
András N. Spaan, Thomas Henry, Willemien J.M. van ... Host & Microbe, Volume 13 Supplemental...
14
Cell Host & Microbe, Volume 13 Supplemental Information The Staphylococcal Toxin Panton-Valentine Leukocidin Targets Human C5a Receptors András N. Spaan, Thomas Henry, Willemien J.M. van Rooijen, Magali Perret, Cédric Badiou, Piet C. Aerts, Johan Kemmink, Carla J.C. de Haas, Kok P.M. van Kessel, François Vandenesch, Gérard Lina, and Jos A.G. van Strijp
Transcript of András N. Spaan, Thomas Henry, Willemien J.M. van ... Host & Microbe, Volume 13 Supplemental...

Cell Host & Microbe, Volume 13
Supplemental Information
The Staphylococcal Toxin Panton-Valentine Leukocidin Targets Human C5a Receptors
András N. Spaan, Thomas Henry, Willemien J.M. van Rooijen, Magali Perret, Cédric Badiou, Piet C. Aerts, Johan Kemmink, Carla J.C. de Haas, Kok P.M. van Kessel, François Vandenesch, Gérard Lina, and Jos A.G. van Strijp
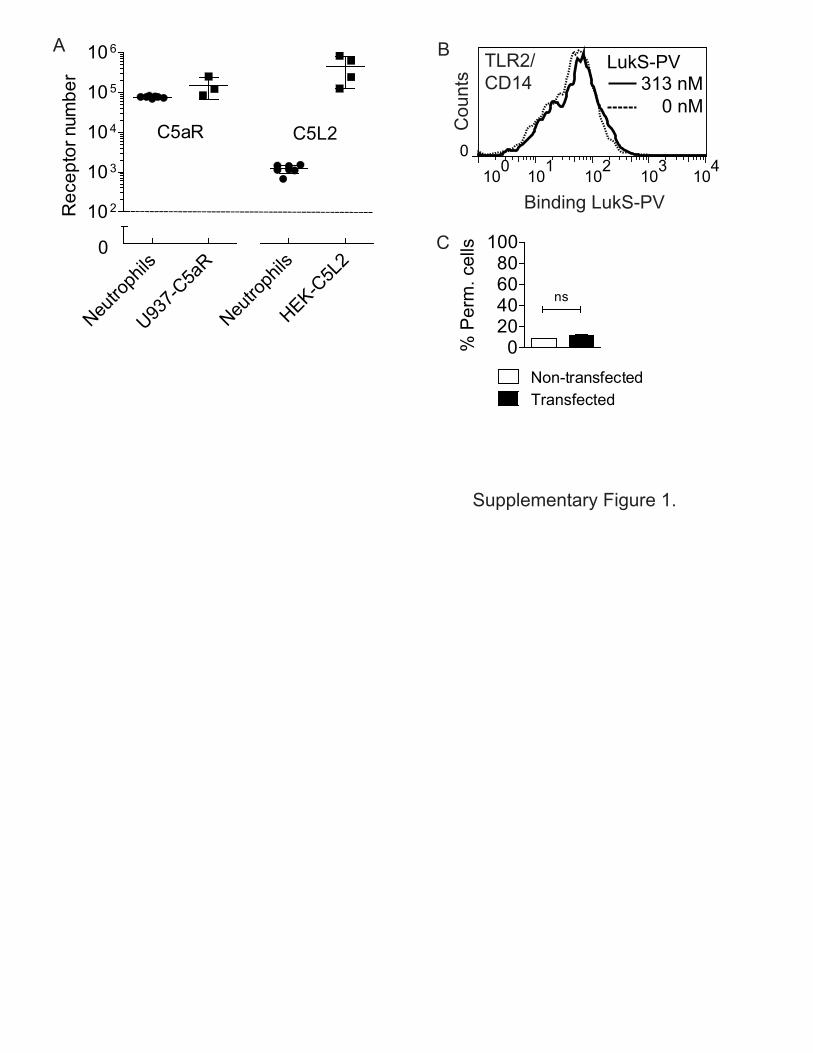
Supplementary Figure 1.
Neutro
phils
U937-C
5aR
102
103
104
105
106
C5aR
Rec
epto
r num
ber
Neutro
phils
HEK-C5L
2
C5L2
0
A
Binding LukS-PV
TLR2/CD14
Cou
nts
020406080
100
ns
% P
erm
. cel
ls
TransfectedNon-transfected
B
C
313 nM 0 nM
LukS-PV

Figure S1 (Related to Figure 1). (A) Receptor expression levels of stably transfected cell lines in relation to freshly isolated neutrophils from healthy donors. (B, C) The TLR2/CD14 Receptor complex does not mediate PVL-induced pore formation. (B) No binding of LukS-PV to HEK cells stably co-transfected with TLR2 and CD14. (C) No PVL induced pore formation (concentration PVL 625 nM) in transfected HEK cells. Permeable cells are DAPI-positive cells. Bars express the standard deviation, with n=2. Histograms depict a representative example.

