoons, null models to test patterns of co-occurrence of macrophyte species were used. The results showed that indeed assemblages are generally organized non-randomly. However random assemblages may appear in specific circumstances and it depends on the degree of connectivity between the lagoon and the river during the flood pulse phase. There was a tendency of floods to disorganize macrophyte assemblages during high waters in lagoons that are not connected to the main river. This suggests that macrophyte assemblages in the tropics may be very dynamic. More studies focusing on these aspects in tropical freshwater ecosystems would certainly contribute to the debate about the nature of communities.
5.2. Factors affecting assemblage composition
Local assemblages are composed of species contained in the regional species pool,
although long distance species may be brought by migrant birds, one of the main natural causes for macrophyte dispersion. Nowadays, humans are also important vectors of species introductions, and species brought from distant regions or even other continents by this way may affect local assemblages dramatically if they have strong competitive ability (see section “Macrophytes in populations” and “Macrophytes as weeds” in this chapter). In addition to arrival from other places, local communities are also largely determined by seed banks, especially when recovering from disturbances of flooding or drying. In any instance, the local environment filters out species from the pool creating a community.
Environmental (physical and chemical) factors affect the physiology of individual plants, potentially leading to consequences for whole populations. If the stress and/or disturbance caused by these factors are long or strong enough, communities may also be affected. Thus, in general, the same environmental factors affecting macrophyte populations will also influence communities as a whole. As a consequence, community attributes, such as diversity, dominance and functional traits also change according to morphometry, sediment and water physicochemistry. We discuss below some of the most important abiotic factors affecting community attributes, focusing mainly on examples from tropical aquatic ecosystems.

In temperate wetlands, the relative importance of environmental filters that determine species composition were estimated as follows: hydrology (50%), fertility, salinity and disturbance (15% each) and competition, grazing and burial (<5% each). Although these numbers are just estimates, the high importance given to hydrology highlights that this factor may be particularly important in the tropics, where there are a great number of large and medium-sized rivers with active floodplains experiencing relatively natural water level fluctuations (quite different from temperate regions where most large rivers are regulated). Good examples of such relatively pristine ecosystems in the tropics are the Amazon and its main tributaries, Orinoco and Paraná-Paraguay (Gran Pantanal included) in South America, the Congo basin in Africa, and the Magela floodplain in Northern Australia. Rivers like those, together with their wetlands, usually have well developed, relatively pristine, aquatic plant communities with rich associated faunas.
Hydrology is also considered a primary determinant of plant communities structure in river channels. For example, in the Mary River (Australia), differences in discharge, influencing intensity of disturbance pressure upon the system, largely determine plant community composition in the channel: Myriophyllum verrucosum and M. variifolium dominate at high discharge while Vallisneria nana, Potamogeton crispus and P. perfoliatus occupy intermediate positions in the disturbance gradient.
Changes in hydrology are usually accompanied by several other factors that affect macrophyte communities. Changes in discharge often produce drastic alterations in nutrient concentrations, underwater light, and water flow, in most unregulated systems with annual alterations in water level: all producing increased stress or disturbance impacting the community of plants present. In the Amazon floodplain, for example, macrophyte assemblages respond clearly to water level fluctuations which may reach up to 10 meters within a single year (see “Macrophytes in Populations” in this chapter). In this floodplain, following a seasonal cycle, a terrestrial community composed of several herbaceous species of fast growth are successively substituted by another community of rooted aquatic plants, and then by another comprising free floating species. Similarly, in the Magela wetland (tropical Australia), the structure of 8 communities composed by aquatic herbs was strongly affected by water level fluctuations and some of them change drastically according to it. For example, in the “Eleocharis sedgeland”, Eleocharis spp. dominate during the wet season, but are replaced by annual herbs during the dry season. The importance of water level fluctuation for community structure was also experimentally demonstrated in Australia, where water regime primarily determined which species germinated from the seed bank and survived in the vegetation. In another detailed experiment carried in Australia, it was shown that depth, duration and frequency of flooding all affected plant community development in some way. However, the major factor determining plant community composition was not the total duration, frequency or depth of flooding, but the duration of individual flooding events. Among the attributes measured in this investigation, we can cite species diversity, which showed a gradual decline under longer durations of flooding in a treatment containing intermittent wetland seed banks. Overall, the effects of water level fluctuation upon aquatic plant communities are complex, and full evaluation of the influence of time, duration, frequency and other attributes linked to floods requires an experimental approach, still lacking in the tropics.
Giving the importance of hydrology for macrophyte communities, changes in natural water level regime usually lead to conspicuous changes in plant communities. Such changes are of especial concern in tropics, where large reservoirs usually regulate the flow,

modifying the natural plant habitats existing downstream. We can cite as an example an increase of submerged plant colonization (with dominance by the native Egeria najas and E. densa, and more recently by the exotic Hydrilla verticillata) in response to flow regulation and increase in underwater light in a floodplain of the Upper Paraná River, Brazil. Similarly, following river regulation leading to the presence of water even during dry periods, an emergent plant community dominated by Typha domingensis developed in the in the Old Hadejia River, Nigeria.
Although water level fluctuations are typical of river floodplain systems, this factor is not peculiar to these ecosystems. Lakes and reservoirs are also affected by water level fluctuation, which can strongly influence the composition of macrophyte assemblages. In the Itaipu Reservoir (Brazil/Paraguay), an assemblage composed mainly of submerged species was abruptly changed after a water level drawdown that lasted 3 months. The rooted submerged Egeria najas was the most frequent species before the drop in water level but its frequency was reduced by c. 90%, taking almost 4 years for recovery after water levels rose again. However, in some arms of this reservoir, the recover of water level was immediately followed by fast growth of free-floating species (Eichhornia crassipes, Salvinia herzogii and Pistia stratiotes), probably in response to increase in water nutrients. This phenomenon resembles what happens in tropical reservoirs after first filling: inputs of nutrients, together with high temperatures, often favor the development of a community dominated by free-floating species, which may be considered the first macrophyte communities in the ontogeny of a reservoir. This phenomenon has been recorded in several tropical reservoirs, like Lake Kariba (Zimbabwe/ ZambiaAfrica), where enormous growths of Salvinia molesta developed immediately after the reservoir’s first filling, and in Serra da Mesa, South America, where a community dominated by S. auriculata and P. stratiotes initially established in several arms. On the other hand where nutrient outburst is lower, for example in the case of new reservoirs flooding a desert landscape, free-floating plants are not favored in this way, and development of the aquatic plant community commences with submerged species (e.g. Lake Nasser, Egypyt).
As indicated by these examples, nutrients are also key chemical factors affecting the composition of macrophyte assemblages. Long term studies in Lake Nasser (Egypt) showed conspicuous changes in submerged macrophyte community mediated by physico-chemistry. The main factors mediating changes were conductivity, magnesium, calcium, nitrate and nitrite, and hydrosoil calcium and phosphate, which favored the development of the invasive Myriophyllum spicatum which, in turn, caused alterations in plant communities. In the Neotropics, water and sediment chemistry have also been considered key for macrophyte community structure, both in natural as well as in man-made aquatic ecosystems. In the Upper Paraná River floodplain (Brazil), both sediment chemistry (especially Fe), together with water phosphorus, were important determinants of macrophyte diversity while in the Itaipu Reservoir, water phosphorus proved to be a major driver of macrophyte community, leading to domination by several free-floating species. The examples above, taken from very different aquatic ecosystems from Africa and South America, highlight the importance of trophic status as a determinant of aquatic plant composition in tropical region.
Giving that macrophytes clearly respond to water and sediment chemistry, the use of these plants as bioindicators has been proposed. This is especially valid because macrophytes do not actively move, are easy to see and identify and readily sampled. Although presence-absence data can be used for such purposes, macrophytes are extremely

