
Antitumoral effects of calcitriol in basal cell carcinomas...
31
1 Antitumoral effects of calcitriol in basal cell carcinomas involve inhibition of Hedgehog- signaling and induction of vitamin D receptor-signaling and differentiation Anja Uhmann 1 *, Hannah Niemann 1 , Bérénice Lammering 1 , Cornelia Henkel 1 , Ina Heß 1 , Frauke Nitzki 1 , Anne Fritsch 1 , Nicole Prüfer 1 , Albert Rosenberger 2 , Christian Dullin 3 , Anke Schraepler 4 , Julia Reifenberger 5 , Stefan Schweyer 6 , Torsten Pietsch 7 , Frank Strutz 8 , Walter Schulz-Schaeffer 9 , Heidi Hahn 1 1 Institute of Human Genetics, 2 Department of Genetic Epidemiology, 3 Department of Diagnostic Radiology, 4 Central Animal Facility, 6 Department of Pathology, 8 Department of Nephrology and Rheumatology, 9 Department of Neuropathology, University of Goettingen, Germany; 5 Department of Dermatology, University of Duesseldorf, Germany; 7 Department of Neuropathology, University of Bonn, Germany *Corresponding author: Anja Uhmann, Institute of Human Genetics, University of Goettingen, Heinrich-Düker-Weg 12, 37073 Göttingen, Germany; Phone: +49 551 3914100; Fax: +49 551 396580; email: [email protected] This work was supported by grants of the Deutsche Forschungsgemeinschaft UH 228/2-1 and UH228/2-2 to AU and HA 2197/5-1 FOR942 to HH. The authors state no conflict of interest. Running title: Calcitriol inhibits growth of basal cell carcinoma Key words: Hedgehog/Patched1-signaling, 1α,25(OH) 2 D 3 , calcitriol, antitumor therapy, basal cell carcinoma Word count: 3616; 5 Figures on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422
Transcript of Antitumoral effects of calcitriol in basal cell carcinomas...

1
Antitumoral effects of calcitriol in basal cell carcinomas involve inhibition of Hedgehog-
signaling and induction of vitamin D receptor-signaling and differentiation
Anja Uhmann1*, Hannah Niemann1, Bérénice Lammering1, Cornelia Henkel1, Ina Heß1,
Frauke Nitzki1, Anne Fritsch1, Nicole Prüfer1, Albert Rosenberger2, Christian Dullin3, Anke
Schraepler4, Julia Reifenberger5, Stefan Schweyer6, Torsten Pietsch7, Frank Strutz8, Walter
Schulz-Schaeffer9, Heidi Hahn1
1Institute of Human Genetics, 2Department of Genetic Epidemiology, 3Department of
Diagnostic Radiology, 4Central Animal Facility, 6Department of Pathology, 8Department of
Nephrology and Rheumatology, 9Department of Neuropathology, University of Goettingen,
Germany; 5Department of Dermatology, University of Duesseldorf, Germany; 7Department of
Neuropathology, University of Bonn, Germany
*Corresponding author:
Anja Uhmann, Institute of Human Genetics, University of Goettingen, Heinrich-Düker-Weg
12, 37073 Göttingen, Germany; Phone: +49 551 3914100; Fax: +49 551 396580; email:
This work was supported by grants of the Deutsche Forschungsgemeinschaft UH 228/2-1 and
UH228/2-2 to AU and HA 2197/5-1 FOR942 to HH.
The authors state no conflict of interest.
Running title: Calcitriol inhibits growth of basal cell carcinoma
Key words: Hedgehog/Patched1-signaling, 1α,25(OH)2D3, calcitriol, antitumor therapy, basal
cell carcinoma
Word count: 3616; 5 Figures
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

2
Abstract
Activation of the Hedgehog (Hh)-signaling pathway due to deficiency in the Hh receptor
Patched1 (Ptch) is the pivotal defect leading to formation of basal cell carcinoma (BCC).
Recent reports provided evidence of Ptch-dependent secretion of vitamin D3-related
compound, which functions as an endogenous inhibitor of Hh-signaling by repressing the
activity of the signal transduction partner of Ptch, Smoothened (Smo). This suggests that Ptch
deficient tumor cells are devoid of this substance, which in turn results in activation of Hh-
signaling. Here we show that the application of the physiologically active form of vitamin D3,
calcitriol, inhibits proliferation and growth of BCC of Ptch mutant mice in vitro and in vivo.
This is accompanied by the activation of the vitamin D receptor (Vdr) and induction of BCC
differentiation. In addition, calcitriol inhibits Hh-signaling at the level of Smo in a Vdr-
independent manner. The concomitant antiproliferative effects on BCC growth are stronger
than those of the Hh-specific inhibitor cyclopamine, even though the latter more efficiently
inhibits Hh-signaling. Taken together, we show that exogenous supply of calcitriol controls
the activity of two independent pathways, Hh- and Vdr-signaling, which are relevant to
tumorigenesis and tumor treatment. These data suggest that calcitriol could be a therapeutic
option in the treatment of BCC, the most common tumor in humans.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

3
Introduction
The Hh-signaling pathway regulates cell differentiation, organ patterning, and cell
proliferation (1). In the absence of a ligand the activity of the Hh-pathway is inhibited due to
an interaction between the Hh receptor Ptch and its partner Smo. Binding of Hh to Ptch,
inactivating Ptch mutations, or activating Smo mutations may suspend this inhibition,
resulting in the transcription of target genes including Gli1 (1). Thus aberrant (e.g. mutation-
driven) Hh-signaling results in tumor formation (2). One prominent example is BCC, the most
frequent tumor entity in humans, with active Hh-signaling due to mutations in Ptch.
Today the inhibition of the Hh-pathway is considered as a promising strategy in the treatment
of these tumors. Thus specific Smo inhibitors such as cyclopamine and GDC0449 have been
tested in several small, non-randomized clinical trials (2). GDC0449 was recently shown to
elicit antitumoral effects in 18 out of 33 patients with locally-advanced or metastatic BCC (3).
It remains to be elucidated if these promising results will be confirmed in a prospective,
randomized and controlled study and if Hh targeting alone will be sufficient. In this context, a
recent report describes resistance to GDC0449 due to a therapy-associated Smo mutation (4).
As with other tumors, therapy of those associated with abnormal Hh-signaling may require
targeting of additional signaling pathways.
Vitamin D3 and its derivatives (e.g. EB1089; Paricalcitriol) are known to have antitumoral
effects on different cancer types (5) including squamous cell carcinoma (6, 7) or on
hyperproliferative skin diseases such as psoriasis (8, 9). These effects comprise G0/G1 arrest,
cellular differentiation, induction of apoptosis and modulation of inflammation or of different
signaling pathways in tumor cells, as well as inhibiting tumor angiogenesis (5). Until now, the
antitumoral effects of vitamin D3 are explained by binding of the biologically active form of
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