0
20
40
60
80
100
10 100 1000CHIPS (nM)
% P
erm
eabl
e ce
llsA C CHIPS
(nM) PVL (nM)
Ratio CHIPS:PVL
Recombinant proteins 500.00 2.34 213.67
5% o/n CCY supernatant 18.65 177.29 0.11
5% o/n IMDM supernatant 24.16 0.29 83.31
5% o/n TSB supernatant 0.19 11.43 0.02
Supplementary Figure 2
B
0 5 10 150
500
1.000
1.500
2.000
24
CCY
Time (hours)
Prot
ein
(nM
)
0 5 10 15 24
IMDM
Time (hours)0 5 10 15 24
LukS-PVCHIPS
TSB
Time (hours)
0
2000
4000
6000
8000
CHIPS-PVL+
CHIPS+PVL+
PI F
luor
esce
nce
(AU
)ED
CCYIM
DMTSB
0
20
40
60
80
100 U937U937-C5aR
% P
erm
eabl
e ce
lls

Figure S2 (Related to Figure 2). CHIPS is produced in insufficient amounts to compete with PVL for interaction with the C5aR. (A) Pore formation inhibition in U937-C5aR cells by different concentrations of CHIPS (PVL concentration 2.34 nM, incubation time 30 min). (B) Expression over time of CHIPS and LukS-PV by S. aureus USA300-LAC in CCY, IMDM and TSB broth. Detection of proteins by ELISA. (C) Molar ratio of CHIPS versus PVL needed to protect cells from PVL-induced pore formation using recombinant proteins based on panel A. Ratio of CHIPS versus PVL in 5% overnight supernatant as detected in panel B and used in panel C. (D) Pore formation in U937-C5aR cells induced by 5% overnight CCY, IMDM or TSB based bacterial supernatant containing CHIPS and PVL as shown in panel C (incubation time 30 min). (E) Pore formation in PMA-differentiated THP-1 cells induced by 0.2% overnight supernatant of various PVL-positive clinical S. aureus isolates with or without the gene encoding CHIPS (incubation time 180 min). Bars express the standard deviation, with n=3.

C5a Bgr
HumanNeutro
αC5aRIso
HumanNeutro
Binding αC5aR antibody
MouseNeutro
Binding C5a
RabbitNeutro
RabbitNeutro
MouseNeutro
MouseNeutro
MacaqueNeutro
MacaqueNeutroC
ount
s
Supplementary Figure 3
Cou
nts
A B

