
ANTIBACTERIAL EFFECTS OF NITRITE IN URINE
58
From the Department of Surgical Science, Section of Urology and the Department of Physiology and Pharmacology Karolinska Institutet, Stockholm, Sweden ANTIBACTERIAL EFFECTS OF NITRITE IN URINE Stefan Carlsson M.D. Stockholm 2005
Transcript of ANTIBACTERIAL EFFECTS OF NITRITE IN URINE

From the Department of Surgical Science, Section of Urology and the Department of Physiology and Pharmacology Karolinska Institutet, Stockholm, Sweden
ANTIBACTERIAL EFFECTS
OF NITRITE IN URINE
Stefan Carlsson M.D.
Stockholm 2005

All previously published papers were reprinted with permission of the publishers. Printed by Repro Print AB, Box 21085 100 31 Stockholm, Sweden © Stefan Carlsson, 2005 ISBN 91-7140-237-3

To Anne, Emma, Adam and Fabian

”Hela bollen måste ligga stilla”
Ralf Edström

Antibacterial effects of nitrite in urine
Stefan Carlsson, MD From the Department of Surgical Science, Section of Urology
and the Department of Physiology and Pharmacology Karolinska Institutet, Stockholm, Sweden
Abstract
Urinary tract infections (UTI`s) are among the most common conditions causing individuals to seek medical care. In addition, catheter-associated infections account for most of the hospital-acquired UTI´s and is also a major source of resistant nosocomial pathogens. The majority of bacterial strains causing UTI have a nitrate-reducing capacity and since the 1920s, when the urinary nitrite test was first developed, this dipstick assay has been an important component of modern UTI diagnosis. In this thesis we have expanded the role of nitrite beyond diagnosis of infection and explored its role as an antimicrobial agent. From earlier studies it is known that acidification of nitrite results in the formation of nitric oxide (NO) and other reactive nitrogen oxides, which are toxic to a variety of microorganisms. The aim of the present thesis was to investigate NO formation and possible antibacterial effects of acidified nitrite-containing urine. We also sought to determine the antibacterial effect of a two-step procedure in which bacteria were first fed with nitrate to form nitrite, followed later by acidification of the urine. Finally we wanted to evaluate in vitro a novel concept for intravesical delivery of antibacterial NO via the retention balloon of a urinary catheter. NO formation was measured in headspace gas using a chemiluminiscence-technique, while antibacterial effects were evaluated by turbidity measurements and by viable counts. We show that large amounts of the antimicrobial gas NO are generated in mildly acidified nitrite-containing human urine and this NO formation is greatly enhanced by the addition of ascorbic acid. Furthermore, we show that mildly acidified nitrite-containing human urine has potent antimicrobial activity against three of the most common urinary pathogens and this inhibitory effect is further increased by ascorbic acid. We also found, that Escherichia coli self destructs when it is first allowed to generate nitrite and then transferred to mildly acidified urine. Translated into clinical terms, this could imply that ingestion of nitrate followed some hours later by acidification of urine could be a new approach for treatment of UTI. Finally, potent antibacterial effects were also observed in infected urine when the nitrite-derived nitrogen oxides were delivered via the retention balloon of a urinary catheter. In conclusion, we describe potent antibacterial effects of acidified nitrite-containing urine and also suggest a novel attractive alternative to prevent catheter-associated urinary tract infections. Key words: nitrite, nitrate, nitric oxide, urinary tract infection, catheter-associated urinary tract infection, antimicrobial, Escherichia coli. ISBN 91-7140-237-3

LIST OF PUBLICATIONS
This thesis is based on the following papers which are referred to in the text by their Roman numerals (I-IV):
I. JO Lundberg, S Carlsson, L Engstrand, E Morcos, NP Wiklund and E Weitzberg. Urinary nitrite: More than a marker of infection. Urology, 50:189-191, 1997.
II. S Carlsson, NP Wiklund, L Engstrand, E Weitzberg and JO Lundberg. Effects of pH, nitrite, and ascorbic acid on nonenzymatic nitric oxide generation and bacterial growth in urine. Nitric Oxide: Biology and Chemistry, 5:580-586, 2001.
III. S Carlsson, M Govoni, NP Wiklund, E Weitzberg and JO Lundberg. In vitro evaluation of a new treatment for urinary tract infections caused by nitrate-reducing bacteria. Antimicrobial Agents and Chemotherapy, 47: 3713-3718, 2003.
IV. S Carlsson, E Weitzberg, NP Wiklund and JO Lundberg.
Intravesical nitric oxide delivery for prevention of catheter-associated urinary tract infections. Antimicrobial Agents and Chemotherapy, (in press May 2005).

CONTENTS ABBREVIATIONS................................................................................................................................... 9
INTRODUCTION ................................................................................................................................... 10 URINARY TRACT INFECTIONS ................................................................................................................ 10
Aetiology ....................................................................................................................................................... 10 Bacterial virulence ....................................................................................................................................... 11 Host defense mechanisms ............................................................................................................................ 11 Diagnosis ...................................................................................................................................................... 12 Treatment and Antimicrobial resistance. .................................................................................................... 13 Prophylaxis and Acidification of urine ....................................................................................................... 13 Catheter-associated urinary tract infection and Biofilms ........................................................................... 14
NITRIC OXIDE....................................................................................................................................... 15 The history of nitric oxide. ........................................................................................................................... 15 Enzymatic formation of NO ......................................................................................................................... 16 Nonenzymatic formation of NO................................................................................................................... 16 Nitrate in medicine ....................................................................................................................................... 17 Metabolism of nitrate/nitrite ........................................................................................................................ 18 Chemistry of NO, nitrate and nitrite ............................................................................................................ 18 Beneficial effects of NO ............................................................................................................................... 20 Beneficial effects of acidified nitrite............................................................................................................ 21 NO in inflammation and host-defense ........................................................................................................ 22 NO and inflammation in the lower urinary tract ........................................................................................ 23 Bacterial defense against RNIs.................................................................................................................... 24
AIMS...................................................................................................................................................... 25
MATERIAL AND METHODS ................................................................................................................ 26 STUDY SUBJECTS................................................................................................................................. 26 CHEMICAL ANALYSIS OF NO, NITRITE AND NITRATE ............................................................................... 26
Analysis of NO in gaseous phase by chemiluminescence ........................................................................... 26 Quantification of nitrite and nitrate by chemiluminescence ...................................................................... 27 Quantification of nitrite by capillary electrophoresis ................................................................................. 28
BACTERIAL CULTURES AND MEDIA ........................................................................................................ 28 ANALYSIS OF BACTERIAL GROWTH........................................................................................................ 29
Turbidity measurements............................................................................................................................... 29 Viable counts measurements ....................................................................................................................... 30
STATISTICS .......................................................................................................................................... 31 RESULTS AND COMMENTS ............................................................................................................... 32
NO FORMATION FROM NITRITE IN URINE ................................................................................................ 32 EFFECTS OF NITRITE ON BACTERIAL GROWTH ........................................................................................ 33
Effects of sodium nitrite ............................................................................................................................... 33 Bacterial growth after pre-incubation with nitrate ..................................................................................... 35 Delivery of antibacterial nitrogen oxides via a catheter retention balloon ................................................ 37
GENERAL DISCUSSION...................................................................................................................... 38 ACIDIFIED NITRITE AND ANTIBACTERIAL EFFECTS IN URINE ..................................................................... 38
What is the mechanism of action?............................................................................................................... 38 Bacterial defense systems............................................................................................................................. 40
POSSIBLE CLINICAL USEFULNESS ......................................................................................................... 41 Treatment of UTI ......................................................................................................................................... 41 Prevention of CAUTI ................................................................................................................................... 43
CONCLUSIONS .................................................................................................................................... 45
ACKNOWLEDGEMENTS ..................................................................................................................... 46
REFERENCES ...................................................................................................................................... 48

9
ABBREVIATIONS ABU asymptomatic bacteriuria BCG bacillus calmette-Guérin CAUTI catheter-associated urinary tract infection CE capillary electrophoresis CFU colony-forming units cGMP cyclic guanosine monophosphate cNOS constitutive nitric oxide synthase E. coli Escherichia coli EDRF endothelium-derived relaxing factor eNOS endothelial nitric oxide synthase HNO2 nitrous acid H2O2 hydrogen peroxide HPLC high-performance liquid chromatography IL-8 interleukin 8 iNOS inducible nitric oxide synthase L-NAME NG-nitro- L-arginine-methylester L-NMMA NG-monomethyl-L-arginine LPS lipopolysacharides MBC minimum bactericidal concentration MIC minimum inhibitory concentration mRNA messenger ribonucleic acid NADPH nicotineamide adenine dinucleotide phosphate NO2
- nitrite
NO3- nitrate
NO2 nitrogen dioxide N2O3 di-nitrogen trioxide N2O4 di-nitrogen tetroxide nNOS neuronal nitric oxide synthase NO nitric oxide NOS nitric oxide synthase O2
- superoxide OH hydroxyl radical ONOO- peroxynitrite PDE-5 phosphodiesterase type-5 PMNL polymorphonuclear leukocytes PHOX phagocyte oxidase ppm parts per million ppb parts per billion pO2 partial pressure of oxygen RNI reactive nitrogen intermediates ROS reactive oxygen intermediates RSNO S-nitrosothiols sGC soluble guanylyl cyclase TMP trimethoprim SMX sulfamethoxazole SOD superoxide dismutase UTI urinary tract infection

10
INTRODUCTION
Urinary tract infections Urinary tract infection (UTI) is defined as an inflammatory response of the urothelium to
bacterial invasion [1]. Acute uncomplicated UTI with an estimated incidence of 150 million
yearly on a global basis is among the most common conditions causing individuals to seek
medical care [2]. At the age of about thirty, 50% of all women report having had a UTI. In
contrast, only 20% of men in their seventies have experienced a UTI [3, 4] Approximately
25% of women who have had an episode of acute cystitis develop recurrent UTI [5]. UTI
affects up to 10% of the childhood population and is an important cause of morbidity [6]. It is
a heterogeneous disease which can be sub-divided into a variety of clinical conditions ranging
from the asymptomatic presence of bacteria in the urine to severe, sometimes life-threatening,
infections of the kidney with septicaemia [7].
Aetiology
Microorganisms reach the urinary tract by the ways of the ascending route and via the
haematogenous or lymphatic routes. The ascending route via urethra from periurethral
colonization is the most common pathway [8]. This explains the greater frequency of UTI in
women since the female urethra is short and situated in close proximity to the vaginal
vestibule and rectum. It also explains the increased risk of infection following bladder
catheterization or instrumentation. Other risk factors, apart from gender and the use of
catheters/surgical instruments, are vaginal intercourse, spermicide use and underlying
conditions affecting the urinary tract such as diabetes and pregnancy. Obstruction of urinary
flow (e.g. because of prostate enlargement) and diminished ability to empty the bladder (e.g.
because of spinal cord injury, decreased bladder contractility or neuropathy) also increase the
risk of UTI [4]. Haematogenous infections of the urinary tract can be caused by
Staphylococcus aureus, Candida species or Mycobacterium tuberculosis. These infections are
uncommon in normal adults, but can occasionally occur in immunocompromised patients and
in neonates [9, 10]. UTI via the lymphatic route is probably not of importance, but may occur
in unusual circumstances such as a severe bowel infection or retroperitoneal abscess [10, 11].
Escherichia coli (E. coli) is by far the most common cause of UTI, accounting for 80-85 % of
community-acquired infections and 50% of hospital-acquired infections. Other gram-negative
enterobacteriaceae including Proteus, Klebsiella and gram-positive Enterococcus faecalis

11
and Staphylococcus saprophyticus are responsible for the remainder of most community-
acquired infections [12, 13]. In hospital-acquired UTI E. coli is still the leading cause of UTI,
but other bacteria such as Pseudomonas aeruginosa, Klebsiella species and Proteus mirabilis
are seen more frequent as causative agents [14].
Bacterial virulence
Uropathogenic bacteria are selected from the faecal flora by presence of virulence factors that
enable them to adhere to and colonize the perineum and urethra and migrate to the urinary
tract [15, 16]. These virulence factors include adhesins, siderophores, toxins, polysaccharide
coating, proteases and invasins. However, no single virulence factor is common to all clinical
UTI isolates and none of these factors is absolutely necessary for development of UTI
pathogenesis [17]. The more compromised the natural defense mechanism (e.g. obstruction,
bladder catheterization, diabetes with poor blood glucose control), the fewer virulence factors
are needed for any bacterial strain to induce infection. This is supported by the observation
that bacteria isolated from patients with a complicated UTI frequently fail to express virulence
factors [17-19]. There has also been suggested that if a bacterial strain had a doubling time of
about 50 minutes or less in urine, it could maintain itself in the normal bladder without
adhering [20].
Host defense mechanisms
Flow of urine through the urinary tract and voiding are effective natural bladder defenses
against infection [8, 21]. In addition, low pH, urea and high urine osmolality can be inhibitory
to bacterial growth. Both concentration of urea and urine osmolality are decreased by urine
dilution, which has led some authors to suggest that diuresis actually could favour bacterial
proliferation [22-24]. The asymptomatic bacteriuria (ABU) strains do not activate a host
response because these strains stop expressing adherence factors once they are established in
the urine thereby avoiding the antibacterial defense [25]. In contrast, the uropathogenic
strains, continue to adhere and activate the uroepithelial cells to release cytokines that recruit
polymorphonuclear leukocytes (PMNL) to the site of infection. During their passage across
the mucosa , the PMNL kill the bacteria [26]. The cytokine interleukin-8 (IL-8) seems to be
the main driving force for the PMNL to cross the urinary tract epithelium and the emergence
of leukocytes in the urine, known as pyuria , is a classical sign of UTI [25]. The attachment of
a uropathogenic strain may result in exfoliation of host bladder epithelial cells as part of an
innate host defense system [27]. Another host defense strategy is to produce soluble receptor

12
molecules that compete with bacterial adhesins and block bacterial attachments [28]. The
vaginal flora, dominated by Lactobacillus species, is an additional important host defense
system against UTI. Particularly the Lactobacillus species that produce hydrogen peroxide
(H2O2) seems to have a protective effect [29]. Lactobacilli have also been shown to generate
an acidic vaginal pH, which appear to be of importance for lowering the rate of UTI [30].
Unfortunately, use of antibiotics and intravaginal antimicrobials including antimycotics and
spermicides reduces the vaginal flora and increases susceptibility to UTI [31].
Diagnosis
Urine cultures are still the golden standard to diagnose UTI as it has been for decades,
although the number of bacteria considered relevant for diagnosis has been changed since the
classical “Kass criteria” of 105 bacteria/ml were described in 1956 [32]. Stamm and co-
workers showed in 1982 that a count of ≥ 105/ml identified only about 50% of dysuric
women with bacteriuria, whereas a count of ≥ 102/ml had a predictive value of 0.88 for
diagnosing a probable urinary tract infection [33]. The diagnostic criteria for UTI in acute
uncomplicated cystitis in women is ≥ 103 colony-forming units (CFU) of uropathogen/ml of
mid-stream sample of urine according to EAU guidelines [2, 34]. Since the 1920s, when the
urinary nitrite test was first developed, this dipstick assay has been an important component
of modern UTI diagnosis [35, 36]. Most dipstick assays will indicate positive nitrite (change
of colour) when urinary nitrite levels exceed 7-25 µM, as a result of bacterial nitrate-reducing
capacity [37, 38]. Today the common urine dipstick test in use detects nitrite and leukocyte
esterase. These tests are rapid and cheap alternatives, but show less accuracy compared with a
quantitive culture [39]. Another clear disadvantage with the dipstick assays is that they do not
give information about the exact species that caused the UTI or the antimicrobial
susceptibility profiles. The majority of bacterial strains causing UTI have a nitrate-reducing
capacity, like for example E coli, Pseudomonas aeruginosa and Proteus mirabilis.
Staphylococcus saprophyticus can also express nitrate-reductase, but normally very low levels
[40]. However, streptococci, including strains of Enterococcus faecalis, do not have nitrate
reductase enzymes and lack the capacity to reduce urinary nitrate to nitrite [41, 42]. Recently,
a meta-analysis demonstrated that sensitivities of the combination of both nitrites and
leukocyte-esterase vary between 68 and 88% in different patient groups. The sensitivity was
also analysed in different patient settings and found to be highest in studies carried out in
family medicine (90%). The sensitivity of the urine dipstick test for nitrite alone was 45-60%
with higher level of specificity (85-98%) [43]. Reasons for false negative results using