4
vitamin D3, calcitriol (1α,25-dihydroxy vitamin D3), to the Vdr and the subsequent regulation
of Vdr-bound genes.
Calcitriol is produced from vitamin D3 by two hydroxylation steps in the liver and kidney,
respectively, and to a lesser extent in other organs and in tumor cells (5). Through binding to
Vdr, calcitriol regulates the transcription of Vdr target genes (5). This so-called “genomic”
calcitriol/Vdr-signaling regulates a variety of physiological processes including cellular
differentiation, especially in the skin (10, 11), proliferation, and apoptosis, and can be
monitored by measuring transcription of the calcitriol metabolizing enzyme 24-hydroxylase
(5). In addition, calcitriol elicits rapid, so-called “non-genomic” (i.e. transcription-
independent) effects such as calcium influx (5).
Recent data suggest a crosstalk between vitamin D3 and Hh-signaling. By medium transfer
experiments Bijlsma et al. provided first evidence for a Ptch-dependent secretion of vitamin
D3 compounds. They also showed that vitamin D3 inhibits the Hh-pathway at the level of Smo
(12). This suggests that, in addition to or instead of a direct protein-protein interaction, Ptch
may repress Smo via secretion of a vitamin D3 derivative (12).
The possibility of a Ptch-dependent secretion of a vitamin D3 derivative with Smo-inhibitory
properties opens new perspectives for therapies of tumors that arise due to mutations in Ptch.
One would expect that inactivation of Ptch results in a disrupted secretion of this derivative.
This should result in a potential deprivation of the vitamin D3 derivative, and in lack of Smo-
inhibition. If this hypothesis is true, the concerted action of active Hh- and inactive Vdr-
signaling may be the driving forces leading to enhanced cell proliferation, compromised
differentiation and ultimately to tumor formation. It follows that it should be possible to revert
or at least to impede these processes by administration of the respective vitamin D3 derivative.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

5
Here we investigated the effect of the biologically active vitamin D3 derivative calcitriol on
Vdr- and Hh-signaling, growth, apoptosis and differentiation of Ptch-deficient BCC cells in
vitro and in vivo using the Ptchflox/flox Rosa26CreERT2+/- (Ptchflox/flox ERT2+/-) mouse model
for BCC (13). In addition, we compared its effects with those of cyclopamine and sought to
unravel the molecular mechanisms underlying the calcitriol-mediated effects on Hh-signaling.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

6
Material and methods
Compounds
Calcitriol (Sigma-Aldrich, Germany) and cyclopamine (Toronto Research Chemicals Inc.,
Canada) were dissolved in ethanol (EtOH). Final concentrations for in vitro experiments are
indicated in the respective experiments and correspond to those normally used in cell culture
(5, 14, 15). For in vivo use, calcitriol was diluted individually for each animal in 20 µl
EtOH/1200 µl sterile sunflower oil (Sigma-Aldrich) to obtain a final concentration of 40 or
100 ng/kg in 50 µl.
Animals and treatment of tumor-bearing Ptchflox/flox ERT2+/- mice with calcitriol
Conditional Ptchflox/flox ERT2+/- mice were randomized into 2 groups and BCC were induced in
all animals by intramuscular injection of 100 µg tamoxifen as described (13, 16). Tumors of
this animal model lack the expression of wt Ptch alleles and are therefore deficient in Ptch
(13, 17). Starting points of the daily intraperitoneal (i.p.) treatment with 100 ng/kg calcitriol
or vehicle were day 0 or 60 after BCC induction. Treatment of each cohort was conducted
until day 90 after BCC induction. For RNA isolation and histological examinations skin
samples were collected from tails 24 h after the last calcitriol injection.
Mice were fed with calcium- and phosphate-reduced and vitamin D3-free food (ssniff
Spezialdiaeten, Soest, Germany; E15312-14) (18) one week before and during the injection
period, and for one week thereafter.
All animals were treated and housed in accordance with the German animal protection law.
Measurement of tumor size
BCC size was measured on hematoxylin and eosin (H&E) stained sections using the area
calculation tool of the software CellF (Olympus Soft Imaging Solutions GmbH; Germany).
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

7
Analysis of calcium blood serum values
100 µl of blood were collected from the retroorbital plexus. Serum calcium concentrations
were measured using an O-cresolphthaleine-based assay (cobas, Roche Diagnostics GmbH,
Germany).
Histopathology and Immunohistochemistry
BCC and normal skin (NS) from the tail were embedded in paraffin for histological analyses
or were used for isolation of total RNA. The identity of BCC was confirmed by examination
of H&E stained sections. Paraffin sections were stained using an anti-Ki67- and anti-active
caspase 3-antibody as described (19).
Cell lines and primary cell culture of BCC
The fibroblast cell line Ptchflox/floxERT2+/- was established from dermis of a Ptchflox/floxERT2+/-
mouse. Ptch-/- cells are stable Ptch-deficient cells derived from tamoxifen-treated
Ptchflox/floxERT2+/- fibroblasts. Smo-/- and Vdr-/- fibroblasts have been described in (20) and
(21), respectively. All fibroblast cell lines were maintained in Dulbecco’s Modified Eagle
Medium (DMEM), 10% fetal calf serum (FCS) and 1% penicillin/streptomycine (PS).
The murine BCC cell line ASZ001 was established from UV-induced BCC of Ptchneo12/+ mice
and maintained as described (22). All cell lines have been tested and authenticated before
using by genotyping PCRs on genomic DNA as described (13, 20, 21).
Skin punches were isolated from BCC-bearing Ptchflox/floxERT2+/- mice 30, 40 and 60 days
after tumor initiation by tamoxifen. Punches were maintained for 10 days in culture as
reported (16).
Cell culture experiments
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

8
For gene expression analysis or 5-bromo-2'-deoxyuridine (BrdU) incorporation and caspase
assays, 100,000 or 4,000 cells/well were seeded in 6-well- and 96-well-plates, respectively.
For siRNA-mediated knockdown of Vdr expression Ptchflox/flox ERT2+/- cells were transfected
with Vdr-specific or control siRNA as described below. After 24 h, the cells were washed and
incubated for additional 48 h with medium supplemented with calcitriol, cyclopamine or
ethanol as indicated in the respective experiments.
Cell proliferation was measured after BrdU-pulsing for the last 22 h using a Cell Proliferation
BrdU ELISA (Roche Diagnostics GmbH).
Activity of Caspase 3 and 7 was measured using the Caspase-Glo® 3/7 Assay (Promega) and
a microplate reader (Biotek instruments inc., Bad Friedrichshall, Germany) according the
manufacturer’s instructions.
Shh-N-conditioned medium (Shh-N-CM) or respective control medium (CoM) were obtained
from HEK293-Shh (HEK293-Shh express one of the 3 mammalian Hh proteins Sonic
hedgehog (Shh)) or HEK293 cells, respectively, as described (14).
Knockdown of Vdr expression in Ptch-/- and Ptchflox/floxERT2+/- cells was achieved by using a
Vdr-specific short interfering RNA (siRNA) (5’-CAGGCGGAGCATGAAGCGCAA-3’).
Scrambled siRNA (AllStars negative, Qiagen, Germany) was used as control siRNA. 75 ng
siRNA and 4.5 µl HiPerFect (Qiagen, Germany) were mixed with 100 µl DMEM and
incubated for 10 minutes. Simultaneously, 15,000 cells/well were seeded in DMEM/10%
FCS/2% PS in 24-well-plates. Subsequently, the siRNA/HiPerFect-mix was added to the
cells. After 24 h the cells were incubated with the respective media (as indicated in the
experiments) supplemented with vehicle or 10 nM calcitriol. After additional 48 h the cells
were collected and used for subsequent experiments.
For activation of Hh-signaling pathway in Smo-/- fibroblasts the cells were transfected with a
plasmid expressing hSMO (15, 23) using RotiFect following the manufacturer’s instructions.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