plastic and thus schemes incorporating macrophyte and community traits (e.g., diversity, total biomass, shoot length, number of lateral branches, total root length, number of reproductive structure etc.) can produce good results. Such an approach was applied in several tropical ecosystems, where morphological traits of macrophyte populations present in rivers provided a potentially useful means of assessing river trophic status.
Underwater light regime is another important factor affecting submerged macrophyte communities, which rely on below surface light for photosynthesis. As largely described for temperate lakes the maximum depth of colonization by submerged macrophytes in tropical waterbodies is also directly related to Secchi disk depth or other measures of underwater light attenuation. The presence of submerged plant assemblages is largely determined by underwater light availability, in turn influenced by both inorganic and/or organic turbidity. Giving the seasonality of rainfall in tropical regions, submerged plant communities can be greatly reduced during periods of increasing water level and/or turbidity.
When turbidity is mainly due to the development of phytoplankton, submerged macrophyte assemblages may disappear as a combination of competition for light and nutrients. Shifts of an aquatic ecosystem toward a “turbid state”, dominated by phytoplankton community, or the recovery to a “clear state” dominated by a submerged macrophyte community, fall within the concept of “alternative stable states”. This phenomenon has been extensively studied in temperate ecosystems but it seems that this concept may also be applied for tropical ecosystems, and relevant evidence has been gained for a variety of Neotropical aquatic ecosystems. The question is how fast the shifts occur in tropical ecosystems and what are the mechanisms involved. An interesting transition between states has been described in arid wetlands in Australia. Lakes of these wetlands become turbid and freshwater after flooding, but they become highly saline and clear after drying, which leads to a shift to dominance by a submerged plant community. Interesting here is that contrary to what happens in temperate lakes, where transition between turbid and clear water may be fairly slow, and is mediated by nutrients, in the Australian arid wetlands this transition is sudden and mediated by increasing salt concentration.
It is important to note that understanding the mechanisms behind aquatic plant community shifts is of considerable practical importance especially in tropical, poor countries, where eutrophication is one of the greatest issues threatening waterbodies. The establishment and sustainable maintenance of a stable clear water state in lakes and reservoirs, dominated by submerged macrophytes represents the presence of “good quality” water, contrary to the turbid state, dominated by phytoplankton. Light, together with nutrients, is also responsible for community changes along reservoir chains. Conspicuous changes in macrophyte community occur along the Tietê River (Brazil), which is heavily regulated, with a cascade of 7 reservoirs. This river drains much of the São Paulo super-conurbation and the first reservoirs, ca. 100 km downstream from that city, are highly eutrophic and turbid. These reservoirs are colonized mainly by macrophyte assemblages dominated by free-floating plants (e.g., Eichhornia crassipes, Salvinia spp and Pistia stratiotes). On the other hand, the final reservoirs in the chain are oligo-mesotrophic, with dominance of submerged assemblages (e.g., Egeria najas, E. densa and Ceratophyllum demersum)(Fig. 3).
Wind disturbance (“exposure”) is another important physical factor affecting plant community attributes, acting via production of waves which can increase environmental disturbance intensity in exposed littoral parts, especially of large water bodies, where fetch (the longest uninterrupted distance in an aquatic ecosystem free for wave formation) is

high. Shallow waters are more affected by physical effects of waves and tend as a result to have substrates of larger particle size (less prone to being washed away by wave action). These coarse sediments are poor in nutrients and organic matter, and hence their presence represents an additional stress for plant communities of shallow waters, though the main effect derived from waves is direct injury upon plants. Thus, waves represent a mixture of stress and disturbance. Effects of winds/waves are usually driven both by “fetch”, direction and duration of high wind speeds, and slope of the shoreline. The greater the fetch, the higher the potential for wave exposure to be a significant factor affecting littoral communities. The importance of fetch for plant communities in tropical regions has been demonstrated in large man-made lakes, where fetch negatively affects the diversity of macrophytes and positively the minimum depth of colonization of submerged assemblages. Waves are not formed only by wind, but also by navigation (especially where water is shallow and powered boats or ships are relatively large). Boat-generated waves have been shown to affect community attributes in tropical areas, as for example in the R. Nile Egypt.
Reservoirs cascade0
50
100
150
200
250
300
350
Phos
phor
us (μ
g/L)
, chl
orop
hyll-
a (μ
g/L
)
Secc
hi d
isk
(m)
phosphorus Secchi chlorophyll-a
Figure 3. Upper: reservoir cascade along the Tietê River Brazil with dominance of free floating species in the first reservoirs and submerged species in the lowest ones. Lower: phosphorus, chlorophyll-a and Secchi disk values recorded in the reservoirs (mean values and standard deviations are shown). In the lower figure, B. Bonita is on the right and Três Irmãos on the left side of axis x.

Although we deal above only with abiotic factors, biotic interactions can be important as additional drivers of tropical macrophyte community structure. For example while water regime played a major role in species germination and initiation of reproduction in Australian wetlands, the subsequent reproductive output of macrophytes was controlled by grazing (simulated experimentally by clipping). Maybe the most heavily studied example of macrophyte herbivory is the grass carp (Ctenopharyngodon idella), introduced to many tropical countries as a biocontrol agent against submerged aquatic weeds. Grass carp feed on several species of macrophytes and can change completely the plant community in aquatic ecosystems where it is introduced. However, there are several other large herbivores that feed on macrophytes (see “Importance of macrophytes for ecosystem structure and functioning” in this chapter). These animals potentially affect macrophyte assemblages by preferentially predating specific species, but their effects are poorly known.
In addition to these native herbivores, buffalo are cause of conspicuous changes in macrophyte communities in tropical waterbodies. The elimination of buffalo caused an increase in plant diversity in the Brazilian Pantanal lagoons and changed communities toward increased abundances of Eleocharis spp in Australian floodplains. In the Amazon floodplain, areas subject to buffalo grazing had a decrease in herbaceous plant species richness. It is of special concern that the expectation that once the buffalo were removed the floodplains would re-establish a natural vegetation has not been met. Instead of Phragmites vallatoria (the species recorded in remote times) re-colonizing areas where buffalo were removed, the perennial grass Hymenachne acutigluma has spread and dominated large areas. Giving the strong effects of these animals on macrophyte communities, as shown by these examples, and their presence in many tropical countries, environmental agencies should have special policies concerning their control.
Animals may also affect the structure of plant communities at smaller scales (centimeters or meters) not only by grazing. Fish and other animal activities may cause disturbances that affect macrophyte community structure at the scale of small patches. It has been demonstrated in temperate lakes that a dynamic patch structure in submerged vegetation occurs as a response to disturbance caused by nesting fish. Although we do not know of any study using this approach in tropical aquatic ecosystems, this is a potential area of inquiry in tropics, giving the great number of animal species (e.g. waterfowl and fish) utilizing littoral areas for nesting or refuge.
5.3. Biodiversity and Endemism Information about global patterns of macrophyte diversity is extremely scarce. In part,
this limitation is due to the definition of “aquatic macrophyte”, since many species are amphibious and colonize transitional zones. In early investigations, comparisons among regions surveyed in neotropics with others in temperate regions suggested that the highest diversity was found at cool temperate latitudes. In a comparison of highland streams in the Ecuadorian Andes with physically and chemically similar lowland streams in Denmark, for example, it was found that, at least for submerged species, the tropical habitats supported lower richness. If true, these results would indicate aquatic macrophytes as the only group of organisms for which the normal latitudinal gradient of diversity (i.e. declining with increasing latitude) was not valid. However, an extensive survey carried using several international data-bases (primarily the Royal Botanical Garden, Kew – UK) has recently changed this picture. According to this