Figure S3 (Related to Figure 4). Neutrophils from different species express similar levels of C5aR. (A) Fluorescent C5a binding (240 nM) to human, macaque and rabbit neutrophils. (B) Monoclonal antibody binding to the C5aR of different species (human and macaque: mouse anti-human C5aR antibody clone S5/1; rabbit: mouse anti-human C5aR antibody clone 347214; mouse: rat-anti-mouse C5aR clone 10/92). Histograms depict a representative example.
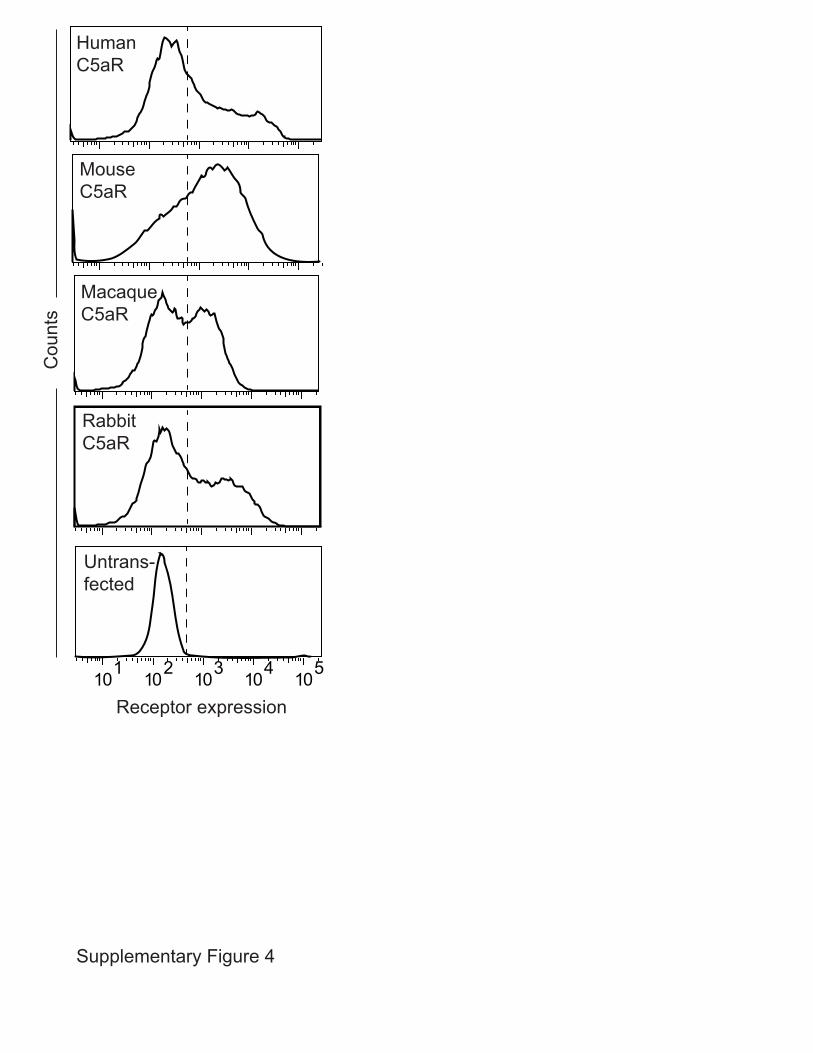
Supplementary Figure 4
HumanC5aR
MouseC5aR
MacaqueC5aR
RabbitC5aR
Cou
nts
Untrans-fected
Receptor expression

Figure S4 (Related to Figure 5 and Experimental Procedures). Receptor expression levels on transient transfected HEK cells. FLAG-tag detection based expression levels of C5aR of different species. Receptor expressing cell populations right of the dashed lines were gated and used for experiments. Histograms depict a representative example.

Untrans-fected
Cou
nts
Cou
nts
Receptor expression
Receptor expression
Untrans-fected
N-C5aR
N-C5L2
N-C5aRCore-C3aRChimera
N-C3aRCore-C5aRChimera
HumanC3aR
A
B
Supplementary Figure 5

Figure S5 (Related to Figure 6 and Experimental Procedures). Receptor expression levels on transient transfected HEK cells. (A) HA-tag detection based expression levels of human N-terminal receptors. (B) FLAG-tag detection based expression levels of human C5aR-C3aR chimeras. Receptor expressing cell populations right of the dashed lines were gated and used for experiments. Histograms depict a representative sample.