9
After 6 h the cells were incubated with media containing 10 nM calcitriol, 5 µM cyclopamine
or ethanol for 48 h.
Data shown are representative for at least three independent experiments each performed in
triplicate.
Reverse transcription, RT-PCR and quantitative real time-PCR-analyses
Total RNA was extracted using TriReagent (Sigma-Aldrich). For skin samples the RNeasy
fibrous tissue mini kit (Qiagen, Germany) was used. Synthesis of cDNA and primer
combinations for amplification of 18S rRNA, Gli1 and Keratin10 (K10) transcripts used for
quantitative real-time PCR (qRT-PCR) were described previously (16, 19). Primer pairs used
for quantitative real time-PCR-(qRT-PCR) analysis of Gli2, Vdr, Cyp24a1 and
transglutaminase 1 (Tgm1) were 5’-GCAAGGTCAAGACTGAGGCTGA-3’/5’-
GCTGCTCCTGTGTGTCATACTCCTT-3’; 5’-AGAACATGTGCTGCTCATGGC-3’/5’-
TCATCTTGGCGTAGAGCTGGTGGCT-3’; 5’- GTGTGGCAAGCGCACACGCT-3’/5’-
CCGTGACAGCAGCGTACAGT-3’ and 5’-GCAGTGGTGTAAATGCAGCTGG-3’/5’-
ATGAGGAGCTCAAGGGCAATGC-3’, respectively.
Amplification of 18S rRNA was performed as an endogenous control for the normalization of
target gene expression. The amount of target and endogenous reference was determined using
the relative standard curve method. Each sample was measured in triplicates. Graphs
represent the mean value of all measurements.
Glycerol-3-phosphate dehydrogenase (mGapd), wildtype (wt) Ptch transcripts (derived from
the non-recombined Ptchflox locus) and Ptchdel transcripts were detected by RT-PCR as
described in (13, 24).
Protein extraction and Western blot
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

10
Ptch-/- fibroblasts were transfected with Vdr-specific siRNA or control siRNA as described.
After 72 h the cells were harvested. Nuclear fractions of the transfected cells and of Vdr-/-
fibroblasts were isolated using the kit NE-PER Nuclear and Cytoplasmatic Extraction
Reagents (Thermo scientific, Rockford, IL, USA) according to the manufacturer’s
instructions. Protein concentrations were measured by Pierce BCA Protein Assay Kit
(Thermo scientific) and 16.5 µg of each protein sample of the nuclear fraction were analysed
by western blot using a rabbit anti-Vdr antibody (1:100; clone 9A7; Abcam, UK) and a mouse
anti-heat shock 70 kDa protein 8 (HSC-70) (1:10,000; Santa Cruz, Germany) in combination
with horseradish peroxidase (HRP)-conjugated goat anti rat IgG (1:10000; Thermo scientific)
and rabbit anti-mouse IgG (1:5000; GE Healthcare, Germany), respectively. Signals were
detected by using ECL+ reagent (GE Healthcare).
Statistics
Mann-Whitney-U testing was performed to determine the significance of the results.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

11
Results
Calcitriol activates the Vdr-pathway, suppresses Hh-signaling and inhibits proliferation
of cultured Ptch mutant BCC cells
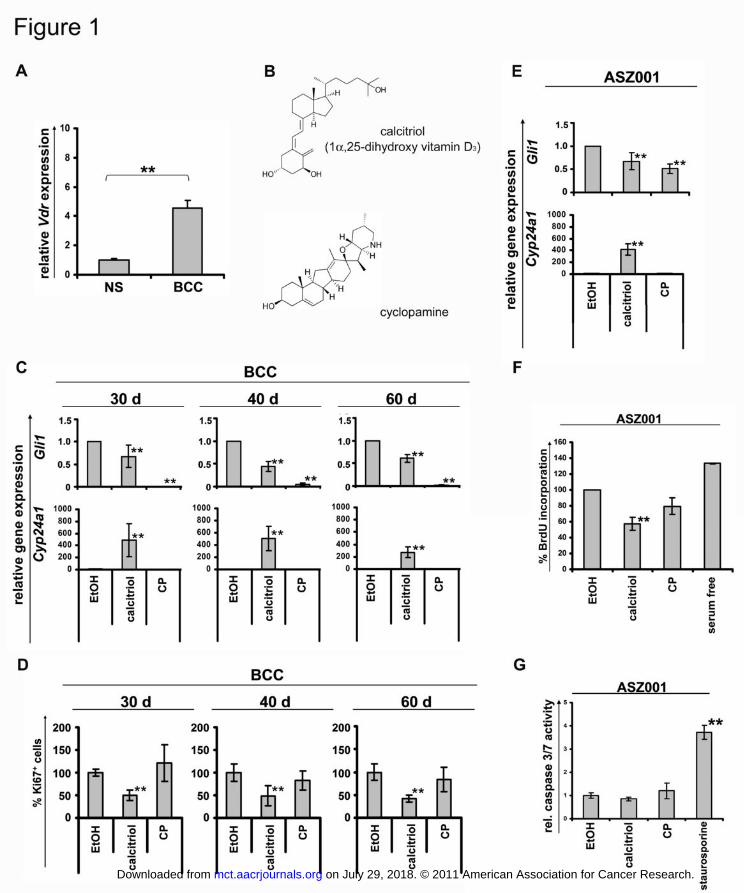
As human BCC (5, 25-27), BCC from Ptchflox/floxERT2+/- mice express elevated Vdr levels
compared to normal murine skin (Fig. 1A).
To test the response of Ptch deficient murine BCC cells to calcitriol in vitro, BCC-bearing
skin punches were collected 30, 40 or 60 days after BCC initiation in Ptchflox/floxERT2+/- mice
(see material and methods). The punches were cultured with 10 nM calcitriol or with 10 µM
cyclopamine for 10 days (the structures of cyclopamine and calcitriol are provided in Fig.
1B). Gli1 expression levels served to monitor Hh-signaling activity. Cyp24a1 (24-
hydroxylase) transcripts were measured to estimate activation of Vdr.
Calcitriol led to a significant induction of Cyp24a1 transcription (Fig. 1C), which was
consistent with the presence of Vdr in BCC (Fig 1A). In addition, calcitriol decreased Gli1
expression, thus indicating an inhibition of the Hh-signaling pathway. As expected,
cyclopamine did not induce Cyp24a1 transcription but repressed Gli1 expression. The
repressive effect was more pronounced than that achieved with calcitriol (Fig. 1C).
Next, we assessed the antiproliferative effects of calcitriol. As revealed by anti-Ki67 antibody
staining, calcitirol inhibited proliferation of tumor cells in BCC-bearing skin punches (Fig.
1D). Most interestingly, the antiproliferative effect of calcitriol was more pronounced than
that achieved with the Hh-specific inhibitor cyclopamine (Fig. 1D).
Similar results were obtained, when the BCC-derived cell line ASZ001 was incubated with 10
nM calcitriol or with 10 µM cyclopamine for 48 h. As shown in figure 1E both substances
efficiently inhibited Hh-signaling. However, only calcitriol significantly inhibited BrdU-
incorporation in these cells (Fig. 1F). Inhibition of proliferation apparently was not
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