survey, vascular aquatic macrophytes are represented by 33 orders and 88 families, with about 2613 species in c. 412 genera. Contrary to what was shown previously, vascular macrophyte species diversity is highest in the Neotropics (984 species), intermediate in the Orient, Nearctic and Afrotropics (664, 644 and 614 species, respectively), lower in the Palearctic and Australasia (498 and 439 species, respectively), and lower again in the Pacific region and Oceanic islands (108 species), whilst only very few vascular macrophyte species have been found in the Antarctica bioregion (12 species). The greatest diversity in the tropics is in part due to the high number of species belonging to the family Podostemaceae (180), which supports solely aquatic species. Considering that temperate regions have been much more heavily investigated than tropical ones, where taxonomic studies are still scarce, it is expected that this difference will increase with higher sampling and taxonomic efforts in the tropics. A good example of this conclusion comes from a survey carried in Neotropics, where plotting the number of studies against the number of species showed a clear tendency of increasing species richness, which is still far from reaching an asymptote (Figure 4). This strongly suggests that the number of macrophyte species will increase with new surveys in the tropics.
0 1 2 3 4 5 6 7 8 9 10 11 12
Studies
0
100
200
300
400
500
600
700
800
900
Cum
ulat
ive
num
ber o
f spe
cies
Figure 4. Cumulative number of species found in 12 studies made in large spatial scales in Neotropical ecosystems.
Aquatic plant endemism seems also to be higher in tropical than other bioregions of
the world. A recent survey found that 395 aquatic macrophyte species (64% of total present) were endemic to the Afrotropical region whilst 604 species (61%) are endemic to the Neotropics. Lower degrees of endemicism are found in the Nearctic (268 species; 42%), Palaearctic (139 species; 28%), Pacific (8 species; 7.4%) and Antarctic (no endemic macrophyte species). Tropical areas also have high endemism at smaller spatial scales. For example, 114 endemic macrophyte and wetland species, sub-species or varieties (about 20% of the total) were recorded in Southern Africa (South Africa, Lesotho, Swaziland, Namibia, Botswana); and 100 species (again, c. 20% of the total) in a region including South Brazil, Uruguay, Paraguay and North Argentina.
Concerning alpha (local) diversity, macrophytes can also exhibit high richness in small areas in the tropics. Within waterbodies of the Upper Paraná River (Brazil), for example, up to 14 species have been recorded inside a 1m2 quadrat (P. Carvalho, unpublished). In small, shallow streams of the Upper Congo Basin, such as the Musola River (Northern Zambia), surveys during 2006 found macrophyte diversity not as high as

this, but still reaching 6 – 7 species per m2 (typically in mixed communities supporting submerged, floating-leaved, and emergent species: including Potamogeton nodosus, Aponogeton vallisnerioides, Nymphaea caerulea, Phragmites australis, Ludwigia repens, Cyperus alopecuroides and Panicum obtusifolium).
A more complete analysis of diversity should encompass different spatial scales and include also beta (or between habitats) diversity, since the gamma diversity (diversity within a large region) is a function of both “between” and “within” habitat diversity. There are few data concerning beta diversity in tropical waterbodies. In the Itaipu Reservoir (Brazil), macrophyte beta diversity was shown to be linearly and positively correlated with habitat heterogeneity, which is in accordance with the general ecological principle that the more different habitats exist along a gradient, the more species will colonize that gradient. A tentative attempt at integrating diversity at different spatial scales was made using studies carried out in a floodplain in Brazil (Fig. 4). More studies using this approach would be useful to compare different tropical and temperate regions in terms of macrophyte diversity, and help to prioritize areas of special interest for biodiversity conservation. This is especially valid for macrophytes, given the important role that these plants have in structuring the waterscape and providing habitat for other biota.
Baia River40 species
n = 8 lagoonsturnover (Beta 1) = 0.10
Canal do Meio Lagoon13 species
turnover (Beta 1) = 0.14
Garças Lagoon18 species
turnover (Beta 1) = 0.21
Clara Lagoon22 species
turnover (Beta 1) = 0.30
Paraná River34 species
n = 6 lagoonsturnover (Beta 1) = 0.23
Ivinheima River16 species
n = 6 lagoonsturnover (Beta 1) = 0.12
Upper Paraná River Floodplain62 species
Paraná BasinSpecies richness undetermined
Turnover among rivers
CA 1
CA
2
BaiaIvinheimaParaná
Figure 4. Studies of macrophyte alpha, beta and gamma diversity carried out in the Upper Paraná River floodplain at different spatial scales. Upper level indicates gamma diversity for the Upper Paraná floodplain; intermediate level shows gamma diversity for lagoons of each river independently and beta diversity among lagoons of each river; lower level shows alpha diversity of three lagoons connected to the Paraná river and beta diversity among stands of each of these lagoons. Coordinate Analysis (right hand side) provides evidence that beta diversity is higher among lagoons connected to the Paraná River.
6. Macrophytes as weeds
Several species of macrophytes have a variety of characteristics that make them
potential aquatic weeds all over the world. These nuisance species usually reproduce vegetatively, grow very fast (see “Macrophytes in Populations” in this chapter), and disperse easily by seeds or vegetative propagules, transported by water, birds or, more recently, by boats, or accidental dispersal via the aquarium trade. Small fragments of