Table S1 (Related to Experimental Procedures). Bacterial strains used in this study to produce recombinant proteins. All genes were cloned, omitting the region predicted to encode the signal peptide, as determined on the SignalP V3.0 World Wide Web Prediction Server (http://www.cbs.dtu.dk/services/SignalP/). Protein S. aureus strain Reference LukS-PV ATCC 49775 (Perret et al., 2012) LukF-PV ATCC 49775 (Perret et al., 2012) CHIPS Newman (de Haas et al., 2004) CHIPS31-121 Newman (Haas et al., 2005) FLIPr-Like MW2 (Prat et al., 2009)
Table S2 (Related to Experimental Procedures). Primer sequences used in this study to produce plasmids for transient transfections or for qRT-PCR (F: Forward primer; R: Reverse primer; TM: transmembrane domain; N: N-terminus). Restriction enzyme recognition sites are underlined. FLAG tag sequences are double underlined.
Receptor Primers Human C5aR. Accession nr.: NM_001736
Fwd: 5’-CCGGAATTCATGGACTACAAGGACGACGACGACAAGATGAACTCCTTCAATTATACC-3’ Rev: 5’-GCTCTAGACTACACTGCCTGGGTCTTCT-3’
Human C5L2. Accession nr.: NM_001271749
Fwd: 5’-CCGGAATTCATGGACTACAAGGACGACGACGACAAGATGGGGAACGATTCTGTCAGCTACG-3’ Rev: 5’-GCTCTAGACTACACCTCCATCTCCGAGACC-3’
Human C3aR. Accession nr.: BC020742
Fwd: 5’-CGGGATCCATGGACTACAAGGACGACGATGACAAGATGGCGTCTTTCTCTGCTGAGACC-3’ Rev: 5’-GCTCTAGATCACACAGTTGTACTATTTC-3’
Human Chimera N-C3aR / TM-C5aR
Fwd: 5’-CGGGATCCATGGACTACAAGGACGACGATGACAAGATGGCGTCTTTCTCTGCTGAGACC-3’ Rev: 5’-GACCAAGGCCAGGATTACTGGGGGCTCATTCCATG-3’ Fwd: 5’-ATTGAGCCCCCAGTAATCCTGGCCTTGGTCATCTTTG-3’ Rev: 5’-GCTCTAGACTACACTGCCTGGGTCTTCT-3’
Human Chimera N-C5aR / TM-C3aR
Fwd: 5’-CCGGAATTCATGGACTACAAGGACGACGACGACAAGATGAACTCCTTCAATTATACC-3’ Rev: 5’-GACCATGGAGAGAATGTCTGGAACACGCAGCGTGTTAG-3’ Fwd: 5’-TGCGTGTTCCAGACATTCTCTCCATGGTCATTCTC-3’ Rev: 5’-GCTCTAGATCACACAGTTGTACTATTTC-3’
Mouse C5aR (C57BL/6). Accession nr.: BC125643
Fwd: 5’-CCGGAATTCATGGACTACAAGGACGACGACGACAAGATGGACCCCATAGATAACAGCAG-3’ Rev: 5’-GCTCTAGACTACACCGCCTGACTCTTCC-3’
Macaque C5aR (M. fascicularis). Accession nr.: EHH59724
Fwd: 5’-CCGGAATTCATGGACTACAAGGACGACGACGACAAGATGAACTCCTTCAATTATACC-3’ Rev: 5’-GCTCTAGACTACACTGCCTGGGTCTTCT-3’
Rabbit C5aR (N.Z. White). Accession nr.: AF068680
Fwd: 5’-CCGGAATTCATGGACTACAAGGACGACGACGACAAGATGGAAAATAGCACGTACGATTACACC-3’ Rev: 5’-GCTCTAGATCACACCGCTTGGCACTTGTCG-3’
Human N-C5aR Fwd: 5’-TAGTGAAGATCTATGAACTCCTTCAATTATACCACCC-3’ Rev: 5’-AGTACCATCGATGATGTCTGGAACACGCAGCG-3’
Human N-C5L2 Fwd: 5’-TAGTGAAGATCTATGGGGAACGATTCTGTCAGCTACG-3’ Rev: 5’-CCATCGATGGCCACGCGCAGCGGGTCGA-3’
Human C5aR (qRT-PCR)
Fwd: 5’-GGACACTATGGCCCAGAAGA-3’ Rev: 5’-CTGGGAAAGACAGGCAACAT-3’
Human C5aR (qRT-PCR)
Fwd: 5’-CACCCACTCCTCCACCTTTGAC-3’ Rev: 5’-GTCCACCACCCTGTTGCTGTAG-3’

Supplemental Experimental Procedures Quantification of CHIPS and PVL expression in supernatants. Expression of CHIPS and PVL in the supernatant of S. aureus USA300-LAC was measured at multiple time points of growth in casein hydrolysate and yeast extract (CCY) medium (Lipinska et al., 2011; Woodin, 1959), tryptic soy broth (TSB), or Iscove’s Modified Dulbecco’s Medium (IMDM). CHIPS and LukS-PV were detected in supernatant by specific ELISA techniques as described elsewhere (Badiou et al., 2008; Badiou et al., 2010; de Haas et al., 2004). Quantification of PVL expression was based on LukS-PV detection. Clinical S. aureus isolates. Clinical S. aureus strains isolated from patients hospitalized for severe community-acquired pneumonia were collected during prospective studies that were previoulsy described (Gillet et al., 2002; Sicot et al., 2013). The genomic content of the clinical isolates was analysed by commercial microarray (StaphyType, Alere Technologies) as previously described (Albrecht et al., 2011), following manufacturer's instructions to determine the presence or absence of the PVL- and CHIPS-encoding genes. To generate supernatant, single hemolytic colonies were grown overnight in CCY broth (Lipinska et al., 2011; Woodin, 1959). Supernatants were sterilized by a 0.2 μm filter diluted 1/625 before addition onto PMA-differentiated THP-1 macrophages in the presence of propidium iodide (PI) (5 µg/ml). Pore formation was assessed by measuring PI-incorporation on a Tecan fluorimeter after 180 min incubation. Receptor expression determination on neutrophils of different species. Freshly isolated human, macaque and rabbit leukocytes were incubated in a total volume of 50 μL at 5x106 cell/mL with 240 nM FITC-labeled C5a (Bachem). FITC-labeled C5a was produced as decribed before (de Haas et al., 2004; Prat et al., 2009). In a total volume of 50 μL at 5x106 cell/mL, human and macaque leukocytes were incubated with mouse anti-human C5aR antibody clone S5/1 (BioLegend), followed by FITC-conjugated goat-anti-mouse antibody (Dako). Rabbit leukocytes were incubated with PE-conjugated mouse anti-human C5aR antibody clone 347214 (R&D Systems). Murine leukocytes were incubated with Alexa488-conjugated rat-anti-mouse clone 10/92 (AbD SeroTec). Fluorescences were detected by flow cytometry.