12
accompanied by an increase in apoptosis, as calcitriol had no effect on caspase 3/7 activity
(Fig. 1G).
Together, these results show that calcitriol efficiently inhibits the Hh-signaling pathway and
activates Vdr-signaling in BCC. In contrast to cyclopamine only calcitriol mediated
antiproliferative effects in BCC, even though cyclopamine more efficiently inhibits Hh-
signaling.
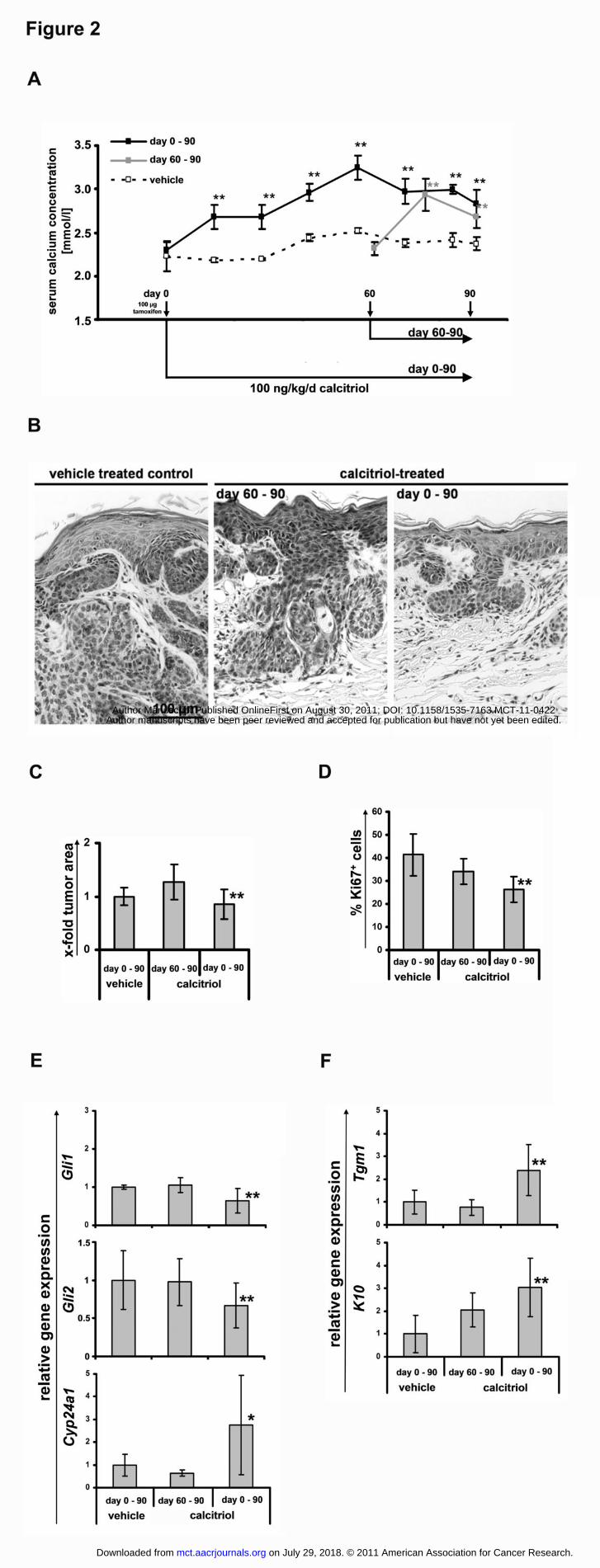
Calcitriol treatment of Ptch mutant mice inhibits the Hh-signaling pathway and growth
of BCC and stimulates BCC differentiation
Next the in vivo antitumoral effects of calcitriol were tested in the Ptchflox/floxERT2+/- mouse
model for BCC. In this model, where all mice develop full-blown BCC 90 days after
activation of ERT2 recombinase by tamoxifen (13), treatment can be commenced at a
specified time after tumor induction and at a defined age of the animals.
Preliminary studies, in which mice were treated daily with 40 ng/kg calcitriol starting at the
day of tamoxifen injection for 90 subsequent days (days 0-90), did not result in an induction
of Vdr-signaling or differentiation (data not shown). Therefore, we increased the daily
calcitriol dose to 100 ng/kg. Calcitriol treatment was started either immediately (day 0) or 60
days after BCC initiation (n60-90d=6; n0-90d=4). Vehicle treated animals (nvehicle=17) served as
controls. The treatment was continued until day 90, when all mice were sacrificed (see Fig.
2A).
Calcitriol therapy led to a significant increase in serum calcium concentrations (Fig. 2A)
without causing weight loss, hypercalcemia-driven kidney damage or signs of
nephrocalcinosis (data not shown). This indicates that the treatment induced calcitriol-specific
systemic effects without causing toxicity. Tumor areas on H&E-stained skin sections in mice
treated on days 0-90, but not on days 60-90, were significantly decreased when compared to
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

13
the vehicle treated group (Fig. 2B, C). Furthermore, calcitriol inhibited tumor cell
proliferation as measured by Ki67+ cells in tumors treated on days 0-90 (Fig. 2D). This was
accompanied by a significant decreased expression of Hh-pathway target genes Gli1 and Gli2
(28-30) in BCC treated for days 0-90 (Fig. 2E). Consistent with the results from our in vitro
studies no significant increase of caspase 3 positive BCC cells were observed (data not
shown). Moreover, calcitriol treatment resulted in a substantial activation of Vdr-signaling as
revealed by an enhanced expression of the immediate Vdr-target gene Cyp24a1 (Fig. 2E).
Finally, a significantly increased expression of the keratinocyte differentiation markers and
Vdr-target genes Tgm1 and K10 were detected (Fig. 2F). Whereas both differentiation
markers can be induced by active Vdr-signaling, K10 expression also depends on Hh-
signaling activity (10, 29, 31-34).
In summary, our data demonstrate that calcitriol significantly inhibits proliferation and
induces cellular differentiation of Ptch-associated BCC in vivo. Moreover, this response is
accompanied by activation of Vdr- and inactivation of the Hh-signaling pathways.
Calcitriol inhibits Hh-signaling downstream of Ptch but upstream of Gli1
Vitamin D3 has been shown to inhibit Hh-signaling at the level of Smo (12, 35, 36). In order
to investigate whether the mechanism of Hh-pathway inhibition mediated by calcitriol is
similar to that of Vitamin D3 we made use of Ptchflox/floxERT2+/- and Smo-/- fibroblasts.
Ptchflox/floxERT2+/- fibroblasts normally express wt Ptch transcripts from the Ptchflox locus.
Upon tamoxifen-induced activation of ERT2 recombinase the exons 8 and 9 of the floxed
Ptch alleles are excised. This results in the expression of an aberrant, non-functional Ptch
transcript (Fig. 3A).
In Ptchflox/floxERT2+/- fibroblasts both, the treatment with Shh-N-CM or the tamoxifen-induced
Ptch mutation result in activation of Hh-signaling as revealed by induction of Gli1 expression
(Fig. 3B). Co-incubation of Shh- or tamoxifen-treated Ptchflox/floxERT2+/- fibroblasts with
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