submerged macrophytes may travel long distances and establish new populations. In a study involving the Neotropical submerged plant Egeria najas, fast sprouting was found in fragments and even after 3 days in complete dryness, fragments could sprout and emit roots after re-wetting. Indeed, successful invasions by alien submerged species may initiate from fragments or vegetative propagules and may be entirely based on asexual propagation. Some, if not all of the above-mentioned features, are found in free-floating (e.g. Eichhornia crassipes and Salvinia spp) and submerged (e.g. Hydrilla verticillata and Egeria densa) species. The transport of these and other tropical species between basins and mainly between continents make them major nuisance species all over the world. For example, although Hydrilla verticillata is probably native to some countries in Africa, this species was first recorded in South Africa only in 2006 and RAPD analysis showed that invasion originated from Indonesian and Malaysian populations.
Excessive growth of macrophytes may impact aquatic ecosystems in a variety of ways, threatening ecosystem functioning, biodiversity and ecosystem multiple uses by man. Free-floating species like Eichhornia crassipes or Salvinia spp may completely cover the water surface in short time periods in tropical ecosystems, reducing light below the water surface and gas exchange between water and atmosphere. These mechanisms may eliminate submerged macrophytes, and change plankton and fish communities. Submerged species, like Hydrilla verticillata and Egeria densa may also grow and dominate less turbid waters, leading other submerged species to become locally extinct.
Impacts are usually much stronger when plants are introduced to regions where they are not native, since the biological and physical controlling factors may not be present in the new environment. Although mechanisms behind macrophyte invasions in tropical ecosystems are poorly known, the literature about plant invasion records and their impacts upon freshwater ecosystems is abundant for this region. We will explore some of these examples below.
Natural habitats have been severely threatened in all tropical areas. Introduced species may affect other native ones and change the assemblage structure. Examples in South America are the Gran Pantanal (one of the largest wetlands in the world), and the Paraná River floodplain in Brazil. In the first ecosystem the Australian grass Panicum repens and the African Brachiaria subquadripara are invading several areas, especially those suffering great disturbances by human interference. These two species are dominant over native macrophytes and their expansion is a great concern for the extremely rich aquatic and terrestrial biodiversity of this ecosystem. More recently (c. 2005), the submerged Hydrilla verticillata was recorded in South American waters, specifically in the Tietê River (Brazil). A few months later, this plant was found in natural areas of the Paraná floodplain, downstream from Tietê and in less than two years this became the most frequent submerged species, developing high biomasses (up to 500 g DM m-2) in the Paraná River channel and its floodplain backwaters. Similarly to the Pantanal, the expansion of this submerged species in this floodplain is of great concern, given its extreme competitiveness and potential to dislodge other native species. Other examples come from tropical Australian floodplains, which have also changed in recent decades due, in part, to invasions of species such as Salvinia molesta and Urochloa mutica, and Hymenachne amplexicaulis (this one reducing plant diversity and abundance of several macroinvertebrate orders); and from Lake Nasser (Egypt), where Myriophyllum spicatum invasion changed submerged plant assemblages.

One of the biggest problems affecting a natural lake in Africa in recent years was the mass invasion of Lake Victoria (Uganda, Kenya, Tanzania) and its upstream catchment of the Kagera River (Rwanda) by Eichhornia crassipes during the 1990s. This caused major problems for fisheries and water transport on the lake, and severely threatened hydro-electricity generation in Uganda. The problem was brought under control by a major biological control operation, coordinated by the Lake Victoria Environmental Management Program, in all three riparian countries. An inspection report in 2003, for the World Bank, which funded the program concluded that the operation : “...has in general reduced water hyacinth coverage by 80 – 90% in most of the formerly-infested areas. This outcome is a biologically-self sustaining one, based primarily on the successful introduction of two host-specific natural enemies of water hyacinth (the weevils Neochetina eichhorniae and Neochetina bruchi), by means of a rearing and release programm with major community involvement. However, there remains a residual problem of waterhyacinth hotspot areas, both external and internal to the Lake, which requires to be addressed. Most of these hotspots (which occupy relatively small areas of the Lake) are associated with nutrient-polluted areas of the lake, or inflowing rivers”.
Together with changes in macrophyte assemblages, invasions also affect other aquatic communities that depend on macrophytes for shelter or feeding. However, not always is it possible to show effects of invasive species upon other aquatic communities and contradictory results after invasion are also been recorded. In a tropical floodplain in Australia, the invasion of para grass (Urochloa mutica) had no significant detrimental effects on aquatic macroinvertebrates living among or beneath vegetation and this finding was attributed to similarity in growth form and structure of this species with other native ones. An investigation about water hyacinth Eichhornia crassipes in a Zimbabwe impoundment showed that fish density and diversity were higher in areas colonized by these plants but diversity of zooplankton was higher in unvegetated areas. However, most studies assessing the impact of invasions usually lack data from before invasions and/or are time limited, which may impair definite conclusions about the role of invasive species in tropical ecosystems.
Together with these more “ecological” impacts, excessive growth may impair water uses by man. Fishing, navigation, tourism and, more recently, energy production, are activities severely impacted by excessive growth of submerged and free-floating species in the tropics. In addition, water plant habitats may be the favorite habitat for several disease vectors (e.g. malaria) bringing an additional concern in tropical countries.
It is interesting to note that in contrast to most tropical and temperate areas, where it is non-native species that typically become nuisance aquatic weeds, in South American reservoirs native plants are the most troublesome species. In the Jupiá Reservoir (Brazil), the most troublesome species are the submerged Egeria densa, E. najas and Ceratophyllum demersum, although the emergent Typha domingensis and the free floating Eichhornia crassipes and Pistia stratiotes also cause problems. Massive quantities of these plants impair energy production especially during the rainy season: from 1990 to 1999, 1016 protection screens were damaged and had to be changed, at the Jupiá Dam, and from 1994 to 2001, 54,044 m3 of plants were removed from these screens. Energy production has to be stopped during screen exchanges and even before these plants clogging screens reduce water entering turbines, which also contributes to reduced electricity generation. Together with costs associated with plant removal, this means that millions of dollars are lost by the electricity company. In Ecuador, several reservoirs are almost completely covered by

Eichhornia crassipes, which may reach up to 30 kg FW m-2. Differently from the Brazilian reservoirs, navigation with small boats by local people, who use the reservoirs as main transport routes, is the most important activity affected in Ecuador. Still in Africa, concern about blockage of intake of turbines has been arisen in Koka Reservoir (Ethiopia). 7. Potential use of water macrophytes:
In spite of its limited economic value in the modern world, the high number of species and biomass distribution of aquatic macrophytes in tropical inland aquatic ecosystems have motivated their use in a vast array of commercial, technological and cultural activities. Such activities have important ramifications in many formal and informal economic sectors of tropical developing countries. Aquatic macrophytes are used in the control of eutrophication and environmental pollution, provisioning of food, soil fertilization, pharmaceutical products and aesthetical and spiritual services such as ornamentation and cultural manifestations. Their benefits to man also resides in wild-life conservation practices, where they provide food for animals, protections for spawning fish and other species (see “Importance of macrophytes for ecosystem structure and functioning” in this chapter). In the following we attempt to highlight the many possibilities and the success of the use of aquatic macrophytes in the tropical region.
7.1. Cultural and economic use The use of aquatic macrophytes by humans is widespread in many parts of the world.
Their aesthetic and material importance to the peoples of the orient, the Near East and the early European civilizations is revealed in Sanskrit, Chinese, Greek and Roman literature and in the appearance of several species as ornamentation in ancient architecture, painting, and metalwork. However, in the tropical region Indians were certainly the first who utilized aquatic plants in their subsistence activities. The high number of different life forms of aquatic macrophytes found in tropical inland waters allowed Indians to use them to distinct purposes. Since some aquatic plants match flexibility and resistance they allowed produce useful and durable handicraft products. For example, broad leaved aquatic plants such as Typha spp were routinely used to manufacture a variety of artifacts such as sleeping mats and baskets where Indians could store food and other utensils. Such plants were also used to construct roofs for the tents and ropes which could be used in a variety of ways. The petiole of slender plants such as Nymphaea spp were used to produce wears and adorns which are used both routinely and in folkloric and cultural manifestations.
Most of the uses of aquatic macrophytes discovered by Indians perpetuated over the time to modern civilizations and nowadays configure an important contribution to the economy of tropical developing countries. For example, aquatic plants-made artifacts such as bracelets, wears and sculptures are nowadays commercialized in the internal but mainly in the external market. Live or died macrophytes are also used as decorative and aesthetical products, such as macrophyte flowers arranges. Macrophytes handicrafts have been increasingly exported for developed countries and are also an important cultural item commercialized to tourists who visited tropical countries. Such commercial activities are important both because they constitute an important economic subside but also because they disseminate the cultural richness of tropical developing countries worldwide.