13
dipstick test for nitrite i.e., negative nitrite assay in urine with more than 105 nitrate-reducing
organisms/ml could be urinary pH below 6.0, urobilinogen, lack of dietary nitrate or intake of
ascorbic acid [37]. Also high frequency of micturition might be a reason for false negative
results, since an incubation at 37◦ C of four to six hours is preferable for generation of
detectable nitrite in infected urine samples [41].
Treatment and Antimicrobial resistance.
In treatment of uncomplicated cystitis, a short course of antibiotics are highly effective and
also desirable because of the improved compliance that they promote, their low cost and the
low frequency of adverse reactions. According to American guidelines, trimethoprim-
sulfamethoxazole (TMP-SMX) for three days is considered the current standard therapy [44].
TMP alone and other fluoro-quinolones (ofloxacin, norfloxacin, ciprofloxacin, and fleroxacin)
are equivalent to TMP-SMX [44]. Pivmecillinam and nitrofurantoin are alternative oral drugs,
especially in situations in which fluoroquinolones are not indicated [45, 46]. In Sweden,
clinical practice guidelines have been developed by the STRAMA-group, which recommends
pivmecillinam or nitrofurantoin or TMP for seven days as a standard treatment of
uncomplicated cystitis in women [47]. However, there are other Swedish studies supporting
short-term treatment for 3-5 days of uncomplicated lower urinary tract infections in women
[48, 49]. A growing problem of worldwide concern is the increasing resistance of pathogens
to conventional antibiotics [50]. In UTI much of the increase in bacterial resistance is in acute
uncomplicated cystitis with increasing TMP-SMX and β-lactam resistance. Of more concern,
however, are the emerging issues of fluoroquinolone resistance and multidrugresistance
among community-acquired urinary isolate. Important strategies to help slow the progression
of resistance is a judicious use of antibiotics and to develop novel methods for the prevention
of UTI [51].
Prophylaxis and Acidification of urine
Urinary acidification has long been used as an aid in the treatment and prevention of urinary
tract infections. Various agents have been used in attempts to lower urinary pH, these include
mandelic acid, gluconic acid, ammonium chloride, methionine and ascorbic acid (vitamin C)
[52-55]. Urinary acidification with vitamin C has traditionally been used as a household
remedy, although evidence to support its efficacy has largely been anecdotal and any
antibacterial action still remains unclear [56]. Effective acidification of urine has been poor
and antibacterial effects variable [57-59]. Methenamine hippurate, a weak base that slowly

14
hydrolyzes in acidic urine to ammonia and the nonspecific antibacterial formaldehyde, are
often used for the prevention of urinary tract infection. However, there is not enough evidence
to conclusively support the use of methenamine hippurate for UTI prophylaxis [60]. Today
most prophylaxis of UTI consists of antimicrobial therapy. The increasing prevalence of E.
coli isolates that are resistant to antimicrobial agents, however, has stimulated interest in
novel, non-antibiotic methods for preventing UTI. Natural compounds like cranberry products
have been more popular and used as a household remedy for treating and preventing UTI
[61]. It has been postulated that acidification of urine is an important means by which
cranberry products might protect against UTI. However, effective acidification of urine
through cranberry ingestion has not been proved [62]. The mechanism of action is likely to be
a tannin (proanthocyanidin) that block adherence of P-fimbriated E. coli [63]. Although there
are some clinical trials that have shown positive effects with cranberry ingestions in
preventing UTI a Cochrane review in 2001 concludes that there is no compelling evidence to
recommend cranberries for prevention of UTI [64].
Catheter-associated urinary tract infection and Biofilms
Catheter-associated urinary tract infection (CAUTI) accounts for most of the hospital-
acquired UTI [65]. In medical intensive care units in the US as much as 95% of urinary tract
infections were associated with urinary catheters [66]. CAUTI is also a major source of
resistant nosocomial pathogens [67]. The incidence of bacteriuria in catheterized patients
varies between 3% and 10% per day [68]. A biofilm is a collection of microbial organisms on
a surface that is surrounded by an extra cellular matrix composed primarily of poly-saccharide
materials. Urinary catheters can develop biofilm in two ways. The extra luminal route, by
direct inoculation at time of catheter insertion or by migration in the mucous sheath
surrounding the catheter, is the most common way (about 70%) [69]. Adherent bacteria
aggregate, multiply and form a biofilm within a few hours [70]. Microorganism may also
enter the catheter from the internal lumen of the catheter, but this intra luminal route
presupposes a contamination of the closed drainage system [71]. The biofilm provides a
protective environment for the microorganisms and make them less susceptible to
antimicrobial agents. The reason for this poor effect of antimicrobial agents is that the
bacteria in the biofilm grow much slower than bacteria growing freely within the urine and
also that the extra cellular matrix prevent penetration of antimicrobial agents into the biofilm
[72]. Another problem with biofilm production is that freely growing bacteria found in
cultures obtained from catheterized patients may not reflect bacterial population growing

15
within the biofilm [73]. Guidelines recommend aseptic technique during catheter insertion
and stress the importance of using a closed urinary drainage system to minimise the
development of CAUTI [74]. Other approaches in preventing these infections include
systemic antibiotics and anti-infective catheters [70] . Silver alloy catheters have been used
and the result of a meta-analysis indicates that they likely prevent bacteriuria and despite a
higher cost also seem economically efficient when used in patients receiving indwelling
catheterization for 2 to 10 days [75, 76]. Impregnation with salicylic acid and even
electrification have been showed to reduce bacterial adhesion to urethral catheters [77, 78].
Additional methods are suprapubic catheters and clean intermittent catheterization.
Suprapubic catheters are mainly used in patients requiring long-term catheterization and
several trials have indicated that patients have lower risk of bacteriuria and higher rate of
satisfaction compared with those using indwelling catheters [79]. Clean intermittent
catheterization is associated with a decreased risk for UTI compared with long-term
indwelling catheters [80]. Despite these preventive efforts CAUTIs are very common and
result in substantially increased health care costs and leads to substantial morbidity and
mortality with data suggesting an almost threefold increase in mortality even when comorbid
conditions and other factors are accounted for [81, 82].
Nitric Oxide The history of nitric oxide. In 1980, Furchgott and co-workers showed that acetylcholine-induced relaxation of blood
vessels was dependent on the endothelium and as a consequence the endothelium-derived
relaxing factor (EDRF) was discovered [83]. Murad and co-workers had three years earlier
shown that nitroglycerine and nitroprusside cause relaxation of smooth muscle cells via
activation of the enzyme soluble guanylyl cyclas (sGC) which increase cyclic guanosine
monophosphate (cGMP) [84]. Murad and co-workers then found that Nitric Oxide (NO) was
the mediator of this cGC activation and, in 1983, that EDRF induced relaxation in smooth
muscle cells via increased cGMP [85, 86]. Eventually in 1987, Ignarro and Moncada with co-
workers independently showed that NO is identical to EDRF [87, 88]. With these Nobel Prize
rewarded discoveries (Furchgott, Ignarro and Murad in 1998), NO went from being regarded
merely an atmospheric pollutant to a highly significant biological molecule. The discovery of
NO led to an explosion of research regarding the biological significance of this small diatomic
molecule with, until today, more than 65 000 publications on the topic.

16
Enzymatic formation of NO
Mammalian cells synthesize NO enzymatically from the amino acid L-arginine and molecular
oxygen by nitric oxide synthase (NOS) [89] (fig 1). This enzymatic reaction is nicotineamide
adenine dinucleotide phosphate (NADPH)-dependent and results in formation of equimolar
amounts of L-citrulline [90]. NOS also needs the cofactors heme, flavin adenine dinucleotide,
flavin mononucleotide and tetrahydrobiopterin [91]. At present, three main isoforms of the
enzyme have been identified, out of which two are expressed constitutively and one isoform
is inducible. The isoforms of the enzymes were named after the tissue in which they were
initially found [92]. The two constitutively expressed (cNOS) are neuronal NOS (nNOS) and
endothelial NOS (eNOS) and activation of these enzymes requires calmodulin, which in turn
is controlled by the intracellular concentration of Ca2+ [93, 94]. Within seconds from
stimulation with agonists such acetylcholine or bradykinin, cNOS produce femtomolar to
picomolar concentration of NO. The third isoenzyme, iNOS, is on the other hand independent
of free Ca2+and its expression can be induced by bacterial products such as lipopolysaccharide
and proinfammatory cytokines (e.g. tumour necrosis factor-α, interleukin-1β and interferon-γ)
a process involving transcription factors (e.g. nuclear factor-κB) and thereby increasing iNOS
mRNA [95, 96]. The time that is required for iNOS mRNA and protein to be synthesized
results in a delay of several hours between cell activation and NO synthesis. Once iNOS is
expressed, which can happen in a wide variety of human cells, it can produce nanomolar
concentration of NO and this production is sustained over a prolonged period of time.
Nonenzymatic formation of NO
Chemical synthesis of NO in vivo was first described 1994 when Benjamin and co-workers
and Lundberg and co-workers independently showed NO production in the stomach, which
relies on the secretion of nitrate in saliva and bacterial conversion to nitrite on the tongue with
reduction to NO by stomach acid [97, 98]. The dorsal surface of the tongue harbours a
specialized flora of symbiotic nitrate-reducing facultative anaerobic bacteria [99].
Nonenzymatic formation occurs in the stomach at low pH, when the nitrite ion (NO2-) is
converted to nitrous acid (HNO2), which subsequently decomposes to various nitrogen oxides
including NO in the parts per million (ppm) range [97, 98] (fig 1).

17
Fig 1. Enzymatic and nonenzymatic NO formation.
Apart from pH, this nonenzymatic formation depends on pO2, proximity to heme containing
proteins, redox state and thiol concentration [100]. Reducing agents, like ascorbic acid, will
further enhance this reaction. Ascorbic acid and ascorbate ion will generate NO over a wide
pH range in aqueous solution by rapid reduction of nitrous acid (HNO2) and formation of
dehydroascorbic acid [101] (fig 1). In addition to the stomach, in vivo non-enzymatic
formation of NO has also been shown in the oral cavity, the heart, and on the skin [99, 102-
104]. The enzyme nitrate reductase is not found in humans, but a number of commensal
bacteria inhabiting mucosal surfaces have multiple nitrate reductases and can catalyse the
reduction of nitrate to nitrite especially during oxygen limitation. There are now increasing
data suggesting that these nitrate-reducing commensals have a true symbiotic role in
mammals [105, 106]. Other enzymes in humans, that can reduce nitrite to NO are
mitochondrial enzymes, xantineoxidase and cytochrome P450 [107-110]. In addition, deoxy-
hemoglobin has recently been shown to reduce nitrite to NO and thereby causing
vasodilatation in the human circulation [111].
Nitrate in medicine
During the 19th century inorganic nitrate was a very common component in many galenic
preparations and was widely used in treatment of different disorders (eg. oedemas, pain and
inflammation) [112]. In the early decade of the 20th century it was shown that inorganic
nitrate could cause methaemoglobinaemia in infants and its therapeutic use clearly fell into
Enzymatic NO formation L-Arginine + NADPH + O2 L-Citrulline + NO + NADP+
NOS
Nonenzymatic NO formation NO2
- + H+ HNO2 Nitrite Nitrous acid
2HNO2 N2O3 + H2O Nitrous acid Dinitrogen trioxide
N2O3 NO + NO2 Dinitrogen trioxide Nitrogen dioxide
Ascorbic acid + 2HNO2 Dehydroascorbic acid + 2NO + 2H2O Nitrous acid

18
disgrace [113]. Organic nitrates, like glyceryl trinitrate and isorbididinitrate on the other hand,
have been used therapeutically for over 100 years and are still in use in the treatment and
prophylaxis of angina pectoris [114]. Since around 1950 when inorganic nitrate was suspected
to be associated with development of gastric cancer its bad reputation grew even worse [115].
The metabolism of nitrate and nitrite can result in N-nitrosoamines which are carcinogenic in
cell cultures and animal models [116]. However, despite numerous studies, there is still no
clear evidence in humans for a link between inorganic nitrate intake and gastric cancer and
most studies show no relationship between a high intake of nitrate and gastric cancer in
humans [117-120]. More recent studies even indicate the opposite, that in humans the
bacterial metabolism of nitrate to nitrite and the subsequent formation of biologically active
nitrogen oxides could in fact be beneficial [105, 121].
Metabolism of nitrate/nitrite
Green vegetables such as lettuce and spinach, root vegetables such as beetroot and drinking
water are the main sources of exogenous nitrate intake [122]. Exogenous intake of nitrite is
much smaller, but nitrite can be found in some food as a ingredient in red meat curing giving
cured meat and hot dogs their red colour (nitrosylated myoglobin) and protecting it from
oxidation and spoiling [123]. Nitrite is particularly effective against the pathogen Clostridium
Botulinum [124]. About 90% of ingested nitrite comes from nitrate in saliva and less than
10% comes from food [125]. The main source of endogenous nitrate in mammals is the L-
arginine-NO-pathway, which in fasting humans constitutes the major source of nitrate/nitrite
in plasma [126]. After ingestion, nitrate is absorbed from the stomach and proximal small
intestine into plasma, where it mixes with endogenously synthesized nitrate [127]. It is then
concentrated, by a factor of ten, from the plasma into the saliva [128]. By this entero-salivary
circulation of nitrate, about 25% of plasma nitrate is actively taken up by the salivary glands
and secreted with saliva [129]. About 1/5 of this recycled nitrate (approximately 5% of the
total ingested nitrate) is converted to nitrite by oral cavity organisms [130]. Studies with
isotope labelled administrated nitrate have shown that only 60% is recovered in the urine.
Consequently, the exact fate of all the nitrate in the body is still unresolved [131].
Chemistry of NO, nitrate and nitrite
The gas NO, with a molecule weight of only 30 Dalton, is certainly one of the smallest
biological mediators. Due to it small size, uncharged character and lipid solubility it diffuses
easily and rapidly over biological membranes. NO is also a free radical in that it has an

19
unpaired electron and is therefore extraordinary labile, with a half-life of only about 1-5
seconds in most biological systems [132]. NO exhibits a low level of solubility in water
(approximately 9 times more lipid soluble than water soluble) and when inserted into a
physiological buffer, the major part of the gas immediately appears in the headspace [133].
Despite being a radical the reactivity of NO under physiologically circumstances is quite
limited to either a reaction with other radicals, with oxygen or with transition metals [134].
The in vivo reaction of NO and superoxide (O2-), that forms peroxynitrite (ONOO-), is an
example of two relatively unreactive free radicals that will produce a much more reactive
species (fig 2). Peroxynitrite is a potentially cytotoxic substance that can be protonated to
peroxynitrous acid (OONOH), which in turn dissociates to oxidizing hydroxyl radical (OH)
and nitrogen dioxide radical (NO2) within seconds at physiological pH [135] (fig 2).
However, eukaryotic cells contain large amounts of superoxide dismutas (SOD), an enzyme
that keep concentration of O2- remarkably low [136]. High concentrations of gaseous NO can
react with oxygen to form NO2, a poisonous gas called “brown gas” (fig 2). However, the
reaction is of second order with respect to NO which means that it will be slower when NO
concentrations are low for example in vivo (normally 100-300 nM and no more than 1-3 µM
during inflammation). Much more common in biological fluids, with oxygen present, is the
rapid oxidation of NO to nitrite (NO2-) or nitrate (NO3
-) [137]. One of the most important
examples of a reaction with metals is when NO interacts with the heme group in the enzyme
sGC and thereby increases cGMP, which in turn mediates many of the biological actions of
NO [138]. Another example of reaction with metals occurs in the blood, where NO reacts
with Hb(Fe2+)O2 to form NO3- and methaemoglobin. This is suggested to be an important
pathway for NO elimination [139] (fig 2). NO can also react with thiol groups allowing the
formation of S-nitrosthiols (RSNOs) that may represent a physiologically important source of
NO [140]. Acidification of nitrite will generate nitrous acid (HNO2), which will
spontaneously yield di-nitrogen trioxide (N2O3), NO and nitrogen dioxide (NO2) [100]. The in
vivo chemistry of these reactive nitrogen intermediates (RNIs), many of which have
biological effects, is very complex and still not fully characterized [141, 142]. Until recently,
the above mentioned oxidation of NO to the inert and stable end products nitrite and nitrate
was generally accepted as a mechanism for inactivation of NO. However, several recent
studies have shown that different pathways exist to recycle nitrite back into bioactive NO in
blood and tissue [97, 98, 102, 108, 110, 111, 143].