14
calcitriol significantly inhibited Gli1 expression (Fig. 3B). Similar results were obtained with
the validated Smo inhibitor cyclopamine (Fig. 3B). These data show that similar to
cyclopamine, calcitriol inhibits Hh-signaling downstream of Ptch.
The effects of calcitriol were also tested in Smo-/- cells, which express basal Gli1 levels. Due
to lack of Smo, Hh-signaling cannot be activated in these cells by incubation with Shh-
conditioned medium (37). However, in these cells Hh-pathway activity can be restored by
transfection with a hSMO expression plasmid.
As shown in figure 3C, calcitriol did not influence basal Gli1 expression levels in Smo-/- cells
(Fig. 3C). However, calcitriol efficiently inhibited Hh-signaling after restoration of Hh-
pathway activity upon transfection with hSMO. These results are similar to those obtained
with cyclopamine (Fig. 3C) and show that calcitriol normally inhibits Gli1 expression at the
level of Smo.
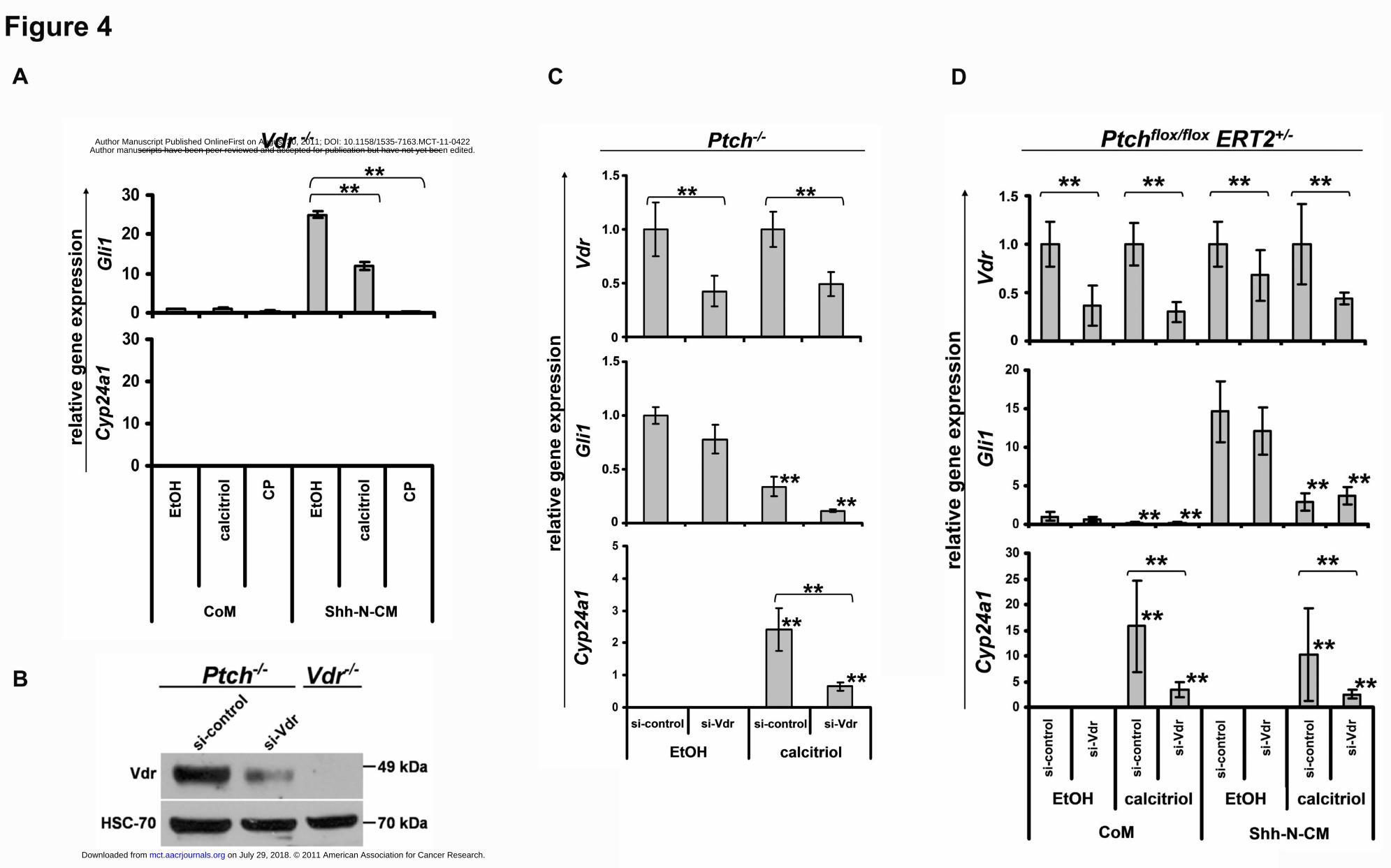
Calcitriol inhibits Hh-signaling in a Vdr-independent manner
Next, we tested whether calcitriol-mediated inhibition of the Hh-signaling pathway is
independent of Vdr-signaling. For this purpose we used Vdr-/- fibroblast (21). In these cells
Vdr-signaling is completely abrogated, since incubation with calcitriol does not result in the
induction of the Vdr-target Cyp24a1 (Fig. 4A). As revealed by Gli1 expression, incubation of
the cells with Shh-N-CM resulted in induction of Hh-pathway activity (Fig. 4A), which was
significantly inhibited by addition of calcitriol or by the control substance cyclopamine (Fig.
4A). Similar results were achieved in Ptch-/- or Shh-N-CM-treated Ptchflox/flox ERT2+/- cells
after siRNA-mediated Vdr knockdown. Efficient downregulation of Vdr expression was
verified by western blot (Fig. 4B), qRT-PCR and by a significantly lower Cyp24a1 expression
after calcitriol treatment in comparison to the controls (Fig. 4C, D). Whereas Vdr knockdown
per se did not significantly attenuate Hh-pathway activity, treatment with calcitriol resulted in
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

15
a downregulation of Gli1 expression (Fig. 4C, D). These data show that calcitriol-mediated
inhibition of the Hh-signaling pathway occurs independently of the Vdr.
Taken together, these data demonstrate that calcitriol inhibits Hh-pathway activity
downstream of Ptch at the level of Smo in a Vdr-independent manner.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