Some macrophyte species also have been used with industrial purposes. In Brazil many species of Typha spp are used in the production of cellulose, which subsidize the paper industry and many other activities.
7.2. Water Gardering The cultivation of decorative aquatic plants may have arisen as an incidental feature
of the ancient arts of pisciculture and landscape horticulture, activities that can be traced back to at least 2500 B. C. Since formal lotus pools became an essential feature of the gardens of Buddhist temples, an emphasis was placed on the aesthetic, contrasting the utilitarian, value of aquatic macrophytes. Water has always been the image of garden designs, once water provides the irrigation and due to the pleasure of the calm magnificence of lakes and pools, enhancing the visual impact of building and trees and creating a peaceful environment.
Despite all life forms can be commonly observed in water gardens, more decorative and imposing types received a greater emphasis. Notably emergent plants are those from the genus Typha, Cyperus and numerous aroids; floating-leaved species such as Nymphaea spp, Nymphoides spp; and free-loating plants like Eichhornia spp, Pistia stratiotes and Salvinia spp are also extensively used. Tropical macrophytes are used to decorate water gardens and fish aquariums both in tropics and temperate countries. Due to this practice, the increasing exportation of macrophytes collected in the tropics to European countries, redirected the attention of temperate botanists to poorly understood macrophyte genera, and incidentally helped to reveal several previously undescribed species.
7.3. Medical Use One of best-known species, accorded a therapeutic value by several races, is Acorus
calamus, the sweet flag, of which the rhizome has been used medicinally since at least the time of Hippocrates (c. 460-377 B.C). Since then, many other references to the medicinal use of macrophytes can be found in ancient literature. The medicinal application of aquatic plants is still common in many places where native customs persists. In the tropics, Indians also used aquatic plants as medicine for a large variety of illnesses. For example Pistia stratiotes was used for Indians as a treatment for skin hurts and infections and until nowadays have been used in the central region of Brazil in the treatment of urinary tract infections. The rhizome of the species Acorus calamus was used to combat eyes illness and toothache.
The use of plants in popular medicine is very common in tropical developing countries at least for more isolated populations living in rural areas where the heritage of the knowledge of popular medicine has been maintained and the access to ordinary medicines are more difficult. Consequently, the use of aquatic plants for this purpose is also evident diversified. Although still scarce, there is a growing body of studies in tropical countries which have attempted to improve the potential use of aquatic macrophytes in human medicine. Aquatic macrophytes are particularly interesting for this purpose because they harbor a vast chemical arsenal against herbivory, such as phenol and anti-oxidants which are also chemical substances of interest to human medicine. The study of such phytotherapycs has shown they may have important antimicrobial properties. Such studies have started to be carried out in tropical countries and are a promising field in tropical

medicine. However, the lack of communication among limnologists, botanists, pharmaceutics and medicine scientists are responsible for the slow advances in the use of macrophytes in human medicine are important obstacles to be overcame in tropical regions.
7.4. Source of food Aquatic macrophytes present organs which due to their accumulated food reserves are
of potential nutritional value to man: among then seeds, fruits and swollen vegetative perennating organs are the most important. A variety of fruits and seeds are rich in oil, starch or protein and can be eaten raw, or dried and ground to flour which can be baked with water or milk to give a kind of bread or cake. Numerous rhizomes and tubers are similarly rich in carbohydrates, especially starch, sugar and mucilage, and are wholly edible when raw or cooked. The foliage of many macrophytes provides acceptable salad ingredients or cooked vegetable dishes. In Amazon, Indians used to utilize water hyacinth ashes as salt and in the Northeast region of Brazil, the leaves of the macrophyte Typha domingensis are often used in salads due to its high nutritional value and good palatability. The rhizome of many other macrophytes is also used in the production of cookies, cakes and other products. In Brazil, one of the most used macrophyte with nutritional value to man is the watercress (Nasturtium sp), and it is often used in fresh or cooked dishes.
Due to its high nutritional value, aquatic macrophytes are used as food for animals as well. Leaves of Typha domingensis are often used to feed the cattle in periods of drought in many Brazilian States and thus, there is a great potential in use aquatic plants to substitute ordinary animal feed. In pisciculture aquatic macrophytes can be used as fertilizers to the water, enhancing fish production as ultimate effect, or as an alternative source of protein. In this context, there is a general agreement that the use of aquatic macrophytes as food source in aquiculture activities must be intensified worldwide.
7.5. Eutrophication and Pollution control Although aquatic macrophytes have been used for a variety of ways their use in the
eutrophication and pollution control are among the most disseminated in tropical countries. The high growth rates of aquatic macrophytes have long been viewed as a problem in the management of impacted aquatic inland ecosystems, since the accumulation of plant biomass can diminish the multitude of the aquatic ecosystem services (see “Macrophytes as weeds” in this chapter). It has been a critical environmental problem in tropical developing countries where the growth of aquatic macrophytes is high during the whole year and sewage treatment is still deficient with the most of sewage entering tropical inland waters without treatment. However, the traits that made tropical aquatic macrophytes an environmental problem in eutrophic systems are the same which made them useful tools in the management of eutrophication and pollution. The high growth rates and fast nutrient assimilation allow aquatic macrophytes to be used as environmental filters in the treatment of wastewater. Several studies have emphasized the use of aquatic macrophytes for this purpose and the results have demonstrated that there are significant differences among plant species, plant biotope (e.g., submerged, floating leaves, emergent) and plant diversity. In general floating macrophytes such as Eichornia spp, Salvinia spp and Pistia stratiotes are the most used species in sewage treatment because their photosynthetic activity and thereby their nutrient assimilation are not affected by the water turbidity which is generally higher