20
Fig 2. NO reaction with oxygen, superoxide and oxyhaemoglobin. Nitrate reduction by
bacterial nitrate reductase (NR).
Beneficial effects of NO
NO is a unique messenger molecule. In the autonomic nervous system NO functions as a
major non-adrenergic non-cholinergic neurotransmitter. Moreover, NO has a particularly
important role in relaxation of smooth muscle in the gastrointestinal and urogenital tracts and
NO is responsible for the smooth muscle relaxation of corpus cavernosus leading to penile
erection. These important smooth muscle relaxations are mediated by the reaction of NO with
sGC leading to increased cGMP [83, 144, 145]. Sildenafil (Viagra®), a drug that inhibits
phosphodiesterase type-5 (PDE-5) and thereby inhibit the degradation of cGMP leading to a
prolonged effect of cGMP and better erection, has revolutionised the treatment of erectile
dysfunction [146]. In the urinary tract of animal models, NOS activity has been found in the
urothelium, smooth muscle, striated muscle, nerves and blood vessels [147]. In addition, NO
is an important mediator in dilation of bladder neck, urethra and urethral sphincter during the
micturation reflex [148]. NO is also a potent vasodilator and inhibitor of platelet aggregation
and adhesion [149-151]. Inhibitors of NO synthases have been very useful to explore the
biological activities of NO. Two L-arginine analogues, NG-monomethyl-L-arginine (L-
Nitric oxide oxidation 2NO + O2 2NO2
Nitric oxide oxygen Nitrogen dioxide NO + O2
- OONO- Nitric oxide Superoxide Peroxynitrite OONO- NO3
-
Peroxynitrite Nitrate OONO- + H+ HONOO OH + NO2 Peroxynitrite Peroxynitrous acid Hydroxyl radical Nitrogen dioxide NO + Hb(Fe2+)O2 NO3
- + MetHb(Fe3+) Nitric oxide Oxyhaemoglobin Nitrate Methaemoglobin
Nitrate reduction
NO3
- + e- + 2H+ NO2- + H2O
Nitrate Nitrite NR

21
NMMA) and NG-nitro- L-arginine-methylester (L-NAME), that lack selectivity for the NOS
isoforms, have been shown to increase blood pressure in normotensive humans, confirming
that NO is important in the regulation of vascular tone [152, 153]. There are also several NO-
donors currently being used in various types of experimental studies [154]. Some of these
different NO-donors appear to be of interest for their potential therapeutic applications, for
example the diazeniumdiolates (formerly NONOates), that have been used in studies
regarding inhibiting restenosis after angioplasty, preparing thromboresistant medical devices,
reversing vasospasm, and relieving pulmonary hypertension [155]. Another very exciting
group of NO-donors, are the NO-non-steroidal anti-inflammatory (NO-NSAIDs) drugs, which
are characterized by a reduction in gastrointestinal side effects [156, 157]. Since NO is also
thought to play a role in detrusor instability, a NO-releasing and prostaglandin synthesis-
inhibitory-drug has been used in patients with neurogenic bladder instability with promising
results [157].
Beneficial effects of acidified nitrite.
Dietary nitrate has an important role in protection against ingested pathogens. After a high
nitrate containing meal, like a portion of lettuce, the levels of nitrite and nitrate in saliva
increase. When this saliva enters the acidic environment in the stomach, it results in an
increased NO concentration from basal levels of 20 parts per million (ppm) to > 400 ppm.
The acid environment alone is not sufficient for killing of pathogenic bacteria in the stomach.
However, after mixture of salivary nitrite in gastric juice, most pathogenic bacteria are killed
within a hour in vitro [105, 127]. Dietary nitrate and the above described acidified-nitrite-
derived NO are also of great importance as a gastroprotective agent. Bjorne and co-workers
have shown in a rat in vivo model that mucosal blood flow and mucus secretion increased
after luminal application of nitrite-rich saliva, whereas saliva from a fasting individual had no
effect [158]. Acidified nitrite has antimicrobial effect on periodontal bacteria, which might
explain some of the protective effects of normal saliva against dental caries [159].
Commensal skin bacteria have been shown to produce NO by the reduction of sweat nitrate to
nitrite and its subsequent conversion to NO by the acidic environment [103]. In vitro studies
have shown that acidified nitrite is microbiocidal to common cutaneous pathogens [160]. As
described earlier, acidification of nitrite results in production of a complex mixture of
nitrogen oxides including nitrous acid (HNO2), di-nitrogen trioxide (N2O3), nitrogen dioxide
(NO2) and NO. All these nitrogen oxides may act as nitrosating agents and can rapidly react
with reduced thiols to form nitrosothiols. Nitrosothiols as well as many other of these nitrogen

22
oxides can kill microbes and therefore the exact mechanism responsible for the above
described in vivo antibacterial effects of acidified nitrite is not fully understood and needs to
be further investigated.
NO in inflammation and host-defense
In 1983, it was shown that that E. coli lipopolysaccharides (LPS) stimulates urinary nitrate
excretion and it was proposed that this mammalian nitrate biosynthesis was the result of
oxidation of reduced nitrogen compounds [161]. In 1985, Marletta and Stuehr reported that
LPS-stimulated peritoneal macrophages from mice, synthesize nitrite and nitrate from an
unknown precursor molecule [162]. Three years later the same group showed that NO, was
produced and that L-arginine, was the precursor [162, 163]. Independently in 1987, Hibbs and
co-workers, showed that the anti-tumour effect of macrophages were abolished when arginine
was removed from the medium [164]. In addition to macrophages, cytokine-induced high-
output NO synthesis, has been found in most somatic cells involved in cell-mediated immune
reactions [165]. Today, NO is clearly regarded as a central component of innate immunity and
an effective antimicrobial agent [166]. On the other hand, the large amounts of NO resulting
from increased iNOS expression, may also contribute to the morbidity of infection by acting
as vasodilator, myocardial depressant and cytotoxic mediator [167]. Not all of the host–
derived NO is synthesized by NOS. As described earlier, dietary nitrate can be reduced by
oral bacteria to nitrite and then in the acidic environment in the stomach further reduced to
NO and other RNIs may also be formed. The first line of host defense is the innate immune
response, where phagocytic cells are very important components. In addition to the iNOS
pathway that generate NO radicals, phagocytic cells use another important antimicrobial
system, the NADPH phagocyte oxidase (PHOX) pathway responsible for superoxide (O2-)
generation. Superoxide (O2-) is like hydrogen peroxide (H2O2) and hydroxyl radical (OH)
intermediate reduction products of oxygen en route to water and they are commonly called
“reactive oxygen species” (ROS) [168]. The microbial targets of ROS and RNI are mainly
thiols, metal centres and DNA. RNI mainly inhibit respiration and interfere with DNA
replication, whereas ROS work mainly through direct DNA damage. The antimicrobial
actions of RNI are more complex than those of ROS and multiple cellular targets are almost
certainly involved, although the exact mechanism for the antimicrobial effects are still not
known [167, 169, 170]. ROS/ RNI and phagocytic cells represent nonspecificity in the
immune system and are effective killers of microbes and essential for clearing invading
microbes from an infected area. Phagocytes are quick to sacrifice themselves and surrounding

23
host tissue to prevent microbial metastasis, thus resulting in further tissues damage [169] .
Increased NO production in vivo has been described in many chronic inflammatory processes,
such as for example cystitis [171], asthma [172], colitis [173] and rheumatoid arthritis [174].
NO has been suggested to be very useful as a non-invasive marker of several inflammatory
diseases, although the role of NO in chronic inflammatory processes remains to be elucidated
[175]. However, there are now studies questioning whether the sustained iNOS expression
seen in many chronic inflammatory diseases, for example psoriasis, is actively producing
significant amounts of NO in vivo and recent evidence, in fact, suggest that lack of NO
formation in this disease is of importance in the pathophysiological process [176].
NO and inflammation in the lower urinary tract.
In studies of the mouse urinary tract, as discussed earlier, it has been shown that the
uropathogenic strains adhere to the uroepithelial cells and activate them to release cytokines.
The cytokines recruit polymorphonuclear leukocytes (PMNL) that clear the urinary tract from
the infection [177]. In humans, increased iNOS activity has been detected during UTI from
neutrophils in the urine and the iNOS activity persist for a period of time after initiation of
antibiotic treatment [178]. Luminal NO in the urinary bladder is increased in patients with
cystitis of different aetiology [179]. Cytokines, as well as bacillus calmette-Guérin (BCG),
have been shown to increase iNOS activity in vitro in human uroepithelial cells [180, 181].
However, studies with UTI in iNOS-deficient mice have shown that there were no differences
in bacterial clearance or persistence between mice with intact iNOS capacity compared with
the deficient genotype [182]. It has been shown that human bladder smooth muscle cells
respond to LPS and inflammatory cytokines with increased iNOS activity and collagen type
III expression. The authors suggest that iNOS may be a critical mediator of the bladder wall
fibrotic response to chronic UTI [183]. In ifosfamide induced hemorrhagic cystitis in mice,
NO produced by iNOS, is suggested to be responsible for urothelial damage and inflammation
[184]. In conclusion, the exact role of NO in human UTI is not clear, but taking the well-
known antimicrobial properties into account it is very likely that NO in some way enhance
local host defense in the bladder.

24
Bacterial defense against RNIs.
Since protection against ROI have been studied for over 100 years, detoxification-enzymes
like microbial catalase, SOD and peroxides that convert ROS to less toxic species have been
well documented [185]. In contrast, until about seventeen years ago, RNI production in
mammals was not thought to exist, because RNI was regarded to be too toxic [169].
Knowledge about resistance to RNI is therefore a relatively new concept. However, some
pathways for microbial defense against RNI have now been identified and include scavenge
of RNIs, suppressed RNI production, repair of nitrosative damage and enzymatically
detoxification of RNIs. The most studied bacterial enzymatic detoxification defense to NO
and other RNIs are in E coli, where nitrite reductases and NO reductases, as well as a
flavohaemoglobin, have been characterized [105, 186]. Studies in E coli have also
demonstrated specific antioxidant regulons. One example is a family of approximately 12
genes (even including the SOD gene) called the soxRS regulon, which may protect bacteria
against NO [187].

25
AIMS
The general aim of this thesis was to investigate antibacterial effects of nitrite in urine.
The specific aims were:
• To investigate the changes in NO formation in relation to different concentrations of
nitrite and different pH-levels with or without ascorbic acid in normal and infected
urine.
• To study antibacterial effects of acidified nitrite and ascorbic acid against three
common urinary pathogens at different urinary pH levels.
• To evaluate the minimal bacteriostatic and the minimal bactericidal concentrations of
acidified nitrite and ascorbic acid and to compare the effects with conventional
antibiotics.
• To evaluate the antibacterial effect of a new two-step procedure in which bacteria are
first fed with nitrate, followed by acidification of the urine.
• To evaluate the in vitro antibacterial effect of a novel concept for delivery of NO via a
catheter retention balloon into the surrounding urine.

26
MATERIAL AND METHODS
Study subjects In paper I urine was collected from 8 patients (41-70 years old) with bacteriuria as confirmed
by urinary cultures. Urine was also collected from 5 healthy subjects before and after
ingestion of vitamin C ( 2g/day) for 2 days.
In paper III 8 healthy subjects mean age 37 years (range 25-47) fasted overnight. Basal
urinary samples were collected and then the subjects ingested sodium nitrate (10 mg/kg)
dissolved in 150 mL water. Urinary samples were collected at 1, 2 and 3 hours.
Chemical analysis of NO, nitrite and nitrate Analysis of NO in gaseous phase by chemiluminescence (paper I-II and IV)
In this thesis we have used two chemiluminescence NO analysers when measuring gaseous
NO. In paper I and II CLD 700, Eco Physics, Dürnten, Switzerland was used and in paper
IV, Aerocrine AB, Stockholm, Sweden was used. Measurement of gaseous NO with
chemiluminescence, is based on the reaction of NO with an excess of ozone (O3) to produce
nitrogen dioxide (NO2), which is partly excited (NO2*). When NO2
* returns to its ground state
the excess energy is released as a photon (hv) and light (luminescence) in the 640-3000 nm
wavelength range is emitted, which is measured by a sensitive photomultiplier tube. The
intensity of this luminescence is proportionally converted into an electrical signal and
displayed as NO levels with a response time of less than 0.7 s. The chemiluminiscence assay
is extremely sensitive for NO and has a detection limit of 1 part per billion (ppb) and a linear
response for concentrations between 1-100 000 ppb. The assay also shows a high specificity
for NO without interference from other nitrogen oxides [188]. When ≥50 ml of NO-
containing gas (0.5-20 ppm) was injected into the NO analyser the recovery of the NO signal
was 100%. In this thesis we made use of the well documented properties of NO, as a gas with
poor solubility in water and extremely high diffusibility. Thus, when NO is injected into a
physiological buffer, >80% immediately appears in the gas space (head space) [133]. NO has
also been shown to diffuse through the inflatable retention balloon of a urinary catheter [133,
179, 189]. In paper I and II, urinary samples (10 ml) were incubated at 37◦ C in closed
syringes with a headspace of 50 ml. After 30 min, the headspace gas was removed and
immediately injected into the chemiluminescence analyser (CLD 700, Eco Physics, Dürnten,
Switzerland). Ambient NO levels were below 5 ppb. In paper I the urinary samples consisted

27
of 10 ml infected urine or 10 ml control urine with or without addition of 100 µM sodium
nitrite and NO release was measured at different pH levels. NO was also measured before and
after ingestion of vitamin C (2g/day) for 2 days in 10 ml urinary samples with addition of 100
µM sodium nitrite at different urinary pH levels (pH 4-7). In paper II NO was measured in
10 ml urinary samples consisting of sodium nitrite (50-500 µM) with or without addition of
vitamin C (10 mM) at different urinary pH levels (pH 4.5-6.0). Ambient NO levels were
below 5 ppb in all experiments.
In paper IV An all-silicone catheter (Argyle®, Sherwood Medical, Tullamore, Ireland) was
placed in a bladder model (50 ml flask) and the retention balloon was filled with 10 ml of
saline containing ascorbic acid (10 mM) and sodium nitrite (5 mM) at pH 2.5. The flask was
then closed and synthetic NO-free air was flushed via an inlet at a rate of 4 L/min. Headspace
NO concentration was continuously measured from an outlet by a rapid-response chemi-
luminescence system (Aerocrine AB, Stockholm, Sweden) at room temperature (20° C ).
Ambient NO levels were below 5 ppb in all experiments.
Quantification of nitrite and nitrate by chemiluminescence
In paper III and paper IV nitrite and/or nitrate sample concentrations were determined by
chemiluminescence after reductive cleavage and subsequent determination of the NO released
into the gas phase. The samples were directly introduced via a gas-tight syringe into a
reduction solution of a micro reaction purge vessel coupled with a condenser and heating
jackets unit (Sievers, Boulder, CO, USA). The condenser jacket temperature was controlled
by a continuous flow of cold water while the temperature of the heating jacket was controlled
by a flow of warm water regulated by a constant-temperature circulating bath (MGW Lauda
M3). Nitrogen gas, at flow of 192 mL/min, was used as the carrier gas of NO. The flow could
be adjusted with a needle valve integrated with the purge vessel and the outlet of the gas
stream was passed through a scrubbing bottle containing sodium hydroxide (1 M, 0°C) in
order to trap traces of acid before transfer into the NO analyzer. A chemiluminescence NO
analyser (Aerocrine AB, Stockholm, Sweden) was used to display the NO signals and collect
the data, which were further manipulated with Origin for Windows, Version 7.0 (Microcal,
Northampton, MA) and reported as area under the curve. Nitrite was determined according to
Feelish et al [190]. The reducing mixture, consisting of 45 mmol/L potassium iodide (KI) and
10 mmol/L iodine (I2) in glacial acetic acid, was kept at a constant temperature of 56 °C and
continuously bubbled with nitrogen gas. The amount of nitrite in a given sample was
quantified by simple subtraction of the peak areas of sample aliquots pretreated with

28
sulfanilamide from that of untreated aliquots (10% (v/v) of a 5% solution of sulfanilamide in
1N HCl is added to the biological sample (final concentration 29 mmol/L) and incubated for
15 min at room temperature. Under these conditions, nitrite reacts with sulfanilamide to form
a stable diazonium ion that is not converted to NO. Nitrate was reduced to NO with a solution
of Vanadium(III)chloride in hydrochloric acid 1N (saturated solution) at 95°C. Since
Vanadium(III)/HCl will also convert nitrite to NO the amount of nitrate was quantified by
subtraction of the nitrite concentration calculated before.
Quantification of nitrite by capillary electrophoresis
Capillary electrophoresis (CE) has been used in the analysis of several ions and
micromolecules, because of its high efficiency of separation and short analysis time compared
to other available techniques such as high-performance liquid chromatography (HPLC). In
paper I CE was used to analyse urinary nitrite concentrations in 8 patients with bacteruria.
After collecting midstream urine the samples were stored at -80◦ C until analysis. The CE
system used was Hewlett Packard capillary electrophoresis system (HP 3D, Waldbrunn,
Germany) with UV detection at 214 nm. The background electrolyte consisted of 25 mM
sodium sulphate and 5% NICE-Pack OFM Anion-BT, dissolved in freshly brought Milli-Q
water. Samples were injected hydrodynamically by 30 mbar for 60 seconds onto extended
light path capillaries (104 cm effective lenght and 75 µm ID). The analysis was done at an
applied negative voltage of 300 kV/m and the temperature of the capillaries was set to 40◦ C.
Data were analyzed with a HP 3D CE Chem Station data system. Standard curves for nitrite
was plotted. Before being used for the first time, capillars were conditioned with sodium
hydroxide (1 and 0.1 M) and the background electrolyte for 5 min each while the temperature
of the capillary was set to 60◦ C.
Bacterial cultures and media
The strains used in this thesis were E. coli U1106024 (paper IV) isolated from a patient with
urinary tract infection and reference strains E. coli ATCC 25922 (paper I-IV) , P. aeruginosa
ATCC 27853 (paper II), S. saprophyticus SS2 (paper II), all obtained from the department
of Clinical Microbiology, Uppsala, Sweden. In paper III E. coli RK 4353 and a RK 4353
mutant, lacking all three known nitrate reductase enzymes were used [191]. The RK 4353
strains were a generous gift from Dr. JA. Cole, Birmingham, UK. Before each experiment