16
Discussion
The active form of vitamin D3, calcitriol, is rapidly gaining importance in oncology due to its
antiproliferative and differentiation-inducing effects combined with its low toxicity (5).
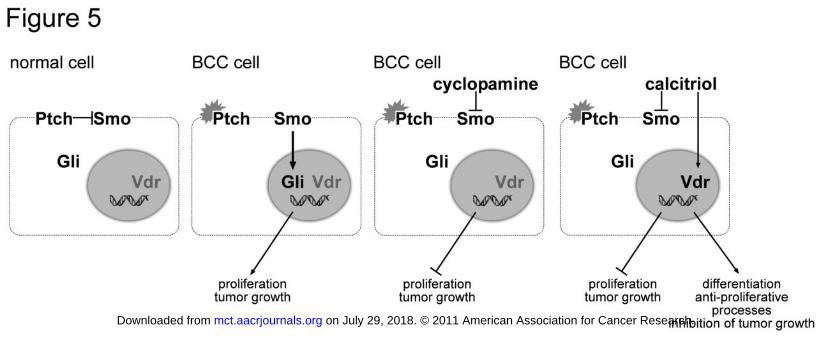
Our work extends the range of calcitriol tumor targets to those induced by an abnormal
activity of the Hh-signaling pathway (summarized in Fig. 5). Specifically, we show that
calcitriol inhibits growth of BCC induced by deletion of the Hh receptor Ptch. This is
accompanied by the expression of relevant cell differentiation markers. Our data suggest that
calcitriol could be a valuable supplement or even alternative to the established treatments of
BCC, the most common tumor in humans, associated with aberrant Hh-pathway activity.
As assessed by reduced Gli1 transcription, calcitriol inhibits canonical Hh-signaling
independently of Vdr-signaling and downstream of Ptch (Fig. 5). An obvious molecular target
of this Vdr-independent effect of calcitriol is Smo, since Smo-deficient cells (unlike those
reconstituted with Smo or Ptch-deficient ones) show no decreased Gli1 transcription in
response to this substance. A similar observation has been made for the inactive form of
calcitriol, vitamin D3 (12). Nevertheless, whether calcitriol directly binds to Smo should be
addressed in future studies.
Besides inhibition of Hh-signaling pathway, calcitriol inhibits proliferation and increases the
expression of skin differentiation marker in BCC. The latter effects are also induced by
calcitriol in skin of patients with hyperproliferative skin diseases such as psoriasis (6-9, 34,
38). Calcitriol treatment specifically results in inhibition of proliferation of psoriatic skin and
induction of differentiation of keratinocytes (9, 39, 40). Similarly to BCC, psoriatic skin also
expresses high levels of the Hh-target gene Gli1 (41). These facts raise the question whether
calcitriol mediates its antiproliferative effects and differentiation stimuli via activation of Vdr-
signaling or rather via inhibition of the Hh-signaling pathway. A clue might come from our
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

17
present study: Our in vitro study shows that calcitriol has a significantly stronger
antiproliferative effect on BCC than the pure Smo inhibitor cyclopamine, even though the
latter substance inhibits Hh-signaling more efficiently (see Fig. 1D and 1F). Together with the
fact that both calcitriol (present study) and cyclopamine (42) inhibit BCC proliferation in
vivo, it is possible that calcitriol exerts its antiproliferative effects via both signaling pathways
(e.g. by inhibition or activation of Hh- or Vdr-signaling, respectively). On the other hand,
differentiation of calcitriol-treated BCC is probably unrelated to inhibition of the canonical
Hh-signaling pathway because cylopamine never has been reported to induce the expression
of keratinocyte differentiation markers in BCC. More likely, calcitriol induces BCC
differentiation via Vdr-signaling, which is supported by the increased expression of the Vdr-
target genes Cyp24a1 and Tgm1 (see Fig. 2F).
Antiproliferative and Hh-signaling inhibitory properties have also been described for the
inactive form of calcitriol, vitamin D3, on murine BCC in vitro and in vivo (35). In contrast to
calcitriol, vitamin D3 did not affect differentiation in BCC, although it was topically applied at
high concentrations. Supposedly the treatment period using vitamin D3 (i.e. 30 days) may
have been too short to induce Vdr-signaling and thus a differentiation response. This
suggestion is based on a comparison with our study, in which a 30 days calcitriol application
also had no significant effects on these processes (see BCC treated for days 60-90, Fig. 2B-E).
Finally, a new model of tumorigenesis driven by Ptch-deficiency may emerge from our study.
According to a recent work, Ptch might function as an efflux pump for vitamin D3-related
compounds with Hh-inhibitory properties (12). A deficiency of this compound due to Ptch
inactivation would pathologically activate Hh-pathway and reduce Vdr-signaling. Vice versa
application of this compound should result in inhibition of Hh-pathway and activation of Vdr-
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

18
signaling (see Fig. 5). Whether this vitamin D3-related compound is calcitriol remains to be
analyzed in the future.
Taken together the application of calcitriol holds promises as an effective anti-cancer drug in
the treatment of BCC. Due to its dual effects on both Vdr- and Hh-signaling it may be
superior to substances which solely target the Hh-signaling pathway. Calcitriol treatment may
also be superior to application of vitamin D3, which has to be metabolized before activating
Vdr-signaling. The benefits of topical application of calcitriol in treatment of BCC have to be
tested in the future.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

19
Acknowledgement
We are grateful to Stefan Wolf and Susan Peter for excellent animal care. We thank J.
Taipale, University of Helsinki, Finland for provision of Smo-/- fibroblasts, E. Epstein, UCSF,
USA for the BCC cell line ASZ001, R. Toftgard, Karolinska Institute, Huddinge, Sweden for
hSMO plasmid, Steven Johnsen, University of Göttingen, Germany for proofreading and for
comments on the manuscript and Leszek Wojnoswki, University of Mainz, Germany for
comments on the manuscript.
Grant support
This work was supported by grants of the Deutsche Forschungsgemeinschaft UH 228/2-1 and
UH228/2-2 to AU and HA 2197/5-1 FOR942 to HH.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

20
References
1. Hooper JE,Scott MP. Communicating with Hedgehogs. Nat Rev Mol Cell Biol 2005; 6: 306-17.
2. Yang L, Xie G, Fan Q,Xie J. Activation of the hedgehog-signaling pathway in human cancer and the clinical implications. Oncogene 2009; 29: 469-81.
3. Von Hoff DD, LoRusso PM, Rudin CM, Reddy JC, Yauch RL, Tibes R, et al. Inhibition of the hedgehog pathway in advanced basal-cell carcinoma. N Engl J Med 2009; 361: 1164-72.
4. Yauch RL, Dijkgraaf GJ, Alicke B, Januario T, Ahn CP, Holcomb T, et al. Smoothened Mutation Confers Resistance to a Hedgehog Pathway Inhibitor in Medulloblastoma. Science 2009; 326: 572-4.
5. Deeb KK, Trump DL,Johnson CS. Vitamin D signalling pathways in cancer: potential for anticancer therapeutics. Nat Rev Cancer 2007; 7: 684-700.
6. Hershberger PA, Modzelewski RA, Shurin ZR, Rueger RM, Trump DL,Johnson CS. 1,25-Dihydroxycholecalciferol (1,25-D3) inhibits the growth of squamous cell carcinoma and down-modulates p21(Waf1/Cip1) in vitro and in vivo. Cancer Res 1999; 59: 2644-9.
7. Majewski S, Skopinska M, Bollag W,Jablonska S. Combination of isotretinoin and calcitriol for precancerous and cancerous skin lesions. Lancet 1994; 344: 1510-1.
8. Stewart DG,Lewis HM. Vitamin D analogues and psoriasis. J Clin Pharm Ther 1996; 21: 143-8.
9. van de Kerkhof PC. An update on vitamin D3 analogues in the treatment of psoriasis. Skin Pharmacol Appl Skin Physiol 1998; 11: 2-10.
10. Bikle DD. Vitamin D and the skin. J Bone Miner Metab 2010; 28: 117-30.
11. Haussler MR, Whitfield GK, Haussler CA, Hsieh JC, Thompson PD, Selznick SH, et al. The nuclear vitamin D receptor: biological and molecular regulatory properties revealed. J Bone Miner Res 1998; 13: 325-49.
12. Bijlsma MF, Spek CA, Zivkovic D, van de Water S, Rezaee F,Peppelenbosch MP. Repression of smoothened by patched-dependent (pro-)vitamin D3 secretion. PLoS Biol 2006; 4: e232.
13. Zibat A, Uhmann A, Nitzki F, Wijgerde M, Frommhold A, Heller T, et al. Time-point and dosage of gene inactivation determine the tumor spectrum in conditional Ptch knockouts. Carcinogenesis 2009; 30: 918-26.
14. Chen JK, Taipale J, Young KE, Maiti T,Beachy PA. Small molecule modulation of Smoothened activity. Proc Natl Acad Sci U S A 2002; 99: 14071-6.
15. Taipale J, Chen JK, Cooper MK, Wang B, Mann RK, Milenkovic L, et al. Effects of oncogenic mutations in Smoothened and Patched can be reversed by cyclopamine. Nature 2000; 406: 1005-9.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