in wastewater. Furthermore, the management of floating macrophytes is easier to perform when plants need to be harvested. However, emergent plants such as Typha spp. and Juncus spp are also efficient in nutrient removal, but their affects are indirect, since the most of nutrient is assimilated by periphytic algae and bacteria which grow attached to plant leaves. In addition, emergent plants are more efficient in adsorb heavy metals due the fast adsorption of metals to organic matter and their post precipitation in the sediment in the particulate form. Another important aspect is the ability of aquatic macrophytes to reduce the number of thermo-tolerant bacteria, ubiquitous in domestic sewage and potentially pathogen. In a Brazilian coastal lagoon (Rio de Janeiro State), aquatic macrophytes can reduce in more than 99% the number of thermo-tolerant bacteria present in the domestic sewage. Therefore, there is a great potential in use aquatic macrophytes to diminish the concentration of nutrients, heavy metals and pathogens bacteria in the sewage, reducing substantially the budget to construct ordinary wastewater treatment stations and the ecological and sanitary damage of sewage discharge in aquatic ecosystems.
8. Bibliography
Ali, M.M., Murphy, K.J., and Abernethy, V.J. (1999). Macrophyte functional variables v. species assemblages as predictors of trophic status in flowing waters. Hydrobiologia 415, 131-138. [This paper brings an interesting example of how to use macrophytes as bioindicators in tropical ecosystems].
Ali, M. M., Mageed, A. A., and Heikal, M. (2007). Importance of aquatic macrophyte for invertebrate diversity in large subtropical reservoir. Limnologica 37, 155-169. [In this paper the authors show the effects of environmental factors on macrophytes and their importance of aquatic biota].
Araújo-Lima, C. A. R. M., Forsberg, B. R., Victoria, R., and Martinelli, L. A. (1986). Energy sources for detritivorous fishes in the Amazon. Science 234, 1256–1258. [This is one of the first investigations showing that detritivorous fish in Amazon rely on phytoplankton, and not macrophytes, to obtain their energy].
Cook, C. D. K. (1996). Aquatic plant book. SPB Academic Publishing, The Hague, The Netherlands. [This is the most classic book about macrophyte diversity and systematic].
Cook, C. D. K. (2004). Aquatic and wetland plants of Southern Africa. Backhuys Publ., Leiden. 281 pp. [This is an important book about diversity of aquatic plants in Africa].
Chambers, P. A., Lacoul, P., Murphy, K. J., and Thomaz, S. M. Global diversity of aquatic macrophytes in fresh-waters. Hydrobiologia, in press. [This is the most recent survey about diversity of macrophytes, and it also includes aspects of evolution, adaptations and use of these plants by man].
Duarte, C. M., Planas, D., and Peñuelas, J. (1994). Macrophytes, taking control of an ancestral home. In: Margalef, R. (ed.). Limnology Now: A Paradigm of Planetary Problems. Elsevier. pp. 59-79. [In this chapter, the authors discuss the main adaptations of submerged plants, as well as their role in nutrient cycling and food webs].
Esteves, F. A. (1998). Fundamentos de Limnologia. Interciência/Finep, Rio de Janeiro. 575p. [This book about tropical limnology includes a chapter synthesizing aspects of macrophyte ecology in tropics].

Friis, E. M., K. R. Pedersen & P. R. Crane, 2001. Fossil evidence of water lilies (Nymphaeales) in the Early Cretaceous. Nature 410: 357-360. [Evidences of the early aquatic plants are discussed in this paper].
Finlayson, C. M. (2005). Plant ecology of Australia´s tropical floodplain wetlands: A review. Annals of Botany 96, 541-555. [This is an excellent review about ecology of macrophytes in tropical wetlands in Australia].
Junk, W. J., and Piedade, M. T. F. (1993). Biomass and primary-production of herbaceous plant communities in the amazon floodplain. Hydrobiologia 263, 155-162. [The primary production and succession of several species of plants along water level fluctuations are shown in this study carried in Amazon].
Keddy, P. A. (2000). Wetland ecology: Principles and conservation. Cambridge University Press, Cambridge. 614. [This is a classical book containing all aspects of ecology of macrophytes in wetlands].
Madsen, T. V., and Sand-Jensen, K. (1991). Photosynthetic carbon assimilation in aquatic macrophytes. Aquatic Botany 41, 5-40. [This is an excellent review about plant strategies to use inorganic carbon from the aquatic medium].
Morison, J. I. L., Piedade, M. T. F., Müller, E., Long, S. P., Junk, J. W., and Jones, M. B. (2000). Very high productivity of the C4 aquatic grass Echinochloa polystachya in the amazon floodplain comfirmed by net ecosystem CO2 flux measurements. Oecologia 125, 400-411. [In this paper, besides measurements of production, several comparisons with previous works are shown].
Oliveira, A. C. B., Soares, M. G. M., Martinelli, L. A., and Moreira, M. Z. (2006). Carbon sources of fish in an Amazonian floodplain lake. Aquatic Sciences 68, 229-238. [This paper highlights the importance of macrophytes for fish for Amazonian fishes and brings a survey of literature about this topic].
Pieterse, A. H., and Murphy, K. J. (1990). Aquatic weeds: the ecology and management of nuisance aquatic vegetation. Oxford University Press, Oxford. 593p. [This is a classical book discussing the main species of weeds, their ecological strategies, problems caused and methods of control; there are several examples from tropics].
Pott, J. P., and Pott, A. (2000). Plantas aquáticas do Pantanal. Embrapa, Brasília. 404p. [The systematic, biodiversity and ecology of aquatic macrophytes in Pantanal, Brazil, are considered in this book].
Sculthorpe, C. D. (1985-reprint). The biology of aquatic vascular plants. Koeltz Scientific Books, Königstein. 610p. [This is the most classical and complete book about biology and adaptations of aquatic plants].
Thomaz, S. M., and Bini, L. M. (2003). Ecologia e manejo de macrófitas aquáticas. Eduem, Maringá. 341p. [In this book, aspects of ecology and management of tropical macrophytes are presented].
Wetzel, R. G. (2001). Limnology: Lake and river ecosystems. Academic Press, San Diego. 1006p. [This is a classical book about general limnology, including an excellent chapter about ecology of littoral regions].