29
bacteria were grown aerobically in Mueller-Hinton broth for 6 hours at 37° C, resulting in 2-5
x 108 colony-forming units (CFU)/mL.
All growth experiments in this thesis were carried out in pooled urine. Midstream urine was
collected from healthy subjects (paper I: 5 subjects, paper II: 8 subjects, paper III: 10
subjects, paper IV: 4 subjects) pooled, divided into batches (50 ml) and immediately frozen
(-20 C) until use.
Analysis of bacterial growth
Turbidity measurements
The basic concept for turbidity measurements is that bacteria scatter light in proportion to
their number. The turbidity or optical density of a suspension of cells is directly related to the
cell mass or cell number, after construction and calibration of a standard curve. The method is
efficient, reproducible and has been widely used as a rapid screening method for large number
of antimicrobial agents [192, 193]. In paper I-II the bacterial strains were grown in Mueller-
Hinton broth for 6 hours at 37° C, resulting in 2-5 x 108 colony-forming units (CFU)/mL. The
strains were diluted to a bacterial density of 106 CFU/mL in the pooled urine. After 2 hours of
incubation at 37° C in a closed tube (in pooled urine with various amounts of nitrite, with or
without addition of vitamin C and at various urinary pH-levels), 40 µL of the bacterial
suspension was transferred to micro well plates containing 360 µL sterile filtered control
urine (pH 6). After this dilution (1/10) and transfer to the recovery medium (filtered control
urine, pH 6), bacterial growth was measured continuously for 10 hours (paper I) and 20
hours (paper II) by vertical photometry (optical density) at a wavelength of 540 nm in a
computerized incubator for bacteria (Bioscreen C, Labsytems, Helsinki, Finland). In paper
III the bacterial culture was diluted to a bacterial density of 2-5 x 106 CFU/mL in flat-bottom
micro well plates (96 wells of 300 μL) containing the pooled urine and various amounts of
nitrite, with or without addition of vitamin C and at various urinary pH-levels. Note that in
paper III, there is no use of a recovery medium. Bacterial growth was measured continuously
for 20 hours at 37° C by vertical photometry (optical density) at a wavelength of 540 nm in a
computerized incubator (Molecular Devices Spectra Max 340 Sunnyvale, CA). Minimum
inhibitory concentration (MIC) was defined as the lowest concentration at which no visible
growth had taken place after 20 hours. MICs of nitrite and ascorbic acid were determined. We
also determined MIC values for nitrite in combination with a fixed concentration of ascorbic

30
acid (10 mM). MIC was also determined for nitrofurantoin and trimethoprim using the same
urine.
Viable counts measurements
Viable count involve spreading (plating out) a sample of a culture on a nutritient agar surface.
The sample is diluted in a non-toxic diluent (e.g. phosphate-buffered saline) before plating.
Each viable unit grows and forms a colony. Each colony that can be counted is called a CFU
and the number of CFU´s is related to the viable number of bacteria in the sample. An
excellent correlation has been shown by Domínguez and co-workers between antibacterial
effects by turbidity (Bioscreen C) and viable counts methods for ciprofloxacin and standard
strains of E. coli and P aeruginosa [194].
In paper III we used the viable count method for determination of bactericidal activity of
various amounts of nitrite, with or without addition of vitamin C and at various urinary pH-
levels in the pooled urine. In paper IV we used the viable count method for determination of
bactericidal activity of a novel in vitro concept of delivery of NO via a catheter retention
balloon into the surrounding urine. An all-silicone catheter (Argyle, Sherwood Medical,
Tullamore, Ireland) was placed in the flask and the retention balloon was filled with 10 ml of
saline containing ascorbic acid (10 mM) and sodium nitrite (5 mM). The acidity of the
solution was adjusted to pH 2.5 using hydrochloric acid (3 M). Ascorbic acid and nitrite were
prepared and mixed immediately before administration. Ascorbic acid solution alone (pH 2.5)
was used in control catheters. After filling the retention balloons the catheter was gently
pulled outwards and fixed at the neck whereby the flask-opening was sealed off. Then the
flasks were turned up-side-down and incubated at 37 degrees for 24 h.
After 20 hours (paper III) or 24 hours (paper IV) of bacterial growth/incubation (at 37° C,
10 μL urine was diluted in series with PBS (pH 7.3) and transferred to agar plates. In separate
experiments (paper IV) we also tested shorter exposure times of 5, 30, 60 and 120 min. The
agar plates were incubated for 24 hours and then a viable count was performed (counting
CFU/ml). MBC (Minimum Bactericidal Concentrations) was defined as the concentration
where at least 99.9% (a reduction of > 3 log CFU/ml) of the original inoculate was killed.
In paper IV we also compared the effects of nitrite with another NO-releasing compound
DETA NONOate, which is considered to be a rather pure NO donor. In these experiments the
sodium nitrite was replaced with DETA NONOate (0.05, 0.5 or 5 mM) and all other test
parameters were identical. DETA NONOate was prepared immediately before the start of the
experiment. We then used the viable count method for determination of bactericidal activity

31
as described above. To exclude any antibacterial effect of the parent compound (a polyamine),
the same experiment was also performed with DETA NONOate solution that had been
prepared and stirred for 72 h in open air to release all NO (complete NO release was
confirmed with chemiluminescence showing that the NO signal at 24 h was <0.1 % of the
initial peak level).
Statistics Experimental data are expressed as mean ± S.E.M (paper I-II) and as mean ± S.D. (paper III
–IV). Statistical significance was tested according to Mann-Whitney U test (paper I). For
calculation of statistical difference between repeated measurements analysis of variance
(ANOVA) was used (paper II).

32
0
1
2
3
4
5
6
6,05,5
5,04,5
BasalAA
50+AA100+AA
500+ AA
log
NO
(ppb
)
RESULTS AND COMMENTS
NO formation from nitrite in urine (paper I-II, IV)
Infected urine may contain considerable amounts of nitrite as a result of bacterial nitrate
reductase activity. We sought to determine whether NO was generated from acidified nitrite-
containing urine. We found that at a basal urinary pH (5.5-6.5), NO formation from both
control urine and infected urine was low. In contrast, large amounts of NO were generated
from infected urine (containing 8 to 400µM nitrite) when the urine was acidified to pH 4.5-
5.0 and from acidified non-infected urine if nitrite (100 µM) was added. Urinary NO
formation was strongly pH dependent in the presence of nitrite and was markedly enhanced
by addition of ascorbic acid. We also found that increasing concentrations of nitrite (0-500
μM) caused a dose-dependent enhancement of NO formation that again was greatly enhanced
by the addition of ascorbic acid (Fig 3 and 4).
Fig 3. Urinary NO formation (ppb) in urine from different concentrations of sodium nitrite (0-
500 µM) with the addition of ascorbic acid (AA ,10mM) or from AA alone, at varying urinary
pH values (4.5-6.0).

33
pH 6.0 pH 5.5 pH 5.0 pH 4.5
log
NO
ppb
1
2
3
4
5
6nitritenitrite + AA
Fig 4. NO formation in urine from nitrite (500 µM) at different urinary pH values (4.5-6.0) with
and without the addition of ascorbic acid (AA ,10mM).
We also studied whether a nitrite/ascorbic acid-containing solution when placed in a catheter
retention balloon would generate NO that could be measured outside the balloons. Indeed,
NO did traverse the silicone membrane and measured levels peaked initially at about10 ppm
in the headspace gas. If the balloon was filled with nitrite alone (pH 7) or ascorbic acid alone
(pH 2.5) no NO signal was noted (NO < 0.001 ppm). These results shows that a tiny
uncharged gas such as NO can easily diffuse through the silicon membrane of the retention
balloon of a silicone urinary catheter (Argyle®), which is in accordance with previous findings
when a similar catheter was used as a sampling device [133, 179, 189].
Effects of nitrite on bacterial growth Effects of sodium nitrite (paper I-III)
Urine is an excellent culture medium for bacteria that cause urinary tract infections [32].
Indeed, at a basal urinary pH and even in slightly acidic urine the bacterial growth was very
good. In contrast, the growth of E. coli, P. aeruginosa and S. saprophyticus was dose-
dependently inhibited by nitrite in acidified urine. The inhibitory effect of nitrite was greater
at lower pH. Addition of ascorbic acid (10 mM) further enhanced the inhibition of bacterial
growth by nitrite. Increasing concentrations of nitrite (0-500 μM) with a fixed concentration
of ascorbic acid (10mM) caused a dose-dependent inhibition of E.coli growth which
correlated to the NO formation observed (Fig 5). Note that by this method the bacteria are
transferred to a recovery medium after 2 hours of nitrite exposure and thus we are studying
post-antibiotic effects.
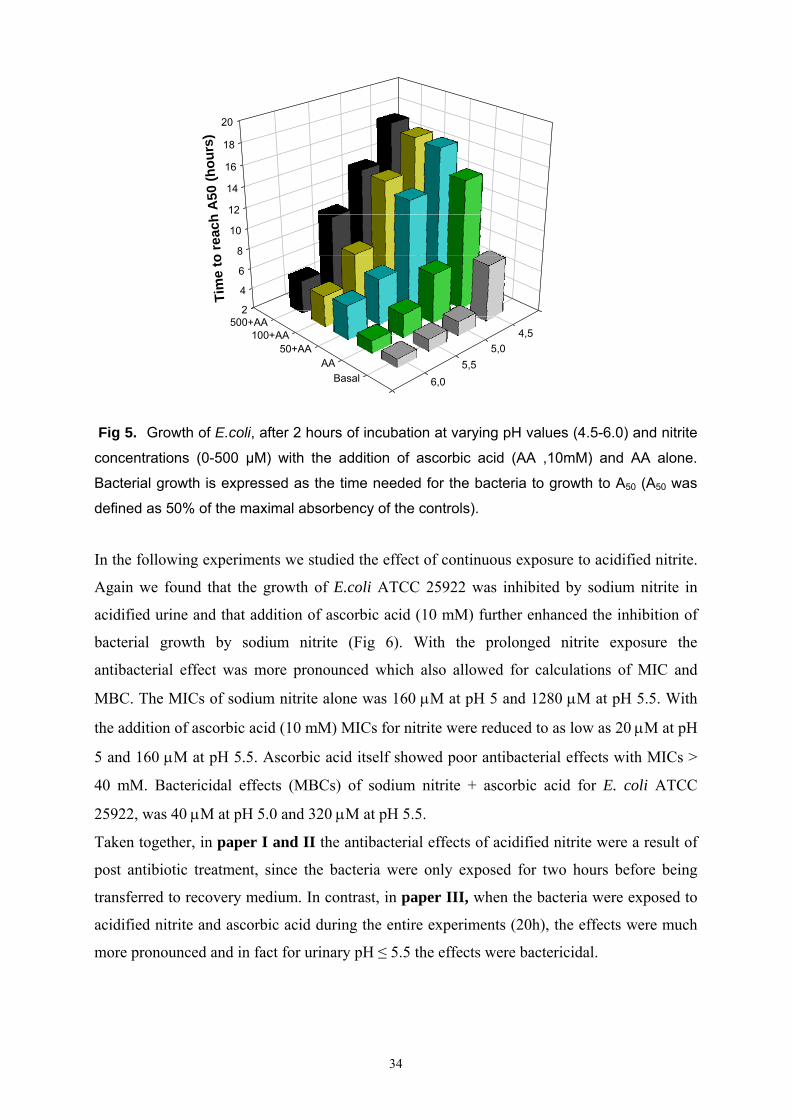
34
Fig 5. Growth of E.coli, after 2 hours of incubation at varying pH values (4.5-6.0) and nitrite
concentrations (0-500 µM) with the addition of ascorbic acid (AA ,10mM) and AA alone.
Bacterial growth is expressed as the time needed for the bacteria to growth to A50 (A50 was
defined as 50% of the maximal absorbency of the controls).
In the following experiments we studied the effect of continuous exposure to acidified nitrite.
Again we found that the growth of E.coli ATCC 25922 was inhibited by sodium nitrite in
acidified urine and that addition of ascorbic acid (10 mM) further enhanced the inhibition of
bacterial growth by sodium nitrite (Fig 6). With the prolonged nitrite exposure the
antibacterial effect was more pronounced which also allowed for calculations of MIC and
MBC. The MICs of sodium nitrite alone was 160 μM at pH 5 and 1280 μM at pH 5.5. With
the addition of ascorbic acid (10 mM) MICs for nitrite were reduced to as low as 20 μM at pH
5 and 160 μM at pH 5.5. Ascorbic acid itself showed poor antibacterial effects with MICs >
40 mM. Bactericidal effects (MBCs) of sodium nitrite + ascorbic acid for E. coli ATCC
25922, was 40 μM at pH 5.0 and 320 μM at pH 5.5.
Taken together, in paper I and II the antibacterial effects of acidified nitrite were a result of
post antibiotic treatment, since the bacteria were only exposed for two hours before being
transferred to recovery medium. In contrast, in paper III, when the bacteria were exposed to
acidified nitrite and ascorbic acid during the entire experiments (20h), the effects were much
more pronounced and in fact for urinary pH ≤ 5.5 the effects were bactericidal.
2
4
6
8
10
12
14
16
18
20
6,05,5
5,04,5
BasalAA
50+AA100+AA
500+AA
Tim
e to
reac
h A
50 (h
ours
)

35
Fig 6. Growth of E.coli in urine at pH 5.5 with different amounts of sodium nitrite (20-160 µM)
and a fixed concentration of ascorbic acid (AA, 10 mM).
Bacterial growth after pre-incubation with nitrate (paper III)
Having confirmed that exogenous nitrite had antibacterial effects in urine, we wanted to go on
and study if nitrite generated by the bacteria themselves could be turned against them in the
form of toxic RNIs. We first studied how much nitrate that would accumulate in urine after
ingesting 10 mg/kg of body weight sodium nitrate (corresponds to the amounts found in about
300g of spinach). Basal nitrate levels in urine were about 0.7 mM in fasting individuals and
these levels increased about 10-fold already one hour after ingestion of sodium nitrate. Now
knowing the amount of nitrate that could easily be achieved in urine, we continued with
studying how much nitrite the nitrate reducing bacteria, E. coli ATCC 25922, could form
from this nitrate. For comparisons we used a E. coli mutant that lacked all nitrate reducing
enzymes. When E. coli was incubated for 20 hours in basal urine (nitrate 0.7 mM) about 2 µM
nitrite was generated. When the basal urine had been supplemented with 1 mM sodium nitrate
the nitrite formation increased to 370 µM. Maximum formation of nitrite was seen after
supplementation with 3 and 10 mM sodium nitrate (2470 and 2400 µM nitrite respectively).
When the mutant lacking all three nitrate reductases was incubated for 20 hours with
maximum substitution of sodium nitrate (10 mM) the formation of nitrite was negligible. Note
that resulting nitrite concentrations after transferring the cultures to the acidic urine in the
second part of the experiment were 1/10 of the values above, in other words 37, 247 and
240µM nitrite respectively.
Time (hours)
0 5 10 15 20
Opt
ical
den
sity
0,10
0,15
0,20
0,25
0,30
0,35
0,40
Basal20 µM nitrite + AA40 µM nitrite + AA80 µM nitrite + AA160 µM nitrite +AA

36
Fig 7 Growth of two strains of E. coli. A control strain and a mutant lacking nitrate reductase.
The bacteria were first grown at pH 7 with sodium nitrate (10mM). After 20 hours the cultures
were transferred to acidified urine (pH 5.0) and bacterial survival was monitored by viable
counts.
Having shown that E. coli ATCC 25922 was able to produce significant amounts of nitrite
from urinary nitrate, we then continued with studying whether acidification in a second step
would result in antibacterial effects. We first incubated E. coli in urine at pH 7.0 with 0-10
mM sodium nitrate added, which resulted in accumulation of nitrite in the medium.
Subsequent transferring of the nitrite-rich culture to acidic urine resulted in a marked decrease
in viable counts. This effect was dose-dependent with more effective killing rates at higher
pre-incubation levels of sodium nitrate. At the highest level of nitrate (10 mM), a final
reduction of more than 3 logs CFU/ml (bactericidal effect) was observed after transfer to
acidic medium. If bacteria were pre-incubated in basal urine without addition of sodium
nitrate, no inhibition was noted when the culture was transferred to the acidic urine. This
indicates that a normal diet in a patient treated with urinary acidification is probably
insufficient to excrete enough nitrate in the urine to achieve antibacterial effects.
In experiments with the mutant lacking nitrate reductases and a control strain both strains
grew similarly well at pH 7.0 with 10 mM sodium nitrate added. When transferring the
cultures to acidic urine containing ascorbic acid the control strain was effectively killed while
the mutant retained full viability (fig 7). Taken together, in paper III we have shown that E.
Control strain
Nitrate reductase mutant
20 h
pH 7.3
20 h
pH 5
Overnight Culture
Urine + nitrate Nitrite↑↑
20 h
pH 7.3
20 h
pH 5
Overnight culture
Urine + nitrate Nitrite≈0
Time hours0 5 10 15 20
log
CFU
/ml
2
3
4
5
6
7
8
9
10
Time hours0 5 10 15 20
log
CFU
/ml
2
3
4
5
6
7
8
9
10