21
16. Nitzki F, Zibat A, König S, Wijgerde M, Rosenberger A, Brembeck FH, et al. Tumor stroma-derived Wnt5a induces differentiation of basal cell carcinoma of Ptch mutant mice via CaMKII. Cancer Res 2010; 70: 2739-48.
17. Calzada-Wack J, Kappler R, Schnitzbauer U, Richter T, Nathrath M, Rosemann M, et al. Unbalanced overexpression of the mutant allele in murine Patched mutants. Carcinogenesis 2002; 23: 727-734.
18. Banach-Petrosky W, Ouyang X, Gao H, Nader K, Ji Y, Suh N, et al. Vitamin D inhibits the formation of prostatic intraepithelial neoplasia in Nkx3.1;Pten mutant mice. Clin Cancer Res 2006; 12: 5895-901.
19. Ecke I, Rosenberger A, Obenauer S, Dullin C, Aberger F, Kimmina S, et al. Cyclopamine treatment of full-blown Hh/Ptch-associated RMS partially inhibits Hh/Ptch signaling, but not tumor growth. Mol Carcinog 2008; 47: 361-72.
20. Ma Y, Erkner A, Gong R, Yao S, Taipale J, Basler K, et al. Hedgehog-mediated patterning of the mammalian embryo requires transporter-like function of dispatched. Cell 2002; 111: 63-75.
21. Sun J, Kong J, Duan Y, Szeto FL, Liao A, Madara JL, et al. Increased NF-kappaB activity in fibroblasts lacking the vitamin D receptor. Am J Physiol Endocrinol Metab 2006; 291: E315-22.
22. Xie J, Aszterbaum M, Zhang X, Bonifas JM, Zachary C, Epstein E, et al. A role of PDGFRalpha in basal cell carcinoma proliferation. Proc Natl Acad Sci U S A 2001; 98: 9255-9.
23. Xie J, Murone M, Luoh SM, Ryan A, Gu Q, Zhang C, et al. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature 1998; 391: 90-2.
24. Uhmann A, Dittmann K, Nitzki F, Dressel R, Koleva M, Frommhold A, et al. The Hedgehog receptor Patched controls lymphoid lineage commitment. Blood 2007; 110: 1814-23.
25. Mitschele T, Diesel B, Friedrich M, Meineke V, Maas RM, Gartner BC, et al. Analysis of the vitamin D system in basal cell carcinomas (BCCs). Lab Invest 2004; 84: 693-702.
26. Kamradt J, Rafi L, Mitschele T, Meineke V, Gartner BC, Wolfgang T, et al. Analysis of the vitamin D system in cutaneous malignancies. Recent Results Cancer Res 2003; 164: 259-69.
27. Reichrath J, Kamradt J, Zhu XH, Kong XF, Tilgen W,Holick MF. Analysis of 1,25-dihydroxyvitamin D(3) receptors (VDR) in basal cell carcinomas. Am J Pathol 1999; 155: 583-9.
28. Ikram MS, Neill GW, Regl G, Eichberger T, Frischauf AM, Aberger F, et al. GLI2 is expressed in normal human epidermis and BCC and induces GLI1 expression by binding to its promoter. J Invest Dermatol 2004; 122: 1503-9.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

22
29. Regl G, Kasper M, Schnidar H, Eichberger T, Neill GW, Ikram MS, et al. The zinc-finger transcription factor GLI2 antagonizes contact inhibition and differentiation of human epidermal cells. Oncogene 2004; 23: 1263-74.
30. Regl G, Neill GW, Eichberger T, Kasper M, Ikram MS, Koller J, et al. Human GLI2 and GLI1 are part of a positive feedback mechanism in Basal Cell Carcinoma. Oncogene 2002; 21: 5529-39.
31. Stark HJ, Breitkreutz D, Limat A, Ryle CM, Roop D, Leigh I, et al. Keratins 1 and 10 or homologues as regular constituents of inner root sheath and cuticle cells in the human hair follicle. Eur J Cell Biol 1990; 52: 359-72.
32. Beck B, Lehen'kyi V, Roudbaraki M, Flourakis M, Charveron M, Bordat P, et al. TRPC channels determine human keratinocyte differentiation: new insight into basal cell carcinoma. Cell Calcium 2008; 43: 492-505.
33. Lee SC, Ikai K, Ando Y,Imamura S. Effects of 1 alpha,25-dihydroxyvitamin D3 on the transglutaminase activity of transformed mouse epidermal cells in culture. J Dermatol 1989; 16: 7-11.
34. Bikle DD. The vitamin D receptor: a tumor suppressor in skin. Discov Med 2011; 11: 7-17.
35. Tang JY, Xiao TZ, Oda Y, Chang KS, Shpall E, Wu A, et al. Vitamin d3 inhibits hedgehog signaling and proliferation in murine Basal cell carcinomas. Cancer Prev Res (Phila) 2011; 4: 744-51.
36. Bruggemann LW, Queiroz KC, Zamani K, van Straaten A, Spek CA,Bijlsma MF. Assessing the efficacy of the hedgehog pathway inhibitor vitamin D3 in a murine xenograft model for pancreatic cancer. Cancer Biol Ther 2010; 10:
37. Varjosalo M, Li SP,Taipale J. Divergence of hedgehog signal transduction mechanism between Drosophila and mammals. Dev Cell 2006; 10: 177-86.
38. Bikle DD. Vitamin D and skin cancer. J Nutr 2004; 134: 3472S-3478S.
39. Bikle DD. 1,25(OH)2D3-modulated calcium induced keratinocyte differentiation. J Investig Dermatol Symp Proc 1996; 1: 22-7.
40. Gottlieb AB. Therapeutic options in the treatment of psoriasis and atopic dermatitis. J Am Acad Dermatol 2005; 53: S3-16.
41. Endo H, Momota Y, Oikawa A,Shinkai H. Psoriatic skin expresses the transcription factor Gli1: possible contribution of decreased neurofibromin expression. Br J Dermatol 2006; 154: 619-23.
42. Tas S,Avci O. Rapid clearance of psoriatic skin lesions induced by topical cyclopamine. A preliminary proof of concept study. Dermatology 2004; 209: 126-31.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