Appendix 1. Bibliography used but not cited. Agostinho AA, Gomes LC & Julio Júnior HF (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas.
Eduem, Maringá. pp. 260-279 Agostinho AA, Gomes LC & Pelicice FM (2007) Ecologia e Manejo de Recursos Pesqueiros em Reservatórios do Brasil. Eduem,
Maringá Agostinho AA, Thomaz SM, Gomes LC & Baltar SLSMA (2007) Aquatic Ecology (in press) Albright T, Moorhouse T & McNabb T (2002) The abundance and distribution of water hyacinth in Lake Victoria and the Kagera River
basin, 1989 – 2001. USGS/ EROS Data Center/ Clean Lakes Inc., Sioux falls, SD / Martinex, CA, USA. Albright T, Moorhouse T & McNabb T (2002) The abundance and distribution of water hyacinth in Lake Victoria and the Kagera River
basin, 1989 – 2001. USGS/ EROS Data Center/ Clean Lakes Inc., Sioux falls, SD / Martinex, CA, USA. Ali MM, Hamad A, Springuel IV & Murphy KJ (1995) Archiv für Hydrobiologie 133, 107–128 Ali MM, Murphy KJ & Abernethy VJ (1999a) Hydrobiologia 415, 131-138 Ali MM, Murphy KJ & Langendorff J (1999b) Hydrobiologia 415, 93-100 Baldwin DS & Mithcell AM (2000) Regulated Rivers-Research & Management 16, 457-467 Barbieri R, Esteves FA & Reid JW (1984) Verh Internat Verein Limnol 22, 1631-1635 Benedito-Cecílio E & Araújo Lima CARM (2002) Journal of Fish Biology 60, 1603-1607 Bianchini Jr. I. (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas. Eduem, Maringá. pp. 85-126 Bini LM (1996) Arquivos de Biologia e Tecnologia 39, 715-721 Bini LM, Thomaz SM, Murphy KJ & Camargo AFM (1999) Hydrobiologia 415, 147-54 Bini LM, Thomaz, SM, Souza DC (2001) Archiv für Hydrobiologie 151, 511-525 Bortolotto IM & Guarim GN (2005) Acta Botanica Brasilica 19, 331-337 Brendonck L, Mães J, Rommens W, Dekeza N, Nhiwatiwa T, Barson M, Callebaut V, Phiri C, Moreau K, Gratwicke B, Stevens M, Alyn
N, Holsters E, Ollevier F & Marshal B (2003) Archiv für Hydrobiologie 158, 389-405 Camargo AFM & Esteves FA (1988) Acta Limnologica Brasiliensia 1, 273-298 Camargo AFM & Esteves FA (1995) In: Tundisi JG, Bicudo CEM & Matsumura-Tundisi T (Eds.), Limnology in Brazil. ABC/SBL, Rio
de Janeiro. pp. 137-149 Camargo AFM & Esteves FA (1996) Archiv für Hydrobiologie 158, 423-432 Camargo AFM, Pezzato MM, Henry-Silva GG & Assumpção Am (2006) Hydrobiologia 570, 35-39 Carney HJ, Binford MW, Marin RR & Goldman CR (1993) Hydrobiologia 251, 39-47 Carpenter SR & Lodgre DM (1986) Aquatic Botany 22, 21-32 Carpenter SR & Mccreary (1985) Aquatic Botany 22, 21-32 Carrilo Y, Guarin A & Guillot (2006) Aquatic Botany 85, 7-15 Carvalho FT, Veline ED, Frazzatto EF, Angos FM & Peruchi M (2003) Planta Daninha 21, 101-104 Carvalho P, Bini LM, Thomaz SM, de Oliveira LG, Robertson B, Tavechio WLG & Darwisch AJ (2001) Acta Scientiarum 23, 265-273 Carvalho P, Thomaz SM & Bini LM (2005) Brazilian Journal of Biology 65, 767-770 Casanova MT & Brock MA (2000) Plant Ecol 147, 237-250 Cavenaghi AL, Velini ED, Galo MLBT, Carvalho FT, Negrisoli E, Trindade MLB & Simionato JLA (2003) Planta Daninha 21, 43-52
Chambers PA & Kalff J (1985) Canadian Journal of Fisheries and Aquatic Sciences 42, 701-709 Colares IG & Colares EP (2002) Brazilian Archives of Biology and Technology 45, 67-72 Conserva A & Piedade MTF (1998) Verh Internat Verein Limnol 26, 994-995 Cook CDK (1999) Perspectives in Plant Ecology, Evolution and Systematics 2/1, 79–102 Cook, CDK (2004). Aquatic and wetland plants of Southern Africa. Backhuys Publ., Leiden Crosslé K & Brock MA (2002) Aquatic Botany 47, 43-56 Crow GE (2003) Aquatic Botany 44, 229-258 de Filippo R (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas. Eduem, Maringá. pp. 281-297 Delariva RL, Agostinho AA, Nakatani K & Baumgartner G (1994) Revista UNIMAR 16, 41-60 Dibble ED, Killgore KJ & Harrel SL (1996) In Miranda LE & DeVries DR (Eds), Multidimensional approaches to reservoir fisheries
management. American Fisheries Society, Symposium 16, Bethesda, Maryland, pp. 357-372 Dibble ED, Thomaz SM, Padial AA (2006) Journal of Freshwater Ecology 21, 239-247 dos Santos A & Esteves FA (2004) Brazilian Arvhives of Biology and Technology 47, 281-290 Douglas MM & O’Connor RA (2003) Freshwater Biology 48, 962-971 El-Sayed AFM (1992) Fish Management 23, 167-173 Engelhardt KA & Ritchie ME (2001) Nature 411, 487-489 Essa MA (1997) Egyptian Journal of Aquatic Biology of Fisheries 1, 19-34 Esteves, FA (1998) In: Esteves, FA (Ed), Ecologia das Lagoas Costeiras do Parque Nacional da Restinga de Jurubatiba e do Município
de Macaé (RJ). Nupem/UFRJ, Macaé. pp. 63-90 Feild TS & Aren NC (2007) Plant, Cell and Environment 30, 291–309
Feldmann T & Nõges P (2007) Aquatic Botany 87, 15-21 Fernandez OA, Murphy KJ, Cazorla AL, Sabbatini MR, Lazzari MA, Domaniewski JCJ & Irigoyen JH (1998) Hydrobiologia 380, 15-25 Friis EM, Pedersen KR & Crane P (2001) Nature 410, 357-360 Gaspar da Luz KD, Fugi R, Abujanra F, Agostinho AA (2002) Acta Scienturam 24, 427-432 Goes BJM (2002) River Research and Applications 18, 81-95 Grillo MT (1993) MSc thesis, PPGB, UFRPE Guarin NG (1984) Ver. UFMT 4, 45-50 Hahn NS, Fugi R & Andrian IF (2004) In: Thomaz SM, Agostinho AA & Hahn NS (Eds) The upper Paraná River and its floodplain.
Backhuys, Leiden. pp. 247-269 Helbing UW, Esteves FA, Tilzer MM & Stabel HH (1986) Acta Limnologica Brasiliensia 1, 611-637