Supplemental References:
Albrecht, N., Jatzwauk, L., Slickers, P., Ehricht, R., and Monecke, S. (2011). Clonal replacement of epidemic methicillin-resistant Staphylococcus aureus strains in a German university hospital over a period of eleven years. PLoS One 6, e28189. Badiou, C., Dumitrescu, O., Croze, M., Gillet, Y., Dohin, B., Slayman, D.H., Allaouchiche, B., Etienne, J., Vandenesch, F., and Lina, G. (2008). Panton-Valentine leukocidin is expressed at toxic levels in human skin abscesses. Clin Microbiol Infect 14, 1180-1183. Badiou, C., Dumitrescu, O., George, N., Forbes, A.R., Drougka, E., Chan, K.S., Ramdani-Bouguessa, N., Meugnier, H., Bes, M., Vandenesch, F., et al. (2010). Rapid detection of Staphylococcus aureus Panton-Valentine leukocidin in clinical specimens by enzyme-linked immunosorbent assay and immunochromatographic tests. J Clin Microbiol 48, 1384-1390. de Haas, C.J., Veldkamp, K.E., Peschel, A., Weerkamp, F., Van Wamel, W.J., Heezius, E.C., Poppelier, M.J., Van Kessel, K.P., and van Strijp, J.A. (2004). Chemotaxis inhibitory protein of Staphylococcus aureus, a bacterial antiinflammatory agent. J Exp Med 199, 687-695. Gillet, Y., Issartel, B., Vanhems, P., Fournet, J.C., Lina, G., Bes, M., Vandenesch, F., Piemont, Y., Brousse, N., Floret, D., et al. (2002). Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 359, 753-759. Haas, P.J., de Haas, C.J., Poppelier, M.J., van Kessel, K.P., van Strijp, J.A., Dijkstra, K., Scheek, R.M., Fan, H., Kruijtzer, J.A., Liskamp, R.M., et al. (2005). The structure of the C5a receptor-blocking domain of chemotaxis inhibitory protein of Staphylococcus aureus is related to a group of immune evasive molecules. J Mol Biol 353, 859-872. Lipinska, U., Hermans, K., Meulemans, L., Dumitrescu, O., Badiou, C., Duchateau, L., Haesebrouck, F., Etienne, J., and Lina, G. (2011). Panton-Valentine leukocidin does play a role in the early stage of Staphylococcus aureus skin infections: a rabbit model. PLoS One 6, e22864. Perret, M., Badiou, C., Lina, G., Burbaud, S., Benito, Y., Bes, M., Cottin, V., Couzon, F., Juruj, C., Dauwalder, O., et al. (2012). Cross-talk between S. aureus leukocidins-intoxicated macrophages and lung epithelial cells triggers chemokine secretion in an inflammasome-dependent manner. Cell Microbiol. Prat, C., Haas, P.J., Bestebroer, J., de Haas, C.J., van Strijp, J.A., and van Kessel, K.P. (2009). A homolog of formyl peptide receptor-like 1 (FPRL1) inhibitor from Staphylococcus aureus (FPRL1 inhibitory protein) that inhibits FPRL1 and FPR. J Immunol 183, 6569-6578. Sicot, N., Khanafer, N., Meyssonnier, V., Dumitrescu, O., Tristan, A., Bes, M., Lina, G., Vandenesch, F., Vanhems, P., Etienne, J., et al. (2013). Methicillin resistance is not a predictor of severity in community-acquired Staphylococcus aureus necrotizing pneumonia-results of a prospective observational study. Clin Microbiol Infect 19, E142-E148. Woodin, A.M. (1959). Fractionation of a leucocidin from Staphylococcus aureus. Biochem J 73, 225-237.


