37
coli is effectively killed when it is first allowed to generate nitrite in nitrate rich urine at
neutral pH and then transferred to mildly acidified urine. In contrast, a mutant lacking a
nitrate reducing capacity, retained full viability by this procedure.
Delivery of antibacterial nitrogen oxides via a catheter retention balloon (paperIV)
A model of a catheterized urinary bladder was used to study a novel approach for intravesical
delivery of an antibacterial agent. Bacteria were grown in urine outside of the retention
balloon thereby simulating a catheterized bladder with infection. When the catheter balloon
was filled with ascorbic acid without nitrite (pH 2.5), the bacterial counts of the reference
strain had increased from 2.5x106 to 2.7x108 CFU/ml and the clinical isolate strain had
increased from 9.0x105 to 2.0 x108 CFU/ml after 24 h. In contrast, when using test catheters
with ascorbic acid and nitrite (pH 2.5) both tested strains were effectively killed (fig 8).
To further try to pinpoint the compound responsible for antibacterial effects of acidified
nitrite we also studied the effects of a pure NO donor. A 24 h exposure to the NO-donor
DETA NONOate was completely bacteriostatic at 5 mM while 0.5 and 0.05 mM had no effect
on bacterial growth. Thus, in paper IV we describe a novel concept, which allows for
intravesical delivery of antibacterial gases into the urinary bladder model via the catheter
retention balloon, resulting in bacteriostatic and bactericidal effects in the urine.
Fig 8. Effects of test (5 mM nitrite + 10 mM ascorbic acid) and control (ascorbic acid alone)
catheters on the growth of two strains of E coli in a model of urinary tract infection (pH 6.1).
log
CFU
/ml
1
2
3
4
5
6
7
8
9
Reference controlReference test Clinical isolate controlClinical isolate test
0 24h

38
GENERAL DISCUSSION
Acidified nitrite and antibacterial effects in urine Nitrite has been used in the meat curing process since the 1920s because it produces a
desirable bright pink colour and also enhances flavour. In addition, in 1934 it was shown by
Tanner and Evans that the growth of the anaerobic bacteria of the genus Clostridium was
inhibited by this process [195]. Since 1941, when Tarr showed that bacteria were susceptible
to low concentration of nitrite provided the meat was sufficiently acid, the antimicrobial
effects of acidified nitrite have been used to prevent from botulism [196]. Later it was shown
that addition of ascorbic acid enhanced the inhibition of Clostridium botulinum by nitrite and
that antibacterial effects of nitrite in meat involves generation of NO [197, 198]. However,
not until 1994 when Benjamin/Lundberg and co-workers demonstrated NO generation in the
stomach from acidified nitrite, it was appreciated that this chemical reaction was of
importance even in vivo [97, 98].
In this thesis we have shown (paper I) that large amounts of the antimicrobial gas NO is
formed in mildly acidified nitrite-containing human urine and this NO formation is greatly
enhanced by the addition of ascorbic acid. Furthermore, we have shown (paper I and II) that
mildly acidified nitrite-containing human urine has potent antimicrobial activity against three
of the most common urinary pathogens and this inhibitory effect is further increased by
ascorbic acid. We have also shown (paper III), that E. coli is effectively killed when it is first
allowed to generate nitrite and then transferred to mildly acidified urine. Finally we have
suggested (in paper IV) a new approach to prevent CAUTI by intravesical delivery of
antibacterial compounds generated from nitrite and ascorbic acid via the catheter retention
balloon.
What is the mechanism of action?
It is obvious from this thesis that acidified nitrite together with ascorbic acid shows
antimicrobial effects in urine. However, the exact chemical nature of the toxic nitrite-derived
compound is not known and neither is the exact mechanism by which bacterial killing occurs.
Dykhuizen and co-workers, showed bactericidal effects of acidified nitrite in the stomach
against several common gut pathogens. They claimed that the antimicrobial effect is due to an
additive contribution of reactive intermediates of nitrogen [199]. The chemistry of acidified
nitrite is very complex and a variety of reactive nitrogen intermediates (RNIs) and other
nitrogen oxides are generated directly or after reactions with other compounds.
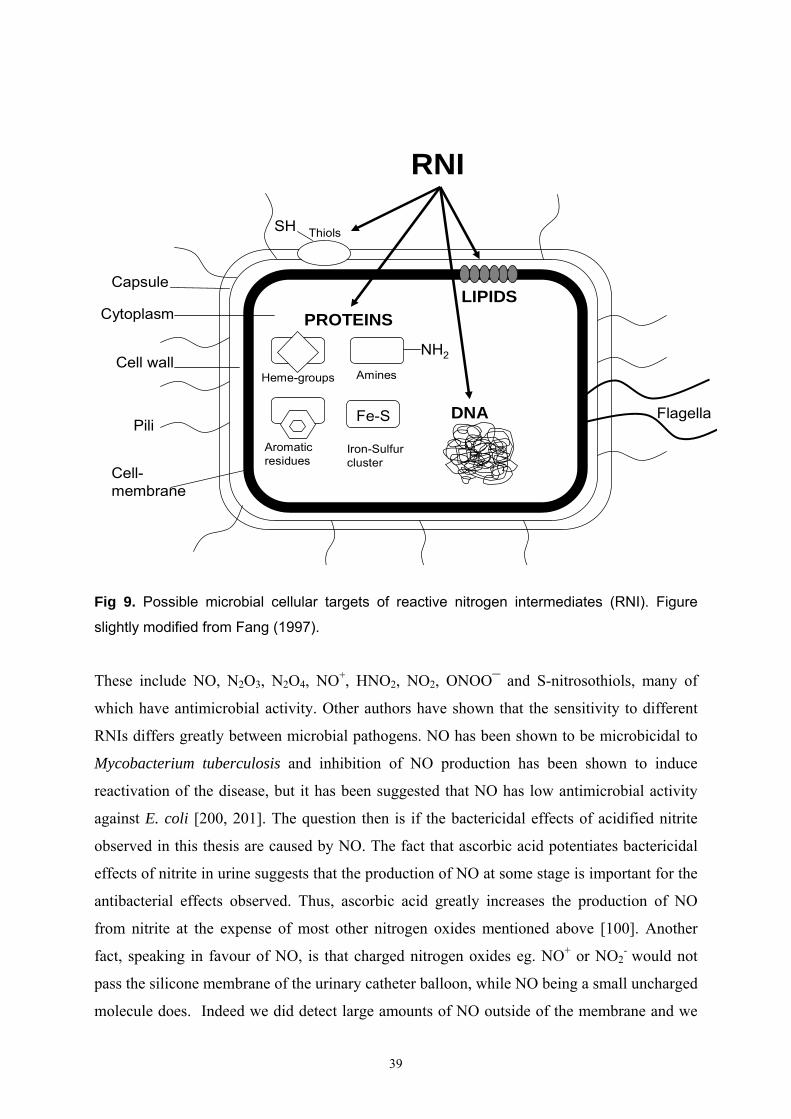
39
PiliFlagella
Capsule
DNA
Cell wall
Cytoplasm
Cell-membrane
LIPIDS
SH
PROTEINS
RNI
Thiols
NH2
Fe-S
Heme-groups
Aromaticresidues
Amines
Iron-Sulfurcluster
Fig 9. Possible microbial cellular targets of reactive nitrogen intermediates (RNI). Figure
slightly modified from Fang (1997).
These include NO, N2O3, N2O4, NO+, HNO2, NO2, ONOO¯ and S-nitrosothiols, many of
which have antimicrobial activity. Other authors have shown that the sensitivity to different
RNIs differs greatly between microbial pathogens. NO has been shown to be microbicidal to
Mycobacterium tuberculosis and inhibition of NO production has been shown to induce
reactivation of the disease, but it has been suggested that NO has low antimicrobial activity
against E. coli [200, 201]. The question then is if the bactericidal effects of acidified nitrite
observed in this thesis are caused by NO. The fact that ascorbic acid potentiates bactericidal
effects of nitrite in urine suggests that the production of NO at some stage is important for the
antibacterial effects observed. Thus, ascorbic acid greatly increases the production of NO
from nitrite at the expense of most other nitrogen oxides mentioned above [100]. Another
fact, speaking in favour of NO, is that charged nitrogen oxides eg. NO+ or NO2- would not
pass the silicone membrane of the urinary catheter balloon, while NO being a small uncharged
molecule does. Indeed we did detect large amounts of NO outside of the membrane and we

40
did observe bactericidal effects in the urine outside of the catheter balloon. Yet another
observation favouring NO as a candidate of the nitrite effect is that the pure NO-donor DETA
NONOate, had antibacterial effects in the same system. In addition, the antibacterial effect
and the amount of NO produced from acidified nitrite when tested in a Mueller-Hinton
medium instead of urine, were clearly decreased (data not shown). The exact mechanism
whereby NO and other products from acidified nitrite inhibit bacteria is still not settled but
multiple cellular targets are most certainly involved (Fig 9) including DNA, surface proteins
and key enzymes in the respiratory chain [167]. The effects of RNIs on DNA could be
interaction with DNA repair system as well as direct inactivation of ribonucleotide reductase
by NO [202].
Bacterial defense systems
Some urinary pathogens e.g. Proteus species have the capacity of producing ammonia from
urea thereby increasing urinary pH to levels where nitrite is not converted to toxic nitrogen
oxides. Once the higher pH has been established acidification of urine is more difficult [203].
This seems to be an effective mechanism developed by some pathogenic bacteria to protect
against acid/nitrite induced damage. Helicobacter pylori for example, has the unique ability to
thrive within the human stomach and has a urea-specific channel that opens as the pH of the
medium drops and cytoplasmatic urease is able to gain access to urea [204]. Obviously, as
shown in this thesis, when pH falls to very low levels (4.5 and below) urinary bacterial
growth is retarded even in the absence of nitrite indicating also direct effects of acidity itself.
At the pH levels more commonly found in mildly acidified urine (5-5.5) however, the effect
of pH itself was only minor and required the presence of nitrite/ascorbic acid for more
pronounced antibacterial effects (paper II-III).
An other bacterial defense mechanism found in many bacteria, including E. coli, is the flavo
haemoglobins, an ancient family of proteins that have been shown to effectively bind NO
thereby protecting the bacteria from nitrosative stress [186, 205]. Studies in E coli have also
demonstrated specific antioxidant regulons (soxRS regulon), which may protect bacteria
against NO when induced [187]. Indeed, E. coli with deficient soxRS locus, was more
sensitive to NO-dependent killing [206]. Another important defense alternative for E coli,
especially under low oxygen tensions, is the cytochrom c nitrite reductase (NrfA), a bacterial
enzyme that may further reduce NO to other less toxic nitrogen metabolites [207]. In addition,
many denitrifying bacteria, including Pseudomonas species, possess NO reductase enzymes
(NOR), which convert NO to nitrous oxide and ultimately to nitrogen [208]. There are data

41
showing that a mutation of NO reductase gene was lethal to Pseudomonas stutzeri during
anaerobic growth in the presence of nitrate or nitrite [209]. The pH optimum for the nitrate
reductase is high (about pH 8) and it probably operates less effectively at a low pH [210].
Obviously, it is of value for the bacteria not to reduce nitrate to nitrite under acidic conditions,
as nitrite will automatically convert to toxic nitrogen oxides thereby leading to self-
destruction. Other compounds that can help bacteria to detoxify NO or related compounds
include glutathione, homocysteine and superoxide dismutase [167]. Taken together there are
many different possible bacterial defense mechanisms against RNIs, but apparently they were
all insufficient in our experiments. One can speculate that it is the rapid kinetics of NO release
from acidified nitrite described in this thesis, that prevent the bacteria from up regulating
these defensive pathways.
Possible clinical usefulness
Treatment of UTI
Acidification of urine e.g. with vitamin C has been used as a household remedy for urinary
tract infections although the mechanism of action remains uncertain. We here speculate that
some of these effects are related to generation of bactericidal nitrogen oxides from urinary
nitrite. In this process Vitamin C has a dual action; it helps to acidify the urine and in addition
its intrinsic reducing capacity greatly enhances nitrite reduction. The reported clinical effects
of Vitamin C are however not very consistent and there may be several reasons for this. First,
vitamin C itself does not seem to be a very effective urinary acidifier and sometimes it
completely fails to reduce urinary pH [57-59]. In addition, in cases where acidification of
urine might have been effective, the nitrite concentration in urine may have been to low. As
shown here vitamin C itself had poor antibacterial effect and acidity itself did not have a
major influence on bacterial growth in urine. However, with the further addition of nitrite
bacteria were effectively killed in this medium. Therefore, to refine and improve the clinical
effects of vitamin C we probably need to create conditions with sufficient concentrations of
nitrite in urine in combination with a consistent and effective acidification of the urine. Is this
then possible to achieve in vivo? The urinary ascorbic acid concentration (10mM) used in this
thesis is physiologically achievable and in fact, similar to those obtained after ingestion of 1-2
gram of vitamin C daily [211]. Urinary pH normally varies between 5 and 7 and urinary pH
levels as low as 4.6 have been reported after treatment with drugs used for urinary
acidification [212]. Urinary nitrite in patients with nitrite-positive urinary tract infection

42
varies between 10-700 μM [213]. Thus, in some cases these amounts would be sufficient
provided that the urine is made acidic and contain enough ascorbic acid. In addition, we could
supplement the diet with nitrate to further increase substrate supply to the bacteria in the urine
as shown here. Already one hour after ingestion of 10 mg/kg sodium-nitrate (which
corresponds to what is found in about 300g spinach), nitrate levels in urine increased to a
concentration that is well above what the bacteria’s need to generate enough nitrite.
Now, knowing that sufficient amounts of nitrite, ascorbic acid and low urinary pH is
physiologically achievable. How can we accomplish this in the clinical setting? For this
concept to work we need first to obtain high nitrite levels in urine. Then, we probably need to
lower the urinary pH quite rapidly in order to convert the nitrite into toxic RNI`s. Thus, a
sequential two-steps procedure may be preferable. A medicinal product resulting in sufficient
urinary nitrate levels would be useful to start with. The dose of nitrate ingested and the time
interval between nitrate intake and acidification of the urine will probably be critical. In this
thesis (paper III) we used a pre-incubation period of 20 h in nitrate-rich urine. With this long
incubation time maximum amounts of nitrite are generated. For practical reasons a shorter
incubation time will be necessary in a clinical setting which may result in less nitrite
generated. A potential problem is therefore that nitrite levels could be to low to effectively kill
the bacteria when the urine is acidified. There are several ways of acidifying the urine, but
even if pH can be sufficiently lowered it will likely be important that this process is rather
rapid so that bacteria will not have time to up regulate defense mechanisms. In fact, studies
with ammonium chloride (50 mg/kg) showed a significant decrease in urinary pH (4.7-5.4)
already about 4 hours after a single dose [214]. Various other agents for acidifying the urine
have been used, these include mandelic acid, gluconic acid and methionine [52-55].
Another potential problem is that infected urine often has higher pH and therefore
acidification is probably more difficult. On the other hand, considering the potent antibacterial
effect of acidified nitrite, a decrease in urinary pH below a critical level during a limited
period, could be sufficient to inhibit bacterial growth if nitrite levels are sufficiently high. In
this thesis we used pooled urine from many different volunteers on a normal western diet. It is
likely that the antimicrobial activity of acidified nitrite in urine will vary because of different
composition of urine e.g. in relation to diet. Also, urinary osmolality may influence the
antibacterial effects of nitrite in acidified urine. One great advantage with the sequential
procedure described here compared to many traditional antibiotics is that the bacteria in the
gastro-intestinal tract are probably much less, if at all affected. On the other hand, a
disadvantage with the concept is that it will only work in bacteria with nitrate reductases.

43
Bacteria that do not reduce nitrate e.g. Enterococcus faecalis will not be sensitive to this
treatment similarly to the E. coli nitrate reductase mutant used in paper III.
Taken together, we have suggested a new concept for treatment of urinary tract infections
caused by nitrate-reducing bacteria. This involves a two-step procedure were bacteria are first
fed with nitrate followed some hours later by acidification of the urine. According to this
theory the bacteria self-destruct by producing nitrite which is later turned against them in
form of toxic RNI`s when urine is acidified. Clearly, controlled in vivo studies are needed to
evaluate the clinical potential for this antimicrobial regime and to test the timing of the
therapeutic interventions (nitrate intake, ascorbic acid intake and urinary acidification).
Prevention of CAUTI
CAUTI continues to be large problem in modern medical care and this condition is associated
with significant higher costs, morbidity and even higher mortality. Immense amount of work
has been done trying to prevent these infections, for example the development of silver alloy
catheters, catheters with salicylic acid impregnation, immersion of catheters in antibiotic
solution, closed drainage systems and even electrification of catheters [70, 77, 78]. Despite
these efforts CAUTI continues to be a major problem in modern health care.
In this thesis (paper IV) we describe a new concept, which allows for direct intravesical
delivery of antibacterial gases into the urinary bladder via the catheter retention balloon. It is
tempting to speculate that this regime could be useful to treat and to prevent CAUTI`s. Is
then this concept applicable in vivo? Again a key question is if the in vitro concentrations of
nitrite, ascorbic acid and the pH used in this set up are effective in vivo also without causing
troublesome side-effects. The concentrations of ascorbic acid (10 mM) used here were the
same as described above. Natural acidification of salivary nitrite in the stomach, continuously
exposes the gastric mucosa to similar amounts of nitrogen oxides. Furthermore, a nitrite
concentration of about 3 mM, which is similar to what we used in this set up, is allowed as an
ingredient in meat curing. Thus, these antibacterial compounds are expected to have fairly
low acute toxicity to host cells in the concentrations used here. Whether a prolonged exposure
of the bladder to these RNSs could favour formation of nitrosamines, a group of potentially
carcinogenic chemicals, must indeed be further studied. However, the addition of ascorbic
acid as one component in the retention balloon likely inhibits the total formation of
nitrosamines [215]. The urinary pH in the bladder-model was not influenced by the acidic
solution in the intact catheter balloon. Even if the balloon broke by an accident, the small
amount (10ml) of solution with pH 2.5 would likely have low acute toxicity to the bladder

44
mucosa since probably at least some amount of residual urine is present thereby diluting and
buffering the acidic solution.
In paper IV we examined two different strains of bacteria. Although not studied here, it is
likely that also other urinary tract pathogens are sensitive to this procedure. Thus, from the
results presented in this thesis it is known that RNSs generated from acidified nitrite have
anti-bacterial effects against several of the most common urinary pathogens including also P.
aeruginosa and S. saprophyticus.
A potential advantage with this regime is that NO will likely diffuse easily and rapidly into
the biofilm generated by the bacteria. This biofilm normally provides a protective
environment for the microorganisms and thereby making them less susceptible to
antimicrobial agents. Naturally, this assumption has to be tested. Many other questions remain
to answer before we can consider using this method in the clinic. For example we need to
determine the durability of the antibacterial effect as well as, the exact dosing of nitrite and
ascorbic acid and the risk of bacterial resistance. Nevertheless, this simple concept of
substituting the content of the retention balloon of a urinary catheter with an active diffusible
drug instead of normal saline, is an attractive new alternative to the existing methods used to
treat and prevent CAUTI`s. With this novel method one could easily administrate repeated
individual doses of the active drug at chosen intervals without disturbing the integrity of the
closed drainage system. Considering the fact that CAUTIs are still very common and result in
substantially increased morbidity and health care costs, new approaches for their prevention
are highly warranted.