Henriques RPB, Araujo DSD, Esteves FA & Franco AC (1988) Acta Limnologica Brasiliensia 2, 783-802 Henry-Silva GG & Camargo AFM (2006) Scientia Agricola 63, 433-438 Herrera-Silveira JA, Medina-Gomes I & Colli R (2002) Hydrobiologia 475/476, 91-98 Houston WA & Duivenvoorden LJ (2002) Marine and Freshwater Research 53, 1235-1244 Hutchinson GE (1975) A treatise on limnology. v 3. Limnological botany, Wiley, New York
Irgang BE & Gastal Jr CVS (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas. Eduem, Maringá. pp. 163-169
Jacobsen D & Terneus E (2001) Aquatic Botany 71, 281-295 Junk WJ & Piedade MTF (1993) Hydrobiologia 263, 155-162 Junk WJ, Bayley PB & Sparks RE (1989) Canadian Special Publish Fisheries Aquatic Science 106, 110-127 Lewis WM, Hamilton SK, Rodriguez MA, Saunders JF & Lasi MA (2001) Journal of the North American Benthological Society 20, 241-
254. Lopes CA, Benedito-Cecilio E & Martinelli LA (2007) Journal of Fish Biology 70, 1649-1659 Lopes-Ferreira C (1998) In: Esteves, FA (Ed), Ecologia das Lagoas Costeiras do Parque Nacional da Restinga de Jurubatiba e do
Município de Macaé (RJ). Nupem/UFRJ, Macaé. pp. 375-390 Maberly SC, Madsen TV (2002) Functional Plant Biology 2-3, 393-405 Mackay SJ, Arthington AH, Kennard MJ & Pusey BJ (2003) Aquatic Botany 77, 169-186 Madeira PT, Coetzee JÁ, Center TD, White EE & Tipping PW (2007) Aquatic Botany 87, 176-180 Maltchik L & Pedro F (2001) Biotropica 33, 566-572 Marcondes DAS, Mustafá AL & Tanaka RH (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas.
Eduem, Maringá. pp. 299-317 Martins D, Marchi SR & Costa NV (2003) Planta Daninha 23, 351-358 Martins RP & Borges JC (1999) Biotropica 31, 530-534 McCourt RM, Delwiche CF & Karol KG (2004) Trends in Ecology and Evolution 19, 661-666 Milne JM, Murphy KJ & Thomaz SM (2006) Hydrobiologia 570, 19 - 25 Mitchell DS, Pieterse AH & Murphy KJ (1990) In: Pieterse AH. & Murphy KJ (Eds) Aquatic Weeds. Oxford, OUP. pp 351-354 Motta RL & Uieda VS (2005) Austral Ecology 30, 58-73 Murphy KJ & Barret PRF (1990) In: Pieterse AH & Murphy KJ (Eds) Aquatic weeds. The ecology and management of nuisance aquatic
vegetation. Oxford Science, Oxford Publications. pp. 136-173 Murphy KJ & Pieterse AH (1990) In: Pieterse AH & Murphy KJ (Eds) Aquatic weeds. The ecology and management of nuisance aquatic
vegetation. Oxford Science, Oxford Publications. pp. 222-227 Murphy KJ (1990) In: Pieterse AH & Murphy KJ (Eds) Aquatic weeds. The ecology and management of nuisance aquatic vegetation.
Oxford Science, Oxford Publications. pp. 228-237 Murphy KJ (2003) LVEMP Water Hyacinth Component Regional Stocktaking Report: Regional Executive Summary. Report to the World
Bank. 5 pp. Murphy KJ, Dickinson G, Thomaz SM, Bini LM, Dick K, Greaves K, Kennedy M, Livingstone S, McFerran H, Milne J, Oldroyd J &
Wingfield R (2003) Aquatic Botany 77, 257 - 276 NACA/FAO (2000) Desenvolvimento da Aquacultura para Além de 2000: A Declaração de Banguecoque e Estratégia. Conferência
sobre aquacultura no Terceiro Milênio, 20- 25 NACA, Banguecoque e FAO, Roma Nahlik AM & Mitsch WJ (2006) Ecological Engineering 28, 246-257 Nogueira F & Esteves FA (1990) Acta Limnologica Brasiliensia 3, 617-632 Nogueira F, Esteves FA & Prast AE (1996) Hydrobiologia 328, 199-205 Ochiel G, Mailu A, Gitonga W & Njoka S (2001) Proc. 1st IOBC Water Hyacinth Working Group, 115 – 118 Oliveira NMB, Sampaio EVSB, Pereira SMB & Moura Júnior AM (2005) Planta Daninha 23, 363-369 Pagioro TA & Thomaz SM (1999) Acta Limnologica Brasiliensia 11, 157-171 Pagioro TA & Thomaz SM (1999) Hydrobiologia 411, 45-51 Pease AA, Davis JJ, Edwards MS & Turner TF (2006) Freshwater Biology 51, 475-486 Pedralli G (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas. Eduem, Maringá. pp. 171-188 Pelicice FM, Agostinho AA & Thomaz SM (2005) Acta Oecologica-International Journal of Ecology 27, 9-16 Piedade MTF, Junk WJ & Long SP (1991) Ecology 72, 1456-1463 1991 Pierini SA & Thomaz SM (2007) Brazilian Archives of Biology and Technology (in press) Pierini SA, Sgorlon D & Thomaz SM (2006) Acta Scientiarum 28, 31-34 Poi de Neiff AS & Casco SL (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas. Eduem, Maringá. pp.
127-144 Pompêo MLM & Henry R (1998) Verh. Internat. Verein. Limnol. 26, 1871-1875 Pompêo MLM & Moschini-Carlos V (1996) Limnetica 12, 17-23 Porter JL, Kingsford RT & Brock MA (2007) Plant Ecol 188, 215-234 Pott VJ, Bueno NC, Pereira RAC, Salis SM & Vieira NL (2989) Acta Botanica Brasilica 3, 153-168 Santamaria L (2002) Acta Oecologia-International Journal of Ecology 23, 137-154 Santos JE, Esteves FA, Menezes CF & Barbieri R (1986) Acta Limnologica Brasiliensia 1, 341-362 Santos JE, Gazarini LC, Lacava PM & Maintinguer (1983) Anais do Seminario Regional Ecologia 3, 127-133 Scheffer M (1990) Hydrobiologia 200, 475-486 Springuel I, Ali MM & Murphy KJ (1990) Proceedings European Weed Research Society 8th International Symposium on Aquatic
Weeds, 199 - 201. EWRS, Wageningen, The Netherlands Takeda AM, Souza-Fraco GM, Melo SM & Monkolski A (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas
aquáticas. Eduem, Maringá. pp. 243-260 Talling JF & Lemoalle J (1998) Ecological dynamics of tropical inland waters. Cambridge University Press, Cambridge Taniguchi GM, Bicudo DC & Sennab PAC (2004) Brazilian Archives of Biology and Technology 47, 961-971 Thomaz SM & dos Santos AM (2001) Macrófitos acuáticos en los embalses del Proyecto Trasvases Manabi: evaluación de la
colonización y propuestas de manejo. Universidade Estadual de Maringá, Technical Report. 39p.

Thomaz SM, Bini LM & Pagioro TA (2003) In: Thomaz SM & Bini LM (Eds), Ecologia e manejo de macrófitas aquáticas. Eduem, Maringá. pp. 319-341
Thomaz SM, Bini LM, Pagioro TA, Murphy KJ, dos Santos AM & Souza DC (2004) In: The Upper Paraná River and its Floodplain: Physical aspects, Ecology and Conservation. Thomaz SM, Agostinho AA & Hahn NS (Eds), Backhuys, Leiden. pp. 331 - 352
Thomaz SM, Dibble ED, Evangelista LR, Higuti J & Bini LM (2008) Freshwater Biology (in press) Thomaz SM, Pagioro TA, Bini LM & Murphy KJ (2006) Hydrobiologia 570, 53-59 Tomas WM & Salis SM (2000) Studies on Neotropical Fauna and Evironment 35, 165-172 Vono V & Barbosa FAR (2001) Environmental Biology of Fishes 61, 371-379 Wetzel RG (1995) Freshwater Biology 33, 83-89 Williams AE, Hecky RE & Duthie HC (2007) Aquatic Botany 87, 94-96 Wilson JRU, Ajuonu O, Centere TD, Hill MP, Julien MH, Katagira FF, Neuenschwander P, Njoka SW, Ogwang J, Reeder RH & Van T (2007) Aquatic Botany 87, 90-93
















![Environmental Protection Division...marine aquatic ecosystem, including phytoplankton, zooplankton, benthos, macrophytes and fish. “aquatic life water use” [AW] means the use of](https://static.fdocuments.in/doc/165x107/600a50d3a2bb500f19706d9f/environmental-protection-division-marine-aquatic-ecosystem-including-phytoplankton.jpg)