23
Figure legends
Figure 1: Calcitriol inhibits proliferation and the Hh-signaling pathway of BCC-bearing
skin punches and of the BCC cell line ASZ001.
(A) Vdr expression levels of BCC (n=3) of Ptchflox/floxERT2+/- mice 90 days after tumor
induction compared to normal skin (NS; n=3). (B) Chemical structures of calcitriol and
cyclopamine. (C) Gli1 and Cyp21a1 expression levels of cultured BCC-bearing skin punches
isolated from Ptchflox/floxERT2+/- mice 30, 40 or 60 days after BCC induction. (D) Ki67+ BCC
cells of the respective punches. (E) Gli1 and Cyp21a1 expression levels and (F) BrdU-
incorporation in ASZ001 cells. (G) Caspase 3/7 activities of ASZ001 cells. Cells treated with
500 nM staurosporin served as positive controls. The punches and the ASZ001 cells were
incubated with vehicle (ethanol, EtOH), calcitriol or cyclopamine (CP) as indicated in the
text. Gli1 expression and caspase 3/7 activities are shown in relation to the respective vehicle
treated controls. Ki67+ BCC cells and BrdU-incorporation are represented as percentage of
respective vehicle treated controls. Asterisks: p<0.05; error bars: mean+/-SD.
Figure 2: Calcitriol treatment of BCC-bearing mice inhibits tumor growth and the Hh-
signaling pathway, and induces expression of differentiation markers. (A) Treatment
scheme and calcium serum levels of BCC-bearing Ptchflox/floxERT2+/- mice. BCC were induced
in Ptchflox/floxERT2+/- mice by tamoxifen. Treatment with 100 ng/kg/d calcitriol started either
directly (0-90) or 60 (60-90) days after tumor induction and was continued until day 90 when
all mice were sacrificed. Serum calcium levels of the respective BCC-bearing Ptchflox/flox
ERT2+/- mice were measured before (day 0) and during therapy. Serum calcium levels of
vehicle-treated mice served as controls (nvehicle=17; n60-90d=6; n0-90d=4). (B) H&E stainings,
(C) relative tumor areas of H&E stained BCC and (D) percentage of Ki67+ BCC cells of
vehicle- or calcitriol-treated mice. (E) Gli1, Gli2, Cyp24a1 and (F) Tgm1 and K10 expression
levels from vehicle or calcitriol-treated BCC-bearing mice. Values of vehicle-treated controls
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

24
for Gli1, Cyp24a1, Tgm1 and K10 expression were set to 1. One Asterisk: p=0.06; two
Asterisks: p<0.05; error bars: mean+/-SD.
Figure 3: Calcitriol inhibits the Hh-signaling pathway downstream of Ptch but upstream
of Gli1. (A) Ptch ablation in tamoxifen-treated Ptchflox/flox ERT2+/- fibroblasts after incubation
with 10 µM tamoxifen for 48 h. Wt Ptch transcripts (derived from the non-recombined
Ptchflox locus) and Ptchdel transcripts were detected by PCR using the primer pair
mPtc11/mPtc7R as described in (13, 24). cDNA derived from 12.5 days old wild-type mouse
embryos (E12.5 Ptch+/+) was used as a control. (B) and (C) Gli1 and Cyp24a1 expression
levels of (B) Ptchflox/floxERT2+/- and (C) Smo-/- fibroblasts after treatment with vehicle (EtOH),
calcitriol, or cyclopamine (CP). Ptchflox/floxERT2+/- fibroblasts were cultured in either Shh-N-
conditioned medium (Shh-N-CM) or control medium (CoM) with or without 10 µM
tamoxifen as indicated. Smo-/- fibroblasts were either transfected with a hSMO expressing
plasmid or with empty vector as indicated. Asterisks: p<0.05; error bars: mean+/-SD; ntc: no
template control.
Figure 4: Calcitriol inhibits the Hh-signaling pathway in a Vdr-independent manner.
(A) Gli and Cyp24a1 expression of Vdr-/- fibroblasts after treatment with vehicle (EtOH),
calcitriol, or cyclopamine (CP) and Shh-N-conditioned medium (Shh-N-CM) or control
medium (CoM). (B) Analyses of the Vdr protein level of nuclear extracts of si-Vdr or si-
control transfected Ptch-/- and Vdr-/- fibroblasts by western blot. Detection of HSC-70 protein
served as control. (C) and (D) Gli1, Cyp24a1 and Vdr expression of Ptch-/- and Shh-
stimulated Ptchflox/floxERT2+/- fibroblasts after si-Vdr or si-control transfection and calcitriol
treatment. Gli1 expression of Ptch-/- fibroblasts and Vdr expression of Ptch-/- and Ptchflox/flox
ERT2 cells are shown in relation to the respective vehicle-treated control. Asterisks: p<0.05;
error bars: mean+/-SD.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

25
Figure 5: Model for the dual function of calcitriol in Ptch-associated BCC. Normally Ptch
inhibits its signaling partner Smo, thereby regulating the activity of the Hh-signaling pathway
(normal cell). Mutations of Ptch lead to a constitutive activation of the Gli transcription
factors, which results in cell proliferation and tumor formation (BCC cell). The known Smo-
inhibitor cyclopamine inhibits Hh-pathway in the Ptch-mutant cells and thus Hh-related
processes involved in tumor growth (cyclopamine-treated BCC cell). In contrast, calcitriol
inhibits Hh-pathway activity and additionally activates Vdr-signaling (calcitriol-treated BCC
cell). Consequently, calcitriol not only inhibits tumor-relevant processes mediated by Hh
signaling, but also induces anti-proliferative effects and differentiation processes via the Vdr-
signaling pathway.
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422

Published OnlineFirst August 30, 2011.Mol Cancer Ther Anja Uhmann, Hannah Niemann, Bérénice Lammering, et al. receptor-signaling and differentiationinhibition of Hedgehog-signaling and induction of vitamin D Antitumoral effects of calcitriol in basal cell carcinomas involve
Updated version
10.1158/1535-7163.MCT-11-0422doi:
Access the most recent version of this article at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/early/2011/08/25/1535-7163.MCT-11-0422To request permission to re-use all or part of this article, use this link
on July 29, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 30, 2011; DOI: 10.1158/1535-7163.MCT-11-0422




![Vitamin D in PdPregnancy and Infancy · 1,25-dihydroxy vitamin D = calcitriol [1,25(OH)2D] assessment (hormonal form) ↓ Calcitriol binds to vitamin D receptor (VDR) to regulate](https://static.fdocuments.in/doc/165x107/5ffb3ce26517d830b10f5d09/vitamin-d-in-pdpregnancy-and-125-dihydroxy-vitamin-d-calcitriol-125oh2d.jpg)