45
CONCLUSIONS
• Large amounts of NO were formed in mildly acidified nitrite-containing infected
human urine and in acidified control urine after addition of nitrite. This NO formation
was strongly pH dependent and was greatly increased by the addition of ascorbic acid.
• The growth of E. coli, P. aeruginosa and S. saprophyticus was dose-dependently
inhibited after two hours of incubation in mildly acidified nitrite-containing urine.
This bacteriostatic effect was greater at lower pH and was markedly potentiated by
ascorbic acid.
• Prolonged exposure of to physiological achievable concentrations of sodium nitrite
plus ascorbic acid resulted in bactericidal effects. In comparison with conventional
antibiotics tested in this same model nitrite seemed to be at least equally potent.
• A two-step procedure where E coli. was first fed with nitrate followed later by
acidification of the urine, resulted in effective bacterial killing. This killing was
dependent on the nitrite generated by the bacteria themselves.
• A novel concept for intravesical delivery of NO via a catheter retention balloon was
tested in vitro. The urinary pathogen E. coli was effectively killed by this procedure.

46
ACKNOWLEDGEMENTS
I wish to express my sincere gratitude to all those who have helped and supported me throughout these studies. I especially thank:
Professor Jon Lundberg, my brilliant tutor, for helping me during all phases of my work and for your countless numbers of inspiring and innovative “pilot-experiments”. For giving to me the huge responsibility to continue the “urine track” from your dissertation, for your excellent scientific guidance in combination with always being enthusiastic and encouraging. For your great patience when I during some periods did other important things in life such as being on paternity leave and practising robotic surgery. Professor Peter Wiklund, my co-tutor and chairman at my department, for introducing me to the fascinating field of nitric oxide research. For your unique triad of qualities; being an excellent scientist, an extraordinary talented surgeon and having a great personality. For always having answers and moving forwards. For encouraging me to give lectures in front of many people. For teaching me robotic surgery and for never switching off your mobile phone. My co-authors Mirco Govoni, Eddie Weitzberg Lars Engstrand, Edward Morcos for stimulating discussions and enjoyable collaboration. Peter Ekman, for supporting me to do science, for being an excellent speaker and for giving us at the Urology Department inspiring scientific summaries (veckobrev) from all over the world. Lars Norlen, for being such a great support in all kinds of clinical problems, for being a very skillful surgeon with an extraordinary talent for teaching this noble art and for employing me at the clinic. Ann-Charlotte Kinn and Jan Sehlin, for being my tutors during my residency at the Department of Urology and for your support and excellent clinical guidance. My lab-colleagues: Daniel Törnberg, Håkan Björne, Helena Marteus, Jörgen Palm, Max Herulf, Wilhelm Zetterquist, Emmelie Jansson, Claudia Reinders, Tanja Sobko, Helen Sylvin Roelinde Middelveld, Katarina Hallen, and Nasrin Bavand, for making work so enjoyable. My former and present colleagues at the Urology Department involved in the NO-research group, Ingrid Ehren, Abbi Hosseini, Olof Jansson and Lotta Renström-Koskela, for support and advice during international scientific conferences. Margareta Stensdotter and Carina Nihlen, for excellent technical assistance. Carina, I would never have been able to finish my thesis in 2005 without you! Torgny Kolmert, for taking special good care of me when I nine years ago did my first attempts in Urology procedures.

47
My colleagues and excellent robotic assistants Andreas Nilsson, Martin Jonsson, Andreas Thorstensson and Ramin Gaffarpour, for all the fun we have together, for taking very good care of my robotic-patients before and after surgery, making it possible for me to write this thesis. Guys, I will repay when you write your thesis! My colleagues in the prostate-cancer group, Stephan Brändstedt, Leif Haendler, Magnus Hellström and David Kudren and my other colleagues at the clinic Pelle Larsson, Elisabeth Farrelly, Daniela Volz, Marianne Brehmer and Eric Borgström, for support and understanding. Berit Björk and Annika Broström, for being helpful with administrative matters and support. Marie Dahlgren, for always helping me in the best way with administrative matters concerning salary and vacation (Marie, you know how much I hate filling in forms!) Carl-Eric and Sonja, my parents for your love and constant support in everything I have been doing in my life and for your indispensable help with the children. Sudha, my sister for being there and caring for every one around you and always supporting me and my family. Gunnar and Aase, for being the best parents-in-law you could ever dream of. Lars, my neighbour for “magic computer-skills” when my computer crashed, some weeks before printing deadline of my thesis. Finally, I would like to thank Anne, my wife and my “dream-team” friend for endless amounts of patience and love. Emma, Adam and Fabian, our wonderful children for being a constant source of joy in life.

48
REFERENCES 1. Walsh R, Vaughan, Wein, Campbell´s Urology. 2002. Eight Edition: p. 516. 2. Stamm WE and SR Norrby, Urinary tract infections: disease panorama and
challenges. J Infect Dis, 2001. 183 Suppl 1: p. S1-4. 3. Kunin CM, Urinary tract infections in females. Clin Infect Dis, 1994. 18(1): p. 1-10;
quiz 11-2. 4. Foxman B and P Brown, Epidemiology of urinary tract infections: transmission and
risk factors, incidence, and costs. Infect Dis Clin North Am, 2003. 17(2): p. 227-41. 5. Finer G and D Landau, Pathogenesis of urinary tract infections with normal female
anatomy. Lancet Infect Dis, 2004. 4(10): p. 631-5. 6. Craig JC, Urinary tract infection: new perspectives on a common disease. Curr Opin
Infect Dis, 2001. 14(3): p. 309-13. 7. Rubin RH, ED Shapiro, VT Andriole, RJ Davis, and WE Stamm, Evaluation of
new anti-infective drugs for the treatment of urinary tract infection. Infectious Diseases Society of America and the Food and Drug Administration. Clin Infect Dis, 1992. 15 Suppl 1: p. S216-27.
8. Sobel JD, New aspects of pathogenesis of lower urinary tract infections. Urology, 1985. 26(5 Suppl): p. 11-6.
9. Schlager TA, Urinary tract infections in children younger than 5 years of age: epidemiology, diagnosis, treatment, outcomes and prevention. Paediatr Drugs, 2001. 3(3): p. 219-27.
10. Walsh R, Vaughan, Wein, Campbell´s Urology. 2002. Eight Edition: p. 522. 11. Hooton TM, Pathogenesis of urinary tract infections: an update. J Antimicrob
Chemother, 2000. 46 Suppl 1: p. 1-7; discussion 63-5. 12. Hooton TM, Practice guidelines for urinary tract infection in the era of managed care.
Int J Antimicrob Agents, 1999. 11(3-4): p. 241-5; discussion 261-4. 13. Kahlmeter G, The ECO.SENS Project: a prospective, multinational, multicentre
epidemiological survey of the prevalence and antimicrobial susceptibility of urinary tract pathogens--interim report. J Antimicrob Chemother, 2000. 46 Suppl 1: p. 15-22; discussion 63-5.
14. Ronald A, The etiology of urinary tract infection: traditional and emerging pathogens. Dis Mon, 2003. 49(2): p. 71-82.
15. Schaeffer AJ, JM Jones, and JK Dunn, Association of vitro Escherichia coli adherence to vaginal and buccal epithelial cells with susceptibility of women to recurrent urinary-tract infections. N Engl J Med, 1981. 304(18): p. 1062-6.
16. Yamamoto S, T Tsukamoto, A Terai, H Kurazono, Y Takeda, and O Yoshida, Genetic evidence supporting the fecal-perineal-urethral hypothesis in cystitis caused by Escherichia coli. J Urol, 1997. 157(3): p. 1127-9.
17. Johnson JR, Microbial virulence determinants and the pathogenesis of urinary tract infection. Infect Dis Clin North Am, 2003. 17(2): p. 261-78, viii.
18. Sandberg T, B Kaijser, G Lidin-Janson, K Lincoln, F Orskov, I Orskov, E Stokland, and C Svanborg-Eden, Virulence of Escherichia coli in relation to host factors in women with symptomatic urinary tract infection. J Clin Microbiol, 1988. 26(8): p. 1471-6.
19. Tseng CC, JJ Wu, HL Liu, JM Sung, and JJ Huang, Roles of host and bacterial virulence factors in the development of upper urinary tract infection caused by Escherichia coli. Am J Kidney Dis, 2002. 39(4): p. 744-52.

49
20. Gordon DM and MA Riley, A theoretical and experimental analysis of bacterial growth in the bladder. Mol Microbiol, 1992. 6(4): p. 555-62.
21. Kaye D, Host defense mechanisms in the urinary tract. Urol Clin North Am, 1975. 2(3): p. 407-22.
22. Asscher AW, M Sussman, WE Waters, RH Davis, and S Chick, Urine as a medium for bacterial growth. Lancet, 1966. 2(7472): p. 1037-41.
23. Dontas AS and P Kasviki-Charvati, Significance of diuresis-provoked bacteriuria. J Infect Dis, 1976. 134(2): p. 174-80.
24. Hadas H, O Medalia, and M Aronson, Differential susceptibility of Escherichia coli and Proteus mirabilis to mouse urine and to urea. J Infect Dis, 1977. 136(1): p. 100-3.
25. Wullt B, G Bergsten, H Fischer, G Godaly, D Karpman, I Leijonhufvud, AC Lundstedt, P Samuelsson, M Samuelsson, ML Svensson, and C Svanborg, The host response to urinary tract infection. Infect Dis Clin North Am, 2003. 17(2): p. 279-301.
26. Svanborg C, G Bergsten, H Fischer, B Frendeus, G Godaly, E Gustafsson, L Hang, M Hedlund, D Karpman, AC Lundstedt, M Samuelsson, P Samuelsson, M Svensson, and B Wullt, The 'innate' host response protects and damages the infected urinary tract. Ann Med, 2001. 33(9): p. 563-70.
27. Schilling JD, MA Mulvey, and SJ Hultgren, Dynamic interactions between host and pathogen during acute urinary tract infections. Urology, 2001. 57(6 Suppl 1): p. 56-61.
28. Eden CS, R Freter, L Hagberg, R Hull, S Hull, H Leffler, and G Schoolnik, Inhibition of experimental ascending urinary tract infection by an epithelial cell-surface receptor analogue. Nature, 1982. 298(5874): p. 560-2.
29. Redondo-Lopez V, RL Cook, and JD Sobel, Emerging role of lactobacilli in the control and maintenance of the vaginal bacterial microflora. Rev Infect Dis, 1990. 12(5): p. 856-72.
30. Reid G and AW Bruce, Low vaginal pH and urinary-tract infection. Lancet, 1995. 346(8991-8992): p. 1704.
31. Foxman B and RR Frerichs, Epidemiology of urinary tract infection: I. Diaphragm use and sexual intercourse. Am J Public Health, 1985. 75(11): p. 1308-13.
32. Kass EH, Asymptomatic infections of the urinary tract. Trans Assoc Am Physicians, 1956. 69: p. 56-64.
33. Stamm WE, GW Counts, KR Running, S Fihn, M Turck, and KK Holmes, Diagnosis of coliform infection in acutely dysuric women. N Engl J Med, 1982. 307(8): p. 463-8.
34. Johnson JR and WE Stamm, Diagnosis and treatment of acute urinary tract infections. Infect Dis Clin North Am, 1987. 1(4): p. 773-91.
35. L.T. HAM, The nitrite reaction as an indicator of urinary infection. J Urology, 1929. 22: p. 521.
36. Nickel JC, Management of urinary tract infections: historical perspective and current strategies: Part 2--Modern management. J Urol, 2005. 173(1): p. 27-32.
37. James GP, KL Paul, and JB Fuller, Urinary nitrite and urinary-tract infection. Am J Clin Pathol, 1978. 70(4): p. 671-8.
38. Soler Rodriguez F, MP Miguez Santiyan, and JD Pedrera Zamorano, Evaluation of reagent strips for the rapid diagnosis of nitrite poisoning. J Anal Toxicol, 1992. 16(1): p. 63-5.
39. Lohr JA, Use of routine urinalysis in making a presumptive diagnosis of urinary tract infection in children. Pediatr Infect Dis J, 1991. 10(9): p. 646-50.
40. Wathne B, B Hovelius, and PA Mardh, Causes of frequency and dysuria in women. Scand J Infect Dis, 1987. 19(2): p. 223-9.

50
41. Powell HR, DA McCredie, and MA Ritchie, Urinary nitrite in symptomatic and asymptomatic urinary infection. Arch Dis Child, 1987. 62(2): p. 138-40.
42. Kaji S, [Experimental study on the role of bacteria from urinary tract infections on the bladder carcinogenesis]. Fukuoka Igaku Zasshi, 1994. 85(10): p. 294-308.
43. Deville WL, JC Yzermans, NP van Duijn, PD Bezemer, DA van der Windt, and LM Bouter, The urine dipstick test useful to rule out infections. A meta-analysis of the accuracy. BMC Urol, 2004. 4(1): p. 4.
44. Warren JW, E Abrutyn, JR Hebel, JR Johnson, AJ Schaeffer, and WE Stamm, Guidelines for antimicrobial treatment of uncomplicated acute bacterial cystitis and acute pyelonephritis in women. Infectious Diseases Society of America (IDSA). Clin Infect Dis, 1999. 29(4): p. 745-58.
45. Nicolle LE, Pivmecillinam in the treatment of urinary tract infections. J Antimicrob Chemother, 2000. 46 Suppl 1: p. 35-9; discussion 63-5.
46. Hooton TM, Fluoroquinolones and resistance in the treatment of uncomplicated urinary tract infection. Int J Antimicrob Agents, 2003. 22 Suppl 2: p. 65-72.
47. Lundbergh P, Clinical pratice guidelines developed by STRAMA-group in Stockholm: Urinary tract infections in primary care. Läkartidningen, 2001. 98: p. 4198-4201.
48. Norrby SR, Short-term treatment of uncomplicated lower urinary tract infections in women. Rev Infect Dis, 1990. 12(3): p. 458-67.
49. Molstad S, LG Burman, and S Ferry, [Uncomplicated lower urinary tract infection: 3-day therapy is recommended!]. Lakartidningen, 1999. 96(39): p. 4212-4.
50. Wise R, T Hart, O Cars, M Streulens, R Helmuth, P Huovinen, and M Sprenger, Antimicrobial resistance. Is a major threat to public health. Bmj, 1998. 317(7159): p. 609-10.
51. Gupta K, Emerging antibiotic resistance in urinary tract pathogens. Infect Dis Clin North Am, 2003. 17(2): p. 243-59.
52. Murphy FJ, S Zelman, and W Mau, Ascorbic acid as a urinary acidifying agent. 2. Its adjunctive role in chronic urinary infection. J Urol, 1965. 94(3): p. 300-3.
53. McDonald DF MG, Bacteriostatic and acidifying effects of methionine, hydrolyzed casein, and ascorbic acid on the urine. N Engl J Med, 1959. 261: p. 803-806.
54. Rotoski, Ueber den bactericiden einfluss der acidität des harus auf die cystitserreger. Dtsch. Med. Wschr., 1898. 24: p. 235-236 and 249-252.
55. Clark, Escherichia coli bacilluria under ketogenic treatment. Proc. Staff Meet.,Mayo Clin., 1931. 6: p. 605-608.
56. Quinn EL aKEE, Principles in the long Term Management of Chronic infection of the Urinary Tract. Biology of Pyelonephritis, Boston little, Brown and Co., 1960.: p. 663-672.
57. Barton CH SM, Thomas R, Vaziri ND, Byrne C and Ryan G, Ineffectiveness of Intravenous Ascorbic Acid as an Acidifying Agent in Man. Arch Intern Med, 1981. 141.: p. 211-212.
58. Nahata MC SL, lampman T, et al, Effects of ascorbic acid on urine pH in man. Am j Hosp Pharmacol, 1977. 34: p. 1234-1237.
59. Travis L DW, Mintz A and Assemi M, Urinary acidification with ascorbic acid. The Journal of Pedriatrics, 1965.: p. 1176-1178.
60. Lee B, T Bhuta, J Craig, and J Simpson, Methenamine hippurate for preventing urinary tract infections. Cochrane Database Syst Rev, 2002(1): p. CD003265.
61. Avorn J, M Monane, JH Gurwitz, RJ Glynn, I Choodnovskiy, and LA Lipsitz, Reduction of bacteriuria and pyuria after ingestion of cranberry juice. Jama, 1994. 271(10): p. 751-4.

51
62. Howell AB, Cranberry proanthocyanidins and the maintenance of urinary tract health. Crit Rev Food Sci Nutr, 2002. 42(3 Suppl): p. 273-8.
63. Foo LY, Y Lu, AB Howell, and N Vorsa, The structure of cranberry proanthocyanidins which inhibit adherence of uropathogenic P-fimbriated Escherichia coli in vitro. Phytochemistry, 2000. 54(2): p. 173-81.
64. Jepson RG, L Mihaljevic, and J Craig, Cranberries for preventing urinary tract infections. Cochrane Database Syst Rev, 2001(3): p. CD001321.
65. Krieger JN, DL Kaiser, and RP Wenzel, Urinary tract etiology of bloodstream infections in hospitalized patients. J Infect Dis, 1983. 148(1): p. 57-62.
66. Richards MJ, JR Edwards, DH Culver, and RP Gaynes, Nosocomial infections in medical intensive care units in the United States. National Nosocomial Infections Surveillance System. Crit Care Med, 1999. 27(5): p. 887-92.
67. Maki DG and PA Tambyah, Engineering out the risk for infection with urinary catheters. Emerg Infect Dis, 2001. 7(2): p. 342-7.
68. Haley RW, TM Hooton, DH Culver, RC Stanley, TG Emori, CD Hardison, D Quade, RH Shachtman, DR Schaberg, BV Shah, and GD Schatz, Nosocomial infections in U.S. hospitals, 1975-1976: estimated frequency by selected characteristics of patients. Am J Med, 1981. 70(4): p. 947-59.
69. Daifuku R and WE Stamm, Association of rectal and urethral colonization with urinary tract infection in patients with indwelling catheters. Jama, 1984. 252(15): p. 2028-30.
70. Cormio L, P La Forgia, D La Forgia, A Siitonen, and M Ruutu, Bacterial adhesion to urethral catheters: role of coating materials and immersion in antibiotic solution. Eur Urol, 2001. 40(3): p. 354-8; discussion 359.
71. Stamm WE, Catheter-associated urinary tract infections: epidemiology, pathogenesis, and prevention. Am J Med, 1991. 91(3B): p. 65S-71S.
72. Donlan RM, Biofilm formation: a clinically relevant microbiological process. Clin Infect Dis, 2001. 33(8): p. 1387-92.
73. Warren JW, Catheter-associated urinary tract infections. Infect Dis Clin North Am, 1997. 11(3): p. 609-22.
74. Pratt RJ, C Pellowe, HP Loveday, N Robinson, GW Smith, S Barrett, P Davey, P Harper, C Loveday, C McDougall, A Mulhall, S Privett, C Smales, L Taylor, B Weller, and M Wilcox, The epic project: developing national evidence-based guidelines for preventing healthcare associated infections. Phase I: Guidelines for preventing hospital-acquired infections. Department of Health (England). J Hosp Infect, 2001. 47 Suppl: p. S3-82.
75. Saint S, JG Elmore, SD Sullivan, SS Emerson, and TD Koepsell, The efficacy of silver alloy-coated urinary catheters in preventing urinary tract infection: a meta-analysis. Am J Med, 1998. 105(3): p. 236-41.
76. Plowman R, N Graves, J Esquivel, and JA Roberts, An economic model to assess the cost and benefits of the routine use of silver alloy coated urinary catheters to reduce the risk of urinary tract infections in catheterized patients. J Hosp Infect, 2001. 48(1): p. 33-42.
77. Farber BF and AG Wolff, The use of salicylic acid to prevent the adherence of Escherichia coli to silastic catheters. J Urol, 1993. 149(3): p. 667-70.
78. Shafik A, The electrified catheter. Role in sterilizing urine and decreasing bacteriuria. World J Urol, 1993. 11(3): p. 183-5.
79. Sethia KK, JB Selkon, AR Berry, CM Turner, MG Kettlewell, and MH Gough, Prospective randomized controlled trial of urethral versus suprapubic catheterization. Br J Surg, 1987. 74(7): p. 624-5.

52
80. Jamil F, Towards a catheter free status in neurogenic bladder dysfunction: a review of bladder management options in spinal cord injury (SCI). Spinal Cord, 2001. 39(7): p. 355-61.
81. Platt R, BF Polk, B Murdock, and B Rosner, Mortality associated with nosocomial urinary-tract infection. N Engl J Med, 1982. 307(11): p. 637-42.
82. Saint S, Clinical and economic consequences of nosocomial catheter-related bacteriuria. Am J Infect Control, 2000. 28(1): p. 68-75.
83. Furchgott RF and JV Zawadzki, The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature, 1980. 288(5789): p. 373-6.
84. Katsuki S, W Arnold, C Mittal, and F Murad, Stimulation of guanylate cyclase by sodium nitroprusside, nitroglycerin and nitric oxide in various tissue preparations and comparison to the effects of sodium azide and hydroxylamine. J Cyclic Nucleotide Res, 1977. 3(1): p. 23-35.
85. Rapoport RM, MB Draznin, and F Murad, Endothelium-dependent relaxation in rat aorta may be mediated through cyclic GMP-dependent protein phosphorylation. Nature, 1983. 306(5939): p. 174-6.
86. Katsuki S, WP Arnold, and F Murad, Effects of sodium nitroprusside, nitroglycerin, and sodium azide on levels of cyclic nucleotides and mechanical activity of various tissues. J Cyclic Nucleotide Res, 1977. 3(4): p. 239-47.
87. Palmer RM, AG Ferrige, and S Moncada, Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature, 1987. 327(6122): p. 524-6.
88. Ignarro LJ, GM Buga, KS Wood, RE Byrns, and G Chaudhuri, Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc Natl Acad Sci U S A, 1987. 84(24): p. 9265-9.
89. Leone AM, RM Palmer, RG Knowles, PL Francis, DS Ashton, and S Moncada, Constitutive and inducible nitric oxide synthases incorporate molecular oxygen into both nitric oxide and citrulline. J Biol Chem, 1991. 266(35): p. 23790-5.
90. Palmer RM and S Moncada, A novel citrulline-forming enzyme implicated in the formation of nitric oxide by vascular endothelial cells. Biochem Biophys Res Commun, 1989. 158(1): p. 348-52.
91. Knowles RG and S Moncada, Nitric oxide synthases in mammals. Biochem J, 1994. 298 ( Pt 2): p. 249-58.
92. Alderton WK, CE Cooper, and RG Knowles, Nitric oxide synthases: structure, function and inhibition. Biochem J, 2001. 357(Pt 3): p. 593-615.
93. Knowles RG, M Palacios, RM Palmer, and S Moncada, Formation of nitric oxide from L-arginine in the central nervous system: a transduction mechanism for stimulation of the soluble guanylate cyclase. Proc Natl Acad Sci U S A, 1989. 86(13): p. 5159-62.
94. Mayer B, K Schmidt, P Humbert, and E Bohme, Biosynthesis of endothelium-derived relaxing factor: a cytosolic enzyme in porcine aortic endothelial cells Ca2+-dependently converts L-arginine into an activator of soluble guanylyl cyclase. Biochem Biophys Res Commun, 1989. 164(2): p. 678-85.
95. Stuehr DJ, NS Kwon, SS Gross, BA Thiel, R Levi, and CF Nathan, Synthesis of nitrogen oxides from L-arginine by macrophage cytosol: requirement for inducible and constitutive components. Biochem Biophys Res Commun, 1989. 161(2): p. 420-6.
96. Tayeh MA and MA Marletta, Macrophage oxidation of L-arginine to nitric oxide, nitrite, and nitrate. Tetrahydrobiopterin is required as a cofactor. J Biol Chem, 1989. 264(33): p. 19654-8.

53
97. Benjamin N, F O'Driscoll, H Dougall, C Duncan, L Smith, M Golden, and H McKenzie, Stomach NO synthesis. Nature, 1994. 368(6471): p. 502.
98. Lundberg JO, E Weitzberg, JM Lundberg, and K Alving, Intragastric nitric oxide production in humans: measurements in expelled air. Gut, 1994. 35(11): p. 1543-6.
99. Duncan C, H Dougall, P Johnston, S Green, R Brogan, C Leifert, L Smith, M Golden, and N Benjamin, Chemical generation of nitric oxide in the mouth from the enterosalivary circulation of dietary nitrate. Nat Med, 1995. 1(6): p. 546-51.
100. Weitzberg E and JO Lundberg, Nonenzymatic nitric oxide production in humans. Nitric Oxide, 1998. 2(1): p. 1-7.
101. Bartsch H, H Ohshima, and B Pignatelli, Inhibitors of endogenous nitrosation. Mechanisms and implications in human cancer prevention. Mutat Res, 1988. 202(2): p. 307-24.
102. Zweier JL, P Wang, A Samouilov, and P Kuppusamy, Enzyme-independent formation of nitric oxide in biological tissues. Nat Med, 1995. 1(8): p. 804-9.
103. Weller R, S Pattullo, L Smith, M Golden, A Ormerod, and N Benjamin, Nitric oxide is generated on the skin surface by reduction of sweat nitrate. J Invest Dermatol, 1996. 107(3): p. 327-31.
104. Benjamin N, S Pattullo, R Weller, L Smith, and A Ormerod, Wound licking and nitric oxide. Lancet, 1997. 349(9067): p. 1776.
105. Lundberg JO, E Weitzberg, JA Cole, and N Benjamin, Nitrate, bacteria and human health. Nat Rev Microbiol, 2004. 2(7): p. 593-602.
106. Gladwin MT, Haldane, hot dogs, halitosis, and hypoxic vasodilation: the emerging biology of the nitrite anion. J Clin Invest, 2004. 113(1): p. 19-21.
107. Zhang Z, DP Naughton, DR Blake, N Benjamin, CR Stevens, PG Winyard, MC Symons, and R Harrison, Human xanthine oxidase converts nitrite ions into nitric oxide (NO). Biochem Soc Trans, 1997. 25(3): p. 524S.
108. Millar TM, CR Stevens, N Benjamin, R Eisenthal, R Harrison, and DR Blake, Xanthine oxidoreductase catalyses the reduction of nitrates and nitrite to nitric oxide under hypoxic conditions. FEBS Lett, 1998. 427(2): p. 225-8.
109. Li H, A Samouilov, X Liu, and JL Zweier, Characterization of the magnitude and kinetics of xanthine oxidase-catalyzed nitrate reduction: evaluation of its role in nitrite and nitric oxide generation in anoxic tissues. Biochemistry, 2003. 42(4): p. 1150-9.
110. Kozlov AV, B Dietrich, and H Nohl, Various intracellular compartments cooperate in the release of nitric oxide from glycerol trinitrate in liver. Br J Pharmacol, 2003. 139(5): p. 989-97.
111. Cosby K, KS Partovi, JH Crawford, RP Patel, CD Reiter, S Martyr, BK Yang, MA Waclawiw, G Zalos, X Xu, KT Huang, H Shields, DB Kim-Shapiro, AN Schechter, RO Cannon, 3rd, and MT Gladwin, Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med, 2003. 9(12): p. 1498-505.
112. Adelon, Dictionnaire Abrege des Sciences Medicales.Panckoucke, Paris., 1824. 113. N.M. EGBaK, Transient methemoglobineia following administration of ammonium
nitrate. Medical Clinics of North America, 1929. 12: p. 1489-1496. 114. Mangione NJ and SP Glasser, Phenomenon of nitrate tolerance. Am Heart J, 1994.
128(1): p. 137-46. 115. J.M. MPNaB, The production of malignant primary hepatic tumours in the rat by
feeding dimethylnitrosamine. British Journal of Cancer, 1956. 10: p. 114-122. 116. Archer MC, Mechanisms of action of N-nitroso compounds. Cancer Surv, 1989.
8(2): p. 241-50.

54
117. Forman D, S Al-Dabbagh, and R Doll, Nitrates, nitrites and gastric cancer in Great Britain. Nature, 1985. 313(6004): p. 620-5.
118. Knight TM, D Forman, R Pirastu, P Comba, R Iannarilli, PL Cocco, G Angotzi, E Ninu, and S Schierano, Nitrate and nitrite exposure in Italian populations with different gastric cancer rates. Int J Epidemiol, 1990. 19(3): p. 510-5.
119. McKnight GM, CW Duncan, C Leifert, and MH Golden, Dietary nitrate in man: friend or foe? Br J Nutr, 1999. 81(5): p. 349-58.
120. van Loon AJ, AA Botterweck, RA Goldbohm, HA Brants, JD van Klaveren, and PA van den Brandt, Intake of nitrate and nitrite and the risk of gastric cancer: a prospective cohort study. Br J Cancer, 1998. 78(1): p. 129-35.
121. Dejam A, CJ Hunter, AN Schechter, and MT Gladwin, Emerging role of nitrite in human biology. Blood Cells Mol Dis, 2004. 32(3): p. 423-9.
122. Ysart G, P Miller, G Barrett, D Farrington, P Lawrance, and N Harrison, Dietary exposures to nitrate in the UK. Food Addit Contam, 1999. 16(12): p. 521-32.
123. Pegg RB and F Shahidi, Unraveling the chemical identity of meat pigments. Crit Rev Food Sci Nutr, 1997. 37(6): p. 561-89.
124. Pierson MD and LA Smoot, Nitrite, nitrite alternatives, and the control of Clostridium botulinum in cured meats. Crit Rev Food Sci Nutr, 1982. 17(2): p. 141-87.
125. Archer DL, Evidence that ingested nitrate and nitrite are beneficial to health. J Food Prot, 2002. 65(5): p. 872-5.
126. Rhodes P, AM Leone, PL Francis, AD Struthers, S Moncada, and PM Rhodes, The L-arginine:nitric oxide pathway is the major source of plasma nitrite in fasted humans. Biochem Biophys Res Commun, 1995. 209(2): p. 590-6.
127. McKnight GM, LM Smith, RS Drummond, CW Duncan, M Golden, and N Benjamin, Chemical synthesis of nitric oxide in the stomach from dietary nitrate in humans. Gut, 1997. 40(2): p. 211-4.
128. Tannenbaum SR, M Weisman, and D Fett, The effect of nitrate intake on nitrite formation in human saliva. Food Cosmet Toxicol, 1976. 14(6): p. 549-52.
129. Spiegelhalder B, G Eisenbrand, and R Preussmann, Influence of dietary nitrate on nitrite content of human saliva: possible relevance to in vivo formation of N-nitroso compounds. Food Cosmet Toxicol, 1976. 14(6): p. 545-8.
130. Walker R, The metabolism of dietary nitrites and nitrates. Managing risks of nitrates to humans and the environment. The Royal Society of Chemistry, Cambridge, UK., 1999: p. 250-258.
131. Wagner DA, VR Young, SR Tannenbaum, DS Schultz, and WM Deen, Mammalian nitrate biochemistry: metabolism and endogenous synthesis. IARC Sci Publ, 1984(57): p. 247-53.
132. Thomas DD, X Liu, SP Kantrow, and JR Lancaster, Jr., The biological lifetime of nitric oxide: implications for the perivascular dynamics of NO and O2. Proc Natl Acad Sci U S A, 2001. 98(1): p. 355-60.
133. Chung SJ and HL Fung, Identification of the subcellular site for nitroglycerin metabolism to nitric oxide in bovine coronary smooth muscle cells. J Pharmacol Exp Ther, 1990. 253(2): p. 614-9.
134. Wallace JL and MJ Miller, Nitric oxide in mucosal defense: a little goes a long way. Gastroenterology, 2000. 119(2): p. 512-20.
135. Beckman JS, TW Beckman, J Chen, PA Marshall, and BA Freeman, Apparent hydroxyl radical production by peroxynitrite: implications for endothelial injury from nitric oxide and superoxide. Proc Natl Acad Sci U S A, 1990. 87(4): p. 1620-4.
136. Beckman JS and WH Koppenol, Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. Am J Physiol, 1996. 271(5 Pt 1): p. C1424-37.

55
137. Ignarro LJ, JM Fukuto, JM Griscavage, NE Rogers, and RE Byrns, Oxidation of nitric oxide in aqueous solution to nitrite but not nitrate: comparison with enzymatically formed nitric oxide from L-arginine. Proc Natl Acad Sci U S A, 1993. 90(17): p. 8103-7.
138. Marletta MA, Nitric oxide synthase: aspects concerning structure and catalysis. Cell, 1994. 78(6): p. 927-30.
139. Goretski J and TC Hollocher, Trapping of nitric oxide produced during denitrification by extracellular hemoglobin. J Biol Chem, 1988. 263(5): p. 2316-23.
140. Foster MW, TJ McMahon, and JS Stamler, S-nitrosylation in health and disease. Trends Mol Med, 2003. 9(4): p. 160-8.
141. Grisham MB, D Jourd'Heuil, and DA Wink, Nitric oxide. I. Physiological chemistry of nitric oxide and its metabolites:implications in inflammation. Am J Physiol, 1999. 276(2 Pt 1): p. G315-21.
142. Fukuto JM, Chemistry of nitric oxide: biologically relevant aspects. Adv Pharmacol, 1995. 34: p. 1-15.
143. Modin A, H Bjorne, M Herulf, K Alving, E Weitzberg, and JO Lundberg, Nitrite-derived nitric oxide: a possible mediator of 'acidic-metabolic' vasodilation. Acta Physiol Scand, 2001. 171(1): p. 9-16.
144. Holmquist F, H Hedlund, and KE Andersson, Characterization of inhibitory neurotransmission in the isolated corpus cavernosum from rabbit and man. J Physiol, 1992. 449: p. 295-311.
145. Ignarro LJ, PA Bush, GM Buga, KS Wood, JM Fukuto, and J Rajfer, Nitric oxide and cyclic GMP formation upon electrical field stimulation cause relaxation of corpus cavernosum smooth muscle. Biochem Biophys Res Commun, 1990. 170(2): p. 843-50.
146. Goldstein I, TF Lue, H Padma-Nathan, RC Rosen, WD Steers, and PA Wicker, Oral sildenafil in the treatment of erectile dysfunction. Sildenafil Study Group. N Engl J Med, 1998. 338(20): p. 1397-404.
147. Ho MH, NN Bhatia, and O Khorram, Physiologic role of nitric oxide and nitric oxide synthase in female lower urinary tract. Curr Opin Obstet Gynecol, 2004. 16(5): p. 423-9.
148. Ehren I, H Iversen, O Jansson, J Adolfsson, and NP Wiklund, Localization of nitric oxide synthase activity in the human lower urinary tract and its correlation with neuroeffector responses. Urology, 1994. 44(5): p. 683-7.
149. Moncada S and A Higgs, The L-arginine-nitric oxide pathway. N Engl J Med, 1993. 329(27): p. 2002-12.
150. Ignarro LJ, Endothelium-derived nitric oxide: actions and properties. Faseb J, 1989. 3(1): p. 31-6.
151. Radomski MW, RM Palmer, and S Moncada, An L-arginine/nitric oxide pathway present in human platelets regulates aggregation. Proc Natl Acad Sci U S A, 1990. 87(13): p. 5193-7.
152. Broere A, AH Van Den Meiracker, F Boomsma, FH Derkx, AJ Veld, and MA Schalekamp, Human renal and systemic hemodynamic, natriuretic, and neurohumoral responses to different doses of L-NAME. Am J Physiol, 1998. 275(6 Pt 2): p. F870-7.
153. Haynes WG, JP Noon, BR Walker, and DJ Webb, L-NMMA increases blood pressure in man. Lancet, 1993. 342(8876): p. 931-2.
154. Moncada S, A Higgs, and R Furchgott, International Union of Pharmacology Nomenclature in Nitric Oxide Research. Pharmacol Rev, 1997. 49(2): p. 137-42.
155. Keefer LK, Progress toward clinical application of the nitric oxide-releasing diazeniumdiolates. Annu Rev Pharmacol Toxicol, 2003. 43: p. 585-607.

56
156. Fiorucci S, E Antonelli, L Santucci, O Morelli, M Miglietti, B Federici, R Mannucci, P Del Soldato, and A Morelli, Gastrointestinal safety of nitric oxide-derived aspirin is related to inhibition of ICE-like cysteine proteases in rats. Gastroenterology, 1999. 116(5): p. 1089-106.
157. Burgaud JL, JP Riffaud, and P Del Soldato, Nitric-oxide releasing molecules: a new class of drugs with several major indications. Curr Pharm Des, 2002. 8(3): p. 201-13.
158. Bjorne HH, J Petersson, M Phillipson, E Weitzberg, L Holm, and JO Lundberg, Nitrite in saliva increases gastric mucosal blood flow and mucus thickness. J Clin Invest, 2004. 113(1): p. 106-14.
159. Allaker RP, LS Silva Mendez, JM Hardie, and N Benjamin, Antimicrobial effect of acidified nitrite on periodontal bacteria. Oral Microbiol Immunol, 2001. 16(4): p. 253-6.
160. Weller R, RJ Price, AD Ormerod, N Benjamin, and C Leifert, Antimicrobial effect of acidified nitrite on dermatophyte fungi, Candida and bacterial skin pathogens. J Appl Microbiol, 2001. 90(4): p. 648-52.
161. Wagner DA, VR Young, and SR Tannenbaum, Mammalian nitrate biosynthesis: incorporation of 15NH3 into nitrate is enhanced by endotoxin treatment. Proc Natl Acad Sci U S A, 1983. 80(14): p. 4518-21.
162. Stuehr DJ and MA Marletta, Mammalian nitrate biosynthesis: mouse macrophages produce nitrite and nitrate in response to Escherichia coli lipopolysaccharide. Proc Natl Acad Sci U S A, 1985. 82(22): p. 7738-42.
163. Marletta MA, PS Yoon, R Iyengar, CD Leaf, and JS Wishnok, Macrophage oxidation of L-arginine to nitrite and nitrate: nitric oxide is an intermediate. Biochemistry, 1988. 27(24): p. 8706-11.
164. Hibbs JB, Jr., Z Vavrin, and RR Taintor, L-arginine is required for expression of the activated macrophage effector mechanism causing selective metabolic inhibition in target cells. J Immunol, 1987. 138(2): p. 550-65.
165. Hibbs JB, Jr., Infection and nitric oxide. J Infect Dis, 2002. 185 Suppl 1: p. S9-17. 166. Vallance BA, W Deng, M De Grado, C Chan, K Jacobson, and BB Finlay,
Modulation of inducible nitric oxide synthase expression by the attaching and effacing bacterial pathogen citrobacter rodentium in infected mice. Infect Immun, 2002. 70(11): p. 6424-35.
167. Fang FC, Perspectives series: host/pathogen interactions. Mechanisms of nitric oxide-related antimicrobial activity. J Clin Invest, 1997. 99(12): p. 2818-25.
168. Fang FC, Antimicrobial reactive oxygen and nitrogen species: concepts and controversies. Nat Rev Microbiol, 2004. 2(10): p. 820-32.
169. Nathan C and MU Shiloh, Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc Natl Acad Sci U S A, 2000. 97(16): p. 8841-8.
170. Schapiro JM, SJ Libby, and FC Fang, Inhibition of bacterial DNA replication by zinc mobilization during nitrosative stress. Proc Natl Acad Sci U S A, 2003. 100(14): p. 8496-501.
171. Lundberg JO, I Ehren, O Jansson, J Adolfsson, JM Lundberg, E Weitzberg, K Alving, and NP Wiklund, Elevated nitric oxide in the urinary bladder in infectious and noninfectious cystitis. Urology, 1996. 48(5): p. 700-2.
172. Alving K, E Weitzberg, and JM Lundberg, Increased amount of nitric oxide in exhaled air of asthmatics. Eur Respir J, 1993. 6(9): p. 1368-70.
173. Lundberg JO, PM Hellstrom, JM Lundberg, and K Alving, Greatly increased luminal nitric oxide in ulcerative colitis. Lancet, 1994. 344(8938): p. 1673-4.

57
174. Ueki Y, S Miyake, Y Tominaga, and K Eguchi, Increased nitric oxide levels in patients with rheumatoid arthritis. J Rheumatol, 1996. 23(2): p. 230-6.
175. Lundberg JO, Airborne nitric oxide: inflammatory marker and aerocrine messenger in man. Acta Physiol Scand Suppl, 1996. 633: p. 1-27.
176. Suschek CV, O Schnorr, and V Kolb-Bachofen, The role of iNOS in chronic inflammatory processes in vivo: is it damage-promoting, protective, or active at all? Curr Mol Med, 2004. 4(7): p. 763-75.
177. Haraoka M, L Hang, B Frendeus, G Godaly, M Burdick, R Strieter, and C Svanborg, Neutrophil recruitment and resistance to urinary tract infection. J Infect Dis, 1999. 180(4): p. 1220-9.
178. Wheeler MA, SD Smith, G Garcia-Cardena, CF Nathan, RM Weiss, and WC Sessa, Bacterial infection induces nitric oxide synthase in human neutrophils. J Clin Invest, 1997. 99(1): p. 110-6.
179. Ehren I, A Hosseini, M Herulf, JO Lundberg, and NP Wiklund, Measurement of luminal nitric oxide in bladder inflammation using a silicon balloon catheter: a novel minimally invasive method. Urology, 1999. 54(2): p. 264-7.
180. Morcos E, OT Jansson, J Adolfsson, G Kratz, and NP Wiklund, Endogenously formed nitric oxide modulates cell growth in bladder cancer cell lines. Urology, 1999. 53(6): p. 1252-7.
181. Jansson OT, E Morcos, L Brundin, JO Lundberg, J Adolfsson, M Soderhall, and NP Wiklund, The role of nitric oxide in bacillus Calmette-Guerin mediated anti-tumour effects in human bladder cancer. Br J Cancer, 1998. 78(5): p. 588-92.
182. Poljakovic M and K Persson, Urinary tract infection in iNOS-deficient mice with focus on bacterial sensitivity to nitric oxide. Am J Physiol Renal Physiol, 2003. 284(1): p. F22-31.
183. Austin PF, AJ Casale, MP Cain, RC Rink, and SJ Weintraub, Lipopolysaccharide and inflammatory cytokines cause an inducible nitric oxide synthase-dependent bladder smooth muscle fibrotic response. J Urol, 2003. 170(2 Pt 1): p. 645-8.
184. Ribeiro RA, HC Freitas, MC Campos, CC Santos, FC Figueiredo, GA Brito, and FQ Cunha, Tumor necrosis factor-alpha and interleukin-1beta mediate the production of nitric oxide involved in the pathogenesis of ifosfamide induced hemorrhagic cystitis in mice. J Urol, 2002. 167(5): p. 2229-34.
185. Horsburgh MJ, SJ Wharton, M Karavolos, and SJ Foster, Manganese: elemental defence for a life with oxygen. Trends Microbiol, 2002. 10(11): p. 496-501.
186. Poole RK and MN Hughes, New functions for the ancient globin family: bacterial responses to nitric oxide and nitrosative stress. Mol Microbiol, 2000. 36(4): p. 775-83.
187. Nunoshiba T, T deRojas-Walker, JS Wishnok, SR Tannenbaum, and B Demple, Activation by nitric oxide of an oxidative-stress response that defends Escherichia coli against activated macrophages. Proc Natl Acad Sci U S A, 1993. 90(21): p. 9993-7.
188. Archer S, Measurement of nitric oxide in biological models. Faseb J, 1993. 7(2): p. 349-60.
189. Herulf M, T Ljung, PM Hellstrom, E Weitzberg, and JO Lundberg, Increased luminal nitric oxide in inflammatory bowel disease as shown with a novel minimally invasive method. Scand J Gastroenterol, 1998. 33(2): p. 164-9.
190. Feelisch M, T Rassaf, S Mnaimneh, N Singh, NS Bryan, D Jourd'Heuil, and M Kelm, Concomitant S-, N-, and heme-nitros(yl)ation in biological tissues and fluids: implications for the fate of NO in vivo. Faseb J, 2002. 16(13): p. 1775-85.
191. Potter LC, PD Millington, GH Thomas, RA Rothery, G Giordano, and JA Cole, Novel growth characteristics and high rates of nitrate reduction of an Escherichia coli

58
strain, LCB2048, that expresses only a periplasmic nitrate reductase. FEMS Microbiol Lett, 2000. 185(1): p. 51-7.
192. Odenholt-Tornqvist I, Studies on the postantibiotic effect and the postantibiotic sub-MIC effect of meropenem. J Antimicrob Chemother, 1993. 31(6): p. 881-92.
193. Nilsson AI, OG Berg, O Aspevall, G Kahlmeter, and DI Andersson, Biological costs and mechanisms of fosfomycin resistance in Escherichia coli. Antimicrob Agents Chemother, 2003. 47(9): p. 2850-8.
194. Dominguez MC, M de La Rosa, and MV Borobio, Application of a spectrophotometric method for the determination of post-antibiotic effect and comparison with viable counts in agar. J Antimicrob Chemother, 2001. 47(4): p. 391-8.
195. Evans Ta, Zbl. Bakt. Abt II, 1934. 91: p. 1. 196. Tarr, Bacteriostatic Action of Nitrates. Nature, 1941. 147(3727): p. 417-418. 197. Tompkin RB, LN Christiansen, and AB Shaparis, Enhancing nitrite inhibition of
Clostridium botulinum with isoascorbate in perishable canned cured meat. Appl Environ Microbiol, 1978. 35(1): p. 59-61.
198. Reddy D, JR Lancaster, Jr., and DP Cornforth, Nitrite inhibition of Clostridium botulinum: electron spin resonance detection of iron-nitric oxide complexes. Science, 1983. 221(4612): p. 769-70.
199. Dykhuizen RS, R Frazer, C Duncan, CC Smith, M Golden, N Benjamin, and C Leifert, Antimicrobial effect of acidified nitrite on gut pathogens: importance of dietary nitrate in host defense. Antimicrob Agents Chemother, 1996. 40(6): p. 1422-5.
200. Long R, B Light, and JA Talbot, Mycobacteriocidal action of exogenous nitric oxide. Antimicrob Agents Chemother, 1999. 43(2): p. 403-5.
201. Pacelli R, DA Wink, JA Cook, MC Krishna, W DeGraff, N Friedman, M Tsokos, A Samuni, and JB Mitchell, Nitric oxide potentiates hydrogen peroxide-induced killing of Escherichia coli. J Exp Med, 1995. 182(5): p. 1469-79.
202. Lepoivre M, F Fieschi, J Coves, L Thelander, and M Fontecave, Inactivation of ribonucleotide reductase by nitric oxide. Biochem Biophys Res Commun, 1991. 179(1): p. 442-8.
203. C RRaS, The biochemical and serological relationship of organism of genus Proteus. J. Bacterial, 1945. 49: p. 419.
204. Athmann C, N Zeng, T Kang, EA Marcus, DR Scott, M Rektorschek, A Buhmann, K Melchers, and G Sachs, Local pH elevation mediated by the intrabacterial urease of Helicobacter pylori cocultured with gastric cells. J Clin Invest, 2000. 106(3): p. 339-47.
205. Poole RK, MF Anjum, J Membrillo-Hernandez, SO Kim, MN Hughes, and V Stewart, Nitric oxide, nitrite, and Fnr regulation of hmp (flavohemoglobin) gene expression in Escherichia coli K-12. J Bacteriol, 1996. 178(18): p. 5487-92.
206. Nunoshiba T, T DeRojas-Walker, SR Tannenbaum, and B Demple, Roles of nitric oxide in inducible resistance of Escherichia coli to activated murine macrophages. Infect Immun, 1995. 63(3): p. 794-8.
207. Poock SR, ER Leach, JW Moir, JA Cole, and DJ Richardson, Respiratory detoxification of nitric oxide by the cytochrome c nitrite reductase of Escherichia coli. J Biol Chem, 2002. 277(26): p. 23664-9.
208. Watmough NJ, G Butland, MR Cheesman, JW Moir, DJ Richardson, and S Spiro, Nitric oxide in bacteria: synthesis and consumption. Biochim Biophys Acta, 1999. 1411(2-3): p. 456-74.

59
209. Braun C and WG Zumft, Marker exchange of the structural genes for nitric oxide reductase blocks the denitrification pathway of Pseudomonas stutzeri at nitric oxide. J Biol Chem, 1991. 266(34): p. 22785-8.
210. Kay CJ and MJ Barber, Assimilatory nitrate reductase from Chlorella. Effect of ionic strength and pH on catalytic activity. J Biol Chem, 1986. 261(30): p. 14125-9.
211. Brandt R, KE Guyer, and WL Banks, Jr., Urinary glucose and vitamin C. Am J Clin Pathol, 1977. 68(5): p. 592-4.
212. R.C. Y, The effect of pH on the growth of bacteria in urine. J Urology, 1940. 44: p. 699-713.
213. Mizutani T, N Akinaga, and H Takatera, Urinary nitrite in Japanese patients with urinary tract infections. Sci Total Environ, 1989. 80(2-3): p. 161-5.
214. Cherek DR, WC Lowe, and TT Friedman, Effects of ammonium chloride on urinary pH and cigarette smoking behavior. Clin Pharmacol Ther, 1981. 29(6): p. 762-70.
215. Tannenbaum SR, JS Wishnok, and CD Leaf, Inhibition of nitrosamine formation by ascorbic acid. Am J Clin Nutr, 1991. 53(1 Suppl): p. 247S-250S.


















